苯環氧乙烷光學異構物對人類淋巴母細胞致毒性與致突變性之研究; Cytotoxicity and Mutagenicity of Styrene Oxide Optical Isomers in Human Cultured Lymphoblastoid Cells
57
0
0
全文
(2) 目. 錄. 中文摘要 .......................................................................................................... 1 英文摘要 .......................................................................................................... 3 第一章 前言 ................................................................................................... 5 第二章 研究目的 ........................................................................................... 6 第三章 文 獻 探 討 ..................................................................................... 6 第一節 苯乙烯 ............................................................................................ 6 第二節 苯環氧乙烷 ................................................................................. 10 第三節 苯環氧乙烷的光學異構物 ........................................................ 13 第四節 hprt 與 tk 基因 ........................................................................... 18 第四章 材料與方法 ..................................................................................... 20 第一節 材料 .............................................................................................. 20 第二節 方法 .............................................................................................. 21 第五章 結. 果 ............................................................................................. 25. 第一節 SO 與 SO 光 學 異 構 物 對 TK6 細胞之毒性 ........................... 25 第二節 SO 與 SO 光 學 異 構 物 對 TK6 細胞之致突變性 ................... 26 第六章 討. 論 ............................................................................................. 28. 第一節 SO 與其光學異構物對 TK6 細胞之毒性 ............................... 28 第二節 SO 與 SO 不同光學異構物致突變性 ..................................... 29 第七章 結論 ................................................................................................. 33 第八章 參考資料 ......................................................................................... 34. i.
(3) 圖表目錄 圖 1. 180μM SO 與 SO 光學異構物誘發 TK6 細胞 ...................... 42 圖 2 .以細胞相對存活率(RCN )評估 SO 與 SO 光學異構物對 TK6 細胞之毒性 ..................................................................................... 43 圖 3. 以細胞群落形成能力(PE)評估 SO 與 SO 光學異構物對 TK6 細胞之毒性 ..................................................................................... 44 圖 4. SO 與 SO 光 學 異 構 物 在 TK6 細胞誘發 hprt. 突變率 ......... 45. 圖 5. SO 與 SO 光 學 異 構 物 在 TK6 細胞誘發 tk fast ..................... 46 圖 6. SO 與 SO 光學異構物對在 TK6 細胞誘發 tk total ................ 47 圖 7. SO 與 SO 光學異構物對在 TK6 細胞誘發 tk slow ................ 48. ii.
(4) 致. 謝. 在結婚生子,工作八年之後,能夠完全放棄工作,全心投入學業 的人並不多,而我即是那幸運人之一。能夠完成學業最主要是來自先 生的支持,雖然因學業與工作的關係,他不在我身邊,但是每次一遇 到困難他必定幫我解決,雖然他的脾氣不是很好,但是真的要非常感 謝他對我這兩年的體諒與支持。另外!我的兩個可愛的孩子,他們也 能夠體諒媽媽陪他們的時間變少了,而且還不能打電動(因為媽媽要 用電腦) ,七月一日即是電腦時間還給他們的日子,雖然他們有少許抱 怨,但至少還很體諒。子瑄、禹心,媽媽畢業了! 學習是無止盡的,這兩年學到很多,包括人與人之間的問題,技 術上之學習。要感謝的人有一拖拉庫,要謝謝賴主任與職安系助教室, 那兒常常是無法解決問題時的去處。謝謝江老師在實驗技術上之指 導,讓我兩年來學到以前從未學過的技術,並常陪我熬夜到半夜 2、3 點。保萱謝謝你常常隨叫隨到,替我解決許多統計問題。還有謝謝陳 昭玲教授,熱心幫我改英文摘要。最後要謝謝我的家人與 Mila 沒有 他們替我接送孩子並照顧他們,使我無後顧之憂,沒有他們我想我無 法如此順利完成學業。 最後希望還在奮戰的老公,你也加油,2 年後我們等著你拿到博 士學位。. iii.
(5) 中文摘要 苯 乙 烯 (styrene) 為 常 用 化 工 原 料 , 在 生 物 體 內 經 由 cytochrome P- 450(CYP -450) 代 謝 成 苯 環 氧 乙 烷 (styrene oxide;SO),SO 具 有 兩 種 不 同 光 學 異 構 物 (R - SO 與 S- SO)。 SO 已 被 證 實 為 動 物 致 癌 物 , 在 許 多 生 物 系 統 中 可 以 誘 發 細 胞 毒 性 與 突 變 性 , 然 而 目 前 仍 不 清 楚 R - SO 或 S- SO 何 者 扮 演 較 重 要 的 角 色 。 本 研 究 之 目 的 是 研 究 SO 與 其 光 學 異 構 物 在 人 類 淋 巴 母 細 胞 (human lymphoblastoid cell;TK6)之 相 對 致細胞毒性與致突變性。 細 胞 經 不 同 濃 度 的 SO 與 SO 其 光 學 異 構 物 處 理 24 小 時 後,藉 著 測 量 細 胞 恢 復 正 常 生 長 所 需 時 間 (growth delay) 與 細 胞 群 落 形 成 能 力 (plating effecticiency ; PE) , 來 探 討 SO 與 其 不 同 光 學 異 構 物 之 相 對 細 胞 毒 性 。 結 果 顯 示 SO 與 其 光 學 異構物所誘發的細胞毒性會隨著劑量的增加而增加,且呈現 劑 量 反 應 關 係。在 120μ M 濃 度 之 R- SO、S- SO 與 (R+S)- SO (racemic styrene oxide ) 處 理 後 ,TK6 細 胞 的 相 對 存 活 率 依 次 為 50.55、56.25 與 51.24%,180μ M 濃 度 下 分 別 為 12.7、 14.0 與 13.95。 R- SO、 S- SO 與 (R+S)- SO 對 TK6 細 胞 誘 發 的細胞毒性沒有達到統計上之顯著差異。 利. 用. hypoxanthine. guanine. phosphoribosyl- transferase ( hprt) 與 thymidine kinase (tk) 基 因 為 突 變 基 因 指 標 , 探 討 SO 與 其 光 學 異 構 物 的 致 突 變 性 。 結 果 顯 示 SO 與 其 光 學 異 構 物 在 TK6 細 胞 誘 發 的 h p r t 與 tk 基 因 突 變 機 率 均 呈 現 良 好 之 劑 量 反 應 關 係 , 在 TK6 細 胞 S- SO 所 誘 發 之 hprt 基 因 突 變 率 趨 勢 高 於 R - SO (p<0.05) 。 在 120μ M 濃 度 下 R - SO、 S- SO 與 (R+S)- SO 所 引 發 hprt 基 因 的 突 變 率 分 別 為 26.6 ×10 - 6 、 33.0 ×10 -6 與 1.
(6) 35.1 ×10 - 6,在 180μ M 濃 度 下 誘 發 的 hprt 基 因 突 變 率 分 別 為 32.1 ×10 - 6、50.0 ×10 -6 與 45.0. ×10 - 6。 120μ M 濃 度. 下 R - SO、 S- SO 與 (R+S)- SO 對 tk-fast 基 因 ( tk-fast) 之 致 突 變 率 分 別 為 23.05 ×10 - 6、 30.95 ×10 -6 與 28.33 ×10 - 6, tk -slow 基 因 ( tk-slow) 之 致 突 變 率 分 別 為 41.07 × 10 - 6 、 28.01 ×10 -6 與 31.72 ×10 -6 , 但 是 都 沒 有 達 到 統 計 上 之 顯 著差異(p>0.05)。 綜 合 上 述 結 果 顯 示 SO 與 其 光 學 異 構 物 對 TK6 細 胞 會 誘 發 細 胞 毒 性 與 突 變 性 , 雖 然 有 劑 量 反 應 關 係, 但 是 其 間 的 差 異 並 未 達 到 統 計 上 之 意 義 ; 因 此 R - SO 與 S- SO 可 能 在 SO 誘發的細胞毒性與突變性過程中佔有同樣重要地位。. 2.
(7) 英文摘要 Styrene is widely used in the production of plastics. It is metabolic activated by cytochrome P- 450 to styrene- 7, 8 - oxide (SO) with two enantiomeric isomers, R- SO and S-SO.. SO is a. known animal carcinogen and probable human carcinogen.. It is. unclear which isomer plays a more important role in SO - induced cytotoxicity and mutagenicity. In this study the cytotoxicity and mutagenicity of R- SO, S- SO and racemic styrene oxide ((R+S) - SO) were determined in the human lymphoblastoid cell line (TK6). After exposure to various concentrations (60~180μM ) of SO and its isomers for 24 hours, treated cells were counted and plated to determine the relative survival (RS) by the growth delay curve and the plating efficiency (PE)measurement,. respectively.. SO. and. SO. isomers. cytotoxic to TK6 cells in a dose-dependent manner.. were. The RS of. 120μ M R- SO,S- SO and (R+S) - SO -treated cells were 50.6, 56.3 and. 51.2%,. respectively.. At. 180 μ M. R- SO,S- SO. and. (R+S) - SO-treated cells were 12.7 14.0 and 14.0%, respectively. However, there is no statistically significant difference among SO and its optical isomer-induced cytotoxicity. The relative mutagenic potential of the SO and its isomers was. determined. at. hypoxanthine. guanine. phosphoribosyl-transferase ( hprt) and thymidine kinase (tk) gene in TK6 cells.. The mutant frequencies at the hprt and tk gene. induced by SO and its isomers were dose-dependent. more mutatgenic at hprt gene than R- SO (p<0.05).. S- SO was. Treatment of. 120 μ M R- SO, S- SO and (R+S) - SO induced the hprt gene mutation frequencies of 26.6, 32.96 and 35.1 ×10 - 6 , respectively.. 3.
(8) Treatment of 180 μ M R- SO, S- SO and (R+S)- SO induced hprt gene. mutation. respectively.. frequency. of. 32.1,. 50.0. and. 45.0. × 10 - 6 ,. Treatment of 120μ M R- SO, S- SO and (R+S)- SO. induced tk-fast mutant frequency of 31.0, 23.1 and 28.3× 10 - 6 , respectively. Treatment of 120 μ M R- SO, S-SO and (R+S)- SO induced tk- slow mutant frequency of 28.0, 41.1 and 31.7×10 - 6 , respectively. No statistically significant difference among R- SO, S- SO. and. (R+S) - SO- induced. tk-fast. and. tk- slow. mutant. optical. induced. frequencies was found. Results. of. this. study. show. SO. and. SO. cytotoxicity and mutagenicity in a dose dependent relationship,. but they were not significal.. Taken together, these data. suggest that R- SO and S- SO may play a similar important role in (R+S)-SO-induced cytotoxicity and mutagenicity.. 4.
(9) 第一章. 前言. 苯 乙 烯 (styrene) 在 塑 膠 製 品 工 業 中 , 為 製 程 中 不 可 缺 少 的 原 料 , 而 且 使 用 量 逐 年 升 高 。 Styrene 在 生 物 體 內 的 代 謝 主 要 經 由 細 胞 內 質 網 上 的 細 胞 色 素. CYP- 450. (cytochrome - P450 mediated monooxygenase) 酵 素 代 謝 為 為 苯 環 氧 乙 烷 (styrene oxide ; SO) , SO 再 經 由 體 內 去 毒 性 酵素作用後排出體外。 在 動 物 實 驗 已 證 實 SO 具 有 致 癌 性 , 而 其 致 癌 的 生 物 機 轉 目 前 為 止 仍 不 清 楚 。 有 關 SO 在 動 物 實 驗 與 人 體 暴 露 調 查 之 相 關 研 究 很 多 , 其 探 討 內 容 包 括 , SO 代 謝 的 相 關 酵 素 、 SO 之 細 胞 毒 性 、 半 致 死 劑 量 ( L D 5 0 = 1.5 nmol/egg )(Vainio et al.,1977)、SO 在 D N A 上 產 生 的 DNA 共 價 鍵 結 物 ( DNA adduct)、 SO 的 致 突 變 性 、 人 體 暴 露 在 styrene 作 業 環 境 下 之 勞 工 所 造 成 的 姊 妹 染 色 體 交 換 ( sister chromatid exchange; SCE)與 染 色 體 變 異 (chromosomal aberration; CA) 等 。 而 SO 具 有 兩 種 光 學 異 構 物 ( R -styrene oxide ; R- SO 與 S-styrene oxide ; S- SO), 然 而 對 於 SO 的 光 學 異 構物產生的細胞毒性與致突變性之探討並不多。 本 研 究 首 次 以 人 類 淋 巴 母 細 胞 (TK6) 來 探 討 SO 光 學 異 構 物 在 人 類 細 胞 的 致 細 胞 毒 性 與 致 突 變 性。利 用 TK6 細 胞 生 長 情 形 與 細 胞 群 落 形 成 能 力 來 測 試 細 胞 毒 性 , 並 且 使 用 hprt (hypoxanthine guanine phosphoribosyl- transferase) 基 因 與 tk (thymidine kinase ) 基 因 為 標 的 基 因 進 行 突 變 性 的 測 試 , 這 些 結 果 有 助 於 進 一 步 瞭 解 SO 與 其 光 學 異 構 物 之 相 對 致細胞毒性與致突變性。. 5.
(10) 第 二 章 研究目的 比較不同光學異構物(R-SO 與 S-SO)對人類淋巴母細 胞(human lymphoblastoid cell ;TK6)的致細胞毒性與致突變 性,進而了解 SO 光學異構物在人類細胞中何者扮演較重要 的角色。. 第三章. 文 獻 探 討. 第 一 節 苯乙烯 ( 一)苯乙烯之物化特性 styrene 是 一 種 重 要 的 工 業 用 化 學 單 體 , 廣 泛 應 用 於 塑 膠 、 樹 脂 、 塗 料 等 化 學 工 業 。 據 估 計 1992 年 一 年 中 styrene 的 產 量 約 1600 萬噸( Scott,1994 ) ,以 民 國 七 十 三 年 為 例 , 我 國 全 年 使 用 之 styrene 約 高 達 22 萬 公 噸 ( 經濟部,1979) 。 styrene 又 名 vinylbenzene , 1- phenylethylene (CAS No.100 -42- 5 ) 是 無 色 或 淡 黃 色 的 液 體 , 其 密 度 為 0.9059 g/ml, 分 子 量 104.1 6 g/mol,沸 點 為 145 .14 ℃,其 分 子 結 構 如 附 圖 9( Pfaffli and Saamamene , 1993) 。 styrene 在 水 中 的 溶 解 度 很 低 , 在 25℃ 中 約 為 3mM (Howard et al .,1989) , 但 是 可 溶 於 醇 類 ( alcohol)、酯類( ether)、丙酮( acetone ) 及二硫化碳(carbon disulfide)(ATSDR,1990)。 styrene 在 空 氣 中 會 和 氫 氧 基 及 臭 氧 反 應 , 其 半 衰 期 約 為 3.5 ∼ 9 小 時 ( Howard, 1989) 。 在 空 氣 中 經 陽 光 照 射 或 是 環 境 溫 度 超 過 200℃ 時 會 產 生 聚 合 反 應 ( Keith et al.,1987) 。 6.
(11) 低 濃 度 之 styrene 氣 體 有 芳 香 氣 味 , 高 濃 度 時 有 令 人 厭 惡 之 惡臭味。. ( 二)苯乙烯的毒性與致癌性 暴 露 在 styrene 環 境 下 , 會 有 自 覺 性 的 頭 痛 、 疲 倦 、 眩 暈、嗜睡、抑鬱、精神不易集中的感覺,其他自覺症狀有平 衡障害、反應時間延遲與眼、鼻、呼吸道和皮膚的刺激等症 狀。(Seppalainen et al., 1976)。 動 物 暴 露 在 含 有 styrene 的 空 氣 中 , 並 不 會 造 成 癌 症 , 但 是 對 於 暴 露 在 styrene 工 作 環 境 中 是 否 會 造 成 基 因 的 傷 害 , 看 法 並 不 一 致 。 將 大 鼠 與 小 鼠 長 時 間(2 年)暴 露 在 含 有 styrene 的 空 氣 中 ( 50ppm~100ppm) 發 現 , 其 肝 臟 細 胞 中 每 10 8 核. 酸 序 列 , 在 guanine 的 O 6 位 置 上 產 生 的 DNA. adduct 約 有 100 個 , 而 作 者 認 為 每 10 8 核. 酸 中 有 200 ∼. 500 個 DNA adduct 還 不 會 使 小 鼠 與 大 鼠 發 生 腫 瘤 , 因 此 他 認 為. styrene. 是 一 個 非 常 弱 的 親 電 子 性 (weakly. electrophilic activity) 之 化 學 物 質 , 其 與 D N A 之 結 合 力 不 大 , 而 且 在 大 鼠 細 胞 血 漿( plasma ) 中 半 衰 期 只 有 20 分 鐘 , 而 且 產 生 之 DNA adduct 的 量 很 低 ( Otteneder and Lutz, 1999)。 1978 年 有 學 者 對 31 位 勞 工 進 行 生 物 檢 測 與 作 業 環 境 測 定 , 在 空 氣 中 檢 測 到 styrene 含 量 為 10 及 40ppm,mandelic acid 在 尿 液 中 之 含 量 約 為 40 及 100 mg/L,雖 然 有 14 位 勞 工 其 mandelic acid 的 代 謝 產 物 在 尿 液 中 之 含 量 約 為 正 常 人 的 36 倍 , 也 發 現 他 們 的 染 色 體 缺 損 現 象 , 但 是 彼 此 間 並 沒 有 劑 量 反 應 關 係 , 相 同 的 情 況 也 發 生 在 SCE 現 象 中 。 而 作 者 認 為 空 氣 中 styrene 含 量 與 染 色 體 缺 損 之 現 象 , 沒 有 劑 量 反 應 7.
(12) 關 係 之 原 因 可 能 是 styrene 不 是 直 接 造 成 染 色 體 有 缺 損 之 主 要 物 質 , 可 能 有 其 他 因 素 造 成 染 色 體 缺 損 ( Scott and Preston,1993)。 以工廠中非現場作業勞工作為低暴露組,以實驗室研究 員 作 為 未 暴 露 組, 分 析 9 位 暴 露 在 styrene 環 境 中( 7 個 月 ) 之 勞 工 的 周 邊 血 液 淋 巴 球 與 顆 粒 球 ( granulocytes ) 中 O 6- guanine DNA adduct 與 DNA strand breaks。研 究 顯 示 , 利用. 32. P- postlabelling 方 法 檢 測 DNA adduct, 發 現 暴 露 在. styrene 環 境 下 之 勞 工,在 guanine 上 O 6 位 置 所 產 生 之 DNA adduct 明 顯 高 於 低 暴 露 組 (p<0.0001) 。 在 h p r t 基 因 突 變 率 方 面 雖 然 暴 露 組 ( 17.5 ×10 - 6 ) 比 低 暴 露 組 ( 15.7 ×10 - 6) 突 變率稍高,但是並沒有達到統計上之顯著差異,而暴露組 (18.7 × 10 - 6) 之 突 變 率 比 未 暴 露 組 (11.8. × 10 - 6) 高. ( P<0.021)。因 此 作 者 認 為 暴 露 在 styrene 工 作 環 境 中 之 勞 工 , 可 能 會 引 發 基 因 傷 害 , 高 暴 露 者 的 hprt 基 因 突 變 率 並 沒 有 明 顯 高 於 低 暴 露 組 , 但 明 顯 高 於 未 暴 露 組 (Vodicka et al.,1995)。 在 27 個 獨 立 研 究 之 調 查 顯 示,有 11 個 報 告 指 出 暴 露 在 styrene 環 境 中 之 勞 工 , 染 色 體 變 異 有 明 顯 高 於 非 暴 露 者 之 趨 勢 , 12 個 研 究 微 核 ( micronuclei; MN ) 現 象 中 有 4 個 研 究 有 明 顯 差 異,13 個 研 究 SCE 現 象 中 有 3 研 究 有 明 顯 差 異。 總 計 在 52 個 獨 立 研 究 中,有 18 個 研 究 認 為,暴 露 在 styrene 環 境 中 的 勞 工 有 染 色 體 傷 害 的 形 成 ( Scoot and Preston, 1994)。 在 許 多 流 行 病 調 查 發 現 ,styrene 會 使 致 癌 的 風 險 提 高 , 包 括 直 腸 癌 、 淋 巴 瘤 、 喉 頭 癌 等 。 Vodicka 指 出 styrene 作 業勞工中因白血病與淋巴瘤而死亡人數有升高之趨勢,在一 8.
(13) 項針對歐洲地區大規模流行病學調查研究指出,在塑膠成型 工業中之勞工因為癌症而死亡的比例有增加,而且在部分塑 膠成型工業勞工族群中,工作超過一年者得到白血病與淋巴 瘤 的 危 險 性 有 小 幅 度 升 高 之 趨 勢 ( Vodicka et al., 1995)。由 以 上 結 果 顯 示 styrene 是 否 對 人 體 造 成 基 因 傷 害 , 在 暴 露 勞 工中之研究與流行病學之調查都尚未有定論。 文獻指出澳洲、比利時、芬蘭、瑞士、英國和美國設定 勞 工 暴 露 在 styrene 環 境 中 之 標 準 為 100 ppm, 丹 麥 、 挪 威 及 日 本 為 50 ppm, 捷 克 及 東 德 為 47 ppm, 波 蘭 為 24 ppm, 匈 牙 利 為 1 ppm (Scandinavian,1979 )。我 國 行 政 院 勞 工 委 員會公告之勞工作業環境空氣中有害物質容許濃度標準, styrene 暴 露 之 時 量 平 均 容 許 濃 度 (PEL - TWA) 為 50 ppm,短 時 間 時 量 平 均 容 許 濃 度 (PEL- STEL) 為 62.5 ppm ( 行 政 院 勞 工 委 員 會,1995)。從 上 述 顯 示 各 國 所 訂 標 準 styrene 之 容 許 濃 度 標 準 有 差 異 , 但 因 其 體 內 代 謝 物 SO 可 能 誘 發 人 類 產 生 致 癌, 因 此 styrene 的 容 許 濃 度 有 日 漸 下 降 之 趨 勢 。 styrene 本身的細胞毒性很低,超出生物檢測所能偵測到之範圍,必 須藉由生物體內代謝後才會產生毒性(Scott et al.,1994)。. ( 三)苯乙烯的代謝 styrene 主 要 在 肝 臟 進 行 代 謝,而 少 部 分 在 肺 臟 、 腎 臟 、 腸 、 皮 膚 、 淋 巴 系 統 與 紅 血 球 等 其 他 組 織 進 行 代 謝 (Leibman et al.,1968,Belvedere and trusi,1981)。 在 1993 年 Nakajima 等 人 利 用 體 外 試 驗 發 現 ,在 動 物 體 內 參 與 代 謝 的 CYP - 450 酵 素 有 物 種 上 的 差 異,因 此 在 不 同 物 種 間 所 造 成 之 細 胞 毒 性 與 致 突 變 性 也 可 能 會 不 同( Nakajima et al.,1993) 。文 獻 指 出 在 大 鼠 體 內,參 與 styrene 代 謝 為 SO 9.
(14) 的 CYP -450 酵 素 主 要 為 CYP 2E1 、 CYP 2B1、 CYP1A 1/2 與 CYP 2C 11; 在 人 體 中 參 與 styrene 代 謝 為 SO 的 酵 素 主 要 有 CYP 2E1 , CYP 2B6 、 CYP 1F1 等 (Gadberry et al.,1996) 。 而 C arlson 利 用 α- naphthoflavone ( α- NF ) 與 α- methylbenzylaminobenzotriazole ( MBA) 分 別 抑 制 CYP 2B 與 CYP 1A 之 活 性 , 發 現 CYP 2B 與 CYP 1A 並 不 是 參 與 代 謝 styrene 的 主 要 酵 素 , 人 體 中 主 要 參 與 代 謝 styrene 的 主 要 酵 素 為 CYP 2E1 (Carlson et al.,1998) 。 顯 示 不 同 物 種 間 參 與 styrene 代 謝 為 SO 的 酵 素 並 不 相 同 , 縱 使 同 一 物 種,在不同器官中其酵素種類與含量也會有差異。 Styrene 經 CYP- 450 代 謝 為 SO 後 , 在 生 物 體 內 經 由 glutathione. ( GST) 酵 素 或. S- transferases. epoxide. hydrolase ( EH) 酵 素 進 行 去 毒 性 作 用 , 再 隨 著 尿 液 排 出 體 外 。 SO 在 體 內 經 由 GST 酵 素 會 產 生 硫 醇 酸 ( mercapturic acid) ,經由 EH 酵 素 作 用 其 代 謝 產 物 為 苯 乙 烯 甘 油( styrene glycol) 或 苯 乙 醇 酸 (mandelic acid; MA) , 其 代 謝 簡 圖 見 附 圖 2.(Barale,1991)。. 第二節. 苯環氧乙烷. ( 一)苯環氧乙烷的致毒性與致癌性 SO 曾 被 報 導 會 造 成 生 殖 毒 性 , 例 如 會 導 致 大 鼠 preimplantation loss 頻 率 增 加,且 對 於 懷 孕 的 大 鼠 有 增 加 體 內 缺 鈣 的 現 象 , SO 也 會 造 成 早 期 懷 孕 的 動 物 胎 兒 的 毒 性 , 但是並沒有足夠證明會造成畸形兒(Liodbohm,1993)。 SO 經 由 動 物 實 驗 證 實 , 會 引 發 老 鼠 產 生 癌 症 , 包 括 餵 食 SO 會 造 成 老 鼠 的 胃 癌( forestomach tumours)(Lijinski, 10.
(15) 1986),而 皮 膚 接 觸 SO 會 造 成 皮 膚 癌。SO 在 1994 年 被 IARC (International Agency for Research on Cancer) 歸 類 為 2 A group 致 癌 物 ( 動 物 實 驗 已 被 證 實 會 造 成 癌 症 , 但 是 人 類 流 行 病學調查還沒有足夠證據證實會致癌)。. ( 二 ) DNA 共 價 鍵 結 物 ( DNA adduct) Styrene 代 謝 為 SO 後,SO 結 構 中 的 Oxirane ring 會 攻 擊 D N A, 形 成 DNA adduct, DNA adduct 可 被 D N A 修 補 酵 素 修 補 , 未 修 復 之 DNA adduct 在 DNA 複 製 時 可 能 產 生 錯 誤配對或造成複製程序被終止 ,進一步造成基因突變或細胞 死亡。 SO 會 攻 擊 D N A 許 多 位 置 , 於 體 外 試 驗 顯 示 在 guanine 中 N -7、N 2 與 O 6 等 位 置 都 會 產 生 DNA adduct, 其 發 生 比 例 依 次 為 72:23:3.7。SO 除 了 攻 擊 guanine 位 置 而 產 生 DNA adduct 外 , 也 會 在 adenine 的 N -1 及 N 6 、 cytosine 的 N 4 、 N-3 及 O 2 與 thymine 的 N- 3 等 位 置 產 生 DNA adduct (Bastlova et al.,1996)。 SO 主 要 藉 著 Oxirane ring 的 α 與 β 碳 與 DNA 結 合( 附 圖 1) ,DNA 與 α 碳 位 置 結 合 後 , 其 型 態 較 類 似 環 狀 芳 香 環 , DNA 與 β 碳 位 置 結 合 後 較 親 脂 性 。 而 α / β 碳 結 合 所 佔 的 比 例 會 受 到 親 核 性 物 質 不 同 而 影 響 , 研 究 顯 示 , 在 guanine 中 N-7 位置上,與 α /β 碳 位 置 結 合 的 比 例 為 47/53;N 2 位 置 上 只 會 與 α 碳 位 置 結 合 ; O6 位 置 上 與 α /β 碳 位 置 結 合 的 比 值 為 4.41(H emminki and Vodicka ,1995), 此 現 象 顯 示 , 在 guanine 不 同 的 位 置 上 , 與 Oxirane ring 之 結 合 位 置 也 有 所 差異。 在 所 有 產 生 DNA adduct 的 核 11. 酸 中 , 發 生 於 guanine.
(16) 中 N -7 上 的 Adduct 比 例 最 高,但 是 guanine 中 N- 7 並 不 參 加 DNA 複 製 時 的 鹼 基 配 對 , 但 是 其 可 能 因 為 自 發 性 或 修 補 酵 素 的 作 用 而 產 生 去 鹼 基 位 置( abasic site ) ,進 而 造 成 DNA 複 製 終 止 或 錯 誤 配 對 。 另 外 guanine 中 O 6 位 置 形 成 DNA adduct 的 機 率 雖 然 比 較 低 , 但 是 因 為 這 個 位 置 會 參 與 D N A 的 鹼 基 配 對,因 此 在 O 6 位 置 所 形 成 之 D N A adduct 對 基 因 突 變的影響可能佔有重要地位。 利 用 alkaline elution 技 術 , 測 量 白 血 球 中 DNA strand breaks 之 情 形,能 夠 有 效 的 偵 測 到 styrene 低 濃 度( 20ppm) 下 所 造 成 的 DNA 損 傷( Walles et al.,1993 ) ,一 項 調 查 研 究 顯 示 , 大 鼠 肝 臟 細 胞 細 胞 質 ( hepatocytes ) 暴 露 3mM 之 styrene 經 3 小 時 ,會 造 成 DNA strand breaks,而 SO 則 只 需 0.3 mM 就會造成 DNA strand break(Barale,1991)。. ( 三)苯環氧乙烷之基因毒性 在許多針對哺乳類細胞之體外試驗與動物體內 ( mouse ) 試 驗 顯 示 SO 確 實 會 造 成 染 色 體 異 常 之 現 象 (Barale,1991)。 SO 會 導 致 人 類 周 邊 血 液 淋 巴 球 細 胞 、 Chinese hamster 細 胞 中 染 色 體 傷 害( chromosome damage;C A)( Scott and Preston, 1994 )。 將 人 類 周 邊 血 液 之 淋 巴 球 細 胞 分 離 , 並 暴 露 在 100μ M 之 SO 中 , 經 22∼ 72 小 時 , 即 有 明 顯 SCE 增 加 之 情 形 ( C hakrabarti et al.,1997)。 以 SO 濃 度 200μ M 下 處 理 不 同 人 類 周 邊 血 液 之 淋 巴 球 細 胞 24 小 時 , 而 其 引 發 的 hprt 基 因 突 變 率 分 別 為 15.0 ×10 - 6 /cell)約 為 未 處 理 組 的 3.6 倍 (Bastlova et al .,1995)。以 SO 濃度( 200 ∼ 400 μ M) 下 處 理 人 類 周 邊 血 液 之 淋 巴 球 細 胞 6 天 , 而 其 引 發 的 hprt 12.
(17) 基 因 突 變 率 ( 10 ∼ 20 × 10 - 6 /cell ) 約 為 未 暴 露 組 的 4 倍 (Bastlova et al.,1995)。因 此 作 者 推 測 長 時 間 暴 露 在 styrene 之 環 境 下 之 作 業 勞 工 , 比 高 濃 度 短 時 間 的 暴 露 , 所 引 發 hprt 基因突變與 DNA adduct 會較嚴重(Bastlova et al.,1995)。. 調 查 研 究 顯 示 , 在 人 類 白 血 球 細 胞 中 加 入 styrene ( 2.6 mM )經 72 小 時 後 , 在 淋 巴 球 中 誘 發 染 色 體 異 常 之 現 象 , 而 在 相 同 實 驗 中,SO 的 劑 量 只 要 在 0.7 mM 即 可 看 到 染 色 體 異 常現象(Barale,1991)。 對 styrene 暴 露 的 勞 工 , hprt 基 因 突 變 與 DNA adduct 進 行 研 究 , 發 現 他 們 的 hprt 基 因 突 變 率 約 為 一 般 非 暴 露 者 的 1.5 ∼ 2 倍 , 而 且 在 3∼ 4 次 的 採 樣 中 , hprt 基 因 突 變 的 突 變 率高於非暴露者,但是只有一次達到統計上之顯著意義,並 且 同 時 發 現 DN A adduct 也 明 顯 增 加 , 但 是 在 4 次 之 採 樣 分 析 顯 示 , hprt 基 因 突 變 與 DNA adduct 並 沒 有 呈 現 良 好 的 關 係 , 顯 示 hprt 基 因 突 變 可 能 會 受 到 其 他 許 多 不 明 原 因 之 影 響,包 括 飲 食 、 生 活 習 慣 等 因 素 影 響( Lambert et al.,1995 )。. 第 三 節 苯環氧乙烷的光學異構物 ( 一)苯環氧乙烷光學異構物之致毒性 有 關 於 R - SO 與 S- SO 之 致 毒 性 的 文 獻 並 不 多 , 在 細 菌 與 動 物 實 驗 顯 示 , 不 同 物 種 對 不 同 SO 光 學 異 構 物 , 所 造 成 的 細 胞 毒 性 結 果 並 不 一 致 。 作 者 利 用. Salmonella. typhimurium TA100 經 不 同 SO 光 學 異 構 物 暴 露 後 , R - SO 處 理 的 細 胞 存 活 比 率 為 陰 性 對 照 組 的 76%, S- SO 為 2%, 13.
(18) 此 結 果 顯 示 : S- SO 對 細 菌 的 毒 性 比 R - SO 大 (Pagano et al.,1982), 而 Seiler 於 1990 年 以 相 同 的 細 胞 與 測 試 方 法 , 其所得到之結果也相同(Seile,1990)。 Gadberry 等 人 將 R - SO 與 S- SO 以 注 射 方 式 處 理 小 鼠 24 小時後,藉著測量小鼠肝臟與肺臟釋放之酵素的活性,作為 毒 性 大 小 之 指 標 , 結 果 顯 示 R - SO 處 理 的 老 鼠 , 肝 臟 所 釋 放 出 的 酵 素 活 性 比 S- SO 處 理 的 酵 素 活 性 高 , 雖 然 大 部 分 都 沒 有 達 到 統 計 上 之 顯 著 意 義 , 在 R - SO 與 (R+S)- SO 處 理 的 肝 臟 其 釋 放 出 來 SDH 的 酵 素 活 性 明 顯 高 於 S- SO 處 理 組 ( Gadberry et al.,1996) 。Watabe 也 認 為 在 S- SO 較 容 易 被 大 鼠 肝 臟 中 mEH 酵 素 分 解 , 所 以 R- SO 處 理 的 大 鼠 肝 細 胞 之 毒 性 可 能 高 於 S- SO(Watabe and Yoshikawa , 1981 )。 此 顯 示 R- SO 對 肝 臟 產 生 的 毒 性 大 於 S- SO, 而 這 些 老 鼠 所 測 試 的 結 果 恰 與 細 菌 之 毒 性 結 果 相 反 ( Pagano et al .,1982) 。. 1 .CYP-450 酵 素 之 影 響 將 phenobarbital( PB) 與 β- naphthoflavone (β -NF) 注 射 到 大 鼠 體 內 , 分 別 誘 發 CYP 2B 與 CYP1A 酵 素 , 以 PB 與 β-NF 注 射 之 老 鼠 其 肝 臟 微 粒 體 (microsome)與 styrene 反 應 後 , 所 產 生 的 R- SO/S- SO 分 別 為 0.92 與 1.25 , 而 沒 有 經 過 藥 物 注 射 的 老 鼠 , 其 R- SO/S- SO 為 0.65。 此 現 象 顯 示 CYP 2B 與 CYP1A 酵 素 雖 然 不 是 主 要 代 謝 SO 之 酵 素 , 但 是 在 生 物 體 內 之 CYP- 450 中 某 些 酵 素 活 性 增 加 時,會 造 成 不 同 SO 光 學 異 構 物 生 成 率 不 同 , 因 此 生 成 率 高 者 , 其 所 造 成 的 毒 性 也 可 能 較 高 (Foureman e t al.,1988),但 是 SO 光 學 異 構物的細胞毒性會受到其他去毒性酵素之影響。. 14.
(19) 2 .GST 酵 素 之 影 響 GST 酵 素 對 不 同 SO 光 學 異 構 物 之 結 合 力 有 2 種 不 同 看 法。H iratsuka 等 人 於 1989 將 老 鼠 肝 臟 細 胞 質 (cytosol) 與 SO 反 應 後 , 測 試 SO 光 學 異 構 物 與 GSH 的 鍵 結 物 , 結 果 顯 示 R- SO 與 GST 酵 素 的 結 合 力 約 為 S- SO 的 1.8 倍 , 顯 示 大 鼠 中 之 肝 臟 內 GST 酵 素 對 R - SO 代 謝 速 率 比 S- SO 快 (Hiratsuka et al.,1989) 。 老 鼠 體 內 測 試 顯 示 , 將 SO 光 學 異 構 物 注 射 到 老 鼠 體 內 , 發 現 有 28% 的 R- SO 會 與 GST 酵 素 作 用 , 產 生 R- SO 的 mercapteria acid 代 謝 產 物 , S- SO 則 約 為 19%(Linhart et al.,1998) , 而 在 Watabe 於 1982 年 也 有 類 似 之 報 告 (Watabe et al.,1982) , 這 些 結 果 都 顯 示 R - SO 與 GST 酵素可能有較高之結合力。 Foureman 於 1988 年 對 老 鼠 肝 臟 微 粒 體 ( microsome)進 行 體 外 試 驗, 其結果與上述 2 位 研 究 者 看 法 相 反 。他 利 用 [ 1 4C ]SO 處 理 肝 臟 微 粒 體,測 量 SO 與 肝 臟 中 GST 的 鍵 結 物, 發 現 R- SO/S- SO 的 比 例 為 0.65 , 此 結 果 顯 示 老 鼠 肝 臟 中 GST 酵 素 與 S- SO 的 結 合 力 較 高 。 將 老 鼠 注 射 β - NF, 會 降 低 體 內 CYP- 450 酵 素 活 性 , 此 時 將 肝 臟 取 出 , 萃 取 肝 臟 微 粒 體 , 但 是 當 微 粒 體 中 EH 酵 素 活 性 被 CHO (cyclohexene oxide)抑 制 後 , SO 不 同 光 學 異 構 物 大 部 分 經 由 GST 酵 素 進 行 去 毒 性 作 用 ,GST 酵 素 與 SO 不 同 光 學 異 構 物 作 用 , 發 現 R- SO/S- SO 的 比 例 為 1.25 (Foureman et al.,1988 )。 此 結 果 顯 示 與 SO 光 學 異 構 物 與 GST 酵 素 之 結 合 力 也 會 受 到 CYP-450 在生物體內活性的影響。 有學者萃取老鼠肝臟細胞之酵素,發現老 鼠 肝 臟 細 胞 GST 3 - 3,3- 4,4- 4( 被 分 類 為 mu 酵素)對 R - SO 的 結 合 力 較 強 , 其 中 尤 其 以 GST3- 3 對 R- SO 具 有 高 度 之 結 合 力 , 而 15.
(20) G S T 1-1, 1- 2, 2- 2( 被 分 類 為 α 酵 素 ) 與 S- SO 結 合 力 較 強 。 而 以 化 學 物 質 誘 發 老 鼠 產 生 肝 癌 中 的 主 要 GST 酵 素 (GST 7-7) , 發 現 其 與 S- SO 結 合 力 較 強 。 此 現 象 更 明 顯 顯 示,不同的 GST 酵 素 對 SO 光 學 異 構 物 之 結 合 力 不 同 , 而 在 人 體 肝 臟 中 酵 素 與 SO 之 結 合 力 大 小 依 次 為 GSTµ > GST π >GST α~ε (Hiratsuka et al.,1989)。. 3 .Epoxide hydrolase( EH) 酵 素 之 影 響 在細菌方面之研究顯示,利用從細菌中純化出 recombinant 所 產 生 之 EH 水 解 酵 素 來 水 解 SO,發 現 EH 酵 素 會 將 R - SO 先 水 解 , 而 R - SO 水 解 約 90%時 , S- SO 才 開 始 快 速 與 EH 酵 素 進 行 作 用 , 因 此 認 為 EH 酵 素 對 R- SO 結 合力較高(Spelberg et al.,1998)。 SO 除 了 經 由 GST 進 行 去 毒 性 作 用 外 , 還 會 經 由 EH 進 行 水 解 , 學 者 在 EH 酵 素 對 SO 光 學 異 構 物 之 結 合 力 也 有 不 同 的 看 法 。 有 學 者 將 (R+S)- SO 注 射 到 老 鼠 體 內 經 24 小 時 , 並 同 時 給 予 TPCO(3,3,3- tr ichloropropene oxide) 阻 斷 EH 代謝路徑,測量小鼠肝臟與肺臟釋放之酵素活性,因為肝細 胞 與 肺 細 胞 被 破 壞 , 會 分 別 大 量 釋 放 出 SDH (serum sorbitol dehydrogenase) 與 LDH (lactate dehydrogenase) 、 GGT (gamma - glutamyl. transpeptidase) 酵 素 。 先 以. BSO. (buthionine sulfoxamine) 抑 制 GST 酵 素 之 活 性 後 , 再 注 射 (R+S) - SO, 肝 臟 與 肺 臟 中 這 些 酵 素 的 量 卻 沒 有 明 顯 增 加 , 此 結 果 顯 示 阻 斷 GST 酵 素 之 活 性 並 不 會 對 肝 與 肺 細 胞 造 成 傷 害 。 因 此 作 者 認 為 EH 代 謝 路 徑 為 SO 之 主 要 代 謝 途 徑 (Watabe et al.,1982,Gadberry et al.,1996),此 結 果 與 細 菌 方面之結果相反。 16.
(21) 有 學 者 也 發 現 小 鼠 肝 臟 細 胞 中 EH 之 酵 素 活 性 就 比 中 國 倉 鼠 高 ( Norppa,1979)。在 1989 年 Drummond 等 人 與 1987 年 Korn 等 人 偵 測 人 體 中 styrene 經 由 EH 代 謝 其 mandelic acid 尿 中 代 謝 物 , 推 估 R - SO 與 S- SO 的 比 例 在 0.5 ~2.2 之 間 ( D rummond et al.,1989 , Korn et al.,1987)。 因 此 不 同 物 種 中 去 毒 酵 素 的 含 量 可 能 不 同 , 對 SO 光 學 異 構 物 之 結 合 力 也 可 能 不 同,因 此 會 影 響 SO 光 學 異 構 物 對 細 胞 毒 性 反 應。. ( 二)苯環氧乙烷光學異構物之致突變性 1982 年 Pagano 等 人 利 用 Salmonella typhimurium TA100 進 行 Ames test 顯 示 , R - SO 致 突 變 率 高 於 S- SO, 而(R+S)-SO 之突變性介於兩者中間(Pagano et al.,1982)。. 老 鼠 骨 髓 細 胞 中 觀 察 C A 與 SCE 之 情 形 顯 示 , S- SO 的 致 突 變 性 大 於 R - SO (Sinsheimer et al.,1993) 。 在 細 胞 株 (C hinese hamster V79)方 面 之 研 究 顯 示 : S- SO 造 成 SCE 的頻率高於 R-SO (von der hude et al.,1992)。. 而影響光學異構物致突變之因子除了不同光學異構物與 去 毒 性 酵 素 之 反 應 外 , SO 光 學 異 構 物 與 DNA 形 成 D N A adduct 也 是 一 個 重 要 因 子 。 Seiler 在 1990 年 的 報 告 中 指 出 , 不 同 光 學 異 構 物 所 造 成 的 DNA adduct 有 所 不 同 , 因 此 認 為 不 同 型 態 的 DNA adduct 在 經 過 D N A 修 復 與 複 製 過 程 中 , 所 造 成 基 因 突 變 的 頻 率 也 會 不 同 ( Sinsheimer et al.,1993)。 Latham 於 1993 年 在 體 外 以 human N -ras 61 codon 中 Adenine 結 構 上 的 N 6 位 置 合 成 R- SO 與 S- SO 的 DNA 17.
(22) adduct,將 此 具 有 DNA adduct 的 gene 植 入 細 菌 內,經 DNA 複 製 後 測 試 點 突 變 ( 附 圖 3), 結 果 發 現 只 有 在 S ( 6 1. , 2). 位置上. 之 DNA adduct 會 引 發 約 1.6 % 的 A→ G 的 突 變,而 且 認 為 除 了 DNA adduct 形 成 的 位 置 會 影 響 突 變 率,另 外 在 adduct 附 近 之 序 列 也 會 影 響 突 變 率,影 響 突 變 率 的 因 子 包 括 形 成 D N A adduct 的能力、D N A 的 修 復 能 力 與 DNA 聚 合. 錯誤判斷所. 造成的錯誤配對能力(Latham et al.,1993)。. 第 四 節 hprt 與 tk 基 因 1 .hprt 基 因 hprt 基 因 位 於 X 染 色 體 上,具 有 9 個 exon 與 8 個 intron, 約 有 4 0 0 0 0 base pair , 因 此 只 要 在 hprt 基 因 上 發 生 單 一 突 變 就 可 以 被 選 擇 性 試 劑( 6- Thioguanine )偵 測 出 來,而 因 為 hprt 基 因 不 是 細 胞 生 長 的 必 需 基 因 ( essential gene ), 為 偵 測 突 變 之 良 好 標 的 基 因 。 hprt 基 因 已 經 被 定 序 , 因 此 再 配 合 基因序列分析 就可以明確檢測出點突變或基因缺失的種類與 位 置 (Casciano et al.,1999) , 經 由 DNA adduct 分 析 可 以 推 測出造成基因突變的可能的來源。. 2 .tk 基 因 tk 基 因 因 為 突 變 位 置 不 同 , 導 致 生 長 速 度 的 表 現 也 不 同,第 一 種 為 tk 基 因 突 變 後,細 胞 生 長 比 較 快 之 型 態 ( tk-fast growing mutants ; tk-fast) , 其 所 測 得 之 突 變 率 為 點 突 變 或 小 段 基 因 缺 失 或 嵌 入 ; 另 一 種 tk 基 因 突 變 會 使 tk 基 因 的 兩 旁 (flanking) 造 成 缺 失 , 使 得 細 胞 長 的 比 較 慢 ( tk -slow growing mutants ; tk- slow), 主 要 可 以 測 得 DNA 大 片 段 的 18.
(23) 缺 失 ( deletion)。 tk 基 因 的 突 變 很 容 易 被 Trifluorothymine deoxyriboside ( TFT ) 選 擇 性 試 劑 所 篩 選 。 tk 基 因 能 夠 測 量非常廣泛的突變,包括點突變與嚴重的染色體改變或對偶 基 因 的 缺 失 ( allele loss ), 可 以 說 是 一 個 良 好 的 基 因 指 標 (Honma et al.,1997)。. 綜合前人之研究結果顯示,SO 光學異構物在不同生物 種類中,其扮演的致毒性與致突變的角色可能會有所差異, 而 且 不 同 研 究 對 不 同 SO 光 學 異 構 物 所 造 成 的 細 胞 毒 性 與 致 突變性看法並不一致,而目前仍沒有以人類細胞來探討 R- SO 或 S- SO 誘 發 細 胞 毒 性 與 突 變 率 的 相 關 研 究 報 告 。 我 們 首 次 以 人 類 淋 巴 球 細 胞 對 SO 與 SO 光 學 異 構 物 所 造 成 之 細 胞 毒 性 與 致 突 變 性 進 行 探 討 , 希 望 能 進 一 步 瞭 解 SO 與 其 光學異構物對人類淋巴球細胞之相對細胞毒性與致突變性。. 19.
(24) 第 四 章 材料與方法 第 一 節 材料. ( 一)細胞來源 本 實 驗 使 用 人 類 淋 巴 母 細 胞 株 (human lymphoblastoid cell line ; TK6 ) , 此 細 胞 株 購 自 新 竹 食 品 工 業 發 展 研 究 所 菌 種保存中心(CCRC 60107 )。 ( 二)藥品 styrene oxide、R- phenyloxirane (R-styrene Oxdie) 與 S- phenyloxirane (S-styrene Oxide ) 購 自 Fluka 公 司 。 配 置 RPMI 1640 medium 粉 末 ( 美 國 Gibco BRL),在 RPMI 1640 medium 中 加 入 Sodium Bicarbonate (NaHCO3)( 美 國 Sigma) 配 置 為 培 養 液,並 以 1N HC l 調 整 pH 值 為 7.2 ∼ 7.4, 分 裝 為 450 ml 置 於 4 ℃ 備 用 , 使 用 前 加 入 penicillin , streptomycin (P/S) , 200mM L-Glutamine , ( 美 國 Gibco BR ) 、 9% Donor. horse (HS) ( 英 國 Selborne Biological. Services ) 與 1 % H EPES buff er ( 義 大 利. Biological. industries)。 降 低 突 變 背 景 值 所 需 之 HAT supplement ( 40μ M. aminopterin , 1.6. mM. thymidine , 10mM. sodium. hypoxanthine )、HT supplement ( 1.6 mM thymidine,10mM sodium. hy poxanthine ) ( 美 國. Gibco. BRL ). 與. 2-Deoxycytidine( 美 國 Sigma ) 。 篩 選 h p r t 基 因 突 變 之 選 擇 性 試 劑 為. 2- Amino -6- mercaptopurine. ( 6- TG ), 篩 選. tk. 6- thioguanine. 基 因 突 變 之 選 擇 性 試 劑 為. Trifluorothymine deoxyriboside (TFT) (美國 Sigma)。 20.
(25) 細 胞 計 數 所 需 之 染 劑 為 0.4% Trypan Blue Stain( 美 國 Gibco BRL )。 冷 凍 細 胞 所 需 之 sterile dimethyl-sulfoxide (DMSO) ( 美 國 Sigma) 。 細 胞 培 養 瓶 與 所 有 15ml、 50ml 無 菌 試 管 , 與 過 濾 RPMI 1640 medium 所 需 之 500 ml 無 菌 濾 杯 均 購 自 美 國 Corning 公 司 。 96 孔 平 底 培 養 盤 購 自 美 國 Costar 公司。. 第 二 節 方法 ( 一) 細胞培養 TK6 細 胞 株 利 用 含 有 9% H S,1% 的 L- Glutamine,1 % penicillin /streptomycin 及 1 % HEPES buffer 之 RPMI 1640 細 胞 培 養 液 培 養 , 將 細 胞 株 置 於 含 有 5% CO 2 的 37 ℃ 恆 溫 培 養 箱 內 培 養 , 將 細 胞 培 養 在 1.5 ∼ 2.2 X10 5 cell/ml,每 天 更 換 細 胞 培 養 液,將 細 胞 維 持 在 對 數 生 長 期( log phase)。. ( 二) 細胞株解凍與保存. 1 .細 胞 解 凍 將 以 液 態 氮 保 存 之 細 胞 凍 乾 管 ( vial) 在 37℃ 水 浴 槽 中 回 溫 約 一 分 鐘 , 使 它 溶 解 後 , 將 細 胞 放 入 15ml 的 無 菌 試 管 中 , 緩 慢 加 入 細 胞 培 養 液 , 充 分 混 合 均 勻 後 以 200g 離 心 4 分 鐘 ( 1100 rpm), 去 除 上 層 液 , 再 加 入 細 胞 培 養 液 , 並 使 用 pipet 將 離 心 後 所 剩 之 pellet 進 行 混 合 , 混 合 後 取 出 放 入 25cm2 培養瓶中。. 21.
(26) 2 .細 胞 保 存 log phase 的 細 胞 以 200g 離 心 4 分 鐘 離 , 去 除 上 層 液 後,加 入 含 有 1 2 %之 無 菌 DMSO 之 細 胞 培 養 液,使用 pipet 進 行 混 合 , 取 1ml 放 入 凍 乾 管 中 , 迅 速 放 到 - 80℃ 冰 箱 中 進 行降溫,24 小時後再移至液態氮桶中保存。. ( 三)細胞計數 利用血球計數器計數之,其蓋玻片重量將樣品厚度壓 為:0.1mm , 總 樣 品 體 積 為( V)=1mm. ×1mm. ×0.1mm. = 0.1(mm) 3 = 0.1(10 -1cm) 3 =10 -4ml 所 計 數 之 細 胞 為 10 4 /ml。 依 不 同 細 胞 濃 度 調 整 所 計 數 之 格 數 , 使 每 次 計 數 之 細 胞 均 在 100 ∼ 300 cell, 以 確 保 細 胞 計 數 之 精 確 性 。 取 培 養 瓶 中 之 細 胞 液 100μ l 放 到 well 中 , 加 入 10 μ l 的 0.4% trypan blue stain 染 色 ( 美 國 Gibco BRL), 以 100X 顯 微 鏡 ( OLYMPUS, IX- 70 ) 觀 察 , 死 細 胞 之顏色為藍色,計數活細胞數。. ( 四)細胞毒性測試. 1 .未 處 理 組 SO 都 必 須 利 用 DMSO 配 置 成 不 同 的 濃 度 而 才 能 進 一 步 處 理 細 胞,因 此 未 處 理 組 之 細 胞 及 加 入 等 量 的 DMSO 試 劑 為 溶劑控制組(control)。 2 .相 對 細 胞 數 目 (relative cell number; RCN) 去 除 SO 的 細 胞 培 養 液 後 , 每 天 計 算 其 細 胞 數 , 並計 算 22.
(27) 其 Td 值,若 Td 回 到 正 常 值 (14~18), 則 處 理 組 累 計 細 胞 生 長 的 倍 數 /控 制 組 累 計 細 胞 生 長 的 倍 數,即 可 以 得 到 細 胞 相 對 毒性 RS% 。. 3 .細 胞 群 落 生 成 率 (plating efficiency; PE) 將 細 胞 經 CHAT 處 理 後 , 分 別 處 理 SO、 R- SO 與 S- SO 並 將 蓋 子 旋 緊 , 培 養 在 37℃ 的 培 養 箱 中 , 經 24 小 時 後 , 離 心 (200g, 4mins) 去 除 R - SO 與 S- SO, 並 以 10ml 之 RPMI medium 1640 進 行 清 洗 , 再 離 心 (200g, 4min) , 去 除 上 層 液,加 入 細 胞 培 養 液,將 細 胞 轉 置 在 75c m2 的 培 養 瓶 中 培 養。 去 除 styrene oxide 的 細 胞 培 養 液 後,利 用 細 胞 培 養 液 進 行 連 續 稀 釋,將 細 胞 接 種 到 96 well 的 平 板 中,使 每 個 well 中 的 細 胞 數 控 制 在 2 或 4 cell ( 依 低 劑 量 暴 露 到 高 劑 量 ), 經 12- 15 天 觀 察 細 胞 生 長 結 果 , 可 以 計 算 出 細 胞 生 成 率 , 將 處 理 組 之 細 胞 生 成 率 / 控 制 組 細 胞 生 成 率,即 可 得 到 細 胞 相 對 毒 性(relative survival;RS). ( 五 ) hprt 與 tk 基 因 突 變 率 測 試 (Liber and Thilly , 1982,Skopek,1993) 取 Td 值 正 常 的 TK6 細 胞,加 入 CHAT(Deo xycytidine 、 HAT supplement 與 HT supplement) 之 培 養 基 後 , 去 除 上 層 液 ( CHAT. 之 成 分 ) , 加 入 含 有. HT. supplement. 與. Deoxycytidine (CHT) 之 細 胞 培 養 液 , 經 24 小 時 培 養 後 (37 ℃ , 5% CO 2 ) 加 入 細 胞 培 養 液 , 以 血 球 計 數 器 計 算 細 胞 每 天 增加之情況,以測得 Td 值。 以 細 胞 培 養 液 處 理 2∼ 3 天 內 , 細 胞 之 td 值 必 須 在 正 常 23.
(28) 範圍內(表 示 細 胞 回 到 正 常 的 對 數 生 長 期), 在 此 時 進 行 細 胞 生 成 率 測 試 與 突 變 率 測 試 , 即 可 得 知 細 胞 中 背 景 突 變 率 。細 胞 經 SO 處 理 24 小 時 , 以 離 心 方 法 去 除 SO 後 , 經 3~4 天 後 , 將 細 胞 轉 植 到 96 well plate ( 2cell/well/150μ l 細 胞 培 養 液 )中,置 於 5% CO 2,37℃ 恆 溫 培 養 相 中 培 養。經 12-15 天 培 養 , 計 算 細 胞 生 長 結 果 , 可 以 得 到 未 添 加 TFT 選 擇 性 試 劑 之. PE 。 另 外 將 細 胞 轉 植 到. 96. well. plate. ( 20000cell/well/150 μ l 細 胞 培 養 液 ) 中 , 此 細 胞 培 養 液 中 含 有 1μ g/ ml. Trifluorthymine deoxy- riboside (TFT) 之 選. 擇 性 試 劑 , 可 以 有 效 篩 選 tk-fast 基 因 突 變 率 , 將 培 養 盤 置 於 5% CO 2 , 37℃ 恆 溫 培 養 相 中 培 養 。 經 12- 15 天 培 養 , 計 算 細 胞 生 長 結 果 , 即 可 得 到 tk -fast 基 因 突 變 率 的 細 胞 突 變 率 , 經 10 天 後 , 再 加 入 1μ g/ ml TFT 之 選 擇 性 試 劑 , 即 可得到 tk slow 基因突變率(附圖 4)。 利 用 上 述 之 方 法 , 細 胞 以 SO 處理後,約 6∼ 7 天後,再 加 入 1μ g/ ml 6- TG 之 選 擇 性 試 劑 , 即 可 得 到 TK6 細 胞 中 hprt 基因之突變率. ( 六)統計分析方法 利用無母數分析(Wilcoxon signed rank test)方法分 析。. 24.
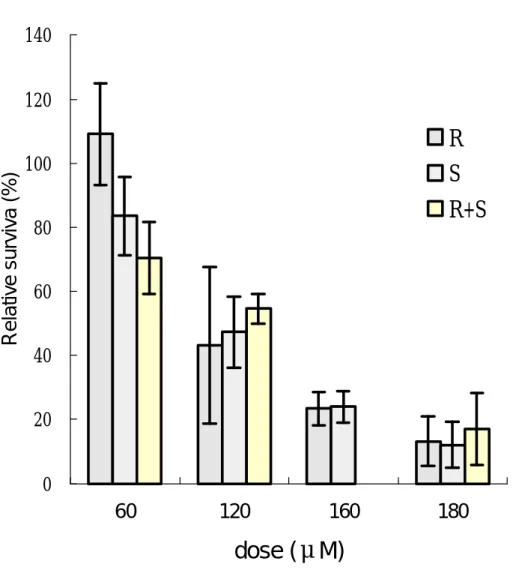
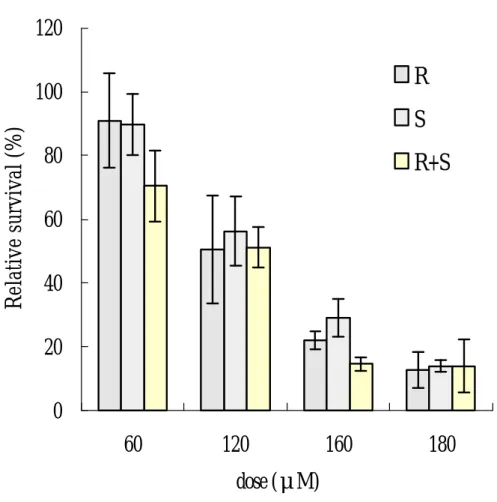
(29) 第五章 結. 果. 第 一 節 SO 與 SO 光 學 異 構 物 在 TK6 細 胞 之 致 細 胞 毒 性 ( 一 ) 細 胞 生 長 恢 復 情 形 (growth delay) 以 不 同 劑 量( 60、120、160 與 180μ M)的 R- SO、S- SO 與 (R+S)- SO 處 理 TK6 細 胞 經 24 小 時 後 , 180 μ M 濃 度 之 R- SO、 S- SO 與(R+S) - SO 處 理 之 細 胞 顯 示 , 細 胞 生 長 恢 復 情 形 明 顯 比 control 組 慢 , 在 暴 露 72 小 時 後 , R- SO 處 理 的 細 胞 生 長 倍 數 為 原 來 的 3.2 倍 , S- SO 為 原 來 的 2.6 倍 , (R+S) - SO 為 原 來 的 3.0 倍 ( 圖 1), 由 此 顯 示 SO 與 SO 學 異構物對 TK6 細胞所產生之細胞相對毒性沒有差異。. 以 不 同 劑 量( 60、120、160 與 180μ M)的 R- SO、S- SO 與 (R+S)- SO 處 理 TK6 細 胞 經 24 小 時 , 每 日 計 算 細 胞 生 長 倍 數,在 細 胞 培 養 48 小 時 後,control 組 細 胞 增 加 為 原 來 7.36 倍 , 依 暴 露 劑 量 增 加, 細 胞 有 生 長 遲 緩 之 現 象 , 顯 示 有 劑 量 反應關係(圖 2)。. SO 與 其 光 學 異 構 物 對 TK6 細 胞 之 毒 性 顯 示 , R - SO、 S- SO 與(R+S) - SO 三 者 間 對 TK6 細 胞 之 毒 性 沒 有 達 到 統 計 上之顯著差異。. ( 二 ) 細 胞 群 落 形 成 能 力 (PE) 利 用 細 胞 群 落 形 成 能 力 (PE) 試 驗 顯 示,隨 著 R - SO、S- SO 與 (R+S)- SO 劑 量 的 增 加 , 所 引 發 的 細 胞 毒 性 也 隨 著 增 加 , 25.
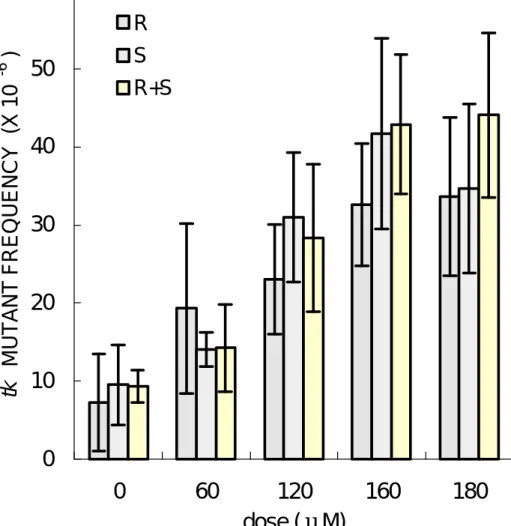
(30) 而 且 呈 現 劑 量 反 應 關 係 。以 120 μ M 濃 度 之 R - SO、S- SO 與 (R+S) - SO 處 理 TK6 細 胞 , 其 細 胞 相 對 存 活 率 依 次 為 50.55、 56.25 與 51.24% ( 圖 4)。 由 以 上 結 果 顯 示 , 三 者 間 對 TK6 細 胞 的 毒 性 結 果 並 無 差 異,此 結 果 與 細 胞 生 長 恢 復 情 形 之 結 果相似。. 第 二 節 SO 與 SO 光 學 異 構 物 在 TK6 細 胞 之 致 突 變 性 ( 一 ) hprt 基 因 突 變 率 Pagano 與 Seiler 認 為 R - SO 對 細 菌 細 胞 致 突 變 率 明 顯 比 S- SO 高 , 此 顯 示 SO 光 學 異 構 物 對 細 菌 細 胞 內 之 致 突 變 機制,顯然與老鼠細胞和人類淋巴球細胞不同。 TK6 細 胞 之 hprt 基 因 背 景 突 變 率 為 1.0 ∼ 18.8 ×10 - 6 , 隨 SO 暴 露 劑 量 ( 60、120 與 180 μ M),hprt 基 因 的 突 變 率 有 增 加 之 趨 勢 ( 圖 4)。 在 120 μ M 濃 度 下 R - SO、 S- SO 與 (R+S) - SO 所 引 發 hprt 基 因 的 突 變 率 分 別 為 25.1 ×10 -6 、 24.1 ×10 - 6 與 35.0 ×10 - 6 , 在 180μ M 濃 度 下 引 發 hprt 基 因 突 變 率 分 別 為 29.8 ×10 - 6 、 50.4 ×10 - 6 與 45.0 ×10 - 6 。 R- SO 與 S- SO 對 TK6 細 胞 之 突 變 機 率 有 劑 量 反 應 關 係 , 而 且 S- SO 之 整 體 突 變 機 率 有 高 於 R - SO 之 趨 勢 , 並 且 達 到 統 計 上 之 顯 著 不 同( p<0.05) 。此 結 果 顯 示 S- SO 可 能 引 發 TK6 細胞中 hprt 基因突變率略高 R-SO。. ( 二 ) tk-fast 基 因 突 變 率 tk -fast 基 因 突 變 率 結 果 顯 示 , TK6 細 胞 的 tk-fast 基 因 之 背 景 突 變 率 為 0.6 ∼ 18.9 ×10 - 6 。 隨 著 SO 劑 量 增 加 , 26.
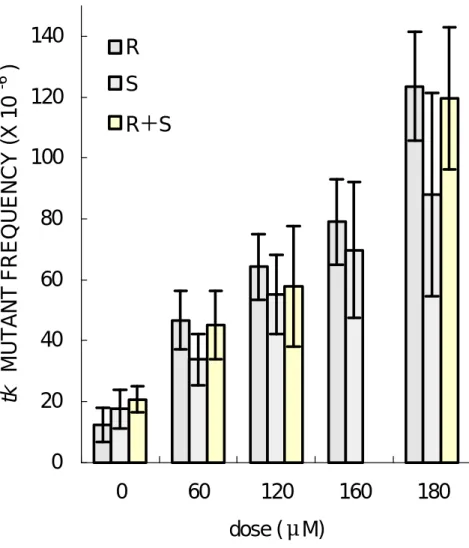
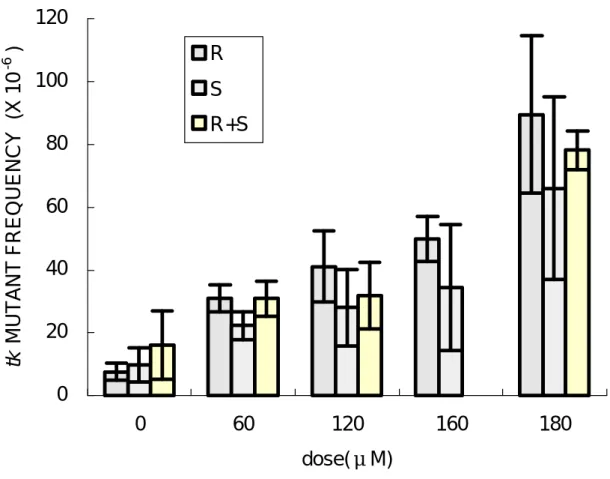
(31) tk -fast 基 因 突 變 機 率 也 增 加 ( 圖 5), 但 是 彼 此 間 沒 有 達 到 統 計 上 之 顯 著 差 異( p>0.05) ,此表示 R - SO 與 S- SO 在 TK6 細胞所誘發之 tk-fast 基因突變率並無差異。. ( 三 ) tk-slow 基 因 突 變 率 在 TK6 細 胞 中 tk- slow 基 因 之 背 景 突 變 率 在 2.3~23.7 ×10 - 6 之間, tk-s l o w 之 基 因 突 變 率 隨 著 SO 劑 量 增 加 呈 現 良 好 劑 量 反 應 關 係 。在 120 μ M 劑 量 作 為 比 較 ,R- SO 引 發 tkslow 基因之突變率為 41.1 × 10 -6,S-SO 為 28.0 × 10 -6 , (R+S) - SO 為 31.7 ×10 - 6 , 而 180 μ M 劑 量 下 R - SO 之 突 變 率 為 89.5 ×10 - 6 , S- SO 為 66.0 ×10 - 6 ( 圖 6), (R+S) - SO 為 78.2 ×10 - 6 ( 圖 7)。 此 突 變 率 之 結 果 明 顯 比 tk -fast 基 因 突 變 率 高 出 許 多 , 而 且 R - SO 對 TK6 細 胞 中 tk-slow 基因致突變率高於 S-SO。. ( 四 ) tk-total 基 因 突 變 率 在 TK6 細 胞 中 tk - toatl 基 因 之 背 景 突 變 率 在 12.3 ×10 - 6 ∼ 20.1 ×10 -6 之 間 , tk- total 之 基 因 突 變 率 隨 著 SO 劑 量 增 加 呈 現 良 好 劑 量 反 應 關 係 。 以 120 μ M 的 劑 量 作 為 比 較 , R- SO 引 發 tk- total 基 因 之 突 變 率 為 64.1 ×10 - 6 , S- SO 為 55.2 ×10 - 6 ,(R+S) - SO 為 57.8 ×10 - 6 , 而 180μ M 劑 量 下 R- SO 之 突 變 率 為 123.6 ×10 - 6 , S- SO 為 87.8 ×10 - 6 ( 圖 6),(R+S) - SO 為 119.5 ×10 - 6 。 在 三 次 獨 立 實 驗 中 的 結 果 顯 示 , 在 60、120 與 180μ M 三 種 劑 量 測 試 中 R - SO 之 突 變 率都比 S-SO 高,但是沒有達到統計上之顯著差異。 27.
(32) 第六章 討. 論. 第 一 節 SO 與 SO 光 學 異 構 物 對 TK6 細 胞 之 毒 性 在 TK6 細 胞 分 別 暴 露 SO 與 其 光 學 異 構 物 24 小 時 後 , 以 顯 微 鏡 觀 察 細 胞 型 態 , 以 計 數 細 胞 存 活 率 時 , 發 現 R - SO 比 S- SO 處 理 的 細 胞 完 整 性 差 , 而 且 細 胞 數 明 顯 減 少 , 死 細 胞 數 明 顯 較 多 。以 細 胞 生 長 遲 緩 情 況 與 細 胞 群 落 形 成 能 力 探 討 細 胞 相 對 毒 性 , 統 計 方 法 分 析 結 果 顯 示 , R - SO 致 細 胞 毒 性 沒 有 比 S- SO 高,細 胞 相 對 毒 性 無 法 分 析 出 R - SO 比 S- SO 致 細 胞 毒 性 高 之 原 因 可 能 是 在 計 數 細 胞 時 , 因 R- SO 處 理 的 細胞完整較差,因此遺漏許多細胞,是否造成低估細胞數之 情形,則有待進一步確認。. 在 細 菌 方 面 對 認 為 S- SO 的 細 胞 毒 性 大 於 R - SO,有 學 者 認 為 是 R- SO 較 容 易 被 細 菌 中 之 EH 酵 素 所 水 解 (Pagano et al.,1982)所 造 成 。 相 反 的 在 老 鼠 肝 臟 測 試 發 現 R - SO 對 肝 臟 細 胞 之 毒 性 大 於 S- SO, 而 且 認 為 去 毒 性 酵 素 中 以 SO 主 要 依 賴 EH 酵 素 進 行 代 謝 。 另 外 有 報 告 指 出 , 在 體 外 老 鼠 肝 臟 細 胞 測 試 與 檢 測 老 鼠 尿 液 中 SO 的 代 謝 物 mercapturic acid 都顯示,R- SO 與 GST 酵 素 之 結 合 力 比 S- SO 強( H iratsuka et al.,1989,Watabe et al.,1982)。 而 Watabe 的 研 究 指 出 R- SO 大 部 分 經 由 GST 酵 素 進 行 去 毒 性 作 用 , S- SO 則 經 由 EH 酵 素 進 行 代 謝。由 此 可 見 R - SO 與 S- SO 對 細 胞 的 毒 性 看 法 並 不 一 致 。 本 實 驗 雖 然 在 統 計 分 析 中 , 無 法 比 較 出 SO 不 同光學異構物對細胞之毒性大小,但是細胞在暴露後,以顯 微 鏡 觀 察 之 結 果 , R - SO 的 細 胞 毒 性 有 大 於 S- SO 之 趨 勢 。 28.
(33) 研 究 顯 示 TK6 細 胞 中 之 GST 酵 素 活 性 為 AHH -1 細 胞( 人 類 淋 巴 母 細 胞 ) 的 1/5(Cres pi et al.,1985),而 且 有 報 告 指 出 , 中 國 倉 鼠 V79 細 胞 株 中 之 EH 酵 素 活 性 比 小 鼠 高 ( Scott et al.,1994 )。由 此 顯 示 在 不 同 物 種 中 細 胞 內 所 含 之 去 毒 性 酵 素 的量與酵素活性並不相同,因此可能造成不同物種間細胞毒 性 的 差 異 。 由 於 去 毒 性 酵 素 一 旦 與 SO 結 合 之 後 , SO 即 失 去 對 細 胞 的 毒 性 與 致 突 變 性 , 因 此 TK6 細 胞 中 EH 與 GST 兩 個 去 毒 性 酵 素 的 含 量 與 不 同 SO 光 學 異 構 物 之 作 用 , 值 得 進一步探討。. 研 究 顯 示 以 200μ M 之 SO, 處 理 V79 細 胞 株 3 小 時 , 其 細 胞 存 活 率 接 近 100% , 顯 示 在 此 劑 量 下 , SO 對 V79 細 胞 株 並 沒 有 造 成 毒 性 ( N ish et al.,1984 )。 而 以 200 μ M 的 SO 處 理 人 類 周 邊 淋 巴 球 細 胞 24 小 時 , 其 細 胞 存 活 率 約 為 40%( Bastlova et al.,1995) 。本 實 驗 以 180μ M 的 (R+S)- SO 處 理 TK6 細 胞 經 24 小 時 , 細 胞 存 活 率 為 13%。由 以 上 之 結 果 顯 示 ,TK6 細 胞 對 SO 造 成 之 細 胞 毒 性 比 V79 細 胞 與 人 類 周邊淋巴球細胞靈敏度高。. 第 二 節 SO 與 SO 不 同 光 學 異 構 物 之 致 突 變 性. ( 一 ) hprt 基 因 突 變 率 Sinsheimer 於 1993 年 對 老 鼠 骨 髓 細 胞 之 SCE 頻 率 與 CA 測 試,結 果 顯 示 S- SO 的 突 變 率 大 於 R - SO( Sinsheimer et al.,1993) ,而 本 實 驗 顯 示 S- SO 所 誘 發 TK6 細 胞 的 h p r t 基 因 突 變 率 也 有 高 於 R- SO 趨 勢 , 並 且 達 到 統 計 上 之 顯 著 差 異 29.
(34) ( p=0.038 ), 此 結 果 表 示 S- SO 可 能 比 R- SO 更 容 易 引 起 point mutation、frameshift 及 small deletion。 Nish 指 出 , SO 為 一 種 弱 致 突 變 物 質 ( N ish et al.,1984 ) , 但 是 在 環 氧 化 物 中 EB( 1.2- epoxybutene )以 125 μ M 處 理 對 TK6 細 胞 24 小 時 , 誘 發 hprt 基 因 突 變 率 為 2.5 ×10 - 6 , 此 結 果 顯 示 EB 對 TK6 細 胞 之 致 突 變 性 明 顯 比 SO 小。文 獻 指 出 100 μ M 處 理 V79 細 胞 3 小時, hprt 基 因 突 變 率 增 加 為 原 來 的 <1 × 10 -6 ( Nish et al.,1984 )。 Bastlova 於 1995 年 之 結 果 顯 示 ,SO 在 100μ M 劑 量 下 , 誘 發 人 類 周 邊 血 液 淋 巴 球 細 胞 之 突 變 率 為 11.3 ×10 - 6 , 約 為 control 的 1.5 倍。而 且 在 本 實 驗 中,在 2 次 獨 立 實 驗 背 景 突 變 率 (1~3 × 10 -6 ) 較 低 之 情 況 下 , R- SO 與 S- SO 以 120 μ M 誘 發 hprt 基 因 突 變 率 ( 19∼ 27 ×10 - 6 ) 約 為 c o ntrol 的 8∼ 10 倍 左 右 。 以 上 結 果 顯 示 SO 對 TK6 細 胞 所 造 成 致 突 變 率 比 V79 細 胞 與 人 類 周 邊 血 液 淋 巴 球 細 胞 高 出 很 多 。 因 此 SO 是 否 為 弱 致 突 變物質仍有待進一步證實。. 曾 有 文 獻 指 出 , TK6 細 胞 中 h p r t 基 因 背 景 突 變 率 約 在 2.5 ×10 - 6(Cochrane and Skopek,1994),正 常 人 之 背 景 突 變 率 在 1.4 ∼ 22.6 ×10 -6 之 間 ( H ou et al.,1995)。 本 實 驗 TK6 細 胞 之 背 景 突 變 率 範 圍 在 1.0 ∼ 16.5 ×10 - 6 , 背 景 突 變 率 一 直 沒 有 很 穩 定 之 原 因,可 能 是 在 處 理 CHAT 降 低 背 景 突 變 率 時 ,CHAT 添 加 三 天 後 , 需 加 入 CHT 處 理 一 天 , 此 時 細 胞 應 再 度 回 到 正 常 代 謝 途 徑 ( de novo pathway 與 savage pathway) , 但 是 由 於 操 作 之 關 係 , 延 遲 了 CHT 加 入 之 時 間 , 導致細胞背景突變率升高之原因。. 30.
(35) ( 二 ) tk 基 因 突 變 率 hprt 基 因 為 在 X 染 色 體 上 可 以 偵 測 點 突 變 及 小 段 之 基 因 缺 失 , 而 由 於 tk 基 因 位 於 體 染 色 體 上 , 而 且 可 以 偵 測 到 大 段 基 因 缺 失 , 因 此 本 研 究 首 次 探 討 SO 與 其 光 學 異 構 物 對 TK6 細 胞 中 tk 基 因 所 誘 發 之 突 變 率 。 結 果 顯 示 隨 著 SO 與 SO 光 學 異 構 物 劑 量 增 加 呈 現 良 好 劑 量 反 應 關 係 , 因 此 證 明 SO 確 實 會 誘 發 tk 基 因 突 變 。 Von der Hude 於 1992 年 發 現 , 以 S- SO 誘 發 Chinese Hamster V79 細 胞 株 產 生 SCE 的 頻 率 雖 然 略 高 於 R- SO , 但 是 並 沒 有 達 到 統 計 上 之 顯 著 差 異 。 Watabe 於 1981 年 以 Ames test 對 細 菌 之 測 試 也 發 現 R - SO 與 S- SO 的 致 細 胞 突 變 率 沒 有 差 異 ( Watabe et al.,1981 )。 在 一 次 的 獨 立 實 驗 之 結 果 顯 示 , tk- total 背 景 突 變 率 為 5~22 ×10 - 6 之 狀 況 下 , 沒 有 看 出 SO 不 同 光 學 異 構 物 對 TK6 細 胞 致 突 變 之 差 異。此 結 果 顯 示 R - SO 與 S- SO 有 可 能 對 TK6 細 胞誘發相同的 tk-toatl 基因突變率。. 本 實 驗 結 果 顯 示 S O 與 SO 光 學 異 構 物 對 tk-fast 基 因 突 變 率 並 沒 有 達 到 統 計 之 顯 著 差 異,此 結 果 顯 示 R - SO 與 S- SO 誘 發 TK6 細 胞 中 tk -fast 基 因 之 突 變 率 有 可 能 沒 有 差 異 。 在 tk -slow 基 因 之 突 變 率 明 顯 高 出 tk-fast 基 因 , 而 且 在 三 次 之 獨 立 實 驗 中 , R - SO 造 成 TK6 細 胞 中 tk -s l o w 基 因 之 突 變 率 比 S- SO 高,顯 示 R - SO 造 成 TK6 細 胞 中 較 大 段 之 基 因 突 變。. 研 究 顯 示 在 human N -ras codon 61 中 Adenine 結 構 上 的 N 6 位 置 合 成 R - SO 與 S- SO 的 DNA adduct,經 細 菌 DNA 複 製 後 測 試 點 突 變 , 結 果 發 現 只 有 在 S(61. , 2). 位 置 上 之 DNA. adduct 會 引 發 約 1.6 % 的 A→ G 突 變 , 由 此 顯 示 不 同 光 學 異 31.
(36) 構 物 所 造 成 之 DNA adduct 並 不 一 定 會 造 成 突 變 。SO 與 SO 光學異構物細胞突變率受到許多因素之影響,包括去毒性酵 素 之 作 用 、DNA adduct 形 成 之 能 力 、DNA 修 補 酵 素 對 DNA adduct 之 修 補 與 DNA adduct 被 DNA 聚 合. 誤認之機率所. 影響,甚至在突變基因旁邊之基因序列也會影響基因突變率 (Latham et al.1993)。. tk -slow 基 因 突 變 率 與 細 胞 毒 性 有 相 同 趨 勢 ( R - SO ≧ S- SO ), 雖 然 都 沒 有 達 到 統 計 上 之 顯 著 意 義 , 但 是 tk-s l o w 基因突變率主要在測試較大段的基因突變或缺損,因此顯示 較 大 段 基 因 突 變( tk-solw 基 因 )率 比 hprt 或 tk-fast 基 因 的 突 變 率 高 。 雖 然 只 要 發 生 突 變 都 會 使 hprt 酵 素 或 tk 酵 素 功 能 喪 失 , 但 是 因 為 在 tk 基 因 旁 邊 有 許 多 生 長 必 須 之 基 因 , 因 此 SO 與 其 光 學 異 構 物 對 TK6 細 胞 所 造 成 大 段 基 因 缺 損 之 情 況,可 能 比 點 突 變 或 小 段 基 因 缺 損 之 影 響 較 大,尤其是 R - SO 對 TK6 細胞大段基因缺損之影響可能比 S-SO 重要。. 因 此 本 實 驗 選 擇 hprt 基 因 與 tk 基 因 作 為 指 標 基 因,以 測 試 SO 光 學 異 構 物 之 致 突 變 性 , 希 望 能 將 具 有 基 因 突 變 之 細 胞 收 集 , 日 後 將 進 一 步 分 析 DNA 序 列 以 建 立 突 變 圖 譜 , 探 討 SO 誘 發 hprt 基 因 突 變 之 主 要 突 變 位 置 與 所 發 生 的 突 變 種 類,以進一步瞭解 SO 與其光學異構物之分子突變機制。. 32.
(37) 第七章. 結論. 本 實 驗 比 較 R - SO、 S- SO 與(R+S) - SO 對 TK6 細 胞 的 毒性與致突變性,以無母數分析方法進行分析,其結果顯示 R- SO 與 S- SO 之 致 細 胞 毒 性 未 達 到 統 計 上 之 顯 著 差 異 , 顯 示在 SO 光學異構物間,對 TK6 細胞之毒性並無差異。. S- SO( 60,120 與 180 μ M)對 TK6 細 胞 hprt 的 突 變 率 比 R- SO 高 ( p<0.05 )。 在 tk-fast 基 因 上 不 同 光 學 異 構 物 之 致 突 變 性 經 統 計 分 析 結 果 , 沒 有 達 到 顯 著 差 異( p>0.05 )。 tk - total 與 tk -solw 基 因 之 致 突 變 性 顯 示 , 在 三 次 獨 立 實 驗 中 S- SO 的 致 突 變 性 都 雖 有 高 於 R - SO 的 趨 勢 , 但 是 仍 然 沒 有 達到統計上之顯著差異(p>0.05),. 以 上 結 果 顯 示 R - SO、S- SO 與 (R+S)- SO 三 者 對 人 類 淋 巴球細胞之致毒性與致突變性並沒有顯著差異,造成彼此間 沒 有 差 異 之 原 因 為 , 不 同 SO 光 學 異 構 物 對 TK6 細 胞 之 致 突 變 , 除 了 酵 素 影 響 外 , 還 會 受 到 不 同 SO 光 學 異 構 物 與 D N A 之結合力、DNA 修補與 DNA 複 製 時 被 誤 認 之 機 率 所 影 響 , 而 也 可 能 是 R- SO 與 S- SO 彼 此 間 互 相 拮 抗 , 導 致 R - SO 與 S-SO 之致細胞突變率無差異之原因。. 33.
(38) 第八章. 參考資料. 1.Agency for Toxic Substance and Disease Registry (ATSDR). (1990) Toxicological Profile for styrene, U.S. Public Health service, 7-75. 2.Baker.R.A. (1963) Threshold odor of organic. Journal American Water Workers Association. 55, 913-916. 3.Barale Roberto.(1991) The genotic toxicology of styrene and styrene oxide. Mutation Research.257, 107-126. 4.Bastlova, T., Vodicka, P., Peterkova, K., Hemminki, K., Lambert, B. (1995) styrene oxide- induced HPRT mutations, DNA adducts and DNA strand breaks in cultured human lymphocytes. Carcinogenesis. 16(10), 2357-62. 5.Belvedere, G. and Trusi, F.(1981) styrene oxidation to styrene oxide in human blo od erythrocytes and lymphocytes, Research. Commununication Chemistry Pathology Pharmacology., 33,273-282. 6.Bolt, H.M. (1993) Interspecies differences in metabloism and kinetics of 1,3- butadiene, isobutene and styrene IARC Scientific Publications, No.127, 37-44. 7.Budavari, S., O'Neil, M.J., Smith A, Heckelman, P.E. (Eds.) (1989)The Merck Index, 11th ed. Merck & Co., Inc., Rahway, NJ, p.1397. 8.Carlson, G.P. (1997) Effects of inducers and inhibitors on the microsomal metabolism of styrene to styrene oxide in mice. Journal of Toxicology and Environmental Health, 51, 447-488. 9.Carlson, G.P. (1998) Metabolism of styrene oxide to styrene glycol by mouse liver and lung. Journal of 34.
(39) Toxicology and Environmental Health. 53(1), 19-27. 10.Carlson, G.P., Hynes, D.E., and Mantick, N.A. (1998) Effects of inhibitors of CYP1A and CYP2B on styrene metabolism in mouse liver and lung microsomes. Toxicology Letters. 98(3), 131-7. 11.Casciano, D.A., Aidoo A., Chen T., Mittelstaedt, R.A., Manjanatha, M.G. and Heflich R.H.(1999) Hprt mutant frequency and molecular analysis of Hprt mutantions in rats treated with mutagenic carcinogens. Mutation Research.431, 389-395. 12.Chakrabarti, S., Zhangand, X.X., Richer, C.L. (1997) Influence of duration of exposure to styrene oxide on sister chromatid exchanges and cell-cycle kinetics in culture human blood lymphocytes in vitro. Mutation Research.395, 37-45. 13.Cochrane, J.F and Skopek, T.R. (1994) Mutagenicity of butadiene and its epoxide metabolites:I. Mutagenic potential of 1,2 -epo xybutene,1,2,3,4 - diepoxybutane and 3,4 -epoxy- 1,2,- butanediol in cultured human lymphoblasts.Carcinogenesis.15(4),713-717. 14.Crespi, C.L., Seixas, G.M., Turner, T.R.,Ryan, C.G. and Penman,B.W.(1985) Mutagenicity of 1,2- dichloroethane and 1,2- dibromoethane in two human lymphoblastoid cell lines. Mutation Research.142, 133-140. 15.Drummond, L., Caldwell, H, J., Wilson, K. (1989) The metabolism of ethylbenzene and styrene to mendelic acid stereochemical consideration.Xenobioticam,19,199-207. 16.Foureman, G.L., Harris, C., Guengerich, F.P. and Bend, J.R. (1988) Stereoselectivity of styrene Oxidation in 35.
(40) Microsomes and in Purified Cytochrome P-450 Enzymes from Rat Liver. The Journal of Pharmacology and Experiment Therapeutics, 248(2), 492-497. 17.Gadberry, M.G ., DeNicola, D.B. and Carlson,G.P. (1996) Pneumotoxicity and hepatotoxicity of styrene and styrene oxide. Journal of Toxicology and Environmental Health. 48(3), 273-94. 18.Hallier, E., Goergen, H.W. et., al. (1994) Intervention study on the influence of reduction of occupatonal exposure to styrene on sister chromatid exchanges in lymphocytes. International Archives of Occupational & Environmental Health, 66(3), 167-72. 19.Hemminki, K. and Vodicka,O. (1995) styrene:form characterisation of DNA adduct to application in styrene -exposed lamination works. Toxicology Letters. 77,153-161. 20.Hiratsuka, A., Yokoi,A., Iwata,H., Watabe,T., Satoh,K., Hatayama,I. and Sato, K. (1989) Glutathione conjugatiuon of styrene 7,8 -oxide enantiomers by major glutathione transf erase isoenzymes isolated from rat livers.Biochemical Pharmacology.38(24),4405-4413. 21.Honma, M., Hayashi, M. and Sofuni T.(1997) Cytotoxic and mutagenic responses to X-rays and chemical mutagens in normal and p53- mutated human lymphoblastoid cells. Mutation Research,374,89-98 22.Horvath, E., Pongeracz,K., Rappapport,S. and Bodell,W.J. (1994) 32P postlabelling detection of DNA adduct in mononuclear cells of workers occupationally exposed to styrene. Carcinogenesis.15,1309-1315. 23.H ou,S.- M.,. Falt,S.. and 36. Steen,A.- M.(1995). HPRT.
(41) mutant frequency and GST M1 genotype in non-smoking heathy individuals. Environment mol.mutagenesis.25,97-105. 24.Howard, P.H.(1989) Handbook of Environmental Fate and Exposure Data for Organic Chemicals, Vol I,490-498. 25.Keith, L.H. and Walters, D.B. (Eds.). (1987) Compendium of Safety Data Sheets for Research and IndustrialChemicals, Part VI ; VCH Publishers, Deerfield Beach, 3178-3179. 26.Korn,M., Wodarz,R., Drysch,K., and Schmachl,W. (1987) Stereometablism of styrene in man. Urinary excretion of chiral styrene metabolites.Arch.Toxicol.60,86-88. 27.Kramer,A., Pudil,J., Frank,H., Oesch,F. and Glatt,H.(1993) Some substrates and inhibitors of cytosolic epoxide hydrolase induce sister-chromatid exchanges in mammalian cells ,but do not induce gene mutations in salmonella typhimurium and V79. Mutation Research.290, 165-174. 28.Latham,G.J., Zhou,L., Harris,C.M., Harris,T.M., Lloyd,R.S. (1993) The replication fate of R - and S- styrene chirality of the lesion and the local sequence context. Journal of Biological Chemistry, 268(31), 3427-34. 29.Leibman, K.C. and Ortiz E. (1968) styrene epoxide An intermediate in microsomal oxidation of styrene to its glycol. Pharmacologist,.203. 30.Lijinski, W.(1986) Rat and mouse fore stomach tumors induced by chronic oral administrate of styrene oxide.J.Natl.Cancer inst 77,471-476.. 37.
(42) 31.Linhart I., Smejkal J.and Mladkova I.(1998) Stereochemical aspects of styrene biotransformation. Toxicology Letters. 94(2):127-35. 32.Nakajima,T., Elovaara,E., Gonzalez,F.J., Gelboin,H.V., Vainio,H. and Aoyama,T. (1993) Characterization of the human cytochrome P- 450 isozymes responsible for styrene metabolism IARC Scientific Publications No.127,01-108. 33.Nishi,Y., Hasegawa,M.M., Taketomi,M., Ohkawa,Y. and Inui,N.(1984) Comparison of 6- thioguanine- resistant mutation and sister chromatid exchanges in chiness hamster V79 cells with forty chemical and physical agents. Cancer research. 44,3270-3279. 34.Otteneder, M. and Lutz,W.K.(1999) Correlation of DNA adduct levels with tumor incidence:carcinogenic potency of DNA adduct. Mutation Research.424, 237-247. 35.Pagano, D.A., Yagen,B., Hernandez,O., Bend,J.R., Zeiger,E. (1982) Mutagenicity of (R) and (S) styrene 7,8- oxide and the intermediary mercapturic acid metabolites formed from styrene 7,8- oxide. Environmental Mutagenesis, 4(5), 575-84. 36.Pfaffli, P. and Saamanen A.(1993) The occupation scene of styrene. International Agency for Research on Cancer, Lyon. 15-44. 37.Rink,R., and Janssen,D.B.(1998) Kinetic mechanism of the Enantioselective Conversion of styrene oxide by Epoxide hydrolase from Agrobacterium radiobacter AD1.Biochemistry.37,18119-18127. 38.Scandinvian Expert group on documentation -styrene. (1979) 38. limit Arbete. value Och.
(43) Halsa,14,36. 39.Scott,D. (1993) Cyto gene tic studies of workers exposed to styrene,a review.In Sorsa,M.,Peltonen,K., Vainio,H. and Hemminki, K. (eds). Butadiene and styrene , Assessment of Health Hazards. International Agency for Research on Cancer,Lyon.275-286. 40.Scott,D., and Preston,R.J.(1994).A re- evaluation of the cyto gene tic effects of styrene . Mutation Research,318,175-203. 41.Seppalainen, A. and Harkonen,H. (1976) Neurophysiological finding among workers Occupation exposure to styrene.Scand Journal Work Environ Health, 3,140-146. 42.Seiler J.P.(1990) Chirality- dependent DNA reactivity as the possible cause of the differential mutagenicity og the two components in an enantiomeric pair of epoxides. Mutation Research.245:168-169. 43.Sinsheimer,J.E., Chen,R., Das,R.K., Hooberman, B.H., Osorio,S. and You,Z. (1993) The genotoxicity of enantiomeric aliphatic epoxides. Mutation Research.298,197-206. 44.Spelberg,J.H.L., Rink,R., Kellogg,R.M. and Janssen,D.B.(1998) Enantio-selestivity of recombinant epoxide hydrolase from Agrobacterium radiobacter Tetrahedron,Asymmetry 9,459-466. 45.U.S. Environmental Protection Agency. Health and Enviro - nmental. (1984) Effects Profile for styrene. Office of Research and Development, U.S. EPA, Washington, D.C. ECAO-CIN-P103. 46.Vainio, H., Hemminki, K. and Elovaara E. (1977) 39.
(44) Toxicity and styrene and styrene oxide on chick embryos.Toxicology, 8.(3)319-325. 47.Von der hude,W.,Carstensen S.,Gurtler R.,Gunter O. (1992) Structure- activity relationships of eopxides:induction of sister -chromatid exchanges in V79 cells by enantiomeric epoxides. Mutation Research.278:289-297. 48.Vodicka,P., Bastlova,T., Vodickova,L., Peterkova,K., Lambert,B. and K.Hemminki.(1995) Biomarkers of styrene exposure in lamination workers,levels of O6- guanine DNA adducts, DNA strand breaks and mutant frequencies in the hypoxanthine guanine phosphoribosyl- transferase gene in T- lymphocytes. Carcinogenesis, 16(7), 1473-1481. 49.Walles, S.A.A., Edling, C., Anundi,H., Johanson,G. (1993) Rxposure dependent increase in DNA single strand breaks in leucocytes from workers exposed to low concentrations of styrene.British Journal of Industrial Medicine,.50,570-574. 50.Wtabe, T., Ozawa, N. and Yoshikawa, K. (1981). Stereochemistry in the oxidative metabolism of styrene by hepa microsomes.Biochem.Pharmacol,.30,1695-1698. 51.Watabe, T., Ozawa, N. and Yoshikawa, K. (1982) . Studies on metabolism and toxicity of styrene .V.The metabolism of styrene,reacmic,R-(+), and S- (-) - phenyloriranes in the rat, Journal Pharmacobio-Dyn.5,129-133. 52.行 政 院 勞 工 委 員 會 (1995). 勞 工 作 業 環 境 空 氣 中 有 害 物 質 容許濃度標準. 53.國 貿 局 海 關 總 稅 務 司 署 .(1979~1984). 進 出 口 貨 品 統 計 月 40.
(45) 報. 54. 經 濟 部 統 計 處 .(1979~1984). 中 華 民 國 台 灣 工 業 生 產 統 計月報. 41.
(46) 30 R S R+S control. RCN (Fold). 25 20 15 10 5 0 0. 1. 2. 3. DAY 圖1. 180μM SO 與 SO 光學異構物誘發 TK6 細胞 之生長遲緩現象. 42.
(47) 140. Relative surviva (%). 120. R S. 100. R+S. 80 60 40 20 0 60. 120. 160. 180. dose (μM). 圖 2.以 細 胞 相 對 存 活 率( RCN)評 估 SO 與 SO 光 學 異 構物對 TK6 細胞之毒性. 43.
(48) 120. R. Relative survival (%). 100. S 80. R+S. 60 40 20 0 60. 120. 160. 180. dose (μM). 圖 3. 以 細 胞 群 落 形 成 能 力( PE)評估 SO 與 SO 光 學 異 構 物 對 TK6 細胞之毒性. 44.
(49) R. -6. hprt MUTANT FREQUENCY (X 10 ). 70. S. 60. R+S 50 40 30 20 10 0 0. 60 120 dose(μM). 180. 圖4. SO 與 SO 光學異構物在 TK6 細胞誘發 hprt 突變率. 45.
(50) 60 R ). S. tk MUTANT FREQUENCY (X 10. -6. 50. R+S. 40. 30. 20. 10. 0 0. 60. 120 160 dose (μM). 180. 圖 5 SO 與 SO 光學異構物在 TK6 細胞誘發 tk fast 基因突變率. 46.
(51) R. tk MUTANT FREQUENCY (X 10. -6. ). 140 120. S R+S. 100 80 60 40 20 0 0. 60. 120. 160. 180. dose (μM). 圖 6. SO 與 SO 光學異構物對在 TK6 細胞誘發 tk total 因突變率. 47.
(52) tk MUTANT FREQUENCY (X 10 -6 ). 120 R 100. S R+S. 80 60 40 20 0 0. 60. 120. 160. 180. dose(μM). 圖 7 SO 與 SO 光學異構物對在 TK6 細胞誘發 tk slow 基因突變率. 48.
(53) 表 1. SO 與 SO 光學異構物對 TK6 細胞之致突變率 R-SO Dose (μM). S-SO. (R+S)-SO. Mutant Frequencies (X10 -6). Hprt 基因 control. 8.86. 9.31. 11.05. 60. 16.68. 21.83. 26.1. 120. 26.56. 32.96. 35.03. 180. 32.08. 50.04. 45.03. control. 7.25. 9.50. 9.30. 60. 19.33. 14.05. 14.25. 120. 23.05. 30.95. 28.33. 180. 33.67. 34.64. 44.08. control. 7.55. 9.71. 15.98. 60. 30.86. 22.25. 30.81. 120. 41.07. 28.01. 31.72. 180. 89.52. 66.04. 78.18. control. 12.23. 17.53. 20.65. 60. 46.68. 33.80. 45.10. 120. 64.13. 55.17. 57.80. 180. 123.58. 87.84. 119.50. tk-fast. tk-slow. tk-total. 49.
(54) (A). H CH. C H. (B). (C). H. O. O. CH CH2. CH CH2 H. α. β. 附圖 1 SO 與 SO 光學異構物之化學結構 (A)styrene (B)R-SO (C)S-SO. 50.
(55) Styrene CYP-450. SO. DNA adduct. ((R+S)-SO) Epoxide. 51. glutathione. hydrolase. S-transferases. (EH). (GST) mercapturic acid. styrene glycol mandelic acid. 附圖 2. Styrene 之活化與去活化路徑. Mutation.
(56) 1. 2. 3. 5’-C GGA CAA GAA G-3’ 60. 61. 62. H C. H CH2OH. C. NH. NH N. N. N. N N. S-isomer. CH2OH. N. N. N. R-isomer. 附 圖 3. SO 光 學 異 構 物 與 人 類 N-ras 基 因 codon. 61 上之 adenine 中 N7 位置形成之 DNA adduct (Latham et al.,1993).
(57) 人 類 淋 巴 母 細 胞 ( T K 6 )經 C H A T 處 理 7 2 小 時 去 除 C H A T 後 , 繼 續 培 養 3-4 天 以 R - S O 、 S-SO 或 S O 處 理 2 4小 時. 53. 離心後去除化學物質 (2) 細 胞 毒 性. (1) 突 變 率 以正常培養基培養. 立 刻 接 種 於 96 3∼ 4天. 6∼ 7天. 接種於含 6-T G 之 培 養 基 中 12∼ 1 5天. 接種於含 TFT之 培 養 基 中. well plate 12 ∼ 1 5天. RCN. 10 天. 再 加 TFT. 計算細胞增加倍數. 以 細 胞 群 落 (c o l o n y ) 數 目 計 算 細 胞 相 對 毒 性 ( R S). 10 天. 以 細 胞 群 落 (c o l o n y ) 數 目 , 計 數 突 變 率. 附圖 4. 測量細胞毒性與突變率之實驗流程.
(58)
數據
+3

相關文件
For the primary section, the additional teaching post(s) so created is/are at the rank of Assistant Primary School Master/Mistress (APSM) and not included in calculating the
So, we develop a tool of collaborative learning in this research, utilize the structure of server / client, and combine the functions of text and voice communication via
In the table boldface line was the S curve means students and dotted line was the P curve means problem.
In this study, ohmic contact is formed by Ni convert to NiO X after annealing, according to the results from x-ray photoelectron spectroscopy and the Cserveny’ s
Notice that if the dx in the notation for an integral were to be interpreted as a differential, then the differential 2xdx would occur in (1) and so, formally, without justifying our
T transforms S into a region R in the xy-plane called the image of S, consisting of the images of all points in S.... So we begin by finding the images of the sides
6 《中論·觀因緣品》,《佛藏要籍選刊》第 9 冊,上海古籍出版社 1994 年版,第 1
To proceed, we construct a t-motive M S for this purpose, so that it has the GP property and its “periods”Ψ S (θ) from rigid analytic trivialization generate also the field K S ,
