國立臺灣大學生物資源暨農學院森林環境暨資源研究所 碩士論文
Department of Bioresources and Agriculture College of Forestry and Resource Conservation
National Taiwan University Master Thesis
轉殖小枯草桿菌纖維素酶對於雲杉紙漿 吸附、降解與磨漿促進效能
Effect of Recombinant Paenibacillus Cellulase on Adsorption, Degradation and Refining Assistance of Spruce Pulps
廖婉均 Wan-Jyun Liao
指導教授:柯淳涵 博士 Advisor: Chun-Han Ko, Ph.D.
中華民國 100 年 6 月
June, 2011
誌謝
此篇研究謹獻給我的老師──柯淳涵教授。當初若非老師的邀約,學生也不 會再往上攻讀研究所,也不會有這篇論文誕生,感謝老師對我大學四年及碩士班 兩年的指導,我是個任性的人,好在老師願意包容,不時鼓勵著我,讓我能在兩 年取得碩士學位。能夠當老師的學生,是我的運氣,感謝的話太多太多,反而說 不盡實。
也謝謝我的母親和父親,辛勤工作支持我唸書。老家那裡不少同學國中畢業 就必須負擔家計,爸媽卻願意栽培我到今日,早出晚歸,餐桌上沒有餘肉,而我 從來不缺書錢。等我完成學業,一定會讓他們吃上世上最美味的食物、穿最好的 衣服、住最好的房子,聊表他們的養育之恩。
研究所期間,受到許許多多老師、學長學姊及同學們的照顧。我的實驗最大 得助於楊秉元及陳芳瑾學長,謝謝他們的指導;另外還有說話很賤但卻會聽我抱 怨又能點出我實驗盲點的伯亨學長;連續好幾個星期七點起床,無怨無尤,帶我 去離心的藍婷(小白)學姊;總是很溫柔,帶了許多點心餵養我的晶瑩學姊;博
學多聞,可以諮詢多國語言又是美食專家的明訓學長;實驗室 HPLC 專家,晚上
八九點還空腹帶我實驗的政喬學長;還有我研一舉目無親的時候幫我找到房子,
在生活照顧我一整年的品叡學姊及錦儀學姊,另外也感謝阿璧學姊對我的種種照 料。我也很慶幸能遇到同屆四個好同學,都是帥哥大美女,我很幸福。特別是和 千瑩去華紙實驗,都是她在罩我,和她討論過後總是能解開紙漿實驗的難題;智 遠教會我許多實驗技巧,非常可靠;嘉介在百忙之中還大力幫忙聚戊醣實驗,沒 有他們,就不能順利完成研究。另外,只要看到蕙菁,就讓人心情愉悅。研二這 年來了可愛的學弟妹,慧儀、佳興、恆賓,有他們在,日子生色不少。
實驗室以外,還得助於各方幫助,尤其是中華紙漿花蓮廠各位大哥大姊,無 條件出借化學實驗室的王大哥和帶我煮漿的傅大哥、美麗的菁姊,還有生管室、
物理實驗室其他前輩,期間總是麻煩他們許多事情,很抱歉也很感謝。也謝謝王 國鼎學長千里將木片空運來臺。
我要感謝的人真的太多了,最後也只能謝天了。
摘要
本研究探討 Paenibacillus 所生產之纖維素酶於生質能與造紙生產的作用,以
此特定纖維素酶與雲杉紙漿纖維素反應,研究其吸附、水解效能及和木質素 (lignin)、酵母添加物 (yeast extract) 的交互作用,以及探討酵素處理對於磨漿的纖 維性質變化。
研究顯示,與水解作用無關的雜蛋白質會優先吸附於紙漿纖維外部的木質 素,而外加的酵母抽出物會增強雜蛋白吸附於木質素現象。纖維素酶水解系統會 受到以下複雜因子影響:與水解作用無關的雜蛋白質、紙漿木質素和纖維素含量。
實驗分別加入纖維素酶劑量5 IU 及 50 IU 與不同木質素漿樣反應,相對於原漿聚
合度,其高木質素 (12.4%) 漿樣之 LODP 僅降解至 82.3 % 與 62.1 % 的程度,而 低木質素 (1.2%) 漿樣之 LODP 則降至 69.7 % 與 60.7 %。此外,添加酵母抽出物,
可使雜蛋白質與木質素作用而讓水解蛋白正確吸附在目標物纖維素,因而增加了 水解酶的效率,改變水解機制。
另一方面,纖維素酶的前處理將改變磨漿紙漿性質。酵素前處理,會使磨漿 纖維表面產生變化,增加纖維對水的親和力,如紙漿濾水性質及纖維水合程度;
此外,以微纖維分析儀 (Morfi) 觀察微纖維尺度的紙漿纖維,比較無酵素處理的 控制組與有酵素前處理組別,可知纖維素酶前處理會造成較寬的纖維、扭結角度 (kink angle) 較小、較少漿絮 (small fines)、更多破裂纖維和增加微纖維長度的比 例。經酵素處理後,紙力也產生變化。透過掃描式顯微鏡 (SEM) 可發現纖維素酶 前處理會增加磨漿纖維表面粗糙度 (coarseness)。
關鍵詞:酵素吸附、酵素水解、聚合度、磨漿、微纖維分析
ABSTRACT
This article focused on two sides of action recombinant Paenibacillus cellulase on spruce kraft pulps. One is about the study of interaction of cellulase and pulp on different conditions among four kinds of lignin contents and adding yeast extract.
Another subject would discuss the refined fiber properties with the cellulase-pretreatment.
We observed that irrelevant proteins preferentially adsorbed on pulp surface with higher lignin contents than cellulase, and the addition of yeast extract strengthened the above trend. The cellulase hydrolysis rates were influenced by various factors on this system: the presence of irrelevant proteins, pulp lignin, and cellulose contents. The high lignin sample (12.4%) with cellulase dosages of 5 IU and 50 IU would degrade DP of 82.3 % and 62.1 %; and the low lignin sample with same enzyme treated would degrade DP of to LODP to 69.7% and 60.7% of original LODP values. Addition of yeast extract further promoted preferential adsorption of irrelevant proteins, and increased the hydrolysis rates by cellulase and altered the mode of hydrolysis.
On the other hand, the pretreatment of cellulase through refined would raise the pulp properties. The enzyme modified the interfacial properties of fibers, increasing the affinity for water, such as pulp drainability and fiber hydration. Moreover, the enzymatic pretreatment also changed the distribution of microfiber (Morfi), causing wider fiber, less kink angle and small fines, more broken ends and rate in length of microfiber. The strength properties of the pulp had change, and the SEM also showed that the fiber would increase the coarseness with cellulase treated.
Keywords: Adsorption; Hydrolysis; Degree of Polymer; Refined; Morfi
CONTENTS
口試委員會審定書 ...i
誌謝 ... ii
摘要 ... iii
Abstract ...iv
Contents ... v
List of Figures ... vii
List of Tables ...ix
I Introduction……… ... 1
II Literature Reviews ... 3
(I) Endoglucanase and Paenibacillus campinanesis BL 11 ... 3
(II) Saccharification of Biofuels ... 4
1. Adsorption ... 4
2. Hydrolysis ... 5
3. Lignin ... 6
4. Additives ... 7
(III) Refinig of Pulp Stock ... 8
1. PFI ... 8
2. Scanning Electron Microscope (SEM) ... 9
III Materials and Methods ... 10
(I) Microorganism and Growth Measurements ... 10
(III) Expression of the Recombinant Endoglucanase ... 10
(III) Fiber Preparation and Chemical Analysis ... 10
(IV) DEDD Bleaching ... 11
(V) Characterization of Pulps ... 12
(VI) Cellulase Activity ... 13
(VII) Enzymatic Hydrolysis of Pulp ... 13
(VIII) Additive of Yeast Extract ... 13
(IX) Endoglucanase Accessibility and Digestibility ... 14
(X) Enzyme Treatment ... 15
(XI) Pulp Refining and Handsheets Properties ... 15
(XII) Morfi ... 16
(XIII) Scanning Electron Microscope (SEM) ... 16
IV Results and Discussion ... 17
(I) Impacts of Lignin Contents and Yeast Extract Addition on the Interaction between Spruce Pulps and Crude Endoglucanase ... 17
1. Effects on the Accessibility of Proteins... 17
2. Effects on the Preferential Adsorption of Irrelevant Proteins ... 22
3. Effects of Pulp Lignin Contents on Hydrolysis ... 24
4. Effects of Yeast Extract Addition on Hydrolysis... 28
(II) Impact of Endoglucanase on Refining of Spruce Kraft Pulp ... 32
1. Effect on the Drainability and Fiber Hydration ... 32
2. Pulp Biorefining and Effect on Pulp Properties ... 34
3. Fibers of Morfi Analysis ... 35
4. Fibers of SEM Analysis ... 38
V Conclusions… ... 40
VI References.. ... 41
LIST OF FIGURES
Fig. 1 Effect of yeast extract addition on the accessibility of protein (crude recombinant Paenibacillus endoglucanase) on four spruce pulps at 4℃ ... 19 Fig. 2 Effect of yeast extract addition on specific endoglucanase activity of
proteins in supernatants ... 23 Fig. 3 Effect of pulp lignin contents on the reduction of pulp DP by crude
recombinant Paenibacillus endoglucanase at different dosages during hydrolysis at pH 7, 40℃ ... 25 Fig. 4 Effect of pulp lignin contents on the release of reducing sugars by crude
recombinant Paenibacillus endoglucanase at different dosages during hydrolysis at pH 7, 40℃ ... 27 Fig. 5 Effect of yeast extract addition on the reduction of pulp DP (degree of
polymerization) during hydrolysis at pH 7, 40℃ ... 29 Fig. 6 Effect of yeast extract addition on the release of reducing sugars during
hydrolysis at pH 7, 40℃ ... 29 Fig. 7 Effect of PFI-refining revolutions with control (0 IU), 1 IU, 2.5 IU and 5 IU
endoglucanase dosage on freeness ... 33 Fig. 8 Effect of PFI-refining revolutions with control (0 IU), 1 IU, 2.5 IU and 5 IU endoglucanase dosage on water retention value ... 33 Fig. 9 Effect of of PFI-refining revolutions with control (0 IU), 1 IU, 2.5 IU and 5 IU endoglucanase dosage on pulp strength ... 34 Fig. 10 Morfi fiber width of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions ... 35
Fig. 11 Morfi fiber curl of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions ... 36 Fig. 12 Morfi kink angle of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions ... 36 Fig. 13 Morfi fines length of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions. ... 37 Fig. 14 Morfi fines area of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions ... 37 Fig. 15 Broken ends and rate in length of microfiber versus PFI-resolutions treated with control (0 IU), 1 IU, 2.5 IU and 5 IU ... 38 Fig. 16 Scanning electron microscopic images of PFI revolutions 0, 2000 and 4000 surface image of control and endoglucanase treated (5 IU) fibers. ... 39
LIST OF TABLES
Table 1 Chemical properties of four spruce pulps ... 17 Table 2 Effects of pulp lignin contents and yeast extract addition on the maximum adsorption capacity and equilibrium constants of the protein onto four spruce pulps ... 21 Table 3 Effects of pulp lignin contents and cosages of endoglucanase applied on
degradation rate constants (k) and leveling-fff degree of polymerization (LODP) during hydrolysis of four spruce pulps ... 21 Table 4 Effects of yeast extract addition and dosages of endoglucanase applied on degradation rate constants (k) and leveling-off degree of polymerization (LODP) during hydrolysis of spruce pulps LIG 12.4 and LIG 1.2 ... 31
I Introduction
Today, people have known that manufacturing industries would produce much pollution endangering our environment and the biological process is superior to the new methods. Cellulases, one of most effective materials of biological production including biofuels and papermaking, have been studied by many scholars for a long time. In the previous studies, the Paenibacillus campinasensis BL 11, whose strain was isolated from a high temperature, alkaline environment at the washing section rich in black liquor after pulping and had been identified with phylogenic and physiological measures, could possess the cellulases (Ko et al., 2007; Ko et al., 2010b).
Biomass is a renewable resource that can be used for the sustainable production of biofuels. In the past, the sources of biomass were most from corns and soybeans, both of which are major foods in world, and would cause food crisis. To avoid the defect, the abundant forestry resource would be a good choice.
The process of producing biobfuels involved two steps: enzymatic hydrolysis converting cellulose of wood into reducing sugar and fermentation converting reducing sugar to bioethanol. At the first step, bioconversion of cellulose to soluble sugars and glucose is catalyzed by a group of enzymes named as cellulases (Bhat, 2000). Cellulase could be least three different types: cellobiohydrolase (CBH, EC 3.2.1.91), β-glucosidase (EC, 3.2.1.21) and endoglucanase (EG, EC 3.2.1.4). Endoglucanase (1,4-β-D-glucan-4-glucanohydrolase) randomly hydrolyzes the β-1,4 bonds within the cellulose molecule. As the cellulose depolymerization performed by endoglucanases is the rate-limiting step for the whole hydrolysis and plays the central role in the saccharification process (Lynd et al., 2002; Zhang et al., 2006; Himmel et al., 2007).
The adsorption kinetics and cellulase activities were affected along different
hydrodynamic materials, including lignin (Galbe and Zacchi, 2002; Berlin et al., 2006;
Hubbell and Ragauskas, 2010) and additive yeast extract (Ander et al., 2008; Ramírez et al., 2008; Mamma et al., 2009; Ouyang et al., 2010). This study will discuss the impacts of lignin contents and yeast extract addition on the interaction between spruce pulps and endoglucanase.
Besides bioethanol production, cellulases have a wide range of applications, including food products, animal feed, and in the textile and chemical industries, as well as waste management, the medical/pharmaceutical industry, protoplast production, and pollution treatment (Bhat, 2000). A particular area of interest is the pulp and paper industry (Bajpai, 1999) in which microbial cellulases were reported to improve fiber properties (Dienes et al., 2006).
In addition to biofuels, this study will confer that how endoglucanase pretreatment influence the refined fiber of spruce pulp. Only enzyme pretreatment has limit effect on fiber surface, but through refined by PFI (Papir-og fiberinstituttet), the mechanical treatment might improve the effect of biological pretreatment, such as internal and external fibrillation, fines formation, fiber shortening or cutting and fiber curling or straightening (Gil et al., 2009; Ko et al., 2010b). It would be described elsewhere that the result of refining and cellulase treating influence the characteristics of microfibers by freeness, water retention value (WRV), Morfi and scanning electron microscope (SEM).
II Literature Reviews
(I) Endoglucanase and Paenibacillus campinanesis BL 11
Cellulases could be categorized into: endoglucanase (1,4-β-D-glucan-4- glucanohydrolase; EC 3.2.1.4), exoglucanase (1,4-β-D-glucan-glucohydrolase;
cellobiohydrolase; EC 3.2.1.74), and β-glucosidase (β-D-glucoside glucohydrolase; EC 3.2.1.21) (Lynd et al., 2002). The endoglucanases randomly hydrolyze the β-1,4 bonds within the cellulose molecule and they are the rate-limiting step for the whole hydrolysis (Himmel et al., 2007). Cellulases could be applied in the pulp and paper industries (Bajpai, 1999); the identification of cellulases and their potential are available but still have some obstacles for commercial and industrial developments and applications (Jeffries, 2008; Lecourt et al., 2010). Several members of the genus Paenibacillus secrete diverse assortments of extracellular polysaccharide hydrolyzing enzymes and their xylanolytic and cellulolytic systems are gradually being identified (Pason et al., 2006; Wang et al., 2008; Akaracharanya et al., 2009; Maki et al., 2009;
Waeonukul et al., 2009; Pason et al., 2010). A novel recombinant endoglucanase, derived from Paenibacillus campinasensis BL11 (Ko et al., 2007), demonstrated exceptional thermostability and wider pH adaptability was expressed by E. coli and weight loss in gampi and paper mulberry fibers after enzyme treatment validate the potential of this treatment in fiber degumming; in other words, a high-temperature alkaline environment is the best source in which to isolate micro-organisms and associated enzymes for industrial applications that are specific to such environments (Ko et al., 2011). The cellulases produced from Paenibacillus campinasensis BL11 had showed capability of pulp refining assistance (Ko et al., 2010b).
(II) Saccharification of Biofuels 1. Adsorption
The adsorption of cellulases onto lignocellulose is necessarily the first step of the hydrolytic process and corresponds to a phase transfer of the free enzyme (in solution) to the insoluble substrate (solid phase) (Henrissat, 1994). An early recognized feature of the adsorption of cellulases onto cellulose is that certain enzymes adsorb more closely than others and that the tight-binding enzymes are the most efficient for the hydrolysis of cellulose (Klyosov, 1990).
Enzyme adsorption was followed by analyzing the activity and the protein amount of the individual free enzymes in the hydrolysis supernatant. Most enzymes adsorbed quickly at early stages of the hydrolysis and remained bound throughout the hydrolysis, although the conversion reached was fairly high (Várnai et al., 2011).
The presence of cellulose-binding module (CBM) has been found to have a profound role in the adsorption of cellulases on lignocelluloses. The cleavage of CBM results in a strongly reduced enzyme activity toward insoluble cellulose while the enzyme activity on soluble substrates is fully retained (Tomme et al., 1988; Nidetzky et al., 1994).
Similarly, the absence of CBM leads to decreased adsorption on lignin (Palonen et al., 2004).
Bansal et al. (2009) indicated that the exact mechanism of cellulases on lignocellulosic biomass is not completely understood, various factors affecting the enzymatic rates and activities. Different assumptions regarding rate limiting factors and basic substrate–enzyme interactions.
In this study, the adsorption of endoglucanase from our lab before the hydrolysis of spruce pulps with four different lignin contents and yeast extract. The adsorption of the enzymes was followed by two different methods: by measuring the individual enzyme
activities and by quantifying the protein amount on Bradford method, revealing distinct differences in the adsorption behavior of the enzymes on the different substrates and with the additive.
2. Hydrolysis
Cellulases are a group of proteins that degrade cellulose to glucose by hydrolyzing β-1,4-glucosidic bonds in the polysaccharide molecule. Secreted by bacteria or fungi, they are a crude mixture of several enzymes: endoglucanases (EG), exoglucanases, and β-glucosidases. All three types of enzymes exhibit different activity on cellulose.
Endoglucanases cut at random amorphous sites of the cellulose chain, producing new reducing (also called reducing power) or non-reducing ends and oligosaccharides of various lengths. Exoglucanases attack the reducing or non-reducing ends of the cellulose chain, liberating glucose or cellobiose as major products; and β-glucosidases remain in the solution hydrolyzing small chain reducing sugars to glucose (Lynd et al., 2002; Verenich et al., 2007).
The major obstacles in the enzymatic hydrolysis of lignocellulose into sugars are related to the crystalline structure of cellulose and to the complexicity of the raw materials, as the presence of hemicellulose and lignin in the pretreated raw material (Schmidt et al., 1995; Berlin et al., 2006; Várnai et al., 2010). Softwood is generally considered among the most difficult and recalcitrant raw materials for the enzymatic hydrolysis (Galbe and Zacchi, 2002). The reducing sugar yield hold significant index of cellulose hydrolysis. Enzymatic saccharification of cellulose is generally described as a heterogeneous reaction system in which cellulases in an aqueous environment react with the insoluble, macroscopic and structured cellulose, containing highly ordered and less ordered regions (Arantes and Saddler, 2010; Jäger et al., 2010). Before the final product
such as glucose was released, an enzyme-substrate reaction intermediate must form.
Hence, the model structures employed to describe the enzyme-catalyzed hydrolysis of lignocellulose are generally based on modifications of Michaelis-Menten type kinetics.
However, the models are often combined with adsorption kinetics (Bansal et al., 2009;
Andrić et al., 2010) and degradation constant (Keller et al., 2000). The degraded kinetic constant (k) can be an index of lignocellulose DP degrading rate and levelling-off degree of polymerization (LODP) can indicate the utmost degraded DP of reacted pulp (Calvini, 2005).
In this study, the enzymatic hydrolysis ability would detected by reducing sugar and DP. Moreover, the k constant and LODP would be calculated from DP.
3. Lignin
Lignin, the major component of lignocellulosic biomass, is mainly responsible for retarding enzymatic hydrolysis by forming a physical barrier surrounding cellulose microfibrils, and many studies indicated that lignin is the major components limiting enzyme infiltration into the cell walls. The complex component also compasses cellulose microfibrils, preventing the access of cellulases to the cellulose chains (Fan et al., 1980; Yang and Wyman, 2004; Kristensen et al., 2008; Siqueira et al., 2011). The study of Lawoko et al. (2006) demonstrated that lignin was linked through covalent bonds to all the major polysaccharides in the wood cell wall, by arabinoglucuronoxylan, galactoglucomannan, glucomannan, pectin and cellulose. It was also evident that the lignin polymer cross-links various polysaccharides to each other. Such a network structure played an important mechanical role in the properties of the secondary xylem from wood. Furthermore, the progressive degradation of lignin-carbohydrate networks might be important in technical wood processes, such as chemical pulping. Lignin
content would affect the pulp cellulosic hydrolysis, and reduce drop in DP from the cellulose (Hubbell and Ragauskas, 2010).
A mostly negative correlation between lignin contents of pretreated biomass and the accessibility and degradability of cellulase had been found by using cellulase mixture (Boussaid and Saddler, 1999) or cellobiohydrolase (CBH) (Jeoh et al., 2007; Kumar and Wyman, 2009). It has also been argued that the lignin content lower than 5% (g/g) results in a dramatic reduction in cellulose digestibility (Ishizawa et al., 2009). Although the adsorption process has been usually combined in models of enzymatic hydrolysis (Andrić et al., 2010), correlation between the accessibility onto pretreated biomass and degradability of various cellulase components still has not been completely clarified (Gerber et al., 1997; Kumar and Wyman, 2009). Non-productive adsorption between lignin and cellulase is also generally regarded as a major factor to impede the degradability of cellulase (Boussaid and Saddler, 1999; Ishizawa et al., 2009; Várnai et al., 2010). In order to alleviate the non-productive adsorption by lignin, extraneous materials have been brought into the saccharification processes.
4. Additives
Additives are additional materials that do not belong to the original cellulosic-hydrolysis system. The mechanism of additives, including bovine serum albumin (BSA) (Yang and Wyman, 2006), Tween 20 (Zheng et al., 2008), various polyethylene glycols (PEG) (Börjesson et al., 2007; Zhang et al., 2010) and yeast extract, improves cellulases hydrolysis efficiency of lignocellulose.
One of the vital additives of enzymatic hydrolysis is bovine serum albumin (BSA). A capacity for BSA could adsorb even more cellulase, which found adsorption of a substantial amount of cellulase on both lignin and cellulose. Adding BSA with cellulase
resulted in some promise in both rates and yields, and after hydrolysis began also enhanced rates whether pretreated fiber was treated with BSA prior to hydrolysis (Yang and Wyman, 2006). In addition, Tween 20 is also a common additive. The effect of the surfactant Tween 20 on the hydrolysis of different cellulosic fibers was investigated and related to the cellulose fiber structure. It was found that this non-ionic surfactant enhanced the enzymatic saccharification of highly crystalline cellulose (Mizutani et al., 2002). Moreover, polyethylene glycols can raise the hydrolysis yield due to the following two effects, hydrophobic interactions and hydrogen bonds between PEG and lignin. The phenolic hydroxyl group of lignin can interact with the ether oxygen bond (-O-) to form a hydrogen bond (Börjesson et al., 2007; Zhang et al., 2010).
Since yeast extract is commonly present in the simultaneous saccharification and fermentation processes (SSF) (Ballesteros et al,. 2004), its effect on the accessibility and degradability of the crude endoglucanase was also examined. Molecular weight reduction and reducing sugar were monitored for the extents of hydrolysis (Ouyang et al., 2010). The cellulase hydrolysis was related to the degree to which this saccharification was enhanced by the presence by additives which block the cellulases from binding to the lignin. Therefore, more cellulases could participate in lignocellulosic hydrolysis (Ander et al., 2008; Ramírez et al., 2008; Mamma et al., 2009).
(III) Refinig of Pulp Stock 1. PFI
Cellulases were the most efficient enzymes as refining helpers (Gil et al., 2009).
During papermaking, the bleached pulp must be mechanically produced to obtain a certain reduced paper web drainage (defined as freeness) to improve paper web
formation and flexibility (Kamaya, 1996; Mutjé et al., 2005), and the cellulase pretreatment of the pulp could result in noticeable savings in electricity of power, while maintaining the strength properties of the pulp (Bajpai et al., 2006).
The cellulases have effective ability to alter fiber morphology, and could increase drainability (Schopper-Riegler method) and water retention value (WRV) of bleached pulp. They could improve inter-fiber bonding in the refining process. However, the drainability and paper strength results have not always been consistent; also, cellulases are often thought to have a harmful impact on paper properties (Oksanen et al., 2000;
García et al., 2002; Pala et al., 2002; Cadena et al., 2010).
2. Scanning Electron Microscope (SEM)
A scanning electron microscope (SEM) is a type of electron microscope that images a sample by scanning it with a high-energy beam of electrons in a raster scan pattern. The objectives of using SEM were to explore the morphological change on the surface of fiber, to compare at the microscopic level. Visual observation revealed differences in fiber fracture between treated and control samples (Garmaroody et al., 2011; Shi et al., 2011). High resolution SEM analysis of the fiber surface was the methods of choice.
The images should be interpreted in view of possible differences in the fiber fracture mechanism caused by the cellulase treatment (Suchy et al., 2009). In this study, the morphology will be examined by SEM which showed that differences of refined fibers between control and enzyme pretreated.
III Materials and Methods
(I) Microorganism and Growth Measurements
The thermo-alkaline Paenibacillus campinanesis BL 11 used in this study was isolated at 90℃, pH 9 environment in a kraft pulp mill (Ko et al., 2007). Chemicals were obtained from Sigma (St. Louis, USA) or Merck (Darmstadt, Germany). This strain was routinely plated on Luria-Bertani (LB) plate and incubated at 37℃ for 1 day.
(II) Expression of the Recombinant Endoglucanase
One colony of the expression strain transformed by pETCMC2 insertion is inoculated into 2 ml of Luria-Bertani medium containing 100 μg of ampicillin/ml and allowed to grow overnight at 37°C in a rotary shaker. The overnight culture is then transferred to 30 ml of the same medium and grown to an A600 value of 0.4-0.5. Protein productions are induced by the addition of isopropyl-β-D-thio-galactopyranoside (IPTG) to a final concentration of 0.1 mM and grown overnight at 28°C, after which time the cells are harvested by centrifugation, washed, and disrupted by sonication in a phosphate buffered saline (PBS) buffer (20 mM sodium phosphate, 150 mM NaCl, pH 7.4). Clear lysate of the extracts is loaded on a Ni-NTA agarose (QIAgen) volume. The resin is then washed twice with wash buffer (300 mM NaCl, 50 mM NaH2PO4, 25-30 mM imidazole, pH 7.0) and the protein is eluted by adding 200 µl elution buffer (300 mM NaCl, 50 mM NaH2PO4, 100 mM imidazole, pH 7.0).
(III) Fiber Preparation and Chemical Analysis
Norway spruce (Picea abies) wood chips were obtained from the Heiberg Experimental Forest, State University of New York, College of Environmental Science
and Forestry, Tully, New York, USA (Ko et al., 2010a). The kraft spruce pulps obtained by different chemical treatments. The delignification of spruce chips was carried out according to the procedure described by kraft method with 21% and 25% NaOH and sodium sulfite mixed liquor (w/w with respect to O.D. material, sulfide 25 ± 1) and 700 g spruce chips (O.D.), then cooking at constant temperature of 150 min. The liquor to solid ratio was changed and fixed at 4 due to the experimental device used and temperatures 165℃were tested. All the experiments were conducted in a 1 L reactor, in which the heating time to reach the constant temperature was 150 min. Then, the pulp made in the chemical condition with 21% NaOH and sodium sulfite mixed liquor was treated by oxygen delignification. Oxygen delignification was conducted with 1.2%
(w/w) NaOH (O.D.) pulp under 4 kg/cm2 oxygen pressure raised to 99℃ within 60 min, then maintained isothermally for next 15 min (O15). Fully bleached pulps were prepared from oxygen bleached pulps by using a commercial DEDD bleaching sequence.
(IV) DEDD Bleaching
DEDD bleaching (Wong et al., 2001) was carried out using chlorine dioxide (ClO2) that obtained from an industrial source contained 10% active chlorine as ClO2. The first chlorine dioxide stage (D0) was conducted at 10% pulp consistency, 50°C and for 1 h.
The kappa factor is defined as active chlorine multiple (A.C.M.) (Reeve, 1996), which is a predetermined quantity representing the percentage of active chlorine divided by the kappa number of the pulp (Reeve, 1996). The reason for defining the kappa factor is to normalize bleaching agent usage with respect to pulp lignin contents in industrial practice. The reason for defining the active chlorine values is to normalize weight-based usage for various bleaching agents with respect to chlorine molecular weight in
industrial practice. Alkaline extraction stage (E) were performed at 10% consistency, 70°C for 1 h, and with the charge of NaOH equivalent to one half of the initial active chlorine charge. The second chlorine dioxide stages (D1) were carried out at 10%
consistency with 1% ClO2 solution, at pH 4, 70°C for 3 h. The third chlorine dioxide stages (D2) were carried out at 10% consistency with 1% ClO2 solution, at pH 4, 70°C for 3 h.
(V) Characterization of Pulps
After pulp making, the obtained pulps were washed several times through a wire until obtaining a clear filtrate and characterized in terms of yield, kappa number, residual lignin, holocellulose and pentosan. The cooking yield was calculated as the ratio of the weight of O.D. material after washing to that of initial raw material. The residual lignin was determined from both the Klason lignin and the soluble lignin measured by UV absorption of a filtrate specimen at 280 nm (TAPPI method UM 250). The viscosity of pulp (g in mPa.s) dissolved in a cupriethylene-diamine solution was determined according to TAPPI standard (T230 om-99). These values were then converted into degrees of polymerization (DP) thanks to the following relation proposed by Sihtola et al. (1963):
DP= [0.75 (954Log10η – 325)1.105] (1)
The values of both rate constant (k) and LODP have been calculated with the aid of a non-linear curve fitting software (Calvini, 2005):
(1/DP - 1/DP0) = (1/LODP-1/DP0) × (1-e-kt) (2)
(VI) Cellulase Activity
The cellulase was tested in a fixed volume, containing diluted enzyme dosage, 4%
CMC (Carboxymethyl cellulose, pH 7) in the Tris buffer (0.5 M, pH 7) contained 100 mM Tris buffer, 20 mM CaCl2 and 0.04% (v/v) Tween 20 (Bio Basic). The tubes were incubated for 20 min at 40◦C, and the reaction was terminatedly added to dinitrosalicylic acid (DNSA) including 1% (w/v) dinitrosalicylic acid (Sigma), and 0.4 M sodium hydroxide solution (10%, v/v) and 30% (w/v) potassium sodium tartrate tetrahydrate (Sigma), and the tubes were boiled for 10 min, then cooling. The absorbance then was measured at 540 nm (Miller, 1959; König et al., 2002).
(VII) Enzymatic Hydrolysis of Pulp
The crude endoglucanase from P. campinasensis BL 11 were used to hydrolyze cellulose in spruce kraft pulp. Enzymes were added to make 5, 10, 20 and 50 U/g pulp.
Experiments were carried out in heat-resisting plastic bags containing 1 g of kraft pulp (O.D.) and a total liquid volume of 10 mL, with pulp consistence 10% (i.e., cellulase diluted in Tris buffer of pH 7) for 0, 1, 2, 4, 6, 8 and 24 h at 40◦C in a thermostatic water bath. At periodic time intervals, glucose and DP were measured.
(VIII) Additive of Yeast Extract
0.25 g yeast extract (Fluka) was added to 5 mL 50 mM tris buffer, pH 7, to make a 5
% solution, and then kraft pulps (12.43% and 1.24% lignin content) at 20% (w/v) were added. After mixing completely, the final pulps consistence was 10% and yeast extract concentration was 25% to the pulps (w/w). The hydrolysis steps were same to foregoing method “Enzymatic hydrolysis of pulp”.
(IX) Endoglucanase Accessibility and Digestibility
To determine enzyme accessibility, adsorption on three pulps was performed in 2 mL 0.1 M sodium acetate, 20 mM CaCl2, 0.04 % (w/v) Tween 20, pH 6 at 4℃ to avoid hydrolysis. The substrate concentration was 0.1% (w/v), with the dosages for 5, 10, 15, 20, 30, 40, 50 mg enzyme per gram O.D. pulp. The mixtures were loaded in 2.5 mL centrifuge tubes; then turned end-over-end on a home-made rotator. Tubes in triplicates were removed over 1 h and then centrifuged at 5,000 rpm. The adsorbed enzymes were determined by the difference between the amounts of initially added protein and free protein in the supernatant assayed by the Bradford method.
Kumar and Wyman (2009) indicated that adsorption parameters (maximum adsorption capacity [σ] and equilibrium constant [Kd]) were determined by non-linear regression of the adsorption data to the Langmuir expression, using Sigma Plot software (ver 10.0, SPSS Inc., Chicago):
[CE] = (σ × [St] × [Ef]) / (Kd + [Ef]) (3)
where [CE] is the amount of adsorbed enzyme in mg/mL, [Ef] is the free enzyme concentration in mg/mL, σ is the maximum adsorption capacity in mg/mg substrate, [St] is the substrate concentration in mg/mL, and Kd is the equilibrium constant = [C][E]/[CE] in mg of enzyme/mL (Kumar and Wyman, 2009).
To determine enzyme adsorption-desorption kinetics during hydrolysis, the dosage at 6 mg enzyme per gram O.D. pulp was chosen with the above solution at 40℃. Tubes in triplicates were removed over 1 h to 48 h to quantify the adsorbed enzymes. The digestibility was measured by the change of intrinsic viscosity and the release of reducing sugars. Intrinsic viscosities were analyzed following ISO 5351: 2004 standard
method. The released reducing sugars were measured by the dinitrosalicylic acid (DNSA) method.
(X) Enzyme Treatment
The pulp was treated with 1 IU, 2.5 IU and 5 IU of cellulases and endoglucanases per gram of oven-dry (O.D.) pulp. This treatment was carried out with a pulp suspension having a consistency of 10% at 40℃, in beakers, with continuous mechanical agitation.
The following reaction times were tested for 1 h. The pH was adjusted to 7 with Tris buffer for cellulases treatment.
(XI) Pulp Refining
The fully bleached spruce pulp was place in a polyethylene bag at 10% consistency at pH 7.0, 40℃ for 1 h, simulating actual mill operating conditions. Constant pH values were monitored for the filtrates of the pulp-enzyme mixtures before and after enzyme treatment. Crude 38-kDa Cel-BL11 cellulase directly from cell lysate after the disruption of E. coli was applied at levels of 2.5 IU and 5 IU per gram of oven-dried pulp (O.D.). After treatment, the reaction mixtures were washed, diluted up to 400 mL with cold water and subjected to freeness tests. The recovered pulp was beaten at 10%
consistency in a PFI (Papir-og fiberinstituttet) mill, following ISO 5264-2 (ISO, 2002), homogenized in a disintegrator at 1.2% consistency for 2 min and subjected to freeness measurements.
Freeness values of pulp were measured by TAPPI method T227-om04 (TAPPI, 2004) and expressed as Canadian Standard Freeness (CSF) values. The fiber hydratation was determined using the water retention value (WRV), according to the method described by Silvy et al. (1968). This method consists of the soaking of the pulp samples in water
with further centrifugation, and the WRV was calculated from the following equation:
WRV [%] = [(Ww–Wd)/Wd]×100, where Ww is the mass of the wet sample after centrifugation, and the Wd is that after wet sample drying at 105℃ to constant weight.
Handsheets of 75 ± 2 g/m2 grammage were prepared on Rapid-Köhten equipment according to ISO 5331 and tested mechanically in accordance with the following standards: tensile index, ISO 1924; burst index, ISO 2758; tear index, ISO 1974; folding endurance, ISO 5626. Experimental errors were calculated as prescribed by the respective standards.
(XII) Morfi
Morfi (TECHPAP, 10 rue de Mayencin 38400 Saint Martin d’Hères) was used in this experiment to analyzed the morphology of fibers. 30 mg pulp samples were put in 1 L plastic beacker with water to 0.3% consistency, and to analyze using Morfi. Morfi provides the distribution of fines area, fines length, fiber distribution, and fiber width.
Fiber length was 200-10,000 μm, fiber width was 5-75 μm, fines length was defined shorter than 200 μm, and width of fines was defined as shorter than 5 μm.
(XIII) Scanning Electron Microscope (SEM)
The sample of spruce fibers before tested would be formed very thin papers; then, the fibers were coated with gold to provide electrical conductivity. SEM was used to analyze fiber morphology using and acceleration voltage of 20 kV. The tested fibers were randomly chosen, and the better image has a whole fiber in the picture center.
Their dimensions were measured using software (Shi et al., 2011).
IV Results and Discussion
(I) Impacts of Lignin Contents and Yeast Extract Addition on the Interaction between Spruce Pulps and Crude Recombinant Paenibacillus Endoglucanase
Chemical compositions of the pulps were listed in Table 1. Pulp samples were coded with their total lignin contents as follows: LIG 12.4, LIG 8.29, LIG 6.22, and LIG 1.24.
The result of statistics showed the chemical properties of four pulps have some difference, analyzed by Scheffé's method.
Table 1. Chemical properties of four spruce pulps.
Chemical properties
Sample code
LIG 12.4 LIG 8.3 LIG 6.2 LIG 1.2 Pulp lignin content (%) 12.43 ± 0.13 8.29 ± 0.14 6.22 ± 0.12 1.24 ± 0.03
Holocellulose (%) 83.69 ± 0.10 89.50 ± 0.08 93.45 ± 0.05 98.72 ± 0.01 α-cellulose (%) 69.81 ± 0.09 70.54 ± 0.09 78.71 ± 0.02 83.73 ± 0.04 Pentosan (%) 8.83 ± 0.11 8.53 ± 0.13 6.65 ± 0.09 5.98 ± 0.08
DP 3876 ± 85 3226 ± 63 2462 ±37 2135± 26
1. Effects on the Accessibility of Proteins
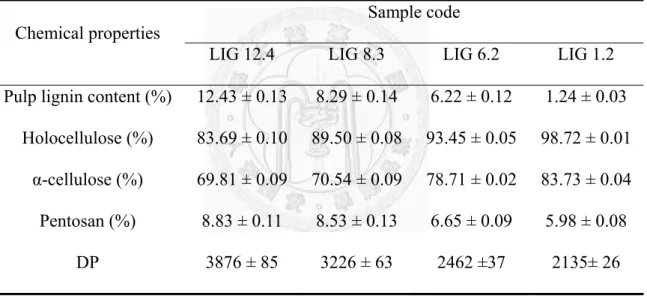
The effect of lignin contents and added yeast extract addition on the accessibility of proteins (crude endoglucanase) onto to the four spruce pulps at 4oC was shown in Fig. 1.
The results at 4oC demonstrated that the pulp with more lignin contents accommodated more proteins as shown Fig. 1A. The above observation contradicts the findings that biomass with more cellulose favored cellulase adsorption, according to several studies using purified cellulases (Chernoglazov et al., 1988), mixed cellulase (Ooshima et al.,
1990; Boussaid and Saddler, 1999), purified CBH (Ishizawa et al., 2009), and commercial endoglucanase (Ko et al., 2011). The above discrepancy could be explained:
CMCase activity of the purified and crude recombinant Paenibacillus endoglucanase was measured at 250 IU/mg (Ko et al., 2010b) and 35.2 IU/mg (this study) at pH 7 and 40oC. So it could be assumed that the mass of irrelevant proteins were 6.1 times more than the purified endoglucanase. In contrast to the behaviors of mostly pure cellulases used in the above studies, Fig. 1A described the adsorption behavior of the mixture of less endoglucanase and much more irrelevant proteins derived from cell rupture. Figure 1A showed that the interaction between irrelevant protein and overall pulp surfaces dominated those between pure endoglucanase and cellulose surface.
The addition of yeast extract facilitated even more protein adsorption onto pulps with more lignin contents, as shown in Fig. 1B (Yang and Wyman, 2006). Yeast extract, composted of amino acid and peptides, also could be regarded as of a mixture of electrolyte and polyelectrolyte. Hence, the above finding was consistent to the effect on increasing protein adsorption by the addition of bovine serum albumin (BSA, also a polylelectolyte) (Yang and Wyman, 2006) and polyethylene glycol (PEG) (Ouyang et al., 2010).
0.00 0.02 0.04 0.06 0
2 4 6 8 10 12
0.00 0.02 0.04 0.06 Adsorbed protein (mg g-1 substrate)
Free protein in the supernatant (mg mL-1)
A B
Fig. 1. Effect of yeast extract addition on the accessibility of protein (crude recombinant Paenibacillus endoglucanase) on four spruce pulps at 4oC. Panel A: Controls with closed symbols. Panel B: Yeast extract with open symbols. Legends of pulps with different lignin contents: LIG 12.4 (★), LIG 8.3 (■), LIG 6.2 (▲), LIG 1.2 (●).
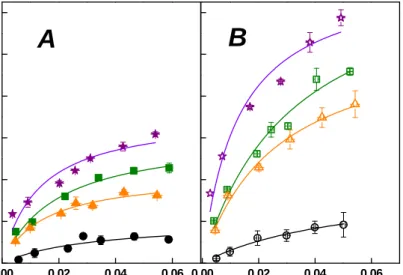
The adsorption parameters were also estimated by nonlinear regression of adsorption data for the protein (very crude endoglucanase) and four spruce pulps with or without yeast extract addition action, using the Langmuir equation. As illustrated in Table 2, the adsorption data of the protein mostly followed the Langmuir relationship. The results of Table 2 showed that the addition of yeast extract significantly increased the maximum adsorption capacities of the protein onto all four pulps. The maximum adsorption capacities [σ] in Table 3 were much lower than published values of treated biomass by leading treatments (Kumar and Wyman, 2009), although in same order of magnitude.
The above observation might be due to much smaller specific surface areas of complete softwood fibers used in the study, when compared with much smaller fibers obtained by leading treatments (Kumar and Wyman, 2009). In general, the maximum adsorption capacities [σ] decreased with pulps of decreasing lignin contents. The above trend matched increasing equilibrium constants [Kd] with pulps of decreasing lignin contents.
The discrepancy between parameters of LIG 8.3 and LIG 6.2 might due to two competing interactions for irrelevant protein/overall pulp surfaces and pure endoglucanase/cellulose surface. Addition of yeast extract increased the maximum adsorption capacities [σ] and equilibrium constants [Kd] of all four pulps; the interaction between amino acid/peptides and pulp surface might create more available binding sites for the proteins.
Table 2. Effects of pulp lignin contents and yeast extract addition on the maximum adsorption capacity (σ) and equilibrium constants (Kd) of the protein onto four spruce pulps.
Sample code
σ Kd R2
Control Y. E. Control Y. E. Control Y. E.
LIG 12.4 7.14 13.93 0.0129 0.0128 0.9002 0.9624 LIG 8.3 6.19 14.87 0.0195 0.0325 0.9790 0.9760 LIG 6.2 4.29 11.20 0.0150 0.0265 0.9720 0.9922 LIG 1.2 2.02 3.82 0.0317 0.0519 0.8275 0.9712
Table 3. Effects of pulp lignin contents and dosages of endoglucanase applied on degradation rate constants (k) and leveling-off degree of polymerization (LODP) during hydrolysis of four spruce pulps.
Sample code
Enzyme dosage (IU/g. o. d. p.)
5 10 20 50
k LODP k LODP k LODP k LODP
LIG 12.4 0.2132 3192 0.2514 2783 0.2958 2624 0.3250 2410 LIG 8.3 0.3141 2147 0.3287 2148 0.3736 2069 0.3856 1982 LIG 6.2 0.2518 1996 0.2913 1949 0.3179 1810 0.3352 1550 LIG 1.2 0.1815 1489 0.1841 1441 0.1863 1392 0.1940 1297
2. Effects on the Preferential Adsorption of Irrelevant Proteins
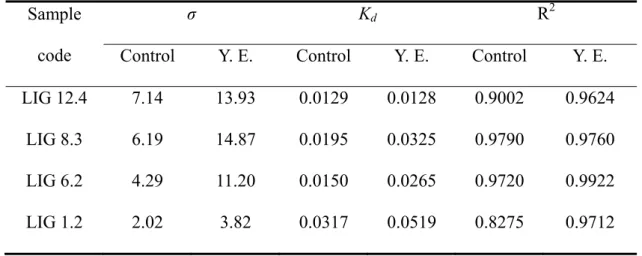
To further verify the different adsorption of endoglucanase and of irrelevant proteins, the specific endoglucanase (CMCase) activities of supernatant portion during adsorption were separately measured at pH 7 and 40oC. Yeast extract couldn’t interfere with the Bradford assay, or with the DNSA assay. Since the CMCase activity of crude recombinant Paenibacillus endoglucanase was measured at 35.2 IU/mg at pH 7 and 40oC, the preferential adsorption of irrelevant proteins could be indicated by the more than 35.2 IU/mg activity values of proteins. The effect of yeast extract addition on specific endoglucanase activity of proteins in supernatants is shown in Fig. 2. Figure 2A showed specific endoglucanase activity of proteins in supernatants without yeast extract addition. With sample LIG 12.4, the specific endoglucanase activities of proteins in supernatants were higher than 35.2 IU/mg. The above trend was more so at lower total proteins loading. However, an opposite trend was shown for LIG 1.2: the specific endoglucanase activity of proteins in supernatants was slightly lower than 35.2 IU/mg at low protein loading. Interestingly, the specific endoglucanase activities of proteins in supernatants for all pulps more or less converged at around 35.2 IU/mg. It could be deduced that the interaction between cellulose and endoglucanase dominated for the low lignin pulp (LIG 1.2) at low protein loading. On the other hand, the interaction between pulp surface and irrelevant proteins dominated for the high lignin pulps at low protein loadings. Protein concentrations were quite dilute at low protein loadings, and the collisions between endoglucanase/irrelevant proteins and pulp surface were fewer. In the above case, not every collision led to successful adsorption. However, the discretion between endoglucanase/irrelevant proteins was diminished at higher protein loading, since there were too many collisions between both endoglucanase/irrelevant proteins and pulp surfaces.
0.01 0.02 0.03 0.04 0.05 0.06 30
35 40 45 50 55 60 65
0.01 0.02 0.03 0.04 0.05 0.06
A
Specific endoglucanase activity of protein in supernatant(IU/mg)
Free protein content in the supernatant (mg/mL)
B
Fig. 2. Effect of yeast extract addition on specific endoglucanase activity of proteins in supernatants. Panel A: Controls with closed symbols. Panel B: Yeast extract with open symbols. Legends of pulps with different lignin contents: LIG 12.4 (★), LIG 8.3 (■), LIG 6.2 (▲), LIG 1.2 (●).
Figure 2B shows the specific endoglucanase activity of proteins in supernatants with yeast extract addition. Since a lot of amino acid and peptide (with much smaller molecular weights) was brought by yeast extract addition, the surfaces of all pulps were further attached by yeast extract to facilitate the adsorption by irrelevant proteins. Again, it showed that the specific endoglucanase activities of proteins in supernatants converged at different levels. The final levels matched the lignin contents of each pulp sample. Since the higher specific endoglucanase activities of proteins in supernatants were exhibited, it could be assumed that more endoglucanase were still un-adsorbed at higher lignin loading at the presence of yeast extract.
3. Effects of Pulp Lignin Contents on Hydrolysis
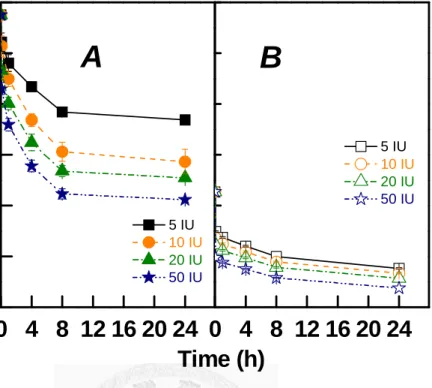
Effects of pulp lignin contents on the reduction of pulp DP by crude recombinant Paenibacillus endoglucanase at different dosages during hydrolysis at pH 7, 40oC are shown in Fig. 3 for samples LIG 12.4 and LIG 1.2. Table 3 lists degradation rate constants (k) and leveling-off degree of polymerization (LODP) during hydrolysis obtained by regression with Eq. 2. As indicated by Table 1, DP values of LIG 12.4 and LIG 1.2 were 3876 and 2135 prior to hydrolysis. 5 IU and 50 IU p. g. o. p. degraded DP of LIG 12.4 to LODP at 3192 and 2410, which is equivalent to 82.3 % and 62.1 % of original LODP values. 5 IU and 50 IU p. g. o. d. p. degraded DP of LIG 1.2 to LODP at 1489 and 1297, equivalent to 69.7 % and 60.7 % of original LODP values. Higher lignin content of LIG 12.4 made it more difficult to be degraded by Paenibacillus endoglucanase than LIG 1.24. Since there was no CBH and β-glucosidase in this system, LODP could not be reduced further.
0 4 8 12 16 20 24 1000
1500 2000 2500 3000 3500 4000
0 4 8 12 16 20 24
5 IU 10 IU 20 IU 50 IU
DP
A B
5 IU 10 IU 20 IU 50 IU
Time (h)
Fig. 3. Effect of pulp lignin contents on the reduction of pulp DP by crude recombinant Paenibacillus endoglucanase at different dosages during hydrolysis at pH 7, 40oC. Panel A: LIG 12.4. Panel B: LIG 1.2.
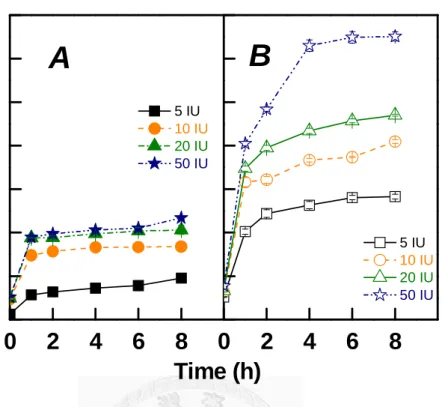
Figure 4 shows the effect of pulp lignin contents on the release of reducing sugars during hydrolysis at pH 7, 40oC. Two different patterns of reducing sugar release are shown by comparison of Figs. 4A and 4B. Since the total crude Paenibacillus endoglucanase was added at 0.14 mg p. g. o. d. p. during hydrolysis, the proteins was loaded in the range of very low concentration in Fig. 2. So it could be assumed that the scenario of Fig. 4A was “minimized un-productive adsorption by lignin, with more un-adsorbed endoglucanase in solution”. On the other hand, the scenario of Fig. 4B was
“maximized productive endoglucanase adsorption by cellulose, with more irrelevant proteins in solution”. So, it could be explained that there was a similar difference between the ranges of DP reduction of LIG 12.4 and of 1.2. More reducing sugars releases shown in Fig. 4B suggests that the degraded cellulose fragments were further degraded, and more cellobiose might form to retard further hydrolysis by crude Paenibacillus endoglucanase (Ko et al. 2010b).
0 2 4 6 8 0
1 2 3 4 5 6 7
0 2 4 6 8
A
R e duc in g s u ga r ( m g/g)
Time (h)
5 IU 10 IU 20 IU 50 IU
B
5 IU 10 IU 20 IU 50 IU
Fig. 4. Effect of pulp lignin contents on the release of reducing sugars by crude recombinant Paenibacillus endoglucanase at different dosages during hydrolysis at pH 7, 40oC. Panel A: LIG 12.4. Panel B: LIG 1.2.
Effects of pulp lignin contents and dosages of endoglucanase applied on degradation rate constants (k) and leveling-off degree of polymerization (LODP) during hydrolysis of four spruce pulps are listed in Table 3. Relations between k, LODP values and enzyme dosages were as expected, but k values of degradations for LIG 8.3 were the highest for all four dosages of endoglucanase applied. The four complicating factors in this system contributed to the above observation: endoglucanase, irrelevant proteins, lignin and celluloses (or lignin-associated microfibril structures). The above observation also suggested that higher degree of delignification might not benefit additionally for a complicated system, such as simultaneous saccharification and fermentation. From a practical perspective, the purest enzymes and the lowest lignin contents of lignocellulosic materials may be not mandatory for a most cost-effective bio-ethanol conversion process.
4. Effects of Yeast Extract Addition on Hydrolysis
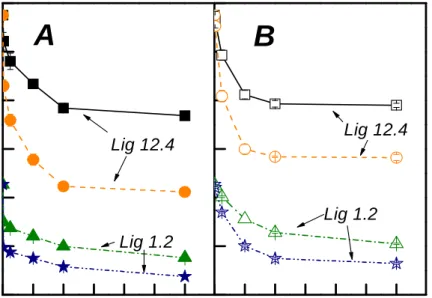
During hydrolysis, effects of yeast extract addition on the reduction of pulp DP and reducing sugar release are shown in Figs. 5 and 6. As shown in Fig. 5, trends of pulp DP reduction were not altered by yeast extract addition.
0 4 8 12 16 20 24 1000
1500 2000 2500 3000 3500 4000
0 4 8 12 16 20 24
Lig 1.2
DP
Time (h)
A
Lig 12.4
Lig 1.2 Lig 12.4
B
Fig. 5. Effect of yeast extract addition on the reduction of pulp DP during hydrolysis at pH 7, 40oC. Panel A: Controls with closed symbols. Panel B: Yeast extract with open symbols. Legends: LIG 12.4 with 5 IU p. o. d. g. p. (■), LIG 12.4 with 50 IU p. o. d. g.
p. (●), LIG 1.2 with 5 IU p. o. d. g. p. (▲), LIG 1.2 with 50 IU p. o. d. g. p. (★).
0 2 4 6 8
0 1 2 3 4 5 6 7
0 2 4 6 8
Lig 12.4 Lig 1.2
R e duc ing s uga r ( m g /g)
Time (h)
A
Lig 12.4 Lig 1.2
B
Fig. 6. Effect of yeast extract addition on the release of reducing sugars during hydrolysis at pH 7, 40oC. Panel A: Controls with closed symbols. Panel B: Yeast extract with open symbols. Legends: LIG 12.4 with 5 IU p. o. d. g. p. (■), LIG 12.4 with 50 IU p. o. d. g. p. (●), LIG 1.2 with 5 IU p. o. d. g. p. (▲) LIG 1.2 with 50 IU p. o. d. g. p.
(★).
Although there was more un-adsorbed endoglucanase in solution (as shown by Fig. 2) by yeast extract addition, the reduced non-productive adsorption by pulp lignin was negated by the presence of yeast extract to interfere with endoglucanase action. The excess yeast extract presented might reduce the collision frequencies between endoglucanase and fiber surfaces in the later stages of hydrolysis.
Figure 6 shows that the trends of reducing sugar release were altered by yeast extract addition. The excess yeast extract present in this system might compete with hydrolyzed cello-oligomers to interact with endoglucanase; hence the reducing sugar releases was decreased. The above finding was more evident in the case of high enzyme dosages with low lignin pulp, LIG 1.2. The degradation rate constants (k) and leveling-off degree of polymerization (LODP) obtained by regression with Eq. 2 of spruce pulps LIG 12.4 and LIG 1.2 are summarized in Table 4.
Table 4 shows that the yeast extract addition increased degradation rate constants (k) for high lignin and high enzyme dosage cases. Comparison with Fig. 2 suggests that more un-adsorbed endoglucanase contributed to the above observation. In Table 4, the degradation rate constant (k) of the low lignin pulp/low enzyme dosage case was not increased by yeast extract addition. The above finding also corresponds well with the only productive cellulase/cellulose adsorption case shown by Fig. 2. The addition of yeast extracts caused slightly higher leveling-off degree of polymerization (LODP) values. Despite the presence of irrelevant protein and yeast extract, the basic mechanism of endoglucanase action remained. The excess yeast extract presented might reduce the collision frequencies between endoglucanase and fiber surfaces at the later stages of hydrolysis.
Table 4. Effects of yeast extract addition and dosages of endoglucanase applied on degradation rate constants (k) and leveling-off degree of polymerization (LODP) during hydrolysis of spruce pulps LIG 12.4 and LIG 1.2.
Sample code
5 IU/g. o. d. p. 50 IU/g. o. d. p.
k LODP k LODP
LIG 12.4 0.4739 2949 0.5987 2412
LIG 1.2 0.1683 1517 0.2886 1323
(II) Effect of Crude Recombinant Paenibacillus Endoglucanase on Refining of Spruce kraft pulp
1. Effect on the Drainability and Fiber Hydration
The effect of cellulases treatments on the pulp and paper properties has been evaluated. The spruce pulp was pre-treated with endoglucanase produced by our laboratory, followed by refining using different PFI revolutions. Pulp and paper properties were determined and compared to those of control samples untreated with cellulase.
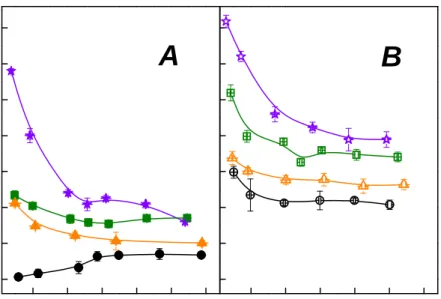
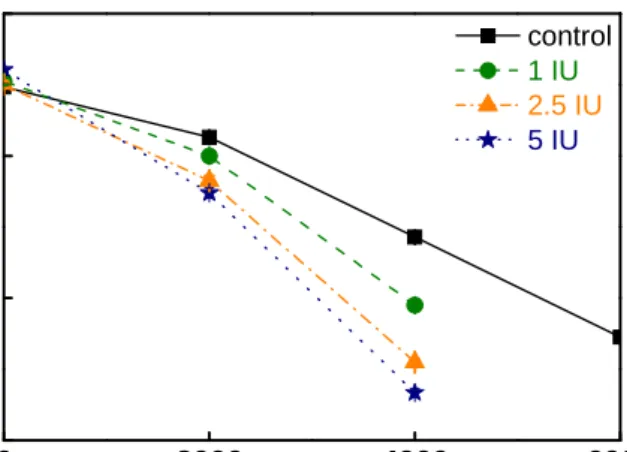
First, the study discussed how the endoglucanase pre-treatment affect the relationship between water and fiber of spruce pulp. The treatment with endoglucanase gave rise to an improvement in pulp behavior involving the refining process, in terms of pulp drainability (CSF) and of fiber hydration (WRV). Figure 7 showed that a decrease of the freeness for the pulps submitted to the endoglucanase treatment relatively to the untreated ones. This effect was more remarkable for refined pulp than unrefined pulp, along with the revolution raising. This index is mainly influenced by fiber external fibrillation and by the amount of fine elements.
Comparison of properties of control and enzyme-treated pulps, unrefined and refined for the same PFI revolutions, showed an increase WRV, which characterizes the water absorption inside the fiber wall, for enzyme-treated pulps (Fig. 8), with similar trend of freeness. Cellulase pretreatment led to an important increase in the WRV of refined sample.
0 2000 4000 6000 200
400 600
800 control
1 IU 2.5 IU 5 IU
Freeness (mL)
Revolutions (rpm)
Fig. 7. Effect of PFI-refining revolutions with control (0 IU), 1 IU, 2.5 IU and 5 IU endoglucanase dosage on freeness.
0 2000 4000 6000
100 120 140 160
control 1 IU 2.5 IU 5 IU
Revolutions (rpm)
WRV (%)
Fig. 8. Water retention value versus PFI-refining revolutions with control (0 IU), 1 IU, 2.5 IU and 5 IU endoglucanase dosage.
2. Pulp Biorefining and Effect on Pulp Properties
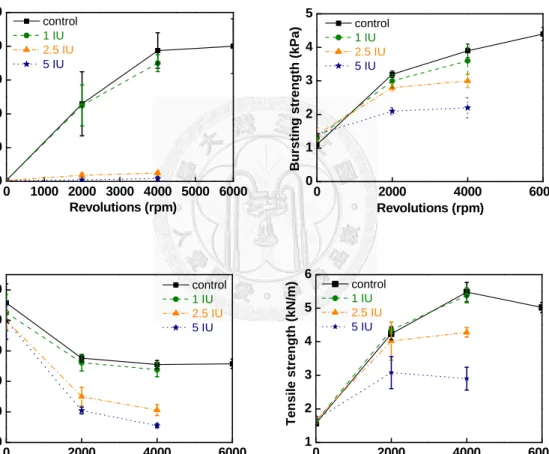
Figure 9 showed pulp properties. It was a pity that the four index losses were observed upon cellulase treatment; in other words, the spruce pulp decreased its physical-mechanical properties by the enzyme. It meant that the cellulase was too effective to degrade the long fiber of spruce, and the study could not ignore the negative influence of enzyme pretreatment on pulp strength.
0 1000 2000 3000 4000 5000 6000 0
400 800 1200 1600 2000
Revolutions (rpm) control
1 IU 2.5 IU 5 IU
Folding (times)
0 2000 4000 6000
0 1 2 3 4 5
Revolutions (rpm) control
1 IU 2.5 IU 5 IU
Bursting strength (kPa)
0 2000 4000 6000
200 400 600 800 1000
1200 control
1 IU 2.5 IU 5 IU
Revolutions (rpm)
Tearing strength (mN)
0 2000 4000 6000
1 2 3 4 5
6 control
1 IU 2.5 IU 5 IU
Revolutions (rpm)
Tensile strength (kN/m)
Fig. 9. Pulp strength of control (0 IU), 1 IU, 2.5 IU and 5 IU cellulase pretreatment of different PFI-resolutions.
3. Fibers of Morfi analysis
Figure 10 to Figure 15 showed that the Morfi (scale in micrometer) displayed the result of refined and unrefined pulp fibers with enzyme pretreatment or without any cellulase treated (control). Mean fiber width was affected by cellulase treatments, and fiber width increased with enzyme pretreatment (Fig. 10). The raising was enhanced when the PFI-resolutions increasing. It said that the refined pulp treated with cellulase would have wider fiber. Mean fines length (Fig. 13) also increased due to that cellulase hydrolyzed most small fines. Moreover, the fiber distribution of kink angle (Fig. 12) declined after cellulase treating. The Morfi also detected more broken ends and higher distribution of microfiber length of the fiber with cellulase pre-treating and refined.
5~17 17~27 27~47 47~67 67~
0 10 20 30 40 50
%
Fiber width (um)
control 0 control 2000 control 4000
5 IU 0 5 IU 2000 5 IU 4000
Fig. 10. Morfi fiber width of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions.
0~24 24~48 48~71 71~95 -- 0
20 40 60 80 100
Fiber curl (%)
%
control 0 control 2000 control 4000
5 IU 0 5 IU 2000 5 IU 4000
Fig. 11. Morfi fiber curl of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions.
0~30 30~55 55~12 120~140 140~
0 10 20 30 40 50
Kink angle
%
control 0 control 2000 control 4000
5 IU 0 5 IU 2000 5 IU 4000
Fig. 12. Morfi kink angle of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions.
1 0 ~ 2 02 0 ~ 3 0 3 0 ~ 4 04 0 ~ 5 0 5 0 ~ 6 0 6 0 ~ 7 07 0 ~ 8 0 8 0 ~ 9 09 0 ~ 1 0 0 1 0 0 ~
0 1 0 2 0 3 0 4 0
%
fin e s le n g th (u m )
c o n tro l 0 c o n tro l 2 0 0 0 c o n tro l 4 0 0 0
5 IU 0 5 IU 2 0 0 0 5 IU 4 0 0 0
Fig. 13. Morfi fines length of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions.
1 0 0 ~ 2 0 0 2 0 0 ~ 3 0 0 3 0 0 ~ 5 0 0 5 0 0 ~ 8 0 0 8 0 0 ~ 1 2 0 01 2 0 0 ~ 1 7 0 01 7 0 0 ~ 2 3 0 02 3 0 0 ~ 3 0 0 03 0 0 0 ~ 3 8 0 0 3 8 0 0 ~
0 1 0 2 0 3 0 4 0
F in e s a re a (u m2)
%
c o n tro l 0 c o n tro l 2 0 0 0 c o n tro l 4 0 0 0
5 IU 0 5 IU 2 0 0 0 5 IU 4 0 0 0
Fig. 14. Morfi fines area of control and 5 IU cellulase pretreatment of 0, 2000 and 4000 PFI-resolutions.