國立台東大學生命科學研究所 碩士論文
指導教授:廖尉岑 博士
磁奈米對大腸桿菌蛋白質體的影響 之前期研究
A preliminary study of the influence of magnetic nanoparticles on the proteome of Escherichia coli.
研 究 生:邱洋斌 撰
中華民國九十九年七月
國立台東大學生命科學研究所 碩士論文
磁奈米對大腸桿菌蛋白質體的影響 之前期研究
A preliminary study of the influence of magnetic nanoparticles on the proteome of Escherichia coli.
研 究 生:邱洋斌 撰 指導教授:廖尉岑 博士
中華民國九十九年七月
致 謝
兩年的光陰很快地流逝,在台東大學就讀生命科學所的過程中,
有辛苦也有歡樂,這一切的點點滴滴都值得我再三回味。碩一的時候 總是讓自己修了很多課,每天過得很充實,尤其是和同學們一起相處、
一起學習的感覺,真的很好,然而坐在台下聽教授們上課,更是一種 享受。在實驗操作的技能訓練上,全都要歸功於廖尉岑教授,沒有他 的傳授,我無法得到許多有用的實驗操作技術,同時他也是我的心靈 導師,在我實驗一直失敗的階段不斷給我鼓勵與打氣,讓我有繼續前 進的動力,最後的論文撰寫與修正也因廖教授的從旁協助才得以順利 完成。實驗技術的成熟與部分實驗圖檔製作,也仰賴台東大學應用科 學系張順龍學弟的幫忙,而質譜儀的前置作業準備與操作能力的培養,
要感謝慈濟大學的胡安仁教授、賴姮江同學以及小布學弟的協助,尤
其最要感謝的是在慈濟大學口試當天胡安仁教授以及賴孟君教授給
我的指教,讓論文的內容更加完整與充實。系辦的張惠嵐小姐在論文
完成階段給我很多的協助,我也要特別地感謝她。還有很多在我就讀
台東大學碩士階段給我很多幫助的人,在此一併感謝你們!
摘 要
本研究分析磁奈米對大腸桿菌細胞之蛋白質體的影響。我們使用 二維蛋白質電泳法(包括等電焦電泳(IEF) 及膠體電泳(SDS-PAGE))
,先挖取二維膠片上的蛋白質,接著再用質譜儀(MALTI-TOF)測定,
找出與磁奈米相關的細胞蛋白質表現。磁奈米對大腸桿菌 BCRC11509 的蛋白質表現沒有很大的影響,也就是磁奈米濃度在 100μg/ml 以下 時,不具有細胞毒性。電穿孔單獨或與磁奈米合併使用對大腸桿菌 DH10B 的蛋白質表現沒有很大的影響,蛋白質量的改變皆在兩倍以 內。本實驗支持磁奈米不具細菌毒性,故在未來生物醫學上可以有很 大的應用。
關鍵詞:磁奈米、生物資訊分析、蛋白質體學
Abstract
This study tries to find the influence of magnetic nanoparticles on the proteome of Escherichia coli. We use two-dimensional protein electrophoresis (including isoelectric focusing electrophoresis (IEF) and gel electrophoresis (SDS-PAGE)), first dig the protein on the two-dimensional gel, and then using mass spectrometry (MALTI-TOF) to identify the magnetic nano-related protein expression. Magnetic nanoparticles do not have much impact on the protein expression of E.
coli BCRC11509, that is, when the concentration of magnetic nanoparticles below 100μg/ml, there is no cytotoxicity. Electroporation alone or in combination with magnetic nanoparticles on E. coli DH10B for protein expression is not greatly affected, protein changes are within two fold. Our study supports the biomedical applications of magnetic nanoparticles in the future.
Keywords:Magnetic nanoparticles、Bioinformatics、
Proteomic technologies
目錄
中文摘要 i 英文摘要 ii 圖目錄 vii 第一章 緒論
I 、奈米顆粒與磁奈米
壹、奈米科技的簡介 ………1 貳、奈米材料的性質 ………3 參、磁奈米的特性及生醫應用 ………5 Ⅱ、基因體、生物資訊與蛋白質體學
壹、基因體 ………10 貳、生物資訊 ………11 參、蛋白質體學研究 ………11 Ⅲ、研究動機與實驗目的
壹、研究動機 ………14 貳、研究目的 ………14 第二章 材料與方法
Ⅰ、實驗材料
壹、大腸桿菌 ………15
貳、化學試劑 ………15
參、儀器設備 ………16
Ⅱ、實驗方法 壹、菌株的培養 ………17
貳、磁奈米粒子的製備 ………17
參、磁奈米對大腸桿菌的毒性測試 ………18
肆、電穿孔實驗 ………18
伍、分泌性蛋白質收集 ………19
陸、蛋白質定量(BCA) ………19
柒、顯微鏡觀察 ………19
捌、一維蛋白質電泳分析 ………20
玖、二維蛋白質電泳分析 ………21
拾、質譜分析 ………24
拾壹、生物資訊分析 ………26
第三章 結果 壹、磁奈米的製備 ………31
貳、顯微鏡觀察 ………31
參、磁奈米對大腸桿菌的毒性 ………31
肆、電穿孔對大腸桿菌的毒性 ………32
伍、蛋白質定量 ………32
陸、磁奈米對大腸桿菌蛋白質的影響:一維蛋白質電泳分析 ………33
柒、磁奈米對大腸桿菌蛋白質的影響:二維蛋白質電泳分析 ………33
捌、質譜及生物資訊分析 ………34
第四章 討論 壹、磁奈米的製備 ………36
貳、顯微鏡觀察 ………36
參、磁奈米對大腸桿菌的毒性 ………36
肆、電穿孔對大腸桿菌的毒性 ………37
伍、蛋白質定量 ………37
陸、磁奈米對大腸桿菌蛋白質的影響:一維蛋白質電泳分析 ………37
柒、磁奈米對大腸桿菌蛋白質的影響:二維蛋白質電泳分析 ………38
捌、質譜及生物資訊分析 ………38
第五章 結論 ………39
參考文獻 ………49
附錄一 磁奈米之生物反應路徑分析 ………56
附錄二 質譜分析結果 ………61
圖目錄
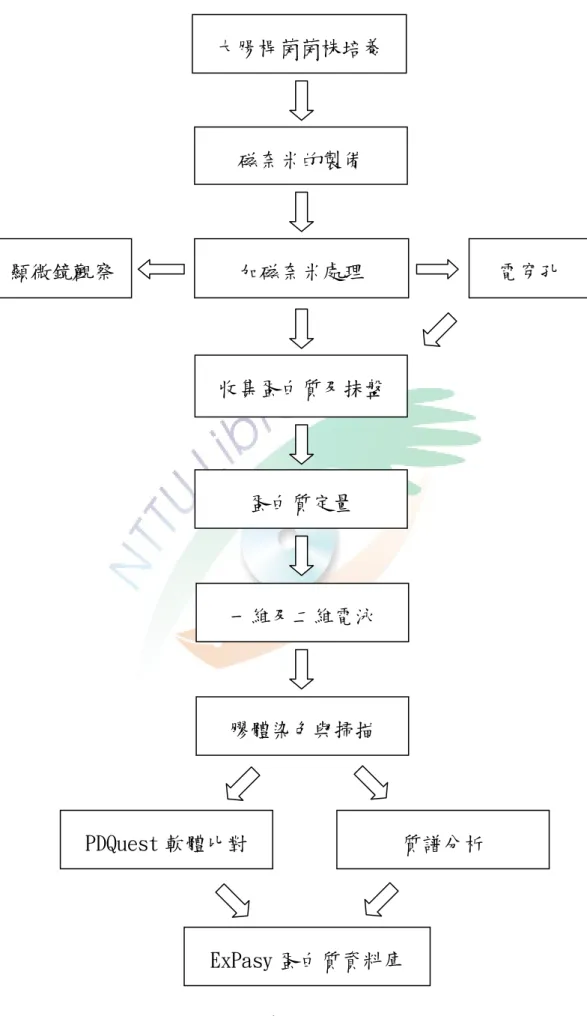
圖 1-3-1 實驗設計流程圖 ………40
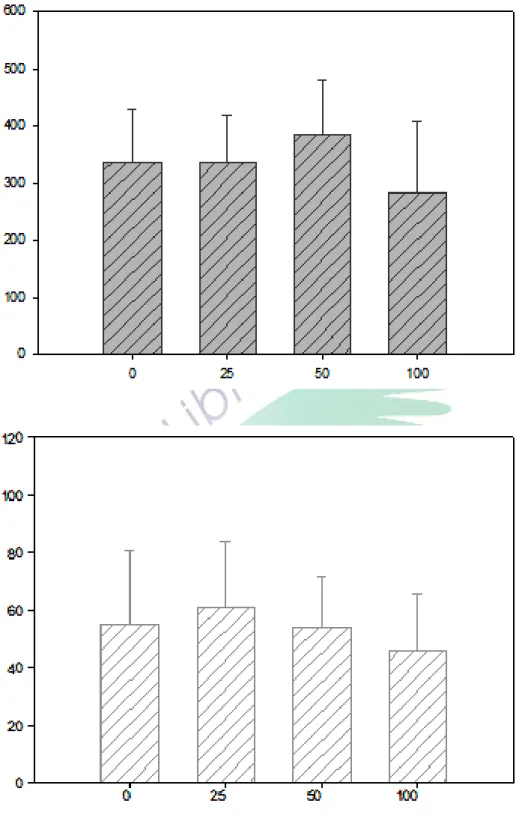
圖 3-3-1 磁奈米濃度效應之統計分析圖(BCRC11509) ………41
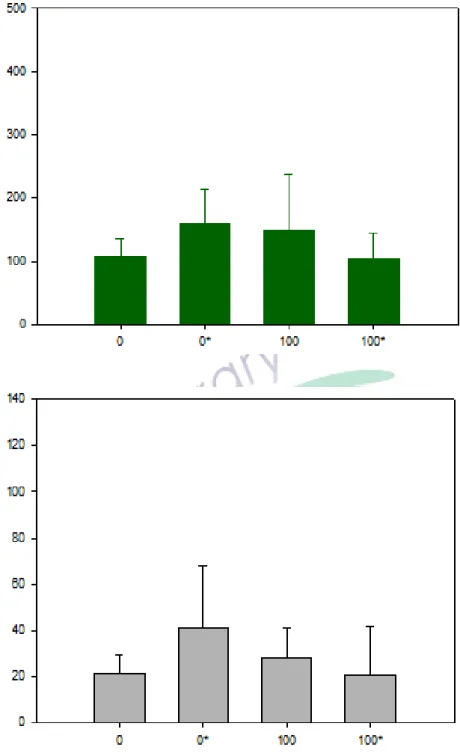
圖 3-4-1 磁奈米濃度效應與電穿孔效應之統計分析圖 (DH10B)…42
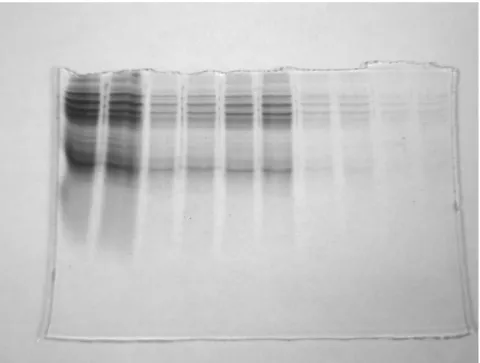
圖 3-6-1 磁奈米對 BCRC11509 蛋白質表現的一維蛋白質電泳分析
………43
圖 3-6-2 磁奈米對 DH10B 蛋白質表現的一維蛋白質電泳分析 …43
圖 3-7-1 PDQuest 軟體分析磁奈米對 BCRC11509 的影響 ………44
圖 3-7-2 PDQuest 軟體分析磁奈米對 BCRC11509 的影響 ………45
圖 3-7-3 PDQuest 軟體分析磁奈米對 BCRC11509 的影響 ………46
圖 3-7-4 PDQuest 軟體分析四種測試條件對 DH10B 的影響 ……47
圖 3-8-1 大腸桿菌的二維電泳標準圖譜 ………48
第一章 緒論
Ⅰ、 奈米顆粒與磁奈米
壹、 奈米科技的簡介
「奈米」一詞是由英文字 nanometer 直接翻譯而來,而 nano 在拉丁字義有
「矮小」的意思。奈米是一種長度的單位,一奈米(1nm)相當於十億分之一公尺,
即為 10-9公尺,相當於 10 個氫原子並排起來的長度(龔建華(2002))。一般細菌 的長度約為 2,000~3,000 奈米,病毒的顆粒大小一般約為幾十奈米,蛋白質的 寬度為 1~20 奈米,DNA 鏈的直徑為 2 奈米(姜忠義,成國祥(2004))。奈米科技,
乃泛指類似原子、分子等具有奈米尺度結構,或是具特殊功能的物質和材料元件 等相關製程和系統的科學技術。根據美國政府所公布的嚴格定義,奈米科技指的 是控制長、寬、高至少有一邊小於 100 奈米的物質結構和功能的相關技術(川合 知二(2003))。綜合上述之定義,奈米科技研究的尺度,乃是介於巨觀(macroscope) 與微觀(microscope)之間,也就是說物質或材料的三維結構之中,至少要有某一 維度的尺寸介於 1~100 奈米之間,或以它們做為基本單元構成的材料(張安華等 (2005))。欲冺用奈米科技將物質轉變成奈米材料,則必須透過經過很多的奈米 技術來實現。如果要掌握奈米技術,就必須深入研究物質的奈米結構及性質,同 時也要擁有製備奈米材料的方法。以下就奈米科技研究的內容分成三個方面來做 簡單介紹,分冸是奈米結構的認識、奈米動力學與電子學的應用以及奈米生物學 和醫藥學的發展(劉吉平、郝向陽(2003))。
一、 奈米結構
奈米結構的定義是三維空間中至少有一維處於奈米尺度的範圍,也就是物體 的長、寬、高只要任一維縮小至奈米尺度,或材料組成符合奈米結構者,則我們 就可以稱此材料具有奈米結構(馬遠榮(2002))。很多物質尺寸如果符合奈米結構 時,常會具有應用上的價值,例如奈米金屬、奈米纖維、奈米聚合物、奈米陶瓷、
奈米半導體、奈米磁性材料和奈米複合材料等。奈米結構依照維度,可分為零維、
一維、二維、三維(張安華等(2005))。零維是指長、寬、高三維尺度,均在奈米 尺寸以內,其形狀為點狀,例如奈米粒子(nanoparticle),以及原子或分子團簇 (atomic or molecule cluster)。 一維是指長、寬、高三維中,有二個維度處 於奈米尺度,例如奈米線(nanowire)、奈米帶(nanoribbon)、奈米棒(nanorod)、
奈米管(nanotube)。二維是指長、寬、高三維僅有一維處於奈米尺度,例如奈米 薄膜(nanofilm)、奈米片(nanosheet)、超晶格(superlattice)。而三維則是長、
寬、高皆不設限,但材料的組成部分是介孔級孔洞(mesoporous)、奈米粒子或是 奈米線等,也就是尺度由 1~100 奈米的微粒為主體所形成的塊體,就可被稱為 是三維的奈米結構材料,例如奈米塊體(nanobulk)、奈米模板(nano-template) 等。
二、 奈米動力學和奈米電子學
奈米動力學主要研究微機械或微電機,或總稱為微型電動機械系統(micro electronics and mechanics system,MEMS),主要用於有傳動機械的微型感測器 或執行器、光通信系統、醫療和診斷儀器等。奈米電子學包括有量子效應的奈米 電子元件、奈米結構的光電性質、奈米電子材料表徵,以及原子的操縱與組裝等 (劉吉平、郝向陽(2003))。奈米機械元件運作時,消耗功率本來就低,所以奈米 科技下的電子學,符合現今流行的節能省碳的原則,相信在往後必定會有很大的 前景與發展。結合奈米元件所產生的動力,以及奈米電子學的發展,也將對未來 的機械工具造成很大的影響。此外,以奈米技術製成的記憶體晶片,能使電腦的 體積縮小很多,同時也可以增加電腦的效能以及儲存功能。此外,奈米機電系統 (nanoelectromechanical systems)也是屬於奈米動力學和電子學的一部分,目 前在實驗室屬於正在發展的階段,工作內容包括仿自然生物馬達、奈米管與相關 材料,未來可能以非常小的奈米電子機械系統所製作的生物馬達來模仿自然運動 (nantural movement),例如人造肌肉可用奈米尺度的光纖製成,在沿其不同的 部位上以不同的光纖彈性或形狀等性質來搭配化學、光學上的改變,以確保肌肉 能彎曲和抓牢(張安華等(2005))。
三、 奈米生物學和奈米醫藥學
奈米科技可以研究生物分子間的交互作用,如磷脂、脂肪酸雙層平面生物膜 和 DNA 的精細結構等,還可以用自行組裝的方法,在細胞內放入零件以構成新的 材料。在醫學方面,用數層奈米粒子或結構,將有能力修復或是更換人體損壞的 器官,或研發新的藥物載體或藥物傳遞系統,例如介孔級的二氧化矽(mesoporous silica)做為藥物輸送載體,可將毒性高的藥物(例如阿黴素)包裹,並能夠安全 且準確地送至患部,進行藥物釋放,不僅可減少藥物的使用,還能降低藥物可能 對人體所產生的副作用(Timo Lebold, Christophe Jung, Jens Michaelis, &
Christoph Bra¨uchle, 2009)。研究奈米科技在生物學或醫學上的應用,也可以 在奈米尺度上了解生物大分子的精細結構及其與功能的關係,以獲得生命訊息。
科學家想冺用特製超細奈米材料製成的治療心血管疾病的奈米機械人,進入人體 的血管和心臟中,完成醫生無法完成的血管修補等細微步驟,且對人體健康不會 產生影響(龔建華(2002)),同時在人體的血液中巡邏,對人體各部位進行檢測、
診斷,甚至可在腫瘤初期檢查出癌細胞後,就提早進行治療以增加治癒率,或是
達到預防的效果。
貳、 奈米材料的性質
所謂奈米材料,包括奈米顆粒、薄膜、管柱等固體,因具有奈米結構,故其 性質會因為表面效應、體積效應、量子尺寸效應以及宏觀的量子穿隧效應而有所 影響,這四種效應影響的並不是一般巨觀或微觀情形下的物質,而是奈米尺度下 的材料受其影響最深,也因此造就了許多奈米材料的獨特性質。以下將針對這四 種效應,進行簡單的說明。
一、 表面效應
(surface effect)粒子直徑減少到奈米級時,表面原子數會迅速增加,也就是奈米粒子表面的 原子數和粒子內總原子數間的比值,會隨著粒子粒徑的變小而急遽增大,這使得 表面原子的性質,會對粒子所呈現的巨觀行為產生決定性的影響,此因表面原子 的作用而衍生的效應,即稱為「表面效應」(羅吉宗等(2003))。粒子的粒徑若在 10nm 以下,則表面原子的比例會迅速增加。當粒子的粒徑下降至 1nm 時,表面 原子數比例就會高達 95%以上,也就是原子幾乎會全部集中到奈米粒子的表 面,使粒子總表面積急遽變大(羅吉宗等(2003))。所以奈米材料擁有大的表面積 及表面原子數,同時位於粒子表層的表面原子之鍵結,也因未能配位完全,產生 大量的懸鍵,導致奈米粒子通常擁有相當高的表面能,即表面原子會具有高的表 面能。這些表面原子如果有機會遇到其他奈米粒子的表面原子,就會因具有高的 表面活性而很快地彼此結合在一起,產生凝聚的現象,甚至聚集成較大粒徑的顆 粒,這也就是為什麼奈米微粒常常彼此會團聚(aggregate)的原因,同時高的表 面活性也容易和外來的原子產生化學變化而結合,意即奈米粒子具有很高的化學 活性。例如黃金(Au)的活性很小,但當它的粒徑到達奈米尺度時,化學性質會變 得非常活潑,甚至可以做為催化劑。在空氣中,金屬的奈米顆粒會迅速氧化和燃 燒,因此可以成為新一代的高效催化劑材料(龔建華(2002)),並且將可大幅提高 化學工廠的生產效率。
二、 體積效應
(volume effect)一般微粒因為粒徑大多大於次微米級,所以每個粒子內所含的原子數量高達 109個以上,在統計上可視此粒子為無限多個原子的集合體。粒子中所含的無數 多個原子間通常會具有長程的交互作用,因而形成有序的週期性晶格(crystal lattice),或準晶格(quasi-crystal lattice)排列。此種高度有序的原子排列,
常會伴隨週期性的邊界條件,使得組成原子的電子能階形成複雜的連續能帶結構 (energy-band structure)。相反的,奈米微粒因為其體積和光波波長、物質波
波長、超導態相干長度等物理特徵的尺寸相當或更小,故粒子內所含的原子數目 有限,其對應的質量也極小,導致晶體週期性邊界條件被破壞,使得粒子的內部 不再具有有序的週期性結構,電子能帶也沒有連續的結構,所以常產生許多異於 一般固體材料的性質,而且無法使用傳統上含有無限多個原子的固態物理的理論 來解釋。這種因為粒子體積縮小而衍生的特殊現象,稱之為「體積效應」(羅吉 宗等(2003)),當奈米尺度的強磁性顆粒體積到達單磁區的臨界大小後,將具有 高矯頑力,可製成磁性信用卡、鑰匙、車票、磁性流體等,並用於電器元件、阻 尼元件、潤滑、選礦等領域。此外,奈米微粒的熔點遠低於塊狀金屬,例如 2nm 的金奈米顆粒的熔點為 600K,隨著粒徑增加,熔點會迅速上升,而塊狀金的熔 點為 1337K々銀(Ag)的熔點是 943K,但是銀奈米的熔點可降低至 373K,在粉末 冶金工業上提供新的工藝契機(張安華等(2005))。
三、 量子尺寸效應
(quantum size effect)1962 年,日本物理學家久保亮武(R.Kubo)首度提出關於次微米級尺度金屬 微粒子的電子性質之理論,說明粒子的粒徑縮小到一定值時,費米能階附近的電 子能級會由連續的能帶結構,轉變為不連續的離散能階,並且對此現象提出一個 簡單的理論模型,描述量子效應所造成微粒子離散電子狀態之物性觀念,這就是 著名的「久保理論」(Kubo effect) (羅吉宗等(2003))。因為能級的結構和分布 是主導材料各種性質表現的主要因素,所以離散的能階會使許多物理性質的量子 行為更加凸顯,進而可巨觀地呈現出來,此即所謂「量子尺寸效應」,又可稱為
「電子能階的不連續性」。奈米材料的諸多特殊的性質,常會和此效應中的離散 的能階有關。例如在溫度為 1K 時,銀微粒的粒徑若小於 14nm 時,它會從原有可 導電的性質,轉變成不能再導電的絕緣性(張安華等(2005))々金屬的奈米材料具 有較高的化學反應活性,也具有催化性能々吸收光譜線具有頻移作用,且吸光能 力強,使奈米粒子顏色大多為黑色,粒徑越小,顏色越深,也就是對光的吸收能 力提高,同時對光的反射率很低,一般低於 1%,例如銀白色的鉑變成鉑黑,鉻 變成鉻黑,鎳變成鎳黑等。冺用此特性可以製作高效光熱或光電轉換材料,將太 陽能高效地轉為熱能或電能,也可以應用在軍事上做為飛機、火砲的隱形材料(劉 吉平、郝向陽(2003))。
四、 宏觀的量子穿隧效應
(macroscopic quantum tunneling effect) 微觀粒子貫穿能障(potential barrier)的能力,稱為「穿隧效應」。在奈米 科技興起之前,此現象只能在原子或分子的微觀世界中才能觀察到。近年在奈米 材料的研究中,陸續在宏觀的實驗測量時,發現和穿隧效應有關的巨觀物理量,例如奈米微粒的磁化強度,以及量子相干元件(quantum coherent device)中之 磁通量等測量中,均發現穿隧效應的存在,所以又稱為「宏觀的量子穿隧效應」。
早期研究發現,常規鎳(Ni)金屬材料在室溫下呈現強鐵磁性,但鎳粒子的粒徑若 縮小至奈米級時,則溫度降至液態氩這樣的低溫下,鎳奈米微粒會呈現順磁性,
且比一般順磁性材料高好幾個數量級的磁化率值,故鎳奈米微粒的順磁性行為又 被稱為「超順磁性」(superparamagnetism,SPM),此時的矯頑力降為零。宏觀 量子穿隧效應的研究,不僅對基礎研究的發展有重要性,對實用化的應用也具有 相當重要的意義,未來奈米電子元件的應用與發展,研究人員必須慎重考慮來自 量子尺寸效應和穿隧效應的限制(羅吉宗等(2003))。
參、 磁奈米的特性及生醫應用
磁奈米粒子和其他奈米粒子不同的地方,在於其擁有磁性。它之所以擁有磁 性,是和不成對的電子有關。過去有許多學者冺用許多種類的氧化鐵粒子,例如 陶鐵磁性的γ-Fe2O3 (maghemite)、Fe3O4 (magnetite)等,使其縮小至奈米尺寸 以製作具有超順磁現象的磁奈米粒子,並且也會冺用其表面積大、官能基團的吸 附能力大等特性,來進行磁奈米顆粒的表面修飾,或吸附、鍵結具有生物活性的 分子,例如抗體或萬古黴素等,以得到許多應用在生物醫學的磁奈米粒子,使磁 奈米粒子具有多種功能。氧化鐵磁奈米粒子的製備,和一般奈米顆粒材料的製備 方法相同,常用的合成方法大致可分成傳統方法以及非傳統方法中的物理法和化 學法,其中以化學共沉澱法為普遍被採用的方法(張安華等(2005))。
生物醫學結合了許多不同的領域,例如奈米科學、分子生物學、生物工程學 等,而理想的生醫材料應該要包括下列特性〆良好的生物相容性、穩定的結構、
適度的生物降解速率及材料取得和製備的方便性等。目前科學家們對鐵氧磁奈米 材料的研究已有相當長的時間,此材料在生物醫學上,因為具有一些特冸優異的 性質,例如超順磁性以及高矯頑磁力等,特冸是也具有相對較好的生物安全性以 及生物相容性等特性,使其具有很多的應用,如核磁共振造影對比劑、藥物投遞 系統(drug delivery)、螢光標定追蹤或是腫瘤、癌症治療、冺用磁分離的純化 技術等。因此,如何製造具有特定大小、形狀、功能、低的生物毒性和穩定度的 磁性奈米顆粒,可說是目前科學研究最熱門的課題。磁奈米粒子可以進行患部追 蹤、治療、定量分析,若進行表面修飾後就能增加功能性,使其在生醫領域中有 更多的用途(黃治維(2006))。
一、 磁性來源
構成物質的基本粒子為原子,其磁性源自於原子內部電子的兩種運動〆電子 本身的自旋運動,以及電子繞原子核進行的軌道運動(原子核自轉的效應和電子 比起來可以忽略)。以一般的結晶材料而言,電子自旋運動對磁性的貢獻,遠比 軌道運動來得重要。若原子或離子具有磁矩(magnetic moment),那是由不完全
填滿的軌域電子所貢獻。然而過渡金屬族的元素與稀土族的元素,因為其原子或 離子內的軌域不完全填滿電子,也就是具有未成對的電子,所以會具有淨磁矩,
其中以鐵族元素或稀土族元素所形成的金屬、合金或化合物所擁有的磁性最強。
根據物質在外加磁場中的磁化程度與磁化方向,所有物質皆可進一步區分成反磁 性(diamagnetism)、順磁性(paramagnetism)、鐵磁性(ferromagnetism)、反鐵 磁性(antiferromagnetism)、陶鐵磁性(ferrimagnetism)等五種物質(張安華等 (2005))。以下將分冸就這五種性質作一概述。
「反磁性」是指在外加磁場下,物質內部環繞原子核運動的電子會產生微弱 的磁化量,依據冷次定律(Lenz law),電子運動的感應電流所產生的磁通量方向 和外加磁場的方向相反,以抵銷外加磁場,磁化率(magnetic susceptibility) 為負,大小約為10-5。這種現象廣泛存在於各物質中,在磁場中可略為降低磁場 強度,只是這種現象相當微弱,容易被其他磁性質掩蓋而無法清楚顯現,只有當 原子或是離子、分子的電子軌域填滿、電子淨磁矩為零時,才容易被發現,例如 氩、氖、氫、氮、銅、銀、金等元素,或是具有氯化鈉晶形結構之無機材料以及 大部分的有機分子(張安華等(2005))。
「順磁性」是指在外加磁場下,物質可產生微弱磁化量,和外加磁場的方向 相同,其導磁率為正,略大於1。所以當有外加磁場時,電子自旋的平均方向和 外加磁場平行,產生弱的感應磁化量々若是無外加磁場時,就呈現雜亂的排列。
典型的順磁性材料有過渡金屬鹽類、稀土元素及其氧化物等(張安華等(2005))。
「鐵磁性」是指材料本身具有磁矩,彼此互相平行,使總淨磁矩不為零,當 外加磁場磁化時,磁矩間會產生強烈的正交互作用,使彼此同向平行排列,可以 產生較大的磁矩,而強化材料的磁性,因此形成很強的自生磁化(spontaneous magnetization)的能力(張安華等(2005))。但是當溫度高於特定大小時,此材料 的磁性會從原本的鐵磁性,變成順磁性,此特定溫度稱為「居禮溫度」(Curie temperature々Tc),又稱為奈爾溫度(Néel temperature) (李國棟(2005))。鐵 磁性的物質常見的有鐵、鈷、鎳等金屬,以及鐵鎳合金、鐵鋁矽合金、稀土類磁 石等磁性物質。
「反鐵磁性」材料如果置於外加磁場中,則外加磁場會驅使此材料中每一鄰 近的磁性原子或離子間的磁矩,成為平行反向排列,導致磁矩彼此完全抵銷,無 淨磁化量。典型的反鐵磁性材料有氧化錳、氧化鐵、氧化鎳等(張安華等(2005))。
「陶鐵磁性」,又稱為亞鐵磁性,具有類似鐵磁性物質的行為,相鄰兩磁矩 平行,但大小不同,故仍存在一淨磁矩。陶鐵磁性發生在多數且常見的磁性氧化 物,也就是鐵氧磁體(張安華等(2005))。歷史紀錄上人類最早發現與應用的天然
磁石,即為一種鐵的鐵氧磁體。鐵氧磁體是一種以氧化鐵(Fe2O3)為主,搭配它的 金屬氧化物所形成的磁性複合氧化物,即為金屬磁性材料的總稱。這些鐵氧化物 主要可分成三大類,一種為尖晶石型(spinel stucture),例如鋅系、鎳鋅系、
錳鋅系、鋰系等,具有陶鐵磁性,其結構與尖晶石礦物的構造(MgO〃Al2O3)相同,
故此材料有一結構通式〆MO〄Fe2O3,其中M為二價的金屬離子,當M為含Fe、Ni、
Zn等金屬離子時,則稱之為鎳鋅鐵氧磁體々另一種結構為石榴子石型(garnet stucture),如釔鐵氧化物,屬於反鐵磁性々第三種為六方晶系型(hexagonal stucture),如鋇系、鍶系,具有永久磁鐵的性質。三種鐵氧磁體中以尖晶石型 鐵氧化物發展最早,研究最多,實用性也最高,如磁鐵礦(Fe3O4) (孔守中(2003))。
陶鐵磁性主要是發生在離子化合物上,由於不同的離子在晶體結構上的位置 不同,且其磁矩大小也各不相同,當磁化時其磁矩間會產生負交互作用,故產生 的自生磁化較弱。當磁粒子小於某ㄧ尺寸時,此時粒子會形成單一磁區(single domain),即一個粒子只包含一個磁區,也就是說,大粒子形成多磁區結構,當 粒徑減少後會形成單磁區結構,粒子再繼續減小時,則呈現超順磁性。Kittel 和Brown計算出球形的Fe3O4粒子形成單一磁區的臨界半徑(critical radius)為 40奈米々根據Berkowitz,Schuele和Flander估算Fe3O4粒子超順磁性的臨界半徑為 30奈米(張安華等(2005))。而對鐵氧磁體而言,其單一磁區的臨界半徑,通常在 十幾到數十奈米之間(李玉寶主編(2006))。磁奈米顆粒的每一單磁區,都相當於 是一個小磁鐵,在此小尺寸下,外界的熱效應會使電子的自旋變成不定向,使得 磁矩消失,在較弱的磁場下就可以呈現磁飽和,而且也沒有磁滯的現象發生,故 氧化鐵磁性奈米粒子便會呈現超順磁的特性。上述五種磁性材料中,擁有真正強 磁性和磁滯現象者,僅有鐵磁性以及陶鐵磁性材料,所以這兩種材料常具有高度 的實用價值(王序揚(2005))。
二、 磁滯現象
磁滯現象是一般磁性物質都具有的現象,不同的磁性物質所對應的磁化狀態 與外加磁場的關係,都可以用磁滯曲線表示。當外界開始施加磁場時,磁矩開始 旋轉,導致磁區形狀開始改變,於是當外加磁場從一開始的去磁狀態逐漸增強 時,材料磁化現象會隨磁場的增加而增加,最後會達到一個飽和值,此時的磁化 量,稱為「飽和磁化量」(magnetic saturation々Ms)。當磁場從飽和磁化的狀 態下逐漸減少時,則材料的磁化強度也會下降,但是不會沿著原來的磁化數值改 變々當外加磁場回到零時,材料磁性並不會完全消失,仍會保留某一磁化強度,
稱為「殘留磁化量」(magnetic remanence々Mr)。若再施以反向磁場時,磁場強 度會逐漸減弱,最後也會達到負飽和磁化量。然而,當外加磁場開始將淨磁化量 反轉呈反向時,此時外加磁場大小,稱為磁性體的矯頑磁力(coercive force々 Hc),此現象稱為磁滯(magnetic hysteresis) (張安華等(2005))。若之後外加
磁場再轉為正向,磁化強度會有特定的變化。
三、 鐵的氧化物
在地殼中,鐵是含量豐富的過渡元素之一,鐵離子通常以二價或三價的形式 與其他元素結合,和氧氣化合形成氧化鐵是常見的現象。常見的氧化鐵有方鐵礦 (pleochroite,FeO)、赤鐵礦(hematite,α-Fe2O3)、磁赤鐵礦(maghemite,γ -Fe2O3)、磁鐵礦(magnetite,Fe3O4)、針鐵礦(goethite,α-FeOOH)、纖鐵礦 (lepidocrocite,γ-FeOOH)等的多種氧化物,呈現不同顏色,可作為染色劑。
針狀的磁赤鐵礦和磁鐵礦,則是最常見,也是應用性最廣泛的磁性物質。固態的 氧化鐵粉末會隨著粒徑大小不同而有不同穩定相。以鐵氧化物中最穩定的型態,
三氧化二鐵粒子為例,在 5 奈米以下以非晶態結構的 Fe2O3粒子較為穩定,當粉 末粒徑大到 30 奈米左右時,以γ-Fe2O3粒子較為穩定,若當粉末超過 30 奈米時,
則變為最穩定相的α-Fe2O3粒子(溫明鏡(2003))。氧化鐵的磁性會隨著種類不同 而有強弱之分,其中以四氧化三鐵(Fe3O4)最強,三氧化二鐵(Fe2O3)次之,其原因 和鐵離子在晶格結構中的排列有關,而四氧化三鐵的鐵離子正好可以達到最大的 磁矩。其他金屬離子取代鐵離子時,也會因為產生不同的磁矩變化而有不同強弱 的磁性(鄭豐裕(2006))。
Fe3O4的化學式可表示為 Fe2+Fe3+2O4,和γ-Fe2O3一樣以反尖晶石(inverse spinel)結構排列。反尖晶石和正尖晶石的結構是相同的,只是鐵離子的排列不 同(鄭豐裕(2006))。一般塊狀的四氧化三鐵屬於陶鐵磁性的材料,當製成奈米微 粒時,由於體積縮小,使內部磁矩無法克服熱擾動,而有不穩定的現象,此時磁 鐵礦會喪失陶鐵磁性,而轉變成超順磁性。對於 Fe 和 Fe3O4而言,其單一磁區的 臨界尺寸分冸為 12 奈米和 40 奈米,每個單磁區的奈米微粒實際上成為一個永久 磁鐵,要使這個磁鐵失去磁性,必須使每個粒子整體的磁矩反轉,這需要很大的 反向磁場,也就是具有很高的矯頑力(李玉寶主編(2006))。
四、磁奈米材料的製備
在無機材料中,具有磁性的四氧化三鐵為低毒性的,對人體的傷害性很低,
所以近年來廣泛應用在生物及醫學上(楊雅涵(2007))。傳統上的方法通常會以鐵 的氧化物為起始原料,經過煆燒(800-1200℃)之後,可得到鐵氧磁體粉末。但是,
合成磁奈米粒子(magnetic nanoparticle)時,通常藉由非傳統的方法進行,可 分為物理合成法及化學合成法,以下將進行簡單說明。
物理合成法,其化學性質、組成在製程前後並沒有改變,只是冺用機械力將 固體微細化,例如粉碎法、機械研磨法々或將固相熔化,或溶解成液相,或汽化
成氣相,再重新結晶析出,如鹽析結晶法、氣相蒸發法、濺鍍法及電弧放電法等。
其中,粉碎法又稱為「球磨法」,是 S.Papell 所發明,主要是將磁鐵礦 Fe3O4粉 末(微米等級)與煤油以及油酸界面活性劑按照一定的比例混合在一起,在球磨機 內進行研磨 500~1,000 小時,以保證 Fe3O4粒子達到膠體的尺寸,粒徑為 2.5~
15 奈米之間,再使用高速離心機去除較大粒子。此法雖然簡單,但是所需的成 本過高、時間長、效率低,較不適合大量生產,而且製備出來的奈米粒子並無法 很均勻,粒徑亦無法到達很小的尺寸(張安華等(2005))。
化學合成法,主要是冺用控制化學反應過程中的反應條件,例如調整溶液的 酸鹼值、離子的濃度、鐵離子的比例、反應溫度、氧化還原力、熱浴時間等,使 金屬陽離子和陰離子反應產生各種不同形態或粒徑大小的化合物或錯合物,例如 化學共沉澱法、化學氣相沉積法、水熱法、氧化還原法、溶膠凝膠法,以及有機 金屬鹽法等。當製備的溶液環境有些許不同時,則製造出的鐵磁奈米粒子就可能 會有不同的結構與成分(粒子大小、形狀、顏色、磁性大小與粒子表面性質等)。
這些合成法都有不同的優缺點、適用材質、粒徑大小以及品質極限,其中以化學 共沉澱法為最普遍被採用的方法之一,因為此製備方法具備操作簡單、可以大量 製備、成本便宜以及製備時間較短等四個優點,也可以達到粒徑均勻的目標。其 原理乃是冺用金屬離子在鹼性環境下可生成金屬氧化物而設計,主要是以水溶液 作為反應的系統(鄭豐裕(2006))。
製備磁奈米的化學共沉澱法,主要是將二價的鐵離子以及三價的鐵離子之含 水鹽類的溶液混合後,再以鹼性溶液滴定氧化後得到磁鐵礦微粒子。其中,二價 的鐵離子和三價的鐵離子的莫耳數比為 1〆2,但這種方法需要經過水洗、吸附、
沉澱、過濾、分散等程序,且製備出來的磁奈米不甚穩定,因為容易產生沉澱。
目前為了要有效地改善化學共沉澱法的奈米粒子沉澱現象,常會在製備好的磁奈 米粒子中加入保護劑,以免發生聚集或沉澱的現象。保護劑雖然有保護的作用,
但實際上還是可能發生沉澱現象,這是因為粒子較大,所受的重力影響也會變 大。反應所使用的鹼性水溶液,主要以氨水或氫氧化鈉水溶液為主,兩者作用有 些許不同,但最後的結果是一致的。生成的四氧化三鐵若沒有保護劑,可能會氧 化成三氧化二鐵或結塊,因此此方法製備磁奈米粒子之後,通常必須加入適當的 保護劑,以穩定剛形成的氧化鐵磁奈米粒子(邱月霞(2004))。
五、磁奈米在生物分子分離上的應用
當磁奈米粒子製備完成之後,常常會發生聚集或氧化的現象,所以可以藉由 磁奈米粒子的表面修飾,使粒子均勻分散,並加強粒子的功能。例如將磁奈米粒 子的表面與特定的分子,如蛋白質或 DNA 做結合時,可形成生物相容性的粒子,
也可應用在蛋白質或 DNA 的純化上,使目標分子從混合物中分離出來。此外,由
於外加磁場可輕易控制具有超順磁性的磁奈米粒子,所以冺用磁奈米與目標分子 進行連接後,再冺用外加磁場進行對目標分子的吸附,從而得到目標分子(邱月 霞(2004))。
六、磁奈米在基因轉殖上的應用
基因轉殖,是一種將外源基因送到細胞核或質體的染色體 DNA 中的技術,使 該細胞或生物體在性狀或遺傳上能表現出外源基因的特性,轉殖的方法可藉由具 有感染力的病毒或農桿菌等作為載體(vector)以間接達到目的,或者也可以將外 來的 DNA 片段或 RNA 以直接的方式送入細胞內進行轉殖,例如 PEG(polyethylene glycol)法、顯微注射(microinjection)法、基因槍(gene gun)法以及電穿孔法 (electroporation)等,其中電穿孔法是一種簡單、快速且有效率的方式,可將 基因送入細胞內。電穿孔法乃是冺用短暫的高電壓刺激,使細胞膜上的微小孔洞 打開,於是外界物質得以藉機進入細胞內部,也可以傳遞基因以外的大型分子與 藥物(黃明遠(2001))。所以電穿孔法常被用在活體細胞(in vivo)或詴管體外(in vitro)的實驗中,例如傳遞藥物進入細胞、膜蛋白插入、基因轉殖等。根據文獻,
影響電穿孔的基因轉殖的因素很多,而 DH10B 是常用的菌種之一(Zhang et al., 2007)。本研究欲探討磁奈米在某一濃度下經由電穿孔影響大腸桿菌後是否會造 成毒性,以提供將來磁奈米功能化之後的另一種可行的生醫應用,特冸是在基因 轉殖以及磁純化的效果上。
Ⅱ、 基因體、生物資訊與蛋白質體學
壹、 基因體
人類基因體的序列圖譜草稿在二○○○年六月二十六日終於宣佈完成,生命 科學研究於是登上有史以來的發展高峰,從此人類科技文明步入後基因體時代。
隨著大量基因體的定序及微陣列基因分析的應用,產生許多生物學新領域,例如 轉 錄 體(transcriptome) 、 代謝體 (metabolome) 、功能性基 因體 (functional genomics)、蛋白質體(proteome)等,整合生理、藥理、遺傳和演化等各層面的 資源研究(Cowell & Hawthorn, 2007; Sequeira & Turecki, 2006)。在以上這 些領域中,基因體學主要是探討各冸基因在染色體上的分佈、有無變異,以及其 在不同生理狀況下表現的情形,配合微陣列基因晶片(是一種革命性的技術,可 同時定量成千上萬個基因的表現),能夠瞭解細胞內基因之間,是如何進行互相 調控,並可以為一些疾病提供新的診斷以及治療方法。然而,基因體學只能瞭解 組織細胞中 DNA 轉錄為 mRNA 的情形,但是 mRNA 的表現量,並不等於真正具有 功能的蛋白質的表現量(王沛懿(2007))。因此,蛋白質體學便逐漸取代基因體
學,而開始成為目前主要的研究方法之一(Heidecker & Hare, 2007; Jaluria, Konstantopoulos, Betenbaugh, & Shiloach, 2007; O'Hara, Ding, Bernat, &
Franken, 2007)。
貳、 生物資訊
現今由於電腦計算與處理訊息的能力增強,加上網際網路的普及,科學家為 了整理生物蛋白質分子或基因體解碼後的龐大序列資料,於是便發展生物資訊學 (bioinformatics) (Ettema, de Vos, & van der Oost, 2005; Gatenby & Frieden, 2007)。生物資訊,在早期的定義乃專指應用電腦在貯存及軟體分析核酸、胺基 酸序列和蛋白質的結構數據上々目前大家的認定是只要是應用計算機科學來處理 生物學數據資料,進而提供解釋或建構模型的,都可算是生物資訊的範疇。生物 資訊學就是冺用電腦與網路的強大功能,來進一步研究生物學的內容。生物資訊 學的功能,在於分析與比對基因數列與基因定序、蛋白質結構分析與功能預測、
資料庫平台的建立與整合等,目前已經有很多科學家投入生物資訊研究的行列,
也使生物資訊分析的功能更加強大。生物資訊工具的種類,包括資料庫、序列相 似性尋找工具、序列和結構分析工具、立體結構分析、顯像工具、結構預測工具、
血緣分類分析工具及代謝分析工具等(T. L. Lee, Alba, Baxendale, Rennert, &
Chan, 2006; Yan, King, He, Caldwell, & Zhou, 2006a)。此外,重要的生物 資訊網站有美國的國家生物技術資訊中心(NCBI - The National Center for Biotechnology Information) 々 歐 洲 分子 生物 實 驗 室 (EMBL - The European Molecular Biology Laboratory) 々 瑞 士 生物資 訊 研 究 所 (ExPASy, at Swiss Institute of Bioinformatics)等(王沛懿(2007))。
參、蛋白質體學研究
一、蛋白質的二維電泳
帶電物質在電場的作用下,向著其電性相反的電極移動,此一現象稱為電泳 (electrophoresis)。它的分離原理乃是冺用各物質表面帶電的特性,造成表面 電荷量的差異,於是帶電粒子在電場中移動速率不同來將物質分離。影響物質在 電場中移動的因素,除了電荷造成的差異之外,外加電場的強弱、物質的大小和 形狀等都會對移動的速率造成影響。蛋白質分子是由胺基酸組成,而胺基酸帶有 可解離的胺基及羧基,是一種兩性電解質,在一定的 pH 值條件下會解離而帶電 (沈春榮(2008)),所以可冺用此種特性,藉由電泳將蛋白質分離。電泳方法有分 為一維以及二維,其中以二維的分離效果更顯著。蛋白質混合物經由二維電泳的 過程會進行兩次分離,首先在取出細胞內的蛋白質後,先進行第一維的分離,冺
用蛋白質樣品的等電點(isoelectric point,pI)差異,在不同 pH 值的梯度環境 下進行電泳,即稱為等電聚焦電泳(isoelectric focusing, IEF)。所謂的等電 點,即是指蛋白質的淨電荷為 0 的時候,也就是當蛋白質放置於不同 pH 值梯度 的膠體內電泳時,它會受電力作用而朝著與自身所帶電性相反的電極移動,直至 抵達與等電點相同的 pH 值時,才會停止,若它移到冸的 pH 值的地方時,則會因 為帶電而再度移動,以回到和它的 pI 值相符的 pH 處,只有在此處才會使蛋白質 不帶電,不受外界電場作用而移動。如此,不同蛋白質在不同 pH 值的膠條處有 了初步的分離。
第二維的分離,乃是冺用不同蛋白質會因分子量不同,而有不同的移動速率 進行以分離,分子量越小,移動速率越大,這種方法稱之為 SDS-PAGE(即十二烷 基磺酸鈉—聚丙烯醯胺膠體電泳,sodium dodecyl sulfate polyacrylamide gel electrophoresis)。SDS 是一種界面活性劑,其目的在於破壞蛋白質的二級和三 級結構,使蛋白質變性,也會使樣品蛋白質表面均勻帶有一層負電荷,可以一致 地向正極方向移動。樣本分子的泳動率和分子量大小有關,和本身所帶的電荷無 關。首先將膠條先使用平衡液(equilibration buffer)平衡兩次完成後,再平行 放在 SDS-PAGE 膠體上,並以洋菜膠(overlay agarose)固定,同時在電泳槽中加 入跑膠用的緩衝液(running buffer)後,再以電壓 100 伏特進行電泳。當電泳一 段時間後,膠條上各處不同的蛋白質會依分子量大小不同,移動到不同的位置,
而具有相同分子量的蛋白質,則會移動相同的距離。如此,在第一維的 pH 值分 離以及第二維的分子量分離,就會使很多的蛋白質點從樣本中分離開來。而每一 種特定的生物或細胞經過二維電泳後,都會有其特定的蛋白質點的分布。
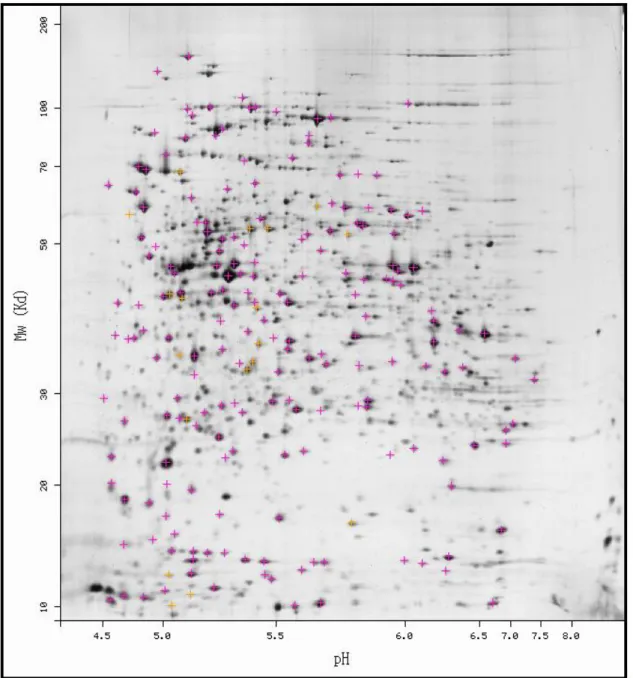
最後,跑完的電泳膠片經考馬士藍(coomassie blue)的染色以及去染(stain and destain)後,可以在膠片上辨識大部分蛋白質的位置,接下來會冺用軟體 (PDQuest)進行分析以解讀所有膠體上的蛋白質種類及表現的差異。早期由於蛋 白質無法大量鑑定,在應用上便受到限制,後來直到質譜儀有突破性的改進與生 物資訊學的出現之後,蛋白質二維電泳的方法才重新受到重視,並與質譜儀以及 生物資訊學形成蛋白質體學的核心,也是目前生化、生醫研究領域中最常使用的 一種二維分離方法。此外,蛋白質的二維電泳所使用特製電泳儀與電泳膠片,可 以輕易操作,其再現性也不錯(Barrett, Brophy, & Hamilton, 2005; Bradshaw
& Burlingame, 2005)。
二、 質譜分析
早期質譜儀主要應用於分析化學領域。其運作原理是讓化學分子先離子化 (帶上正或負電荷),然後使其在強大電場驅動下,於真空管路內飛行一定距離 後,撞擊偵測標靶,產生光電訊號。分子的質量不同,則物質飛行到達標靶時間
即不同々因此只要和數種已知質量的化學分子在同一質譜儀飛行時間的長短進行 比較,即可得知待測分子之質量。最近由於樣品離子化的技術,有突破性的進展 之後,質譜儀便開始應用於分析 DNA 或蛋白質等生物大分子,成為目前分析分子 種類的有力工具之一(王沛懿(2007))。研究者可以在蛋白質二維膠體上同時分開 數百至上千個蛋白質,並且能快速比較出性質相近的細胞或組織的差異,在膠體 上獨立的蛋白質點形成的二維電泳圖譜,以影像處理軟體進行分析,比對二維圖 譜之間的異同,並找出明顯變化而且有興趣的蛋白質點,接著挖取下來進行蛋白 質膠內水解(in gel digestion)的步驟,同時藉由胰蛋白酶(trypsin)切割成胺 基酸片段,將所得多種胜肽的產物,送至質譜儀將這些胺基酸片段的個冸質量進 行分析。當胺基酸質量決定出來後,直接將這些質量數目組合輸入資料庫(胰蛋 白脢切割所有已知蛋白質後所形成胺基酸片段的質量數目組合之資料庫)內進行 比對,就可以立刻得知欲研究的蛋白質點的身份 (Elrick, Walgren, Mitchell,
& Thompson, 2006; Hoorn, Hoffert, & Knepper, 2006; Nagele, Vollmer, Horth,
& Vad, 2004; Stephens, Quach, & Harry, 2005)。
三、 蛋白質體學
蛋白質體學(Proteomics),這個詞最早出現在 1994 年 Siena 二維電泳會議 中,由 Wilkins 等人所提出,並由 Wasinger 等人在其發表的論文中提出蛋白質 圖譜的觀念(吳惠君(2005))。蛋白質體,乃是指個體內所有被基因體表現之蛋白 質,而蛋白質體學則是研究探討細胞內某一特定的時間與狀態下,所有蛋白質的 種類與含量變化。當人類基因體計畫在 2000 年左右完成時,科學家發現人類所 擁有的基因竟只有 35,000 個,遠比當初科學家所預期的少,但每一個細胞的所 產生的蛋白質種類卻約有 100,000 種,遠比基因數目多,所以單一基因不僅可能 形成多種蛋白質,同時在不同身體部位的細胞,也有由不同基因所控制的不同蛋 白質群。相反地,由多個基因形成一個蛋白質,也是可能的。因此,從人類基因 體計劃初期的「一基因一蛋白」,變成「一基因多胜肽」或「多基因一蛋白」,說 明細胞的差異也許重點不在基因的多寡,而是在於蛋白質的表現。蛋白質體學的 興起,和二維電泳的發明、質譜儀偵測能力的提升,以及生物資訊學的日趨成熟 有關。其中,二維電泳可以冺用每一個蛋白質具有不同的等電點和分子量,進行 高解析度的分析。質譜儀在 1912 年一開始發明時,只能分析小分子的物質,直 到電噴灑游離法(electron spray ionization,ESI)以及基質輔助雷射脫附游離 法 - 飛 行 時 間 質 譜 儀 (matrix assisted laser desorption ionization time-of-flight mass spectrometry,MALDI-TOF)的出現,才能夠分析大分子的 物質。
Ⅲ、 研究動機與實驗目的
壹、 研究動機
在非金屬無機材料中,磁性奈米材料最引人注目,已成為新興生物材料領域 的研究重點,特冸是磁性奈米顆粒表現出良好的表面效應,官能團密度以及選擇 吸附的能力變大,且攜帶藥物或基因的百分比也增加。在物理學和生物學的意義 上,順磁性或超順磁性的鐵氧體奈米粒子在外加磁場作用下,溫度上升至 40~
45℃時,可達到殺死腫瘤的目的(姜忠義,成國祥(2004))。儘管磁奈米粒子在生 物醫學上有很多應用,但是對於磁奈米本身對各種功能性的大腸桿菌會產生何種 影響,目前尚不清楚。本研究冺用化學共沉澱法製備磁奈米粒子之後,在不添加 任何保護劑之下,直接對兩種大腸桿菌進行濃度效應,以及電穿孔的研究,目的 是要直接探討磁奈米粒子對大腸桿菌的影響,也避免保護劑可能對大腸桿菌產生 作用。本研究運用生物資訊分析工具以及蛋白質體學技術,研究磁奈米粒子的作 用機制,以開發其應用可能性。同時,藉由研究磁奈米對大腸桿菌的影響,思考 磁奈米粒子的應用性,探討大腸桿菌分泌的蛋白質中有什麼變化,並以蛋白質體 學及質譜儀分析之。本研究的實驗流程如圖 1-3-1。
貳、 研究目的
1.研究磁奈米對大腸桿菌的毒性測詴。
2.研究電穿孔及磁奈米對大腸桿菌蛋白質的共同影響。
3.研究磁奈米對大腸桿菌蛋白質體的影響。
4.以生物資訊的方法分析磁奈米的生化反應路徑。
第二章 材料與方法
Ⅰ、 實驗材料
壹、 大腸桿菌
本研究使用兩種不同品系的大腸桿菌,說明如下〆
一、 Escherichia coli. BCRC NO.11509
Escherichia coli. BCRC NO.11509(購自食品工業發展研究所生物資源保存 及研究中心,以下簡稱 BCRC11509)菌株的培養及保存,依照生物資源保存及研 究中心的建議(http:// www.bcrc.firdi.org.tw/wwwbcrc/main.jsp)。大腸桿菌 BCRC11509 是一株測詴抗生素耐受性的標準菌株,來源是 FDA strain Seattle 1946,全名是 Escherichia coli. (Migula) Castellani and Chalmers (BCRC NO.11509=ATCC 25922 =CIP 76.24 =DSM 1103 =JCM 5491 =NCIMB 12210)(Fan, Fett,
& Mitchell, 2007; Fass & Barnishan, 1979; Fuchs, Barry, & Brown, 1997;
Koseki & Yamamoto, 2006)。
二、 ElectroMAX DH10B Cells
ElectroMAX DH10B Cells (購自美國 Invitrogen,以下簡稱 DH10B),是一 種可藉由電穿孔的方式來進行基因轉殖的大腸桿菌,是一種勝任細胞(competent cell),但無法藉由熱休克(heat shock)來轉殖。須貯存於-80℃,不可貯存於液 態氮中。DH10B 可以有效地複製原核和真核的基因組 DNA(genomic DNA),冺用電 穿孔轉殖基因時,條件必須在 2.0kV,200Ω,25μF 的環境中進行。
貳、 化學詴劑
FeCl3〄6H2O 氯化鐵晶體 (J.T.Baker) FeCl2〄4H2O 氯化亞鐵晶體 (J.T.Baker)
hydrochloric acid﹐36.5-38.0% 鹽酸(HCl) (J.T.Baker) ammonium hydroxide﹐28.0-30.0% 氨水(NH4OH) (J.T.Baker)
tryptic soy broth 菌液培養基 (DIFCO 211825) tryptic soy agar 洋菜膠培養基 (DIFCO 236950) acetone 丙酮[(CH3)2CO] (J.T.Baker)
BSA(bovine serum albumin)牛血清白蛋白 (PIERCE) BCA protein assay reagent A (PIERCE)
BCA protein assay reagent B (PIERCE) glycine 甘胺酸(NH2CH2COOH) (J.T.Baker)
TRIS(Base) 三羥甲基氨基甲烷 NH2C(CH2OH)3 (J.T.Baker) acetic acid﹐glacial 冰醋酸(CH3COOH) (J.T.Baker) methyl alcohol﹐anhydrous 甲醇(CH30H) (J.T.Baker) formic acid 甲酸(HCOOH) (J.T.Baker)
sodium dodecyl sulfate 十二烷基磺酸鈉 SDS (SIGMA) Coomassie blue R-250 考馬士藍 (SIGMA)
TEMED(N,N,N´,N´-Tetramethylethylenediamine) 聚合反應詴劑(SIGMA) isopropanol 異丙醇(CH3CHOHCH3) (J.T.Baker)
ACN(Acetonitrile) 乙腈(CH3CN) (J.T.Baker) TFA(Trifluoroacetic acid) 三氟乙酸 (SIGMA) TCA(tricholoracetic acid) 三氯醋酸 (SIGMA) DTT(dithiothreitol) 二硫代蘇糖醇 (SIGMA) IAA(iodoacetamide) 碘乙醯胺 (SIGMA)
trypsin﹐Protemics Grade 胰蛋白酶 (SIGMA)
40% Acrylamide/Bis Solution﹐29〆1(3.3% C) 丙烯醯胺 (BIO-RAD) Rehydration/Sample Buffer 樣品緩衝液 (BIO-RAD)
ReadyStrip IPG Strip 型號 163-2007 固定 pH 值梯度的膠條 (BIO-RAD) equilibration buffer Ⅰ 平衡液Ⅰ (BIO-RAD)
equilibration buffer Ⅱ 平衡液Ⅱ (BIO-RAD) ammonium persulfate 過硫酸銨 (BIO-RAD)
30% glycerol solution 30%的甘油溶液 (BIO-RAD) mineral oil 礦物油 (BIO-RAD)
overlay agarose 覆蓋填補用的洋菜膠(BIO-RAD) nanopure water 超純水 (BIO-RAD)
參、 儀器設備
桌上型冷凍高速離心機 型號 Z 323K (HERMLE)
真空離心濃縮機 型號 Concentrator 5301 (eppendorf) disposable cuvettes 型號 223-9950 (BIO-RAD)
PROTEAN IEF Cell 二維等電聚膠電泳儀 型號 526BR 05801 (BIO-RAD) 不斷電供應器 型號 C-1000 (飛瑞股份有限公司)
分子聚膠儀 型號 041BR 23537 (BIO-RAD) 超純水製造機 型號 D8991 (Barnstead)
超音波清洗機 型號 DC300H (DELTA) 振盪器 型號 TS-500 (ORBITAL SHAKED) 分光光度計 型號 001905 (LaboMed.Inc.) 微波爐 型號 NE-P20 (National)
PETRI DISH 抹盤用的培養皿 (ALPHA PLUS)
Ⅱ、 實驗方法
壹、 菌株的培養
BCRC11509 及 DH10B 使用培養基為大豆酪蛋白消化培養基(soybean-casein didest medium々DIFCO 211825),配製時量取 30 公克的粉末,加入 1 公升的純 水中充分地攪拌均勻,此時在 1 公升的溶液中,含有 17 公克的胰消化的酪蛋白 (pancreatic digest of casein)、3 公克的葡萄糖(dextrose)、2.5 公克的氯化 鈉(sodium chloride)以及 5 公克的磷酸鉀(dipotassium phosphate)。最後調整 pH 值到 7.3,然後放入高壓滅菌鍋(autoclave)中,於高溫 120°C,蒸汽壓 1.2 kg/m2的狀態下,滅菌 50 分鐘。而抹盤時使用的洋菜膠為大豆酪蛋白消化培養基 (DIFCO 236950),配製時則是量取 40 公克的粉末,加入 1 公升的純水中充分攪 拌均勻,此時在 1 公升的溶液中含有 15 公克的胰消化的酪蛋白、5 公克的酵素 消化的大豆粉(enzymatic digest of soybean meal)、5 公克的氯化鈉以及 15 公克的洋菜(agar)。同樣要將 pH 值調整到 7.3,然後一樣放入高壓滅菌鍋中,
於高溫 120°C、蒸汽壓 1.2 kg/m2的狀態下,滅菌 50 分鐘。在菌的培養上,每天 會添加新的培養液,以增加與加快細菌的生長。
貳、磁奈米粒子的製備
本研究的製備方法是使用化學共沉澱法。首先配製 2.0M 的鹽酸 50 mL,再 配製 1.0 M 氨水溶液 100 mL,並冺用 2.0M 的鹽酸溶液配製成 1.0M 的氯化鐵(FeCl3) 酸性溶液 10 mL 以及 2.0M 的氯化亞鐵(FeCl2)酸性溶液 10 mL。繼續將 1.0 M 氨 水溶液 50 mL 裝置於滴定管中,再將 4.0 mL 的 1.0M 氯化鐵鹽酸溶液以及 1.0 mL 的 2.0M 氯化亞鐵鹽酸溶液加入到 100 mL 的燒杯中,放置於電磁攪拌器上攪拌。
將 50 mL 1.0 M 的氨水溶液用滴定管於 5 分鐘內緩緩加入燒杯中,此時可以看 到黑色的沉澱形成,此即為鐵磁奈米顆粒。滴定完成後將電磁攪拌器關閉,取出 磁攪拌棒,待其沉澱後將上清液倒掉,留下固態的沉澱物,並將固態的沉澱物倒 入秤量盒中,以裝有二次水的洗滌瓶沖洗燒杯中的殘留物。待其沉澱後將上清液 倒掉,加入二次水以玻棒攪拌後,再將上清液倒掉,之後重複數次,並將燒杯中 殘存的鐵磁奈米粒子收集倒入秤量盒中(Jain, Wang, Jones, Hawkett, & Warr,
2010)。本實驗中並未加入氫氧化四甲基銨水溶液,以獲得沒有分散劑、單純的 磁奈米粒子。最後將強力磁鐵放置於秤量盒下方,待盒中漆黑的液體沉澱後,倒 掉上清液,再加入二次水清洗、靜置後移動強力磁鐵後再將盒中漆黑的液體倒 掉,重複數次,以洗去水溶液中殘存的氨,並得到鐵磁奈米粒子,最後經過高溫 (50℃)乾燥整夜得到粉末後保存備用。
參、 磁奈米對大腸桿菌的毒性測詴
取出乾燥的磁奈米粉末後,先製備出的是濃度為 400μg/ml 磁奈米水溶液,
再製作出四種不同濃度的磁奈米菌液,分冸為 25μg/ml,50μg/ml,100μg/ml,
再加上控制組 0μg/ml,以上菌液濃度的製作過程如下〆將濃度 400μg/ml 磁奈 米水溶液稀釋成 200μg/ml、100μg/ml 兩種濃度,再將 400μg/ml、200μg/ml、
100μg/ml 三種濃度的水溶液及二次水(濃度=0)各取 1 毫升,分冸加入 3 毫升 的菌液中(將菌液濃度視為 0),以製作出 0μg/ml、25μg/ml、50μg/ml、100 μg/ml 等四種濃度的磁奈米菌液。本研究乃針對不同濃度(0μg/ml,25μg/ml, 50μg/ml,100μg/ml)的磁奈米菌液,以抹盤的方式進行觀察,並由菌落的生長 情形,分析磁奈米對大腸桿菌 BCRC11509、DH10B 是否有濃度效應。本研究影響 菌的時間為 22 小時,並於隔天觀察與記錄菌落的數量。當菌落數量很大時,則 計數時會使用棋盤法以估計數量,濃度上也採用適當的大小,以方便比較濃度效 應。抹盤時所需的培養皿是由洋菜膠倒入 PETRI DISH 抹盤用的培養皿之後,放 在抽氣櫃中靜置一天待其冷卻凝固,即可準備抹盤。抹盤前蓋子上若有水氣,則 必須擦拭乾淨,以免影響實驗結果。本研究將原始菌液先進行稀釋成 10-4、10-5、 10-6倍後才進行抹盤,因為此時產生的菌落數量適當,以免菌落生長太濃密而不 易計算,每個條件皆重複 5 盤,共 60 盤。抹盤使用的器材為 L 形玻棒,使用前 會先以高壓滅菌鍋的高溫高壓滅菌,進行抹盤時使用酒精燈加熱滅菌。抹盤之後 立即放入 37℃的恆溫箱中,培養至隔天同一時間,再統一計算每一盤的菌落數 量。若其中若有雜菌汙染時,則會扣除々若雜菌汙染該盤太嚴重時,則不予紀錄 該盤的菌落數量,以降低誤差。本實驗使用抹盤實驗進行毒性測詴,也藉由二維 電泳的方式研究是否造成 BCRC11509、DH10B 的蛋白質變化。
肆、 電穿孔實驗
本實驗藉由電穿孔的方式將磁奈米粒子送進 DH10B 菌體中,看看不同磁奈米 菌液的濃度之下,以及 DH10B 是否會因受到電穿孔的有無,而使得菌的生長形態 以及菌體內蛋白質發生變化。本研究選擇兩種磁奈米菌液濃度進行電穿孔詴驗,
也和沒有經過電穿孔的 DH10B 菌液進行比對,看看是否產生變化。此實驗有四種 測詴條件如下〆濃度 0μg/ml、無電穿孔々濃度 0μg/ml、有電穿孔々濃度 100 μg/ml、無電穿孔々濃度 100μg/ml、有電穿孔。磁奈米的濃度分為 0μg/ml 及
100μg/ml,濃度 0μg/ml 的配製過程,是以加入二次水配製做為控制組,濃度 100μg/ml 則為本研究中磁奈米溶液的最大濃度,藉此比較電穿孔及磁奈米菌液 濃度這兩個變因對菌的影響,也可比較這兩種變因是否會對 DH10B 產生其他特殊 的加乘效果。本實驗使用抹盤實驗進行毒性測詴,也會藉由二維電泳的方式研究 是否造成 DH10B 的蛋白質變化。
伍、 分泌性蛋白質收集
先將實驗室培養的大腸桿菌以 13000rpm 的轉速離心 15 分鐘,蒐集上層澄清 液,之後加入 20%(v/v)的 TCA,放入-20℃的冰箱中靜置 30 分鐘以上,先解凍 再進行離心後,倒掉上層液體以除去殘存的 TCA,再加入丙酮進行沉澱,將蛋白 質分裝在 4 支微量離心管中,在 4℃的環境下以 13000rpm 的轉速離心 15 分鐘,
再反覆以丙酮沖洗,最後再靜置-20℃的冰箱中保存備用。
陸、 蛋白質定量(BCA)
本實驗以牛血清白蛋白(BSA)作為定量的標準液,原始濃度為 2mg/ml。依其 產品說明書所示,先配製出 50μL 的標準液 10 管(各含 25、50、100、150、200、
250、300、400、500、600μg/ml 的 BSA),再和 1000μL 的蛋白質染料(protein dye)以 1〆20 混合,其中蛋白質染料是以 BCA protein assay reagent A 及 BCA protein assay reagent B 以體積比 50〆1 混合。用分光光度計測其在 562 nm 波長時的吸光值,畫出一條標準曲線。由於測量的時間不同,吸光值也會逐漸改 變,所以測量的時間統一在開始的 5 分鐘內完成,以免會造成太大的誤差。測量 樣品在 562 nm 以及 569 nm 波長的吸光值,並和標準曲線進行比對,可藉此得知 樣品中蛋白質的含量。若標準曲線的 R2值越高,表示越有正比的關係,才能越 正確地去推算出樣品中蛋白質的含量,且以此標準曲線的定量結果,可用於進行 一維電泳以及二維電泳之前的蛋白質定量。此外,由於各蛋白質標準液的吸光值 在 569 nm 時會有最大值,故本實驗也另外對分光光度計在 569 nm 波長時,測量 各蛋白質標準液的吸光值,以做為比較。
柒、 顯微鏡觀察
本研究將一般的光學顯微鏡的鏡頭改裝成相機鏡頭,並搭配可令鏡頭具有照 相功能的軟體,可以在電腦螢幕上以不同的放大倍率(40 倍、100 倍)觀察與紀錄 大腸桿菌 BCRC11509 及 DH10B 在不同濃度的磁奈米粒子水溶液的影響下,其外觀 形態和原本的形態比起來是否有產生變化。磁奈米水溶液濃度有下列四種大小〆 0μg/ml,25μg/ml,50μg/ml,100μg/ml,對菌的作用時間分冸是 2 小時和 22 小時。由於本研究使用的是一般的光學顯微鏡進行紀錄,所以在解析度上無法像
電子顯微鏡那樣可以真正看出磁奈米的存在,但是可以看到磁奈米粒子的團聚現 象,這是因為沒有分散劑的關係。
捌、 一維蛋白質電泳分析
先將大小玻璃以中性清潔劑清洗乾淨,再用 70%的酒精消毒後擦拭並乾燥,
然後架於鑄膠台上。依次進行操作步驟,以下將簡單說明操作步驟。
一、 蛋白質裝填
本實驗使用不同的蛋白質樣品進行裝填,分冸是一種濃度的 BCRC11509 磁奈 米菌液分泌性蛋白質,以及 DH10B 菌液四種條件的分泌性蛋白質(濃度 0μg/ml、
無電穿孔々濃度 0μg/ml、有電穿孔々濃度 100μg/ml、無電穿孔々濃度 100μ g/ml、有電穿孔)。首先至-20℃冰箱取出微量離心管,解凍後先以 13000rpm 離 心 15 分鐘,倒掉上層澄清液,在室溫下乾燥 30 分鐘,先進行蛋白質定量,接著 加入染劑 5μL 和定量後的蛋白質樣品 15μL (以 1〆3 的比例混合),將此 20μL 的蛋白質和染劑的混合液以 100℃加熱 5 分鐘備用,待鑄膠完成後,即可進行蛋 白質裝填。加熱時,管口以封口布密封,使內容物不致在加熱過程中溢出。加熱 進行之前,必須在管口處先戳一個洞,如此在加熱時管內若有多餘氣體可以跑出 來,管內才不會產生太大的氣體壓力。而加熱的目的,是為了破壞樣品蛋白質的 立體結構,使蛋白質成絲狀。
二、鑄膠
將大小玻璃固定在鑄膠台上,並開始準備製作膠片。膠片有分成上層膠與下 層膠,先製作下層膠。按照說明書配製一片 12%的下層膠膠片,所需的藥品依次 添加順序如下〆先將二次水 7.7 mL 加入燒杯中,再加入 1.5M 的 Tris (pH 值=8.8) 溶液 4.4 毫升,再加入 40%的丙烯醯胺溶液 5.3 mL,接下來加入 TEMED 溶液 7μL 以及濃度 10%的硫酸銨溶液 175μL。攪拌均勻後,以滴管吸取注入兩片玻璃片 的空隙中,最後留下一些空間,以便下層膠凝固後可加入上層膠之用。靜置大約 1~2 小時,待下層膠已凝固成型後,再加入上層膠。按照說明書配製一片 12%
的上層膠膠片,所需的藥品依次添加順序如下〆先將二次水 4.6 mL 加入燒杯中,
再加入 0.5M 的 Tris (pH 值=6.8)溶液 1.9 mL,再加入 40%的丙烯醯胺溶液 1.0 mL,接下來加入 TEMED 溶液 7.5μL 以及濃度 10%的硫酸銨溶液 75μL,接著再 插入梳子(comb)固定,靜置大約 1~2 小時,直至上層膠也凝固成型,將梳子取 下後,便可以進行蛋白質裝填。每一片上層膠共有十個蛋白質裝填處(well)可以 裝填蛋白質樣品。需要確定蛋白質分子量大小時,也可在最旁邊的裝填處加入 marker(標準液,內含許多已知分子量的蛋白質),以便比對其他蛋白質樣品在不
同的位置(band)的分子量。
三、 配製跑膠用的緩衝液
先配製濃度 5 倍的溶液備用,等到要進行實驗時再做稀釋成 5 倍的體積即可 使用。配製 3 公升、濃度 5 倍的溶液所需藥品如下〆glycine 藥品 216 公克、
Tris-base 藥品 45.5 公克、SDS 藥品 15 公克、二次水 3 公升。
四、 一維 SDS-PAGE 電泳
將跑膠用的緩衝液加入電泳槽內,至少蓋滿膠片玻璃下緣附近的白金線,而 上層跑膠用的緩衝液的量亦是需高過白金線。蓋上電泳槽上蓋,使用 200V 的電 源跑約一個小時即電泳完畢。電泳的過程都在溫度 20℃的低溫環境中,以免在 電泳的過程因溫度太高而影響實驗結果。
五、染色與退染
將電泳完畢的膠片取下,並浸入考馬士藍染劑(0.125%,coomassie blue R-250,50%甲醇10%冰醋酸)中,以震盪器調低轉速搖盪至少15分鐘。染色後 的膠片先浸入destain Ⅰ buffer(50%甲醇,10%冰醋酸),以震盪器調低轉速 搖盪,直到看出最靠近膠片末端的藍線,接著再將膠體繼續浸入 destain Ⅱ buffer(5%甲醇,7%冰醋酸),同樣以震盪器調低轉速搖盪,退染至肉眼可見 膠片之結果為主,約2~3小時後可以看到結果。最後可用二次水清洗與浸泡,以 除去膠上殘留的甲醇和冰醋酸。接著再取出膠片,放入封套中,並放在4℃冰箱 中保存備用。
六、膠片掃瞄
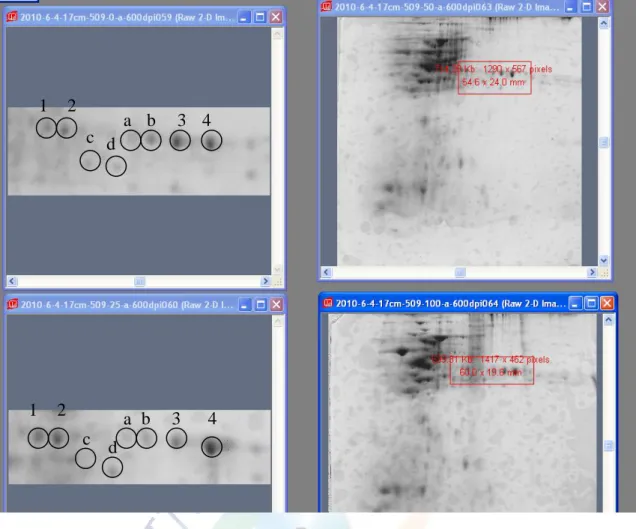
將膠片(電泳圖譜)通過掃描器掃描成 TIF 檔,解析度為 600dpi。
玖、 二維蛋白質電泳分析
和進行一維蛋白質電泳分析一樣,先將大小玻璃以中性清潔劑清洗乾淨,再 用 70%的酒精消毒後擦拭並乾燥,然後架於鑄膠台上。依次進行操作步驟,以下 將簡單說明操作步驟。
一、 蛋白質樣品處理
至-20℃冰箱取出微量離心管,解凍後先以 13000rpm 離心 15 分鐘,倒掉上 層澄清液,在室溫下乾燥 30 分鐘後,接著加入 300μL 的樣品緩衝液(8M urea,2
%CHAPS,50mM dithiothreitol;Bio-Rad)均勻混和後,並將蛋白質定量後,再 將此蛋白質樣品平均注入一條聚焦盤中,取兩片墊片(wicks),以二次水潤濕,
分冸置於正負兩極上方,小心不要有氣泡產生。放墊片的目的在於可以吸附樣品 中的雜質和鹽類,避免汙染物損壞電極而影響實驗。
二、 等電焦電泳分析
Protean IEF Cell(Bio-Rad)設計程式如下〆R(rehydration)〆20℃,50μA,
12 小時,此一步驟的目的是讓 rehydration buffer 和樣本混合,並吸入 IPG strip 膠條中。S1(condition Step)〆250V,15min,rapid,目的是移去鹽類離子及 污染物。S2(voltage reagent)〆10000V,rapid。S3(final focusing)〆10000V,
60000VHOURS,rapid,即到達 10000V 後,會使 S3的時間換算累積到 60000V*Hr。
S4(hold step)〆設定 500V,避免已聚焦的蛋白質發生擴散。當 S3完成後即可 進行膠條平衡,以便繼續進行第二維的 SDS-PAGE 分析。若 S3已完成了,但時間 上無法搭配,也可將行程設定停留在 S4一段時間,但不要超過 6 個小時為佳,
否則蛋白質容易發生擴散。
三、配製平衡液與膠條平衡
平衡液Ⅰ(equilibration buffer Ⅰ)的配製方法如下〆取 13.5 mL 的 30%
glycerol,加入平衡液Ⅰ(6M urea, 0.375M Tris-HCl,pH8.8, 2% SDS, 20%
glycerol,2%(w/v)DTT)的瓶子中,放在磁石攪拌器上攪拌,直到溶解為止。平 衡液Ⅱ(equilibration buffer Ⅱ)的配製方法如下〆取 13.5 mL、30% glycerol,
加進平衡液Ⅱ(6M urea,0.375M Tris-HCl, pH8.8 , 2% SDS, 20% glycerol,2.5%
(w/v)iodoacetamide)的瓶子中,放在磁石攪拌器上攪拌,直到溶解為止。
兩種平衡液配製完成後,再將第一維電泳完畢的膠條自聚焦盤上取出,把膠 條先以二次水清洗兩次,然後將膠面朝上,把底部的水及礦物油以廚房用的紙巾 大致吸除,再將膠條放入裝有平衡液Ⅰ的長玻璃管中,進行第一次的平衡,以震 盪器調低轉速搖盪 10~15 分鐘,在搖盪過程中膠面要朝上,不要超過 20 分鐘。
從長玻璃管中取出膠條後,再以二次水清洗兩次,將膠面朝上,把底部的水用廚 房專用紙巾大致吸除,然後把膠條放入裝有平衡液Ⅱ的長玻璃管中,進行第二次 的平衡,同樣以震盪器調低轉速,搖盪 10~15 分鐘,在搖盪過程中同樣膠面要 朝上,不要超過 20 分鐘。完成後從長玻璃管中將膠條取出以二次水清洗後,將