國立臺灣大學生物資源暨農學院園藝暨景觀學研究所 碩士論文
Department of Horticulture and Landscape Architecture College of Bioresources and Agriculture
National Taiwan University Master Thesis
利用二甲基標記結合質譜技術建立奇異果過敏原之絕對定量分析平台 Establishment of an analytical platform for the absolute quantification of
kiwifruit (Actinidia deliciosa) allergens using dimethyl labeling coupled with mass spectrometry
施奕辰 Yi-Chen Shih
指導教授:許輔 博士 Advisor: Fuu Sheu, Ph.D.
中華民國 107 年 8 月
August, 2018
致謝
三年前若有人問我為什麼要讀研究所,答案會是為了學習研究以及取得 學歷。而現在看來不只如此,更有其他使這段不長不短的時光更有價值的事情。
為此本篇致謝如化石標本,僅能做為一個曾經如此的證物而不足夠代表感謝的全 貌。
人說歸屬感是探索世界的原點,是在這社會的錨。三年來所完成的大小 事無不有第一研究室大家的支持,令人印象深刻的並不是那許多吃飯的場合,而 是進度報告完你一言我一句,積極想幫忙解決實驗困境的那個場景,是與老師討 論實驗規劃的小圓桌,是深夜一起邊聊天邊準備離開的默契。第一研究室像一個 家,能得到關心與感到安全,而其中的大家皆是家人,很幸運能夠身為一員,也 期待這份歸屬感永不消失。
研究能力不只能於學術上有所應用,更像是一個能夠應用於生活的技能,
其中包含了邏輯思考、明辨是非的能力。學習是單純而快樂的過程,不管是與他 人討論研究的意義,找出自己在科學上最獨一無二的價值;全盤審視自身能力與 資源,摸索找尋適合的研究方向;從無數的文獻中找出關鍵的幾篇,並藉由理解 他人的實驗從中反思得到靈感。於學術上承上接後,從零到有步步建造一個屬於 自己的研究,皆是人生中獨一無二極具成就感的感受。
如同實驗上有起有伏,這段時間的情感亦有升有落。從初次見面到不捨 分離,從無法同理到互相幫忙,從形同陌路到能夠互相依靠,敬佩、欣賞、感激、
尊重、驕傲、快樂、不捨、祝福、尊敬、珍惜、期許,情感好或壞都已化為這段 期間人格的成長,為此感到珍惜也為自己的逐漸成熟感到自豪。
感謝參與這人生階段的每一個人,或多或少都造就了現在的我,感謝口 試委員 周志輝 博士、繆希椿 博士、蘇南維 博士以及陳家揚 博士提供各領域 深入的見解,拓展了研究視野上的深度與廣度,感謝讓我再次意識到虛心求教的 重要性。特別感謝恩師 許輔 教授在學術、待人處事、人生方面給予諸多指導與 建議,為成熟有智慧的榜樣,我亦以此為目標,期望有天能成為像老師一樣甚至 更好的人。
羅馬不是一天造成的,更亦不是單憑一人的力量。這本論文的完成是研
究所生涯的一個里程碑,能夠抵達必須感謝所有的一切。不管是科學界前人的努 力,使我能站在巨人的肩膀上努力;國家、社會、學校提供的豐厚資源,讓我有 足夠的材料與能力去完成研究;家人、師長、同學、朋友、夥伴的支持,使我有 足夠的動力去突破困境;對於一切都感到無法回報的感恩,能做的是有一天成為 有成就的人來回報大家。
施奕辰 謹誌於 國立臺灣大學園藝學景觀所 2018 年 7 月 26 日
摘要
食品過敏為重要之食品安全議題,而過敏原之定量法為其管理上所必需的。
市面上最普及的 ELISA 法,方法立基於抗體,亦受抗體特性所限制,具有許多 難以改善之缺點。近年來以質譜為基礎之過敏原定量法逐漸受到重視,因其不僅 能解決 ELISA 法之缺點,同時具有高通量、能提供分子指紋證據等優點。在食 品過敏原中以水果過敏原之定量,因為蛋白質含量低、干擾物質多故較為困難。
奇異果 (Actinidia deliciosa)為國人大量食用之水果,亦為致敏性食物,於歐洲食 物過敏原排名前六,於台灣排名前十,為我國建議標示之食品過敏原。本研究選 擇奇異果作為代表物進行質譜過敏原定量法之開發。
結果分為三部分:第一部分、以奇異果為例之水果過敏原質譜前處理法開發 與優化。開發出兩種適合水果之苯酚蛋白質萃取法,分別為最佳化苯酚蛋白質萃 取法 (optimized phenol extraction method, OPE) 及微量苯酚萃取法 (micro phenol extraction method, MPE),兩者皆具高萃取率。其中 OPE 單次萃取蛋白質總重量 較 MPE 多;而 MPE 之成本低、操作簡便快速 (1 天)、具良好之消化步驟相容 性。同時優化胰蛋白酶水解法與確認二甲基穩定同位素標記法之標記效率與同位 素效應,並將 MPE、胰蛋白酶水解法、二甲基標記法與樣品除鹽法,整合為一套 可連續且低轉換損耗之質譜樣品前處理法。第二部分、建立及確效奇異果過敏原 之質譜絕對定量分析法。利用 quadrupole time of flight mass spectrometer (Q-ToF MS) 結果來進行奇異果目標過敏原之選擇。以 Act d 1、Act d 5、Act d 11 作為目 標物,AD1、AD5、AD11 胜肽分別作為其代表胜肽。再以 triple quadrupole mass spectrometer (QqQ MS)選擇代表胜肽之定量子離子與定性子離子及進行 LC 與 MS 之參數調整。對本法進行確效試驗,AD1H、AD5H 胜肽之定量範圍分別為 1250-125000 ng/mL、2500-250000 ng/mL;準確度(回收率)分別為 82.76%、83.46%;
重複性(變異係數)分別為 6.61%、8.69%;儀器定量極限分別為 0.008 ng/mL、8 ng/mL;儀器偵測極限分別為 0.008 ng/mL、4 ng/mL;兩者皆具良好專一性及線 性,成功建立於多重反應監測模式下之奇異果過敏原質譜分析法。第三部分、將 此法應用於奇異果加工製品中。本研究比較了鮮果、果醬、熱風乾燥果乾、冷凍
乾燥果乾、熱殺菌果泥、高靜水壓果泥、熱殺菌果汁與高靜水壓果汁,八種常見 之奇異果產品於加工後過敏原之消長。此方法成功定量出八個處理組之 Act d 1、
Act d 5 含量,證明了本法適用於上述加工奇異果加工製品中。於長時間加熱之 加工製品中 (果醬與熱風乾燥果乾) 過敏原含量較低,而熱殺菌、高靜水壓或冷 凍乾燥製程對於過敏原含量較無影響。
本研究成功開發出 OPE 與 MPE 兩種水果蛋白質萃取法,並將 MPE、蛋白 水解法、標記法與樣品除鹽法整合優化為一套質譜樣品前處理法,及成功建立奇 異果過敏原質譜絕對定量分析法,並成功將此方法應用於八種奇異果食品中。本 研究可作為未來食品過敏原方法開發之範本,並於未來能應用於各國,協助食品 安全之提升。
關鍵字:二甲基標記、三重四極柱質譜儀、過敏原、絕對定量、美味獼猴桃
Abstract
Food allergy management is one of the main issues in food safety. For efficient management of allergy, an effective analytical method is required. In the past, antibody- based Immuno-assay ELISA was the most popular analytical method for allergen determination, however it has some disadvantages in the use of antibody. Recently, mass spectrometry-based methods gains popularity because it can overcome the disadvantages of antibody-based methods, also possesses advantages such as high throughput, and ability to provide molecular evident etc. Quantification of plant allergens, especially fruit allergens, which have low protein contents, is difficult to achieve, compared to other food allergens. Kiwi fruit (Actinidia deliciosa) is one of the popular fruits in Taiwan. It ranks the second highest among the imported fresh fruits.
Kiwi fruit is also an allergenic fruit, of which allergic popularity is rated sixth among food allergens in Europe and tenth in Taiwan. Many governments suggest allergenic labeling on food label. In our study, we choose kiwi fruit as an example to develop a MS-based quantification method for fruit allergen.
Our results consisted of three major parts. In the first part, we have developed and optimized sample preparation methods for mass spectrometry analysis of fruit allergen using kiwifruit as a model sample. We have developed two fruit optimized phenol
extraction methods, including optimized phenol extraction method (OPE) and micro phenol extraction method (MPE). Both methods had yielded a high extraction efficiency. OPE had a higher protein extraction weight, while MPE had some other advantages such as low cost, fast, simple and was well compatible with further mass spectrometry sample preparation steps. Meanwhile we have optimized trypsin in- solution digestion method and confirmed efficiency as well as isotopic effect of stable isotope dimethyl labeling method. We have also combined different methods of sample preparation steps into one continuous, low intra-method conversion loss process. In the second part, we have established the method of mass spectrometry for allergen quantification of kiwifruits. First, we used the results in the test of Q-ToF to decide the target allergens (Act d 1, Act d 5, and Act d 11) and representative peptides (AD1, AD5, and AD11). Then, we decided the quantitative and qualitative daughter ions of representative peptides on QqQ MS while establishing and optimizing parameters of LC and MS. After validation of our methods, AD1H and AD5H we found to have satisified validation characteristics, including range, accuracy, repeatability, instrumental quantification limit, instrumental detection limit, specificity, and linearity.
We have successfully developed and validated mass spectrometry-based allergen quantification method for kiwifruits under the MRM mode. In the third part, we have compared eight common kiwi fruit products to study the influence of different process
methods on kiwi allergens. We have determined the allergen contents of Act d 1 and Act d 5 in all samples, which indicated that our method was robust and could be applied to some other common fruit products. We found that allergen contents were significantly reduced under long time heat processing.
In conclusion, we have developed two fruit optimized phenol extraction methods, while MPE was combined with rest of the sample preparation methods into one continuous MS sample preparation process. In addition, we have developed a MS-based allergen quantification method for kiwifruits and its derivative products. Our study not only could be an example of analytical method development of food allergen, but also could be applied to improve food safety.
Keywords: dimethylation, QqQ MS, allergen, absolute quantification, Actinidia deliciosa
大綱
致謝 I 摘要 III Abstract V 大綱 VIII 表目錄 XII 圖目錄 XIII 縮寫表 XIV
壹、 前言... 1
食品過敏... 1
食品過敏的成因與發生情形... 1
過敏原種類... 4
常見的食品過敏原... 10
食品過敏與食品安全... 11
食品過敏原閾值... 12
食品過敏原分析方法... 13
1. 以免疫為基礎之食品過敏原分析法... 13
以核酸為基礎之食品過敏原分析方法... 15
以質譜為基礎之食品過敏原分析方法... 16
奇異果... 17
1. 歷史與品種... 17
臺灣奇異果現況... 18
營養價值... 18
產業應用... 19
奇異果過敏... 19
奇異果過敏原種類... 20
質譜技術... 23
用於定量分析之質譜技術... 23
SRM/ MRM 技術之優勢 ... 24
胜肽標記方法... 26
二甲基穩定同位素標記法... 29
利用二甲基同位素標記法搭配 MRM 進行過敏原之絕對定量 ... 32
質譜分析過敏原之萃取法... 33
1. 三氯乙酸-丙酮法 (trichloroacetic acid – acetone, TCA-A) ... 34
2. 苯酚蛋白質萃取法 (phenol protein extraction) ... 34
3. 分子量止透 (cutoff) 過濾法 ... 35
4. 其他... 36
食品化學檢驗方法之確效... 36
1. 分析方法之確效種類... 36
分析方法之確效方法... 37
研究動機與目的... 41
研究架構... 42
貳、 材料與方法... 44
藥品... 44
器材... 44
實驗方法... 45
奇異果產品製作法... 45
奇異果蛋白質萃取法... 45
蛋白質定量法... 48
蛋白質定性法... 48
蛋白質消化法... 49
胜肽標記與除鹽法... 51
蛋白質質譜定性... 51
目標胜肽選擇與合成... 52
超效液相層析三重四極柱質譜儀... 53
方法確效... 54
參、 結果... 55
一、 樣品前處理法... 55
苯酚蛋白質萃取法選擇... 55
苯酚蛋白質萃取法優化... 55
溶液內胰蛋白酶消化法之優化... 57
穩定同位素二甲基標記法效率確認... 57
分析方法建立... 58
目標過敏原之選擇- Q-ToF 複雜蛋白質定序結果 ... 58
代表胜肽選擇... 58
MS 參數設定 ... 59
LC 梯度設定 ... 60
校正曲線... 60
方法確效試驗... 61
不同加工處理對奇異果過敏原之影響... 62
不同加工處理對奇異果過敏原之影響... 62
肆、 討論... 64
一、 樣品前處理法... 64
各萃取法 SDS-PAGE 條帶差異之探討 (圖一) ... 64
苯酚法重複萃取效率探討... 64
苯酚蛋白質萃取法優化... 65
溶液內胰蛋白酶消化法之優化... 67
穩定同位素二甲基標記法效率確認... 69
分析方法建立... 70
Q-ToF 複雜蛋白質定序結果 ... 70
代表胜肽選擇... 71
MS 參數設定 ... 72
標準曲線... 73
不同加工處理對奇異果過敏原之影響... 74
不同加工處理對奇異果過敏原之影響... 74
結論與未來展望... 75
伍、 參考文獻... 76
表目錄
表 一 多次苯酚蛋白質萃取法之萃取效率... 96
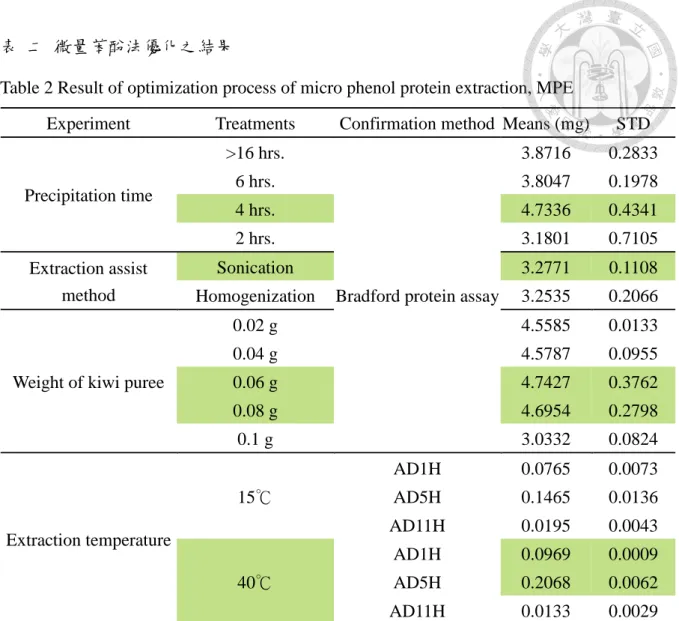
表 二 微量苯酚法優化之結果... 97
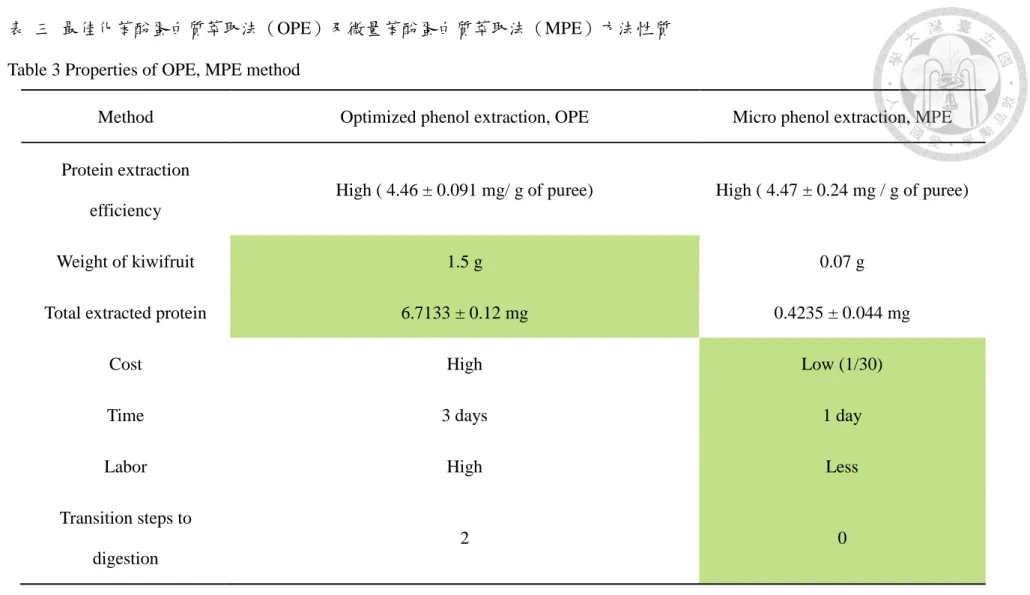
表 三 最佳化苯酚蛋白質萃取法(OPE)及微量苯酚蛋白質萃取法 (MPE)方法性質 ... 98
表 四 胰蛋白酶溶液內消化方法之優化... 99
表 五 複雜蛋白質定序辨認到之奇異果蛋白質... 100
表 六 代表胜肽之篩選條件... 104
表 七 目標過敏原之複雜蛋白質定序辨認到之胜肽結果... 105
表 八 候選胜肽之 BLAST 結果 ... 106
表 九 目標過敏原與代表胜肽之性質... 107
表 十 代表胜肽及其二甲基標記(氫和氘)胜肽之性質... 108
表 十一 代表胜肽之定量子離子性質... 109
表 十二 代表胜肽之定性子離子性質... 110
表 十三 AD1H 之確效結果 ... 111
表 十四 AD5H 之確效結果 ... 112
表 十五 AD11H 之確效結果 ... 113
表 十六 代表胜肽測定方法之確效性質... 114
圖目錄
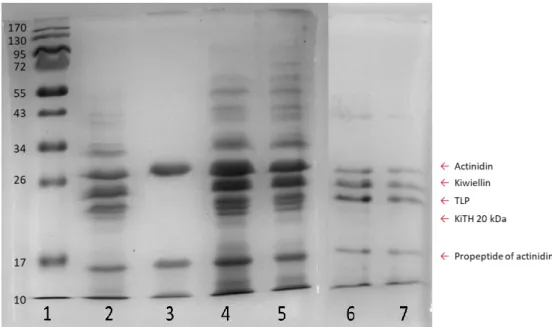
圖 一 不同蛋白質準備法之 SDS-PAGE ... 115
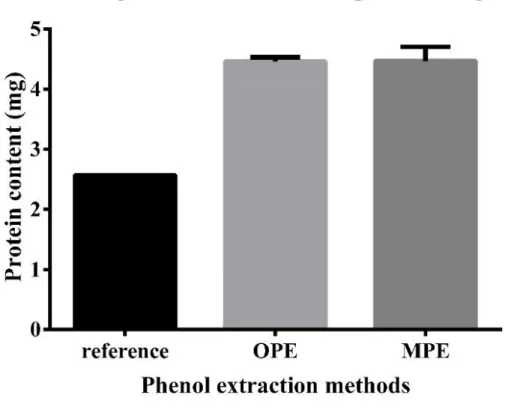
圖 二 文獻之苯酚蛋白質萃取法、最佳化苯酚蛋白質萃取法 (OPE) 及 微量苯酚蛋白質萃取法 (MPE) 萃取效率比較 ... 116
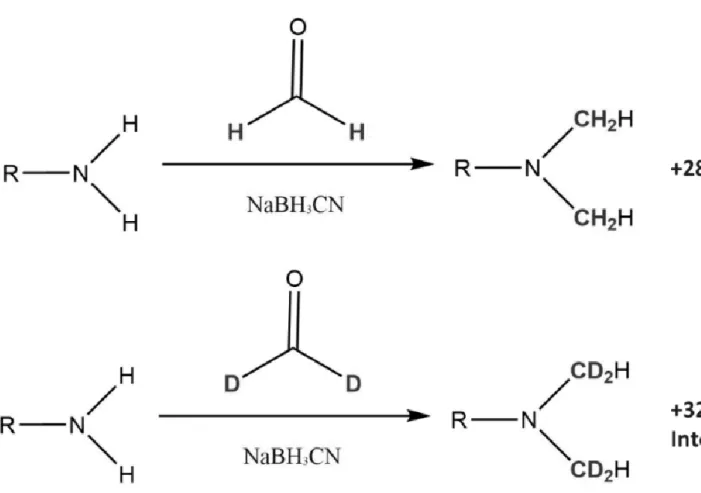
圖 三 二甲基同位素標記法氫與氘標記之反應式及其產物... 117
圖 四 代表胜肽之同位素效應... 118
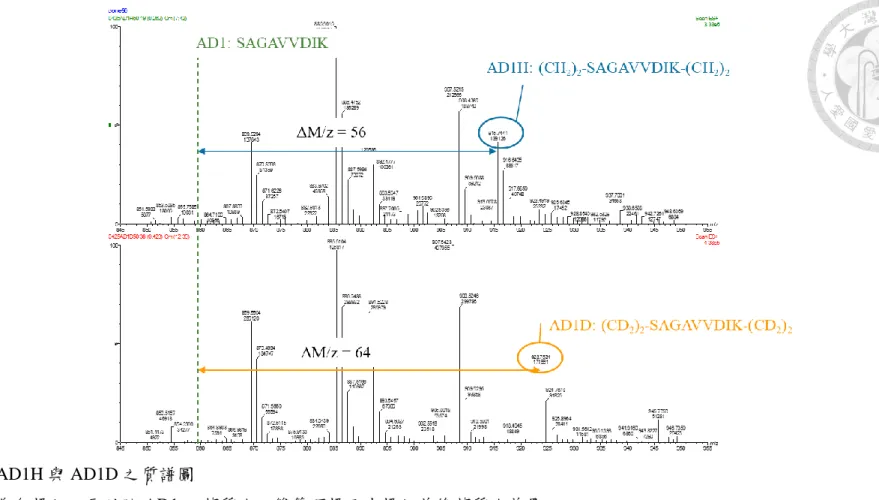
圖 五 AD1H 與 AD1D 之質譜圖 ... 119
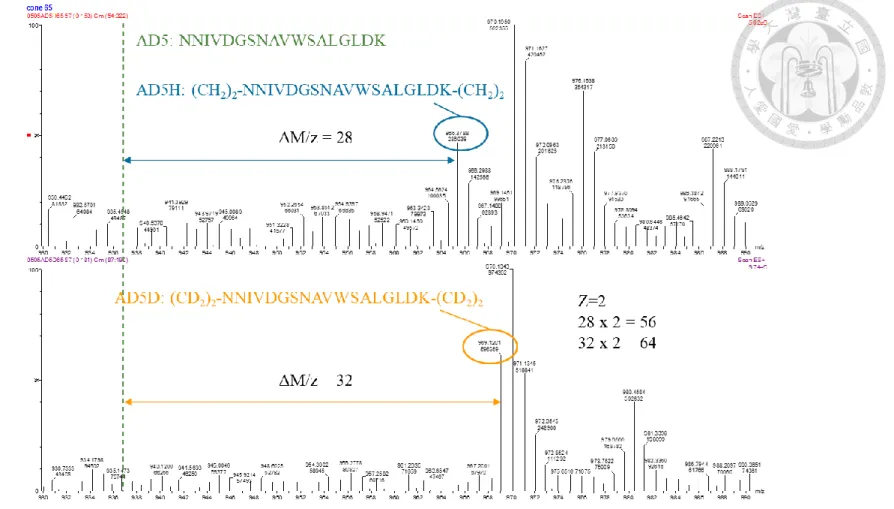
圖 六 AD5H 與 AD5D 之質譜圖 ... 120
圖 七 AD11H 與 AD11D 質譜圖 ... 121
圖 八 蛋白質複雜定序時使用之液相層析梯度圖... 122
圖 九 目標過敏原之複雜蛋白質定序覆蓋率... 123
圖 十 代表胜肽定量子離子之二次質譜圖... 124
圖 十一 代表胜肽定性子離子之二次質譜圖... 125
圖 十二 液相層析梯度圖之殘留量確認... 126
圖 十三 液相層析梯度圖與標記胜肽之滯留時間... 127
圖 十四 AD1H 胜肽之校正曲線與殘差圖 ... 128
圖 十五 AD5H 胜肽之校正曲線與殘差圖 ... 129
圖 十六 AD11H 胜肽之校正曲線與殘差圖 ... 130
圖 十七 AD1H 於適用範圍之線性 ... 131
圖 十八 AD5H 於適用範圍之線性 ... 132
圖 十九 AD11H 於適用範圍之線性 ... 133
圖 二十 每克經過不同加工處理後之奇異果中 Act d 1 之含量 ... 134
圖 二十一 每克經過不同加工處理後之奇異果中 Act d 5 之含量 ... 135
縮寫表
縮寫 全名
AD1 Signature peptide of Act d 1
AD1H(D) Dimethyl hydrogen (deuterium) labeled signature peptide of Act d 1 AD5 Signature peptide of Act d 5
AD5H(D) Dimethyl hydrogen (deuterium) labeled signature peptide of Act d 5 AD11 Signature peptide of Act d 11
AD11H(D) Dimethyl hydrogen (deuterium) labeled signature peptide of Act d 11 AQUA Absolute Quantification
DTT Dithiothreitol
ELISA Enzyme-Linked Immunosorbent Assay ESI Electrospray Ionization
ICAT Isotope-coded Affinity Tag
ICH The International Conference on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use
iTRQ isobaric Tag for Relative and Absolute Quantification IUIS International Union of Immunological Societies MPE Micro Phenol Extraction
MRM Multiple Reaction Monitor OPE Optimized Phenol Extraction
QToF Quadruple-Time of Flight mass spectrometry
SILAC Stable Isotope Labeling by Amino acids in Cell culture TEAB Triethylammonium bicarbonate
QqQ MS Triple Quadruple Mass Spectrometry
壹、 前言
食品過敏
食品過敏的成因與發生情形
食品過敏是指人體對某些特定食品中的蛋白質成份,所產生的非預期免疫反 應 (adverse immune response),而且定義上與食品的不耐受性、藥物反應及毒物 反應都不相同[1-3]。
(1) 食物過敏的發生機轉
食品過敏的發生,通常因為人體的免疫系統錯將食品中某些蛋白質,誤認為 是對人體有害的外來成份,特別是當某些可耐消化的食品蛋白質片段,受到 IgE 抗體所辨識之時,就可能引發後續的免疫反應,特別是區域性的發炎反應,因而 引起食品過敏。食品過敏可就 IgE 的參與與否分為二類,有時此兩類反應可能 同時發生[2, 4, 5]。
a IgE 參與之食品過敏 (IgE-mediated food allergy)
這是傳統認為的食品過敏反應,通常在攝食過敏原後兩小時內發生。當人體 首次接觸外來過敏原時, B 淋巴球受過敏原活化而分化後,製造結構相對應此 外來抗原的 IgE 抗體;當人體再次接觸過敏原時,可辨識此過敏原之特異性 IgE 會與過敏原結合,並進一步與顆粒性球細胞 (肥大細胞、嗜酸性與嗜鹼性白血球) 結合,導致顆粒性球反應並釋放大量例如組織胺之免疫趨化物質,引起食道、胃、
腸道甚至是其他組織例如皮膚與呼吸道的發炎反應[6]。
b 非 IgE 參與之食品過敏 (non-IgE-mediated food allergy)
非 IgE 參 與 之 食品 過 敏 又 稱 為 細 胞參與 之 食 品過 敏 (cell-mediated food allergy),此類食品過敏反應的發生機轉尚不完全清楚,目前已知可能有 T 細胞 或是嗜酸性球的參與[7],也可能與新生嬰兒腸道淋巴組織發育時,飲食所引起的 免疫反應與耐受反應(tolerance),兩者間的平衡有關。非 IgE 參與之食品過敏,
因為涉及 T 細胞及其他細胞的活化,通常發生較慢,於飲食後 4-48 小時後才 發生,例如嬰兒餵食數小時後的出血性腹瀉、腹脹、血便;或是例如成人攝食海 鮮後造成暈眩、噁心、抽續、嘔吐反應。另一方面,食品不耐受性(food intolerance) 症狀與非 IgE 參與之食品過敏類似,但是食品不耐受性,例如乳醣不耐受性及 藥物反應,因未涉及免疫反應,因此並不屬於食品過敏之範疇[8]。
由於腸道同時肩負接觸外物、吸收養分、抵禦病原的複雜功能,人體為了避 免對所有攝取的外來食品蛋白質產生過敏反應,腸道免疫系統應對外來無害之食 品成份具口服耐受性 (oral tolerance),可能與後天調節性 T 細胞 (adaptive Foxp3+CD4+ regulatory T cells, iTreg) 相關,抑制腸道免疫細胞對食品的免疫反 應而產生耐受性[4],以避免發生食物過敏。然而,食品過敏患者不具有或失去口 服耐受性的原因,至今仍未完全清楚。
(2) 食品過敏的臨床症狀與流行病學研究
一般 IgE 參與之食品過敏反應,通常在兩小時內發生,可能引起消化道、皮 膚或呼吸道發炎,並可能發生造成生命危險的全身性過敏性休克 (anaphylaxis),
常見症狀包括花粉食物症候群、食物蛋白誘導大小腸結腸炎、食物誘導直腸結腸 炎、嗜酸性球腸胃病鼻結膜炎、過敏性氣喘、口腔過敏症候群或異位性皮膚炎 (atopic dermatitis) 等[8, 9]。非 IgE 參與之食品過敏的發生較為緩慢,引起小腸 或結腸炎,患者常出現嘔吐、腹部絞痛、腹瀉等症狀;亦有過敏原會同時引發兩 種反應[8]。食品過敏除了本身症狀造成的自身困擾之外,亦會影響病患及其家庭 之整體活力及社交活動[10, 11]。
在流行病學上,約 33%的過敏反應主要是由食物誘發, 且與遺傳、地域、食 物種類、飲食習慣等多種因素有關[12]。大約有 25% 的美國民眾自認對某些食 物過敏,但是經由病史及實驗上可以確認過敏原的比例,在嬰幼兒大約是 2-8%,
在成人大約是 2%[6]。世界過敏組織指出,三歲以下之嬰幼兒約有 8% 對食品過 敏,若是對患有濕疹的兒童而言,可能 30%同時對食品過敏;大約 2.5% 的新 生兒對牛奶過敏,在一歲內對牛奶過敏的幼兒,可能 80% 直到五歲都對牛奶過 敏,25% 直到二十歲仍對牛奶過敏,而且其中 60% 屬於 IgE 參與之食品過敏 反應[13]。另一方面,食品過敏好發於嬰幼兒或是其他過敏疾病之患者,例如嬰 幼兒至少有 35% 發生過 IgE 參與之食品過敏反應, 6% 的幼童發生過由食物引 起的哮喘或是過敏性濕疹,而人類總人口數的 2.5% 對食品過敏[14]。
(3) 交叉反應與食品過敏
免疫系統的特性,在於能夠辨識自身的物質 (self) 與自身以外的異物 (non- self),在生物體內擔任監視與防衛的角色。免疫反應可分為「先天性免疫反應」
和「後天性免疫反應」兩大類,前者不具有抗原特異性,只要有異物存在就能引 發;後者則具有抗原特異性,針對不同的異物產生不同的反應,需要淋巴球的參 與,例如不同的 B 淋巴球株,可分別受特定結構的抗原活化,分化為抗原特異 性的記憶性 B 淋巴球與漿細胞 (plasma cells), 其中漿細胞職司產生抗體,以中和 (neutralize) 或吸附 (opsonize) 異物。
當免疫系統無法鑑別兩種抗原,同時對兩種結構類似的不同抗原均發生反應,
稱為交叉反應 (cross reaction)[15]。交叉反應在免疫與醫學上非常重要,例如金 鈉醫師 (Edward Jenner, 1749~1823) 即是利用免疫系統的交叉反應,以牛的天花 作為抗原免疫個體,讓個體對人的天花產生免疫反應。然而,交叉反應也可能引 起非預期的不良反應,食品過敏就是典型的例子,倘若是致敏 (sensitization) 的 抗原與食品中的蛋白結構相似時,就可能引起食品過敏反應。
臨床上食品過敏的首例於 1942 年被提出,當時發現受花粉過敏原致敏的個 體,有可能對部份的新鮮水果產生過敏症狀[16]。後續的研究發現,此一過敏的
機制與 IgE 抗體有關,個體第一次接觸花粉過敏原時,可能活化 B 淋巴球產生 可辨識花粉過敏原的特異性 IgE; 當人體第二次接觸結構近似的過敏原例如蘋果 Mal d 1 蛋白時,因為花粉過敏原與蘋果 Mal d 1 蛋白的結構類似,因此個體內 特異性的 IgE 便會與蘋果 Mal d 1 蛋白結合,並進一步與顆粒性球細胞 (肥大 細胞、嗜酸性與嗜鹼性白血球) 結合,導致顆粒性球反應並釋放大量例如組織胺 之免疫趨化物質,於是蘋果便可能因為交叉反應,造成個體過敏引起發炎反應。
過敏原種類
現在 Pfam (蛋白質家族, protein family)資料庫共記載了 16,295 個蛋白質家族 [17]。蛋白質家族又再分群組成蛋白質超家族 (superfamily),而某些蛋白質家族 又再細分成蛋白質次家族 (subfamily)。SDAP (https://fermi.utmb.edu)過敏原蛋白 質結構資料庫將現今已知之過敏原分類於 Pfam 資料庫 16,295 家族中的 130 個家 族中。世界上最重要的過敏原蛋白可被粗略分類於 30 – 40 個蛋白質家族中。以 下介紹較重要的幾種家族及其之下之次家族與過敏原。[18]
表 a. 常見過敏原來源及其所屬之蛋白質家族
超家族 (superfamily) 家族 (family) 過敏原來源 醇溶蛋白 (prolamin) 穀醇溶蛋白 (cereal prolamins ) 穀類榖粒
雙 功 能 抑 製 劑 (bifunctional inhibitors)
穀類榖粒
2S 白蛋白 (2S albumins) 堅果、花生、豆類、種子 非特異性脂質傳送蛋白 (non-
specific lipid transfer proteins)
水果、堅果、花生、蔬菜、花 粉、膠乳
EF-hand polcalcins 樹、草、野草花粉
parvalbumins 魚
類肌動蛋白結合蛋白 肌動蛋白結合蛋白 (profilin) 樹、草、野草花粉、蔬菜、水
(profilin-like) 果、膠乳 類 原 肌 凝 蛋 白
(tropomyosin-like)
原肌凝蛋白 (tropomyosin) 甲殼類、軟體類及魚類寄生蟲 海 獸 胃 線 蟲 (Anisakis simplex)、螨類、 蟑螂
cupin 蠶豆球蛋白 (vicilins) 堅果、花生、豆類、種子
豆球蛋白 堅果、花生、豆類、種子
Bet v 1-like Bet v 1 殼斗目花粉、水果、蔬菜、豆
類、堅果
calycin 脂質運載蛋白 (lipocalins) 哺乳類、螨類、蟑螂
double-psi DPBB_1 草類花粉
beta-barrel pollen_allerg_1 草類花粉
(1) 醇溶蛋白家族 (prolamin superfamily)
醇溶蛋白超家族之名字來自於穀類穀粒中醇溶的儲藏性蛋白。此超家族特色 為具有 α-螺旋球狀結構域 (domain),此結構域含有保守的半胱胺酸殘基模式,
通常形成三至五個分子內雙硫鍵。除了保守的半胱胺酸模式之外不同家族間僅存 在少量的序列相似性。此超家族包含穀類醇溶的儲藏性蛋白,如麥膠蛋白 (gliadins)與麥蛋白 (glutenins)及其他雙硫鍵豐富的蛋白質家族,如雙功能抑製劑 (bifunctional inhibitors)、2S 儲藏性白蛋白與非特異性脂質傳送蛋白 (nonspecific lipid transfer proteins)。
(2) 穀醇溶蛋白 (cereal prolamins)家族
穀醇溶蛋白為在穀類穀粒中大量存在之種子儲藏性蛋白,與其他低分子量的
超家族成員相比,其 α-螺旋結構域中插入了一段重複性序列[19]。麥膠蛋白
(gliadins)與麥蛋白 (glutenins)為穀醇溶蛋白家族代表性之成員。麥膠蛋白可溶於
醇類並被分類為α/β-、γ-、ω-麥膠蛋白。麥蛋白為多聚體蛋白彼此間由雙硫鍵結
合。其分為高分子量與低分子量兩群[20]。
小麥(Triticum aestivum)含有一些過敏性穀醇溶蛋白。Tri a 19 為 ω-5 麥膠蛋 白、Tri a 21 為 α/β-麥膠蛋白、Tri a 26 為高分子量之麥蛋白。
(3) 雙功能抑製劑 (bifunctional inhibitors)
與麥醇溶蛋白一樣雙功能抑製劑僅存在於穀類穀粒。這些過敏原藉由呼吸系 統吸入麵粉或藉由消化系統食入含有小麥、大麥或米造成過敏。其為 12 到 16 kDa 之蛋白質,由 4 到 6 個雙硫鍵組合而成[21]。單體、異雙聚體和異四聚體形 態之過敏原藉由其次單元的聚合度來分辨。雙功能抑製劑為造成烘焙師哮喘之主 因,並同時是植物食品之過敏原[22]。
Hor v 15 為大麥之單體 α-澱粉酶抑制劑,Tri a 28 及 Tria a 29 分別為小麥
之二聚體及四聚體α-澱粉酶抑制劑。
(4) 2S 白蛋白
2S 白蛋白為植物主要一群種子儲藏性蛋白質,許多重要之種子或堅果過敏 原為 2S 白蛋白。多數 2S 白蛋白先合成成單鍊的蛋白再被切成一個小的及一個 大的次單元。兩個次單元藉由 4 – 5 個雙硫鍵組合成緊密的 α-螺旋分子[23]。
過敏性 2S 白蛋白包含花生之 Ara h 2 及 Ara h 6、巴西果之 Ber e 1、榛果之 Cor a 14、英國胡桃之 Jug r 1、芝麻子之 Ses i 1 以及黃芥末之 Sin a 1。
(5) 非特異性脂質傳送蛋白 (nonspecific lipid transfer proteins, nsLTP)
nsLTP 之功能為調節囊泡與細胞膜間磷脂質之傳送,許多 nsLTP 也擔任抵抗 防禦真菌及細菌之角色。過敏性 nsLTP 為一大群耐熱及消化之蛋白質[24]。nsLTP 高濃度存在於水果之表皮組織。另外過敏性 nsLTP 存在於堅果、種子、蔬菜及橡 膠樹 (Hevea brasiliensis)之乳汁。
薔薇科水果之 nsLTP 包含蘋果之 Mal d 3 及桃子之 Pru p 3,堅果中具代表 性之過敏性 nsLTP 為榛果之 Cor a 8 與胡桃之 Jug r 3。花粉類 nsLTP 包含 法國梧桐之 Pla a 3 與北艾之 Art v 3。玉米中之 nsLTP 為 Zea m 14。
(6) EF-hand 家族
許多鈣結合蛋白皆具有一個保守性結構域,此域為 12 個胺基酸組成之鈣結 合環,此環包圍於由 12 個胺基酸組成之 α-螺旋之兩側[25]。這些蛋白質之功能 包括訊號傳遞及鈣離子的緩衝或傳送。
(7) Polcalcins 家族
Polcalcins 為 9-kDa 之鈣結合花粉蛋白,其生理功能尚不清楚。通常 polcalcins 含有兩個 EF-hand 結構域,一些 polcalcins 相關之過敏原含有三或四個 EF-hand 結構域[26]。Polcalcins 為少數但高交叉反應性的花粉過敏原來自多個植物的科。
過敏性 polcalcins 包含來自樺樹之單體的 Bet v 4,來自貓尾草的二聚體 Phl p 7 及來自藜 (Chenopodium album)之四聚體 Che a 3。
(8) Parvalbumins 家族
Parvalbumins 為 12 kDa 並含有兩個 EF-hand 結構域的蛋白。他們存在於脊 椎動物快速抽動的肌肉纖維中,在肌肉放鬆時與鈣離子結合。來自魚與兩生類之 parvalbumins 為誘發多數魚類過敏患者的主要過敏原[27]。
過敏性 parvalbumins 包含大西洋鱈魚的 Gad m 1 與大西洋鮭魚的 Sal s 1 與 螃蟹中的 Cyp c 1。
(9) 類肌動蛋白結合蛋白家族 (profilin-like superfamily)
Profilin 為可以在所有的真核細胞中發現的小胞漿蛋白。Profilin 會與單體的 肌動蛋白和其他肌肉蛋白結合,並在細胞移動、胞質分裂及訊號傳遞時調節肌動 蛋白聚合作用之動力。來自高等植物的 profilin 組成了一個蛋白質家族,其蛋白 質間序列非常保守,在親緣關係非常遙遠的成員間序列相似性都至少有 75%[28]。
Profilin 專一之 IgE 反應通常與所有植物來源的同源物產生交叉反應,這些過敏 原被認定為花粉-食物過敏的一個風險因子[29]。
過敏性 profilin 包含北艾花粉之 Art v 4、樺樹花粉之 Bet v 2、橄欖花粉之 Ole e 2、柳橙之 Cit s 2、厚皮甜瓜之 Cuc m 2 及香蕉之 Mus a 1。
(10) 類原肌凝蛋白家族 (tropomyosin-like superfamily)
Tropomyosin 存在於肌肉及非肌肉細胞。於橫紋肌中其調解肌鈣蛋白複體與 肌動蛋白之互動以調節肌肉收縮。Tropomyosin 被認定是動物性食物像是甲殼類、
軟體類及魚類寄生蟲海獸胃線蟲 (Anisakis simplex)之過敏原[30]。Tropomyosin 同 時也是節肢動物 (塵螨、蟑螂)中之吸入性過敏原。Tropomyosin 之序列具很高的 保守性,說明了其在含有 tropomyosin 來源間的經常交叉反應[31]。
印度明對蝦中之 Pen I 1、德國蟑螂中之 Bla g 1 及家塵螨中之 Der p 10 為有 名之過敏性 tropomyosin。
(11) Cupin 家族 (cupin superfamily)
Cupin 超家族為一巨大且蛋白質生理功能差異非常大之超家族,其具有共同 起源並可被追朔從細菌至真核生物包含動物與高等植物。Cupin 超家族下具 57 個 蛋白質家族。最大的兩個家族為 7/8S 與 11S 種子儲藏性球蛋白,其為植物種子 中主要的成分為人類飲食中重要的蛋白質來源且同時為過敏原[32, 33]。
(12) 蠶豆球蛋白家族 (Vicilins family)
這些同三聚體蛋白分子量約 150 – 190 kDa。其次單元之組成差距非常大歸 因於其單體在蛋白質分解及醣基化過程之差異。
過敏性蠶豆球蛋白包含花生 Ara h 1、黃豆 Gly m 5、胡桃 Jug r 2 及芝麻 Ses i 3。
(13) 豆球蛋白家族 (Legumins family)
成熟之豆球蛋白由兩個三聚體形成一個六聚體。
過敏性豆球蛋白包含花生 Ara h 3、黃豆 Gly m 6、巴西堅果 Ber e 2 及蕎麥 Fag e 1。
(14) 類 Bet v 1 家族 (Bet v 1-like superfamily)
類 Bet v 1 超家族含有來自 1,452 個物種之 14,065 種蛋白質,其序列全部與 樺樹 (Betula verrucosa)花粉之主要過敏原 Bet v 1 相關。這些蛋白分散於 14 個蛋
白質家族。這些來自 14 個家族 (其中一個為 Bet v 1 家族)的蛋白都具有相同的 結構,此結構由 7 個反平行的 β-摺板及 3 個 α-螺旋組成[34]。至今過敏原被發現 存在於 Bet v 1 家族下 11 個次家族中的其中 3 個次家族。
(15) 十號致病性相關蛋白質次家族 (Pathogenesis-related protein subfamily 10, PR-10)
PR-10 蛋白質為 Bet v 1 家族下 11 個次家族中的其中一個。這些蛋白之表現 由病原體攻擊或非生物體逆境或由生長調節所引發。PR-10 蛋白於生殖組織中高 濃度表現,像是於花粉、種子及果實。許多樺樹花粉過敏患者對許多水果與蔬菜 產生過敏反應,是因為 Bet v 1 與植物食物中同源過敏原產生 IgE 交叉反應所造 成。多數 Bet v 1 相關之食物過敏原存在於特定植物科別:薔薇科 (蘋果、梨子、
核果)、繖形花科 (西洋芹、胡蘿蔔)、豆科 (黃豆、花生)。
(16) 成熟相關蛋白 /主要乳汁蛋白家族 (ripening-related proteins/major latex proteins family, RRR/MLP family)
RRR/MLP 蛋白家族為另一個 Bet v 1 家族之次家族。奇異果之 Act d 11 為第 一個來自 RRR/MLP 次家族之 Bet v 1 相關過敏原。
(17) 細胞分裂素專一結合蛋白家族(cytokinin-specific binding proteins subfamily, CSBP subfamily)
來自綠豆之 Vig r 6 為第一個來自 CSBP 次家族的 Bet v 1 相關過敏原。
(18) Calycin 家族 (Calycin superfamily)
Calycin 超家族由 16 個家族組成,儘管結構相似此超家族成員間之序列相似 性低。Calycin 結構基於八股 β-桶狀結構並可能與不同配體結合[35]。
(19) 脂質運載蛋白 (Lipocalins)
Lipocalins 為輸水性小分子之運載者,像是脂質、固醇類激素、後膽色素類 (bilins)及類視色素。Lipocalins 摺疊成一八股反平行 β-桶狀結構。此家族之過敏 原包含β-乳球蛋白、哺乳動物皮屑過敏原及細胞質脂肪酸結合蛋白[36, 37]。β-乳
球蛋白為反芻物種之主要乳清蛋白。Bos d 5 為牛乳中主要之過敏原。其他物種 與牛乳蛋白也存在交叉反應。Lipocalins 組成動物皮屑過敏原的一大部分。細胞 質脂肪酸結合蛋白與細胞質外 lipocalins 及 β-乳球蛋白間具遙遠的親緣關係。其 為塵螨與蟑螂中之小部分過敏原。
哺乳動物來源之過敏性 lipocalins 為來自馬之 Equ c 1、牛之 Bos d 2、狗之 Can f 1 與 Can f 2 及貓之 Fel d 4。
(20) Double-psi beta-barrel superfamily, DPBB superfamily
β-桶狀結構為蛋白質結構中常見之元素。不同股數之桶狀結構能夠有不同的 股間連結或拓撲 (topology)。其中由六個 β 股特殊排列而成之拓撲稱為 double- psi beta-barrel (DPBB)[38]。第一組之草類花粉過敏原存在於 DPBB 超家族下的 DPBB_1 家族中。其與 β-胞壁擴張蛋白具很近的親緣關係,兩者皆含有兩個結構 域。第一個結構域為六股的 DPBB 拓撲,與醣苷水解酶家族的 45 個蛋白有遙遠
的親緣關係。第二個結構域由類似免疫球蛋白摺法的兩個堆疊β-摺板組成,此域
同時也稱為第二組草類花粉過敏原結構域[39]。胞壁擴張蛋白之活性通常與生長 細胞之細胞壁鬆動活性相關,包含花粉管穿過柱頭及花柱[40]。第二與第三組草 類花粉過敏原同屬於花粉過敏 1 家族 (Pollen_allerg_1),並與 β-胞壁擴張蛋白之 第二結構域相關。
第一組的草類花粉過敏原為黑麥草之 Lol p 1、貓尾草之 Phl p 1 與草地早熟 禾之 Poa p 1。第二/三類草類花粉過敏原為鴨茅之 Dac g 2 與 Dac g 3、黑麥 草之 Lol p 2 與 Lol p 3 及貓尾草之 Phl p 2 與 Phl p 3。
常見的食品過敏原
食物過敏主要由食物內生性的蛋白過敏原所引起,無論動物性或植物性食物 都可能含有過敏原。因為食品過敏的種類及發生機制不同,因此造成兒童過敏及 成人過敏的食品,也不盡相同。90% 之食品過敏由八種食物群所造成,包含蛋 類、魚類、奶類、花生、貝類、黃豆、堅果類及小麥,為「八大過敏原」[41]。
文獻上,常引發兒童過敏之食物主要有牛奶、蛋、花生、大豆、小麥;而引起成 年人花粉相關食物過敏之食物則有水果、蔬菜、堅果、種子及魚貝類等[42-45]。
榮民總醫院吳子聰教授曾針對台灣常見的食物過敏進行研究,結果顯示蝦、蟹、
牛奶、蛋、花生是台灣地區最易引起過敏反應的第一級食物,第二級致敏食物為 芒果、其他海鮮,第三級致敏食物則是花枝、蛤蠣、魷魚、墨魚、螺、鱈魚、大 豆、小麥、奇異果等[46]。
食品過敏與食品安全
食品過敏 (food allergy) 是重要的食因性疾病 (foodborne illness) 之一,也是 現代非常重要的食品安全議題之一。美國國會在 2004 年通過「食物過敏原與消 費者保護法案」(Food Allergen Labeling and Consumer Protection Act of 2004, FALCPA 2004),後續 FDA 規定自 2006 年開始,所有包裝食品中,若含有主要 過敏原 (major food allergen) 之原料,及部份可能引起過敏的食品添加物,均應 予以標示[1]。除美國之外,在聯合國食品法典委員會 (CAC Food Standards 4.2.1.4)、歐盟(2000/13/EC, 2003/89/EC)、 英國(Directive 2006/142/EC)、日本 (食 品安全法, 食品衛生執行條例)、澳洲、紐西蘭、中國 (預包裝食品標籤通則) 等 國,均對食品過敏成份的標示,有不等的法令規範[1, 2],我國亦於 2014 年發布
「食品過敏原標示規定」,規定於 2015 年 7 月 1 日起市售有容器或包裝之食品含 有下列對特殊過敏體質者致生過敏之內容物,應於容器或外包裝上,顯著標示含 有致過敏性內容物名稱之醒語資訊,須標示之內容物包含蝦、蟹、芒果、花生、
牛奶、蛋及其製品,另公布「食品過敏原標示之建議事項」,建議得自願性標示
相關醒語,包含羊奶、魚類、頭足類、螺貝類、堅果種子類、含有麩質之穀類、
大豆、奇異果及其製品,與產品中亞硫酸鹽類或二氧化硫殘留量總計達 10 毫克 /公斤以上時。鑑於食品過敏對食品安全以及人體健康非常重要,及各國過敏原 強制標示之規定,有必要建立食品過敏原之分析平台。
食品過敏原閾值
現今食品過敏原標示僅以醒語標示過敏原之存在,而無法明確表達其代表之 風險,建立食品過敏原閾值能幫助解決當前法規對於食品過敏原標示之不確定性。
食品過敏原閾值之定義為誘發劑量或能夠引起過敏反應之最低攝食過敏原量 [47]。定義此閾值能為各個不同團體帶來效益:食物過敏患者及其照顧者、食品 產業、政策決定者及政府部門[47-49]。考慮到現行標示法規的不足,定義食品過 敏原閾值為重要之策略。現行法規,以美國「食物過敏原與消費者保護法案」
FALCPA 為例,其「可能含有」、「於同一設備製作」、「於同一工廠製作」之防範 性醒語標示造成了不確定性,因為這些標示無法明確闡明其含有主要過敏原之風 險 [50-53]。數據顯示許多過敏患者因各種原因忽略防範性醒語標示,原因包括:
先前曾安全的食用過此產品因此產生了此產品是安全的這種錯誤的假設;文字之 誤解 (將「可能含有」誤解與「於同一工廠製作」相同);或直接忽略其潛在風險 [51, 54]。這是個潛在的危險情形,考慮到這些產品通常含有百萬分之一水平的過 敏原[51-53, 55]。
然而食品過敏原閾值之建立具有潛在問題,包含食品過敏原閾值的代表性不 足、消費者對於此閾值之接受度不足、社會對於此閾值之信任度不足以及建立過 程之花費過大,其中花費包含評估、偵測及標示花費高於其帶來之利益。目前食 品產業主要依靠酵素免疫分析法 (Enzyme-Linked Immunosorbent Assay, ELISA) 為基礎的過敏原檢驗方法來確效其過敏原控制計畫[51, 52, 55]。儘管於發展及使 用這些過敏原測試方法有大量的進展,仍需投入更多的努力於發展、提升及標準 化過敏原食品之檢驗方法,及降低方法之價格來擴大小型製造商對方法之使用及 接受度[53]。建立食品過敏原閾值能幫助解決當前法規對於食品過敏原標示之不 確定性,然而在建立此閾值系統中仍缺乏價格划算、可信、高通量之標準化的過 敏原分析方法。
食品過敏原分析方法
現今用於偵測食物中潛在過敏原的方法以過敏原本身或者是能夠代表過敏 原存在的標記為目標。這些食品標記通常於過敏食品中具有蛋白質專一性或是為 有特徵之 DNA 片段[56]。以蛋白質為目標之方法通常涉及免疫學步驟,像是放 射性過敏原吸附試驗 (radio-allergosorbent test; RAST)、酵素過敏原吸附試驗 (enzyme allergosorbent test; EAST) 、 火 箭 免 疫 電 泳 法 (rocket immuno- electrophoresis; RIE) 、 免 疫 墨 點 法 (immunoblotting) 及 酵 素 鏈 結 免 疫 吸 附 法 (Enzyme-linked immunosorbent assay; ELISA),現今 ELISA 法被用於例行的食品 分析,因其具高準確性、簡單處理及於步驟標準化有很高的潛力。而以 DNA 為 主之方法則利用聚合酶連鎖反應 (Polymerase chain reaction; PCR)來放大特定之 DNA 片段,藉由即時聚合酶連鎖反應(Quantitative real time polymerase chain reaction; qPCR/qrt-PCR)可以得到高精確度之結果[56]。而於過去幾年間,於開發 食品過敏原分析方法時,開發者使用質譜儀系統以解決現今常用方法之主要缺點 [57]。
1. 以免疫為基礎之食品過敏原分析法
酵素鏈結免疫吸附法 (Enzyme-linked immunosorbent assay; ELISA)為現今最 廣泛使用於例行測定及篩選食品過敏原的分析技術。大體上 ELISA 可依偵測原 理為三明治或競爭法分為兩種形式。三明治式 ELISA (sandwich ELISA; s-ELISA) 利用對目標蛋白(過敏原或食品標記)專一的固定式抗體來捕捉目標分析物,目標
分析物則再被另一抗原專一並帶有酵素標記的抗體辨認,形成「三明治」。於偵
測步驟時此標記於抗體之酵素與特定基質反應產生有顏色之產物,有顏色之產物 之吸光值與分析物之濃度成直接的比例關係。而在三明治法因抗原必須要有一個 以上之抗原決定位,此方法只能應用於大分子像是蛋白質。因此 s-ELISA 為最常 見的食品過敏原免疫偵測法[58]。
儘管 ELISA 具高靈敏性且專一性並能夠產出定量結果[56],但是 ELISA 法 具下述缺點。
(1) 可偵測之過敏原目標受限制
雖然八大過敏原中大多數皆有可利用的 ELISA 套組,但多數的套組只針對 來自單一食品中的單一過敏原,舉例來說,牛奶過敏原之商業 ELISA 套組之偵 測目標僅包含酪蛋白 (casein)、β-乳球蛋白 (β-lactoglobulin)及總過敏原含量 (酪
蛋白及β-乳球蛋白),而其他牛奶中已知的過敏原像是同為主要過敏原的 α-乳球
蛋白 (α-lactoglobulin) 則無被列為目標。
(2) 對抗體暸解程度不足
抗體為主之方法需有單株亦或多株抗體,多數抗體為商業可及,但多數抗體 之特性尚未被清楚了解。使用抗體為主之方法時可能會產生交叉反應,造成潛在 偽陽性結果。複雜之基質可能會包含影響物質像是多酚類或單寧,可能會與蛋白 質或抗體反應或結合。此外食品加工或樣品準備過程可能會修飾過敏原,造成無 法被目標抗體辨認產生潛在偽陰性結果。食品之熱加工過程可能會破壞或創造結 構性抗原決定位,造成抗體辨認上之錯誤[59]。
(3) 不同套組結果差異大
文獻[60-62]比較不同 ELISA 套組間之分析結果,其準確度、精密度甚至適 用範圍皆有明顯缺陷。因缺乏通用之標準品或參考物質,故不同 ELISA 套組間 相比較非常困難。
(4) 不具指紋不能夠鑑定
ELISA 套組針對特定過敏原目標進行偵測或定量,然而受限於其抗體與過 敏原抗原決定位的專一性,其對非目標之蛋白不具分辨性,故 ELISA 法不能用 做鑑定過敏原之方法。
這些不足之處明顯表明了於食品過敏原分析定量方法上需要其他方法之迫 切需求[57]。
以核酸為基礎之食品過敏原分析方法
聚合酶連鎖反應 (PCR)用於擴增一小段已知的 DNA 片段,可能是單個基因,
或僅是某個基因的一部分。一般的聚合酶連鎖反應由 20 到 35 個循環組成,每個 循環包括以下 3 個步驟:
(1) 變性
利用高溫(93-98℃)使雙鏈 DNA 分離。高溫將連接兩條 DNA 鏈的氫鍵打斷。
在第一個循環之前,通常加熱長一些時間以確保模板和引物完全分離,僅以單鏈 形式存在。該步驟時間 1-2 分鐘,接下來機器就控制溫度進入循環階段。
(2) 接合
在 DNA 雙鏈分離後,降低溫度使得引物可以結合於單鏈 DNA 上。此階段的溫 度通常低於引子熔點 5℃。
(3) 延伸
DNA 聚合酶由降溫時結合上的引子開始沿著 DNA 鏈合成互補鏈。此階段的溫 度依賴於 DNA 聚合酶。該步驟時間依賴於聚合酶以及需要合成的 DNA 片斷長 度。
即 時 聚 合 酶 連 鎖 反 應 (quantitative real time polymerase chain reaction;
qPCR/qrt-PCR) 是一種在 DNA 擴增反應中,以螢光染劑偵測每次聚合酶鏈鎖反 應(PCR)循環後產物總量的方法,其偵測範圍廣、靈敏度高、準確、專一及快 速。
qPCR 與 ELISA 法相比因多數情況下 DNA 比起蛋白質較不易受到熱變性影 響,因此偽陰性結果可被避免。然而與蛋白質相同,食品原料可能會與 DNA 反 應並影響 PCR 反應的進行。然而此方法並無法提供蛋白質含量的直接資訊,因 其仍缺乏校正參數來將 DNA 的重量、濃度或套數 (copy number)轉換為蛋白質
[59]。
以質譜為基礎之食品過敏原分析方法
以質譜系統為基礎之食品過敏原分析方法於近年快速且蓬勃發展。質譜儀 (mass spectrometry, MS)主要由三個部分組成:離子源、質量分析器以及偵測器。
最 為 常 見 之 兩 種 離 子 源 為 基 質 輔 助 雷 射 脫 附 電 離 (Matrix-assisted laser desorption/ionization, MALDI)及電灑游離 (electrospray ionization, ESI)。質量分析 器於蛋白質體學最常見的為飛行時間(time of flight, TOF)及離子阱 (ion trap, IT),
於定量上則以三重四極柱(triple quadruples, TQ/QQQ)最為廣泛利用。通常會再搭 配 樣 品 分 離 方 法 像 是 高 / 極 效 液 相 層 析 (High/Ultra performance liquid chromatography, HPLC/UPLC) 或是氣相層析 (gas chromatography, GC),於分析 食品過敏原通常使用極效液相層析。組合不同之層析法、離子源與質量分析器可 得混和式的質譜儀系統,像是 UPLC-ESI-qTOF、MALDI-TOF、UPLC-ESI-IT、
UPLC-ESI-TQ 等系統[59]。
蛋白質體學於質譜儀分析蛋白質時,需一系列的方法步驟包含以蛋白酶消化 蛋白質來產生胜肽並將其引導至質譜儀中進行分析。接著利用生物資訊工具及蛋 白質資料庫來分析質譜圖。藉由荷質比確認胜肽序列,而過敏原蛋白可利用其衍 生之胜肽序列與已知的蛋白相比對而確認。蛋白質定量分析時之步驟將於後面章 節詳細說明。
質譜儀系統具有許多優點,像是簡單的樣品準備方法、快速的分析時間及能 夠同時分析多個過敏原。質譜儀與其他同樣擁有低偵測極限 (limit of detection, LOD)與定量極限 (limit of quantification, LOQ)的方法相比較為強大且穩定,並能 夠簡單的自動化及標準化。另一優點是擁有更明確的標準,使得比較方法間與實 驗室間的結果更為容易。食品熱加工所產生之後轉譯修飾、蛋白質立體結構破壞
可能會影響 ELISA 法的靈敏性與專一性,而質譜法則受影響程度較小,因其偵 測較穩定之蛋白質一級結構。質譜法更能夠提供過敏原的胺基酸序列,並辨認後 轉譯修飾以及蛋白質亞型 (protein isoform)。總結來說,質譜儀系統於近幾年被 應用於開發食品過敏原之分析方法,其能夠解決現在已開發之方法的主要缺點,
如非特異性抗體-抗原反應與未知修飾的問題[59]。
奇異果
1. 歷史與品種
奇異果為獼猴桃屬 (Actinidia) 木質藤蔓植物的可食性果實。獼猴桃屬下有 許多物種與許多品種,各有不同的特性。奇異果又稱獼猴桃,原生於中國南方,
1904 年時奇異果種子被傳教士引進紐西蘭,被當地人稱為中國醋栗 (chinese gooseberry),因其嘗起來像醋栗且來自中國[63, 64]。而現今奇異果中最重要的綠 肉奇異果栽培種 ’Hayward’ 即是那批種子的直系後代,其於 1920 年代由 Hayward Wright 在艾文代爾 (Avondale) 地區發展出來。於 1950 年代紐西蘭開始 將奇異果出口至美國,但因美國正值冷戰時期,基於政治考量名稱奇異果 (kiwifruit) 被 提 出 並 接 受 [63] 。 於 2000 年 時 黃 肉 奇 異 果 栽 培 種 ‘Hort 16A‘ (ZESPRI®) 被引入市場,打破了奇異果只有綠肉的概念。至今許多國家都 有栽種奇異果,像是義大利、中國、智利、法國、希臘、日本與美國[64]。現今 市面上最常見的兩個栽培種為綠肉的美味獼猴桃 Actinidia deliciosa ’Hayward’ 與 黃肉的中華獼猴桃 Actinidia chinensis ‘Hort 16A’, (ZESPRI®)。市面上之其他栽培 種還有 A. deliciosa ‘Summer 3373’、A. chinensis ‘Jintao’或是俗稱奇異果莓的 A.
argute,或極可能於未來上市的 A. eriantha ‘Eriantha 96’ 和’Eriantha 114’[65]。
(1) 美味獼猴桃 Actinidia deliciosa ’Hayward’
栽培種 ’Hayward’ 之果實小呈蛋形,大小近大顆的雞蛋,果皮上有毛並呈現 褐色包覆著果肉,果肉為半透明且鮮綠色,於果肉中包覆著排狀排列的可食黑色 小種子,果肉具有撲鼻酸甜動人的香味[66, 67]。
(2) 中華獼猴桃 Actinidia chinensis ‘Hort 16A’
黃金奇異果栽培種 ‘Hort 16A’ 之形狀特殊,其具有突出的花柱頂,稱為「喙 (beak) 」。果實具有明亮的黃色果肉,包覆於光滑、無毛、青銅色果皮中,果肉 具有香甜和熱帶的味道[66, 67]。
臺灣奇異果現況
於 1980 年代起台灣開始進口奇異果,於市面上首次能購買到此特殊之水果,
於短短 40 年進口量每年持續成長,於 2017 年奇異果年進口重量已到 42,019 公 噸,年進口價值為 124,701 千美元,佔總生鮮冷藏水果進口價值之 16.9 %,為總 生鮮冷藏水果第二名,僅次於蘋果,為農產品進口值之第 24 名。隨新食物之引 入同時亦可能引入新的風險,其中對新食物之過敏症即是其中一個不可忽視之風 險,而在隨著進口量逐年增加,本國國民更亦暴露於奇異果過敏之風險下。
營養價值
與其他常見的水果相比不論是綠肉或黃肉的奇異果皆是營養密度高的水果,
其尤其富含維生素 C、E、K、葉酸、類胡蘿蔔素、鉀、纖維及數種植化素 (phytochemical)[67]。黃金奇異果搭配富含鐵的飲食是幫助輕微鐵元素缺乏的女 性增加的鐵含量的有效策略[68]。規律食用綠色或黃金奇異果可能對血脂組成有 良好的影響,例如增加高密度脂蛋白膽固醇 (HDL-C) 含量、減少總膽固醇和高 密度脂蛋白膽固醇比值 (TC/HDL-C) 及三酸甘油脂 (TG) 含量[69-72],同時具 有抗血栓的效果,如減少血小板聚集[69, 72, 73]。規律食用奇異果亦有抗高血壓 的效果,此效果來自於血管緊縮素轉化酶 (angiotensin-converting enzyme, ACE) 活性的抑制調控[69, 72, 73]。綠色奇異果亦能幫助消化,其能幫助有效率的消化 蛋白質、增加糞便體積與柔軟度及提供潤滑幫助排便[67]。規律食用綠色與黃色 奇異果能增加體內的抗氧化能力,保護身體受到內生性氧化的傷害[72, 74]。連續 四週每日食用黃金奇異果可減少幼童發生感冒或流感類的發生率,且於易發感冒 與流感類症狀的兩個族群(幼童與年長者)中減少感冒或流感類症狀的嚴重性[75,
76]。
產業應用
奇異果被大量應用於食品產業,做為果醬、果凍、糖漿、糕點之原料[77],
而又因其含有奇異果酵素 Actinidin,為一種半胱胺酸蛋白酶,故被利用為食品加 工助劑,例如肉質軟化劑[78, 79]、起司產業之乳凝結劑[80]。做為肉質軟化劑,
actinidin 水解效果雖不如番木瓜酵素 papain,但番木瓜酵素因活性太強常會造成 肉表面之過度軟化問題,此問題亦在鳳梨酵素 bromelain、無花果酵素 ficin 中發 生,故水解效果溫和之 actinidin 具有其優勢,包含無不良風味及不會產生肉面黏 糊現象,然而常需提高活性來達到同等柔軟度[81]。亦有研究成功將奇異果酵素 做為舌面生物膜移除劑[82],
奇異果過敏
將新的物質引入人類飲食中存在著風險,而奇異果就能做為此現象之範例。
自引入人類飲食後,第一起記載之奇異果過敏反應發生於 1981 年[83],至今已成 為十大食品過敏原之一。奇異果過敏之發生頻率於西方人口更高[84]。奇異果過 敏之症狀從輕微之口腔黏膜局部性反應至嚴重之系統性反應,系統性反應特別好 發於幼童[84]。其症狀嚴重性與致敏模式依地域而異,於全歐洲可觀察到許多不 同之致敏模式。奇異果過敏症並非同質單一之病症,於臨床可建立數個不同的子 族群[85]。奇異果過敏可歸因於單一致敏反應 (monosensitization) 或與其他過敏 原產生交叉反應 (cross reaction),主要產生交叉反應之過敏原為樺樹與草類花粉 [86]。
西方之奇異果過敏人口比例較高,於芬蘭研究調查食物過敏的盛行率,報告 中奇異果最常造成不良反應 (38.4%)[87]。而在法國學童中奇異果過敏人數僅次 於對常見之蛋與奶過敏人數[43]。在一個大型的歐洲人口研究調查報告中,奇異 果過敏為於歐洲前六常見之食物過敏[88]。於台灣食品過敏症盛行率之調查研究 結果,在 2086 位食品過敏人口中奇異果過敏之人數為 57 人 (2.7%),名列台灣
前十大食品過敏原之一[46]。而奇異果過敏又會因地區而有所不同,於聯合國 EUROPREVALL 報告中,來自冰島之病患相較於其他地區之病患更易產生嚴重 之奇異果過敏反應,並更常辨認到 Act d 1。在西、中、東歐地區奇異果過敏最常 伴隨樺樹花粉過敏,而相對應地區之患者主要之奇異果過敏原亦為與樺樹花粉相 關的 Act d 8;在南歐則最常伴隨草類花粉,主要過敏原則為 Act d 9,而 Act d 10 亦為此族群中比例較高之過敏原[65, 89]。
而不同之奇異果栽培種亦展現了不同之致敏性,一項於荷蘭與瑞典過敏患者 上進行的研究,其利用了三個不同種六個不同栽培種的奇異果進行針刺試驗 (prick-to-prick test),結果可將奇異果劃分為低致敏性 (Hort16A)、中致敏性 (Summer 3373、Jintao) 與高致敏性 (Eriantha 96、Eriantha 114 及 Hayward) 三類。
而常見之黃金奇異果 Hort16A 及較新之綠奇異果 Summer 3373 與綠奇異果 Hayward 相比較低之致敏性也由食物過敏原激發測試 (food challenge) 確認[90]。
奇異果過敏原種類
奇 異 果 過 敏 原 根 據 世 界 衛 生 組 織 過 敏 原 命 名 小 組 委 員 會 (WHO/IUIS allergen nomenclature sub-committee) 有 13 種,分別為半胱胺酸蛋白酶 (cysteine protease) Act d 1、索馬甜蛋白 (thaumatin-like protein) Act d 2、Act d 3、植物半胱 胺酸蛋白酶抑制劑 (phytocystatin) Act d 4、奇異果素 (kiwellin) Act d 5、果膠甲 酯 酶 抑 制 劑 (pectin methylesterase inhibitor) Act d 6 、 果 膠 甲 酯 酶 (pectin methylesterase) Act d 7、十號致病相關蛋白質 (pathogenesis-related protein, PR-10) Act d 8、肌動蛋白結合蛋白 (profilin) Act d 9、非特異性脂質傳送蛋白 (nsLTP) Act d 10、主要乳汁蛋白/成熟相關蛋白 (major latex protein/ripening-related protein, MLP/RRP) Act d 11、11S 球蛋白 (cupin, 11S globulin) Act d 12 及 2S 白蛋白 (2S albumin) Act d 13。
(1) Act d 1
Act d 1 為半胱胺酸蛋白酶,其 EC 編號為 EC 3.4.22.14,俗稱為奇異果酵素
或 actinidin,來自於番木瓜蛋白酶家族 (papain superfamily)。Actinidin 之成熟形 態包含單一多肽鏈組成之含有三個雙硫鍵的兩個結構域 (domain),其分子量為 23.8 kDa 而等電點為 3.5 [91-93]。Actinidin 有數個亞型蛋白,編碼於十個以上不 同之 mRNA 中,亞型蛋白之預測等電點範圍從酸性 (3.9) 至鹼性 (9.3) 都有[94]。
Actinidin 之活性 pH 範圍 (pH 4 - pH 10) 與基質專一性相當廣,其偏好水解離 胺酸 (Lys) 羧基端之醯胺鍵與酯鍵[77, 95, 96]。Actinidin 不會與樺樹花粉或草類 花 粉 產 生 交 叉 過 敏 反 應 , 故 被 認 為 是 造 成 奇 異 果 單 一 致 敏 反 應 (monosensitization) 的 其中一 個 過敏原 [77, 85, 92, 97, 98] 。 在多數 的 研 究 中 actinidin 被辨識為是奇異果過敏患者的主要過敏原[97-101]。
Actinidin 佔綠奇異果 (Actinidia deliciosa ’Hayward’) 總可溶性蛋白含量之 50%,而在黃金奇異果中 (Actinidia chinensis ‘Hort 16A’) 則相當低[94, 102]。免 疫組織定位研究發現 actinidin 分佈於綠奇異果外果皮小細胞之液胞中[94]。其他 來自番木瓜、鳳梨、無花果等,同為 papain 家族的同源性半胱胺酸蛋白酶,亦會 在特定組織或細胞中高含量的累積[77]。這些酵素的生理角色尚未知,但有人提 出其功能為儲藏性蛋白質或參與昆蟲、植物疾病或逆境的防禦機制[101, 103]。
Actinidin 在熱處理後仍保持其致敏性,此性質為造成攝食含有奇異果之食品 後產生過敏反應之重要特性[77]。儘管某些研究指出 actinidin 在經過腸胃道時容 易被分解[104],但其他研究顯示 actinidin 在經過腸與胃的消化環境後,仍保持其 酵素活性並能幫助消化食物中之蛋白質[105, 106]。
(2) Act d 5
Act d 5 又稱為 kiwellin 最先於綠奇異果中發現,為一種過敏原同時亦是奇 異果蛋白質成分之主要組成之一[107],於黃金奇異果中亦存在,且 kiwellin 是黃 金奇異果中最大量的蛋白質成分。Kiwellin 含有 189 個胺基酸,而於序列之 61 號 位置存在著序列異質性 (sequence heterogeneity),40%之蛋白質分子的 61 號位置 上之酪胺酸 (Tyr) 被組胺酸 (His) 所取代[77]。
Kiwellin 被分割成四個以下的胜肽:kissper、KiTH1、KiTH2、KiTH3[108]。
Kissper 為一段 39 個胺基酸之胜肽,可於奇異果可食部位中大量純化出來,其序 列與 kiwellin N 端前 39 個胺基酸序列完全相同。Kiwellin 為 kissper 之前驅物,
kiwellin 經過 Thr 39 與 Thr 40 間肽鍵斷開而產生 kissper。Kissper 因大量存在於 成熟的奇異果果實中,且同時具有高抗蛋白質水解性,導致其非常有可能影響腸 胃的生理[109]。KiTH 之一級結構與 kiwellin 序列 40 - 189 相同,從綠奇異果中 純化出之 KiTH 與 kissper 化學計量大約相同,故兩者產生之方式可能皆是從 kiwellin 之水解而來[108]。而 KiTH 與 kissper 於黃金奇異果中皆無法被偵測到。
研究觀察到兩者的存在與高含量之 actinidin 存在著關聯性,而後續研究更進一步 證明了 actinidin 造成 kiwellin 之水解而產生 KiTH 和 kissper[77, 108]。
Kiwellin 會被奇異果過敏患者之 IgE 所辨認,而與綠奇異果相似,黃金奇異 果之同源性蛋白質亦展現 IgE 結合能力。利用同一血清測試來自綠奇異果與黃金 奇異果之 kiwellin 蛋白,兩者皆能被血清所辨識,甚至西方墨點法之訊號亦能相 比較,可能意味著 IgE 結合抗原決定位的保守性。KiTH 亦被偵測為 IgE 結合分 子,結果表示 kiwellin 可能含有隱藏性 IgE 結合抗原決定位,在 kissper 被水解後 於 KiTH 上產生;kissper 本身可能即是一個 IgE 結合抗原決定位[77, 108]。而另 有研究發現,此過敏原在不同環境下 (pH、極性) 可能產生體內的結構變化而暴 露出不同的抗原決定位,包含特異性 IgE 的合成/相互作用。使用標準的皮下測 試 (skin prick test, SPT) 發現於 29 個奇異果過敏人口中,有 8 個 (28%) 會對 Kiwellin 產生陽性反應[110]。
(3) Act d 11
Act d 11 為一個 17 kDa 與成熟相關之蛋白質,其於綠奇異果萃取物中之含 量與正常成熟過程與採收後處理 (如乙烯處理) 相關[111]。與 Act d 11 序列與屬 於 Bet v 1 家族 (superfamily) 的主要乳汁 蛋白 / 成熟相關蛋白 (major latex protein/ripening-related protein, MLP/RRP) 家族有最高程度的相似性[112],而與
十號致病相關蛋白質 (pathogenesis-related protein, PR-10) 家族 (Bet v 1 為其中 一員) 有較低程度的序列相似性[111]。多數 Bet v 1 或其相關過敏原中屬於 B 細 胞或 T 細胞抗原決定位的胺基酸序列皆在 Act d 11 中出現,而主要聚集於 p-loop 模體 (motif) 或 C 端結構域 (domain) 上[111]。
在 Bet v 1 被 IgG、IgE 結合的過程中,Glu45扮演了非常重要的角色,而與 其他 MLP/PRPs 相似,Act d 11 與其他 Bet v 1 同源過敏原皆共有此特徵。由 Act d 11 抑制 IgE 結合 Bet v 1 或其同源物之能力,可推測其與 Bet v 1 類似物共抗原 決定位之區域比從序列相似度推測的還要大。Act d 11 在皮下測試 (SPT)、雙盲 合併安慰劑對照組的激發測試 (double blind placebo-controlled food challenge, DBPCFC)、免疫墨點法(immunoblot) 中皆產生 IgE 反應性。Act d 11 能部分抑制 Bet v 1 及其同源物 (Mal d 1、Cor a 1) 與 IgE 之結合。而 Bet v 1 可達到 100%之 對 Act d 11 正反應血清之 IgE 抑制[111]。
質譜技術
用於定量分析之質譜技術
(1) 提取離子色譜圖 (eXtracted Ion Chromatograms, XIC)
最 簡 單 用 於 LC-MS 是 之 定 量 方 法 為 提 取 離 子 色 譜 圖 (eXtracted Ion Chromatograms, XIC),藉由全域的 MS 掃描模式獲得數據,並於資料獲取後進行 數據處理建立目標離子的洗脫曲線。XIC 之波峰高度或面積能夠用於計算分析物 之含量。
(2) 選定離子監測 (selected ion monitoring, SIM)
SIM 為藉由掃描式質譜儀將資料獲取範圍限定於目標離子荷質比之周圍。範 圍越窄 SIM 方法越專一。SIM 比起全域掃描的 XIC 更加靈敏,因為 MS 能夠於 目標範圍內停留更長的時間。數個給定荷質比範圍的離子可以被觀察並累積量化,
定量方式仍然是透過離子之層析圖。
(3) 選擇反應監測 (selected reaction monitoring, SRM)
SRM 為非掃描式的技術,主要應用於三重四極柱(triple-quadrupole, QQQ/TQ) 設備上,藉由碎片化來提高選擇性。於 SRM 實驗中兩個質量分析器被用做靜態 的質量篩選器,用以監測特定前驅物離子之特定碎片離子。結合兩個質量篩選過 程之選擇性與高佔空比 (duty cycle)可得到無可媲美的高靈敏性定量分析方法。
所選擇之前驅離子與碎片離子荷質比所組成的特定一對數值被稱為「躍遷」
(transition). 舉例來說像是 673.5/534.3。
(4) 偽選擇反應監測 (pseudo SRM)
Pseudo SRM 被用來描述以線性離子阱(linear ion trap, LIT)或 QqTOF 來對特 定前驅離子的碎片離子進行一個小範圍的掃瞄。雖然此掃描模式與 SRM 相似,
但其目標碎片離子之訊號為藉由電子式的提取,故也可被視為碎片離子之 SIM。
(5) 多重反應監測(multiple reaction monitoring, MRM)
藉由於同一個實驗的層析時間間快速的循環於多個躍遷,並記錄各躍遷的訊 號來做為洗脫時間的函數,即能偵測複數的 SRM 躍遷。此方法於偵測層析時之 共洗脫物的多個躍遷具有更高的選擇性。這樣的並行式 SRM 被廣泛稱為多重反 應監測 MRM,然而此名稱遭到國際純化學暨應用化學聯合會命名法 (IUPAC nomenclature)所棄用(Provisional Recommendations, August 2006)[113]。
SRM/ MRM 技術之優勢
選擇反應監測 (selected reaction monitoring, SRM)利用三重四極柱(triple- quadrupole, QQQ/TQ) 之獨特性質來進行定量分析。於 SRM 中第一個與第三個 四極柱被用作為篩選器來侷限僅特定目標胜肽離子或胜肽碎片離子之荷質比能 通過,第二個四極柱則被用作碰撞室使用,如圖 a。多個躍遷能夠於分析時被偵 測,並產生出多張以層析時間與訊號強度為座標軸之特定躍遷的層析圖譜。此兩 階段的小窗口質量篩選使得方法具高選擇性能夠將共洗脫的背景有效率的篩選 掉。與其他質譜為基礎的方法不同,QQQ 為主之 SRM 分析並不會記錄全域的質
量質譜圖。SRM 這非掃描式的特性使其靈敏性高於傳統式全域掃描方法的一至 二倍。此外也同時使得其線性反應的動態範圍 (dynamic range)擴大至五倍。使得 SRM 技術能夠偵測於複雜樣品中的低含量蛋白質,此特性於系統性定量研究上 十分重要[113]。
文獻[114]比較同一 LC 系統之 UV 偵測器及質譜儀,在偵測牛乳過敏原上後 之靈敏度為前者之 10 倍以上。且 MRM 可偵測到含量為 femtomole 至 attomole 範圍含量之胜肽,等同於由 ELISA 計算之 ppm 到 ppb 範圍[57]。另一文獻[115]
比較 LC/qTOF 與 LC/MRM,後者之靈敏度高於前者 10 倍。文獻[114]分析牛乳 過敏原添加樣品得到之偵測極限及定量極限分別為 1 與 4 ppm。文獻[116]分析花 生過敏原 Ara h 2 之偵測極限及定量極限分別為 5 及 13 ppm,Ara h 3/4 之偵測極 限及定量極限分別為 1 及 3.7 ppm。文獻[117, 118]分析甲殼類原肌凝蛋白及精胺 酸激酶得到之偵測極限及定量極限分別為 3 及 0.25 nM。文獻[119]利用 LC/MRM 系統偵測到微量之穀物過敏原,偵測極限及定量極限分別介於 0.01 到 0.03 ppm 及 0.01 到 0.1ppm 之間。以上研究證明 MRM 之靈敏度與 ELISA 法相當甚至更 高[57]。具高選擇性、靈敏性、廣的動態範圍及能夠同時分析多樣品使得 QQQ MS 系統搭配 SRM/MRM 技術為解決食品過敏原定量分析之樣品基質複雜、過敏原 含量變化大、過敏原目標多等問題之方法。
圖 a、SRM/MRM 於 QQQ MS 之分析。數個分析物於層析系統中共洗脫出。第一個四極柱以特定之荷質比 篩選掉多數的共洗脫離子,然而因為與目標物有相同的質量藍色的干擾物仍存在。於第二個四極柱分析物 被碰撞為碎片。於第三個四極柱又再過濾掉多數的碎片離子,包含全部的藍色碎片離子,僅留下特定目標