國立臺灣大學生物資源暨農學院園藝暨景觀學系 碩士論文
Department of Horticulture and Landscape Architecture College of Bioresources and Agriculture
National Taiwan University Master Thesis
褐化抑制物對結球萵苣主莖創傷後酚類含量變化 之影響
Effect of Browning Inhibitors on Phenolic Contents in Wound Stem of Iceberg Lettuce
張乃文 Nai-Wen Chang
指導教授:王自存 博士 Advisor:Tsu-Tsuen Wang, Ph.D.
中華民國 105 年 2 月
February, 2016
誌謝
能夠完成碩班的種種挑戰首先最感謝的是我的指導教授 王自存老師於這段 日子的教導與分享,從研究方向的討論到研究架構、實驗方法、實驗材料上的協助、
論文撰寫與修改都耐心地給予指導和啟發,在生活上也常常與我們分享,不論是去 果菜拍賣市場或是內湖花市或是聚餐,於此獻上最深的感謝與敬意。論文口試期間,
特別感謝口試委員 楊雯如老師 與 林淑怡老師的鼓勵、建議和指正,使得本論文 能更加完備,在此獻上感謝。
在實驗上,感謝穎華學姊的各種建議,以及無數次的電話救援;感謝立行彥達 在 HPLC 儀器上所有幫助;謝謝榆蒨、偉晴、偉齊、璧甄學姊在實驗上的討論和建 議,也謝謝嘉祐學長幫忙尋找藥品。在研究期間,感謝處理室所有人的幫助:超級 翻譯機陳昱琬,在這段日子裏的各種當頭棒喝和陪伴;還有萵苣好 partner 楊婕,
在切萵苣和磨萵苣時互相扶持,謝謝你們陪我一起走過做實驗和寫論文的日子,另 外也感謝安迪郭無與倫比的娛樂效果;謝謝學弟瑪莉擔任切菜小助手和口試記錄 員;謝謝歐姊、韋汶、穎華、阿汪學姊,感謝你們的幫忙和在樂華夜市吃炸魷魚的 日子;謝謝伯偉的龍門客棧水餃和歌單,陪伴了不知名的夜晚,也感謝你的各種分 析;謝謝瑄斐和我一起看冷門電影;謝謝 Leo ho、晨崴、國均、藝綸學長,謝謝你 們為我的研究生活增添了色彩。謝謝台大園藝所所有的同學或是在這段日子裏出 現的朋友,和我一起跑步、上課、聊天。
最後,感謝我的家人,爸媽和姊姊,還有臭麻吉,感謝你們經濟上與精神上的 支持,讓我能專注於課業與研究,並且在回家時可以重新歸零,謝謝你們默默地支 持著我。謝謝碩班期間所有的你們,謝謝你們的陪伴和支持,我愛你們。
摘要
結球萵苣 (Lactuca sativa L. var. capitata) 為菊科萵苣屬作物,為國內冬季重要 的外銷蔬菜。結球萵苣採收時由主莖基部切下,採收切面會於創傷後轉為紅褐色,
當結球萵苣以完整葉球販售時,此切面褐化為影響外觀品質與商品價值的重要因 素之一。已知創傷萵苣主莖內,受創傷誘導生成之酚類經酵素氧化後會聚合產生褐 色物質,因而造成主莖切面褐化;以醋酸或含硫氫基化合物處理後,具有延緩主莖 切面褐化之效果,而以含硫氫基化合物處理後會使外觀呈現偏黃色色澤。本研究探 討創傷後主要造成結球萵苣主莖切片褐化的酚類物質種類與其含量變化,並進一 步比較以不同延緩褐化藥劑處理後,其延緩外觀褐化之效果,以及褐化與酚類物質 含量變化之關係。貯藏於 5°C下之結球萵苣主莖切片,於創傷後第二天維管束即出 現褐化徵狀,持續褐化過程中切面逐漸轉為紅褐色,其外觀 a*值變化趨勢與切片 之總酚類含量、總咖啡酸含量變化趨勢相同,主要於創傷第四天以內增加。創傷後 主 要 生 成 酚 類 為 綠 原 酸 (chlorogenic acid) 、 菊 苣 酸 (chicoric acid) 與 異 綠 原 酸 (isochlorogenic acid),為易被萵苣多酚氧化酶氧化的基質,並且造成褐化,同濃度 下的菊苣酸經氧化後造成褐化的程度大於綠原酸。當綠原酸與菊苣酸同時存在時 造成的褐化程度與個別濃度相關,彼此間並無交互關係;類黃酮並非萵苣多酚氧化 酶易催化的基質,但與咖啡酸衍生物同時存在時,類黃酮可被醌類氧化並造成褐 化。主莖切片創傷後首先生成綠原酸,菊苣酸與異綠原酸則是於創傷第二天後開始 生成,三者含量皆於第四天以後達到最高,第四天後綠原酸含量開始下降,其於兩 者含量則維持一定。萵苣主莖切片經 3%醋酸處理後,其外觀可維持白色色澤至貯 藏第八天,貯藏期間總酚類、總咖啡酸含量皆不因創傷而上升,顯示 3%醋酸可抑 制創傷後酚類物質生成,因而具有延緩切面褐化的效果。萵苣主莖切片經含硫氫基 化合物 N-acetyl-L-cysteine (NAC)與 L-cysteine hydrochloride (CySH)以不同濃度處 理後,經 2% NAC 處理之切片於創傷後第四天開始褐化且其外觀 a*值於第四天後 開始上升,第八天切面為橘色;延緩褐化效果較佳的 3% NAC、2% CysH 與 3%
3
CySH 處理,皆且可延緩切片外觀 a*值不上升至第八天,第八天外觀具黃色色澤。
主莖切片經 NAC 與 CySH 處理後,其延緩創傷後綠原酸、菊苣酸、異綠原酸含量 開始上升時間的效果不同,兩種濃度下之 CySH 處理延緩菊苣酸含量上升的效果 皆優於 NAC 處理,且 2% CySH 處理延緩綠原酸含量上升的效果也優於同濃度下 的 NAC 處理,而 CySH 與 NAC 處理對延緩異綠原酸含量上升之效果相同。CySH 處理延緩萵苣主莖切片褐化的效果優於 NAC 處理的原因為其延緩菊苣酸與綠原酸 含量上升的效果較佳。主莖切片經 NAC 與 CySH 處理後,其總類黃酮含量,除了 3% CySH 處理可延緩至第六天後開始上升,其餘三組皆於創傷第四天後開始上升,
與對照組同時。延緩褐化效果較佳的 2% CySH 與 3% NAC 處理組的第八天含量約 為對照組的 5 倍,3% CySH 處理組的含量約為對照組的 9.5 倍,而延緩褐化效果 較差的 2% NAC 處理組則為對照組的 3 倍。由含硫氫基化合物處理結果顯示,此 類化合物處理可藉由延緩咖啡酸衍生物含量上升而延緩外觀褐化,但處理後切片 之類黃酮含量高於對照組,並且貯藏後外觀呈現黃色色澤。
Abstract
Iceberge lettuce (Lactuca sativa L. var. capitata), belonging to the Asteraceae family, is an important cool-season crop for export in Taiwan. Iceberg lettuce is harvested at the bottom of stem and the cut surface turns from white to reddish-brown after storage.
Browning of cut surface causes the quality loss and market value loss of head lettuce during postharvest. Wound-induced phenolic compounds in wound lettuce stem tissue are oxidized to O-quinones by polyphenol oxidase (PPO) and lead to the formation of browning pigment at the cut surface. Researches indicated that acetic acid or sulfhydryl compounds treatment can prevent the browning of cut surface, but sulfhydryl compounds treatment could cause the yellowish appearance of cut surface. In this study, the changes in phenolic compounds content that most contributed to the browning of cut surface after wounding were investigated. Furthermore, the effect of different chemical treatment on alleviating cut surface browning, and the changes in phenolic content of stem disk after wounding were studied in relation to browning. When stem disk of iceberg lettuce were stored at 5°C after wounding, browning appeared first in vascular bundle of stem disk, and then the cut surface turned red-brown. At the same time, the change in a* value of stem disk was related to the total phenolic content and the caffeic acid derivatives content.
Chlorogenic acid, chicoric acid and isochlorogenic acid were the main phenolic compounds that accumulated in the control stem disk after wounding, and were good substrate for iceberg lettuce polyphenoloxidase (PPO) in vitro. After oxidized by PPO, chicoric acid quinone could cause the deeper browning color than chlorogenic acid quinone. During in vitro oxidizing process, chlorogenic acid and chicoric acid were oxidized independently by PPO. However, flavonoids which are not good substrate for iceberg lettuce PPO could enhance the browning because flavonoids could be non- enzymatically oxidized by caffeic acid derivatives quinones when caffeic acid derivatives
5
exist. In the stem disk, chlorogenic acid were synthesized within 2 days after wounding, and then chicoric acid and isochlorogenic were synthesized 2 days after wounding. The content of the three caffeic acid derivatives were peak at 4 days after wounding. Four days after wounding, the content of chlorogenic acid declined, whereas the content of chicoric acid and isochlorgenic acid remained. The appearance of 3% acetic acid treated-stem disk maintained white after 8 days of storage, and the total phenolic content and the caffeic acid derivatives content remain low content after wounding. 3% acetic acid treatment could inhibit the phenolic synthesis which was induced by wounding;
therefore, it was effective on inhibiting browning. After the stem disks were treated with sulfhydryl compounds, including N-acetyl-L-cysteine (NAC) and L-cysteine hydrochloride (CySH) at concentration of 2% and 3%, the color of wound stem disk turned orange or yellow during 8 days of storage. In 2% NAC treatment, when a* value of the stem disk increased at 4 days after wounding, the stem disk appeared browning on cut surface, and the stem disk turned orange at 8 days after wounding. In 3% NAC, 2%
CySH and 3% CySH treatment, which showed better inhibition of the browning, a* value of the stem disk remained low during 8 days of storage, and finally the color of the stem disk turned yellow. The NAC and CySH treatments had different effect on delaying the increase in chlorogenic acid, chicoric acid and isochlorogenic acid content. The CySH treatment could delay the increase in chicoric acid content till 8 days after wounding, while the NAC treatment was less effective. 2% CySH treatment was more effective on delaying the increase in chlorogenic acid than 2% NAC treatment. On the delaying the increase in isochlorogenic acid content, the CySH treatments and the NAC treatments had the same effect. Therefore, the main difference of the effect on delaying browning between CySH and NAC treatment was the ability of delaying the increase in chlorogenic acid and chicoric acid content. 3% CySH treatment delayed the increase in flavonoid
content to 6 days after wounding; In other sulfhydryl compounds treatment and control, the flavonoid content increased at 4 days after wounding. At 8 days after wounding, the flavonoids content in the disk treated with 3% CySH and 3% NAC was 5 times more than that in control; and the flavonoids content in the disk treated with 2% CySH was 9.5 times more than that in control, while the disk treated with 2% NAC treatment was 3 times more than in control. The result of sulfhydryl compounds treatments suggest that it delayed the browning of stem disk by delaying the increase in caffeic acid derivatives content ; however, the treated-disk had higher flavonoid content which affected the browning of stem disk, and finally had yellowish appearance.
7 目錄
誌謝 ... 1
摘要 ... 2
Abstract ... 4
第一章 前言 ... 13
第二章 前人研究 ... 15
一、 萵苣類蔬菜簡介 ... 15
二、 萵苣主莖切面褐化現象 ... 16
三、 創傷與酚類生成 ... 19
四、 影響萵苣主莖褐化程度的因子 ... 23
五、 延緩褐化反應之處理技術 ... 29
六、 結球萵苣內咖啡酸衍生物質測定 ... 31
第三章 材料與方法 ... 33
二、結球萵苣主莖切片與創傷後藥劑處理 ... 34
三、主莖切片外觀褐化變化觀察 ... 35
四、主莖切片酚類組成與含量分析 ... 35
五、萵苣多酚氧化酶於植物體外氧化酚類標準品 ... 36
第四章 結球萵苣主莖切片創傷後酚類之生成與褐化間之關係 ... 38
一、以 HPLC 層析方法分離主莖切片內酚類物質 ... 38
二、確認創傷後萵苣主莖萃取液內主要酚類物質 ... 39
三、萵苣內多酚氧化酶於植物體外氧化酚類物質與褐化之關係... 40
四、創傷後萵苣主莖切片褐化過程中酚類組成變化 ... 44
五、結論 ... 46
第五章 藥劑處理延緩褐化與酚類物質變化之關係 ... 48
一、醋酸處理對創傷萵苣主莖切片中酚類代謝之影響 ... 48
二、NAC 處理對創傷萵苣主莖切片中酚類代謝之影響 ... 49
三、CySH 處理對創傷萵苣主莖切片中酚類代謝之影響 ... 51
四、硫氫基化合物處理延緩褐化效果討論 ... 52
五、結論 ... 55 第六章 總結 ... 90 參考文獻 ... 92
9
圖目錄
圖 1. 咖啡酸衍生物與類黃酮物質生合成途徑 ... 27 圖 2. 不同褐化抑制溶液處理之主莖圓片在 5°C 下貯藏 8 天期間外觀之變化。 .. 57 圖 3. 創傷後於 5°C 下貯藏第四天之萵苣主莖切片萃取液經 HPLC 分離後偵測波長
320 nm 下所得層析圖。(A.為主莖萃取液;B.為標準品綠原酸與菊苣酸) ... 58 圖 4. 創傷後於 5°C 下貯藏第八天之萵苣主莖切片萃取液經 HPLC 分離後偵測波長
320 nm 下所得之層析圖與文獻層析圖比較。(A.為創傷後主莖切片萃取液;B.為 文獻中創傷後萵苣中肋萃取液;C.為文獻中創傷後萵苣中肋萃取液內相對應之 酚類物質(modified from Tomás-Barberán et al., 1997) ... 59 圖 5. 創傷後第八天萵苣主莖切片萃取液經不同 HPLC 梯度分離後之層析圖。(A.
為以 1.2% min-1等梯度進行分離;B.為以修正後梯度進行分離) ... 61 圖 6. 添加綠原酸與菊苣酸標準品於創傷後第四天之對照組主莖切片萃取液,經
HPLC 分離後所得之層析圖。(A.為 40 μg/ml 綠原酸標準品與 20 μg/ml 菊苣酸標 準品;B.為創傷後第四天之對照組主莖切片萃取液;C.為以標準品體積:樣品體 積 (1:1) 添加後之混合溶液)... 62 圖 7. 添加綠原酸與菊苣酸標準品於創傷後第八天 3%CySH 處理組主莖切片之萃
取液中,經 HPLC 分離後所得之層析圖。 ... 63 圖 8. 添加綠原酸與菊苣酸標準品於創傷後第八天 3%NAC 處理組主莖切片之萃取
液中,經 HPLC 分離後所得之層析圖。 ... 64 圖 9. 綠原酸、菊苣酸與類黃酮標準品經結球萵苣葉片內多酚氧化酶氧化後褐化過
程外觀之變化,由上而下分別為經過 0、3、10、25、55 分鐘。 ... 66 圖 10. 不同濃度之綠原酸、菊苣酸與芸香苷標準品經結球萵苣葉片多酚氧化酶於
植物體外氧化後 40 分鐘之 (A) 溶液外觀 (B) 標準品溶液氧化後顏色值。 ... 67 圖 11. 標準品於波長 200-600 nm 間連續吸收光譜。 ... 68 圖 12. 綠原酸標準品 (20 μg/ml) 經萵苣主莖 PPO (30 U/ml) 氧化過程每隔五分鐘
紀錄之吸收光譜變化,(右上)為波長 370-450 nm 間放大。... 69 圖 13. 菊苣酸標準品 (20 μg/ml) 經萵苣主莖 PPO (30 U/ml) 氧化過程每隔五分鐘
紀錄之吸收光譜變化,(右上)為波長 370-450 nm 間放大。... 70 圖 14. 標準品溶液經萵苣葉片 PPO 氧化過程,於反應後 0、15、90、180、300、
... 71
600、900、1200、1800 秒紀錄波長 270-450 nm 間之吸收光譜。(A.為 20 μg/ml 綠 原酸;B.為 20 μg/ml 菊苣酸;C.為 30 μg/ml 綠原酸與 30 μg/ml 菊苣酸之混合溶 液)。 ... 71 圖 15. 標準品溶液經萵苣葉片 PPO 氧化過程,於反應後 0、15、90、180、300、
秒於 (A)波長 320 nm 與 (B) 410 nm 下吸光值變化。 ... 72 圖 16. 標準品溶液經萵苣葉片 PPO 氧化過程,於反應後 0、15、90、180、300、
600、900、1200、1800 秒紀錄波長 270-450 nm 間之吸收光譜。(A.為 20 μg/ml 芸 香苷;B.為 20 μg/ml 綠原酸與 20 μg/ml 芸香苷之混合溶液;C. 為 20 μg/ml 綠原 酸與40 μg/ml 芸香苷之混合溶液) ... 73 圖 17. 標準品溶液經萵苣葉片 PPO 氧化過程,於反應後 0、15、90、180、300、
秒於波長(A) 320 nm、(B) 410 nm、(C) 357 nm 下吸光值變化。 ... 74 圖 18. 創傷後萵苣主莖圓片於 5°C 下貯藏 8 天期間 a*值與 b*值變化。 ... 75 圖 19. 創傷後萵苣主莖圓片於 5°C 下貯藏 8 天期間總酚類含量變化。 ... 76 圖 20. 創傷後萵苣主莖圓片於 5°C 下貯藏 8 天期間綠原酸、菊苣酸、異綠原酸z與
主要咖啡酸衍生物總含量變化。(異綠原酸 z由文獻推測定性,並以綠原酸標準 曲線換算其濃度)... 77 圖 21. 創傷後經 3%醋酸處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間 a*值與 b*值
變化。 ... 78 圖 22. 創傷後經 3%醋酸處理萵苣主莖圓片於 5°C 下貯藏 8 天期間總酚類含量變
化。 ... 79 圖 23. 創傷後經 3%醋酸處理萵苣主莖圓片於 5°C 下貯藏 8 天期間綠原酸、菊苣
酸、異綠原酸z與主要咖啡酸衍生物總含量變化。(異綠原酸z由文獻推測定性,
並以綠原酸標準曲線換算其濃度) ... 80 圖 24. 創傷後經 2%及 3% NAC 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間 a*值
與 b*值變化。 ... 81 圖 25. 經 2%、3% NAC 處理之創傷萵苣主莖圓片於 5°C 下貯藏 8 天期間總酚類含
量變化。 ... 82 圖 26. 創傷後經 2%、3% NAC 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間綠原
酸、菊苣酸、異綠原酸z與主要咖啡酸衍生物總含量變化。(異綠原酸z由文獻推 測定性,並以綠原酸標準曲線換算其濃度) ... 83
11
圖 27. 創傷後經 2%及 3% CySH 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間 a*值 與 b*值變化。 ... 84 圖 28. 經 2%、3% CySH 處理之創傷萵苣主莖圓片於 5°C 下貯藏 8 天期間總酚類
含量變化。... 85 圖 29. 創傷後經 2%、3% CySH 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間綠原
酸、菊苣酸、異綠原酸z與主要咖啡酸衍生物總含量變化。(異綠原酸z由文獻推 測定性,並以綠原酸標準曲線換算其濃度) ... 86 圖 30. 創傷後經 2%、3% CySH 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間主要
總類黃酮含量變化。 ... 87 圖 31. 創傷後經 2%、3% NAC 處理之萵苣主莖圓片於 5°C 下貯藏 8 天期間主要總
類黃酮含量變化。 ... 88
表目錄
表 1. 高效能液相層析*內綠原酸與菊苣酸標準品之標準曲線的迴歸方程式 ... 65 表 2. 標準添加所得之定量誤差 ... 65 表 3. 不同濃度藥劑處理延緩各酚類含量開始上升時間之比較 ... 89
13
第一章 前言
結球萵苣 (Lactuca sativa L. var. capitata, iceberg lettuce ) 俗稱西生菜、美生菜,
為菊科萵苣屬一年生作物。原產於地中海地區,喜冷涼氣候生長,富含多種維生素 及礦物元素,具有適合生食及耐貯運的優點,常為沙拉食材之一。
2013 年全世界萵苣及苦苣之總產量為約 2,489 萬公噸,主要生產國家依次為中國、
美國、印度、西班牙及義大利等國 (FAO, 2015) 。根據聯合國糧食及農業組織 (Food and Agriculture Organization of the United Nations, FAO) 統計,近幾年世界蔬菜生產 量中,萵苣排名約第 11 位,是全球生產量、食用量最高的蔬菜之一。
近年台灣飲食習慣改變,因此對作為沙拉食用的結球萵苣需求量攀升;台灣冬 季適合種植結球萵苣,除可取代十字花科蔬菜生產面積、解決國內冬季蔬菜產量過 剩、價格暴跌的問題,此時期適逢日本國內休耕期,因此具有外銷日本的潛力。2002 年由行政院農業委員會成立計畫,輔導農民成立產銷班,並以策略聯盟方式共同生 產與運銷,供應國內需求以及外銷。2014 年台灣結球萵苣出口達 11,296 公噸,產 值約 28 億新台幣,出口至日本占八成,一成至新加坡,其餘則是出口至韓國、中 國大陸、香港、馬來西亞、阿拉伯聯合大公國 (行政院業委員會農業統計資料查詢)。
目前台灣萵苣外銷至日本的主要對象為大型速食業者,經截切後使用,近年來 期望將完整葉球的萵苣販售至日本超級市場零售,以擴大日本外銷市場,因此貯藏 期間外觀品質的要求提高,其中又以萵苣主莖切面褐化問題最受重視。萵苣採收時 於主莖處切下,因採收造成的機械傷害會誘發萵苣重新合成酚類物質,並於切口附 近被氧化形成褐色物質,使得萵苣於貯藏期間出現切面褐化 (butt discoloration) 的 情形,因而降低商品品質與貯架壽命。
目前減輕萵苣切面褐化的方式可大略分為物理及化學性方法,物理方法可利 用改變貯藏環境的大氣組成或熱處理;化學方法則以藥劑處理為主,常用藥劑為有 機酸、抗氧化劑、半胱胺酸或其衍生物、含鈣化合物等等,許多研究也深入探討減 輕萵苣切面褐化的機制。2000 年 Salveit 提出創傷誘導萵苣組織中酚類代謝改變,
因而造成組織褐化的可能途徑,顯示各種減輕褐化方法可能改變途徑中的某些步 驟。洪 (2014) 以台灣生產之萵苣為材料,藉由物理或化學方法減輕主莖褐化程度,
主莖圓片經藥劑處理後可以減輕貯藏期間的褐化程度,而使用不同藥劑及濃度則 延緩褐化效果不同,其中以半胱胺酸衍生物處理後會改變切面外觀的顏色。本研究 將探討創傷後萵苣主莖切片外觀褐化與其酚類組成及含量之關係,並進一步探討 萵苣主莖切片,經不同減輕褐化藥劑處理後,其總酚類含量變化、萵苣內主要酚類 組成與含量變化,藉此比較不同褐化抑制劑對創傷後萵苣酚類物質與褐化的影響。
15
第二章 前人研究
一、 萵苣類蔬菜簡介
萵苣 (Lactuca sativa L.) 為菊科 (Asteraceae) 萵苣屬一年生草本植物,萵 苣屬 (Lactuca) 其字根 lac 意即如牛奶般的多汁,而 sativa 則是被栽種之意 (Stearn, 1966)。萵苣最早栽種記錄是在地中海區域,其親本刺萵苣 (L. serriola) 原 生地則位於地中海東部地區,喜於冷涼乾燥氣候生長 (Decoteau, 2007)。
(一) 萵苣種類
Nuez 與 Prohens (2008) 綜合萵苣之葉片型態、大小、質地、結球特性以 及莖部形態,將其分為六類:
1. 包被型結球萵苣 (crisphead):學名為 Lactuca sativa var. capitata,又稱 iceberg 或 head lettuce。發育初期葉寬漸大於葉長,葉片形態於 10~12 片本葉期轉為杯狀 (cup-shaped) 並相互重疊,外葉包裹新葉後形成緊實葉球,新葉於葉球內部形 成後使葉球更加緊實。其生長適溫為 10~20℃,適合於台灣 10 月至翌年 4 月栽 培,冬春季日夜溫差有利於結球,遇高溫則不易結球。
2. 抱合型萵苣 (butterhead):又稱 cabbage lettuce。與包被型結球萵苣相似,但其 結球特性會形成較小且較鬆散之葉球,外葉未完全包覆葉球。於歐洲具有日長 敏感度、生長季節不同的栽培種。
3. 羅蔓萵苣 (romaine):學名為 Lactuca sativa var. longifolia,又稱 cos lettuce。
其葉子形態較長且外表粗糙,具有明顯較寬的葉肋,因此有質地相對較脆的特 性。
4. 葉用萵苣 (leaf or leaf bunching lettuce):學名為 Lactuca sativa var. crispa。其栽 培種種類繁多,葉面大小、顏色、葉緣形態多樣;因其開放式生長 (open-growth) 特性,深綠色葉片比例較高,風味較強烈。
5. 嫩莖萵苣 (stem lettuce):學名為 Lactuca sativa var. angustana Hort.或 var.
asparagina Bailey,又稱 celtuce 或 asparagus lettuce。在花芽分化後,莖部抽 長,於開花前採收,其長莖部粗厚且直立,經去皮後可食用。
6. 拉丁萵苣 (latin lettuce):又名 grassé lettuce。與羅曼萵苣相似,皆具有直立向 上生長的特性,但葉長較短且口感較不脆。葉球頂部有時呈現閉合,質地與 bibb- type butterhead 相似。
(二) 台灣結球萵苣生產與出口概況
台灣於 2001 年前種植的萵苣以葉用萵苣為主,近年來因飲食習慣改變,作為 沙拉食用的結球萵苣需求量攀升,主要仰賴美國與澳洲進口;鄰近台灣的國家,如 日本、新加坡、韓國亦為結球萵苣之進口國。台灣冬季適合結球萵苣生長,選擇其 作為冬季裏作蔬菜,除了可以供應國內需求外,因適逢日本休耕期因此具有外銷日 本的潛力 (何,2006)。另外,以結球萵苣取代十字花科蔬菜種植面積,亦可以解決 國內冬季蔬菜產量過剩、價格暴跌之問題。綜合以上因素,2002 年開始由行政院 農業委員會擬定計畫,輔導農民成立產銷班,並以策略聯盟方式共同生產與運銷,
供應國內需求以及外銷 (何,2006)。
2002 年台灣結球萵苣總生產量合計約有一萬五千公噸,雲林縣為主要生產地,
約占總產量之七成。2014 年結球萵苣出口量達 11,296 公噸,產值約為 28 億新台 幣,出口至日本占八成,一成至新加坡,其餘則是出口至韓國、中國大陸、香港、
馬來西亞、阿拉伯聯合大公國 (行政院業委員會農業統計資料查詢)。
二、 萵苣主莖切面褐化現象
褐化為造成完整葉球或輕度加工萵苣於採後貯藏期間品質劣變的主要因素之 一 (Ilker et al., 1977;Mateos et al., 1993)。萵苣在田間採收時會由主莖部分切下並 保留完整葉球,此一採收切口經過一段貯藏期間後會產生褐化的現象。完整葉球的
17
品質判斷指標之一即為觀察主莖切面是否產生褐化徵狀,因此延緩主莖切面褐化 為目前完整萵苣葉球商品販售時之重要目標。
(一) 褐化現象
褐化 (browning) 為園產品產生深色或褐色物質使其外觀褐化的現象,依反應 過程中是否有酵素參與可分為非酵素型褐化 (non-enzymatic browning) 以及酵素 型 褐 化 (enzymatic browning) 兩 類 。 非 酵 素 型 褐 化 包 含 梅 納 反 應 (maillard reaction) 、 抗 壞 血 酸 氧 化 反 應 (oxidation of ascorbic acid) 、 焦 糖 化 反 應 (caramelization) 及單寧氧化等 (Mayer and Harel, 1979),為加工食品中重要之褐化 反應;而酵素型褐化則是造成園產品品質劣變與損耗的重要因素之一 (Vámos‐
Vigyázó, 1981) 。
(二) 酵素型褐化反應
酵素型褐化反應係由於細胞的隔室 (compartmentation) 受到破壞所導致,因 胞器完整性遭受破壞後,在有氧的環境下,存在於植物細胞液胞 (vacuole) 內的酚 類物質與酵素接觸,受到氧化後形成醌類 (quinone) 物質,醌類物質再以非酵素作 用方式與其餘物質共同聚合後,形成黑色素 (melanins) 累積,因而造成園產品褐 化 (Toivonen and Brummell, 2008;Yoruk and Marshall, 2003)。
園產品細胞隔室失去完整性的可能原因為老化、失水、生理障礙或碰傷、創傷 等機械傷害所導致。老化、失水或生理障礙伴隨產生之褐化現象,主要為貯藏過程 中細胞完整性逐漸崩解所導致。園產品在貯藏過程中因失水導致褐化之現象常出 現於果實表皮,如:荔枝、紅毛丹 (rambutan) (梁和柯,2012;Landrigan et al., 1996)。
低溫導致的寒害也會伴隨褐化現象產生,貯藏過程中細胞完整性因受到生理代謝 失調產生的物質影響,而逐漸崩解,因此間接導致酵素型褐化發生,如:發生寒害 的香蕉果皮褐化、發生寒害的鳳梨內部果肉褐化 (Pantastico et al., 1976;薛等,
2009;)。
結球萵苣採收後之生理障礙主要有褐斑病 (brown stain)、粉紅中肋 (pink rib)、
中肋變色 (rib discoloration)與鏽斑病 (russet spotting),分別由不同環境與內在因子 導致。鏽斑病主要是由低濃度乙烯誘導產生 (Morris et al., 1974),褐斑病則為高濃 度二氧化碳誘導產生 (Stewart, 1978),粉紅中肋則是因採收成熟度過高、貯運時間 過久或高溫貯藏導致。
除了因老化、失水、生理障礙導致細胞完整性逐漸崩解,進而發生褐化外,園 產品經碰撞、採收創傷或截切後,傷口附近細胞結構因立即遭受破壞,亦會發生酵 素型褐化。常見的截切水果,如:蘋果、西洋梨,因其組織內具有一定量的酚類物 質,於截切後數個小時內,其截切表面即會產生明顯的褐化現象;而相對酚類含量 較低之萵苣,截切後必須貯藏一段時間後才會出現截切邊緣褐化的現象,主要原因 為萵苣受創傷後會重新合成酚類物質,作為酵素型褐化之基質 (Ke and Saltveit, 1989)。
(三) 多酚氧化酶
Burton (2003) 將具有氧化能力的酵素分為三大種類:和氧酶 (oxygenases) 、 氧化酶 (oxidases) 、過氧化酶 (peroxidases) 。Oxidase以氧分子作為電子接收者,
並產生含氧高活性中間物 (reactive oxygen intermediate),因此具有氧化能力;
Peroxidase以過氧化氫作為電子接收者,於酵素內產生帶有自由基的原血紅素基 (heme-group) ,再利用這些含氧高活性中間物氧化具有還原力的基質。氧化酶 (oxidases) 與過氧化酶 (peroxidases) ,為藉由含氧高活性中間物氧化基質,因此相 對oxygenase的氧化反應較不專一,氧化後的產物也較複雜。
Oxygenase則是利用氧分子,將一個氧原子或氧分子接入反應物的結構中,並 且對反應物較專一。植物內可氧化酚類物質的酵素包含了屬於oxygenase的多酚氧 化 酶 (polyphenol oxidase, PPO, EC. 1.14.18.1) 、 漆 氧 化 酶 (laccase) 、 脂 氧 酶 (lipoxygenase, EC. 1.13.11.12) 以及過氧化酶 (peroxidases, EC. 1.11.1.7),而與園產 品 褐 化 相 關 的 酵 素 主 要 為 多 酚 氧 化 酶 (polyphenol oxidase) 與 過 氧 化 酶
19
(peroxidases) (Toma´s-Barbera´n and Espı´n, 2001)。
過氧化酶 (peroxidases, POD, EC. 1.11.1.7) 參與褐化的程度仍不明瞭。在園產 品內,過氧化酶將酚類物質氧化之能力受到過氧化氫濃度的限制,組織內PPO相對 活性較高,故在一般情形下,PPO氧化酚類物質能力優於POD。但PPO與POD同時 存在時具有協同效果 (synergistic effect),當PPO氧化酚類過程中會產生過氧化氫,
而於此種狀況下,POD即可利用過氧化氫以氧化酚類物質;另外POD也可利用PPO 作用所產生之半醌中間體 (semiquinone intermediate) 代替過氧化氫作為電子接收 者(Murata et al., 2002;Richard-Forget and Gauillard ,1997) 以進行氧化作用。因此 在園產品內主要將酚類氧化之酵素為多酚氧化酶,但仍不能排除過氧化酶可能參 與褐化之影響。
多酚氧化酶 (EC. 1.14.18.1) 為一群可將多酚類氧化成醌類 (o-quinone) 之酵 素,主要存在於類囊體 (thylakoid) 膜上,其三級結構中含有銅離子作為輔基 (cofactor) 構成酵素的活性部位 (active site),催化最佳的pH值範圍為5-7。多酚氧 化酶主要作用的基質為酚類,在有氧環境下將酚類氧化為醌類,可依序分為兩步驟 反應: (1)將單酚羥化為雙酚 (hydroxylation of monophenols to O-diphenols) (2) 將雙 酚氧化為醌類 (oxidation of monophenols to O-quinone)。羥化反應的催化速率相較 氧化反應較慢,產生無色雙酚類;氧化反應則產生有顏色的醌類物質 (Toivonen and Brummell, 2008)。
醌類為具有高活性的親電子分子 (electrophilic molecules),易與自身或其他物 質結合,如氨基酸、胜肽、蛋白質或醣類分子,聚合成高分子聚合物—黑色素,此 為造成園產品褐色、紅色外觀之主要因子 (Mai and Glomb, 2013;Bittner, 2006)。
三、 創傷與酚類生成
酚類物質為酵素型褐化反應之基質,而植物生合成酚類的關鍵步驟酵素—苯 丙胺酸氨基裂解酶 (phenylalanine ammonia lyase, PAL, EC 4.3.1.24; Formerly EC
4.3.1.5),其活性會因植物遭遇逆境而改變 (Dixon and Paiva, 1995)。
Ke 與 Salveit (1989) 指出結球萵苣中肋創傷後,多種酚類含量增加,經氧化後 造成褐化現象,推測受傷的組織會產生創傷信號,此創傷信號會誘導 PAL 活性提 高,進而使酚類含量增加。PAL 活性受影響程度隨距創傷位置增加而減少,且創傷 程度越高則 PAL 活性越高 (Ke and Saltveit, 1989)。
(一) 創傷與創傷信號
創傷為一種植物逆境,當植物細胞或組織遭受破壞後,將影響生理代謝、增加 失水速率,並使微生物較容易由傷口入侵,因此創傷發生後植物會修復受傷組織並 產生防禦反應。植物於傷口處產生創傷信號後,信號傳遞至傷口附近組織以及全株 植物,引發受傷部位的防禦反應 (defense response) 或系統防禦反應 (systemic defense response) 或者兩者皆有之。創傷信號可引發創傷相關基因表現,使受傷組 織復原並進一步誘發防禦機制避免植物再次受到傷害,而創傷誘導的防禦反應包 含:產生不具專一性防禦的酚類、蛋白酶抑制因子 (proteinase inhibitor) 或水解酵 素 (hydrolytic enzyme)、生成植物防禦素 (phytoalexin) 來控制微生物入侵 (León et al., 2001;O'Donnell et al., 1996)。
植 物 中 主 要 的 創 傷 訊 號 為 茉 莉 酸 (jasmonate) 、 甲 基 茉 莉 酸 (methyl jasmonate) 、 寡胜肽系統素 (oligopepetide systemin)、水楊酸 (salicylic acid),另 外如:離層酸 (abscisic aid)、乙烯 (ethylene)、寡糖 (oligosaccharide)、過氧化氫 (hydrogenperoxide) 於不同物種內亦可做為創傷訊號,並且創傷訊號間可能具有交 互關係 (cross talk) ,進而引發不同防禦反應 (León et al., 2001;O'Donnell et al., 1996)。
Ke 與 Salveit (1989) 推測在萵苣中肋受到創傷後,會產生創傷信號使 PAL 活 性上升,並以 PAL 活性受到影響範圍推測此創傷信號以 5 cm•h-1的速率向外擴散 約 2.5 cm,且 PAL 活性受影響程度隨創傷距離增加而減少,並且創傷程度越高則 PAL 活性越高 (Campos‐Vargas and Saltveit, 2002;Kang and Saltveit, 2003;Ke and
21
Saltveit, 1989)。部分植物於創傷後會生成大量乙烯並且誘發酚類生成,而萵苣於創 傷後生成之乙烯量相對其他植物較微量,且其含量於創傷後 24 小時內會回復至創 傷前之生成量 (Ke and Saltveit, 1989)。經 1-MCP (1-methylcyclopropene) 處理的萵 苣於創傷後仍生成大量酚類並產生褐化現象,顯示萵苣於創傷後生成之微量乙烯 並非主要的創傷信號 (Saltveit, 2004)。
創傷後萵苣藉由細胞膜上的磷脂訊號途徑 (phospholipid-signaling pathway) 產生創傷信號。磷脂訊號途徑 (phospholipid-signaling pathway) 為植物產生創傷信 號茉莉酸 (jasmonate acid) 之途徑,對未創傷的完整萵苣外施濃度 1 mM 以下茉莉 酸 (jasmonate) 或 甲 基 茉 莉 酸 (methyl jasmonate) 並 無 法 誘 導 PAL 活 性 上 升 (Campos‐Vargas and Saltveit, 2002);而藉由藥劑 1-phenyl-3-pyrazolidinone (1P3P)、
diethyldithio-carbamic acid (DIECA)、正丁醇 (1-butanol) 或水楊酸 (salicylic acid) 抑制磷脂訊號途徑之中間產物生成後,可以使 PAL 活性及酚類含量不因創傷而上 升,而延遲藥劑施用的時間則會使其抑制效果降低。上述結果顯示萵苣內並非以茉 莉酸與甲基茉莉酸做為創傷信號,而是以磷脂訊號途徑的中間產物做為創傷信號 (Saltveit, 2005)。
(二) 萵苣創傷後 PAL 活性變化
苯丙胺酸氨基裂解酶 (phenylalanine ammonia lyase, PAL, EC 4.3.1.5) 為植物利 用苯丙胺酸 (phenylalanine) 生成 phenylpropanoid 的酵素,為植物生成二次代謝物 酚類物質的關鍵步驟酵素。
PAL 催化苯丙胺酸裂解去氨基化 (deamination),並生成反式肉桂酸 (trans- cinnamic acid),而反式肉桂酸可再被植物轉化成其他酚類物質,如:酚酸 (phenolic acid)、木質素 (lignin)、類黃酮 (flavonoids)、花青素 (anthocyanins)、縮合丹寧 (condensed tannins) 等等。
PAL 為研究植物對逆境反應的重要關鍵酵素。在許多逆境環境下,植物會重
新生合成 PAL,藉此生成相對應的酚類來適應逆境,例如:當植物受到病原菌入侵 時可能會生成香豆素 (coumarins) 或異黃酮 (isoflavones) 防禦,其他逆境如:創 傷、紫外線照射、低溫、乙烯逆境,或是在缺氮、缺鐵、缺磷的情形下,也會生成 相對應的酚類物質 (Dixon and Paiva, 1995)。植物遭遇逆境時,PAL 的表現亦受到 各種與逆境相關的植物賀爾蒙調控,如:乙烯 (ethylene)、茉莉酸 (jasmonate)、甲 基茉莉酸 (methyl jasmonate)、水楊酸 (salicylic acid) 等等 (Lafuente et al., 2003;
Chen et al., 2006)。
經 RT-PCR 分析,羅曼萵苣中肋具有可經創傷誘導的 PAL 基因 (LsPAL1) ,10
℃貯藏下,其轉錄表現於創傷後 12 小時達到最大,並且主要於傷口附近的表皮組 織及維管束組織表現,PAL 活性則接續於創傷後 24 小時達到最大 (Campos et al., 2004)。顯示萵苣於創傷後,創傷信號會誘導 PAL 基因 (LsPAL1) 表現,重新生合 成 PAL,使 PAL 活性上升。
(三) 酚類物質生合成途徑
次級代謝物可分成三類:酚類 (phenolics)、萜類 (terpenoids) 及含氮物質 (nitrogen-containing compounds),次級代謝物在植物內並不直接參與生長與發育,
其功能主要是用來抵禦草食動物或微生物感染、吸引動物幫助受粉或傳播種子、作 為植物間競爭的物質或吸引微生物共生。於高等植物中,大部分酚類物質主要由莽 草酸途徑 (shikimate pathway) 之終點產物—苯丙胺酸 (phenylalanine) 轉換而來,
初 級 代 謝 (primary carbon metabolism) 所 產 生 的 赤 藻 糖 -4- 磷 酸 (erythrose-4- phosphate) 以及磷酸烯醇丙酮酸 (phosphoenolpyruvate) 經由莽草酸途徑後,可轉 換為屬於芳香族胺基酸 (aromatic amino acid) 的苯丙胺酸 (phenylalanine) 與酪氨 酸 (tyrosine)。
苯丙 胺酸 (phenylalanine) 經苯丙胺酸氨基裂解酶 (phenylalanine ammonia lyase, PAL, EC 4.3.1.24; Formerly EC 4.3.1.5) 催化脫去氨基 (amino group) 後生成
23
反式肉桂酸 (trans-Cinnamic acid),此為植物體內最初生成的酚類,因此苯丙胺酸 氨基裂解酶 (PAL) 被視為植物生成酚類的關鍵步驟酵素。反式肉桂酸再經由肉桂 酸 4-羥化酶 (cinnamic acid 4-hydroxylase, C4H) 於苯環上接上一個羥基 (hydroxyl group) 轉變為香豆酸 (p-coumaric acid)。香豆酸 (p-coumaric acid) 可繼續經由苯丙 途徑(phenylpropanoid pathways) 生成木質素 (lignin) 前趨物、 咖啡酸以及咖啡 酰基衍生物 (caffeoyl derivatives),如:綠原酸 (chlorogenic acid);香豆酸輔酶 A (p- poumaric acid-CoA) 經 查 耳 酮 合 成 酶 (chalcone synthase) 即 可 生 成 査 耳 酮 (chalcone) 作為可衍生為類黃酮 (flavonoids) 、花青素 (anthocyanins) 、縮合丹寧 (condensed tannins) 的基本結構 (Taiz and Zeiger, 2006)。
四、 影響萵苣主莖褐化程度的因子
影響園產品創傷後褐化的程度可分為三個因素: (1)氧化基質:酚類的含量與種 類、(2)多酚氧化酶活性、(3)氧化產物的褐化聚合環境
(一) 多酚氧化酶活性與褐化關係
多酚氧化酶 (polyphenol oxidase, PPO) 為園產品內主要進行酵素型褐化反應 之酵素,但PPO活性大小與創傷後褐化程度並無相關性 (Cantos et al., 2001;
Hisaminato et al., 2001)。
Ke與Saltveit (1989) 指出萵苣內PPO活性並不受創傷影響,在萵苣中肋原本組 織內或創傷後組織內的PPO皆有很高的活性,約為PAL活性的100倍且作用快速,
因此創傷後生成之酚類皆足以被PPO氧化。
PPO主要以兩種狀態存在於細胞內,分別為游離態 (soluble form) 或於類囊體 膜上 (thylakoid membrane-bound),於萵苣內以此兩種狀態存在之PPO的比例各占 一半,創傷後於5℃下貯藏七天後只有少部份轉變為游離態,因此PPO存在狀態與 創傷後褐化不具有相關性 (Cantos et al., 2001)。萵苣組織內約有57.5%-72%的PPO
以潛伏狀態 (latent form) 存在,於創傷後,細胞膜降解導致脂肪酸的釋放,並活化 潛伏狀態的PPO,因栽培種的不同,於5℃下約需3.7~6.3天可達到PPO最大活性,
同時間PPO總量並未增加,創傷後並未重新合成PPO,因此在創傷後PPO活性增加 之主要原因為部分潛伏狀態的PPO被活化,即使萵苣中肋於創傷後PPO活性會隨貯 藏期間增加,但品種間褐化程度與PPO活性並沒有相關性 (Mayer, 1986;Cantos et al., 2001)。Hisaminato等 (2001) 也指出萵苣中肋經創傷後三天,PO活性與外觀褐
化程度則無相關性,顯示PPO活性並非影響褐化程度的主要因素 (Cantos et al., 2001;Hisaminato et al., 2001)。
雖然PPO活性並非影響萵苣創傷後褐化程度的主要因素,但為造成褐化的必要 因子,因此仍可以抑制PPO活性達到延緩褐化發生的效果。
(二) 創傷後PAL活性與褐化關係
萵苣原有酚類含量不足以造成嚴重的褐化,因此於創傷後短時間內不會出現 褐化的現象,但因創傷所產生的創傷訊號誘導PAL活性上升,進而使組織內酚類含 量增加,經過多酚氧化酶氧化後,始產生嚴重褐化的現象 (Ke and Saltveit, 1989)。
創傷後PAL活性大小與活性上升所需時間受到創傷程度及貯藏溫度影響,
並且創傷後PAL活性與外觀褐化程度具有相關性 (Hisaminato et al., 2001;López- Gálvez et al., 1996)。萵苣中肋以截切大小作為其創傷程度的區別,將截切大小由 2.5×12 cm縮小至1×0.5 cm,PAL活性在5℃與10℃下分別提高2.5倍與3倍,顯示創 傷程度越高,則其PAL受誘導後活性越高;創傷後在5℃下,PAL活性約於三天內 達到最大,在10℃下則於一天內即可達到最大活性,並且在24小時內下降至原本活 性大小,顯示在10℃下溫度較高時PAL活性變化較快,但活性大小與5℃下差異不大 (López-Gálvez et al., 1996)。
2-Aminoindan-2-phosphonic acid (AIP) 為對PAL具有專一性的競爭型抑制劑,
並且對萵苣PAL也有相同的抑制效果,在4℃下以10 mM AIP溶液浸泡可抑制PAL活 性上升與組織內酚類含量上升,並且可以有效抑制萵苣中肋褐化,顯示控制創傷後
25
酚類生成為抑制褐化的關鍵因素(Hisaminato et al., 2001;Peiser et al., 1998)。
(三) 創傷後萵苣生成酚類種類
創傷後萵苣內 PAL 首先催化生成反式肉桂酸 (trans-cinnamic acid),此為植物 生成最初之酚類,經過苯丙途徑 (phenylpropanoid pathway) 可生成其他同樣為 C6- C3結構的 hydroxycinnamic acid,如: 阿魏酸 (ferulic acid)、芥子酸 (sinapic acid)、
咖啡酸 (caffeic acid),於植物內通常與有機酸酯化或以糖苷 (glycoside) 形式存在,
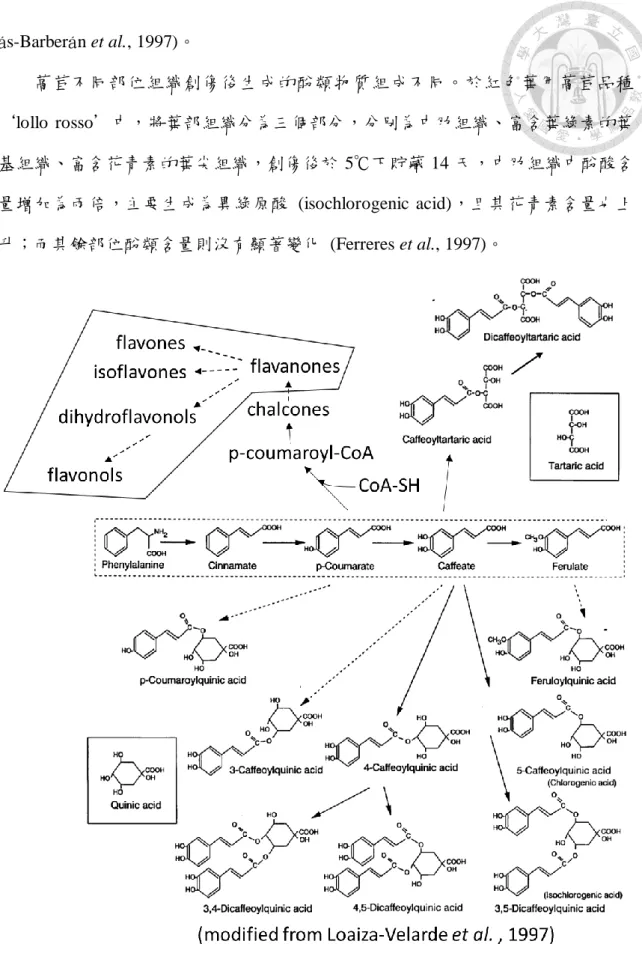
形成 hydroxycinnamic acid derivatives,如:與奎寧酸 (quinic acid) 酯化形成的綠原 酸 (chlorogenic acid) 、與酒石酸 (tartaric acid) 酯化形成的卡夫塔酸 (caftaric acid) 與菊苣酸 (chicoric acid) ,另外也會與蛋白質或細胞壁結合,只有少部分會以自由 酸 (free acid) 的形式存在細胞 (Clifford, 2000)。
萵苣在創傷後生成的主要酚類為咖啡酸衍生物,雖然於結球萵苣葉內原本就 含一定量之類黃酮,但因含量相對較創傷生成之咖啡酸衍生物較少,對創傷引起的 褐化影響不大。結球萵苣葉內之類黃酮主要為榭皮素配醣體 (quercetin glycoside) 與木樨草素配醣體 (luteolin glycoside),約含有 0.3 μg/g FW,創傷後含量會減少 (DuPont et al., 2000;Mai et al., 2013)。
(1) 與奎寧酸酯化之咖啡酸衍生物
綠原酸 (5-O-caffeoylquinic acid, chlorogenic acid) 於許多植物內皆生成,如:番 茄 (Solanum lycopersicum, tomato)、煙草 (Nicotiana tabacum, tobbaco)、蘋果 (Malus domestica, apple)、西洋梨 (Pyrus communis, pear)、咖啡 (Coffea arabica, coffee) 與
朝鮮薊 (Cynara scolymus L., artichoke)等等。綠原酸在各物種內的合成路徑不盡相 同,萵苣內是由 hydroxycinnamoyl-CoA quinate hydroxycinnamoyl transferase (HQT) 利用 caffeoyl-CoA 與 quinic acid 生成綠原酸 (Hisaminato et al.,2001;Ricarda et al.,2004;Steck, 1968;Tomás-Barberán et al.,1997)。在萵苣內綠原酸具有其他同分
異構物 (圖 1),如: neochlorogenic acid (3-O-caffeoylquinic acid)、kryptochlorogenic acid (4-O-caffeoylquinic acid) (Clifford, 2000)。兩個 caffeoyl-CoA 與奎寧酸酯化之咖 啡酸衍生物 (di-esters of caffeic acid))可生成異綠原酸 (3,5-dicaffeoylquinic acid, isochlorogenic acid) (Tomás-Barberán et al.,1997)。
(2) 與酒石酸化酯化之咖啡酸衍生物
菊苣酸 (dicaffeoyltartaric acid, chicoric acid),為酒石酸 (tartaric acid) 與兩個 caffeoyl-CoA 酯化的咖啡酸衍生物,其生合成路徑由一個 caffeoyl-CoA 與酒石酸酯 化產生的 caffeoyltartaric acid,再接上一個 caffeoyl-CoA 即生成菊苣酸 (圖 1)。菊 苣酸與異綠原酸其結構較綠原酸複雜,生合成所需的催化步驟較多。
(3) 咖啡酸衍生物與 PPO 親和性
酚類為 PPO 主要作用的基質,但 PPO 對不同酚類的專一性會因栽培種而異,
例如:不同葡萄栽培種中的多酚氧化酶對咖啡酸 (caffeic acid)、綠原酸 (chlorogenic acid)、兒茶酚 (catechol) 、兒茶素 (d-catechin) 親和性不同 (Yoruk and Marshll, 2003)。由 Km值大小來判斷對基質的親和性,結球萵苣內的 PPO 對綠原酸與咖啡 酸的親和性大於兒茶酚與兒茶素;同時考慮親合性與催化速率,Vm / Km值以綠原 酸最高,為兒茶酚與兒茶素的三倍,以咖啡酸的 Vm / Km值最小,顯示相對類黃酮,
綠原酸為萵苣內極易被 PPO 氧化的酚類 (Altunkaya and Gökmen, 2008)。
(四) 創傷後咖啡酸衍生物含量變化
不同種類的萵苣於創傷後主要生成酚類的種類及組成各不相同。萵苣中肋組 織於創傷後三天,羅曼萵苣、結球萵苣、butter leaf 皆生成 caffeoyltataric acid、菊 苣酸(3,5-dicaffeoyltataric acid, chicoric acid)、綠原酸 (chlorogenic acid)、異綠原酸 (3,5-dicaffeoylquinic acid, isochlorogenic acid)。創傷後三天以結球萵苣生成的總酚 類量最低,其綠原酸含量增加 4.7 倍、異綠原酸增加 3.8 倍、菊苣酸增加 3 倍 (Tom
27 ás-Barberán et al., 1997)。
萵苣不同部位組織創傷後生成的酚類物質組成不同。於紅色葉用萵苣品種
‘lollo rosso’中,將葉部組織分為三個部分,分別為中肋組織、富含葉綠素的葉 基組織、富含花青素的葉尖組織,創傷後於 5℃下貯藏 14 天,中肋組織中酚酸含 量增加為兩倍,主要生成為異綠原酸 (isochlorogenic acid),且其花青素含量也上 升;而其餘部位酚類含量則沒有顯著變化 (Ferreres et al., 1997)。
圖 1. 咖啡酸衍生物與類黃酮物質生合成途徑
Fig. 1. Biosynthesis pathway of caffeic acid derivative and flavonoids
29 五、 延緩褐化反應之處理技術
褐化為造成園產品劣變與商品外觀品質下降的主因之一,除了會降低消費者 購買意願外也可能縮短貯架壽命 (shelf life),甚至降低營養價值,其中截切產品受 到褐化的影響更為直接因此以各種採後處理方法來延緩褐化,。
(一) 調整包裝內氣體成份
氣調貯藏 (control atmosphere storage) 是調節氣體貯藏的簡稱,是指將貯藏環 境內的氣體組成調整至目標比例,使之異於正常大氣的組成。通常藉由添加或移除 特定氣體達到,以降低氧氣濃度或提高二氧化碳濃度來改變園產品原本的代謝速 率 (Kader, 2002)。若是利用包裝材料本身對氣體通透性與園產品呼吸速率達到動 態平衡,並在包裝內形成的特定氣體組成則稱為氣變包裝 (modified atmosphere packaging, MAP)。以氣變包裝使袋內的氧氣濃度維持氧於 0.2-0.5 kPa,可有效控制 截切萵苣褐化,並且不會產生不良風味 (Martinez-Sanchez et al., 2011)。
(二) 熱處理
熱處理為園產品於貯藏前,經過短時間的高溫處理,可分為溫湯、蒸熱及熱風 三種方法,多用於殺死檢疫害蟲或減輕寒害徵狀。植物經熱休克處理後會重新生成 熱休克蛋白 (heat shock protein),而創傷後萵苣經熱處理後會優先生成熱休克蛋白,
使創傷誘導後的 PAL mRNA 無法被轉譯生成,因此熱處理也具有減輕褐化的效果 (Saltveit, 2000;Campos-Vargas et al.,2005)。
(三) 藥劑處理
常見的抗褐化藥劑種類包含抗氧化劑、有機酸、PPO 抑制劑及含鈣化合物。
抗褐化藥劑延緩褐化的效果可能是綜合的結果。抗氧化劑延緩褐化的機制主要是 將多酚氧化酶氧化產生的醌類再還原回雙酚結構,藉此減少醌類累積,因此當抗氧
化劑完全被氧化消耗後,酚類仍會被 PPO 氧化並導致褐化發生;也因此抗氧化劑 延緩褐化的效果會因園產品種類與抗氧化劑濃度不同而改變,常見的抗氧化劑為 抗壞血酸(ascorbic acid)、異抗壞血酸 (isoascorbic acid)、異抗壞血酸鈉 (sodium erythorbate)與含硫氫基化合物,而含硫氫基化合物除了具有還原力外,也具有抑制 PPO 的功能(Kuijpers, 2012; Martinez and Whitaker, 1995)。因 PPO 適合催化的 pH 範圍為 5-7,使用有機酸降低組織內PH 值可使 PPO 活性降低,藉此達到延緩褐化 的效果,常見的有機酸有檸檬酸 (citric acid)、醋酸 (acetic acid)及蘋果酸 (malic acid),而檸檬酸亦具有螯合劑的功能,可使 PPO 活性中心內的銅離子無法作用。
因褐化亦與細胞膜穩定性相關,含鈣化合物主要功能為維持細胞結構穩定來延緩 褐化,並使截切產品維持其脆度與硬度 (Martinez and Whitaker, 1995)。
含硫氫基 (sulfhydryl group, -SH) 之化合物有半胱胺酸 (cysteine, Cys)、麩胱 甘肽 (glutathione)、半胱氨酸鹽酸鹽 (L-cysteine hydrochloride, CysH) 及乙醯半胱 胺酸 (N-acetyl-L-cysteine, NAC),其延緩褐化的機制可能有三種:
(1) 硫氫基與醌類(O-quinone)反應生成無色或淺黃色物質 (Pierpoint,1966 ; Richard et al.,1991;Richard et al.,1992),使醌類無法聚合導致褐化。
如: 綠原 酸經 PPO 氧化生成的醌類與 L-cysteine 反應後可生成 2- cysteinyl-chlorogenic acid (Cabezas-Serrano et al., 2013)。
(2) 本身或與醌類反應的生成物具有抑制 PPO 活性特性
L-cysteine 抑制 PPO 活性程度因濃度而異 (Valero et al., 1991;Ding et al., 1998)。在枇杷內的 PPO,以 1 mM L-cysteine 可抑制 87%的活性
(Ding et al., 1998)。另外,與醌類反應的生成物也可與 PPO 競爭活性部 位,作為 PPO 競爭型抑制物 (competitive inhibition),並抑制 PPO 作 用,使酚類不被氧化成醌類 (Altunkaya and Gökmen, 2008)。
(3) 本身具還原力可將醌類還原回雙酚 (Martinez and Whitaker, 1995)。
31
含硫氫基化合物與醌類反應生成無色物質,避免醌類聚合產生褐化,因此當硫 氫基化合物被完全消耗後,氧化生成的醌類變會開始聚合並造成褐化,也因此特 性,含硫氫基化合物延緩褐化的效果會因園產品種類及施用濃度而異,另外含硫氫
基化合物延緩褐化的效果也會因PH 值改變 (Cabezas-Serrano et al., 2013;Molnar- Perl and Friedman, 1990)。
六、 結球萵苣內咖啡酸衍生物質測定
結球萵苣因生食習慣與做為截切產品的特性,其貯藏後褐化現象有許多研究 探討,如:創傷後褐化、乙烯誘導生理障礙鏽斑病褐化發生,褐化徵狀可出現於結 球萵苣主莖、截切葉片與中肋邊緣,其褐化現象主要為咖啡酸衍生物經酵素氧化後 產生的醌類物質聚合形成,因此萵苣組織內酚類物質組成為研究的方向之一。
Tomás-Barberán 等 (1997) 以甲醇與組織比例 2:1 (v/w) 均質萃取萵苣中肋組 織,經不織布過濾後離心取上清液,上清液於 40℃下濃縮,以 1 ml 50% (v/v) 甲 醇回溶,經 0.45 μm 濾膜過濾後進行 HPLC 分析,HPLC 分析條件為甲醇有機相 與水相添加 5%甲酸進行梯度分離,於波長 326 nm 下偵測結果,顯示結球萵苣中 肋內存在 caffeoyl-CoA 與奎尼酸(quinic acid) 酯化形成的 3-caffeoylquinic acid (neochlorogenic acid)、4-caffeoylquinic acid (kryptochlorogenic acid)、5-caffeoylquinic acid (chlorogenic acid) 、 3,4-dicaffeoylquinic acid 、 3,5-dicaffeoylquinic acid (isochlorogenic acid)、4,5-dicaffeoylquinic acid,以及 caffeoyl-CoA 與酒石酸酯化形 成的 caffeoyltartaric acid、dicaffeoyltartaric acid,但其主要成份為 caffeoyltartaric acid、
綠原酸(chlorogenic acid)、異綠原酸 (isochlorogenic acid) 、菊苣酸 (chicoric acid),
結球萵苣中肋於 5℃下貯藏三天後,綠原酸含量為 15 μg․g-1 FW 增加了 4.8 倍、
異綠原酸含量約為 4 μg․g-1 FW 增加了 3.8 倍、菊苣酸含量約為 10 μg․g-1 FW 增 加了 3 倍,而 caffeoyltartaric acid 含量則增加了 30%。
Hisaminato 等 (2001) 以兩倍重甲醇與組織均質,震盪 10 分鐘後過濾,濾液
進行減壓濃縮,濃縮後加入乙酸乙酯萃取,再進行減壓濃縮,以甲醇回溶後用 Sep- pak C18 層析純化,再經 0.45 μm 濾膜過濾得到樣品進行 HPLC 分離。HPLC 分 析條件為為氰甲烷有機相與水相添加 5%醋酸進行梯度分離,於波長 320 nn 下吸 光,顯示含有主要成份為 caffeoyltartaric acid、綠原酸 (chlorogenic acid) 、異綠原 酸 (isochlorogenic acid) 、菊苣酸 (chicoric acid),經綠原酸標準曲線換算後濃度總 和,萵苣中肋內總酚類含量第 0 天含量約為 10μg․g-1 FW,於 4℃下貯藏三天後總 酚類含量開始上升,第八天含量約為 35μg․g-1 FW,總量增加為 3.5 倍。
兩者分析方法主要相差在於樣品前處理純化步驟與分離使用之流動相不同,
以氰甲烷與水或甲醇與水進行梯度分離後,顯示萵苣中肋創傷後主要生成的酚類 物質為咖啡酸衍生物,包含綠原酸、異綠原酸、菊苣酸。
33
第三章 材料與方法
一、試驗用材料、藥品與儀器 (一) 材料來源
新鮮結球萵苣品種為雲林縣麥寮鄉蔬菜產銷班第 47 班 (台灣生菜村)所栽種 之 6 號,經第二果菜批發市場購入,運回台灣大學園產品處理研究室後放入 1
℃冷藏庫中進行預冷,並待後續處理進行。
(二) 試驗藥品
1. Acetic acid (冰醋酸):皓峯企業股份有限公司。
2. Aluminum Chloride: Sigma。
3. Chlorogenic acid (綠原酸) : Aldrich。
4. Chicoric acid (菊苣酸) : Aldrich。
5. N-acetyl-L-cysteine:Merck。
6. L-cysteine hydrochloride:Merck。
7. Formic acid (甲酸): Sigma-Aldrich。
8. Gallic acid (沒食子酸) : Sigma。
9. Methanol: HPLC grade 及 ACS 試藥級, Macron。
10. Rutin hydrates: Sigma。
11. Sodium dihydrogen phosphate dodecahydrate:
(三) 試驗儀器:
1. 色差儀:德國,Dr LANGE,LMG-160。
2. 切片機:美國,Chef's Choice,Electric food slicer 610。
3. 電子天平:瑞士,Mettler Toledo,AB104-S/FACT。
4. 分光光度計:日本,SHIMADZU,UV-1800。
5. 分光光度計紀錄軟體:日本,SHIMADZU,UVProbe Version 2.42。
6. -20°C 凍箱:美國,Kenmore Model-31205。
7. -80°C 凍箱:日本,Sanyo。
8. 超高速冷凍離心機:日本,HITACHI,CR21GⅢ。
9. 高效能液態層析系統:Intelligent HPLC system LC-800 series,Jasco。
整套系統組成包含六個部分:
a. 幫浦 (pump):日本,Jasco 公司,Model PU-980。
b. 層析管柱 (column):Dikama InspireTM C18 (5μm, 12nm,150 x 4.6 nm) 。
c. 管柱恆溫器 (column oven):台灣,立行科技有限公司。
d. 自動取樣器 (autosampler):德國 SPF 公司,Model S-5200。
e. 紫外光測定儀:日本,Jasco 公司,875-UV。
f. 紀錄軟體:台灣,立行科技有限公司,EC2000。
10. 脫氣用濾紙 (移動相):美國,PALL,FP-Vericel 及 Nylaflo (pore size:
0.45μm,diameter:47mm)。
11. 脫氣用濾紙 (樣品):美國,PALL,PTFE membrane (pore size:0.45μm,
diameter:13mm)。
12. 減壓濃縮系統:
a. 水流抽氣幫浦:日本,Tokyo Rikakikai 公司,Eyela aspirator A-3S b. 恆溫水浴振盪機: 台灣,國洲儀器有限公司,Vapor-mix Model KC-12
二、結球萵苣主莖切片與創傷後藥劑處理
本研究以結球萵苣主莖圓片進行試驗,試驗方法修改自 Tomás-Barberán 等 (1997) 。選用底部未有碰撞傷害或微生物感染之萵苣進行試驗。以不鏽鋼水果刀 將主莖完整的自萵苣取下,利用切片機將底部厚度 0.5 公分部分切除,並將主莖裁 切成厚度 0.5 公分之片狀並保留維管束,以去離子水洗去乳汁,並以擦手紙吸乾切 片表面多餘水分。
將主莖切片分別浸泡於去離子水及不同濃度減輕褐化藥劑中,分別為醋酸 (acetic acid, AcA)、N-乙醯-L-半胱胺酸 (N-acetyl cysteine, NAC)、半胱胺酸鹽酸鹽 (L-cysteine hydrochloride, CysH) ,浸泡五分鐘後取出切片並吸去表面殘餘溶液。將
35
經藥劑處理的切片置於加蓋直徑9公分塑膠培養皿中,並貯藏於5℃下。將藥品秤重 溶解於去離子水定量成指定濃度後,置於4℃冷藏庫備用,處理項目可以分為:
(1)控制組: 浸泡於去離子水中五分鐘
(2)醋酸( AcA)組: 浸泡於3%(v/v)溶液中五分鐘
(3) N-乙醯-L-半胱胺酸 (NAC)組: 浸泡於3% (w/v)及2% (w/v)溶液中五分鐘 (4)半胱胺酸鹽酸鹽 (CysH)組: 分別浸泡於3% (w/v)及2% (w/v)溶液中五分鐘 於藥劑處理後第0、2、4、6、8天進行外觀顏色測量並取出貯藏於5℃的主莖切 片以液態氮急速冷凍,存放於-20凍箱,待進行後續萃取及分析。
三、主莖切片外觀褐化變化觀察
貯藏於 5℃下圓片於藥劑處理後第 0、2、4、6、8 天,拍照並以色差儀 (color pen) 測定主莖切片外觀顏色,結果以 L、a*、b*值表示。
四、主莖切片酚類組成與含量分析 (一) 總酚類萃取
試驗方法參考 Campos-Vargas 和 Saltveit (2002) 之酚類萃取方法,以組織重 量比甲醇體積 1:2 萃取 (w/v)。取 3 克冷凍萵苣主莖切片組織於研缽內經液態氮 研磨至粉狀,將組織粉末置於 50 ml 離心管內,再加入 6 ml 甲醇均勻混和,經 10 分鐘超音波震盪萃取後,以 15000 g 離心 25 分鐘,取上清液至 15 ml 離心管,再 次離心取上清液,置於-20℃下待接續測定。
(二) 總酚類含量測定
試驗方法參考 Loaiza-Velarde 等(1997)、Campos-Vargas 和 Saltveit (2002)之 萵苣組織萃取液內總酚類測定,測定萃取液於波長 320nm 下吸光值。將萃取液以 甲醇等體積稀釋一倍後,以分光光度計測量萃取液於波長 320nm 下吸光值。萃取 液於波長 320 nm 下的吸光值經由綠原酸標準曲線換算為含量,結果以 equivalent chlorogenic acid μg/g FW 表示。
(三) 以 HPLC 分析主要酚類含量變化 1. 萃取液前處理
萃取液於 40℃水浴恆溫震盪機內,減壓濃縮 50 分鐘後以 0.5 ml 甲醇回溶,
經0.45 μm 濾膜過濾得到樣品進行 HPLC 分析。
2. HPLC 分析條件
試驗方法修改自 Tomas-Barberan 等(1997)之分離方法,管柱溫度維持於 40
℃,在流速 1 ml/min 下進行梯度分離。樣品之分析採用紫外光測定儀偵測波長 320 nm 下吸光。測得結果分別以綠原酸與菊苣酸標準品之標準曲線換算濃度,換 算結果分別以μg chlorogenic acid/ g fresh weight 以及 μg chicoric acid/ g fresh weight 表示;異綠原酸的分析結果以綠原酸標準曲線換算。
(1) 移動相: 移動相 A 為去離子水含有 5%(v/v)甲酸 移動相 B 為甲醇含 5%(v/v)甲酸
(2) 分離梯度: 90% 移動相 A 於 18 分鐘內等梯度降至 72% A,再經 14 分鐘 以等梯度降至 65% A,接續於 22 分鐘內再降至 43% A。
(四) 類黃酮含量測定
試驗方法參考 Pekal 與 Pyrzynska (2014),取 0.5 ml 萃取液與 1 ml 2% (w/v) Aluminum Chloride methanol solution 混合均勻,避光靜置反應 10 分鐘後以分光光 度計測定其於 430 nm 之吸光值。以 rutin hydrates 製作標準曲線。換算結果以 μg /g fresh weight 表示。
五、萵苣多酚氧化酶於植物體外氧化酚類標準品 (一) 萵苣主莖內多酚氧化酶之萃取
1. 丙酮粉末:
取 60 g 未經藥劑處理的主莖切片,將組織切碎,以於-20℃下預冷的果汁機內 加入 180 ml 冰冷丙酮,分次以高速均質一分鐘。以墊有濾紙的布氏漏斗經抽氣過
37
濾去除丙酮,再於漏斗中加入 100 ml 冰冷丙酮將破碎的組織攪拌均勻並抽氣過濾,
重覆沖洗組織至丙酮濾液無色且組織中色素均被洗去呈現乳白色為止。將洗淨之 組織倒入培養品內,利用真空乾燥機將組織完全乾燥即得到丙酮粉末,貯藏於-20
℃下。
2. 多酚氧化酶萃取液:
取 0.205 g 丙酮粉末置於燒杯內,加入 40 ml 0.05 M Phosphate buffer (pH 6.8),
在冰浴中攪拌 20 分鐘。經雙層不織布過濾後,用高速離心機離心 10 分鐘 (4℃,
~10,000 xg),離心所得上清液即為酵素粗萃取液,置於冰浴中待用。
(二) 綠原酸、菊苣酸、芸香苷經 PPO 氧化過程之吸收光譜分析
以萵苣主莖內 PPO 分別氧化 20 μg/ml 的綠原酸與菊苣酸,以分光光度計記錄 波長 200-600 nm 間的吸光光譜,每間隔 5 分鐘測定一次連續吸收光譜。
以萵苣葉 PPO 酵素液氧化不同濃度下的酚類標準品溶液及其混合液,以分光 光度計記錄波長 270-450 nm 間的吸光光譜,於反應後 0、15、90、180、300、600、
900、1200、1800 秒紀錄連續吸收光譜。
(三) 酚類標準品經 PPO 氧化後褐化顏色值之測量
於容量為 4 ml 的吸光液槽 (cuvette) 加入 1 ml 萵苣葉 PPO 酵素液與 2 ml 不 同濃度酚類標準品反應並拍照紀錄褐化過程。氧化反應 40 分鐘後褐化外觀維持一 定,於固定光源條件下拍照記錄,以軟體 ImageJ 進行影像顏色分析,將影像轉換 為色彩空間 RGB 數值,再轉換以色彩空間 L*、a*、b*數值表示。
第四章 結球萵苣主莖切片創傷後酚類之生成與褐化間之關係
結球萵苣於田間採收時會由主莖部分切下並保留完整葉球,此一採收切口經 過一段貯藏期間後會產生褐化的現象,並使結球萵苣外觀品質與貯架壽命降低。結 球萵苣主莖創傷後,組織內 PAL 活性會因創傷誘導而上升,因而生合成酚類物質,
當酚類物質經 PPO 氧化後產生之醌類會聚合形成褐色物質,並逐漸造成褐化。本 試驗欲了解創傷後結球萵苣主莖切片於 5℃下貯藏期間,主要產生之酚類物質種類 以及酚類物質含量變化;並藉由植物體外試驗,以結球萵苣多酚氧化酶 (PPO) 氧 化不同酚類標準品溶液以及其混合溶液,進而探討酚類物質種類經 PPO 氧化後,
與造成褐化程度之關係。
一、以 HPLC 層析方法分離主莖切片內酚類物質
萵苣主莖切片創傷後經去離子浸泡五分鐘後貯藏於 5°C 下,其切片外觀於創 傷後第二天即出現維管束褐化,並於第四天外觀褐化程度加劇 (圖 2),顯示與褐化 相關的酚類物質含量於創傷後即開始增加;主莖切片內的總酚類含量 (圖 19) 於 前四天快速增加,第四天後含量上升趨於平緩,與外觀褐化變化相似,而其總類黃 酮含量則是於第四天後上升至第六天,第六天後則維持一定含量。因此進一步以 HPLC 分析創傷後萵苣主莖切片內主要所含的酚類物質種類。
HPLC 層析試驗方法參考 Tomás-Barberán 等 (1997),使用 C18層析管柱,改變 移動相比例進行不同強度的沖提,使用有機相甲醇添加 5%甲酸以及水相添加 5%
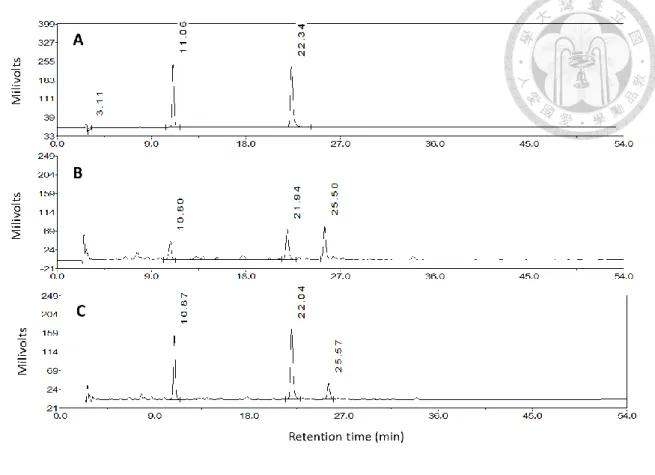
甲酸,以不同比例混合進行梯度分離。對照組外觀於第二天即出現明顯褐化徵狀 (圖 2),並隨貯藏時間增加而褐化加劇,且前四天外觀褐化之變化明顯,顯示與褐 化相關的酚類物質含量於創傷後即開始增加,因此取褐化變化明顯的創傷後第四 天之主莖切片進行 HPLC 組成分析,對照組主莖貯藏後第四天之層析圖中有主要
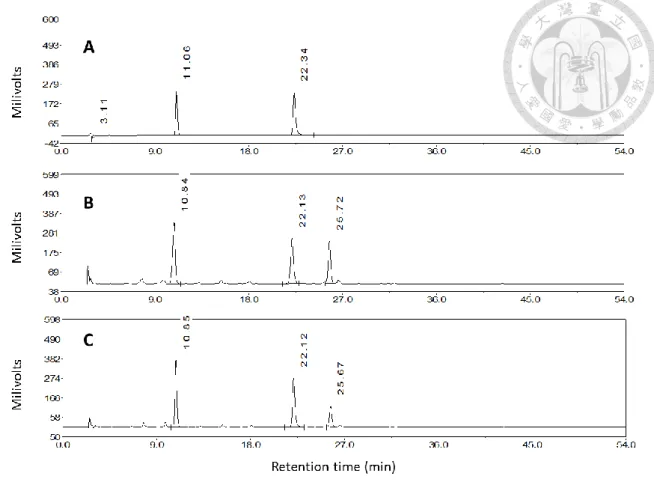
39
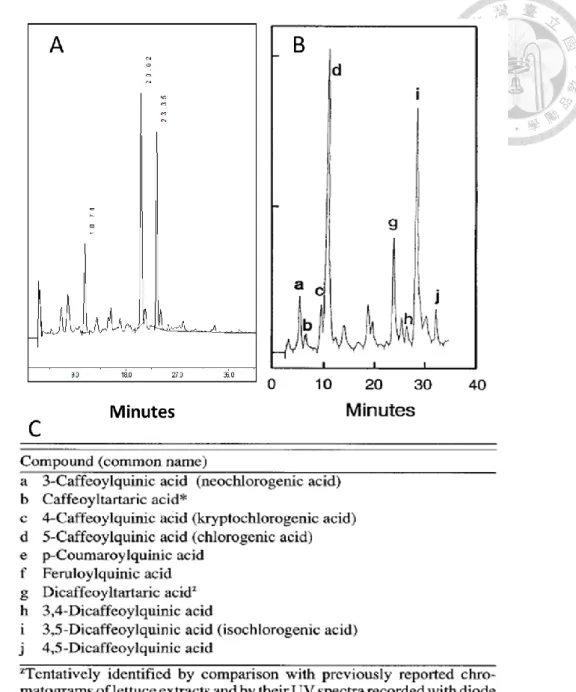
三個訊號,與標準品滯留時間相對照,標準品綠原酸與菊苣酸經層析後其滯留時間 分別為 10.89 分鐘與 22.16 分鐘 (圖 3),與對照組主莖貯藏後第四天之層析圖 (圖 3) 中出現高峰的時間相符合,可確認首先沖提出的前兩個訊號分別為綠原酸與菊 苣酸,而第三個主要訊號由文獻推測應為異綠原酸 (Tomás-Barberán et al.,1997)。
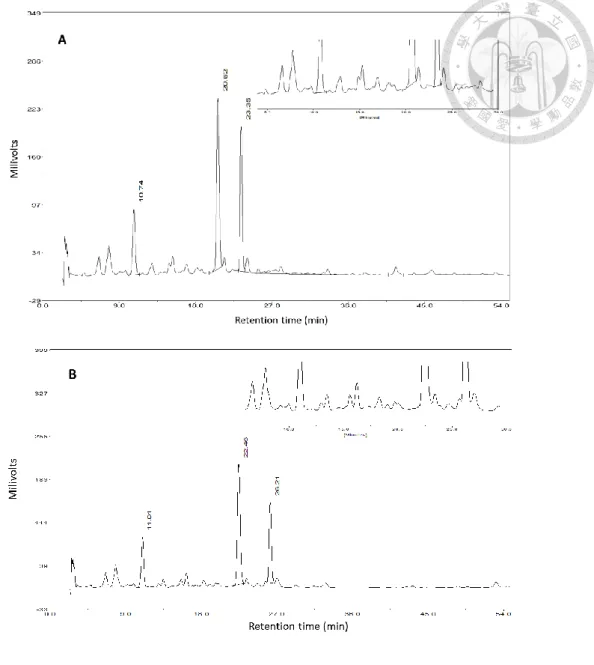
分離起始條件為 10%甲醇相經 40 分鐘等梯度升至 58%甲醇相,甲醇比例以每 分鐘 1.2%上升,對照組貯藏後第八天的萃取液層析圖(圖 4A) 含有三個主要訊號,
經標準品滯留時間比對後沖提出的前兩個訊號分別為綠原酸與菊苣酸,與 Tomás- Barberán 等 (1997) 結果相似,而雖滯留時間相異,但因分子極性差異沖提出的相 對時間比例相近,因此與文獻層析結果(圖 4B)對照後,沖提出的第三個訊號應為 異綠原酸,後續高效能液相層析分析之異綠原酸結果以綠原酸標準曲線換算其含 量。
在主要訊號綠原酸、菊苣酸、異綠原酸間,仍有其於含量較微量的訊號 (圖 4),
可能為異綠原酸的同分異構物 3,4-dicaffeoylquinic acid 或其他未知物質。於主要訊 號間仍有不對稱的波峰,可能為重疊的波峰,因此降低甲醇比例上升速率,由每分 鐘 1.2%改為每分鐘上升 1%,並且分離時間中段的梯度則降為每分鐘 0.5%(圖 4),
沖提強度降低後,綠原酸與菊苣酸間重疊的訊號分離產生新的波鋒,異綠原酸訊號 前則出現新的波鋒,雖主要訊號外的物質含量低,但為減少定量誤差,最後 HPLC 分離梯度修改為起始條件為 10%甲醇,並以甲醇比例每分鐘 1%之速率上升 18 分 鐘,接續以每分鐘 0.5%之速率上升 14 分鐘後,再維持以每分鐘 1%之速率上升 22 分鐘,最後達到甲醇比例為 57%。
二、確認創傷後萵苣主莖萃取液內主要酚類物質
綠原酸與菊苣酸標準品經上述分離條件於 HPLC 內的滯留時間分別為 10.89 分鐘與 22.16 分鐘,與對照組第四天層析圖 (圖 3) 中出現高峰的時間相符合,因