國立臺灣大學理學院海洋研究所 博士論文
Institute of Oceanography College of Science
National Taiwan University Doctoral Dissertation
臺灣東部海域雨傘旗魚之營養階層結構與攝食生態研究 Trophic Size-structure and Feeding Ecology of Sailfish,
Istiophorus platypterus, in Eastern Taiwan Waters
蔡政南 Chung-Nan Tsai
指導教授:孫志陸博士 Advisor: Chi-Lu Sun, Ph.D.
中華民國 103 年 7 月
July, 2014
謝 辭
本篇論文得以順利完成,首先感謝 恩師孫志陸博士及師母 葉素然女 士的悉心指導,在工作及學業雙重之壓力下,仍不斷給與精神勉勵,持續 教導與提攜,讓後學得以在困境下完成本研究,在此致上最深摯的謝意。
本論文審查承蒙臺灣海洋大學環境生物與漁業科學系 李國添講座教 授及 莊守正博士、海洋事務與資源管理研究所 劉光明博士、中央研究院 生物多樣性研究中心 邵廣昭研究員、本校漁業科學研究所 曾萬年名譽教 授、本校生命科學系 丘臺生博士、及中興大學生命科學系 林幸助博士等 先進,對本論文細心審閱與指正,惠賜寶貴建議,使本論文更趨完善,在 此致上最高謝意。
本論文於研究期間承蒙水產試驗所東部海洋生物中心 江偉全博士及 陳淑穎女士協助收集雨傘旗魚與餌料生物樣本,以及資料分類彙整;中央 研究院生物多樣性研究中心 邵廣昭博士及陳宣汶博士協助提供實驗室進 行樣本前置處理作業;環境變遷研究中心 高樹基博士協助穩定同位素樣 本檢測處理;客座專家 Michael K. Musyl 博士協助潤飾英文初稿;本研究 室 曾振德博士、蘇楠傑博士、張以杰博士等協助指導繪圖及統計軟體的 應用,以及學弟妹們協助處理院所相關事務,在此致上萬分謝意。
最後感謝我的家人與親友們,高雄海洋科技大學 陳哲聰前校長及師 母,以及本人服務單位的水利及土木技師同事們,不斷給予關心與支持,
提供本人研究前進的精神動力,本人在此由衷感激,期待本人於進修期間 所學習的專長,得以應用於海事及港灣等公共工程,以及對於推動海洋環 境生態保育與生物資源之永續利用,盡個人棉薄之力,以不負眾師長及各 先進們的厚愛。
蔡政南 2013 年 7 月於臺大海洋研究所
臺灣東部海域雨傘旗魚之營養階層結構與攝食生態研究
中文摘要
過去漁業資源的利用與評估著重於單一魚種的管理模式,為資源永續利用,
FAO提出以生態系為基準之漁業管理方式,其漁業管理方式需考慮物種間的營養階 層結構與能量流,有關雨傘旗魚的族群特徵已累積許多研究成果,但對於食物階層 結構研究卻是完全闕如。本研究為利用雨傘旗魚肌肉之氮、碳穩定同位素來探討其
食物階層結構,經檢測雨傘旗魚肌肉樣本 (n=263) 之氮同位素 (δ15N) 的測量值為
7.51‰至 14.19‰,平均值為 12.05 ± 1.23‰;碳同位素 (δ13C) 的測量值為-22.04‰
至-15.48‰,平均值為-17.68 ± 1.15‰。氮同位素的測量值與雨傘旗魚之體長呈正相 關;碳同位素的測量值與體長呈負相關。各體長組間的雨傘旗魚之估算營養階層,
顯示不同體型之雨傘旗魚分別位於生態系中不同的生態位階,推論影響的因素為餌 料食物來源、棲地環境及洄游路徑之不同。
穩定同位素分析是根據消費者穩定同位素比值與其食物來源之同位素比值相 近的原理,進而推估餌料生物來源貢獻,可以反映生物長期攝食生態的結果。本研
究以雨傘旗魚及其餌料生物之碳、氮穩定同位素,本研究應用Stable isotope mixing
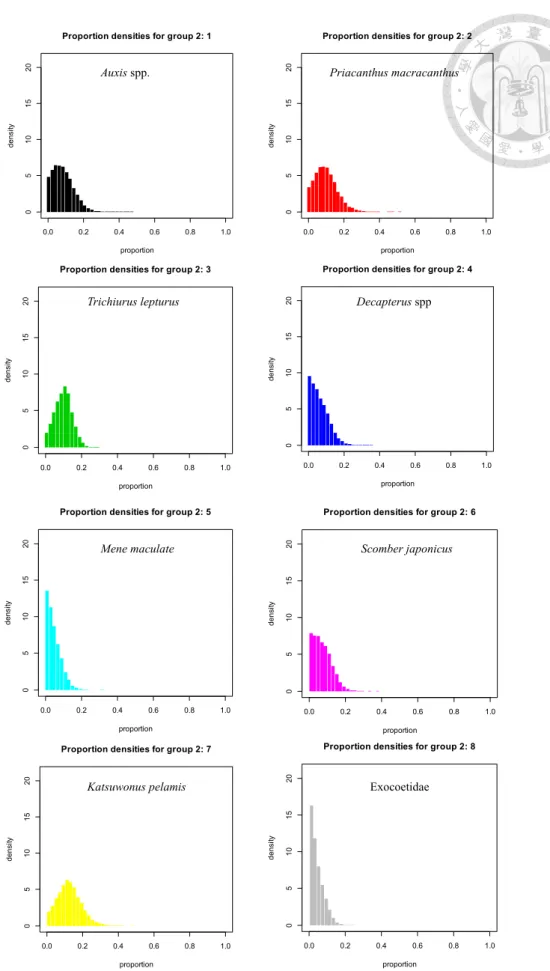
model,來推算雨傘旗魚的餌料生物來源組成,依先前生殖研究的結果,將雨傘樣 本分為幼魚(<140 cm, LJFL )、成熟中(141-180 cm, LJFL)及成魚(>181 cm, LJFL)三 組。研究結果顯示,雨傘旗魚幼魚攝食以烏魴科(15.90%)為最多、其次為眼眶魚 (11.04%)與鯡科(9.62%);而雨傘旗魚成魚攝食以白帶魚(32.63%)為最多、其次為正 鰹(15.82%)與紅目鰱(11.30%)。雨傘旗魚的食物來源組成種類多樣,包含有:頭足 類、沿岸底棲魚類、表水層魚類、大洋中層魚類,以穩定同位素分析結果,雨傘旗 魚食物來源主要為表水層魚類為佔多數。
魚類食物網是海洋生態系中物質循環與能量流動的重要途徑,營養結構則是顯 示能量流動過程中,消費者與被捕食者之間的營養關係。本研究根據胃內容物與穩 定同位素分析推估雨傘旗魚的攝食組成,參考漁獲統計資料,應用Ecopath with Ecosim 建構台灣東部海域生態系統營養結構模型,並經由模型分析系統能量流動
和總體特徵。結果顯示台灣東部海域生態系統各功能群的營養階層範圍為 1 至
4.52,哺乳動物及沙魚佔據了最高的營養階層。利用生態網路分析結果,系統總吞
吐量為 3054.43 t/km2/yr;系統總生產量、總消費量及總呼吸量分別為 1787.29、
824.89 、 528.62 t/km2/yr ; 總 初 級 生 產 量 / 總 呼 吸 量 為 3.13 。 系 統 連 接 指 數 (Connectance Index, CI) 和系統雜食指數 (System Omnivory Index, SOI) 分別為 0..47 和 0.25。本研究初步建立台灣東部大洋海域生態系統模型,探討生態系的結 構與功能,提供未來建置以生態系為基礎的漁業管理相關科學的資訊。經由Ecosim
模式預測各功能組的未來50 年後生物量變動,其結果顯示,如維持目前的漁業規
模,對於海洋哺乳類、鮪魚、旗魚、沙魚等大型掠食性魚種,將會造成資源量的減 少。本研究結果與過去漁獲量統計變動的趨勢相符合,為持續小幅度下滑降低;顯 示目前台灣東部海域漁獲努力量仍偏高,建議應進一步檢討降低。
關鍵字:雨傘旗魚、穩定同位素、營養階層結構、攝食組成、能量流動、
生態系模型。
Trophic Size-structure and Feeding Ecology of Sailfish, Istiophorus platypterus, in Eastern Taiwan Waters
Abstract
Fishery stock assessment and management used to focus on single species method. FAO promoted the ecosystem-based fishery management (EBFM) for the sustainability of living aquatic resources. EBFM need to be taken into account with trophic structure and flows of biomass through species interactions. In present study, nitrogen and carbon stable isotopes analyses were applied to investigate the trophic structure of sailfish. A total of 263 sailfish muscle samples were examined. The δ15N values for sailfish ranged from 7.51 to 14.19 ‰ (mean=12.06‰ ± 1.16) and the δ13C values ranged from -22.04 to -15.48 ‰ (mean=-17.62‰ ± 1.10). The δ15N values were positively related to body length, while δ13C were negatively related to body length. The results revealed the different size class of sailfish occupied different trophic position of pelagic ecosystem.
We postulate that the factors which influence on trophic position of sailfish between young and adult were food sources, habitats and migratory pattern.
Stable isotope ratios of a consumer are related to those of its prey. Stable isotope mixing model was used to estimate diet composition of consumer by mass balance equation. In this study, stable isotope mixing model was applied by carbon and nitrogen stable isotopes of sailfish to estimate the food sources of their prey. The sailfish were segregated by juveniles (<140cm, LJFL), maturing stage (141-180cm, LJFL) and adults (>181cm, LJFL) from previous research. The most important preys for juvenile sailfish were Bramidae (15.90%), Mene maculata (11.04%) and Clupeidae (9.62%). Therefore, adult sailfish feed on Trichiurus lepturus (32.63%), followed by Katsuwonus pelamis (15.82%) and Priacanthus macracanthus (11.30%). In spite of the apparent high prey
diversity of diet composition for sailfish, they feed mainly on epipelagic species in the waters off eastern Taiwan.
Food web is an important element in pelagic ecosystem to transport nutrients and energy flow between consumers and prey. In order to investigate the role of sailfish and to explore the function and structure of pelagic ecosystem in eastern Taiwan waters, we applied Ecopath with Ecosim model inferred from stable isotope analysis, stomach content analysis and fisheries statistical data to perform pelagic trophic model of eastern Taiwan. The results reveal that the trophic level of the functional groups varied from 1.0 to 4.52. Marine mammals and shark are occupied top trophic level. Total ecosystem throughput is 3054.43 t/km2/yr. Sum of all production, sum of all consumption and sum of all respiratory flows are 1787.29, 824.89 and 528.62 t/km2/yr, respectively. Total primary production to total respiration ratio is 3.13. Connectance Index (CI) and System Omnivory Index (SOI) are 0.47 and 0.25, respectively. In this study, we preliminary established pelagic trophic model in eastern Taiwan and the results can provide scientific references for ecosystem-based fisheries management. Ecosim model was applied to predict biomass trajectories of each functional groups in next 50 years. The predicted biomass trends indicate that if we maintain current fishing efforts in eastern Taiwan will reduce most of biomass of top predators in the future (e.g. Marine mammals, tuna, billfish and shark). The results show current fishing efforts in eastern Taiwan is still high, we suggest the fish stock need to be monitored and fishing efforts should be further reduced.
Keywords: sailfish, stable isotopes, trophic structure, diet composition, energy flow, Ecopath model.
臺灣東部海域雨傘旗魚之營養階層結構與攝食生態研究
Trophic Size-structure and Feeding Ecology of Sailfish, Istiophorus platypterus, in Eastern Taiwan Waters
目錄 Table of Contents
中 文 摘 要 --- I Abstract --- III
第 一 章 緒 論 - - - 1
1 . 1 雨 傘 旗 魚 生 態 與 資 源 概 況 - - - 1
1 . 2 相 關 研 究 - - - 5
1 . 3 研 究 目 標 與 論 文 概 要 - - - 8
1.3.1 研究目標 --- 8
1.3.2 論 文 概 要 - - - -- - - 9
第 二 章 雨傘旗魚營養階層結構 --- 11
2.1 前言 --- 11
2 . 2 材 料 與 方 法 - - - 1 5 2.2.1 研究地點 --- --- 15
2.2.2 樣本收集 --- 15
2 . 2 . 3 穩 定 同 位 素 分 析 - - - 1 6 2.2.4 資料分析 --- 18 2 . 3 結 果 - - - 1 9 2 . 3 . 1 體 長 與 同 位 素 之 關 係 - - - 1 9 2 . 3 . 2 體 長 級 別 與 同 位 素 之 關 係 - - - 1 9 2 . 3 . 3 採 樣 區 域 之 差 異 性 - - - 2 0 2 . 3 . 4 季 節 差 異 - - - 2 1 2 . 3 . 5 營 養 階 層 估 算 - - - 2 1 2 . 4 討 論 - - - 2 2
2 . 4 . 1 體 長 與 氮 同 位 素 - - - 2 2 2 . 4 . 2 體 長 與 碳 同 位 素 - - - 2 3 2 . 4 . 3 營 養 階 層 結 構 - - - 2 4 2 . 4 . 4 雨 傘 旗 魚 與 其 他 大 洋 掠 食 者 比 較 - - - 2 5
第 三 章 雨 傘 旗 魚 之 食 性 組 成 及 攝 食 生 態 - - - 2 8 3 . 1 前 言 - - - -- - - 2 8 3 . 2 材 料 與 方 法 - - - 3 1 3 . 2 . 1 樣 本 收 集 - - - 3 1 3 . 2 . 2 胃 內 容 物 分 析 - - - 3 1 3 . 2 . 3 穩 定 同 位 素 分 析 - - - 3 3 3 . 2 . 4 同 位 素 混 合 模 式 - - - 3 4 3 . 3 結 果 - - - 3 6 3 . 3 . 1 胃 內 容 物 分 析 - - - 3 6 3 . 3 . 2 穩 定 同 位 素 特 徵 - - - 3 6 3 . 3 . 3 同 位 素 混 合 模 式 - - - 3 7 3 . 3 . 4 雨 傘 旗 魚 與 餌 料 食 物 階 層 結 構 - - - 3 8 3 . 4 討 論 - - - 4 0 3 . 4 . 1 攝 食 組 成 比 較 - - - 4 0 3 . 4 . 2 同 位 素 混 合 模 式 分 析 - - - 4 1 3 . 4 . 3 雨 傘 旗 魚 攝 食 與 洄 游 - - - 4 2 3 . 4 . 4 雨 傘 旗 魚 與 餌 料 食 物 階 層 結 構 - - - 4 4
第 四 章 臺 灣 東 部 海 域 大 洋 生 態 系 模 式 建 置 - - - 4 5 4.1 前言 --- 45 4.2 材料與方法--- 48 4 . 2 . 1 生 態 系 模 式 之 建 構 - - - 4 8 4 . 2 . 2 資 料 收 集 - - - 5 1 4 . 2 . 3 生 物 群 組 - - - 5 3
4 . 2 . 4 資 料 分 析 - - - 5 5 4.3 結果 --- 58 4 . 3 . 1 臺 灣 東 部 海 域 生 態 模 型 - - - 5 8 4 . 3 . 2 營 養 階 層 結 構 與 能 量 流 動 - - - 5 8 4 . 3 . 3 混 合 營 養 效 應 分 析 - - - 5 9 4 . 3 . 4 生 態 系 總 體 特 徵 - - - 6 0 4 . 3 . 5 生 物 量 變 動 趨 勢 - - - 6 1 4 . 4 討 論 - - - 6 2 4 . 4 . 1 營 養 階 層 結 構 與 能 量 流 動 - - - 6 2 4 . 4 . 2 生 態 系 總 體 特 徵 - - - 6 2 4 . 4 . 3 生 物 量 變 動 趨 勢 - - - 6 3
第 五 章 結 論 與 建 議 - - - 6 6 5.1 結論 --- 66 5.2 建議 --- 70
參 考 文 獻 - - - 7 2 附 圖 - - - 8 6 附 表 - - - 1 2 1
List of Figures
Fig.1. Indo-Pacific sailfish Istiophorus platypterus (Nakamura, 1985).
Fig. 2. Geographical distribution of sailfish.
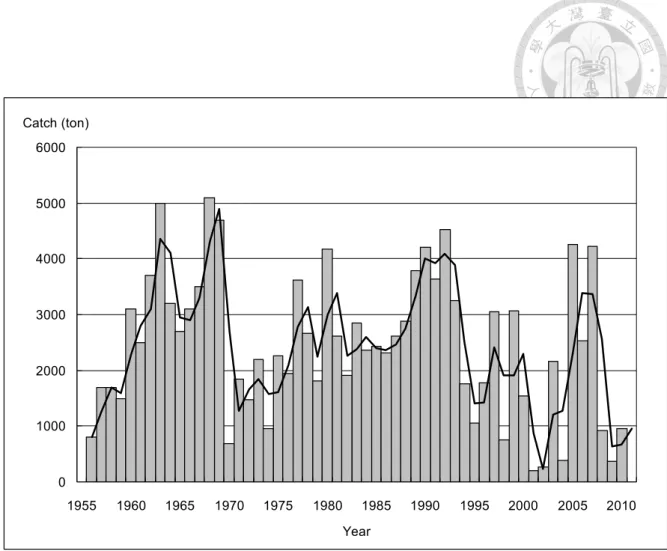
Fig. 3. Catch of Indo-Pacific sailfish in Taiwan from 1956 to 2010 (Data source:
Statistical Yearbook, Taiwan).
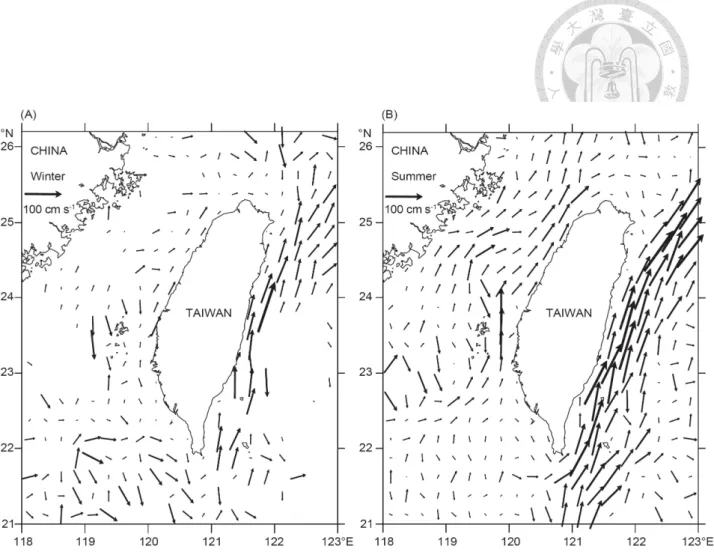
Fig. 4. Current velocity and direction in the surrounding seas of Taiwan during winter (A) and summer (B).
Fig. 5. Map of study area showing locations of sailfish samples taken for stable isotope analysis from eastern Taiwan (oblique lines, n=233) and from South China Sea (cross lines, n=30) during April 2009 and February 2012.
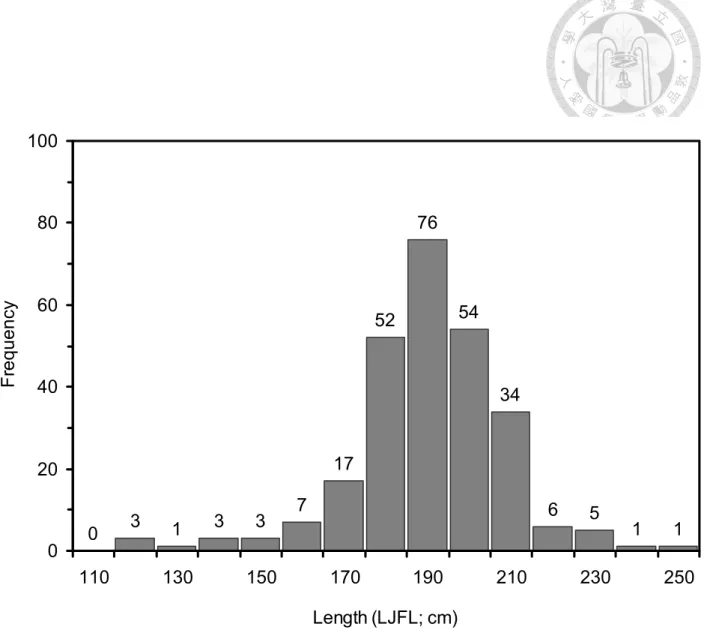
Fig. 6. Lower jaw fork length distribution (by 10 cm intervals) of 263 sailfish off eastern Taiwan water analyzed in this study.
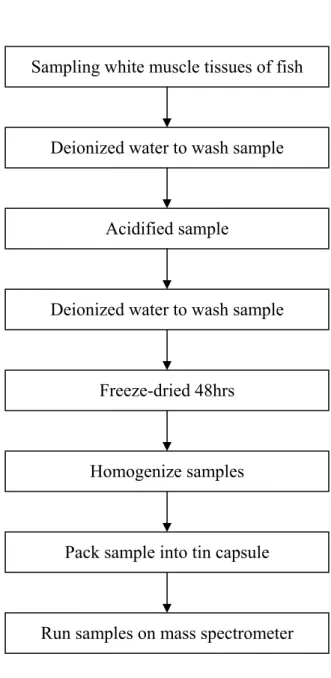
Fig. 7. The flow chart for stable isotope analysis in this study.
Fig. 8. Length of lower jaw to fork length (LLJFL) of sailfish plotted versus δ13C values.
Fig. 9. Length of lower jaw to fork length (LLJFL) of sailfish plotted versus δ15N values.
Fig. 10. Mean carbon isotopic composition (δ13C, ‰; ± S.D.) of each size class of sailfish analyzed in study.
Fig. 11. Mean nitrogen isotopic composition (δ15N, ‰; ± S.D.) of each size class of sailfish analyzed in study.
Fig. 12. Seasonal variation in (a) δ13C and (b) δ15N values (mean±s.d.) for Istiophorus platypterus from 2009 to 2011 (SP, spring; SM, summer; AU, autumn; WN, winter).
Fig. 13. Biplot of δ13C and δ15N values (mean ± S.D.) from each size class of sailfish.
Fig. 14. Relationship between length of lower jaw to fork length (LLJFL, cm) and trophic level (TL), using δ15N concentrations of the herbivorous marine snail
(Chlorostoma argyrostoma argyrostoma Gmelin 1791) 6.24 ‰ ± 0.29 S.D. as a trophic baseline. Also shown are the midpoints of the six size classes of sailfish in the study.
Fig. 15. Map of study area showing position of capture of samples taken for stomach content of sailfish and isotope analyses off eastern Taiwan during April 2009 and February 2012.
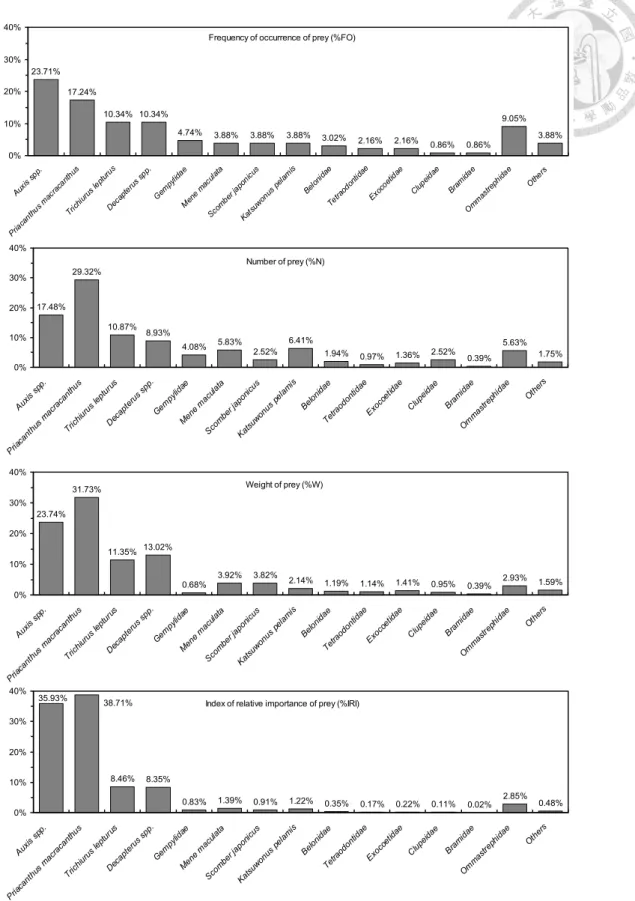
Fig. 16. Percent composition of sailfish stomach contents collected off eastern Taiwan water: frequency of occurrence (%FO), number (%N), weight (%W) and index of relative importance (%IRI) of each prey groups.
Fig. 17. Estimated percentage contributions (mean and 95% credibility intervals) of prey consumed by (a) juvenile (<140cm, LJFL), (b)maturing (141-180cm, LJFL) and (c) adult (>181cm, LJFL) sailfish.
Fig. 18. Estimation of prey group contributions for juvenile sailfish (<140cm, LJFL) by using stable isotopes mixing models.
Fig. 19. Estimation of prey group contributions for maturing sailfish (141-180cm, LJFL) by using stable isotopes mixing models.
Fig. 20. Estimation of prey group contributions for adult sailfish (>181cm, LJFL) by using stable isotopes mixing models.
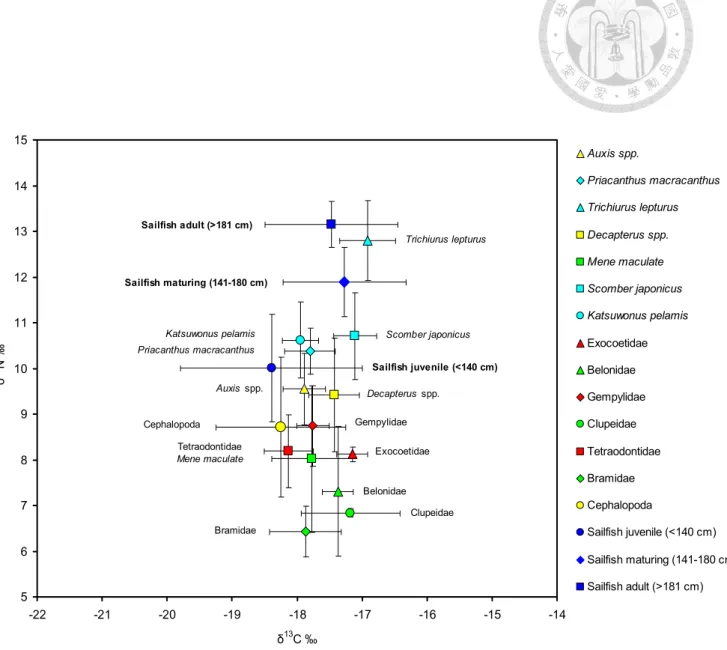
Fig. 21. Biplot of mean δ13C and δ15N values of sailfish (Istiophorus platypterus) and their prey.
Fig 22. Estimated trophic level (TL) (mean ± SD) for sailfish and their pery, using δ15N concentrations of the herbivorous marine snail, Chlorostoma argyrostoma argyrostoma Gmelin 1791, 6.24 ‰ ± 0.29 S.D. as a trophic baseline (Tsai et al., 2014).
Fig. 23. Global trends in the state of world marine fish stocks, 1974-2011 (Data source:
FAO, 2014).
Fig. 24. The study area of eastern Taiwan pelagic ecosystem.
Fig. 25. Annual fishery production and market value for eastern Taiwan from1993 to 2012 (Data source: Fisheries Statistical Yearbook, Taiwan).
Fig. 26. Total number of fishing crafts for eastern Taiwan from 1993 to 2012 (Data source: Fisheries Statistical Yearbook, Taiwan).
Fig. 27. The total tonnage (a) and horsepower (b) of fishing crafts for eastern Taiwan from 1993 to 2012 (Fisheries Statistical Yearbook, Taiwan).
Fig. 28. Annual landing of (a) tuna, (b)billfish, (c)shark for eastern Taiwan from 2002 to 2012 (Fisheries Statistical Yearbook, Taiwan).
Fig.29. Trophic relationships of functional groups in the eastern Taiwan pelagic ecosystem. Green lines indicate the relationships between sailfish and their
preys.
Fig. 30. The trophic flows transmitted through aggregated trophic levels in eastern Taiwan pelagic ecosystem.
Fig. 31. The mixed trophic impact analysis of functional groups in eastern Taiwan pelagic ecosystem.
Fig. 32. Simulation of relative biomass of functional groups for eastern Taiwan pelagic ecosystem.
Fig. 33. Predicted biomass trajectories of functional groups for eastern Taiwan pelagic ecosystem.
Fig. 34. The simulation of catches of main target fishing groups for eastern Taiwan pelagic ecosystem.
List of Tables
Table 1. Statistics of sailfish, Istiophorus platypterus, and corresponding isotopic values by size classes
Table 2. Estimated trophic level for different size classes of sailfish with mean values provided ±SD.
Table 3. Comparison of isotopic values for δ15N (‰) and δ13C (‰) for sailfish and other top predators in pelagic ecosystems.
Table 4. Diet composition of Istiophorus platypterus in eastern Taiwan by stomach content analysis.Length of fish prey items given in fork length, “Others”
indicates unidentified contents.
Table 5. Isotope values of δ13C (‰) and δ15N (‰) for sailfish and their prey items sampled off eastern Taiwan.
Table 6. Estimated percentage contributions (mean and 95% confidence intervals) by stable isotope mixing model of prey consumed by juvenile (<140cm, LJFL) and adult (>180cm, LJFL) sailfish.
Table 7. Functional group components of the eastern Taiwan pelagic ecosystem model.
Table 8. Basic input and output (bold) parameters of the trophic model for eastern Taiwan pelagic ecosystem.
Table 9. Diet composition matrix for all function groups of eastern Taiwan pelagic ecosystem.
Table 10. A tabular model of ecological succession: trends to be expected in the development of ecosystems (modified from Odum, 1969).
Table 11. Ecosystem properties for eastern Taiwan (this study) and East China Sea (Cheng et al., 2009).
Table 12. Change rate in biomass of functional groups after 50 year simulation for eastern Taiwan pelagic ecosystem by using Ecosim model.
第一章 緒論
1.1 雨傘旗魚生態與資源概況
一、分類
雨傘旗魚Istiophorus platypterus (Shaw & Nodder 1792) (如Fig. 1 所示),
英名:sailfish,臺灣俗稱『破雨傘』或『雨笠仔』,在分類學上屬於硬骨魚 綱 (Class Osteichthyes),條鰭亞綱 (Class Actinopterygii),鱸形目 (Order Perciformes) , 鯖 亞 目 (Suborder Scombroidei) , 正 旗 魚 科 (Family Istiophoridae),旗魚屬 (Genus Istiophorus)。 (沈等,1993)。
二、形態特徵
本種體型延長,略呈圓筒形,稍側扁;尾柄細而強壯,具二隆起脊。頭 較長;吻長而尖,呈劍狀突出。口大,微斜裂。頜齒呈絨毛狀齒帶,鋤骨無 齒。體被細長骨質鱗。第一背鰭軟條部顯然較體高為高,呈帆狀;第二背鰭 短小;胸鰭位低,呈鐮刀狀;腹鰭胸位,起點在胸鰭基底下方稍後,向後延 伸至肛門;尾鰭深叉形。體背紫黑色,腹部銀白色,體側無橫帶和圓斑。第 一背鰭藍黑色並有若干黑點;腹鰭藍黑色;尾鰭暗色 (Shao, 2014)。相較於 其它種類旗魚,則其體型較小且呈瘦扁型。
三、地理分布
本種廣泛分布於印度洋及太平洋之熱帶、亞熱帶海域,在西太平洋分布 於45°N 與 40°S 之間,在東太平洋分布在 35°N 與 35°S 之間 (Froese and Pauly, 2014) (Fig. 2)。臺灣各地均有產,尤以東部及南部產量最多 (Shao, 2014)。
四、棲所生態
雨傘旗魚主要棲息於表層水域 (Jolley and Irby, 1979; Boggs, 1992;
Nakano et al., 1997),具有明顯之季節性分佈 (Beardsley et al., 1975; Speare, 1995)。此外,雨傘旗魚的地理分佈亦受到 28 ℃表水溫 (Ovchinnikov, 1966) 與鹽度的影響 (Kuwahara et al., 1982)。在西太平洋雨傘旗魚幼魚及成魚的分 布與黑潮洋流密切相關,在產卵季節時密集成群 (Nakamura, 1985)。雨傘旗 魚在臺灣東部海域產卵 (Chiang et al., 2006a; b),漁獲生產期為 4 月至 10 月 (5 到 7 月為最高峰),雨傘旗魚為表層大目流刺網、延繩釣、鏢旗魚及定置 網的捕獲對象 (Chiang, 2004)。
由標識放流之研究結果顯示,雖然雨傘旗魚在不同海域具有週年洄游或 偏好特定海域習性 (Ortiz et al., 2003),但其族群移動與洄游路線均尚未有確 切之研究結果。雨傘旗魚具獨游性,但當餌料魚群出現時,則會結群圍捕,
集群數量可達3 ~ 30 尾以上 (Voss, 1953)。
雨傘旗魚的攝食對象不具選擇性 (Nakamura, 1949),以大洋性表層魚類 及頭足類為主 (Maksimov, 1971; Eldridge and Wares, 1974),但游至近岸時亦 會 攝 食 岩 礁 區 或 沙 岸 之 底 棲 性 魚 類 、 甲 殼 類 及 頭 足 類 (Voss, 1953;
Rosas-Alayola et al., 2002)。
五、生物學
(一) 雌雄性比
臺灣海域所漁獲的五種旗魚類其雌雄別性比組成皆會隨著成長而有所 改變,於幼魚或未成熟時期,其性比組成並無明顯的變動趨勢,但隨體型的 增長雌魚的比例會逐漸增加,在其他洋區的旗魚類同樣也有呈現此種性比變
動的情形。根據王 (2010) 的報告說明,臺灣海域的劍旗魚、黑皮旗魚、雨 傘旗魚及白皮旗魚,於體長 (下顎至尾叉長;lower jaw fork length;LJFL) 成 長至 150 至 200 公分之後,雌魚比例會逐漸高於雄魚,當體長達約 210 至 250 公分後則幾乎皆為雌魚。
(二) 成長
雨傘旗魚雌魚之估計最大體長 (LJFL) 可達約 340 公分,雄魚則約為 290 公分 (Chiang et al., 2004)。最大記錄體重為 100.2 公斤 (IGFA 2001); 最 大記錄年齡為 13 歲 (Prince et al. 1986)。雌性雨傘旗魚的 50%性成熟體長 (LJFL) 約為 166 公分,而雄魚則約為 151 公分 (Chiang et al., 2004, 2006b)。
六、漁業生產與資源概況
(一) 漁業生產
根據臺灣地區漁業統計年報 (Fisheries Statistical Yearbook, Taiwan),歷 年雨傘旗魚漁獲統計資料 (Fig. 3),臺灣沿近海漁業中雨傘旗魚年漁獲量於 1963 年達 5,000 公噸,高居所有旗魚類漁獲量之冠,隨後逐年間漁獲量變 動,最高為1968 年達 5,100 公噸,隨後 1974 年下跌至 954 公噸,1977 年 與 1980 年則高達 4,175 公噸,但 1981 年起漁獲量變動雖時有增減,但呈 逐漸下降趨勢,至 2001 年漁獲量僅有 201 公噸,然後巨幅振盪變動,至 2010 年為 960 公噸。
以縣市別漁獲量而言,在1965 至 2001 歷年雨傘旗魚漁獲量中,1980 年 代之前以屏東縣漁獲最多,年平均漁獲量佔沿近海雨傘旗魚總漁獲量之 52
%;而至 1980 年代之後,則以臺東縣漁獲量居最多。近二十年來,臺東縣 雨傘旗魚漁獲量年平均漁獲量更高達54 %以上 (Chiang et al., 2006a),為臺
灣最重要之雨傘旗魚漁獲地區。由於旗魚可作為生魚片或魚排之高級食材,
也可以作成魚鬆、魚丸等加工製品,因此,對臺灣不管是沿近海漁業或是遠 洋漁業,都是相當重要的高經濟性魚類。
(二) 資源概況
臺灣沿近海域雨傘旗魚資源量評估,根據Chiang (2004) 之研究,沒有 證據顯示有過度開發的現象,且估算之雌雄別漁獲死亡率也遠低於目標參考 點,而在產卵潛能比例 (spawning potential ratio) 也維持在高水準,因此,
該資源應屬於適度開發階段 (Chiang et al., 2009)。
1.2 相關研究
穩定同位素運用於攝食生態的研究,從1990 年代開始,早期以湖泊及 濕地生態為研究主題,然後再擴展到海岸底棲生態系統,在2000 年代中期 才逐漸運用於大洋性的魚種。雖然穩定同位素的應用越來越廣泛,但應用於 大洋性洄游性魚類的研究,仍處於剛起步發展的階段,本節整理相關研究分 別說明如下:
Young et al. (2006) 針對澳洲東部海域的劍旗魚攝食生態進行研究,研 究結果顯示劍旗魚之氮同位素的數值,比胃內容物內餌料生物之氮同位素的 數值為高,劍旗魚的體長與氮同位素呈現對數線性關係。發現小型劍旗魚以 攝食魚類為主,大型劍旗魚則以攝食頭足類為主,同位素分析的結果與胃內 容物一致。
Menard et al. (2007) 針對西印度洋的黃鰭鮪及劍旗魚,運用穩定同位素 進行攝食生態及洄游研究,探討魚體體長及緯度對於碳、氮同位素的效應。
發現兩種魚都有顯著的體長效應,即氮同位素的數值隨著體長而增加,在劍 旗魚方面則更加明顯,反應出其不同的攝食策略及獨特的生理能力。劍旗魚 比黃鰭鮪可到達更深層的水域,可攝食更大範圍的餌料生物。而黃鰭鮪的幼 魚及成魚,大部分在表水層活動,結果劍旗魚的氮同位素特徵比黃鰭鮪高,
提出兩種魚類之間,分別位於不同營養階層的證據。
Graham et al. (2007) 針對在夏威夷 Oahu 島附近人工集魚器 (Fish aggregating devices: FADs) 的小黃鰭鮪 (23.5-154 cm, FL) 以穩定同位素進 行研究,發現黃鰭鮪在體長45 至 50 cm (約 1.5 kg) 明顯的食性轉變,較小
的幼魚以混合層以淺的浮游生物為食,主要是口足類及十足類等甲殼類,較 大的黃鰭鮪則以魚類,以及具有中層水域垂直洄游的蝦類為主食,作者提出 黃鰭鮪到達體長45cm 以上,才有足夠的行動能力可以攝取更深及更冷水層 的餌料生物。也因為黃鰭鮪攝食範圍及食性的轉變,會影響黃鰭鮪被漁業捕 獲的壓力。
Revill et al. (2009) 利用穩定同位素在澳洲東部海域,探討掠食者與餌料 生物的關係,作者將各種掠食者穩定同位素的特徵,運用群集分析,將掠食 者分為頂級掠食者 (top predator)、中階層魚種 (middle trophic level) 及小型 魚種 (small fishes) 等三大類,發現這些掠食者的體長與氮同位素的數值是 呈線性正相關。另外發現長鰭鮪及黃鰭鮪的同位素的數值,有緯度效應的現 象。
Matich et al. (2010) 針對公牛白眼鮫及鼬鯊的不同組織間的穩定同位素 特徵進行研究,發現公牛白眼鮫在不同組織間 (肌肉、鰭、血液) δ13C值,
在小個體時差異大,隨著體型增大而減少,因為小的公牛白眼鮫部分保留母 體的特徵。其原因為組織的週轉率 (turnover rate) 影響同位素的數值,幼魚 的鰭部位留大部分母體的特徵,因為週轉率較低。而血液的週轉率較高,可 反應出小的公牛白眼鮫的食性特徵 (MacNeil et al., 2005)。 在到達體長 110 cm以後,其差異性趨向平衡。另外, 相同的種類,在不同的生活環境,會 導致isotope的差異 (Post, 2002)。
Olson et al. (2010) 利用穩定同位素探討東太平洋黃鰭鮪與橈足類之間 的食物網關係,發現橈足類的氮同位素的數值,由南到北呈現均勻增加的趨 勢,黃鰭鮪的氮同位素的數值也是呈現相同的趨勢。
Young et al. (2010) 根據從 1992 年至 2006 年澳洲東部海域延繩釣作業 豐度前10 大魚種,檢視其攝食生態及生態區位分離 (niche segregation)。這 些魚類有混合攝食頭足類、甲殼類及魚類。除了小長鰭鮪主要攝食甲殼類為 主,大型劍旗魚與長鰭鮪以攝食頭足類為主,其他各掠食性魚種均以攝食魚 類為主。研究結果發現隨著掠食者的體長增加,其餌料生物的體長也增加。
但是大型的掠食者所攝食的餌料生物範圍較廣,也會攝食較小的餌料生物,
作者根據這些研究結果,提出這些大洋掠食性魚類,在攝食生態區位不同的 證據。
Varela et al. (2013) 利用胃內容物分析及穩定同位素分析,針對大西洋黑 鮪幼魚估算攝食組成,由穩定同位素分析結果,顯示具有年間的變動,大西 洋黑鮪幼魚攝食中營養階層的餌料;然而以胃內容物分析結果,則發現攝食 低營養階層的餌料為主。作者建議結合胃內容物分析及穩定同位素分析,比 單獨採用胃內容物分析更合適。
Logan et al. (2013) 估算在中北大西洋頭足類餌料對表水層大型掠食者 (鮪、旗、鯊魚等) 的貢獻度,結果顯示這些大型掠食者之攝食組成隨空間改 變。劍旗魚及黃鰭鮪的餌料尺寸大小與掠食者尺寸大小相關。並由這些掠食 者的攝食組成,估算出頭足類對於這些表水層大型掠食者的重要性。
1.3 研究目標與論文概要 1.3.1 研究目標
雨傘旗魚是海洋表水層食物網中的頂級掠食者之一 (Kitchell et al., 2006),它在其生態系統中扮演重要的角色。這些上層掠食者的數量減少將 可能對海洋表水層 (epipelagic) 生態系統造成顯著的影響 (Myers and Worm, 2003),導致低營養階層物種的連鎖效應。
雨傘旗魚為印度-太平洋,以及臺灣沿近海重要的經濟性洄游魚類之 一。有關的生物學及資源評估已經有初步的研究成果,其洄游生態也正密集 進行研究中,但是有關食物網結構,營養動力學的相關研究卻闕如。食物連 結是海洋生態系統結構與功能的基本表達形式,質量通過食物鏈及食物網,
轉化爲各營養階層生物之生産力,形成生態系統生物資源量,並對生態系統 的服務和産出,及其動態産生影響。因此,研究魚類食性和食物網的研究,
對於瞭解魚類群落的結構與功能具有重要意義,是建立漁業管理重要的資訊 之一。
本研究以雨傘旗魚的食物階層結構著手開始,先探討雨傘旗魚在種內體 型尺寸之差異性,接著探討雨傘旗魚的攝食組成,及與生態系中相關物種的 關係,經由在海洋生態中雨傘旗魚與餌料生物之攝食關係,逐步建立食物網 結構,探討雨傘旗魚在臺灣東部海域生態系中所扮演的角色、最後根據食物 網結構與能量的關係,嘗試探討建立以生態系為基礎之生態系模型,以期未 來可利用於本資源之適正利用,進而維持生態系的健全發展及資源永續利 用。因此,本論文之研究目標為:
一、以碳、氮穩定同位素分析,探討雨傘旗魚不同體長個體同位素數值之變 化情形,建構雨傘旗魚的營養階層結構,以究明雨傘旗魚在生態系中所 扮演的角色。
二、利用胃內容物分析及穩定同位素分析,探討雨傘旗魚之攝食組成及生 態,建立雨傘旗魚餌料生物的攝食比例與營養階層結構的關係。
三、利用已完成之攝食生態研究、漁獲統計資料及相關生態系模式文獻,建 置雨傘旗魚及其相關物種的生態系模型,推算雨傘旗魚在生態系的能量 流關係,以探討雨傘旗魚生態系之功能與結構,提供未來建置以生態系 為基礎的漁業管理相關科學的資訊。
1.3.2 論文概要
本論文分為五個章節,第一章為歸納整理介紹雨傘旗魚的生物學及生態 概況,穩定同位素在海洋生態的應用,應用穩定同位素建立營養階層與食物 網的關係,逐步建立以生態系為基礎的漁業管理模式,以及本論文之研究目 標。
第二章為以碳、氮穩定同位素分析探討雨傘旗魚的營養階層結構 (Trophic structure),究明雨傘旗魚在生態系中所扮演的角色,不同體長大小 的雨傘旗魚,分別位於不同的營養階層 (Trophic level, TL),並比較分析臺灣 東部與南中國海雨傘旗魚樣本的碳、氮同位素之差異。
第三章為探討雨傘旗魚與其餌料生物之間的攝食營養關係,分別利用胃 內容物分析及碳、氮穩定同位素分析,經由碳、氮穩定同位素質量守衡定律 之同位素混合模式 (Stable isotopes mixing model),來推算雨傘旗魚餌料生物 來源比例;並依據穩定同位素的測量數值,建立營養階層與結構關係。
第四章為利用雨傘旗魚與其餌料生物食物網的關係,參考漁獲統計資 料,以Ecopath with Ecosim (Pauly et al., 2000; Christensen and Walters, 2004) 套裝軟體嘗試建立雨傘旗魚及其相關物種的生態系模式,推算雨傘旗魚生態 系的能量流關係,以探討雨傘旗魚於大洋生態系之功能與結構。本研究初步 建立臺灣東部海域生態系統模型,探討生態系的結構與功能,未來可提供建 置以生態系為基礎的漁業管理相關科學的資訊,以及未來探討臺灣東部海域 漁業資源與環境氣候變遷的關係。
第五章為根據第二章至第四章的研究結果,綜合整體歸納出本篇論文的 結論與摘要,以及提出未來研究的需求與建議,以期本研究的結果,可應用 於雨傘旗魚資源之永續利用,並維護海洋生態系的健全功能。
第二章 雨傘旗魚營養階層結構
2.1 前言
自 20 世紀中期以來,商業性漁業已從太平洋捕撈超過 5,000 萬噸的鮪 魚 (Sibert et al., 2006) ,因而產生漁業改變海洋生態系統的爭論 (e.g. Pauly et al., 1998; Springer et al., 2003; Sibert et al., 2006)。選擇性捕撈去除的食物網 中的大型掠食性動物,將會經由食物階層連鎖而改變食物網結構 (Carpenter et al.,1985; Estes et al., 1998; Pace et al., 1999; Pauly et al., 2001; Springer et al., 2003; Myers et al., 2007)。
由於海洋食物網各物種之間的交互影響錯綜複雜,這使得海洋生態系食 物網研究十分困難,近期研究人員關注於頂級掠食者種群他們的食性和遷移 模式。在食物網研究中,營養物質和能量流動的關係是海洋生態系統動力學 研究的主要問題 (Braga et al., 2012)。穩定同位素方法被有效地用於分析生 態系統的能量流和生物體之間的營養關係。胃內容物方法只提供最近期的覓 食活動的瞬間快照。相較之下,海洋生物體的組織的穩定同位素分析 (Stable Isotope Analysis, SIA) 可以被用來作為一種替代和補充的工具,提供結合時 空及吸收同化的飲訊息 (Fry, 1998; Harrigan et al., 1989; Fantle et al., 1999)。
傳統研究生物食性的方法是消化道內含物法,經由生物在被捕前所攝取 的食物,即消化道內未被消化的食物,來確定生物的食性,其優點是可以直 接觀察測量,而缺點是測量的只是被捕前所攝取食物,且許多水域生物存在 偶食性,無法代表生物長期的食性。而穩定同位素法是根據消費者穩定同位 素比值與其食物相應同位素比值相近的原則,來判斷此生物的食物來源,進 而確定食物貢獻比例,所取樣品是生物體的一部分,即可以反映生物長期生
命活動的結果 (Varela et al., 2013)。在特定的生態系統中, 各種生物種群之間 的攝食關係、營養物質和能量流動是生態學研究的一個重要課題。利用研究 生態系統中有機碳和氮之中的穩定同位素組成動態變化的技術,為解決此一 生態學難題提供了新的研究方法。
穩定同位素是元素自核合成以後,即保持穩定的同位素,是電子數和質 子數相同,但中子數不同的元素,穩定性同位素指是原本就存在於自然界中 的同位素,其性質穩定不易衰變,可以作為標記工具 (Peterson and Fry,
1987)。一般常使用的穩定性同位素以碳同位素 13(13C)、氮同位素 15(15N)、
硫同位素34(34S)為主,原因為這些同位素是地球上主要元素,生物體內含量 多,較容易進行分析。
Fry (1998) 研究發現,動物體內的穩定同位素組成和其所攝食的食物中 的穩定同位素組成相對應。當食物中的碳、氮穩定同位素組成發生變化時,
動物體內的碳、氮穩定同位素組成也發生相應的變化。
因此,通過對動物體內不同組織和可能攝取食物的碳、氮穩定同位素值 的分析比較,就可推斷動物的攝食情況 (DeNiro and Epstein,1978; Balasse, 2002)。由於動物在攝食後對食物的同化作用要經過一段時間,所以動物體 內的穩定同位素值的變化主要反映動物當前一段時期甚至相當長一段時期 內的攝食情況,這主要由動物組織的穩定同位素數值決定 (Cerling et al., 1999)。
胃內含物分析所顯示的為魚類一天的攝食食物組成,而穩定同位素分析 則顯示魚類在攝食棲地長時間累積的食性變化 (DeNiro and Epstein, 1978, 1981; Fry and Sherr, 1984; Peterson et al., 1985);結合胃內含物和穩定同位素
分析可以精確的獲得魚類食性的資料 (Harvey et al., 2002)。生物的穩定同位 素比例值,可以反應本身吸收利用有機物的成分,而呈現出食性長時間的平
均變化 (Hobson, 1999)。很多食性關係的研究,可進一步藉由以穩定同位素
含量的變化,來提供食物來源資料並分析營養階層的關係 (Peterson et al., 1985; Peterson and Fry, 1987)。
目前常使用的穩定同位素,包含碳同位素和氮同位素。穩定性同位素使 用在食物網及營養階層研究應用時,δ13C 和δ34S大致上消費者與餌料的值一 致,δ15N 則隨營養階層增加而增加。氮同位素的分析,為利用動物體內的15N 會隨著消費者營養階層的增加而逐步增加 (Minagawa and Wada, 1984);碳同 位素的分析,則是因為消費者的碳同位素數值,在不同的碳來源之間有顯著 的差異性。一般而言,氮同位素(δ15N) 數值在消費者和食物來源有平均 3.4 (±1.1)‰的關係,而在碳同位素(δ13C) 數值有平均 1~2‰的關係 (DeNiro and Epstein, 1978, 1981; Fry and Sherr, 1984; Minagawa and Wada, 1984)。
過 去 穩 定 性 同 位 素 分 析 主 要 是 利 用 在 一 些 食 物 鏈 較 單 純 的 碎 屑 (detritus) 生態系,或是一些不易利用消化道內含物分析建立的生態系,如溼 地、紅樹林、海藻床、河口等 (Bustamante and Branch, 1996; Kwak and Zedler, 1997; Hayase et al., 1999; Tatrai et al., 1999)。而近幾年研究生態系食物網也大 量使用穩定性同位素分析,來確定生物的營養階層與能量的流向 (Gu et al., 1996a; Jarman et al., 1996; Pinnegar and Polunin, 2000; Weinstein et al., 2000)。
應用碳氮穩定同位素技術,來研究魚類的攝食習性及營養動態,該方法 能穩定、準確地研究低營養層次魚類的食物來源、定量分析所攝食餌料的貢 獻比例等問題,簡化了食物網中捕食者和被捕食者之間的複雜關係,進而確 定生態系統中不同種類間的營養關係,是推估生態系統能量流動的理想工具
之一 (Overman and Parrish, 2001),也能為瞭解海洋生態中各營養階層魚類的 物質循環和能量流動過程,以及為其動態變化研究提供基礎資料。
雨傘旗魚 (Istiophorus platypterus) 是海洋生態系中的頂級掠食者之一 (Kitchellet al., 2006)。在其生態系統中扮演著重要的角色。這些大型掠食者 族群數量的減少,可能對表水層生態系統中的高營養階層物種有顯著的的影 響 (Myers and Worm, 2003) 並導致對低營養階層物種的連鎖效應。臺灣東部 海域雨傘旗魚的產季為每年四月到十月(最高峰在五月至七月),對當地漁 業具有經濟的重要性,雨傘旗魚是主要由沿近海大目流刺網、延繩釣、定置 網、以及鏢旗魚漁法捕獲 (Chiang et al., 2006a)。
儘管雨傘旗魚的資源狀態在臺灣東部海域為適度開發,並有相對低風險 的過度捕撈 (Chiang et al., 2009);根據國際自然保育聯盟 (IUCN) 的紅色名 錄標準,雨傘旗魚族群資源狀態列為 “暫無危機 (least concern, LC)” (Colltte et al., 2011)。為了長期監測資源的狀態和維護生態系統的健康,確立和量化 評估針對的頂級掠食者的營養關係,以及漁業活動對於的表水層生態食物網 結構和功能的影響。亟需迫切建立以生態系統為基礎的漁業管理 (Graham, 2008),來監測這個重要經濟價值的物種。
然而,雨傘旗魚每一個生命階段的營養動態的訊息是不完整的,在其生 命週期的不同階段的旗魚的營養階層也沒有相關的數據。在這項研究中,我 們運用肌肉組織進行了穩定同位素13C和15N分析調查的旗魚的營養階層。這 項研究的主要目的是:(1) 估算為不同大小之旗魚的的營養階層;(2) 以穩 定同位素為基礎建立有關臺灣東部海域旗魚個生活史階段的生態背景基礎 資訊。
2.2 材料與方法 2.2.1 研究地點
本研究所採集雨傘旗魚為來自於臺灣東部海域,在臺灣東部海域週年均 受到黑潮洋流影響,如Fig 4所示,顯示黑潮為北太平洋西部強勁的溫暖洋 流 (海面平均溫度為 24°C),由赤道向北半球流經菲律賓東岸,向北沿臺灣 東岸至日本沖繩海域 (Nitani, 1972),流域約 100 公里寬度,流動過程出現許 多的小尺度渦流,在流經沿海水深100 公尺之海域,因海底地形抬升產生湧 昇流,於沿岸海域形成為具有高度生產力區域 (Makoto, 1989)。
黑潮在到達臺灣東北部陸緣後,主流轉往東北流向日本,支流則流入到 中國東海,與東海水團形成混合區和湧升流 (Fan, 1980; Liu, 1983; Liu and Pai, 1987; Chern and Wang, 1989; Wong et al., 1991; Liu et al., 1992),並且為臺 灣東部海域帶來了豐富的海洋生物資源。許多洄游物種,如:海豚,鰹魚,
旗魚,鮪魚和鯊魚,每年從春到夏季,隨著黑潮洋流進行產卵、攝食和其他 活動。由於黑潮對於西北太平洋的物理和化學的影響重要性,使得附近海域 的漁業生產力較高。在湧升流的高生產率和邊緣混合區的黑潮通常是良好的 漁場,因此,漁業是臺灣東部地區最重要的經濟活動之一。
2.2.2 樣本收集
雨傘旗魚樣本於2009 年 4 月至 2012 年 2 月,在臺灣東部海域 (Fig. 5) 採 集233 個樣本 (雄魚 127 尾與雌魚 106 尾),體長 (LJFL) 111 - 245cm (Fig.
6),體重 4 - 64 公斤,並採集於南中國海捕獲的 30 個樣本 (Fig. 5),進行採 樣地點比較。
下顎尾叉長 (lower jaw fork length, LJFL) 為旗魚類較適合的體長測量 方式,因為旗魚身體細長和側扁,與鮪魚類相比較時,可降低因嘴長度的差 異。根據雨傘旗魚及鮪魚攝食研究 (Arizmendi-Rodríguez et al., 2006; Menard et al., 2006),體長級別以 20cm 為組距,因此,本研究設定體長組距範圍為:
<140, 141~160, 161~180, 181~200, 201~220 與 >221 cm (LJFL) (Table 1)。
至新港魚市場採集回來的雨傘旗魚肌肉樣本,採集尾柄部位白色肌肉,
並採集雨傘旗魚胃內容樣本,帶回實驗室進行分析處理,按同位素質譜儀測 定要求,對樣品進行處理和測試。採集的魚類樣品在- 20℃冰櫃中保存,然 後魚類樣本在實驗室解凍、蒸餾水洗滌後,經實驗室進行分類鑑定及對其胃 含物做分析鑑定後,選擇取樣。分析的樣本取自魚體白色肌肉的部分,因為 白色肌肉中的δ13C 和δ15N 的變異程度較其他組織小 (Pinnegar and Polunin, 1999)。
接著用10% HCl 清洗樣本,排除樣本體表碳水化合物的干擾,再用蒸 餾水洗滌至中性。然後再經由冷凍乾燥處理,冷凍乾燥完成後,再分別取肌 肉樣本約 1 毫克,將樣本裝在錫製的膠囊中,並將膠囊壓實成錫球,其間 不留空氣,錫球即可直接利用同位素分析儀進行分析測定。進行同位素分 析,樣本處理流程如Fig. 7 所示。
2.2.3 穩定同位素分析
穩定同位素分析是在中央研究院環境變遷研究中心進行,儀器首先將錫 球在1000~1050℃下燃燒,燃燒產物經載氣 (氦氣) 通過Cr2O3和活性Cu柱淨 化。這樣,生物樣品中的碳和氮可完全轉化為N2和CO2氣體,再經由質譜儀 測定同位素的組成,分析穩定性同位素的方法參考Pinnegar and Polunin
(2000)、Davenport and Bax (2002)。
為使全世界不同實驗室的測定值能互相比較,因此,穩定同位素分析都 必須使用相同的參考比較標準,相關同位素分析所使用的參考標準乃參閱 Clark and Fritz (2000)。進行譜儀分析時,有關測量的國際標準品比對,δ13C 的國際標準品為Pee Dee Belemnite (PDB),δ15N 則以大氣作標準。穩定性同 位素的比例以千分比為單位,計算公式如下:
3 standard
sample
10 1⎟⎟×
⎠
⎜⎜ ⎞
⎝
⎛ −
= R X R
δ (‰)
其中X 為13C 或15N,R是15N/14N 或13C/12C。
營養階層的估算,根據下列公式:
Trophic level = (δ15Nconsumer – δ15Nbaseline)/TEF + 2
TEF (trophic enrichment factor) 為同位素的富集度,一般根據 Minagawa and Wada (1984) 每一階層TL 約為 3.4‰,本研究根據Vanderklift and Ponsard (2003) 參閱δ15N富集度 (TEF) 之相關文獻 (Caut et al., 2009),採用海水魚 類富集度 (TEF) 2.4 ‰ 為估算值。δ15Nconsumer為消費者之氮同位素數值,
δ15Nbaseline 為營養階層 (TL) 的計算基線,如以基礎生產者為計算基線,其
營養階層 (TL) 為 1。如以基礎消費者為計算基線,則其營養階層 (TL) 為 2,依序來估算接續之消費者的營養階層。本研究採用臺灣東部海域草食性 貝類黑鐘螺 Chlorostoma argyrostoma argyrostoma (Gmelin, 1791) 之δ15N 測 量值 6.24 ‰ ± 0.29 SD (n=5),做為計算營養階層的基線 (TL=2)。
2.2.4 資料分析
δ15N and δ13C 與體長之間的關係,分別採用線性回歸分析 (n=263)。利 用多變方分析 (MANOVA) 來檢定雨傘旗魚肌肉之碳氮同位素,在性別、
體長級別、採樣地點、季節之間是否有差異。當樣本有顯著差異時,以 Tukey’s HSD test (Tukey’s Honestly Significant Difference Test) 來進行事後 檢定分析。季節分析則分為春季(2-4 月)、夏(季 5-7 月)、秋季(8-11 月)及冬 季(11-1 月) 四個季節,統計分析採用R套裝軟體進行分析 (R Development Core Team, 2008)。
2.3 結果
2.3.1 體長與同位素之關係
雨傘旗魚肌肉的δ15N測量值介於 7.51 至 14.19 ‰ (平均=12.05±1.23
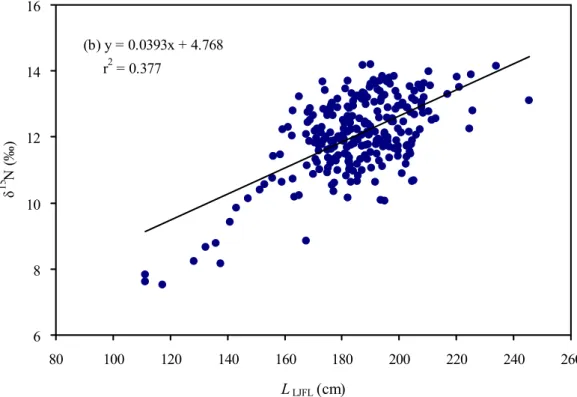
‰);δ13C測量值介於-22.04~15.48 ‰ (平均為-17.68±1.15 ‰) (如Table 1 所 示)。δ15N的數值變化範圍比δ13C較寬廣 (6.68 比 6.56)。雨傘旗魚的肌肉δ13C 和δ15N數值與體長 (LJFL) 之關係。分別如Fig. 8、Fig. 9 所示。
體長和同位素之間的關係以線性迴歸來分析,同位素δ13C與體長呈負相 關:
δ13C =-0.0153 (±0.004)×LJFL-14.777 (± 0.720) [n=263; r=0.242; P <
0.001)],
然而同位素δ15N與體長呈正相關:
δ15N = 0.0393 (±0.003)×LJFL +4.768 (± 0.638) [n=263; r=0.597; P <
0.001)].
在雌雄之間同位素δ13C及δ15N均沒有顯著差異,因此,雌雄樣本合併處 理。本研究則發現不同體長大小的雨傘旗魚,同位素δ15N有顯著差異 (Table 1)。
2.3.2 體長級別與同位素之關係
在本研究中所採集的雨傘旗魚以體長 20 公分為組距,分為 6 個體長級 別,分別從從幼魚 (第 I 級,年齡不超過 2 歲),大型成魚 (第 VI 級,年齡 超過11 歲),如 Table 1 所示,(Chiang et al., 2004)。
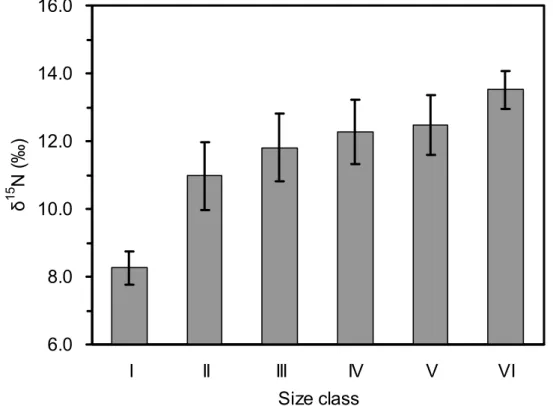
雨傘旗魚每一個體長級別的碳和氮的穩定同位素分別計算,根據掠食者 體型大小來分辨其營養階層之差異。從I級至第VI級,每一個體長級別之間 的δ13C是顯著不同 (ANOVA: F5, 257 = 4.68, P<0.001);每一個體長級別之間的 δ15N也是顯著不同 (ANOVA: F5, 257 = 40.48, P<0.001)。
旗魚的白色肌肉組織的碳同位素值,在各體長級別之間,碳同位素值 δ13C值與體長之間沒有顯著的趨勢 (Fig. 10)。但是雨傘旗魚其氮同位素值 δ15N值與體長級別之間,具有明顯增加的趨勢 (Fig. 11),在各體長級別之間 有顯著差異,顯示年幼的雨傘旗魚氮同位素值為低 (~8.11‰),在大型雨傘 旗魚,氮同位素值為高,具有更豐富的氮同位素 (~14.15‰)。
經 由Tukey HSD test , 碳 同 位 素 (δ13C) 在 體 長 級 別 分 析 為 : I=II=III>IV=V=VI,第I級、II級和III級標本彼此之間相似,在IV,V,VI級 的標本之間也是相似。但是在III和IV級標本中的碳同位素則有顯著差異。氮 同位素 (δ15N) 在體長級別分析為:I<II<III<IV=V=VI,即在體長級別I、II、
III和IV級之間也有顯著差異。在本研究中發現雨傘旗魚其碳及氮同位素值,
在體長 (LJFL) 141〜160 cm 與體長 161〜180 cm之間,有顯著差異。
2.3.3 採樣區域之差異性
本研究利用 MANOVA檢測臺灣東部海域 (n=233) 及南中國海 (n=30) 等兩地的樣本。檢測結果顯示,在臺灣東部海域和南中國海之間的樣本之碳 同位素及氮同位素,沒有顯著差異 (MANOVA, δ13C: F1,256=1.078, P>0.05;
δ15N: F1,256=0.022, P >0.05)。
2.3.4 季節差異
本研究利用 MANOVA 檢測樣品之碳及氮同位素數值與季節性因素的 影響,在臺灣東部海域。我們選取春季為三月至五月,夏季從六月到八月,
秋季九月至十一月,冬季十二月到二月。結果顯示,碳同位素及氮同位素在 季節之間,有顯著性差異 (MANOVA, δ13C: F3,256=12.560, P <0.001; δ15N:
F3,256=5.592, P <0.001)。δ13C在秋季及冬季數值比春季及夏季高,δ15N則顯示
從春季到夏季呈現遞減的趨勢 (Fig. 12)。
2.3.5 營養階層估算
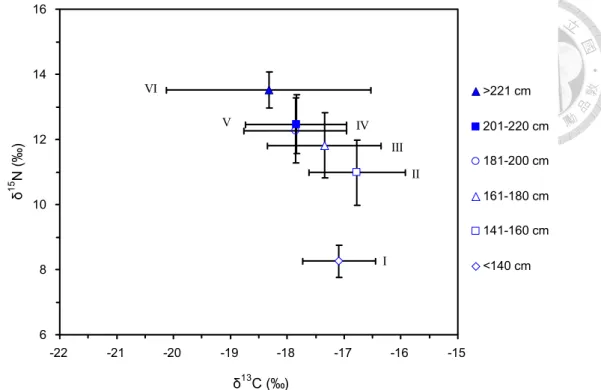
每一個體長組距的雨傘旗魚,其營養階層的估算結果整理如 Table 2。
雨傘旗魚的營養階層隨著體長級別而增加,不同的體長級別,具有不同的營 養階層 (Fig. 13)。體長級別與營養階層之關係如Table 2所示,從第I 級 (年 齡約為2 齡的雨傘旗魚) 其營養階層為 2.84±0.08 (不計算仔稚魚的階段)。在 第VI 級(年齡超過 11 歲的雨傘旗魚) 其營養階層達到 5.03±0.09,顯示大型 高齡的雨傘旗魚,其營養階層達到最高階。所有雨傘旗魚樣本的平均營養階 層為4.42±0.20。體長與營養階層之關係如Fig. 14,迴歸關係為:
TL=0.0193(±0.003)×LJFL+0.762(±0.583) [r=0.95; P < 0.001)]
2.4 討論
2.4.1 體長與氮同位素
本研究結果顯示有關δ15N和δ13C數值與雨傘旗魚體長之間的關係,同位 素δ15N數值對雨傘旗魚體長 (LJFL) 的影響 (Fig. 9) 比δ13C數值更明顯 (Fig.
9) 。
許多物種的食性會隨著整個生命週期而改變,特別是當他們的體型日益 變大,有利於適應獵捕其他大型餌料生物 (Graham et al., 2007)。在一些研究 中,穩定碳和氮同位素的比值,根據物種其生命週期,與在生態系統中利用 餌料食品的變化性,可做為一個物種的食性結構改變的追踪劑 (Loch and
Hily, 2005)。在種內和種間,一般在大型魚比小型魚有較高的δ15N訊息。大
型魚提供更多機會攝食大型的餌料生物,而且選擇更多樣的餌料生物的種類 (Davenport and Bax, 2002)。魚體內具有較高δ15N訊息,一般在他們的攝食或 吃其他物種的具有較高比例的δ15N信號 (Bulman et al., 2001)。
我們的研究結果顯示,在雨傘旗魚白色的肌肉組織中δ15N測量值與體長 間距的關係,在體長 141〜160 cm與 161〜180 之間有顯著的差異。這種模 式顯示了積累相當的15N,這可能解釋為: (a) 魚體的生理變化或 (b) 同位 素 來 源 的 改 變 或 是 在 個 體 發 育 期 間 攝 食 餌 料 同 位 素 經 由 組 織 轉 換 率 (turnover rate) 而變化 (Hesslein et al., 1993; Overman and Parrish, 2001)。在雨 傘旗魚隨著體型增加,體內明顯的保留較重的氮同位素(15N),可能是由於攝 食習性和棲息地的變化,而不是生理上的變化 (Cocheret de la Moriniere et al., 2003; Graham et al., 2007) 。
在 魚 類 方 面 , 體 長 效 應 對 於 餌 料 資 源 的 利 用 已 經 有 相 關 的 研 究 (Pinnegar and Polunin, 2000; Renones et al., 2002; Bode et al., 2004),體長效應 與攝食棲地的改變有關 (Werner and Gilliam, 1984)。Ménard et al. (2007) 發 現在西印度洋黃鰭鮪在尾叉長 (FL) 範圍 39-164 cm,δ15N值變異低。然而,
Sarà and Sarà (2007) 發現在大西洋黑鮪 (Thunnus thynnus) 隨著年齡的增 長,δ15N值越來越高,在 13 歲的成魚,重量在 225 公斤達到最高值。因為 大西洋黑鮪其活存的時間更長,體型比黃鰭鮪的最大尺寸較大。Graham et al.
(2007) 發現在夏威夷的Oahu島的黃鰭鮪,根據δ15N值和胃內容物,體長約於
40cm,存在營養發育過程的轉變。但是一般在黃鰭鮪營養發育過程沒有明 顯的轉變特徵 (Olson and Boggs, 1986; Buckley and Miller, 1994; Ménard et al., 2006),除非是非常小的黃鰭鮪個體 (Maldeniya, 1996)。
在臺灣東部地區(西北太平洋),胃內容物分析顯示,雨傘旗魚以的大 洋表層性魚類為主食 (特別是花鰹類,Auxis spp.) (Chiang et al., unpublished data)。在本研究中,我們發現的雨傘旗魚的攝食生態與鮪魚和劍旗魚類似,
是機會主義捕食者,能夠利用各種不同種類的餌料生物資源 (Chase, 2002)。
隨著他們成長,其攝食的習性也發生改變。理論證實,生物體的營養階層,
經由攝食,可以根據不同的個體發育,而導致生物體在食物網內的位置的轉 變 (Werner and Gilliam, 1984) 。
2.4.2 體長與碳同位素
在各體長間距之間,其平均δ13C數值的差異性不大 (Fig. 10),而δ15N與 δ13C相比,δ15N數值隨著營養階層而增加,較δ13C數值顯著 (DeNiro and Epstein, 1981)。
由雨傘旗魚的δ13C範圍的高變異性 (Fig. 10),顯示在西北太平洋這些開 放的海洋生態系統的雨傘旗魚,未來可能用來釐清棲息攝食的區域地點或追 踪魚群的洄游運動。δ13C可用來解釋初級生產者的原始來源,這些同位素,
也可以幫助研究分析在攝食習性的變化,無論是短期,長期或永久性的。這 些變化甚至可能與季節的變化,反映的浮游植物豐度有關 (Michener and Kaufman, 2007)。在肌肉組織隨著經由一段時間出現攝食習性的差異,所顯 現出不同的同位素特徵是在特定區域停留一段時間的證據 (Graham et al., 2007)。
在雨傘旗魚體長小於160 cm (LJFL) 的樣本,顯示δ13C數值的變異較小 (Fig. 10),我們可以推論,這些個體在海洋表水層生態系統,它們居住在同 一水域的棲息地,並尚未進行大範圍長距離的洄游。雨傘旗魚體長超過160 cm (LJFL)的樣本,顯示δ13C數值的變異較大,推論這些體型較大的雨傘旗 魚,分別從不同的棲息地洄游在臺灣東部海域。
Hobson (1999) 提出一個新論點:“你是什麼地方洄游來的”,針對傳統 著名的穩定同位素生態學論點“你是你吃什麼食物所組成的”(DeNiro and Epstein, 1976),進行補充說明。因此,在動物組織內之穩定同位素比率,可 應用來分辨魚類在不同區域洄游和調查它們的攝食生態 (Menard et al., 2007;
Graham et al., 2009)。
本研究分析結果發現,雨傘旗魚同位素數值有季節性差異,推測主要原 因為攝食魚種來源的差異。在春、夏季節時,臺灣東部海域黑潮洋流較強,
優勢魚種為:飛魚、花鰹類、鬼頭刀等魚種。而在秋冬季節時,臺灣東部海 域黑潮洋流較弱,優勢魚種為:白帶魚、紅目鰱、鯖魚等。因雨傘旗魚餌料 食物來源之優勢魚種的季節變化,而造成雨傘旗魚同位素數值產生季節變
化。另一個季節性產生差異的原因,為雨傘旗魚季節洄游所產生的差異,這 一部分因為雨傘旗魚週年及季節洄游資訊尚不完備,有待後續標識放流及相 關研究來進行補充說明。
2.4.3 營養階層結構
雨傘旗魚顯示有寬廣的營養階層,從 class I (2 歲幼魚) 估算約為TL 2.84 到 class VI (11 歲的成魚) 約為TL 5.03 的營養階層,以黑鐘螺 (Chlorostoma argyrostoma argyrostoma Gmelin, 1791) 肌肉δ15N值 6.24‰,為 TL=2 估算基準 (本研究),如Table 2 所示。 Class V和VI之營養階層分別為 4.59 和 5.03,這些大型雨傘旗魚,他們位於大洋生態系食物網的頂部。在這 本研究中估算全體雨傘旗魚的平均TL是 4.42±0.20,與Fishbase資料庫以食性 組成分析估算雨傘旗魚的營養階層 TL 4.5±0.8 相似 (Froese and Pauly, 2014)。營養階層隨著體型成長而增加,通常是由於在體內之重同位素的累 積增加 (Lindsay et al., 1998; Jennings et al., 2002)。本研究也支持這個論點,
雨傘旗魚隨著體型的增大,營養階層及δ15N也隨之增加。
2.4.4 雨傘旗魚與其他大洋掠食者比較
我們比較雨傘旗魚和其他地區大洋生態系統的頂級掠食者,其δ15N(‰)
和δ13C(‰)的同位素數值的差異,整理如Table 3 所示。在大洋生態系中之 大部分頂級的掠食動物均具有寬廣的δ15N數值的變化。在本研究中的雨傘旗 魚為 7.5〜14.2‰;在澳大利亞西部海域的劍旗魚為 9.1〜15.7‰,紅肉旗魚 為 9.2〜15.2‰ (Revill et al., 2009);在地中海的大西洋黑鮪為 7.2〜13.1‰
(Sara and Sara, 2007);在西印度洋的黃鰭鮪為 10.2〜15.2‰,劍旗魚為 11.8
〜16.2‰ (Menard et al., 2007)。
在許多魚類之氮穩定同位素δ15N數值已被證明,隨著體型尺寸大小和年 齡的變化,營養階層跟著改變 (e.g. Lindsay et al., 1998; Vander Zanden et al., 2000; Overman and Parrish, 2001; Jennings et al., 2002)。根據不同尺寸魚類之 氮同位素富集的研究顯示,魚類在群聚中之營養階層大部分取決於生物體尺 寸的大小 (Jennings et al. 2001, 2002)。因此,觀測到從體型小到體型大的魚 體,氮同位素數值有很大的差異,顯示其攝食餌料生物來源之營養階層狀態 的差異。
在本研究中發現體型較大的雨傘旗魚的δ13C數值 (-22.0 ~ -15.5‰),與在 澳大利亞西部海域的劍旗魚 (-23.9 ~ -17.1‰) 及紅肉旗魚 (-23.4 ~ -14.3‰) (Revill et al., 2009) 相比,他們也同樣有範圍較寬廣的δ13C數值的變化幅度。
但是,在地中海的大西洋黑鮪 (-18.9 ~ -17.2‰) (Sara and Sara, 2007);在印 度洋西部的黃鰭鮪 (-17.4 ~ -15.2‰) 和劍旗魚 (-17.4〜15.0‰),(Menard et
al., 2007) 他們表現出範圍差距較小的δ13C數值的變化。
動物個體之間的δ13C數值有很大的差異,顯示他們具有不同的食物來源 或其食物網的不同基礎生產者。因此,這些頂級掠食者為機會主義捕食者和 高度洄游魚類種群,使他們的δ13C數值有很大的變異。
穩定同位素分析常用於研究在海洋生態系統中的營養傳遞的關係 (Peterson and Fry, 1987; Kwak and Zedler, 1997; Fantle et al., 1999; Peterson, 1999)。本研究結果顯示,可運用穩定同位素分析來估算雨傘旗魚的營養階 層。隨著漁業在全世界捕獲許多大洋性魚類族群,穩定同位素分析提供研究 魚類其環境生態學的重要工具。本研究建議未來增加雨傘旗魚食物網中之低 階和中階營養階層的餌料生物族群的研究,著種於穩定同位素分析和直接採
樣。
為了瞭解生態系的轉變,以及頂級掠食者的種群動態,受到人類利用和 氣候變化的影響,長期監測生態系中物種營養關係的變化是有必要的。本項 研究的結果可提供建構生態系統模式的參數,並可能提供未來進一步運用穩 定同位素來分析雨傘旗魚棲息地與洄游的時空分析之基礎資料。
第三章 雨傘旗魚之食性組成及攝食生態
3.1 前言
在自然界中各物種之間複雜的交互作用,可利用食物網做為一個架構,
以協助生態學家觀察在自然界中各物種之間的網路聯結 (Vander Zanden and Rasmussen, 1996)。 海洋食物網是建構海洋生態系統發展的基礎,也是海洋 生態學基礎理論研究的主要內容之一。海洋食物網研究為經由對海洋魚類及 其食物關係的分析,瞭解其物種間之關係,闡明各營養階層的能流途徑,可 為海洋生態系統復育、改善海洋生態環境、保護海洋生物多樣性,並進一步 合理開發利用海洋生物資源提供科學依據。
Elton (1927) 定義了營養金字塔 (The Eltonian Pyramid) 並提出營養階 層的概念,反映動物在食物迴圈中的位置;Linderman (1942) 根據湖泊水域 攝食之研究發展了有關於營養階層 (Trophic level, TL) 的定量計算,Odum et al. (1975) 提出用小數形式更能確切表達攝食關係,並確定初級生產者的營 養級為1,Pauly et al. (1998) 根據全球漁獲物平均營養階層的分析,提出捕 撈導致海洋食物網平均營養級下降 (fishing down marine food web)。
以營養階層為線索進而研究建構海洋食物網,將有助於釐清各種繁雜的 海洋動物,在食物循環的營養關係及所處位置,而以漁獲物平均營養階層及 各重要群落之營養階層可用於評估海洋食物網的健康狀態(Pauly et al., 1998),進而評估分析可應用於人類活動及全球環境變遷變化對海洋生態系 統的影響。目前,以氮同位素技術應用於海洋魚類營養階層及漁獲物平均營 養階層方面的研究已經日益普及,並逐漸得到推廣應用 (Post, 2002)。
雨傘旗魚為隨機的掠食者 (Rosas-Alayola et al., 2002),其餌物生物的豐 度和分佈也會改變他們的關鍵覓食棲息地點。任何頂級掠食者資源量的減少 都可能改變食物網在海洋生態系統動力學和營養結構 (Paine, 1966; Hinke et al., 2004),然而在餌料生物群聚的變化也可能影響海洋的頂級掠食者的資源
量 (Rosen & Trites 2000)。確認物種之間相互作用的營養關係,是朝向瞭解
和模擬生態系統的動態特性的重要一步。這些研究越來越重要,可用來進行 全球氣候變化對資源之改變研究,以及評估以生態系為基礎之動態影響。
攝食營養研究是有助於生態系統的分析,進而推論生物資源消長的過程 和漁業相互作用影響 (Lopez et al., 2010)。利用自然生物追蹤劑分析,例如:
碳及氮穩定同位素常用於攝食生態研究,經由食物網分析,可建構一個能量 流路徑的可靠方法 (Cabana and Rasmussen, 1996; Post, 2002)。後續可以用於 評估不同物種的攝食消費量和生物量,量化捕食與被捕食的關係,並分析生 態系統的各組成物種間的能量流,用以進行評估在群聚結構中對資源的衝 擊,隨後可應用於海洋生態系統的綜合管理。
胃內容物分析廣泛應用於營養動態研究,用來衡量生態區位重疊及建構 食物網,雖然受到隨機性攝食及不同消化率而產生誤差。胃內容物分析可提 供短期間內之詳細的攝食組成,餌料尺寸、分布、消化率等 (Chipps and Garvey, 2007)。穩定同位素可以追蹤長期的食性,但是會受到組織週轉率的 影響 (Gannes et al., 1998)。利用自然的生物追蹤劑,例如同位素碳 (δ13C) 及 同位素氮 (δ15N) 的比值,常被用來進行生態研究,建立追蹤食物網及估算 能量流更穩固的方法 (Cabana and Rasmussen, 1996; Post, 2002)。捕食者的氮 同位素值,雖然無法表示特定種類的攝食訊息,但當與基層消費者的營養階 層相比,則可提供攝食及營養關係之整合的觀點 (Post, 2002)。
雖然墨西哥及東太平洋雨傘旗魚,已完成食性及胃內容物分析,但在西 太平洋地區的食性訊息仍欠缺。先前的Tsai et al. (2014) 已於臺灣利用雨傘 旗魚穩定同位素分析,發現體長顯著影響氮同位素值,這表明雨傘旗魚之攝 食和營養階層,隨著不同體型大小而轉變。然而,旗魚和它們的獵物之間的 關係仍有待探討。在本研究中,我們經由胃內容分析,並利用穩定同位素分 析和擴展先前的研究,使用多種食物來源與雙同位素之穩定同位素混合模型 (Stable isotope mixing model ) (Parnell et al., 2010),來估算雨傘旗魚之攝食組 成和攝食生態。這些信息可以依序被用來估計雨傘旗魚捕食不同獵物之消費 率及生物量。此外,捕食和能量關係可以被量化,用以評估資源在群聚上的 影響,未來可以應用於結合以生態系統為基礎之管理。
海洋魚類攝食生態和食物網研究的最終目的,乃為海洋漁業資源永續利 用提供科學依據,實現以生態系統為基礎的漁業管理 (Ecosystem based fisheries management) (Pikitch et al., 2004)。本研究以臺灣東部海域生態系統 (marine pelagic ecosystem) 中的雨傘旗魚及其餌料魚類做為研究標的,應用 穩定氮同位素技術,計算各種餌料生物的營養階層,分析臺灣東部海域雨傘 旗魚的攝食種類與比例,探討該海域魚類種間的營養關係,以建構魚類群聚 中捕食主要的連結關係,本研究以描述在海洋生態系統中主要生態群組類別 的連結,目的為建置本海域雨傘旗魚與相關餌料生物之營養連結關係及營養 動力學之資訊。