Brain, music, and non-Poisson renewal processes
Simone Bianco,1Massimiliano Ignaccolo,1Mark S. Rider,2Mary J. Ross,3Phil Winsor,4and Paolo Grigolini1,5,6 1Center for Nonlinear Science, University of North Texas, P.O. Box 311427, Denton, Texas 76203-1427, USA
2
Integrative and Health Psychology, 700 West Wall Street, Suite 200, Grapevine, Texas 76051, USA 3
Neurotherapy Lab, Department of Rehabilitation, Social Work and Addiction, University of North Texas, P.O. Box 310829, Denton, Texas 76203-0829, USA
4
Center for Audio-Visual Experiment Microelectronics and Information Systems Research Center, National Chiao-Tung University, 1001 Ta Hsueh Road, Hsin-Chu, Taiwan 30050
5
Istituto dei Processi Chimico Fisici del CNR, Area della Ricerca di Pisa, Via G. Moruzzi, 56124 Pisa, Italy 6
Dipartimento di Fisica, “E.Fermi”-Università di Pisa, Largo Pontecorvo, 3 56127 Pisa, Italy 共Received 1 December 2006; published 21 June 2007兲
In this paper we show that both music composition and brain function, as revealed by the electroencepha-logram共EEG兲 analysis, are renewal non-Poisson processes living in the nonergodic dominion. To reach this important conclusion we process the data with the minimum spanning tree method, so as to detect significant events, thereby building a sequence of times, which is the time series to analyze. Then we show that in both cases, EEG and music composition, these significant events are the signature of a non-Poisson renewal process. This conclusion is reached using a technique of statistical analysis recently developed by our group, the aging experiment共AE兲. First, we find that in both cases the distances between two consecutive events are described by nonexponential histograms, thereby proving the non-Poisson nature of these processes. The corresponding survival probabilities⌿共t兲 are well fitted by stretched exponentials 关⌿共t兲⬀exp (−共␥t兲␣), with 0.5⬍␣⬍1.兴 The second step rests on the adoption of AE, which shows that these are renewal processes. We show that the stretched exponential, due to its renewal character, is the emerging tip of an iceberg, whose underwater part has slow tails with an inverse power law structure with power index=1+␣. Adopting the AE procedure we find that both EEG and music composition yield⬍2. On the basis of the recently discovered complexity match-ing effect, accordmatch-ing to which a complex system S withS⬍2 responds only to a complex driving signal P
withP艋S, we conclude that the results of our analysis may explain the influence of music on the human brain.
DOI:10.1103/PhysRevE.75.061911 PACS number共s兲: 87.19.La, 05.40.⫺a, 02.50.⫺r, 89.75.Hc I. INTRODUCTION
The study of neuronal systems is a challenge for statistical physics, insofar as experimental evidence is proving that the ordinary Poisson paradigm is inadequate to deal with these complex systems. According to some neurophysiologists the neurons are renewal关1兴 and they are markedly non-Poisson
关2兴. More precisely, the experimental evidence of in vitro
observations coupled with analysis of in vivo spiking pat-terns indicate that single neurons are fundamentally non-Poisson processes关3兴. In the literature the implicit
assump-tion is frequently made that even if the spiking activity of a single neuron is not Poissonian, the activity of a set of many neurons is Poissonian. This assumption may lead us to con-clude that the human brain is a Poisson system. The authors of the paper of Ref. 关4兴 proved that this assumption is
in-valid. In this paper, through the electroencephalogram共EEG兲 analysis we reach the conclusion that the human brain is not a Poisson system, in line with the theoretical remarks of Ref. 关4兴. It is important to point out that in this paper the term
non-Poisson process indicates a strong deviation from the exponential decay. This kind of non-Poisson behavior im-plies, as we shall see, the emergence of fat tails with an inverse power law behavior, although for many reasons, ranging from the finite time observation to the influence of spurious random fluctuations, in the long-time region these tails are truncated.
In this paper we apply the same statistical analysis to music composition. There is a wide agreement that music composition can be thought of as a complex signal. It is convenient to quote the seminal work by Voss and Clarke 关5兴. These authors have found in fact that music composition
yields a 1 / f noise spectrum, which is generally regarded as a complexity manifestation关6–9兴.
The power of music to evoke emotions is well known, but only recently, as discussed in the short review of Ref.关10兴, it
has attracted the attention of neuroscientists关11兴. How does
the communication between music and the brain take place? The authors of an interesting paper关12兴 have recently
stud-ied the brain response of musicians and nonmusicians to mu-sic listening and have found that mumu-sicians yield a higher degree of the gamma band synchrony. A more recent study of these investigators关13兴 has supported the theory that phase
synchronization is a significant marker in human cognition 关14兴.
The phase synchronization established by the authors of Refs. 关12,13兴 is one of the interesting properties of chaos
synchronization关15兴, which is in fact attracting the
increas-ing interest of neuroscientists关16兴. According to the seminal
work of Ref.关17兴 both driving system 共the music
composi-tion兲 and response system 共the brain兲 are dynamic systems with strange attractors共for instance, Rössler systems 关18兴兲.
In this paper we analyze both EEG data and music com-position with the joint use of two techniques, the minimum spanning tree 共MST兲 approach and the aging experiment
共AE兲. Both EEG and music composition data are expressed as multidimensional vectors Vi, with i = 1 , . . . , N. The
differ-ent compondiffer-ents of these vectors correspond to the informa-tion afforded by different electrodes in the EEG case, and timber, pitch, harmony, melody, rhythm,…etc., in the case of music composition. In this representation the existence of events is not evident. Thus to reveal their presence we build the MST proposed by Kruskal 关19兴 and we study its time
evolution: From time to time the topology of this MST un-dergoes an abrupt change that we interpret as an event. The probability of an event occurrence at a time distancefrom the preceding event is found to be a stretched exponential. With the help of the AE method we prove this process to be renewal. The conclusion of this paper is that both EEG and music composition are non-Poisson renewal processes, gen-erated by an inverse power law distribution with index ⬍2. This condition apparently departs from those conditions considered to be essential for synchronization, and so for the brain to be sensitive to music. We note, however, that it has been shown关20,21兴 that while a non-Poisson complex
sys-tem does not respond to harmonic perturbation关22,23兴, it is
very sensitive to the influence of a complex perturbation with the same complexity. The authors of Refs. 关20,21兴 denoted
this effect as the complexity matching 共CM兲 phenomenon. Therefore, the non-Poisson renewal condition is compatible with the transmission of information from a complex system to another, and, on top of that, thanks to the CM effect, might allow us to explain the influence of music on the brain.
The outline of this paper is as follows. In Sec. II, with the help of Appendix A, we show under which condition a sur-vival probability共SP兲 with the form of a stretched exponen-tial can be considered a non-Poisson renewal process with power law index ⬍2. In Sec. III we illustrate the MST method. Sections IV and V illustrate how we reached the conclusion that both EEG and music composition data are manifestations of non-Poisson renewal processes with ⬍2. In Sec. VI we concisely review the CM effect so as to support the main conclusion, illustrated in Sec. VII, that it is possible to tune music composition to the brain so as to make the brain respond to it.
II. STRETCHED EXPONENTIAL AS A TRUNCATED MITTAG-LEFFLER FUNCTION
The purpose of this section is to derive the following con-clusions:共i兲 A particular class of stochastic processes leads, in an appropriate limit, to a renewal process with SP de-scribed by a Mittag-Leffler共ML兲 function; 共ii兲 The ML func-tion has the smalllimiting form of a stretched exponential, exp(−共␥␣兲), with␣⬍1, and a largepower law limit;共iii兲 The power law index is related to the stretched exponential parameter␣ by Eq.共4兲; 共iv兲 One can deduce the power law
index from the smallstretched exponential fit, even in situ-ations where there is no information about the long tail in the data recorded. This is important for the main purpose of this paper. In fact, in Secs. IV and V we shall prove that EEG’s and complex sounds, respectively, are examples of such a process because they are renewal processes and are well fit-ted by a stretched exponential. By means of Eq.共4兲 we shall
derive the hidden information on.
The ML function关24兴 is attracting an ever increasing
in-terest in the literature of complex fluids such as liquid crys-tals, glass-forming liquids, and polymeric and colloid sys-tems关25兴. A remarkable property of the ML function is that
the complex susceptibility produced by the ML relaxation function yields the Cole-Cole experimental form 关26兴. For
this interesting property the reader can consult also Refs. 关25兴, 关27兴, and 关28兴.
The ML function establishes a bridge between the stretched exponential behavior for short time and an inverse power law in the long-time limit关29兴. This property is
im-portant, since it may settle the controversy between the ad-vocates of stretched exponentials and those of inverse power laws. In the case of the financial market, the authors of Ref. 关30兴 have found that the ML function affords a very good
fitting of experimental data. There is a problem with the tails, however, insofar as the lack of sufficiently rich statistics make noisy the time region where the ML fat tail should appear关31兴. The authors of Ref. 关31兴 used the aging
experi-ment to make an inverse power law behavior distinctly emerge from the noisy background of the long-time regime. Here we show an approach to the ML proving that in some noisy conditions only the stretched exponential portion of the ML function can remain visible. Our theoretical ap-proach refers to the time distance between two consecutive events, rather than a molecular relaxation process关26兴: The
motor driving our process is a physical generator of events. Let us assume that a natural-time scale exists, where this physical generator of events is a Poisson process with rate
rⰆ1, so as to make the discrete time representation virtually
indistinguishable from the continuous time picture. The time distance between two consecutive events produced by this generator is given by the waiting time distribution density
P共n兲, which has the following exponential form:
P共n兲 = r exp共− rn兲. 共1兲
To generate the sequence of events of interest for this paper, we operate as follows. We record the activity of this physical generator at each and every time step of its natural time n. At the same time, following the prescription of the subordina-tion theory关32兴, we turn the time n into the continuous time
t共n兲, by setting
t共n + 1兲 − t共n兲 =n. 共2兲
It is straightforward to prove 共see the Appendix兲 that the adoption of an inverse power law with no truncation and power index as subordination function, together with the renewal assumption for the events production and the condi-tion rⰆ1, creates a SP with the form of a ML funccondi-tion of order␣, denoted by the symbol ⌿SP, whose analytical
ex-pression is ⌿SP共t兲 ⬅ E␣„− 共␥t兲␣… =
兺
n=0 ⬁ 共− 1兲n 共␥t兲␣n ⌫共␣n + 1兲, 共3兲 with␣=− 1, 共4兲 and␥ is a time scale given by Eq. 共A8兲. In the case 0⬍␣
⬍1 it is well known that the ML function admits two limit-ing conditions, namely,
E␣„− 共␥t兲␣… ⬃共␥1
t兲␣ t→ ⬁, t ⬎
1
␥, 共5兲
that is, an inverse power law, and
E␣„− 共␥t兲␣… ⬃ exp„− 共␥t兲␣… t → 0, t ⬍1
␥, 共6兲
which is a stretched exponential.
To take into account that the data under study are finite, it is convenient 关32兴 to select for the subordination function 共兲 an inverse power law with index, which is exponen-tially truncated at tⲏ1/⌫, where ⌫⬍␥. As an effect of this choice共see the Appendix兲 we get for the function ⌿SP共t兲 a
resulting form virtually indistinguishable from a stretched exponential, in the intermediate time scale where the depar-ture from the exponential form is significant. This derivation of the stretched exponential is different from the one recently proposed by the authors of Ref.关33兴. However, a comparison
between our approach and that of these authors is not quite appropriate: in fact, Magdziarz and Weron关33兴 aim at the
same purpose as that of the earlier work of Ref. 关26兴, the
explanation of the nonexponential Cole-Cole relaxation, and do not afford prescriptions to evaluate the distribution of the time distances among consecutive renewal events, which is the main trust of this section.
The subordination process is realized by means of a ran-dom prescription and, consequently,⌿SP共t兲 is expected to fit
the renewal condition. We shall assess this property by means of the aging experiment of Sec. IV. We shall also establish a connection between the stretched exponential exp(−共␥t兲␣), with ␣⬍1 and the power index by means of the relation of Eq.共4兲 generated by the subordination
proce-dure. Since the subordination is realized with a function
共兲, which is a truncated inverse power law, the resulting process in the long-time scale does not violate the ergodic condition. Thus the Poisson behavior, and the consequent lack of aging, as we shall see by means of the statistical analysis of both EEG data and music composition, is recov-ered. The same limitations are shared by many other com-plex processes, see, for example, 关32兴, due to the obvious
fact that an exact inverse power law behavior is an idealiza-tion that would imply the infinite size of the systems under study. The study of this idealized condition is useful to shed light into the transient behavior before the eventual Poisson condition.
III. MINIMUM SPANNING TREE APPROACH AS A GENERATOR OF EVENTS
In this Section we introduce an algorithmic procedure that will allow us to process the data at our disposal so as to generate events and, consequently, time series to analyze. The method is based on the famous minimal spanning tree
共MST兲 algorithm 关34兴, which we now briefly introduce.
To define the MST approach we closely follow the argu-ments of Ref.关35兴. Imagine a data set consisting of N
col-umns, each column representing the signal recorded by an electrode, or the timber, pitch, note, …, etc., of the music composition. Taking two columns, say x and y, we define the correlation coefficient between the two columns as follows:
xy=
兺
k=0 t 共xk−具x典兲共yk−具y典兲 xy , 共7兲where the quantities具x典 andxare, respectively, the average and the standard deviation of the values that x takes over the interval关0,t兴. Consider now the following quantity:
x
˜ =x −具x典
x
共8兲 and assume that in the interval关0,t兴 there are t values of the vector x˜, namely, x˜k, k = 1 , . . . , t. The distance between two
columns x˜ and y˜ over the time interval t is easily obtained adopting the well known formula of the Euclidean distance
dxy 2 =储x˜ − y˜储2=
兺
k=1 t 共x˜k− y˜k兲2. 共9兲Moreover,兺k=1t ˜xk2= 1. Therefore, Eq. 共9兲 becomes
dxy 2 =
兺
k=1 t 共x˜k 2 + y˜k2− 2x˜k˜yk兲 = 2 − 2兺
k=1 t x ˜k˜yk. 共10兲The last sum on the right-hand side of Eq.共10兲 is the
corre-lation coefficient between x and y of Eq.共7兲 at the time t. It
is therefore possible to define the distance between the col-umns x and y as
dxy=
冑
2共1 −xy兲. 共11兲 Since −1⬍xy⬍1, then 0⬍dxy⬍2. Moreover, dxy fulfills the property of a distance关35兴.Through the distance dxywe now introduce the MST ap-proach over the time interval t. In a connected graph of N objects, weighted through the distances dxy, the MST is the tree with N − 1 links for which the total sum of the edges is minimum. In the literature there are many algorithms to cre-ate the MST. Here we select the method proposed by Kruskal 关19兴. This method consists of the following steps: we sort the
distances in increasing order; we select the shortest distance and draw an edge between the associated nodes; we go to the next distance and draw edges; if an edge creates a loop we erase it; we continue drawing lines until all the N columns are represented.
In this paper we make a dynamical use of the MST. We build the MST over a time window of length t, then we move to the next共nonoverlapped兲 window and build the MST cor-responding to the new position. An event is defined when-ever a change in the distribution of links occurs. This way of proceeding allows us to define events and, consequently, the distribution of waiting times between two consecutive, events. Moreover, when building the distribution of the
num-ber of edges, we do not label our electrodes, but simply look at the shape of the distribution, that essentially characterizes the topology of the MST. If distributions at subsequent times do not exactly coincide, we consider it to be a signal of a change in the global properties of the brain, and indicate this as our critical event.
The MST has been recently adopted in the literature in an increasing number of papers on different complex systems, from networks 共e.g., 关36兴 among the others兲 and financial
markets共e.g., 关37兴 among the others兲 to neurophysiological
processes 关38兴. Particularly, we quote the recent paper of
McDonald et al.关39兴. In this paper the MST approach has
already been used and applied to financial time series, but in a different way. In fact, the authors of Ref.关39兴 monitor the
survival ratio of edges in time, that is, the ratio between the number of edges connected to a certain node at time 0 and the same quantity computed at a later time t, and find that it decays nonexponentially.
According to Ref.关39兴, the MST, by definition, is more
suitable to study positively than negatively correlated sys-tems. Indeed, negatively correlated electrodes contribute dif-ferently to the MST than positively correlated electrodes. In our analysis this criticism only relatively applies, as we are interested in the changes of the MST topology in time, i.e., in the changes of the correlations with time.
In the next section we shall analyze the statistical proper-ties of the time series obtained in the aforementioned way.
IV. ANALYSIS OF EEG
Digital EEG data was collected on five subjects, utilizing a Scan LT-40 amplifier, manufactured by NeuroScan Medical Systems, El Paso, Texas. The Scan LT-40 is an FDA ap-proved medical device for the collection of digital EEG. On-line monitoring of EEG was provided by NeuroScan Medical Systems 1.2 software. Offline evaluation and the removal of artifacts from the EEG record and the conversion of the data to ASCII files were accomplished using NeuroGuide 2.2.6 software. An electrode cap from Electrocap International Inc. was used to provide standardized electrode placement. Digi-tal EEG data was collected from 19 locations using the in-ternational 10-20 system of electrode placement. A reference electrode was placed on each earlobe to provide a linked ears montage for the physical reference of the scalp recordings. The impedance of the respective earlobe reference electrodes was maintained within 1 K⍀ of each other. All other elec-trode impedances were maintained at 1 or 2 K⍀ relative to amplifier input impedance with no more than 1 K⍀ of vari-ance between any of the electrode contacts. The amplifiers used to acquire the EEG were calibrated with sine waves before the acquisition of EEG. The EEG data was digitized at a rate of 250 samples per second. Before analog to digital conversion, antialiasing was achieved by a low-pass filter built into the software. The EEG was visually inspected on-line during acquisition to monitor for artifact. When neces-sary, data collection was stopped to identify and remove per-sistent sources of artifact such as muscle tension. The subjects included two healthy subjects, one subject with chronic back pain, and two subjects with mental depression.
Subjects were medication-free during the data acquisition. The average number of data collected is 26 141, correspond-ing to about 104 seconds of record. The minimum number of data for a single subject is 10 961, corresponding to about 43 seconds of recording, for one of the healthy subjects, while the maximum number of data for a single subject is 56 544, corresponding to about 226 seconds, for the subject with chronic back pain. Only two states were considered for the subjects, namely, eyes open共EO兲 and eyes closed 共EC兲. The acquisitions were made in the same conditions for all the subjects.
The MST approach, discussed in the previous section, is applied to the set of 19 columns that the data acquisition method affords. The time series of MST topological changes is obtained and, to be consistent with the theoretical results of Sec. II, the SP⌿共t兲 of the time distances between two consecutive MST topological changes is evaluated. The length of the time window inside which the MST is evalu-ated has been chosen to be t = 3 samples, corresponding to 0.012 seconds. In Fig.1 we show the SP relative to the in-dividual with back pain in the EO condition. We fit the re-sulting curve with a stretched exponential of the form of Eq. 共6兲 with parameters␥= 0.205 and ␣= 0.595.
Similar results have been obtained for the SPs of all the other individuals at our disposal. Using Eq.共4兲, we find that
the values of for the EO condition lie between 1.595± 0.005 and 1.960± 0.013, while for the EC condition, between 1.746± 0.018 and 1.974± 0.024.
The results of Fig.1 suggest that the process is strongly non-Poissonian. However, the mere analysis of the distribu-tion of waiting times is not enough to establish the real na-ture of the process. In fact, as pointed out in Refs.关40,41兴, a
modulated Poisson process as well can have as an outcome a nonexponential waiting time distribution. In order to assess if the time series under study is produced by a genuinely non-Poisson renewal process, the authors of Ref.关41兴 propose the
renewal aging experiment共AE兲. According to this procedure, in addition to the waiting time density共兲, an aged waiting time density exp共, ta兲 must be evaluated. This is done as
follows: Let 兵ti其 be the series of absolute time obtained
0.001 0.01 0.1 1 0.1 1 10 100 1000 Ψ(τ) τ Ψ e-(γ τ)α
FIG. 1. The SP of MST topological changes for the individual with back pain in the EO condition. The curve is fitted using a stretched exponential of the form of Eq.共6兲 with␥=0.205 and ␣
through the prescription indicated in the previous section. For each time ti the first time of the sequence at a distance
from ti equal to or larger than ti+ ta is recorded. This time
will be tk, with k⬎i. The time distance共ti, ta兲=tk−共ti+ ta兲 is
considered. The procedure is repeated for all the times of the sequence兵ti其, and the sequence of these recorded time
dis-tances is used to generate the distribution densityexp共, ta兲.
Moreover, the following quantity is evaluated:
ren共,ta兲 =
冕
0 ta dynum共+ y兲 K共ta兲 , 共12兲where K共ta兲 is a suitable normalization constant andnum共t兲
is the numerical waiting time distribution corresponding to
ta= 0.
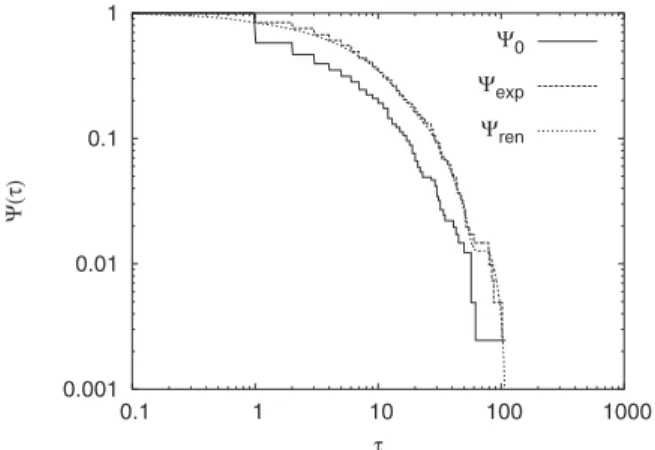
To establish the renewal character of the process by means of the AE it is convenient to plot the corresponding SP’s, ⌿ren共, ta兲, ⌿exp共, ta兲, and the SP of age ta= 0,
indi-cated as⌿0. In Fig. 2the three SPs are compared, and it is found that ⌿exp and ⌿ren virtually coincide. According to
Ref.关41兴 this good accordance is the numerical evidence of
the renewal nature of the process.
Moreover, this result confirms the theoretical framework of Sec. II, allowing us to obtain the index of the inverse power law distribution from the index of the stretched expo-nential, namely, from Eq. 共4兲. Since all the values of␣ are smaller than 1 we conclude that⬍2, and therefore that the human brain is a nonergodic system. It is important to repeat here what we already pointed out in Sec. II, that in the long run, as all the anomalous processes occurring in nature, the ergodic properties, as well as the Poisson condition and the associated lack of aging, are recovered.
How can we explain the emergence of this stretched ex-ponential of renewal origin? An anonymous referee pointed to us that there may exist a dominant structure and that the spanning trees appearing in the processed data may produce short and long waiting times according to their distance from the dominant structure. If there is a systematic alternation of short and long times, we end up in a model recently dis-cussed in Ref. 关40兴. In this case there would be a drastic
reduction of renewal aging. If on the contrary, the process is not a systematic alternation of short and long waiting times, it becomes indistinguishable from the non-Poisson model that we propose with the theory of Sec. II.
Now we have to address an important issue, concerning the physical origin of the complexity property revealed by our analysis. We share the opinion of Haken 关42兴 that the
brain global behavior is an emergent property produced by a synergetic process of synchronization 关43兴. As a
conse-quence, the different electrodes are correlated and the MST of Sec. III detects the emergence of this global property. The electrodes on which the MST analysis is based are single units, like the interacting columns of a surface growing as an effect of random deposition 关32兴. Due to the cooperation
with the other columns the single columns inherit the com-plexity of the whole growing surface关32兴.
On the basis of this observation, we are tempted to make the conjecture that the single electrodes inherit the global complexity. It is important to stress that Buiatti and his co-workers 关44兴 have recently obtained results confirming this
conjecture, with a method of analysis based on the observa-tion of a single EEG. Our conjecture is also compatible with the results found by the authors of Ref.关45兴. To understand
the connection between the results of this section and those found by the authors of Ref.关45兴 the reader should consult
the recent article of Ref.关46兴. The authors of this paper have
studied the dynamic approach to fractional Brownian motion 共FBM兲, as expressed by
d
dtx共t兲 =共t兲, 共13兲
where共t兲 is a fluctuating velocity with memory, namely, a correlation function with slow tails. The dynamical approach to FBM proves that the variable x in the asymptotic time regime shares the same properties as the traditional form of FBM. They have also proved that the origin recrossing of
x共t兲 generates a non-Poisson renewal process with the power
related to the FBM scaling coefficient H by
= 2 − H. 共14兲
Note that the adoption of the more realistic model of the interacting columns of an interface growing as an effect of random deposition of particles would produce saturation and also a truncation of the inverse power law waiting time dis-tribution, a property similar to the long-time Poisson behav-ior revealed by Figs.1and2. However, if the observation is limited to the short-time region, the condition of Eq. 共14兲
applies.
The analysis of the authors of Ref. 关45兴 rests on 共t兲,
rather than on x共t兲. We are convinced in fact that the single electrodes yield a signal that has to be interpreted as the variable x of Eq.共13兲. Thus to relate the results of Ref. 关45兴
to the results of this section, we must adopt Eq.共14兲. This
procedure generates⬍2. With this interpretation in mind, we find that also the analysis of Ref. 关45兴 proves that
indi-viduals whose signal was recorded in the EC condition is closer to= 2 than the individuals in the EO condition. 0.001 0.01 0.1 1 0.1 1 10 100 1000 Ψ(τ) τ Ψ0 Ψexp Ψren
FIG. 2. The AE analysis on the time series of Fig. 1, for ta = 30. The aging is present and compatible with the renewal condi-tion.共ta= 30兲.
Stressing this result is a way for us to draw the attention of the reader to the fact that the discovery of non-Poisson renewal events, made possible by the method adopted in this paper, does not yield results conflicting with the work of other groups. In TableI, we present a sample of the results of our analysis on our group of individuals. We remind that two of them were healthy, one had back pain, and two had mental depression. The values ofrefer to the same subject, in the two conditions. We see that the parameters of individuals in the EC condition are significantly closer to 2 than the parametersof patients in the EO condition. In TableIIwe report a sample of the results of Ref.关45兴 共healthy
individu-als only, TableI兲 in terms of, by means of the rule of Eq. 共14兲. By comparing the results of TableIwith those of Table
IIwe reach the conclusion that the results of our analysis are compatible with those of Ref. 关45兴. It is necessary to stress
that this result is not statistically significant, given the small size of our sample, but it represents indeed a trend, compat-ible with the results present in the literature. More research work is necessary to be done to confirm the presence of this interesting effect.
Before ending this section, we want to stress that the main result of this paper, namely, the surprising complexity match-ing between brain and complex sounds that we shall discuss in Sec. V, does not depend in any way on the conjecture we make about the emergence of⬍2 in the single electrodes. This conjecture would have the effect of explaining the
find-ings of Refs. 关44,45兴, but, if proved wrong, would not
weaken the validity of the main result of this paper. V. ANALYSIS OF MUSIC COMPOSITION
The music composition data that we analyze in this sec-tion have been produced by means of a virtual instrument 共software synthesizer兲 suitably designed 关47,48兴 to produce
physiological effects 关49,50兴. The instrument is capable of
generating abstract sonic textures that are free from overt cultural influences. Moreover, through the use of presets, a record can be kept of all parameters of consequence in the generation of the musical material. This feature is important for purposes of correlating the output sonic textures with, for instance, biological data from EEG recordings. In this way, researchers can precisely pinpoint areas of interest in biologi-cal data for analysis of and comparison with the generative parameters of the sound structures. Designed to allow control over the redundancy of time-point and pitch共frequency兲 pat-terns in a hierarchical framework, the virtual instrument sonic textures can be gradually morphed between constant states 共stable兲 regimes and chaotic 共complex兲 regimes via controls built into the graphical user interface. Moreover, precise measurement and recording of all generative schemes is possible, as well as synchronization with the EEG data time series. An important characteristic of the instrument is that vertical sonic textures are flexibly configurable with re-spect to the degree of vertical and horizontal redundancy of pitch- and time-space organization within the ongoing sonic flow. Various degrees of complexity can be introduced via presets prior to and during the transmission of audio data to the test subject. For instance, microtonal controls are built into the instrument, so that the researcher can regulate the content of the sonic texture at any given point to create a relative harmonicity shift of the composite sonic data stream. In other words, there is maximum control over the degree of complexity via overlapping, phase-modulated patterning of melodic, harmonic, rhythmic, and texture-density data.
The instrument produces a vector V共t兲, with seven com-ponents, namely, pitch onset time共in milliseconds兲, note fre-quency 共in hertz兲, wave form type, amplitude, articulation, preset number, and oscillator number. All these components are filed and assumed to afford information about the signal. We therefore adopt for the music composition the same pro-cedure applied before for the EEG signal, building also in this case a time series of MST topological changes. The re-sulting SP for a sample record is plotted in Fig.3.
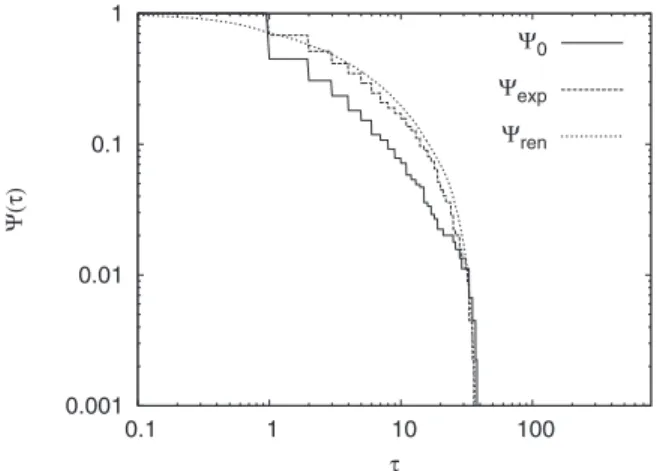
We see from the figure that also in this case the distribu-tion of topological changes produces a nonexponential SP, namely a stretched exponential with parameters ␥= 0.430 and␣= 0.600. As in Sec. IV, a further test is needed to prove that the process is renewal. The results of the AE are plotted in Fig.4.
Also in this case the AE supports the renewal assumption for the process. We conclude that music composition shares the same properties as the EEG signal analyzed in Sec. IV. This suggests that both human brain and music composition, sharing the condition⬍2, are complex systems that, in the absence of the exponential truncation, would violate the er-godic condition关51兴.
TABLE I. This table shows a sample of the results of the analy-sis on our group of individuals in the EO共left side of the table兲 and EC共right side of the table兲 conditions. Results show a larger value of for the EC condition. In parentheses we report the standard deviation. The symbol H indicates a healthy individual, BP the individual with back pain, and D the subjects with mental depression. Id EO EC H1 1.730共0.010兲 1.800共0.010兲 H2 1.740共0.012兲 1.770共0.013兲 BP 1.595共0.005兲 1.746共0.018兲 D1 1.748共0.013兲 1.787共0.014兲 D2 1.960共0.023兲 1.974共0.024兲
TABLE II. This table shows a sample of the results of the analy-sis of Ref.关45兴 on healthy individuals in the EO 共left side of the
table兲 and EC 共right side of the table兲 conditions. The parameter has been obtained according to Eq. 共14兲. Results show a larger
value of for the EC condition. EO EC 1.261 1.888 1.242 1.931 1.294 1.898 1.278 1.877
VI. SHORT REVIEW OF THE CM EFFECT The authors of Refs.关23,52兴 have developed a linear
re-sponse theory that applies to non-Poisson renewal systems. The form of this linear response theory is given by
⌸共t兲 ⬅ 具S共t兲典 =⑀
冕
0
t
dt
⬘
共t,t⬘
兲P共t⬘
兲, 共15兲where S共t兲 denotes the signal produced by the system of
interest,共t,t
⬘
兲 is the linear response function, and P共t兲 isthe external perturbation. In the case of the human brain the signalS共t兲 is a global property emerging from the
synchro-nization of different brain areas, roughly corresponding to the superposition of the signals detected by the electrodes fixed on the patient’s scalp. In the absence of perturbation this global signal is characterized by the non-Poisson re-newal events revealed by the method described in Sec. IV. To transform these data intoS共t兲, we assign alternate signs to
the quiescent time regions between two consecutive events. The complexity of this signal is denoted by means of the power indexS⬍2, emerging from the analysis of Sec. IV.
If the perturbation functionp共t兲 is harmonic the system
does not respond 关23兴. This is a clear sign of complexity,
insofar as a non-Poisson renewal signal cannot be interpreted as the superposition of infinitely many independent pro-cesses. The individual constituents of a set of neurons re-sponsible for any cognitive action are expected to be strictly cooperating with all the others’ constituents. As a conse-quence, a harmonic perturbation triggers a cascade of differ-ent time scales, thereby violating the prescriptions of ordi-nary stochastic resonance processes关23,53兴. Thus, we are led
to make the conjecture that the transmission of information from the perturbing signal to the human brain is determined by the interaction between the renewal events of the perturb-ing signal and the renewal events of the perturbed system. If the perturbing signal does not have any renewal events, as in the case of a harmonic perturbation, there is no response to a weak perturbation.
More recently the authors of Refs. 关20,21兴 have proved
that this conjecture is correct, and that in the case whereP共t兲
is a signal derived from another non-Poisson renewal system with indexP⬍2, the system responds, and the intensity of
the response is maximum when we use the matching condi-tionS=P 关20兴. The authors of Refs. 关20,21兴 have denoted
this effect with the name of complexity matching共CM兲, and they proved that whenP⬍Sthe perturbed system inherits
the perturbation power index. In Sec. V we have proved that music composition is actually a complex signal withP⬍2.
Thus, the existence of the CM effect leads us to conjecture that the reason why the brain is sensitive to music lies on the fact that both the brain and music are non-Poisson renewal systems living in the nonergodic region.
VII. CONCLUDING REMARKS
The literature on complexity is wide, and there are many different proposals to account for complex processes. All these proposals share only one essential property, this being the departure from the canonical exponential distribution of ordinary statistical physics. The authors of Ref. 关41兴 have
pointed out that this departure can be realized by means of quite different physical processes, either nonhomogeneous Poisson processes or homogeneous non-Poisson processes. The different physical origin of these two processes is re-vealed by the AE.
According to the authors of Refs. 关54,55兴 there exists a
close connection between self-organized criticality 关6兴,
su-perstatistics 关56兴, and nonextensive thermodynamics 关57兴.
This analysis coincides with the critical view illustrated by Jensen关58兴. It is very attractive to conjecture that the brain
operates at or near a self-organized critical state关55,59兴. This
corresponds to the recent observation 关43,60兴 that neuron
synchronization is a sort of phase transition involving a close cooperation among the elementary constituents of the neuron set. However, in our opinion, the main limits of these inter-esting theories is that they do not pay attention to the impor-tant role of renewal events, whose objective existence is made compelling by the results of the AE, in both the case of blinking quantum dots 关41兴 and of neuron synchronization
关43兴. 0.001 0.01 0.1 1 0.1 1 10 100 Ψ(τ) τ Ψ e-(γx)α
FIG. 3. The SP of MST topological changes obtained from the music composition. The curve is fitted using a stretched exponential of the form of Eq.共6兲, with␥=0.430 and ␣=0.600 共ta= 100兲.
0.001 0.01 0.1 1 0.1 1 10 100 Ψ(τ) τ Ψ0 Ψexp Ψren
FIG. 4. The AE on the time series of Fig.3. The aging effect is compatible with the renewal assumption.共ta= 100兲.
We think that, although these properties have been over-looked by the majority of the researchers working in the field of complexity, they deserve more attention, and we hope that this paper may serve the important purpose of raising the interest of the investigators in this direction.
If the importance of these non-Poisson renewal events is not recognized, the results of the analysis of this paper on both EEG data and music composition, which are based on a solid method of statistical analysis 关41兴, are
incomprehen-sible. If, on the contrary, we accept the leading idea that complexity rests on the close cooperation of elementary components losing their own identity at the moment of the onset of synchronization, then the fact that both music com-position and the human brain are non-Poisson renewal pro-cesses becomes a natural way of explaining why music ex-erts its influence on the brain.
This conclusion is so important as to deserve further re-marks. First of all, a more appropriate term to denote the music composition analyzed in this paper would be complex
sound. In fact, the main purpose of the music composition
utilized in this work is to affect brain complexity rather than generate emotional and/or aesthetic responses, per se. Thus, to make the composition of these complex sounds more flex-ible and more suitable to the purpose of realizing an efficient transport of information from acoustic excitation to the hu-man brain, thanks to the CM effect关20,21兴, we have to focus
on the realization of a givenP more than on aesthetic
pur-poses. On the basis of the CM effect关20,21兴 the network of
EEG electrodes is expected to inherit the samePas that of
the acoustic excitation if P⬍S, where S denotes the
brain complexity index. We plan to design a real experiment based on recording EEG data from a subject listening to music with an exponentP. We plan also to observe whether
the exponent of the EEG changes if the listener hears songs with different exponents. All of this requires much more work, and the solution of technical as well as conceptual problems. However, we are convinced that the results of this paper are important enough as to trigger further research work for the realization of this important experiment.
ACKNOWLEDGMENTS
We warmly thank Dr. Marco Buiatti for interesting discus-sions, and for communicating to us his results on the analysis of EEG data prior to the publication. S.B., M.I., and P.G. thankfully acknowledge Welch and ARO for financial sup-port through Grants No. B-1577 and No. W911NF-05-1-0205, respectively. This project was approved by the Institu-tional Review Board for the Protection of Human Subjects 共Project No. 2006-47兲 of the University of North Texas. The data were archived from subjects who had given prior writ-ten consent for the confidential use of their test results in future research.
APPENDIX
Let us address the problem of establishing the SP⌿SP共t兲
corresponding to a set of sequences共t兲 prepared at t=0. It is evident that ⌿SP共t兲 =
兺
n=0 ⬁冕
0 t dt⬘
n共t⬘
兲⌿共t − t⬘
兲exp共− rn兲. 共A1兲To explain Eq.共A1兲, let us notice that ⌿SP共t兲 is the
probabil-ity that the physical generator does not produce any further event, after the initial preparation event, until time t. In the natural time scale the probability density of not producing an event is exp共−rn兲. To evaluate ⌿SP共t兲, we have to multiply
exp共−rn兲 by n共t
⬘
兲 and by ⌿共t−t⬘
兲. This is becauseexp共−rn兲 indicates that until time t the physical generator acts n times, thereby making n drawings from the distribu-tion density共兲. Note that共t兲 is also called subordinating function 关32兴. This function is the probability distribution
density affording information on the time distance between two consecutive actions of the physical generator, not neces-sarily producing events. With no drawing 共n=0兲, we get ⌿SP共t兲=⌿共t兲, where ⌿共t兲 is the SP corresponding to 共t兲,
namely, the probability that the physical generator does not act until time t. With n drawings we fill a time interval of length 1+2+¯ +n= t
⬘
⬍t. The factor ⌿共t−t⬘
兲 ensuresthat no drawing occurs in between t
⬘
and t. The functionn共t
⬘
兲 denotes the probability that n drawings from thedis-tribution共兲 occurred, the last of which occurred exactly at time t
⬘
. Due to the renewal nature of this process we haven共t兲 =n−1共t兲丢1共t兲, 共A2兲
where丢 indicates time convolution and1共t兲=共t兲. The Laplace transform of⌿SP共t兲, ⌿ˆSP共u兲, is expressed as
a function of the Laplace transform of共t兲,ˆ 共u兲, as follows: ⌿ˆSP共u兲 =
1
关1 −ˆ 共u兲exp共− r兲兴
1
u关1 −ˆ 共u兲兴. 共A3兲
To obtain the previous expression, still no hypothesis on the form of共兲 has been done. If 共兲 is an exponential, the subordination exerts no physical effect on the SP, as its form remains exponential. Let us see the case of nonexponential subordination function. With straightforward algebra, and as-suming rⰆ1, Eq. 共A3兲 becomes
⌿ˆSP共u兲 = 1 u + r⌽ˆ共u兲 , ⌽ˆ共u兲 = uˆ 共u兲 1 −ˆ 共u兲 , 共A4兲
that is, the Laplace transform of
d
dt⌿SP共t兲 = − r
冕
0t
⌿SP共t − t
⬘
兲⌽共t⬘
兲dt⬘
, 共A5兲and ⌽共t
⬘
兲 is a memory kernel. Note that the assumption r Ⰶ1 is not necessary to generate the time convoluted struc-ture of Eq.共A5兲. It is essentially required to make the naturaltime n compatible with a continuous time representation, thereby yielding for the survival probability in the natural time scale the exponential form exp共−rn兲. An even greater consequence of rⰆ1 is that, as we shall see hereby, this condition makes very extended the stretched exponential regime.
In the case where the memory kernel⌽共t兲 is a delta of Dirac, Eq.共A5兲 makes ⌿SP共t兲 become an ordinary
exponen-tial. To generate a stretched exponential we must make a proper choice of the memory kernel⌽共t兲, and consequently, of the subordination function. Let us assign to the memory kernel in the Laplace space the following form:
⌽ˆ共u兲 =共u + ⌫兲2−, ⬅ 关⌫共2 −兲T−1兴−1, 共A6兲 and let us assume the parameterto be smaller than 2. With this choice the SP becomes, in the Laplace domain,
⌿ˆSP共u兲 =
1
u +␥␣共u + ⌫兲1−␣, 共A7兲
with
␣⬅− 1, ␥⬅ 共r兲1/␣. 共A8兲
If⌫=0, we recognize in Eq. 共A7兲 the well known Laplace
transform of a ML function of order␣ 关see Eq. 共3兲兴. For␣
= 1, the ML function becomes an exponential.
Note that the parameter ⌫ has the important role, as in Ref. 关32兴, of taking into account that we are working with
systems of finite, rather than infinite size. Consequently, we must assign to⌫ a finite value. Thus, let us assume that ⌫ ⬎0 and that ⌫Ⰶ␥Ⰶ1. Note that ␥Ⰶ1 is generated by r Ⰶ1. The condition ⌫⬍␥ is made necessary by the request that the truncation of the fat tail of the subordinating function leaves some sign of the system complexity. However, this condition implies a departure from the pure ML relaxation function, and the form of⌿SPon the time scale at which we
observe the histogram and SP. In the short-time regime ␥ ⰆuⰆ1 it is impossible to neglect the first term in the de-nominator of Eq. 共A7兲, and therefore, considering that ⌫
Ⰶu,
⌿ˆSP共u兲 =
1
u +␥␣u1−␣, 共A9兲
which is the Laplace transform of a stretched exponential. On the contrary, if the condition⌫ⰆuⰆ␥ applies, the first term in the denominator of Eq.共A7兲 can be neglected, and
the expression for the SP reads ⌿ˆSP共u兲 =
1
␥␣u1−␣, 共A10兲
which, thanks to the Tauberian theorem, is the Laplace trans-form of an inverse power law SP. In this condition, the fat
ML function tail becomes visible. If ⌫ⱗ␥, namely, ⌫ is moderately smaller than␥, Eq.共A7兲 shows that the inverse
power law never appears.
Let us now find analytically the form of the subordination function 共t兲. If we adopt the expression for the memory kernel of Eq.共A6兲, we obtain
ˆ 共u兲 = ⌽ˆ共u兲 u +⌽ˆ共u兲 = 1 1 + u ⌽ˆ共u兲 = 1 1 + u 共u + ⌫兲2− . 共A11兲 Again, two regimes clearly appear depending on the param-eters involved in Eq. 共A11兲: if ⌫ⰆuⰆ1, then Eq. 共A11兲
becomes
ˆ 共u兲 ⯝ 1 −1
u−1, 共A12兲
that is, the Laplace transform of an inverse power law distri-bution density. If we explore the regime u⬍⌫, Eq. 共A11兲
reads
ˆ 共u兲 ⯝ 1
1 + u
⌫2−
, 共A13兲
that is, the Laplace transform of an exponential distribution density, implying that the subordination function 共t兲 is an inverse power law, truncated exponentially in the long time limit.
In conclusion, in the case of systems of finite size the fat tail of the subordination function is truncated. In the case where r is not very small, and consequently,␥is only mod-erately small, the stretched exponential regime is not very extended and the adoption of a truncated subordination func-tion generates a ⌿SP共t兲, with a distinct inverse power law
tail. If rⰆ1, and consequently, the stretched exponential re-gime is very extended, a truncated subordination function may have the effect of canceling the inverse power law tail of the SP, and the stretched exponential remains the only sign of complexity. However, the subordination function has an inverse power law nature, and its power index is derived from␣through Eq.共A8兲, thereby yielding Eq. 共4兲.
关1兴 C. van Vreewijk, Neurocomputing 38, 417 共2001兲.
关2兴 R. Baddeley, L. F. Abbott, M. C. A. Booth, F. Sengpiel, T. Freeman, E. A. Wakeman, and E. T. Rolls, Proc. R. Soc., Lon-don, Ser. B 264, 1775共1997兲.
关3兴 C. F. Stevens and A. M. Zador, Nat. Neurosci. 1, 210 共1998兲; Y. Sakai, S. Funahashi, and S. Shinomoto, Neural Networks 12, 1181共1999兲; Y. Sakai, ibid. 14, 1145 共2001兲; S.
Shino-moto and Y. Tsubo, Phys. Rev. E 64, 041910共2001兲; J. Feng and P. Zhang, ibid. 63, 051902共2001兲; E. Salinas and T. J. Seinowski, Neural Comput. 14, 2111共2002兲; S. Shinomoto, K. Shima, and J. Tanji, ibid. 15, 2803共2003兲; M. N. Shadlen and W. T. Newsome, J. Neurosci. 18, 3870共1998兲.
关4兴 H. Câteau and A. D. Reyes, Phys. Rev. Lett. 96, 058101 共2006兲.
关5兴 R. F. Voss and J. Clarke, J. Acoust. Soc. Am. 63, 258 共1978兲. 关6兴 P. Bak, C. Tang, and K. Wiesenfeld, Phys. Rev. Lett. 59, 381
共1987兲
关7兴 M. Paczuski and P. Bak, Proceedings of the 12th Chris Engle-brecht Summer School 共1999兲; e-print arXiv:cond-mat/ 9906077共1999兲.
关8兴 M. Paczuski and D. Hughes, Physica A 342, 158 共2004兲. 关9兴 R. Bove, V. Pelino, and L. De Leonibus, Commun. Nonlinear
Sci. Numer. Simul. 11, 678共2006兲.
关10兴 P. E. Andrade and J. Bhattacharya, J. R. Soc. Med. 96, 284 共2003兲.
关11兴 The Biological Foundation of Music, edited by R. J. Zatorre and I. Peretz, Ann. N. Y. Acad. Sci. 930, 1X共2001兲.
关12兴 J. Bhattacharya and H. Petsche, Phys. Rev. E 64, 012902 共2001兲.
关13兴 J. Bhattacharya and H. Petsche, Signal Process. 85, 2161 共2005兲.
关14兴 F. Varela, J. P. Lachaux, E. Rodriguez, and J. Martinerie, Nat. Rev. Neurosci. 2, 229共2001兲.
关15兴 S. Boccaletti, J. Kurths, G. Osipov, D. L. Valladares, and C. S. Zhou, Phys. Rep. 366, 1共2002兲.
关16兴 M. Chavez, M. Besserve, C. Adam, and J. Martinerie, J. Neu-rosci. Methods 154, 149共2006兲.
关17兴 M. G. Rosenblum, A. S. Pikovsky, and J. Kurths, Phys. Rev. Lett. 76, 1804共1996兲.
关18兴 O. E. Rössler, Phys. Lett. 57A, 397 共1976兲. 关19兴 J. B. Kruskal, Proc. Am. Math. Soc. 7, 48 共1956兲.
关20兴 P. Allegrini, M. Bologna, P. Grigolini, and M. Lukovic, e-print arXiv:cond-mat/0608341.
关21兴 P. Allegrini, M. Bologna, P. Grigolini, and B. J. West, e-print arXiv:cond-mat/0612303.
关22兴 I. M. Sokolov, A. Blumen, and J. Klafter, Physica A 302, 268 共2001兲; I. M. Sokolov, Phys. Rev. E 73, 067102 共2006兲. 关23兴 F. Barbi, M. Bologna, and P. Grigolini, Phys. Rev. Lett. 95,
220601共2005兲.
关24兴 G. M. Mittag-Leffler, C. R. Acad. Sci. Paris 137, 554 共1903兲. 关25兴 J.-L. Déjardin and J. Jadzyn, J. Chem. Phys. 123, 174502
共2005兲.
关26兴 K. Weron and M. Kotulski, Physica A 232, 180 共1996兲. 关27兴 R. Metzler and J. Klafter, J. Non-Cryst. Solids 305, 81 共2002兲. 关28兴 W. T. Coffey, J. Mol. Liq. 114, 5 共2004兲.
关29兴 R. Metzler and J. Klafter, Phys. Rep. 339, 1 共2000兲. 关30兴 M. Raberto, E. Scalas, and F. Mainardi, Physica A 314, 749
共2002兲.
关31兴 S. Bianco and P. Grigolini, Chaos, Solitons Fractals 34, 41 共2007兲.
关32兴 R. Failla, P. Grigolini, M. Ignaccolo, and A. Schwettmann, Phys. Rev. E 70, 010101共R兲 共2004兲.
关33兴 M. Magdziarz and K. Weron, Acta Phys. Pol. B 37, 1617 共2006兲.
关34兴 T. H. Cormen, C. E. Leiserson, R. L. Rivest, and C. Stein, Introduction to Algorithms, 2nd ed. 共MIT Press, Cambridge,
MA, and McGraw-Hill, New York, 2001兲.
关35兴 H. E. Stanley and R. N. Mantegna, An Introduction to Econo-physics: Correlations and Complexity in Finance共Cambridge University, Cambridge, UK,2000兲.
关36兴 Z. Wu, L. A. Braunstein, S. Havlin, and H. E. Stanley, Phys. Rev. Lett. 96, 148702共2006兲.
关37兴 G. Bonanno, G. Caldarelli, F. Lillo, and R. N. Mantegna, Phys. Rev. E 68, 046130共2003兲.
关38兴 U. C. Lee, S. Kim, and K.-Y. Jung, Phys. Rev. E 73, 041920 共2006兲.
关39兴 M. McDonald, O. Suleman, S. Williams, S. Howison, and N. F. Johnson, Phys. Rev. E 72, 046106共2005兲.
关40兴 S. Bianco, P. Grigolini, and P. Paradisi, Chem. Phys. Lett. 438, 336共2007兲.
关41兴 S. Bianco, P. Grigolini, and P. Paradisi, J. Chem. Phys. 123, 174704共2005兲; P. Grigolini, The Physical Roots of Complex-ity: Renewal or Modulation?, in Complexity, Metastability and Nonextensivity, edited by C. Beck, G. Benedek, A. Rapisarda, and C. Tsallis 共World Scientific, Singapore, 2005兲, p. 72; P. Allegrini, F. Barbi, P. Grigolini, and P. Paradisi, Phys. Rev. E
73, 046136共2006兲.
关42兴 H. Haken, Int. J. Psychophysiol 60, 110 共2006兲.
关43兴 S. Bianco, E. Geneston, P. Grigolini, and M. Ignaccolo, e-print arXiv:cond-mat/0611035.
关44兴 M. Buiatti, D. Papo, P.-M. Baudonniére, and C. van Vreeswijk, Neuroscience共to be published兲.
关45兴 J. W. Yuan, B. Zheng, C. P. Pan, Y. Z. Wu, and S. Trimper, Physica A 364, 315共2006兲.
关46兴 R. Cakir, P. Grigolini, and A. Krokhin, Phys. Rev. E 74, 021108共2006兲.
关47兴 P. Winsor, Automated Music Composition 共University of North Texas Press, Denton, Texas, 1992兲, p. 145.
关48兴 P. Winsor, Chaos, Solitons Fractals 20, 45 共2004兲.
关49兴 I. Xenakis, Formalized Music: Thought and Mathematics in Composition共Pendragon Press, Stuyvesant, NY, 1992兲. 关50兴 R. Zatorre, Nature 共London兲 434, 312 共2005兲.
关51兴 G. Bel and E. Barkai, Phys. Rev. Lett. 94, 240602 共2005兲. 关52兴 P. Allegrini, G. Ascolani, M. Bologna, and P. Grigolini, Phys.
Rev. Lett共to be published兲; e-print arXiv:cond-mat/0602281. 关53兴 L. Gammaitoni, P. Hänggi, P. Jung, and F. Marchesoni, Rev.
Mod. Phys. 70, 223共1998兲.
关54兴 M. Baiesi, M. Paczuski, and A. L. Stella, Phys. Rev. Lett. 96, 051103共2006兲.
关55兴 U. Harder and M. Paczuski, Physica A 361, 329 共2006兲. 关56兴 C. Beck and E. Cohen, Physica A 322, 267 共2003兲. 关57兴 C. Tsallis, J. Stat. Phys. 52, 479 共1988兲.
关58兴 H. J. Jensen, Self-Organized Criticality, Cambridge Lecture Notes in Physics 共Cambridge University Press, Cambridge, MA, 1998兲.
关59兴 D. R. Chialvo and P. Bak, Neuroscience 90, 1137 共1999兲. 关60兴 K. Wood, C. Van den Broeck, R. Kawai, and K. Lindenberg,