行政院國家科學委員會專題研究計畫 成果報告
大腸桿菌忌熱性腸毒素 B 次單位蛋白質誘發細胞分化機制及
其應用於下痢防治之研究(2/2)
計畫類別: 個別型計畫 計畫編號: NSC91-2313-B-039-003- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 中國醫藥大學中國醫學研究所 計畫主持人: 侯庭鏞 計畫參與人員: 項千芸, 吳世祿 報告類型: 完整報告 處理方式: 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢中 華 民 國 92 年 10 月 30 日
中 文 摘 要 大 腸 桿 菌 下 痢 症 主 要 是 由 大 腸 桿 菌 產 生 的 忌 熱 性 腸 毒 素 ( heat-labile enterotoxin; LT)所造成的。LT 由 A 及 B 次單位蛋白質(LTA;LTB)組成,LTB 與腸上皮受體 GM1 ganglioside 結合後,LTA 方插入細胞膜中,干擾 G 蛋白質, 使水及離子流失,造成下痢,所以 LTB 能否與腸上皮特定受體結合,為大腸桿 菌下痢症最主要的致病步驟。因此本計畫以大腸桿菌 LTB 為基礎,設計一預防 及有效治療腸毒素下痢的疫苗及藥物,並進而探討毒素與細胞之關係,以拓展 LTB 在其他生物醫學之應用。經由聚合 連鎖反應,我們可增殖出長約 1147 bp 的 LT 基因,並在選殖至表現載體後,於三小時內,自 1 公升的菌液中,純化出 1.3 公克具功能性的重組型 LT 及 LTB。LT 及 LTB 先經 GM1酵素連結免疫吸附試 驗及西方墨點法分析,證實重組型 LT 及 LTB 仍具有與 ganglioside 結合的能力及 仍保有其抗原性。進一步將重組型 LTB 應用在下列三部份的實驗中:一、LTB 作為大腸桿菌疫苗可行性之探討。LTB 利用口服方式免疫老鼠,在血清、肺及腸 道中可以誘發高量的 IgA 反應,顯示 LTB 在不需要經過化工包裝的過程下,可 以直接口服,在肺及腸道引發良好的黏膜性免疫反應。二、抗腸毒素下痢分子模 式之建立及其應用於藥物搜尋之研究。我們利用 LTB 與 GM1接合為基礎,設計
competitive binding assay,發現目前應用在治療下痢的乾薑具有顯著抑制 LTB 與
GM1 結合的能力,此外在動物模式中也呈現相同的結果。三、LT 與細胞相互關
係之研究。我們利用實驗室已構築好的重組細胞株,觀察 LT 及 LTB 對轉錄因子 activator protein 1(AP-1)及 nuclease factor-κB(NF-κB)活性的影響,結果顯 示,LTB 似乎可以藉著調控 NF-κB 的活性,影響免疫反應的發生。本計畫的執
行,除了探討利用 LTB 作為疫苗的可行性外,也發展出以 LTB 與 GM1接合為基
礎的抗下痢藥物篩選模式,並藉助 LT 對轉錄因子的影響,瞭解毒素與細胞的相 互作用。
英 文 摘 要
Heat- labile enterotoxin (LT) from enterotoxigenic Escherichia coli is the major virulence factor of piglet's diarrhea. LT is composed of a single A subunit (LTA) and five identical B subunits (LTB). After being released into the jejunum by the bacteria, the LTB recognizes the oligosaccharide portion of GM1 ganglioside molecules on the
surface of epithelial cells, LTA sequentially ADP-ribosylates the Gs protein and results in a massive efflux of fluids from intestinal cells. Thus, the binding of GM1 by
LTB is the initial step of pathogenesis for E. coli-induced diarrhea. In order to develop the anti-enterotoxic diarrhea drug and vaccine, the LT gene was amplified by polymerase chain reaction. LT was further expressed by prokaryotic expression system and purified by immobilized D-galactose resin. The amount of purified protein was recovered approximately 1.3 g from 1 L culture in a 3-h operation. The recombinant LT exhibited a similar antigenicity and GM1-bindind ability as wild-type
LT did. The recombinant LTB was further immunized to mice by an oral route. In order to screen the anti-diarrhea drugs, the LTB/GM1-based competitive assay was
developed. Our results showed that ginger, which was used to treat diarrhea in traditional Chinese medicine, exhibited the competitive effect with LTB. Further study showed that LT and LTB activated the nuclear factor-κB(NF-κB), which played an important role in inflammation. Taken together, the better understanding of the LTB provides a better way in developing the vaccine, bioadjuvant, immune modulator, and drug delivery system in the future.
目 錄 中文摘要---I 英文摘要---II 前言---1 研究目的---2 文獻探討---3 研究方法---7 結果與討論---10 參考文獻---13 計畫成果自評---17 圖表---18
前 言
豬隻的下痢症是養豬業界一種相當重要的疾病,其中大腸桿菌是造成新生 仔 豬 及 哺 乳 期 仔 豬 下 痢 最 重 要 的 病 因 。 腸 毒 性 大 腸 桿 菌 ( enterotoxigenic
Escherichia coli)在吸附到小腸黏膜上皮後,可產生腸毒素,導致小腸上皮細胞
內水份及電解質的流失,最後造成下痢。腸毒性大腸桿菌所產生的腸毒素主要為 忌熱性腸毒素(heat- labile enterotoxin;LT),LT 是一個高分子量的複合物,由一 個 A 次單位(LTA)及五個 B 次單位(LTB)以非共價鍵結的方式組成,其中 LTB 對 存 在 於 所 有 細 胞 表 面 的 glycosphingolipid — GM1 ganglioside ( Gal( β
1-3)GalNAc( β 1-4)(NeuAc( α 2-3))Gal( β 1-4)Glc( β 1-1)ceramide ) 具 有 親 合 性 (Holmgren et al. 1993),而 LTA 具有 ribosyltransferase 的活性,在進入腸細胞質 後,會將 Gsα ribosylation,使 Gsα失去 GTPase 的活性,進而活化 adenylate cyclase,
使得 cyclic AMP(cAMP)增加。增加的 cAMP 會活化 protein kinase A,將 Cl-
channel 磷酸化,而使得氯離子及水分子由腸細胞中流出,造成下痢的發生(Hirst, 1995)。 LT 除了具有作為大腸桿菌疫苗的潛能外,另外一個重要的生物功能是可充 當口服疫苗的重要佐劑成份。LTB 調節免疫反應的機制,可能是 LTB 與接受器 結合後,會導致白血球不同的效應。LTB 與接受器結合後,會誘發巨噬細胞及淋 巴球 CD25(IL-2Rα)的表現,這種誘發與 CD8+ T 細胞的 apoptosis、B 細胞表 面 MHC class II 及 CD4+ T 細胞 CD40L(gp39)的大量表現有關。CD8+ T 細胞 數量的下降,會造成 IFN-γ減少,而抑制 Th1-type cytokine 的釋放,至於 T 與 B 細胞之間的作用,會導致 B 細胞 B7、CD4、ICAM-1 及 CD4+
T 細胞 CD40L 的 增加,因而增加 Th2-type cytokine 的產生(Williams et al., 1999)。
研 究 目 的 為了能有效控制下痢症,同時為了研究細菌毒素與細胞之相互關係,本計 畫以基因工程方法選殖大腸桿菌 LT 基因、大量產製並純化其蛋白質,進一步進 行蛋白質特性分析及免疫反應的研究。因為 LTB 能否與腸上皮特定受體結合, 為大腸桿菌下痢症最主要的致病步驟,因此本計畫以大腸桿菌 LTB 為基礎,設 計一預防及有效治療腸毒素下痢的疫苗及藥物,並進而探討毒素與細胞之關係, 以拓展 LTB 在其他生物醫學之應用。
文 獻 探 討
豬隻的下痢症是養豬業界一種相當重要的疾病。下痢症可分成三種:新生 仔豬下痢、哺乳期仔豬下痢、及離乳後豬隻下痢,其中大腸桿菌是造成新生仔豬 下痢及哺乳期仔豬下痢最重要的病因。在適當的環境及宿主因素下,腸毒性大腸 桿菌(enterotoxigenic Escherichia coli)可藉著特定的致病因子,在小腸上皮增殖,
並導致下痢的發生。大部份腸毒性大腸桿菌可以產生一到數種的線毛,這些線毛 可吸附在小腸的上皮細胞,使大腸桿菌在小腸內增殖。線毛依不同血清反應性及 受體特異性而有不同的區分,目前已有超過三十種以上的線毛被證實(Tacket et al., 1999)。在傳統自家疫苗及一般活菌、死菌菌苗大規模的使用下,易造成腸毒 性大腸桿菌表面線毛的變異,產生新的突變株,而使得疫苗的效力有一定的波動 現象(Siev, 1997)。所以如何以現今對大腸桿菌致病機轉的了解為基礎,設計一 預防及有效治療腸毒素下痢的疫苗及藥物,是一重要之思考方向。 腸毒性大腸桿菌在吸附到小腸黏膜上皮後,可產生腸毒素,導致小腸上皮 細胞內水份及電解質的流失,最後造成下痢。腸毒性大腸桿菌所產生的腸毒素主 要為忌熱性腸毒素(heat- labile enterotoxin;LT),LT 是一個高分子量的複合物, 由一個 A 次單位(LTA)及五個 B 次單位(LTB)以非共價鍵結的方式組成,其 中 B 次單位對存在於所有細胞表面的 ganglioside 具有高親合性(Holmgren et al. 1993),而 A 次單位為產生毒素效用的單位(de Haan et al. 1996a)。腸毒性大腸 桿菌的 LT 與來自於霍亂弧菌(Vibrio cholerae)的霍亂毒素(cholera toxin;CT)
具有高度的同源性,以胺基酸序列分析,其相似度高達 82﹪。而大腸桿菌的 LT 與其他腸炎致病菌如赤痢志賀桿菌(Shigella dysenteriae)的腸毒素,在核 酸 序列、胺基酸序列、及作用機制方面,也都相當的相似( Spangler, 1992;Ruddock et al., 1996),因此發展大腸桿菌類毒素疫苗,除了可大幅改善現有大腸桿菌菌苗 的效力外,也可有效地交叉中和其他細菌的腸毒素,使得下痢症的發生率能達到 有效的控制。 腸毒性大腸桿菌的忌熱性腸毒素 LT 與霍亂弧菌的霍亂毒素 CT 這兩種毒素 都是由一個 A 次單位及 5 個 B 次單位所組成,其中 A 次單位可被蛋白質分解 切割成 A1及 A2兩個片段,這兩片段彼此間以雙硫鍵鍵結(Merritt et al., 1994;
Sixma et al., 1993)。A1片段具有 ribosyltransferase 的活性,在進入腸細胞質後,
會對 Gsα進行 ADP-ribosylation,Gsα因而失去 GTPase 活性,進而活化 adenylate
cyclase,使得 cyclic AMP(cAMP)增加。增加的 cAMP 會活化 protein kinase A, 將 Cl
channel 磷酸化,而使得氯離子及水分子由腸細胞中流出,因而造成下痢的 發生(Hirst, 1995)。A2片段主要作為與 B 次單位連結的橋樑,此外,A2片段的
C 端具有 KDEL motif(在 LT 為 RDEL motif),這段序列與蛋白質由trans-Golgi
被傳送至內質網有關,因此可能可以協助 A1運送到正確的位置,在調節毒素的
免疫反應上,扮演重要角色(Pelham, 1992)。
所形成的 pentamer 是相當穩定的結構,例如在 1 mg/ml 胰蛋白 或蛋白質分解 的切割下,或是極酸的環境下(CT 為 pH 3.9,LT 為 pH 2.0),都無法破壞 pentamer 的結構(Ruddock et al., 1995)。B 次單位的功能是與細胞接受器結合,CT 及 LT 主 要 的 接 受 器 為 廣 泛 地 存 在 於 哺 乳 類 細 胞 表 面 的 glycosphingolipid — GM1
ganglioside ( Gal( β 1-3)GalNAc( β 1-4)(NeuAc( α 2-3))Gal( β 1-4)Glc( β 1-1)ceramide)(Hol et al., 1995),此外,CT 及 LT 也會與 GD1b ganglioside 結合,
而 LT 則尚會與 asialo-GM1、lactosylceramide 及某些特定的 galactoproteins 結合
(Fukuta et al., 1988;Holmgren et al., 1982)。這幾種接受器與 LT 或 CT 結合的 親和力各不相同,因此 LT 及 CT 是否可藉由不同的結合特異性,影響其與細胞 之相互作用,甚至有調節細胞分化及免疫調控的功能,仍待進一步的釐清。 LT 除了可充當大腸桿菌疫苗的標的外,另外一個重要的生物功能是可充當 口服疫苗的重要佐劑成份。佐劑(adjuvant)是能增強免疫反應的物質,可大幅 增加抗原所誘發的免疫反應。許多研究顯示佐劑可影響抗體反應的時間性、親合 性,此外,佐劑也可增加細胞性免疫反應的發展,例如延遲性過敏反應及細胞毒 殺反應等。目前經常使用的佐劑包括鋁膠、油質佐劑等,它們已經被應用於疫苗 的使用上數十年,這些佐劑在注射位置易造成肉芽腫的缺點,此外,因為這些佐 劑的成份或結構太複雜,使得作用機制仍不是很清楚,而無法有效地評估其免疫 機轉。近年來,因遺傳工程技術的進步,許多新一代的疫苗可利用病原的重組蛋 白質、peptide、或是 DNA 作為疫苗型式,但是一般而言,這些重組蛋白或 peptide 單獨使用的免疫原性都不佳,因此為了有效地增加疫苗刺激免疫反應的效果,並 導引疫苗產生特定的免疫反應,有必要開發新一代的佐劑,使得疫苗的免疫反應 更完整,投與途徑更為多樣化。 具 有 佐 劑 功 能 的 生 物 性 分 子 ( biomolecule ) 也 稱 為 生 物 性 佐 劑 (bioadjuvant),生物性佐劑包括目前較為人所知的細胞素(cytokine)及細菌毒 素。細胞素在疫苗的應用方面,具有可以產生適當的免疫反應並導致保護性免疫 反 應 的 特 性 。 許 多 研 究 顯 示 , 細 胞 素 ( 例 如 interleukin 2 、 interferon 及 granulocyte-macrophage colony stimulating factor 等)可以促進 B 細胞及 T 細胞的 活化、增殖、分化。除了量的增加外,細胞素也可誘導特定抗體型式的產生,以 產生更好的保護性。至於在細菌毒素方面,因為大部份的抗原在以黏膜途徑投與 時,其免疫原性都相當差,並且易造成免疫耐受性,此時黏膜佐劑就可克服這個 缺點,輔助抗原被 M 細胞吸收,誘導黏膜性免疫反應的產生(Kagnoff, 1996; McDermott and Bienenstock, 1979)。細菌毒素,包括 CT 及 LT 等,都具有作為黏 膜佐劑的潛能(Komase et al. 1998)。
LT 作為黏膜免疫性佐劑的效用與 A 次單位的 ADP ribosyltransferase 活性有 關(de Haan et al. 1996a),而 B 次單位則可產生 immunological carrier 的效用 (Holmgren et al. 1993)。早在 1972 年,就有報告指出 CT 在靜脈注射後可作為 佐劑引發抗體反應(Northrup and Fauci, 1972),然而直到 1984 年,Elson 及 Ealding 發現 CT 經由口服接種後也具有佐劑的效果(Elson and Ealding, 1984),才顯現
CT 作為免疫學工具的重要性,而這項發現也使得疫苗學家得以將其應用於增加 口服抗原的免疫原性。早期的研究以 CT 作為佐劑使用時,其需使用遠低於造成 老鼠小腸液體流失的劑量(0.5 μg),以避免 CT 的毒性。之後的研究發現,在 這種劑量下,如果再加入 10 μg 純化的 CTB,則可加強 CT 的佐劑效果( Wilson et al., 1990)。若將 keyhole limpet haemocyanin(KLH)與 CT 共同給予動物口服 後,可同步引發抗體及 T 細胞反應(Clarke et al., 1991),T 細胞反應主要與 T helper 2(Th2)-associated cytokines 有關(Wilson et al., 1991),而這項發現與血清中抗 體主要為 IgG1、高量的分泌型 IgA 被產生(Marinaro et al., 1995;Yamamoto et al., 1997a;Vajdy and Lycke, 1992)及 interleukin-4 剔除鼠的反應顯著下降等結果相 符(Vajdy et al., 1995)。至於 LT 在黏膜接種後,所引發的免疫反應較 CT 來得平 均,LT 可同步誘發 Th1、Th2 cytokines 的產生(Takahashi et al., 1996),此外抗 體反應持續的時間性較久,而且在人類及實驗動物上都呈現相同的效果(Vajdy and Lycke, 1992;Hashigucci et al., 1996)。
CT 及 LT 作為佐劑的功能是來自於 A 或 B 次單位的研究,是相當受到爭議 的。將 LTA 的活性中心進行定點突變後,其佐劑效果也隨之降低,顯示 LT 的佐 劑活性是來自於 A 次單位的 ADP-ribosylation(Lycke et al., 1992)。然而,其他 研究發現,缺少 ribosyltransferase 活性的 LT 或 CT 仍保有其佐劑效果,又說明 A 次單位與佐劑的功能無關( Yamamoto et al., 1997a;Douce et al., 1995;Yamamoto et al., 1997b;de Haan et al., 1996b;Douce et al., 1997)。而且,當一篇報告陳述 重組型的 LTB 或 CTB 無法作為佐劑,彰顯 A 次單位的重要性後(Douce et al., 1995),其他相反結果的報告也隨之出現(Douce et al., 1997;deHaan et al., 1998)。 目前認為,A 及 B 次單位對於佐劑的活性有不同的影響,A 次單位可能可以藉 由穩定 B 次單位的結構,或是與 KDEL 接受器結合,影響毒素於細胞內的運送, 而調節免疫反應。至於 B 次單位,則可藉由與細胞接受器的結合,誘發主要的 佐劑效果。 LT 具有免疫調節的功能,可能來自於下列不同機制:可增加抗原被 M 細胞 攝入的機會、增強巨噬細胞及上皮細胞的抗原呈現能力、增加抗原呈現細胞釋放 interleukin 1 及 interleukin 6、增加 T 細胞反應的分化(誘發產生 Th1、Th2 的反 應)、及刺激 B 細胞 isotype switching 等功能(Takahashi et al. 1996;Marchetti et al. 1998;Truitt et al. 1998)。這些特性,可以輔佐疫苗在經黏膜途徑投與時,產生 良好的免疫反應。因此應用細菌毒素作為黏膜性佐劑的研究,將可改善目前針劑 疫苗的缺點,引發良好的黏膜性免疫反應,而有效地防範胃腸道及呼吸道病原的 感染,使得疫苗的免疫反應更為完整,投與途徑更加多樣化。最近的研究指出, 重組型的 LTB 及 CTB 可作為免疫耐受調節者,強化耐受性(Sun et al., 1994;Sun et al., 1996;Bergerot et al., 1997;Williams et al., 1997)。將 CTB 與 myelin basic protein 或胰島素結合後,可以治療大白鼠人工誘發的過敏性腦脊髓炎或 nonobese diabetic mouse 的糖尿病(Sun et al., 1996;Bergerot et al., 1997)。雖然這些發現 並沒有非常精確的機制可以解釋,但將抗原與 B 次單位結合的必要條件下,顯
示 B 次單位可以作為攜帶者,將抗原藉由 GM1接受器,送至誘發耐受性的途徑。 LTB 的免疫原性主要來自與接受器的作用(Nashar et al., 1996)。將小白鼠 以皮下或口服方式接種 LTB, 可引發高量的抗體反應,若 將 LTB 突變株 LTB(G33D)(已失去與 GM1結合的能力)皮下接種小白鼠,只會產生低量的抗體 反應,而在口服後,甚至不能偵測到抗體反應的產生。由試管內分析,B 次單位 可以干擾淋巴結內淋巴球的種類,減少 CD8+ T 細胞的數目,增加 B 細胞的比例。 若將 LTB 及 CTB 加入淋巴細胞中,會造成 CD8+ T 細胞的 apoptosis,這種現象 與 IL-2 receptor α chain(IL-2Rα)有關,而與 Fas 或 tumor necrosis factor α 無關。相似的結果也出現在 CTB 以口服方式後的反應,於 Peyer’s patch 中可發 現 CD8+ T 細胞的減少。LT 與接受器之間的作用對其他白血球族群的影響,目前 並不清楚。CD4+ T 細胞不會發生 apoptosis,但 LT 與接受器之間的作用可以藉由 影響細胞週期,干擾 CD4+ T 細胞的增殖,也會改變 CD4+ T 細胞的 cytokine profile (Bromander et al., 1991)。 LTB 作為免疫調節者的機制,目前認為是在 LTB 與接受器結合後,會導致 白血球族群不同的效應。LTB 與接受器結合後的直接效應,包括巨噬細胞及淋巴 球族群 CD25(IL-2Rα)的誘發,這種誘發與 CD8+ T 細胞的 apoptosis、B 細胞 表面 MHC class II 及 CD4+ T 細胞 CD40L(gp39)的大量表現有關。巨噬細胞的 活化與 IL-10 的釋放有關。CD8+ T 細胞數量下降造成 IFN-γ減少會降低 Th1 cytokine 的釋放,而 T 與 B 細胞之間的作用,會導致 B 細胞 B7、CD4、ICAM-1 及 CD4+
T細胞 CD40L的增加,而增加 Th2 cytokine 的產生(Williams et al., 1999)。 所以由大腸桿菌的 LT 與細胞之相互關係著手研究,將可以對 LT 與免疫調 節及其它生物醫學的應用有更進一步的了解。以細胞層次而言,LT 可影響腸細 胞與淋巴細胞之增殖、分化、與 apoptosis,所以假如可以 LT 對細胞增殖、分化、 與 apoptosis 相關之訊息傳導及基因轉錄調控作為分析起點,應會對 LT 與細胞之 相互關係的機制有更完整的見解,未來才有可能開發 LT 更新之生物醫學應用。
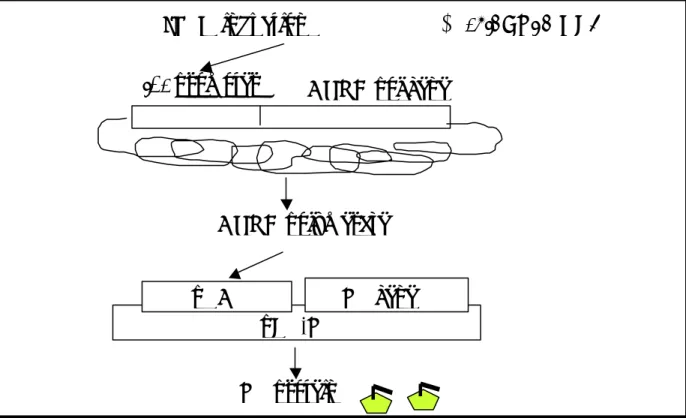
研 究 方 法 一、大腸桿菌 LT 基因及其次單位基因之選殖、表現及功能性分析 (一)大腸桿菌忌熱性腸毒素 A 次單位及 B 次單位基因的選殖 取腸毒性大腸桿菌(enterotoxigenic E. coli)於 37℃增殖培養後,萃取其 DNA,並以設計好的引子(分別位於忌熱性腸毒素 A 次單位及 B 次單位基因 5’端及 3’端),進行 PCR。經增殖後的片段,利用 T-A cloning 的方法,選殖 到質體中,最後經核 酸定序確認(圖一)。 (二)大腸桿菌忌熱性腸毒素 A 次單位及 B 次單位基因之表現及純化 將上述方法所選殖之大腸桿菌忌熱性腸毒素 A 次單位及 B 次單位基 因,利用適當的限制酵素切割,再架接進原核表現載體 pET-28(+)中。經選殖、 定序無誤後,進行下列蛋白質的產製。為了取得大量而且純度很高的忌熱性 腸毒素 A 次單位及 B 次單位蛋白,我們擬利用大腸桿菌表現系統進行蛋白質 的大量產製。將構築好的表現質體送入大腸桿菌 BL21(DE3)pLysS 中,這一 株大腸桿菌內帶有一個嵌在染色體上的 T7 RNA 聚合酵素基因,其表現受 lacUV5 啟動子控制,因此可以用異丙基-β-D-硫代半乳糖 (isopropyl-β -D-thiogalactopyranoside;IPTG)誘導 T7 RNA 聚合酵素的表現,進而大量轉 錄 pET 質體上所帶的基因,產生 RNA 及進一步製造蛋白質。此外,菌體內 帶有一個 T7 溶解酵素( lysozyme)基因的質體,T7 溶解酵素可抑制 T7 RNA 聚合酵素的活性,防止少量的 T7 RNA 聚合酵素在 IPTG 誘導前提早表現外 來的蛋白質,而對細菌產生毒性或抑制其生長(圖二)。表現出來的蛋白質進 一步以D-galactose resin,以親和性色層分析法進行純化。
(三)GM1-酵素連結免疫吸附分析法(GM1-Enzyme- linked Immunosorbent Assay;
GM1 -ELISA)
因忌熱性腸毒素對細胞表面醣脂 GM1 ganglioside 具有極高的親合性
(Spangler, 1992),因此可利用 GM1-ELISA 判定其功能及特性。先將 200 ng
GM1(購自 sigma)經 37 ℃,16 小時固定於 ELISA plate 後用 1% bovine serum
albumin(BSA)遮蔽非特異性結合部位,接著加入純化過且標示 biotin 的 LTB 及 LT 與 GM1進行特異性結合,每做一步驟皆需以 PBS 洗去未結合物,最後 ABTS 經 peroxidase 催化後會產生綠色的受質可在 405nm 測出波長,進而定 出 LTB 及 LT 與 GM1的結合比率。 (四)西方雜合法 將經純化後之 LT 及 LTB 利用蛋白質電泳分析後,轉漬到硝基纖維濾紙 上,再以免疫 LT 的老鼠血清作用,最後加入二次抗體及受質進行呈色。 二、LTB 作為大腸桿菌疫苗可行性之探討 (一)疫苗免疫試驗 這部份是以 4∼5 週齡的小白鼠做為實驗對象,進行大腸桿菌忌熱性腸
毒素 B 次單位蛋白質的接種試驗,以初步評估 B 次單位蛋白質的免疫效果。 將大腸桿菌 B 次單位蛋白質,藉由口服及注射的方式,進行疫苗的接種。於 免疫後不同時間,收集血清及腸洗液,以進行下列免疫反應測定。血清的收 集方法為:由採血管所收集到的血液先平放在 37℃、30 分鐘後,再置於 4℃ 的冰箱中 18 小時,使血液充分凝固收縮後,倒出血清,並在 4℃下以 3000 xg 離心 20 分鐘後收集血清,保存在-20℃備用。腸洗液的收集方法為:採得的 小腸馬上以清水沖洗外部擦乾後,自一端灌入已滅菌好的 PBS,用手將緩衝 液擠到小腸的末端後將腸洗液倒出至 50 ml 的離心管中,以超音波振盪 30 分 鐘後,初步離心 1000 rpm、10 分鐘,取上清液再以 25000 xg 離心 30 分鐘。 上清液再以過濾膜孔徑為 50 kDa 的濃縮離心管濃縮至原體積之 1/10,保存於 -20℃備用。 (二)疫苗免疫反應 全身性及黏膜性免疫反應利用 ELISA 的方式加以評估,以分析抗腸毒素 之 IgG 及 IgA。將 ELISA 平盤先以忌熱性腸毒素抗原吸附後,再加入免疫後 的小白鼠血清或黏膜沖洗液,進行接合反應,最後加入 peroxidase 標示的二 次抗體,利用呈色劑加以呈色,以測定抗體的強度及種類,達到完成大腸桿 菌類毒素疫苗的效力評估。 三、抗腸毒素下痢分子模式之建立及其應用於藥物搜尋之研究 (一)甲醇中藥萃取 將購自中藥行的中藥以 9 克與 30 ml 甲醇的比例做混合,均勻搖晃 48 小時做萃取,離心後取上清液以 0.02 nm 濾網做過濾,取 1 ml 稱乾重其餘存 於–80 ℃待大量搜尋時使用。 (二)競爭性 GM1-ELISA
先將 200 ng GM1(購自 sigma)經 37 ℃,16 小時固定於 ELISA plate 後
用 1% bovine serum albumin(BSA)遮蔽非特異性結合部位,接著將不同濃 度的 GM1與 LTB 於室溫下作用 2 小時,再將混合物加入已吸附 GM1之 ELISA
平盤上,每做一步驟皆需以 PBS 洗去未結合物,最後 ABTS 經 peroxidase 催
化後會產生綠色的受質可在 405 nm 測出波長,進而定出 LTB 與 GM1的結合
比率。
(三)Fluid accumulation assay
利用 LT 毒素會造成生物體腸壁離子通透性失調之原理,使用 female Balb/Cby mice (4-8 weeks,20-25 g 購自國科會),手術前將老鼠飢餓 24 小時, 再以 ketamine/xylazine(2/1 的比例稀釋混合)做腹腔麻醉,在 mice 空腸及迴 腸之間做一小段結紮(約 4~5 cm),再以 28 號針頭將 0.1 μg LT/or herbs(total volume=200 μl)打入結紮部位,使腸液蓄積在 loop 內,24 小時後將老鼠犧 牲並取下結紮部位以 g/cm 的方式計算其 loop 腫脹程度(Hiroshi et al., 2002)。
四、大腸桿菌忌熱性腸毒素與細胞相互關係之研究 (一)重組細胞的構築
為了架構 AP-1/Luciferase 及 NF-êB/Luciferase 報導基因重組細胞株,我 們進行 stable transfection。實驗方法是,將細胞繼代至 25T flask,使細胞於 24 小時內長至約 8 分滿之後,將報導基因質體和具有 neomycin 抗藥性的 pSV3-neo 質 體 分 別 利 用 限 制 AlwN Ⅰ和 EcoRⅠ切成直線形,再利用 SuperFect 進行 co-transfection。Transfection 後 48 小時,以 1:15 的比例繼代 至新的 flask 中,24 小時之後更換含有 400 ng/ml Geneticin(G418)的新鮮培 養液進行篩選。4~7 天後,將細胞繼代至 24 well plate 中,持續給予 400 ng/ml G418 的培養液進行選殖。最後自 24 well plate 中挑選出單一具有抗藥性的 clone,利用 luciferase assay 篩選出 luciferase 活性最強的為重組細胞株。 (二)Luciferase assay
以不同因子刺激重組細胞,測定細胞中 luciferase 的量,以瞭解細胞中 AP-1 及 NF-êB 的活性。依照 Luciferase Assay System(Promega)之說明,取 得細胞萃取物後,以 FB15 luminometer測量 luciferase 的活性,其結果以relative light unit(RLU)來表示。
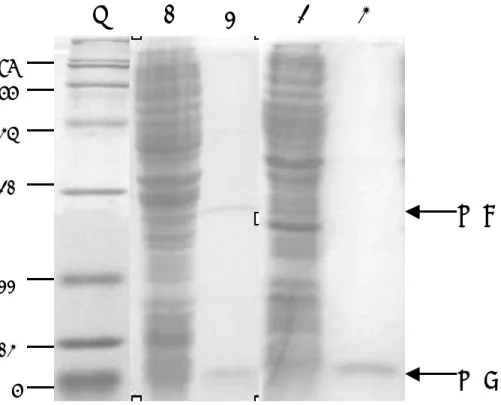
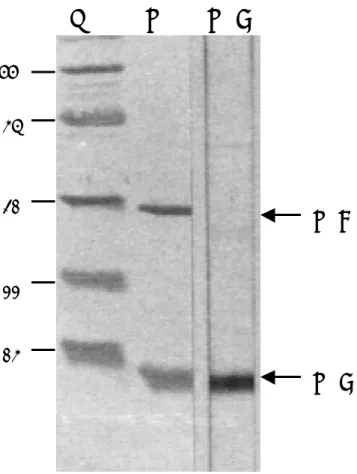
結 果 與 討 論 一、大腸桿菌 LT 基因及其次單位基因之選殖、表現及功能性分析 取腸毒性大腸桿菌於 37℃增殖培養後,萃取其質體 DNA,並以位於 LT 基 因 5’ 端 及 3’ 端 的 一 對 引 子 ( 5’-CGCGGATCCCGATGAAAAATATAAC-3’ ; 5’-ACTGCAGCTAGTTTTTCATACTGATTGCC-3’),進行聚合 連鎖反應。經增 殖後的片段,利用設計於引子上的酵素BamHI 及 XhoI 切割,選殖到原核表現載 體 pET-28(+)中,成為 pET-LT。為了取得大量而且純度很高的 LT,我們利用大 腸 桿 菌 表 現 系 統 進 行 蛋 白 質 的 大 量 產 製 。 將 pET-LT 送 入 大 腸 桿 菌 BL21(DE3)pLysS 中,這一株大腸桿菌內帶有一個嵌在染色體上的 T7 RNA 聚合 基因,其表現受 lacUV5 啟動子控制,因此可以用異丙基-β-D-硫代半乳糖 (isopropyl-β-D-thiogalactopyranoside;IPTG)誘導 T7 RNA 聚合 的表現,進 而大量轉錄 LT 基因,產生 RNA 及進一步製造蛋白質。在經 IPTG 誘導的狀況下, 我 們 可 以 將 細 菌 打 破 , 取 含 有 忌 熱 性 腸 毒 素 的 上 清 液 , 利 用 immobilized D-galactose 親合性色層分析樹脂加以純化(圖三)。在三小時的操作時間內,可 自 1 公升細菌液中,純化出約 1.3 公克之 LT,其純度至少 99﹪。 進一步為了瞭解重組型 LT 及 LTB 的抗原性及功能方面是否與野外型相 同,我們進行西方墨點法及 GM1酵素連結免疫吸附試驗分析。將野外型與重組型 LT 利用蛋白質電泳分析後,轉移至濾紙上,以野外型 LT 所產製的抗體偵測,再 利用標示酵素的二次抗體及受質加以呈色,結果發現抗體可以成功地偵測到野生 型及重組型 LT 及 LTB,顯示重組型 LT 及 LTB 仍保持其抗原性(圖四)。另外, 利用 GM1酵素連結免疫吸附試驗測定並比較野外型與重組型 LT 結合 GM1的能 力,結果顯示隨著蛋白質濃度的增加,吸光值也隨之增加,顯示野外型與重組型 LT 及 LTB 都具備與 GM1結合的能力,而且結合的動力學也非常相似(圖五)。 因此,利用大腸桿菌大量產製的重組型 LT 及 LTB 具備與野外型 LT 相同的特性, 並具有容易生產、容易純化的特性,可以應用於以下的研究。 二、LTB 作為大腸桿菌疫苗可行性之探討 因為 LTB 能否與腸上皮特定受體結合,為大腸桿菌下痢症最主要的致病步 驟,因此我們將 LTB 以口服及注射方式免疫小白鼠,觀察是否可以誘發小白鼠 產生黏膜性或全身性免疫反應,並產生良好的保護性。在本項研究中,我們已將 小白鼠區分為三組進行免疫注射,實驗組為 LTB 口服組、陽性對照組為 LTB 經 傳統油質佐劑混合後之針劑注射組、陰性對照組為生理食鹽水口服組。實驗組每 隻小白鼠給予 12 μg(約 50 μl)LTB;陰性對照組,每隻小白鼠給予約 50 μ l 生理食鹽水;陽性對照組,每隻小白鼠注射 10 μg(約 100 μl)LTB 與油質 佐劑混合之膠體粒子。每組共免疫兩次,間隔為 2 週,最後一次免疫後 2 週,將 動物犧牲,收集血清、肺洗液及腸洗液,再藉由酵素連結免疫吸附試驗偵測 IgA 及 IgG 的濃度。在小白鼠試驗中發現,經由口服的途徑,在血清、肺及腸道中可
以誘發高量的 IgA 反應,與陰性對照組比較分別增加 4.3、5.8 及 6.3 倍,而在 IgG 反應方面,則分別增加 1.7、2.9 及 7.7 倍,顯示大腸桿菌 LTB 在不需要經過化工 包裝的過程下,可以直接口服,在肺及腸道引發良好的黏膜性免疫反應。 三、腸毒素下痢分子模式之建立及其應用於藥物搜尋之研究
因為 LTB 能否與腸上皮特定受體結合,為大腸桿菌下痢症最主要的致病步 驟,因此我們利用 LTB 與 GM1接合為基礎,設計 competitive binding assay,搜尋
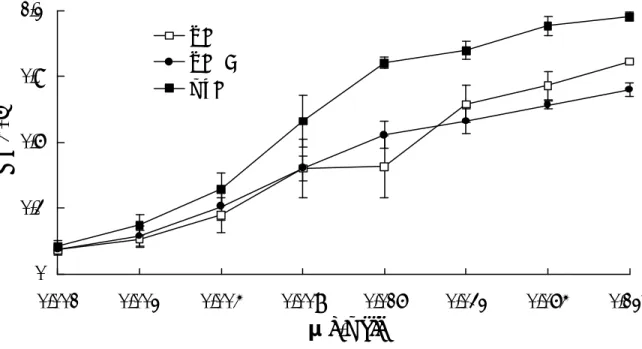
可能抑制下痢之中藥。 為了證明這個實驗的可信度是拮抗劑與受體競爭毒素的模式,首先先建立 標準的 LTB 結合 GM1的劑量曲線。圖七可以發現 LTB 在 0.016 μg/well 時與 200 ng/well GM1的結合達到飽和,接著再利用 GM1進行競爭性 GM1-ELISA 標準曲線 的測定。將不同濃度的 GM1與 LTB 於室溫下作用 2 小時,再將混合物加入已吸 附 GM1之 ELISA 平盤上進行測定,作為陽性對照組的標準曲線(圖六)。結果證 明 GM1能有效競爭游離的 LTB 確立受體與毒素是可被類似結構物競爭的。 由以上可知受體與毒素之間是可被類似結構競爭的,因此我們以甲醇萃取 的中藥為搜尋起點,利用競爭性 GM1-ELISA 分析後發現,有一群中藥的吸光值 曲線較穩定隨中藥濃度增加而下降,將其結果換算成劑量反應,以 50% Inhibitory concentration (IC50)表示,IC50愈低代表此種中藥效果愈好。在此分別以貝母,乾
薑,香薷 IC50較低,皆小於 3mg/ml 即有抑制效果(圖七)。 因為薑科植物常用於做為腸胃道用藥,因此我們選擇乾薑的甲醇萃取液來 做進一步的動物實驗。分別將乾薑萃取液以不同濃度溶於甲醇後,與忌熱型腸毒 素混合後打入已結紮的小白鼠腸腔內,並設立一個甲醇的溶劑控制組,結果顯示 在只給予忌熱型腸毒素的小白鼠腸腔有明顯腫脹,但給予乾薑萃取液與忌熱型腸 毒素混合的組別依藥物濃度的增加其腫脹的程度顯著被抑制下來且成濃度梯度 (圖八),但給予 PBS 的組別沒有改變。證明乾薑萃取液可能是透過干擾忌熱型 腸毒素的活性中心來抑制離子通透性的改變。將剪下的腸段以(Ratio=g/cm)計算 其比率,發現給與 LT 組別誘發約 3.8 倍的腸液堆積在腸腔,乾薑萃取液在動物 體的 IC50約為< 0.24 μg/250 μl。 四、大腸桿菌 LT 與細胞相互關係之研究 目前認為 LTB 調節免疫反應的機制,可能是 LTB 與接受器結合後,導致不 同白血球的增殖或 apoptosis,而引發 Th1 或 Th2-type cytokine 的釋放。當細胞受 到外界刺激時,可藉由 mitogen-activated protein kinase(MAPK)signal transduction pathway 活化轉錄因子 activator protein 1(AP-1),進而調控細胞的生長( Lewis et al., 1998),此外,MAPK 也可以活化另外一類的轉錄因子 nuclear factor-κB(NF-κB),而導致發炎的產生(Baeurele and Baichwal, 1997)。因此,我們利用實驗 室已構築好的重組細胞株,觀察 LT 及 LTB 對 AP-1 及 NF-κB 活性的影響。圖 九顯示,LT 及 LTB 僅會些微的影響 AP-1 的活性,但兩者皆會活化 NF-κB 的活
性,其中又以 LTB 活化的效果最為顯著。此外,LTB 活化 NF-κB 的能力隨濃 度增加而增加,因此由這些結果顯示,LTB 似乎可以藉著調控 NF-κB 的活性, 影響免疫反應的發生。
參 考 文 獻
Auchere, D., Tardivel, S., Gounelle, J.-C., Drueke, T. and Lacour, B. 1998. Role of transcellular pathway in ileal Ca2+ absorption: stimulation by low-Ca2+ diet. Am. J. Physiology 275: G951-956.
Bergerot, I., Ploix, C., Petersen, J., Moulin, V., Rask, C., Fabien, N., Lindblad, M., Mayer, A., Czerkinsky, C., Holmgren, J., and Thivolet, C. 1997. A cholera toxoid- insulin conjugate as an oral vaccine against spontaneous autoimmune diabetes. Nonlinear regression using spreadsheets. Proc. Natl. Acad. Sci. 94: 4610-4614.
Bowen, W.P., and Jerman, J.C. 1995. Trends Pharmacol. Sci. 16: 413-417.
Bromander, A., Holmgren, J., and Lycke, N. 1991. Cholera toxin stimulates IL-1 production and enhances antigen presentation by macrophages in vitro. J. Immunol. 146: 2908-2914.
Clarke, C.J., Wilson, A.D., Williams, N.A., and Stokes, C.R. 1991. Mucosal priming of T- lymphocyte responses to fed protein antigens using cholera toxin as an adjuvant. Immunology 72: 323-328.
Douce, G., Fontana, M., Pizza, M., Rappuoli, R., and Dougan, G. 1997. Intranasal immunogenicity and adjuvanticity of site-directed mutant derivatives of cholera toxin. Infect. Immun. 65: 2821-2828.
Douce, G., Turcotte, C., Cropley, I., Roberts, M., Pizza, M., Domenghini, M., Rappuoli, R., and Dougan, G. 1995. Mutants of Escherichia coli heat-labile toxin lacking ADP-ribosyltransferase activity act as nontoxic, mucosal adjuvants. Proc. Natl. Acad. Sci. 92: 1644-1648.
Elson, C.O., and Ealding, W. 1984. Cholera toxin feeding did not induce oral tolerance in mice and abrogated oral tolerance to an unrelated protein antigen. J. Immunol. 133: 2892-2897.
Fondacaro, J.D. 1986. Intestinal ion transport and diarrheal disease. Amer. J. Physiology 250, G1-8.
Fukuta, S., Magnani, J.L., Twiddy, E.M., Holmes, R.K., and Ginsburg, V. 1988. Comparison of the carbohydrate-binding specificities of cholera toxin and
Escherichia coli heat-labile enterotoxins LT-I, LT-IIa, and LT-IIb. Infect. Immun.
56: 1748-1753
Gabriel, S.E., Davenport, S.E., Steagall, R.J., Vimal, V., Carlson, T., Rozhon, E.J. 1999. A novel plant-derived inhibitor of cAMP- mediated fluid and chloride secretion. Amer. J. Physiology 276: G58-63.
de Haan, L., Feil, I.K., Verweij, W.R., Holtrop, M., Hol, W.G., Agsteribbe, E., and Wilschut, J. 1998. Mutational analysis of the role of ADP-ribosylation activity and GM1-binding activity in the adjuvant properties of the Escherichia coli heat-labile
enterotoxin towards intranasally administered keyhole limpet haemocyanin. Eur. J. Immunol. 28: 1243-1250.
de Haan, L., M. Holtrop, W. R. Verweij, E. Agsteribbe and J. Wilschut. 1996a. Mucosal immunogenicity of the Escherichia coli heat-labile enterotoxin: role of the A subunit. Vaccine 25: 260-266.
de Haan, L., Verweij, W.R., Feil, I.K., Lijnema, T.H., Hol, W.G., Agsteribbe, E., and Wilschut, J. 1996b. Mutants of the Escherichia coli heat-labile enterotoxin with reduced ADP-ribosylation activity or no activity retain the immunogenic properties of the native holotoxin. Infect. Immun. 64: 5413-5416.
Hashigucci, K., Ogawa, H., Ishidate, T., Yamashita, R., Kamiya, H., Watanabe, K., Hattori, N., Sato, T., Suzuki, Y., Nagamine, T., Aizawa, C., Tamura, S., Kurata, T., and Oya, A. 1996. Antibody responses in volunteers induced by nasal influenza vaccine combined with Escherichia coli heat-labile enterotoxin B subunit containing a trace amount of the holotoxin. Vaccine 14: 113-119.
Hirst, T.R. 1995. In “Bacterial Toxins and Virulence Factors in Disease”, Moss, J., Iglewski, B., Vaughan, M., and Tu, A.T., eds, pp.123-184, Marcel Dekker.
Hol, W.G.J., Sixma, T.K., and Merritt, E.A. 1995. In “Bacterial Toxins and Virulence Factors in Disease”, Moss, J., Iglewski, B., Vaughan, M., and Tu, A.T., eds, pp.185-224, Marcel Dekker.
Holmgren, J., Fredman, P., Lindblad, M., Svennerholm, A.M., and Svennerholm, L. 1982. Rabbit intestinal glycoprotein receptor for Escherichia coli heat-labile enterotoxin lacking affinity for cholera toxin. Infect. Immun. 38: 424-433.
Holmgren, J., Lycke, N., and Czerkinsky, C. 1993. Cholera toxin and cholera B subunit as oral- mucosal adjuvant and antigen vector systems. Vaccine 11: 1179-1184.
Kagnoff, M. F. 1996. Mucosal immunology: new frontiers. Immun. Today 17: 57-59. Komase, K., Tamura, S.I., Matsuo, K., Watanabe, K., Hattori, N., Odaka, A., Suzuki,
Y., Kurata, T., and Aizawa, C. 1998. Mutants of Escherichia coli heat-labile enterotoxin as an adjuvant for nasal influenza vaccine. Vaccine 16: 248-254.
Kurkchubasche, A.G., Cardona, M., Watkins, S.C., Smith, S.D., Albanese, C.T., Simmons, R.L., Rowe, M.I. and Ford, H.R. 1998. Transmucosal passage of bacteria across rat intestinal epithelium in the Ussing Chamber: effect of nutritional factors and bacterial virulence. Shock 9: 121-127.
Lycke, N., Tsuji, T., and Holmgren, J. 1992. The adjuvant effect of Vibrio cholerae and Escherichia coli heat-labile enterotoxins is linked to their ADP-ribosyltransferase activity. Eur. J. Immunol. 22: 2277-2281.
Marchetti, M., Rossi, M., Giannelli, V., Giuliani, M.M., Pizza, M., Censini, S., Covacci, A., Massari, P., Pagliaccia, C., Manetti, P., Telford, J.L., Douce, G., Dougan, G., Rappuoli, R., and Ghiara, P. 1998. Protection against Helicobacter
pylori infection in mice by intragastric vaccination with H. pylori antigens is
achieved using a non-toxic mutant of E. coli heat-labile enterotoxin (LT) as adjuvant. Vaccine 16: 33-37.
Marinaro, M., Staats, H.F., Hiroi, T., Jackson, R.J., Coste, M., Boyaka, P.N., Okahashi, N., Yamamoto, M., Kiyono, H., and Bluethmann, H. 1995. Mucosal adjuvant effect of cholera toxin in mice results from induction of T helper 2 (Th2) cells and IL-4. J. Immunol. 155: 4621-4629.
McDermott, M.R., and Bienenstock, J. 1979. Evidence for a common mucosal immunologic system. I. Migration of B immunoblasts into intestinal respiratory and genital tissues. J. Immun. 122: 1892-1898.
Merritt, E.A., Pronk, S.E., Sixma, T.K., Kalk, K.H., van Zanten, B.A., and Hol, W.G. 1994. Structure of partially-activated E. coli heat-labile enterotoxin (LT) at 2.6 A resolution. FEBS Lett. 337: 88-92.
Nashar, T.O., Webb, H.M., Eaglestone, S., Williams, N.A., and Hirst, T.R. 1996. Potent immunogenicity of the B subunits of Escherichia coli heat-labile enterotoxin: receptor binding is essential and induces differential modulation of lymphocyte subsets. Proc. Natl. Acad. Sci. 93: 226-230.
Northrup, R.S., and Fauci, A.S. 1972. Adjuvant effect of cholera enterotoxin on the immune response of the mouse to sheep red blood cells. J. Infect. Dis. 125: 672-673.
Pelham, H.R. 1992. The Florey Lecture, 1992. The secretion of proteins by cells. Proc. Soc. Lond. Biol. Sci. 250: 1-10.
Punyashthiti, K., and Finkelstein, R.A. 1971. Enteropathogenicity of Escherichia coli. Evaluation of mouse intestinal loops. Infect. Immun. 4: 473-478.
Ruddock, L.W., Ruston, S.P., Kelly, S.M., Price, N.C., Freedman, R.B., and Hirst, T.R. 1995. Kinetics of acid- mediated disassembly of the B subunit pentamer of
Escherichia coli heat-labile enterotoxin. Molecular basis of pH stability. J. Biol.
Chem. 270: 29953-29958.
Ruddock, L.W., Webb, H.M., Ruston, S.P., Cheesman, C., Freedman, R.B., and Hirst, T.R. 1996. A pH-dependent conformational change in the B-subunit pentamer of
Escherichia coli heat-labile enterotoxin: structural basis and possible functional
role for a conserved feature of the AB5 toxin family. Biochemistry 35: 16069-16076.
Siev, D. 1997. Interpretation and estimation of relative potency in vaccines. J. Immun. Methods 208: 131-139.
Sixma, T.K., Kalk, K.H., van Zanten, B.A., Dauter, Z., Kingma, J., Witholt, B., and Hol, W.G. 1993. Refined structure of Escherichia coli heat-labile enterotoxin, a close relative of cholera toxin. J. Mol. Biol. 230: 890-918.
Spangler, B. D. 1992. Structure and function of cholera toxin and related Escherichia
coli heat-labile enterotoxin. Microbiol. Rev. 56: 622-647.
Stavric, S., Speirs, J.I., Konowalchuk, J., and Jeffrey, D. 1978. Stimulation of cyclin AMP secretion in Vero cells by enterotoxins of Escherichia coli and Vibrio
cholerae. Infect. Immun. 21: 514-517.
Sun, J.B., Holmgren, J., and Czerkinsky, C. 1994. Cholera toxin B subunit: an efficient transmucosal carrier-delivery system for induction of peripheral immunological tolerance. Proc. Natl. Acad. Sci. 91: 10795-10799.
Sun, J.B., Rask, C., Olsson, T., Holmgren, J., and Czerkinsky, C. 1996. Treatment of experimental autoimmune encephalomyelitis by feeding myelin basic protein conjugated to cholera toxin B subunit. Proc. Natl. Acad. Sci. 93: 7196-7201.
Tacket, C.O., Cohen, M.B., Wasserman, S.S., Losonsky, G., Livio, S., Kotloff, K., Edelman, R., Kaper, J.B., Cryz, S.J., Giannella, R.A., Schiff, G., and Levine, M.M. 1999. Randomized, double-blind, placebo-controlled, multicentered trial of the efficacy of a single dose of live oral cholera vaccine CVD 103-HgR in preventing cholera following challenge with Vibrio cholerae O1 El tor inaba three months after vaccination. Infect. Immun. 67: 6341-6345.
Takahashi, I., Marinaro, M., Kiyono, H., Jackson, R.J., Nakagawa, I., Fujihashi, K., Hamada, S., Clements, J.D., Bost, K.L., and McGhee, J.R. 1996. Mechanisms for mucosal immunogenicity and adjuvancy of Escherichia coli labile enterotoxin. J. Infect. Dis. 173: 627-635.
Truitt, R.L., Hanke, C., Radke, J., Mueller, R., and Barbieri, J.T. 1998. Glycosphingolipids as novel targets for T-cell suppression by the B subunit of recombinant heat-labile enterotoxin. Infect. Immun. 66: 1299-1308.
Vajdy, M., and Lycke, N.Y. 1992. Cholera toxin adjuvant promotes long-term immunological memory in the gut mucosa to unrelated immunogens after oral immunization. Immunology 75: 488-492.
Vajdy, M., Kosco-Vilbois, M.H., Kopf, M., Kohler, G., and Lycke, N. 1995. Impaired mucosal immune responses in interleukin 4-targeted mice. J. Exp. Med. 181: 41-53. Williams, N.A., Hirst, T.R., and Nashar, T.O. 1999. Immune modulation by the
Williams, N.A., Stasiuk, L.M., Nashar, T.O., Richards, C.M., Lang, A.K., Day, M.J., and Hirst, T.R. 1997. Prevention of autoimmune disease due to lymphocyte modulation by the B-subunit of Escherichia coli heat-labile enterotoxin. Proc. Natl. Acad. Sci. 94: 5290-5295.
Wilson, A.D., Bailey, M., Williams, N.A., and Stokes, C.R. 1991. The in vitro production of cytokines by mucosal lymphocytes immunized by oral administration of keyhole limpet haemocyanin using cholera toxin as an adjuvant. Eur. J. Immunol. 21: 2333-2339.
Wilson, A.D., Clarke, C.J., and Stokes, C.R. 1990. Whole cholera toxin and B subunit act synergis tically as an adjuvant for the mucosal immune response of mice to keyhole limpet haemocyanin. Scand. J. Immunol. 31: 443-451.
Yamamoto, S., Kiyono, H., Yamamoto, M., Imaoka, K., Fujihashi, K., Van Ginkel, F.W., Noda, M., Takeda, Y., and McGhee, J.R. 1997a. A nontoxic mutant of cholera toxin elicits Th2-type responses for enhanced mucosal immunity. Proc. Natl. Acad. Sci. 94: 5267-5272.
Yamamoto, S., Takeda, Y., Yamamoto, M., Kurazono, H., Imaoka, K., Yamamoto, M., Fujihashi, K., Noda, M., Kiyono, H., and McGhee, J.R. 1997b. Mutants in the ADP-ribosyltransferase cleft of cholera toxin lack diarrhea genicity but retain adjuvanticity. J. Exp. Med. 185: 1203-1210.
計 畫 成 果 自 評 本年度計畫已達成預期進度,並已完成下列四項預定目標: 一、大腸桿菌 LT 基因及其次單位基因之選殖、表現及功能性分析 我們已選殖大腸桿菌 LT 基因,並利用原核表現系統,在三小時的操作時間 內,自 1 公升細菌液中,純化出約 1.3 公克之 LT,其純度至少 99﹪。重組型 LT 及 LTB進一步利用西方墨點法及 GM1酵素連結免疫吸附試驗與野外型 LT 互相比 較,發現我們所產製的重組型 LT 及 LTB 具備與野外型 LT 相同的抗原性及功能, 並具有容易生產、容易純化的特性,可以應用於以下的研究。 二、LTB 作為大腸桿菌疫苗可行性之探討 在小白鼠試驗中發現,經由口服的途徑,在血清、肺及腸道中可以誘發高量 的 IgA 反應,與陰性對照組比較分別增加 4.3、5.8 及 6.3 倍,而在 IgG 反應方面, 則分別增加 1.7、2.9 及 7.7 倍,顯示大腸桿菌 LTB 在不需要經過化工包裝的過程 下,可以直接口服,在肺及腸道引發良好的黏膜性免疫反應。 三、腸毒素下痢分子模式之建立及其應用於藥物搜尋之研究
我們利用 LTB 與 GM1接合為基礎,設計 competitive binding assay,搜尋可
能抑制下痢之中藥。在我們初步搜尋中藥後,發現其中 11 種中藥材具有可以阻 斷 LTB 與 GM1結合的能力,而由動物試驗中,也證實乾薑可以透過干擾 LT 與腸 上皮細胞的結合,而抑制下痢的發生。 四、大腸桿菌 LT 與細胞相互關係之研究 我們利用實驗室已構築好的重組細胞株,觀察 LT 及 LTB 對 AP-1 及 NF-κ B 活性的影響。由初步的結果顯示,LTB 似乎可以藉著調控 NF-κB 的活性,影 響免疫反應的發生。進一步,我們擬探討 LT 及 LTB 影響轉錄因子活性的訊息傳 導路徑,並探討這些調控機制所代表的生物意義。
圖 一 LT 表現質體的構築。利用聚合 連鎖反應先自大腸桿菌中增殖出 LT 基 因,架接於 pBlueScript 載體上,再轉接入 pET-28 表現載體中,以進行蛋白質的 表現。 LTP1 pBlueScript KS LTA LTB LTM1 BamHI, PstI LT pET-28 LT pET- LT pBKS-LT BamHI, XhoI BamHI, PstI BamHI, XhoI (His)6
圖 二 利用 pET 表現 LT 的模式圖。
IPTG induction
T7 RNA pol gene
lac promoter
T7 RNA polymerase
LT gene
pET-LT
LT protein
pT7
E. coli BL21(DE3)
圖 三 LT 及 LTB 的表現及純化。在加入 IPTG 後,將細菌打破,再利用電泳分 析。Lane 1 及 3 分別代表表現 LT 及 LTB 的大腸桿菌粗萃取物,lane 2 及 4 分別 代表經純化後的 LT 及 LTB。LTA 及 LTB 的位置標示在右邊。蛋白質的分子量 (kDa)表示於左邊。
LTB
LTA
M 1 2 3 4
6 14 22 31 45 66 97圖 四 利用西方雜合法偵測 LT 及 LTB 的抗原性。將純化後的 LT 及 LTB 利用電 泳分析後,轉漬到濾紙上,再利用抗 LT 的老鼠血清進行偵測。LTA 及 LTB 的 位置標示在右邊。蛋白質的分子量(kDa)表示於左邊。
M
LT LTB
LTB
14 22 31 66LTA
45圖 五 GM1酵 素 連 結 免 疫 吸 附 試 驗。酵素連結免疫吸附平盤上先吸附 GM1,再分 別加入不同濃度的大腸桿菌重組型 LT(rLT)、LTB(rLTB)及野生型 LT(wtLT), 最後加入受質,於 405 nm 波長下測定吸光值。所列之數值為三次實驗所得之平 均值±標準偏差。 0 0.3 0.6 0.9 1.2 0.001 0.002 0.004 0.008 0.016 0.032 0.064 0.128
μ
g/well
OD405
r LT r LTB wt LT(A)
(B)
圖 六 Competitive GM1-ELISA 之設定。利用 GM1-ELISA 的方式測出 LTB 結合
GM1的標準曲線(A),再加入不同濃度稀釋的抑制劑 GM1(B)分別對 0.1 μg/ml、 0.2 μg/ml、0.4 μg/ml 的 LTB 做拮抗作用。經受質呈色在 OD405測其波長吸光 值即 LTB 的結合比率以 Y 軸表示,而藥物濃度以 X 軸表示,折線圖代表兩者之 間的關係。 0 0.4 0.8 1.2 1.6 0.00625 0.0125 0.025 0.05 0.1 0.2 Concentration (μ g/ml) OD405 LTB=0.1ug/well LTB=0.2ug/well LTB=0.4ug/well 0 0.4 0.8 1.2 1.6 0.00013 0.00025 0.0005 0.001 0.002 0.004 0.008 0.016 0.032 0.064 0.128 0.256 Concentration (μ g/ml) OD405
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.3830.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.383 0.7661.53 3.066.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.0960.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.0960.191 0.383 0.766 1.533.06 6.1312.5 25 50 100 200 0 0 . 2 0 . 4 0 . 6 0 . 8 1 1 . 2 1 . 4 1 . 6 0.096 0.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.0960.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.3830.766 1.533.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0.096 0.191 0.383 0.766 1.53 3.06 6.13 12.5 25 50 100 200 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 0 . 0 9 60 . 1 9 1 0 . 3 8 3 0 . 7 6 6 1.53 3.06 6.13 12.5 2 5 5 0 100 200 圖 七 抑制 LTB 結合 GM1 前 11 名中藥之劑量反應。將甲醇中藥萃取液利用 GM1-ELISA 的方式與 0.016 μg 的 LTB 混合後反應最後在 O D405波長下呈色之結 果。X 軸代表各溶液層所含成份之濃度,Y 軸代表 LTB 結合 GM1的比率以 OD405
表示。50% Inhibitory concentration (IC50)為抑制 LTB 結合 GM1達一半時所需的藥
物濃度。 絞 股 蘭IC50= 2.3 μg/ml 栝 樓 實IC50=6.7 μg/ml 蓮 子 蕊IC50=2.4 μg/ml 遠 志IC50= 33.2 μg/ml 厚 朴IC50=29.2 μg/ml 淡 豆 鼓IC50= 35.5 μg/ml 車 前 子IC50=32.3 μg/ml 槲 寄 生IC50= 3.4 μg/ml 貝 母 IC50= 1.4 μg/ml 乾薑 IC50= 2.0 μg/ml 香 薷 IC50= 2.1 μg/ml
0 0.05 0.1 0.15 0.2 0.25 0.3 PBS solvent+LT LT 0.24 1.2 5.8 圖 八 乾薑 甲 醇 萃 取 液 對 腸 毒 素 誘 發 腸 液 堆 積 之 抑 制 效 果 。(A)將純化後的忌 熱型腸毒素 1 μg 分別與 solvent、0.24 μg、1.2 μg、5.8 μg 的乾薑甲醇萃取 液萃混合,控制組只有 PBS,在室溫搖晃反應 3 h 後打入 Balb/Cby mice 已結紮 的空迴腸內,24 小時後犧牲取出結紮部位排列圖。(B)將腸子秤重後量結紮長 度並計算其比率(Ratio=g/cm)。所列之數值為三次實驗所得之平均值±標準偏差。 (A) (B) 1. 2. 3. 4. 5. 6. Fluid accumulation (g/cm) 0.24 1.2 5.8 單 位 長 度 ( 公 分 ) LT
+
+
+ + +
Methanol only IC50<0.24 μg/250 μl 1. Concentration (μg/250μl)(A) 0 0.6 1.2 1.8 0.0128 0.064 0.32 0.16 8 40 200 1000 Protein amount (ng)
Relative AP-1 activity
LT LTB (B) 0 1 2 3 4 0.0128 0.064 0.32 0.16 8 40 200 1000 Protein amount (ng) Relation NF-kB activity LT LTB 圖 九 LT 及 LTB 對 AP-1 及 NF-κB 活性的影響。將不同濃度的 LT 及 LTB 加
到含有 AP-1/luciferase(A)或 NF-κB/luciferase(B)的 HepG2 重組細胞中,於
作用 24 小時後,收集細胞,再利用 Luciferase Assay Kit(Promega)測定 luciferase 的活性。結果以實驗組除以對照組的數值表示,所列之數值為三次實驗所得之平 均值±標準偏差。