行政院國家科學委員會專題研究計畫 期中進度報告
子計畫四:探討 JAK-STAT 在氧化態低密度脂蛋白活化小神經 膠細胞的訊息角色及不同氧化程度低密度脂蛋白引起與加
強腦中風傷害的差異性比較(1/3)
計畫類別: 整合型計畫
計畫編號: NSC93-2745-B-038-005-URD
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 臺北醫學大學藥理學科
計畫主持人: 蕭哲志 共同主持人: 許重義 計畫參與人員: 林怡秀
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 6 月 1 日
行政院國家科學委員會補助專題研究計畫 -期中進度報告
探討 JAK-STAT 在氧化態低密度脂蛋白活化小神經膠細胞的訊息角 色及不同氧化程度低密度脂蛋白引起與加強腦中風傷害的差異性比
較(子計畫四) (1/3)
The studies of the role of JAK-STAT on oxidized low-density lipoprotein-induced microglia activation and comparison of the deleterious effects between different oxidative degree of low-density
lipoproteins on stroke (1/3)
計畫類別:整合型計畫
計畫編號:NSC 93-2745-B -038 -005-URD 執行期間:93 年 8 月 1 日至 94 年 7 月 31 日
計畫主持人:許重義教授校長 子計畫主持人:蕭哲志副教授 計畫參與人員:林怡秀技術員
成果報告類型(依經費核定清單規定繳交):精簡報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計 畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位:台北醫學大學醫學院藥理學研究所
中 華 民 國 94 年 5 月 30 日
一、中英文摘要
(IV) 探討 JAK-STAT 在氧化態低密度脂蛋白活化小神經膠
細胞的訊息角色及不同氧化程度低密度脂蛋白引起與加強
腦中風傷害的差異性比較
中樞神經系統之小神經膠細胞(microglia)在腦中風發生過程中腦 細胞損傷或髓鞘發炎氧化性損傷上扮演重要角色。根據文獻顯示氧化 態低密度脂蛋白為造成粥狀動脈硬化症的主因,此變性低密度脂蛋白 藉著血管內巨噬細胞(macrophage)之清除性接受體攝入或引發特定細 胞反應。氧化態低密度脂蛋白不僅對於內皮細胞具細胞致毒性,更可 誘發長期性之慢性發炎反應。本計劃的主要目地為探討氧化態低密度 脂蛋白對腦中風相關發炎性傷害過程之影響性,尤其探討氧化態低密 度脂蛋白活化小神經膠細胞的 iNOS synthase 表現作用機轉,進一步 比較不同來源與氧化程度的低密度脂蛋白之作用性,以了解其作用之 訊息系統。
首先,實驗室完成調整適當初級小鼠小神經膠細胞之分離與培養 條件。從實驗結果發現人類低密度脂蛋白在不同氧化條件與處理下對 於小神經膠細胞之作用程度不同。以鐵離子或空氣氧化人類低密度脂 蛋白不同時間點(6~24 小時),不論透析與否或處理濃度不同(50~100
µg/ml)皆不明顯影響細胞產生一氧化氮或一氧化氮合成酶的表現。以
鐵或銅離子氧化的人類低密度脂蛋白在經過透析處理後對於細胞存 活率具有較明顯的抑制作用。當以銅離子氧化人類低密度脂蛋白 24 小時,不經過透析過程 者,可明顯依濃度效應刺激一氧化氮產生及一氧化氮合成酶之表現。
此刺激一氧化氮產生之作用可被 LY294002 及 SB203580 抑制,而不
被 PD98059 影響,此說明特定氧化之人類低密度脂蛋白刺激小神經 膠細胞之訊息可能藉由 PI3-kinase 或 p38 MAP kinase 之路徑。從這些 結果可提供不同氧化處理之人類低密度脂蛋白對細胞之影響性,並可 確 認 未 來 實 驗 進 行 之 低 密 度 脂 蛋 白 種 類 , 進 而 加 速 有 關 訊 息 (JAK-STATs)研究之進行。
關鍵詞:小神經膠細胞、氧化態低密度脂蛋白、誘發型一氧化氮合成 酶、銅離子
(IV) The studies of the role of JAK-STAT on oxidized low-density lipoprotein-induced microglial activation and comparison of the deleterious effects between different oxidative degree of low-density lipoproteins on stroke
Microglia, the resident macrophage of the brain, mediates immune and inflammatory responses in the central nervous system. Activation of microglia and secretion of inflammatory cytokines associated with the pathogenesis of degenerative diseases. Oxidized low-density lipoprotein (LDL) is believed to play as a critical and important role in various cellular processes involved in the progression of atherogenesis. Therefore, we will especially focus on the role of microglia in the oxLDL-associated stroke. Oxidized LDL is an activator of macrophage and microglia for cytokines and nitric oxide production. Signaling may through its scavenger receptor, and activation of Janus kinase and signal transducer and activator of transcription (JAK-STAT) pathway in microglial cells.
According to our findings, various cation-induced oxidation of LDLs had
different effects. Especially, non-dialyzed copper-catalyzed LDL could induce nitric oxide production and iNOS expression in microglia. It also have less cytotoxic to microglia as compare to ferrous-catalyzed LDL.
Therefore, the mechanisms of nitric oxide synthase or cytokines induction by copper-catalyzed oxidized LDL will be clarified, including JAKs, STATs and phosphatase.
Key words : microglia, oxidized LDL, inducible nitric oside synthase, copper ion
二、源由與目的
腦中風根據定義,就是因為腦血管阻塞或破裂而造成突然或陣發 性的大腦局部或全部的功能失調。腦中風依受損的腦區不同,中風可 以導致不同程度的感覺 (sensory)、運動 (motor)、以及認知障礙 (cognitive functions) (Chen et al., 2002)。由於對中風的病生理並不是完 全透徹瞭解,因此在臨床上處理大腦缺血仍有很多不確定性,一些研 究 指 出 在 中 風 後 造 成 的 神 經 傷 害 包 括 有 : 興 奮 性 毒 性 (excitotoxicity)、鈣離子負荷 (calcium overload)、自由基 (free radicals) 產生以及某些蛋白質合成受到抑制。造成缺血後之低灌流效應的因 子,leukocytes 也參與其過程,在組織缺血後所誘導的發炎反應扮演 重要角色 (Connolly et al., 1996),尤其 microglia 在腦部慢性發炎反應 上扮演相當重要的角色。
Microglia 在 不 同 的 神 經 病 變 中 會 活 化 成 腦 巨 噬 細 胞 (brain macrophage) 執行吞噬功能,此時表面抗原像 Fc 及 CR3 受體會增 加,且伴隨快速誘導組織相容第一型及第二型抗原 (MHC class I & II antigens) 的新合成 (Streit et al., 1988)。所以毫無疑問的 microglia 有 吞噬及免疫的潛能。當中樞神經系統受傷時,microglia 會活化而作 為第一道防禦線。活化過程包括細胞增生、增加免疫表型、 microglia 補充到受傷區域 (Lindholm et al., 1992) 和功能改變,例如釋放 NO 等毒性物質或 cytokines 等發炎物質 (如 IL-1
β, TNFα
等) (Banati et al., 1993)。除此之外,microglia 有很強的抗原表現和明顯的毒性作用。再者,microglia 的活化呈階段性反應;如起初並沒有吞噬作用,過 一段時間,特別是當神經或突觸末端退化時,才會轉變成腦內的吞噬 細胞 (Streit et al., 1988)。在許多發炎反應或敗血症發生的過程中,
microglia 會 釋 放 好 幾 種 cytokines 包 括 IL-1, IL-6, TNFα 和 TGFβ (transforming growth factor β) 等(Dickson et al., 1993);這些
cytokines 都有助於星狀細胞的活化,誘導細胞附著分子(adhesion molecules)表現,增加 T 細胞到受傷區;並且在發炎後期會負責調節 使疾病不至於惡化。同時,microglia 會釋放其它傷害物質,像是蛋 白分解酵素(protease), 反應氧中間物(reactive oxygen intermediates)和 一氧化氮 (NO) (Thery et al., 1991)。而是否不同氧化程度的 ox-LDL 會增加 microglia 表現 ICE 或 iNOS 的活性目前並不清楚。
中樞神經系統發炎時,microglia 會明顯的釋放出 NO 及 TNF-
α
。 此 兩 者 目 前 已 被 證 實 與 發 炎 性 的 脫 髓 鞘 疾 病 (inflammatory demyelinating diseases)如多發性硬化症(multiple sclerosis) (在腦或脊 髓或兩者之間發生發炎性斑狀硬化) 有關 (Parkinson et al., 1997)。而 在中樞神經系統中,microglia 被視為釋放 NO 及 cytokines (TNF-α)
的主要來源。氧化態低密度脂蛋白 (oxidized low-density lipoprotein oxLDL)不僅對於內皮細胞具細胞致毒性,同時也是一個強效平滑肌 增生引發劑,更可誘發長期性之慢性發炎反應。近來文獻也提出 oxLDL 本身與急性冠狀動脈症及急性腦部梗塞發作有相當之一致性 (Tsimikas et al., 2003; Uno et al., 2003)。特別的是 oxLDL 會直接影響 而降低冠狀動脈血流 (Laaksonen et al., 2002) 及直接造成實驗動物 微血管通透度與白血球之沾黏性增高 (Svensjo et al., 2003)。OxLDL 在不同腦部發炎性與退化性疾病扮演相當重要之角色 (Newcombe et al., 1994; Emsley and Tyrrell, 2002)。訊 息 活 化 轉 錄 因 子 (The signal transducer and activator of transcription factors, STAT)一般被認為是生長因子或干擾素所活化而 啟動 DNA 轉錄的重要因子,但目前發現此訊息的活化可藉多種刺激 劑而達成。不同種類的 JAK 與 STAT 受不同訊號刺激而對應選擇性活 化 (Darnell et al., 1994; Zhang et al., 1995)。當相關接受體受活化時,
其 接 受 體 相 接 合 (associated) 的 JAKs (1/2/3) 便 會 磷 酸 化
(autophosphorylation),進而再活化相關 STAT (1/2/3/4/5)。此活化磷酸 化之 STAT 再轉位進入核內啟動基因表現 (Zhang et al., 1995) 此磷酸 化 之 STAT 可 受 去 磷 酸 酵 素 (Src homology 2 domain-containing protein phosphatase 2, SHP2) 所調控(Park et al., 2003)。近來發現腦部 特殊脂肪成份可藉由 JAK-STAT 訊息以誘發 microglia 合成釋放發炎 物質(NO 與 MCP-1) (Kim et al., 2002)。另外 Interleukin 3 同樣可藉 JAK2-STAT5 訊息來活化 microglia 之 CD40 生成 (Bright et al., 2004)。特別的是在不同細胞下 ROS 均可影響 JAK-STAT 訊息活化之 程度 (Simon et al., 1998)。尤其 oxLDL 在內皮細胞可藉 ROS 來活化 JAK2 與 STAT 訊息過程 (Maziere et al., 2001; 1999)。特別是 oxLDL 也藉其接受體而刺激 microglia 引起吞噬與 ROS 之增加 (Newcombe et al., 1994; Coraci et al., 2002)。然而值得注意的是不同來源與氧化程度 的 LDL 似乎在細胞活化上有不同的結果差異 (Carpenter et al., 2003)。
本總計劃的主要目地為進一步探討且假設氧化態低密度脂蛋白 活化小神經膠細胞並誘發 NO 或 cytokines 釋放,且其的作用機轉,
可能藉由 JAK-STATs 訊息系統達成;另外我們亦欲進一步比較不同 來源與氧化程度的低密度脂蛋白在離體細胞通透度與活體動物內引 起與加強局部腦缺血再灌流傷害的影響性。
因此,本次計畫擬針對不同氧化處理的 ox-LDL 探討對 microglia 釋放 cytokines (如 IL-1
β, TNFα
等)或 NO 的差異性以及表現 inducible NO synthase (iNOS) 的作用。透過本期與未來兩年實驗,將有助於瞭 解不同來源與氧化程度的 ox-LDL 對中樞神經系統的 microglia 活化及 腦缺血所引起的腦梗塞反應所扮演的角色及各種不同作用劑對這些 反應的抑制效果。經由這些研究可引領出一個新的治療方向,以期做 為臨床治療的參考依據。三、結果與討論
首先完成設立且調整實驗條件,以具成熟性之實驗過程分離大鼠 小神經膠細胞(rat microglial cells) (Figure 1 and 2)。同時也完成設立人 類低密度脂蛋白(human low-density lipoprotein, LDL)之分離(Figure 3A)與其氧化源包括銅(Cu2+)、鐵(Fe2+)離子與空氣(Air)之氧化方法 (Figure 3B) 。 另 外 , 部 分 氧 化 態 之 LDL (oxidized LDL) 經 透 析 (MWCO:12-14000, SPECTRUM)處理,再進行細胞實驗。以銅離子(5
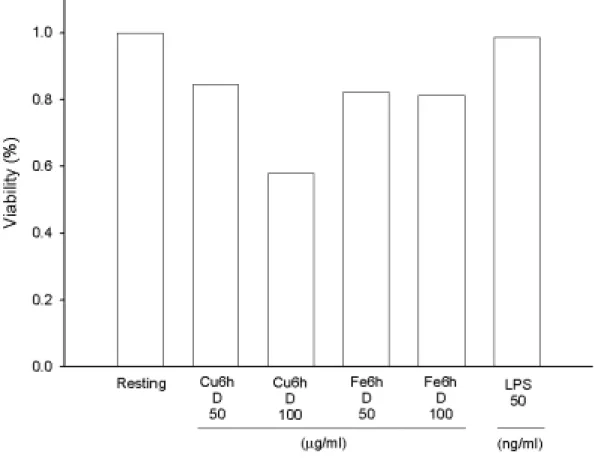
µM)氧化 6 小時之透析後 LDL (100 µg/ml)具明顯降低細胞之存活率
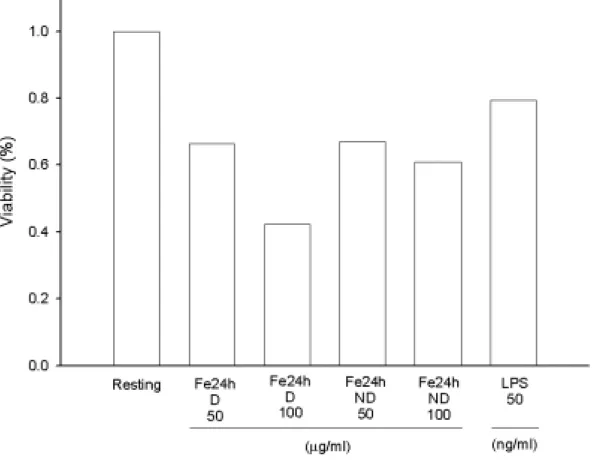
(Figure 4)。而以鐵離子(5 µM)氧化 24 小時之透析後 LDL (100 µg/ml) 較不透析者具較明顯降低細胞之存活率(Figure 4 and 5)。當延長鐵離 子氧化之時間至 48 小時後,oxLDL 隨濃度效應而減少細胞存活率 (Figure 6)。特別的是以非透析處理之 oxLDL (100 µg/ml)影響最大,而原態 LDL 對細胞存活均較氧化態 LDL 為佳(Figure 6)。在相同氧化 時間下(24 小時)且不經透析處理,發現以銅離子較鐵離子處理之 LDL 有較佳之存活率(Figure 7)。內毒素 LPS 在濃度為 50 ng/ml 下影響存 活率則較 oxLDL 為低(Figure 4 and 7)。
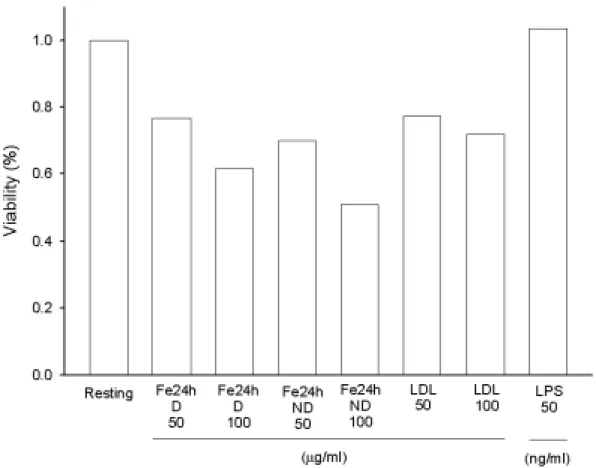
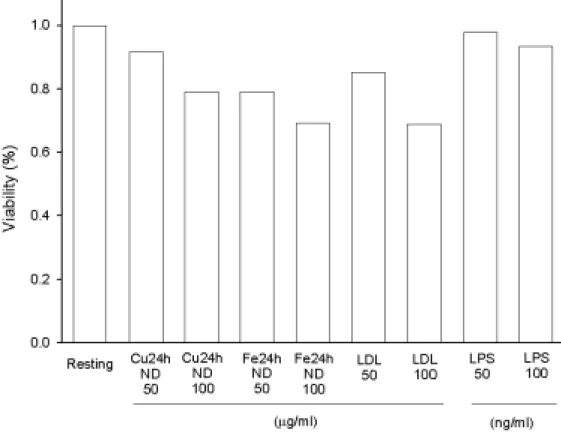
所以,針對小神經膠細胞而言,以不同氧化時間下,銅或鐵離子 處理之 LDL,若經透析則較明顯降低細胞存活率,而銅離子處理之 LDL,若不經透析則有較高之細胞存活率。
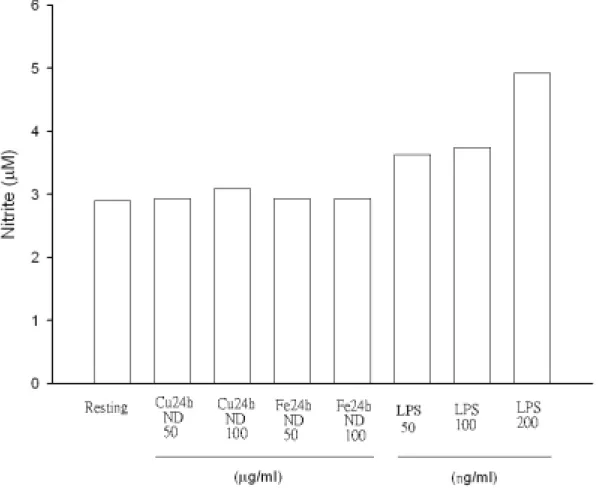
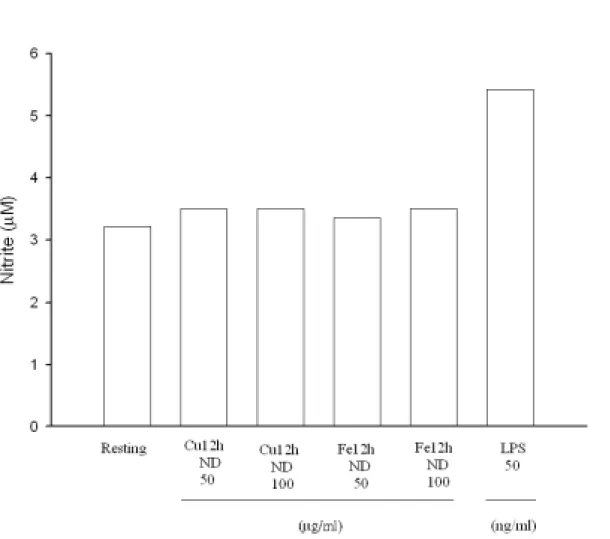
在評估不同氧化處理 LDL 對小神經膠細胞產生一氧化氮(NO)之 實驗上,Figure 8 顯示在以鐵離子氧化 LDL 24 小時後,不論經透析 與否均些微影響一氧化氮之生成,特別是原態 LDL (100
µg/ml)有較
高之產生,而 LPS (50 ng/ml)則最為明顯增加(Figure 8),特別的是若 LDL 氧化溫度降為 25℃,則測量不到上述 NO 些微之增加情形(Figure 9)。另外,若氧化時間縮短為 12 小時,則以銅或鐵離子氧化並透析 之 LDL 同樣不會誘發小神經膠細胞產生一氧化氮(Figure 10)。當以鐵 離子氧化且透析之 LDL 處理下,不同濃度之 oxLDL 並不隨刺激時間 而增加。然而以 LPS (50 ng/ml)處理則發現 NO 產生量隨時間延長而 增加(Figure 11)。特別的是當以銅離子氧化而不經透析處理之 LDL 會 隨濃度及處理時間而增加 NO 之產生量。相反地,鐵離子處理之 LDL 並不具有上述作用。原態之 LDL 在高濃度(100
µg/ml)處理 30 小時後
則有明顯增加 NO 之含量。然而以 LPS (50 與 100 ng/ml)處理下會隨 濃度與刺激時間而增加 NO 之含量(Figure 12)。單純以銅離子處理細 胞並不會引發如銅離子氧化未透析之 LDL 所造成的 NO 生成(Figure 13)。利用不同抑制劑處理發現,LY294002 (PI 3-kinase 抑制劑,5
µM)
與 SB203580 (p38 MAP kinase 抑制劑,5µM)均可抑制由未透析銅離
子氧化 LDL 所誘發細胞 NO 的生成,然而 PD98059 (MEK 抑制劑,5µM)之效果較差(Figure 14)。
綜合上述實驗,我們發現 LDL 在經銅離子氧化後,其不經透析 之條件下,可明顯刺激小神經膠細胞產生 NO 且隨濃度效應與時間效 應而增加。此刺激機轉可能藉由 PI 3-kinase 或 p38 MAP kinase 活化 以達成作用。然而若經透析後則此刺激效應會降低。但鐵離子所造成 oxLDL 在各條件下,其僅些微刺激 NO 之產生。另外,高濃度原態 LDL 或許經長時間在 microglia 處理下可轉成 oxLDL 以刺激產生 NO,但待進一步研究確認。
由銅離子氧化且經透析處理之 LDL 並不含刺激小神經膠細胞產 生 NO,但 LPS (50 ng/ml)可明顯刺激 iNOS 表現(Figure 15)。另一方 面,由鐵離子氧化不同時間比較上,以氧化 24 小時且經透析處理隻 LDL 僅些微刺激小神經膠細胞產生 NO (Figure 16)。由空氣(Air)氧化
且經透析處理隻 LDL 並不刺激細胞產生 NO (Figure 17)。由銅離子、
鐵離子氧化與原態 LDL 比較刺激細胞內 iNOS 的表現,可發現的確 以銅離子氧化且不經透析之 LDL 具有較強誘發小神經膠細胞 iNOS 的表現(Figure 18)。
上述 iNOS 誘發表現之實驗結果發現,的確以銅離子氧化 24 小 時且經透析之 LDL 具有較強的刺激小神經膠細胞之作用,此也印證 NO 生成的實驗結果。綜合以上實驗我們認為以銅離子氧化且不經透 析之 LDL 較不具細胞毒性,而具有刺激小神經膠細胞之作用,其機 轉可能與 PI 3-kinase 或 p38 MAP kinase 有關,是否與 STAT 訊息有關 仍需進一步探討研究。
四、計畫成果自評
此研究計畫為本人第一次取得之國科會能量型多期性研究計 畫。除了進行個人之子計畫實驗外,同時也指導教授其它子計畫研究 人員有關人類低密度脂蛋白之分離與氧化處理。本研究室設立並確實 改善初級細胞之(microglia)分離品質與培養進度,但此大鼠初級細胞 分離與培養成熟使用時間較長(需 9 天),又此腦部細胞並不增殖分 裂,所以每次進行實驗前必須犧牲動物以獲取細胞,實驗進展上相當 花費時間。因此,本實驗室於 2005 年 5 月中已進行採購義大利細胞 庫 BV-2 之細胞株(此細胞株於 2005 年 4 月始於販售),希望以此細胞 株之培養而加快實驗進度。藉由此一系列實驗,我們清楚不同氧化與 透析處理之 LDL 其對細胞作用性有差異。其中以銅離子刺激 24 小時 且不透析之 LDL 具有最明顯之誘發小神經膠細胞之作用。實驗有關 oxLDL 刺激細胞產生 iNOS 之作用與機轉探討,目前正積極進行中。
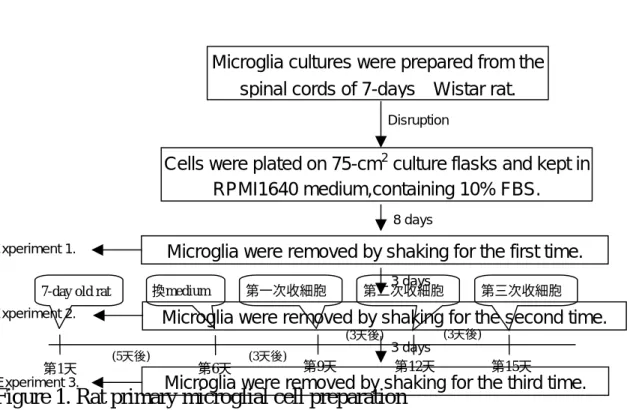
Figure 1. Rat primary microglial cell preparation
(3天後) (3天後)
(3天後) (5天後)
第1天 第6天 第9天 第12天 第15天
7-day old rat 換medium 第一次收細胞 第二次收細胞 第三次收細胞
Microglia cultures were prepared from the spinal cords of 7-days Wistar rat.
Cells were plated on 75-cm2culture flasks and kept in RPMI1640 medium,containing 10% FBS.
Microglia were removed by shaking for the first time.
Disruption
8 days
3 days
Microglia were removed by shaking for the second time.
Microglia were removed by shaking for the third time.
3 days Experiment 1.
Experiment 2.
Experiment 3.
A
B
C
Figure 2. The preparation of primary rat microglial cells. Microgli (Panel B) were harvested from flasks of mixed glial cultures (Panel A) by shaking for 2 hr. Cells were collected by centrifugation and seeded at 5×104 cells/ml. The attached cells (Panel C) were observed after last collection. The collective cells were further cultured in RPMI-1640 supplemented with 100% FBS for 1 day. Cells (5×104 cells/ml) were incubated with indicated oxidized LDL for activation.
(A)
Human LDL isolation
Sequential density gradient ultracentrifugation
(B)
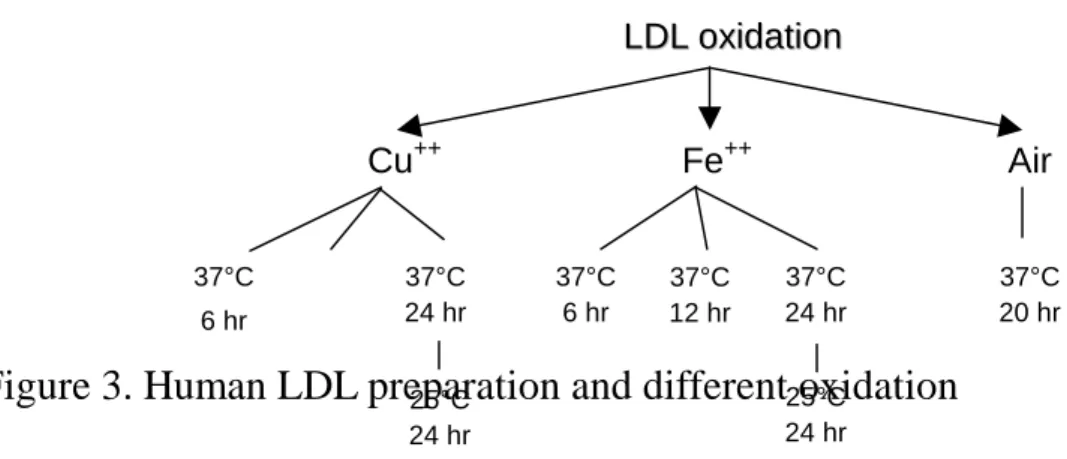
Figure 3. Human LDL preparation and different oxidation
L
LDDLL ooxxiiddaattiioonnCu++ Fe++ Air
37°C 6 hr
37°C 24 hr
37°C 6 hr
37°C 24 hr
37°C 20 hr 37°C
12 hr
25°C 24 hr
25°C 24 hr
Figure 4. Effects of different oxidized LDL on rat microglial cell viability.
Microglia (5×104 cells/ml/well) were treated with oxidized LDL (Cu++- or Fe++-catalyzed LDL for 6 h with dialyzation, D, 50 or 100 µg/ml) for 22.5 hours, then added MTT to medium (final concentration, 0.5 mg/ml) to medium. After 1.5 hours, cells were washed and dissolved with DMSO.
Using the spectrophotometric method by ELISA reader to analyze the absorbance at 550 nm. Percentage of cell viability was calculated as the absorbance of treated cells/control cells×100.
Figure 5. Effects of different oxidized LDL on rat microglial cell viability.
Microglia (5 × 104 cells/ml/well) were treated with oxidized LDL (Fe++-catalyzed LDL for 24 h with or without dialyzation, D/ND, 50 or 100
µg/ml) for 22.5 hours, then added MTT to medium (final
concentration, 0.5 mg/ml) to medium. After 1.5 hours, cells were washed and dissolved with DMSO. Using the spectrophotometric method by ELISA reader to analyze the absorbance at 550 nm. Percentage of cell viability was calculated as the absorbance of treated cells/control cells×100.
Figure 6. Effects of different oxidized LDL on rat microglial cell viability.
Microglia (5 × 104 cells/ml/well) were treated with oxidized LDL (Fe++-catalyzed LDL for 48 h with or without dialyzation, D/ND, 50 or 100
µg/ml) for 22.5 hours, then added MTT to medium (final
concentration, 0.5 mg/ml) to medium. After 1.5 hours, cells were washed and dissolved with DMSO. Using the spectrophotometric method by ELISA reader to analyze the absorbance at 550 nm. Percentage of cell viability was calculated as the absorbance of treated cells/control cells×100.
Figure 7. Effects of different oxidized LDL on rat microglial cell viability.
Microglia (5×104 cells/ml/well) were treated with oxidized LDL (Cu++- or Fe++-catalyzed LDL for 24 h without dialyzation, ND, 50 or 100 µg/ml) for 22.5 hours, then added MTT to medium (final concentration, 0.5 mg/ml) to medium. After 1.5 hours, cells were washed and dissolved with DMSO. Using the spectrophotometric method by ELISA reader to analyze the absorbance at 550 nm. Percentage of cell viability was calculated as the absorbance of treated cells/control cells×100.
Figure 8. Effect of different oxidized LDL on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well) were treated with vehicle or oxidized LDL (Fe++-catalyzed LDL at 37℃ for 24 h with or without dialyzation, D/ND, 50 or 100 µg/ml) for 24 hr. Cell-free supernatant were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (50 ng/ml) was added as positive control.
Figure 9. Effect of different oxidized LDL on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well) were treated with vehicle or oxidized LDL (Cu++- or Fe++-catalyzed LDL at 25℃ for 24 h without dialyzation, ND, 50 or 100 µg/ml) for 24 hr. Cell-free supernatant were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (50, 100 or 200 ng/ml) was added as positive control.
Figure 10 Effect of different oxidized LDL on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well, 5% FBS) were treated with vehicle or oxidized LDL (Cu++- or Fe++-catalyzed LDL at 37℃ for 12 h without dialyzation, ND, 50 or 100 µg/ml) for 24 hr. Cell-free supernatant were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (50 or 100 ng/ml) was added as positive control.
Figure 11. Effect of different oxidized LDL on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well) were treated with vehicle (●) or oxidized LDL (Fe++-catalyzed LDL at 37℃ for 24 h with dialyzation, 50 (▼) or 100 (▽)
µg/ml) for 24 hr. Cell-free supernatant
were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (50 ng/ml, ○) was added as positive control.Figure 12. Effect of different oxidized LDL on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well) were treated with vehicle ( ● ), native LDL (50 or 100 µg/ml, ○ or ▼ ) or oxidized LDL (Cu++-catalyzed LDL at 37℃ for 24 h without dialyzation, 50 (□) or 100 (◆)
µg/ml or Fe
++-catalyzed LDL, 50 (▽) or 100 (■)µg/ml) at 37℃
for 24 h without dialyzation for 24 hr. Cell-free supernatant were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (50 or 100 ng/ml, ◇ or ▲) was added as positive control.
Figure 13. Effect of oxidized LDL or copper on nitrite production in rat microglial cells. Microglia (5×104 cells/ml/well) were treated with vehicle (Resting), oxidized LDL (Cu++-catalyzed LDL at 37℃ for 24 h without dialyzation, ND, 50 or 100 µg/ml) or copper for 24 hr. Cell-free supernatant were assayed for nitrite production (Griess’s method) and cellular lysates were analyzed for iNOS expression. LPS (100 ng/ml) was added as positive control.
Figure 14. Effect of various inhibitors on copper-catalyzed LDL-induced nitrite production. Microglia (5×104 cells/ml/well) were treated with solvent control (0.1% DMSO) or various inhibitors (LY294002, PD98059 or SB203580, 5 µM) for 15 min followed by the addition of oxidized LDL (Cu++-catalyzed LDL at 37℃ for 24 h without dialyzation, 50 or 100
µg/ml) for 24 hr. Cell-free supernatant were assayed for nitrite
production (Griess’s method). LPS (50 ng/ml) was added as positive control.A
B
C
Figure 15. Effect of different oxidized LDL on iNOS expression in rat microglial cells. Cells were treated with either a solvent control or oxidized LDL (Cu++-catalyzed LDL at 37℃ for 24 h or 6 hr with dialyzation, 50 or 100 µg/ml) for 24 hr. LPS (50 ng/ml) was added as positive control. The cellular lysates were analyzed for iNOS expression by Western blotting.
R Cu24-50 Cu24-100 LPS50
R Cu24-50 Cu24-100 LPS50
A
B
Figure 16. Effect of different oxidized LDL on iNOS expression in rat microglial cells. Cells were treated with either a solvent control or oxidized LDL (Fe++-catalyzed LDL at 37℃ for 6 h or 24 hr with dialyzation, 50 or 100 µg/ml) for 24 hr. LPS (50 ng/ml) was added as positive control. The cellular lysates were analyzed for iNOS expression by Western blotting.
R Fe6-50 Fe6-100 LPS50
R Fe24-50 Fe24-100 LPS50
A
B
Figure 17. Effect of different oxidized LDL on iNOS expression in rat microglial cells. Cells were treated with either a solvent control or oxidized LDL (Air-catalyzed LDL at 37 ℃ for 20 h or 6 hr with dialyzation, 50 or 100 µg/ml) for 24 hr. LPS (50 ng/ml) was added as positive control. The cellular lysates were analyzed for iNOS expression by Western blotting.
R Air-50 Air-100 LPS50
R LPS50 Air-50 Air-100
A
B
Figure 18. Effect of different oxidized LDL on iNOS expression in rat microglial cells. Cells were treated with either a solvent control or oxidized LDL (Cu++- or Fe++-catalyzed LDL at 37℃ for 24 h without dialyzation, 50 or 100 µg/ml) for 24 hr. LPS (50 or 100 ng/ml) was added as positive control. The cellular lysates were analyzed for iNOS expression by Western blotting.
R Cu24-50 Cu24-100 LDL50 LDL100 LPS50
R Fe24-50 Fe24-100 LDL50 LDL100 LPS50 LPS100
參考文獻
Banati, R.B., Gehrmann, J., Schubert, P. Kreutzberg, G.W. (1993) Cytotoxicity of microglia. Glia 7: 111-118.
Bright, J.J., Natarajan, C., Sriram, S., Muthian, G. (2004) Signaling through JAK2-STAT5 pathway is essential for IL-3-induced activation of microglia. GLIA. 45: 188-196.
Chen, P., Goldberg, D.E., Kolb, B., Lanser, M., Benowitz, L.I. (2002) Inosine induces axonal rewiring and improves behavioral outcome after stroke. Proc. Natl. Acad. Sci. USA 99: 9031-6.
Connolly, E.S. Jr, Winfree, C.J., Springer, T.A., Naka, Y., Liao, H., Yan, S.D., Stern, D.M., Solomon, R.A., Gutierrez-Ramos, J.C., Pinsky, D.J. (1996) Cerebral protection in homozygous null ICAM-1 mice after middle cerebral artery occlusion. Role of neutrophil adhesion in the pathogenesis of stroke. J. Clin. Invest. 97: 209-16.
Darnell, J.E., Jr, Kerr, I.M., Stark, G.R. (1994) Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science. 264: 1415-1421.
Dickson, D.W., Lee, S.C., Mattiace, L.A., Yen, S.C., Brosnan, C. (1993) Microglia and cytokines in neurological disease with special reference to AIDS and Alzheimer’s disease. Glia 7: 75-83.
Emsley, H.C., Tyrrell, P.J. (2002) Inflammation and infection in clinical stroke. J. Cereb. Blood Flow Metab. 22: 1399-1419.
Kim, O.S., Park, E. J., Joe, E.H., Jou, I. (2002) JAK-STAT signaling mediates gangliosides-induced inflammatory responses in brain microglial cells. J.B.C. 277: 40594-40601.
Laaksonen, R., Janatuinen, T., Vesalainen, R., Lehtimaki, T., Elovaara, I.,
Jaakkola, O., Jokela, H., Laakso, J., Nuutila, P., Punnonen, K., Raitakari, O., Saikku, P., Salminen, K., Knuuti, J. (2002) High oxidized LDL and elevated plasma homocysteine contribute to the early reduction of myocardial flow reserve in healthy adults. Eur. J.
Clin. Invest. 32: 795-802.
Lindholm, D., Castren, E., Kiefer, R., Zafra, F., Thoenen, H. (1992) Tarnsforming growth factor-beta 1 in the rat brain: increase after injury and inhibition astrocyte proliferation. J. Cell Biol. 117:
395-400.
Maziere, C., Alimardani, G., Dantin, F., Conte, M.A., Maziere, J.C. (1999) Oxidizes LDL activates STAT1 and STAT3 transcription factors:
possible involvement of reactive oxygen species. FEBS Lett. 448:
49-52.
Maziere, C., Conte, M.A., Maziere, J.C. (2001) Activation of JAK2 by the oxidative stress generated with oxidized low-density lipoprotein.
Free Radic. Biol. Med. 31: 1334-1340.
Newcombe, J., Li, H., Cuzner, M.L. (1994) Low density lipoprotein uptake by macrophages in multiple sclerosis plaques: implications for pathogenesis. Neutopathol Appl. Neurobiol. 20: 152-162.
Park, E.J., Park, S.Y., Joe, E., Jou, I. (2003) 15d-PGJ2 and rosiglitazone suppress janus kinase-STAT inflammatory signaling through induction of suppressor of cytokine signaling 1 (SOCS1) and SOCS3 in glia. The Journal of Biological Chemistry. 278: 14747-14752.
Parkinson, J.F., Mitrovic, B., Merrill, J.E. (1997) The role of nitric oxide in multiple sclerosis. J. Mol. Med. 75: 174-186.
Simon, A.R., Rai, U., Fanburg, B.L., Cochran, B.H. (1998) Activation of
the JAK-STAT pathway by reactive oxygen species. Am. J. Physiol.
275: C1640-C1652.
Streit, W.J., Kreutzberg, G.W. (1988) Response of endogenous glial cells to motor neuron degeneration induced by toxic ricin. J. Comp.
Neurol. 268: 248-263.
Svensjo, E., Boschcov, P., Ketelhuth, D.F., Jancar, S., Gidlund, M. (2003) Increased microvascular permeability in the hamster cheek pouch induced by oxidized low fensity lipoprotein (oxLDL) and some fragmented apolipoprotein B proteins. Inflamm. Res. 52: 215-220.
Thery, C., Chamak, B., Mallat, M. (1991) Free radical killing of neurons, Eur. J. Neurosci. 3: 1155-1164.
Tsimikas, S., Bergmark, C., Beyer, R.W., Patel, R., Pattison, J., Miller, E., Juliano, J., Witztum, J.L. (2003) Temporal increases in plasma markers of oxidized low-density lipoprotein strongly reflect the presence of acute coronary syndromes. J. Am. Coll. Cardiol. 41:
360-370.
Uno, M., Kitazato, K.T., Nishi, K., Itabe, H., Nagahiro, S. (2003) Raised plasma oxidised LDL in acute cerebral infarction. J. Neurol.
Neurosurg. Psychiatry. 74: 312-316.
Zhang, X., Blenis, J., Li, H. (1995) Require ment of serine phosphorylation for formation of STAT-promoter complexes.
Science. 267: 1990-1994.