兩種附生巢蕨在北台灣亞熱帶潮濕森林的空間分布特性
王相華1) 蘇聲欣2) 張勵婉3,4)
摘 要
附生巢蕨具有特別的生態棲位,也營造出特殊的懸空微棲地環境供樹生的無脊椎動物棲息,與熱 帶、亞熱帶陸域生物多樣性和生態系功能密切相關。本研究於「福山森林動態樣區」所位處的北台灣 亞熱帶潮濕森林內設置3.84 ha樣區並進行附生巢蕨調查。目的為經由分析台灣山蘇花、山蘇花兩種大 型巢蕨的空間分布特性,探討對光照、水分有強烈影響的地形及宿主(樹木)結構因子如何形塑這兩種 巢蕨的空間分布,以了解兩種巢蕨對光照、水分的適應性是否有差別。就地形而言,此兩種巢蕨多集 中分布在棲地水分較高的下坡、溪溝;就樹木宿主上的附生位置而言,兩種巢蕨多附生在宿主的主幹 上,而非樹冠層的枝條上,附生高度集中在離地2~4 m高度,鮮少分布在10 m以上。台灣山蘇花與山 蘇花的空間分布特性近似,即便是在鮮少發生乾旱的福山亞熱帶潮濕森林,台灣山蘇花、山蘇花仍然 多數分布在環境較為濕潤的地形,以及受光量較低的林冠下層樹木主幹。本文由光合作用、水分生理 特性及颱風對森林冠層擾動多個面向角度切入,簡要討論附生巢蕨在此一亞熱帶潮濕森林的空間分布 特性。
關鍵詞:巢蕨、附生植物、潮濕森林、空間分布、地形、樹木寄主。
王相華、蘇聲欣、張勵婉。2020。兩種附生巢蕨在北台灣亞熱帶潮濕森林的空間分布特性。台灣林業 科學35(3):205-16。
1) 林 業 試 驗 所 集 水 區 經 營 組 ,1 0 0 臺 北 市 南 海 路 5 3 號 D i v i s i o n o f F o r e s t Wa t e r s h e d M a n a g e m e n t , Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 10066, Taiwan.
2) 林 業 試 驗 所 森 林 經 營 組 ,100臺北市南海路53號 Division of Forest Management, Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 10066, Taiwan.
3) 林 業 試 驗 所 森 林 保 護 組 ,100臺北市南海路53號 Division of Forest Protection, Taiwan Forestry Research Institute. 53 Nanhai Rd., Taipei 10066, Taiwan.
4) 通訊作者 Corresponding author, e-mail:[email protected]
2020年6月送審 2020年11月通過 Received June 2020, Accepted November 2020.
Research paper
Spatial Patterns of Two Epiphytic Bird's-nest Ferns in a Moist Subtropical Forest, Northern Taiwan
Hsiang-Hua Wang,1) Sheng-Hsin Su,2) Li-Wan Chang3,4)
【Summary】
The aerial micro-habitats created by bird’s-nest ferns provide invertebrate habitats that are dis- tinct from the forest-floor environment, and they contribute to the biological diversity of tropical and subtropical forests. In this study, we set up and surveyed a 3.84-ha sample area in the Fushan Forest Dynamics Plot, which is located in a subtropical moist forest in northern Taiwan. We analyzed the spatial distributions of 2 native bird’s-nest ferns, Asplenium antiquum and A. nidus, to explore how topography and structural factors of host trees, which strongly affect the light conditions and water availability, shape the spatial distributions of these 2 species. The distributions of these 2 bird’s-nest ferns were mostly concentrated in downhill areas and near creeks with higher water availability. In terms of the epiphytic locations on host trees, the 2 bird’s-nest ferns were mostly attached to the trunk of host trees rather than to the branches in the canopy. The epiphytic height of both species was con- centrated at 2~4 m above the ground, and they rarely grew at a height of >10 m above the ground.
The spatial distribution characteristics of A. antiquum and A. nidus were similar. Even in the Fushan subtropical humid forest where drought events are rare, A. antiquum and A. nidus were mostly dis- tributed in downhill areas and near creeks with relatively high water availability and on the lower trunks of host trees with low light levels. The roles of photosynthesis, water physiological charac- teristics, and typhoon disturbances to the forest canopy in affecting the spatial patterns of bird’s-nest ferns in this subtropical moist forest are also discussed.
Key words: bird’s-nest fern, epiphyte, moist forest, spatial pattern, topography, host tree.
Wang HH, Su SH, Chang LW. 2020. Spatial patterns of two epiphytic bird's-nest ferns in a moist subtropical forest, northern Taiwan. Taiwan J For Sci 35(3):205-16.
緒 言
台 灣 的 天 然 林 蘊 育 著 豐 富 的 附 生 植 物 , 多樣的附生植物不僅有助於提升森林動、植物 多樣性,同時也提供極為重要的森林生態系功 能。Hsu等(2002)曾在北台灣福山試驗林進行 附生植物生物量、養分含量調查,發現附生植 物的生物量雖然僅佔該地區森林陸生植物地上 部生物量的1.2%,但卻有高達21~43%的陸生 植物葉部養分儲存在附生植物。由於多數附生 植物的生物量來自於葉部,且葉部養分(相較於
木 材 養 分 )可快速提供生態系中其他生物使用 (Matelson et al. 1993, Nadkarni and Matelson 1992),這表示附生植物在森林生態系養分循 環上佔有重要角色(Hsu et al. 2002)。維管束附 生植物(vascular epiphyte)多數體型較小、不 易受到關注,但種類約佔全球維管束植物種類 的10% (Gentry and Dodson 1987, Nieder et al.
2001),台灣植物誌第二版記錄4,077種台灣原
生維管束植物,其中附生植物有341種(8%),顯
示附生植物亦為台灣原生維管束植物種類的重 要組成。
台灣原生蕨類附生植物有 171種,17種為 鐵角蕨屬(Asplenium)植物(Hsu 2007),鐵角蕨 屬中的大型附生蕨類種類之成熟葉片長度可超 過1 m,根叢(rosette)直徑可達0.5 m (個人觀 察),俗稱鳥巢蕨(bird’s-nest fern)。其懸空葉 片、根叢創造出的特殊微棲地,可提供許多生 物利用,形成有別於喜好森林地面環境的無脊 椎動物群落,因此與陸生生物多樣性密切相關 (Ellwood and Foster 2004, Beaulieu et al. 2010, Díaz et al. 2012)。Ellwood and Foster (2004)的 研究更提出幾個重要結果:(1)附生巢蕨植物的 大小與其蘊藏的的無脊椎動物生物量之間有明 顯相關性;(2)附生巢蕨植株內所發現的無脊椎 動物生物量足以使整個婆羅洲雨林冠層中的無 脊椎動物的估算生物量倍增。
附 生 植 物 與 陸 地 呈 現 隔 離 的 狀 態 , 須 直 接面對離開地面的懸空環境考驗,主要受到光 照強度、水分供給兩項環境壓力 (Adibah and Ainuddin 2011)。附生植物可依據自身的光合作 用、耐旱生理特性,生存在不同光照、水分條 件下的懸空環境,從充滿陽光的開放樹冠到樹 木基部的陰濕環境(Hietz and Briones 2001)。台 灣山區地形崎嶇,致使地形起伏對森林環境的 光照及水分造成極大影響;一般而言,稜線、
上坡環境的地形遮蔽度低,有較強的光照、風 速,棲地較為乾燥,下坡、溪谷環境的地形遮 蔽度高,有較低的光照、風速,棲地較為濕潤 (Su 1987)。因此,我們預估除了附生在宿主上 的棲位條件外,地形因子也會對附生植物的空 間分布造成影響。
北 台 灣 福 山 試 驗 林 有 山 蘇 花 (Asplenium
antiquum)、台灣山蘇花(A. nidus)兩種鐵角蕨屬大型附生巢蕨。山蘇花主要分布在東亞溫帶及 亞熱帶,包括中國、日本、韓國和台灣,台灣 山蘇花廣泛分布於東南亞、澳大利亞東部、夏 威夷、波里尼西亞、印度和非洲東部等熱帶地 區(Huang et al. 1993, Wu et al. 2003)。在台灣,
山蘇花主要分布在中低海拔潮濕森林,台灣山 蘇花常見於低海拔潮濕森林(個人觀察),兩者對
溫度的適應性或有不同,但均同時生長於福山 試驗林附近的亞熱帶潮濕森林。本研究探討廣 泛分布在福山試驗林的兩種鐵角蕨屬大型巢蕨 (山蘇花、台灣山蘇花)的空間分布特性,重點在 分析對光照、水分梯度有強烈影響的地形、宿 主(樹木)空間結構因子如何形塑這兩種巢蕨的空 間分布,並了解兩種巢蕨對光照、水分適應性 是否有差別。
材 料 及 方 法
一、試驗地概述
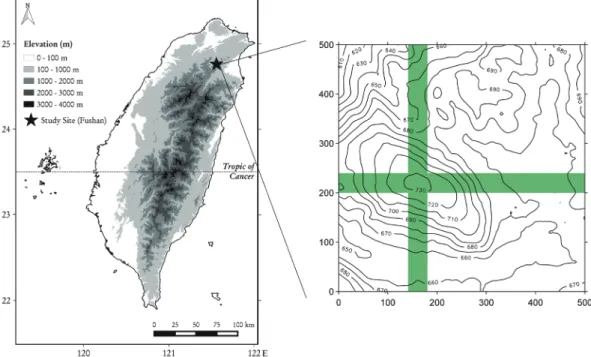
本 研 究 區 域 位 於 新 北 市 烏 來 區 , 鄰 近 福 山 植 物 園 (Fig. 1)。福山植物園設有氣象站,
1994~2019年資料顯示,該地區受冬季東北季風 及夏季颱風、熱對流影響,全年潮濕、多雨,
年平均相對濕度約95%,年均溫18.2度,年均 雨量4,202公釐,其天然植被主要屬於潮濕森林 (moist forest)。
巢 蕨 調 查 樣 區 設 在 福 山 森 林 動 態 樣 區 (Fushan Forest Dynamics Plot)內(Fig. 1)。該 樣區面積為25 ha、海拔高度在650~730 m之 間,首次每木調查完成於2004年,並於2009、
2014、2019年陸續完成三次樣區複查。該動態 樣區為一東西長500 m、南北寬500 m之正方形 樣區,又分割為 625個20×20 m之樣方,並以 南北向為Y座標,東西向X座標,用以定義出樣 方的平面座標編號(X, Y) (Su et al. 2007)。本研 究之巢蕨調查範圍以樣區山頂(所在地點為座標 (7, 10)、(7, 11)、(8, 10)、(8, 11) 4個樣方)為中 心,取樣區X座標7、8線(東西向寬度40 m,南 北向長度 500 m之帶狀範圍)與Y座標10、11線 (南北向寬度40 m,東西向長度500 m之帶狀範 圍)進行調查(Fig. 1),總計調查96個20×20 m樣 方,面積3.84 ha。
二、調查方法
調查期間為2013年3~6月,方法如下:
1.樣區、宿主資料
記錄巢蕨出現樣方座標編號,以及宿主編
號、胸徑、樹高。宿主胸徑採用胸徑尺測量樹
幹離地1.3 m高處的直徑,樹高採用伸縮測高桿 直接測量地面至樹冠頂端的高度。
2. 巢蕨物種
調 查 記 錄 出 現 的 物 種 是 山 蘇 花 或 台 灣 山 蘇花。在野外,區分山蘇花、台灣山蘇花的方 式為觀察葉背孢膜長度,山蘇花的孢膜較長,
從葉背中軸延伸至接近葉緣,台灣山蘇花的孢 膜較短,長度約為葉背中軸到葉緣長度的一半 (Huang et al. 1993)。又因鐵角蕨屬巢蕨的孕性 與植株大小相關(Tsai 2005, Huang et al. 2008),
因此本研究僅記錄叢徑、葉長較大而可觀察到 孢膜的有孕性成熟植株。
3. 附生位置
記錄巢蕨附生在樹木上的高度、區位、方 位。附生高度採用伸縮測高桿進行量測,直接測 量地面至巢蕨附生位置的高度。附生區位分為5 類,由下往上分別為樹木主幹(T)、主幹與一級 枝條交接處(TB1)、一級枝條(B1)、一級枝條與 二級枝條交接處(B1B2)、二級枝條(B2)。附生方
位採用指北針進行量測,以樹木主幹為中心,
量測巢蕨在樹木上的附生方位角,再轉換成北 (N)、東北(NE)、東(E)、東南(SE)、南(S)、西南 (SW)、西(W)、西北(NW)等8個方位。
三、地形數據處理和統計分析 1. 樣方地形運算
進行巢蕨調查的3.84 ha樣區位於25 ha森林 動態樣區內(Fig. 1),樣區曾進行細部地形測量,
因此已知每個20×20 m相交點的高程。我們採用 Su et al. (2010)方法,使用這些測量值以地理運 算方法導出20×20 m樣方的下列地形因子:
(1) 平均高程(Mean elevation):樣方的四個角 落的高程平均值。
(2) 坡度(Slope):由樣方中任意3個角落構成 的4個三角形平面之平均傾斜角(Harms et al. 2001)。
(3) 凹凸度指數(Index of convexity):樣方的 平均高程減去其周邊 8個20×20 m樣方的 平均高程。如為正值表示為凸起的地形,
如為負值表示為低凹的地形 (Valencia et
Fig. 1. Location map of the Fushan Forest Dynamics Plot and the 3.84-ha survey area (in
green) in the plot. Contours on the map are in meters.
al. 2004)。
(4) 坡向指數(Aspect index):本研究將坡向 定 義 為 坡 面 上 最 為 陡 斜 的 方 向 。 計 算 方 式是以樣方四個角落樁點之三維座標值,
以ArcGIS軟體進行坡向運算,以得到各 樣方之坡向值(θ)。由於坡向為一循環資 料 (circular data),數值由0°至360°形成循 環,所以本研究再將坡向以三角函數換算 成四個坡向指數,數值由-1至1:
(a) 北-南向指數(N-S) = cos(θ) (b) 東-西向指數(E-W) = sin(θ) (c) 東北-西南向指數(NE-SW)
= sin(θ+45°)
(d) 東南-西北向指數(SE-NW)
= sin(θ−45°)
2. 統計分析
(1)巢蕨在宿主上的附生位置
將兩種巢蕨於樹上附生高度,每2 m分1級 共6級,再加上5類附生區位及8個附生方位,
分別計算山蘇花及台灣山蘇花在宿主上附生的 高度、區位及方位之數量。為瞭解兩種巢蕨之 附生高度、附生區位分布之有無差異,以隨機 化(逢機)方法進行檢測(randomization test)。
Randomization test的虛無假設(H0)是認為兩組 變數之間並無顯著差異,因此樣本資料可以在 不同組別間互相隨機置換,詳細步驟為針對兩 個 要 檢 測 的 變 數 進 行 樣 本 間 的 隨 機 置 換 排 列 (permutation),並模擬999次,其中之虛無假設 (H0)為兩組變數是獨立不相關的;經過999次隨 機置換後,可計算出機率p值,p值為當虛無假 設為真之下,樣本平均值發生的可能性。當機率
p < 0.05,拒絕虛無假設,顯示兩種巢蕨附生高度、附生區位分布有顯著差異;
p > 0.05則顯示兩種巢蕨無顯著差異(Beh and Lombardo 2014)。
又由於附生方位為循環資料,故使用循環統計 (circular statistics)方法之沃森測試(Watson test)測 試兩種巢蕨附生分布的方位是否有明顯不同。
(2) 巢蕨分布的地形偏好
為 了 解 巢 蕨 ( 山 蘇 花 、 台 灣 山 蘇 花 ) 在 空 間 中 分 布 與 環 境 的 關 係 , 本 研 究 採 用 網 格 式
的 空 間 迴 歸 統 計 : 空 間 自 迴 歸 模 型 ( s p a t i a l autoregressive model, SAR),因取樣之樣方皆 為相鄰,使用空間迴歸統計模型可解決空間資 料所產生的空間自相關性,而對於自變數及依 變數之間的關係作更佳合理的解釋(Ver Hoef et al. 2018)。公式如下:Y = ρWY+Xβ+ε
依變數(Y):山蘇花、台灣山蘇花在各20×20 m 樣方的數量;
自變數(X):包含各項地形因子;
WY為空間變數,ρ和β為迴歸係數,ε是誤差。
由於各地形因子的單位並不相同,在資料分析 前,將各地形因子進行過標準化轉換,以便於 直接比較分析結果之迴歸係數值。
W Y ( 空 間 變 數 ) 為 計 算 空 間 權 重 矩 陣 ( s p a t i a l
weighting matrix),因本研究所取樣方緊緊相 連,且為東西方向及南北方向之規則取樣,本 研究採用Rook contiguity的鄰接矩陣(Getis and Aldstadt 2004)。
結 果
本 研 究 在 3 . 8 4 h a 調 查 區 內 的 2 5 2 棵 喬 木 上 記 錄 到 可 辨 識 種 類 的 山 蘇 花 2 5 4 株 、 台 灣 山 蘇 花 2 1 2 株 , 合 計 4 6 6 株 , 密 度 為 1 2 1 . 4 株 / h a ; 7 4 % 的 巢 蕨 附 生 於 1 0 ~ 4 0 c m 胸 徑 的 樹 上 , 又 以 2 0 ~ 3 0 c m 胸 徑 的 樹 木 所 附 生 數 量 最 多 , 佔 4 0 % , 附 生 樹 種 而 言 , 以 長 葉 木 薑 子 (Litsea acuminata) (26%)、紅楠 (Machilus thunbergii) (17%)、綠樟(Meliosma
s q u a m u l a t a ) ( 9 % ) 、 短 尾 葉 石 櫟 ( P a s a n i a harlandii) (8%)、錐果櫟(Cyclobalanopsis longinux) (7%)等樹種所佔比例最多。兩者的空間分布分析結果如下:
一、巢蕨在宿主上的附生位置
以附生高度而言,超過 80%山蘇花及台灣
山蘇花的附生在高度 2~8 m之間,又以2~4 m
高度較多,少量附生在2 m以下及8 m以上的高
度,10 m以上的高度僅記錄到2株(Fig. 2)。以附
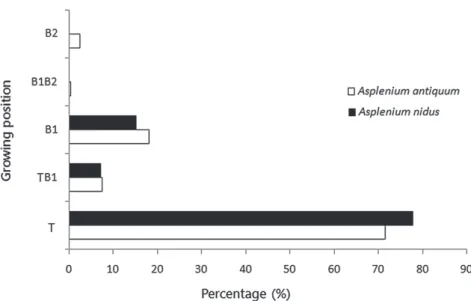
生位置而言,山蘇花和台灣山蘇花大多附生在
樹木的主幹(T),佔山蘇花的71 %、台灣山蘇花
的的78%,但也有約20 %的山蘇花及台灣山蘇 花附生在樹木的1級枝條(B1)和主幹、1級枝條 交 接 處 (TB1),僅有少數山蘇花附生在2級枝 條(B2)及1級枝條、2級枝條交接處(Fig. 3);
以附生在樹上的方位而言,山蘇花及台灣山蘇 花於各個方位的附生數量相當,但山蘇花在西
方 位 的 附 生 數 量 較 多 (Fig. 4)。隨機化檢測結 果顯示,山蘇花與台灣山蘇花在宿主上的附生 高度及附生區位並無顯著差異 (p > 0.05),此 外,循環統計方法之沃森測試(Watson test)結 果,顯示兩種巢蕨之附生分布方位亦無顯著差 異 (p > 0.1)。
Fig. 3. Growing position distributions of Asplenium antiquum and A. nidus on host trees. T, tree trunk; TB1, the junction of tree truck and the first layer of branches; B1, the first layer of branches; B1B2, the junction of the first and second layers of branches; B2, the second layer of branches.
Fig. 2. Height distributions of Asplenium antiquum and A. nidus on host trees.
二、地形對巢蕨分布的影響
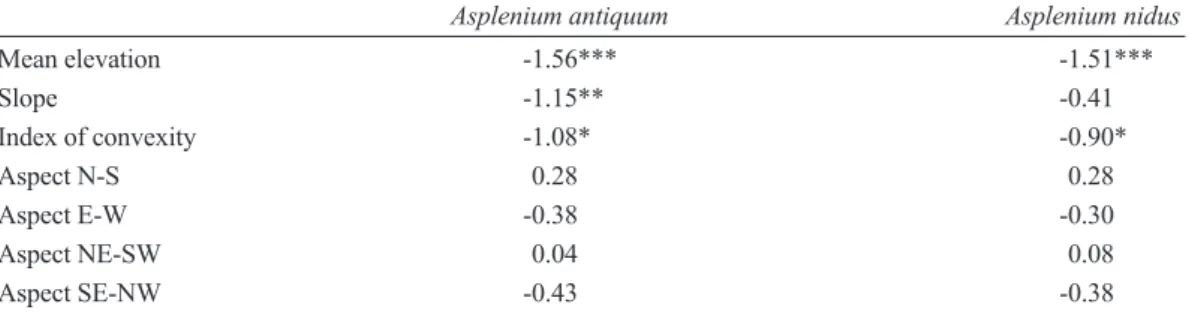
以 空 間 自 相 關 迴 歸 模 型 檢 測 山 蘇 花 、 台 灣山蘇花在樣區中分布與四種地形因子間的關 係(Table 1),結果顯示,海拔高度會對於兩種 巢蕨在樣區中之空間分布皆有顯著的影響 (p <
0.001),海拔高度越高(越接近稜線、山頂),兩 種巢蕨之數量越少;此外,凹凸度對於兩種巢 蕨的空間分布亦具有顯著之影響,地勢越凸起 的區域,巢蕨的數量越少;即兩種巢蕨多分布 於樣區內海拔高度較低的下坡、溪溝地形。另 且坡度僅影響山蘇花的數量,坡度越大數量越 少,並不影響台灣山蘇花在樣區中的分布 (p >
0.05)。而四種地形方位指數對於兩種巢蕨的空 間分布皆無顯著影響(Table 1)。
討 論
就地形而言,本研究調查的兩種大型原生 巢蕨(山蘇花、台灣山蘇花)均喜好分布於樣區內 海拔較低的下坡、溪溝,顯示即使調查區域位於 潮濕森林,山蘇花、台灣山蘇花也多數分布在棲 地水分相對較高的下坡、溪溝地形。除了反應稜 線至溪溝地形變化的海拔高度外,坡度、凹凸度 指數對於樣區兩種巢蕨的空間分布亦有影響,但 不如海拔高度顯著。檢測各地形因子的相關性結 果顯示,樣方海拔高度與凹凸度指數間具成正相 關,表示海拔較高處(稜線、上坡處)的凹凸度指
數通常是正值,微地貌多呈現上凸形態,海拔較 低處(下坡、溪溝處) 的凹凸度指數通常是負值,
微地貌多呈現下凹形態,而兩種巢蕨較喜好分布 於微地貌呈現下凹形態的下坡、溪溝區域。就巢 蕨附生在樹木宿主上的區位而言,本研究發現兩 種巢蕨大多附生在樹木的主幹上,側枝上的數 量較少;而巢蕨附生在樹上的離地高度以2~4 m 最常見,4~8 m 次之,10 m以上僅有兩株。研 究區域(福山森林動態樣區)曾進行樹冠層高度調 查,樹冠層高度集中在11~18 m之間,中位數為 14.5 m (Chen 2012)。比對上述資料後得知,研 究區域的巢蕨較少分布在樹冠層,尤其是樹冠層 的上方。Fayle等(2009)在馬來西亞Pasoh熱帶雨 林的研究也發現,台灣山蘇花的附生區位大多位 於樹木下層(63.4%)。太陽輻射是地表能量平衡 的主要驅動因子,也是影響微氣候的關鍵因素,
包括近地表溫度、蒸發量、濕度和土壤含水量等 (Bennie et al. 2008),地形變化會影響局部區域所 接受到的太陽輻射量,對特定點位的微氣候有重 要影響;受到地形遮蔽效應的影響,下坡、溪溝 陽光輻射量較低,屬於棲地水分較高的地形(Su 1987)。相同地,樹木冠層上方的受光量較高、
濕度較低,而樹木冠層下方的受光量較低、濕度 較大,屬於棲地水分較高的環境。
與樹木相比,蕨類植物的維管束系統較原 始,維管束管道直徑小,水分傳導速度較慢、
氣孔導度低,因光合作用需要經由氣孔進行氣
Fig. 4. Abundances of Asplenium antiquum and A. nidus across different aspects of host trees.
體交換,受限於氣孔導度較低,多數蕨類的最 大淨光合速率(maximal net photosynthetic rate) 及光飽和點(light saturation point)都較木本植 物低(Brodribb et al. 2005),因此蕨類植物通 常生長在林下光度較弱的生境中(Andrade and Noble 1997)。地生蕨類植物可以直接從土壤中 獲取水分 , 但附生蕨類卻要頻繁面對水分供應 短缺的狀況(Zotz and Hietz 2001)。就巢蕨的耐 旱生理而言,新加坡曾進行台灣山蘇花的耐旱 生理研究(Kluge et al. 1989),儘管此試驗最長 乾旱期的持續時間僅為5天,但卻推測台灣山蘇 花是一種耐旱的植物,並歸因於台灣山蘇花在 其混和根系及大量腐植質的根部碎片系統(root- debris system)中可以儲存足夠的水分,克服短 時期乾旱。然而,在強烈的季節性降雨的澳大 利亞東北部,長達幾週的乾旱並不罕見(Tracey 1982),此區域若發生長時間的乾旱會導致台灣 山蘇花枯死並掉落至地上,顯示其無法忍受長 期乾旱(Freiberg and Turton 2002)。Martin等人 (2004)曾測量福山試驗林30種苔蘚、蕨類、蘭 科附生植物的葉片滲透勢(osmotic potential),
發 現 多 數 附 生 植 物 的 葉 片 滲 透 勢 都 很 高 (> -1 MPa),其中山蘇花的葉片滲透勢為-0.94 MPa,
顯示多數附生植物無法採用降低葉片組織滲透 壓方式因應環境的乾旱。我們調閱了林業試驗 所出版的「林業試驗所福山分所氣象水文資料 (1982~1999)」(Lu et al. 2000),發現即使是在 北台灣福山試驗林的潮濕森林環境,在17年間 也經歷兩次月降雨量< 10 mm的乾旱期。由上 述研究結果綜合評估,此類偶發性的乾旱可能
會造成分布在稜線、上坡地形及樹冠層上方巢 蕨的生存壓力,並非其長期適存的環境梯度。
Zhang等(2010)在馬來西亞Pasoh熱帶雨林的研 究亦顯示,台灣山蘇花喜好分布在較為潮溼的 沼澤地(swampy areas)、不喜好分布在山坡地 (hilly areas)。
就 巢 蕨 的 光 合 作 用 生 理 而 言 , Zhang等 (2008)於西雙版納熱帶雨林測得台灣山蘇花的 最大淨光合速率為 3.09±0.03 µmol m
-2 s-1, 光 飽 和 點 為 2 7 1 . 1 9 ± 8 . 4 0 µ m o l m
- 2s
- 1photosynthetic photon flux density (PPFD),
光補償點(light compensation point)為3.94±
0.42 µmol m
-2s
-1PPFD;Wong等(2012)於台灣 南投地區測得培育在10%光量及全光環境下的 山蘇花的最大淨光合速率在4~5 µmol m
-2s
-1之 間,兩者並沒有明顯差別,顯示高光量並無法 提升山蘇花的光合速率。Kuo and Yeh (2015) 分析台灣 180種原生闊葉樹種光合潛力及耐陰 性,並將樹種最大淨光合速率區分為≥ 26.0、
25.9~21.0、20.9~15.0、14.9~12.5、< 12.5 µmol m
-2s
-1等5級;其中之先驅樹種等級,例如 血桐 (Macaranga tanarius)、蟲屎(Melanolepis
m u l t i g l a n d u l o s a ) , 最 大 淨 光 合 速 率 分 別 為31.9及31.0 µmol m
-2s
-1,而耐陰等級樹種如鐵 色 (Drypetes littoralis)、厚殼桂(Cryptocarya
chinensis)則為9.5及9.1 µmol m-2s
-1,都遠高於 Zhang等(2008)於西雙版納熱帶雨林測得台灣山 蘇花的最大淨光合速率(3.09 µmol m
-2s
-1),以及 Wong等(2012)在台灣南投測得的山蘇花最大淨 光合速率(4~5 µmol m
-2s
-1)。上述針對山蘇花、
Table 1. Regression coefficients of the spatial autoregressive model, which tested relationships between the abundances of bird’s-nest ferns (Asplenium antiquum and A.
nidus) and topographic variables (*** p < 0.001, ** p < 0.01, * p < 0.05)
Asplenium antiquum Asplenium nidus
Mean elevation -1.56*** -1.51***
Slope -1.15** -0.41
Index of convexity -1.08* -0.90*
Aspect N-S 0.28 0.28
Aspect E-W -0.38 -0.30
Aspect NE-SW 0.04 0.08
Aspect SE-NW -0.43 -0.38
台灣山蘇花及台灣原生木本植物光合作用研究 數據顯示,台灣山蘇花的耐陰性極高,樹冠上 層的強光照環境並不符合台灣山蘇花的光合生 理特性需求。
本 研 究 結 果 顯 示 山 蘇 花 、 台 灣 山 蘇 花 兩 種 巢 蕨 在 福 山 亞 熱 帶 潮 濕 森 林 的 空 間 分 布 特 性雷同,都喜好分布於環境較為潮濕的地形,
並 附 生 在 受 到 太 陽 輻 射 較 低 的 樹 幹 中 下 段 部 位 。 Fayle等(2009)在樹冠層高度達50~60 m 的 馬 來 西 亞 沙 巴 熱 帶 雨 林 研 究 發 現 , 兩 種 巢 蕨 (A. nidus及A. phyllitidis)所喜好附生的冠層 結構(canopy architecture)區域有明顯差異,
A. nidus多數分布在突出樹(emergent tree)及孔
隙 (gap)區域,對乾旱、強光的適應性較高;
相對的,
A. phyllitidis則喜好分布在相對濕涼的的中低層樹冠。但本研究結果顯示,山蘇花 (A. antiquum)、台灣山蘇花(A. nidus)兩種巢蕨 在福山亞熱帶潮濕森林的空間分布特性雷同,
都喜好分布於潮濕且受光量較低的樹木主幹或 較低高度處。本研究與Fayle等(2009)的研究結 果似乎呈現不同的結果,特別是台灣山蘇花 (A.
nidus)在宿主上的空間分布。Fayle等(2009)的研
究地點為馬來西亞沙巴熱帶雨林,並非颱風盛 行區域,而本研究地點是颱風盛行的區域,會 對森林樹冠造成經常性擾動,強勁風勢會對樹 冠層枝葉造成嚴重擾動(Mabry et al. 1998, Lin et al. 2011, Chang et al. 2020),可能致使巢蕨在 樹冠上層難以長時間穩定附著。再者,大量枝 葉被強風打落後,容易造成樹冠上層枝條的受 光量急劇增加,而依據前述的巢蕨光合生理特 性研究結果,太強的光照處並非巢蕨喜好的環 境棲位,因此不排除經常性的颱風擾動是導致 福山地區的巢蕨主要附生在樹木主幹及樹冠中 下層的重要原因之一。
目前對鐵角蕨屬巢蕨的生理研究資料非常 有限。從前述的光合生理前人研究數據來看,山 蘇花、台灣山蘇花兩者都是耐陰性很高的附生植 物,但目前並沒有針對山蘇花、台灣山蘇花等 巢蕨的溫度、濕度適應性之相關研究文獻。若由 兩者的全球分布、在台灣的海拔分布差異來看,
山蘇花似乎比台灣山蘇花更能適應低溫的環境,
但目前尚未有生理研究文獻資料可以證實。就 濕度而言,雖然蕨類植物多數適合生長在高濕 度的環境,但前述Fayle等(2009)的研究指出:
「不同巢蕨對濕度的喜好會有差異」,而目前 也沒有生理研究文獻資料能證實山蘇花、台灣 山 蘇 花 對 濕 度 的 適 應 性 是 否 有 顯 著 差 別 。 因 此,目前本研究之結果僅能說明這兩種巢蕨都 適合生長在北台灣潮濕森林、海拔高度約700 m 上下的陰濕環境。受限於研究區域的環境梯度 較窄,難以反應兩者確切的溫、濕度適應生理 有何異同,如要釐清此一問題,則有待未來進 行更大環境梯度或是跨區域的研究。
結 論
本研究經由調查,分析山蘇花、台灣山蘇 花兩種大型巢蕨在福山森林動態樣區內的空間分 布特性。就地形而言,兩種巢蕨多集中分布在下 坡、溪溝區域;就樹木宿主上的附生位置而言,
兩種巢蕨多附生在宿主的主幹上,附生高度集中 在離地2~4 m之間,鮮少分布在10 m以上;山蘇 花、台灣山蘇花這兩種巢蕨在福山亞熱帶潮濕森 林的空間分布特性雷同。本研究推測除了光合作 用及水分生理特性外,颱風擾動對福山地區兩種 巢蕨的空間分布可能造成重要影響。
引 用 文 獻