行政院國家科學委員會專題研究計畫 成果報告
登革病毒感染不同抗原呈現細胞對其免疫及凝血功能的影
響(3/3)
計畫類別: 個別型計畫 計畫編號: NSC92-2314-B-006-094- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立成功大學醫學檢驗生物技術學系 計畫主持人: 葉才明 計畫參與人員: 孫睦傑 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 10 月 5 日
Dengue Virus Induces Human Neutrophil Activation and Necrosis
Mu-Jie Suna, Lien-Cheng Chenb, Huan-Yao Leic,Hsiao-Sheng Liuc, Yee-Shin Linc,
Shun-Hua Chenc, Tsuey-Yu Chang d,and Trai-Ming Yeha
a Department of Medical Technology, National Cheng Kung University, b Institute of
Basic Medical Sciences, National Cheng Kung University, c Department of
Microbiology and Immunology, National Cheng Kung University, d Department of
Parasitology, National Cheng Kung University, Tainan, Taiwan, ROC
Short title: Dengue virus and neutrophil Full address:
Dr. Trai-Ming Yeh
Department of Medical Technology, College of Medicine, National Cheng Kung
University, Tainan 701, Taiwan (ROC)
Tel. +886 6 2353535 ext. 5778, Fax +886 6 236 3956, E-Mail:
Abstract
Neutrophils are known to play important role in host defense against pathogens.
Even though neutropenia is common during dengue virus (DV) infection, the role of
neutrophils during DV infection is unclear. In this study, freshly isolated human
neutrophils were incubated with DV to understand the effects of DV and neutrophils
on each other. DV induced the secretion of chemokines such as interleukin 8,
macrophage inflammatory protein (MIP -1α, MIP -1β) and the release of granular enzyme, myeloperoxidase 18 h after incubation. DV-induced neutrophil activation
was further supported by the increased expression of CD11b/CD18 (Mac-1) and
Toll-like receptor 4 on the surface of DV-stimulated neutrophils. In addition, the
phagocytic ability to FITC-carboxylate microspheres (FITC-beads) was increased in
DV-stimulated neutrophils . Co-localization of DV E-antigen with FITC -beads was
found in these neutrophils , indicating that DV may enter neutrophils through
phagocytosis. However, no viral progeny was detected in the supernatants and cell
lysate of neutrophils 18 h after incubation. In addition, the viabilities and apoptosis
of neutrophils were decreased after incubation with DV for 18 h. On the other hand,
DV-stimulated neutrophils was increased in a dose-dependent manner. Therefore,
our results suggest that phagocytosis of DV by neutrophils may help to clear DV
infection. However, DV-induced neutrophil activation and necrosis may also
Introduction
Dengue viruses are mosquito -borne flaviviruses which are widely spread in
tropical and subtropical countries [11]. DV causes a spectrum of disease, from a
relatively benign febrile disease called dengue fever (DF) to a life-threatening illness
called dengue hemorrhage fever and shock syndrome (DHF/DSS) [8]. DF is
characterized by biphasic fever, headache, muscle pain, and rash. DHF/DSS
occurs in a small percentage of dengue patients which is characterized by
hemorrhage and vascular leakage accompanied by severe thrombocytopenia and
hemoconcentration. The pathogenesis of DHF/DSS has not been fully elucidated.
However, abnormal immune responses may be responsible for the progress of DF
to DHF/DSS [16].
Even though DV can infect many different cell types in vitro such as monocytes,
hepatoma, and endothelial cells [17, 12], its primary host target cell is thought to be
monocytes and macrophages. These cells are responsible for the dissemination of
the virus in blood after its initial entry via the mosquito vector [10]. However, the role
not been fully elucidated. Since neutrophils bear an arsenal of weapons against
infection, neutrophils may contribute to the clearance of viral infection as well as to
the pathogenesis of tissue damage [29]. Early clinical and laboratory indicators
suggest neutropenia is a common phenomenon during dengue infection [2, 15].
Furthermore, neutrophil granule enzyme elastase has been found to be significantly
elevated in the sera of dengue patients with shock than in patients without shock
[14]. Therefore, neutrophil activation and degranulation may involve in dengue virus
induced immunopathological process. In this study, the effects of DV and
neutrophils on each other were studied using freshly isolated human neutrophils
incubated with DV in vitro. Results from these studies suggest neutrophils may play
an important role in the clearance of DV as well as in the pathogenesis of DV
Materials and Methods
Reagents
The following reagents were from the indicated sources: fetal calf serum (FCS),
Dulbecco’s modification of Eagle’s minimum essential medium (DMEM),
penicillin-streptomycin-glutamine, and Hank’s balanced salt solution (HBSS) (Gibco
Life Technologies, Grand Island, NY), FITC-carboxylate microspheres (FITC-beads)
(Polysciences, Inc., Warrington, PA), lipopolysaccharide (LPS) of E. coli 0111: B4
(Difco Laboratories, Detroit, MI) , RBC lysis solution (Genera, Minneapolis, MN),
Histopaque-1.077 lymphocyte separation medium and FITC -labeled mouse
monoclonal anti-human CD11b antibody and goat-polyclonal anti-human TLR4
antibody (Santa Cruz Biotechnology, CA), FITC-conjugated mouse-polyclonal
anti-goat IgG antibody (Jackson ImmunoResearch, West Grove, PA), biotin labeled
mouse-monoclonal anti-DV envelop protein (E), non structural protein 1 (NS-1)
antibodies (Dr. Lei’s laboratory), and PE-conjugated streptoavidin (Sigma, St. Louis,
MO).
Dengue type 2 virus strain (PL0146) was propagated in C6/36 cells from
American Type Culture Collection with DMEM supplemented with 2% FCS and
antibiotics. The virus culture medium was harvested after incubation for 5 days, cell
debris was removed by centrifugation, and the virus supernatant was aliquoted and
stored at -70 °C until used. DV titers were determined on BHK-21 cultures under
nutrient agarose and expressed as PFU per milliliter. The UV inactivation of DV
(UV-DV) was conducted in a Stratagene UV-stratalinker apparatus using 1800 mJ
of UV radiation. The heat inactivation of DV (H -DV) was carried out by heating at
56°C for 30 min. No plaque was found when UV-DV or H-DV was used to infect
BHK cells. A mock DV stock was prepared in exactly the same manner as the DV
preparation, with the exception that the C6/36 cells were not infected with DV
(C6/36).
Neutrophil Isolation
Neutrophils were isolated from the peripheral blood of healthy donors. The
blood was collected in citrated tubes and mixed with 5% dextran i n a ratio of 10:1 to
containing the buffy coat was layered over an equal volume of Histopaque-1.077
lymphocyte separation media and centrifuged (x 650g) at 25 °C for 25 min. The
erythrocytes in the resulting pellet were lysed with RBC lysis buffer and removed by
centrifugation (x 450g) at 25°C for 10 min. Purified neutrophils were washed 3 times
with HBSS and re-suspended in RPMI 1640 plus 10% FCS. The cell preparation
contained 98% neutrophils, with less than 1 platelet/100 neutrophils as determined
by cytospin and Liu’s stain after microscopic counting.
Stimulation of Neutrophils
Neutrophils (2x106) were stimulated with either live DV at multiplicity of
infection (MOI) as indicated, UV-DV (with the same MOI as live DV), or 200 ng/ml
LPS and incubated in 1 ml of RPMI 1640 supplemented with 10% FCS and 2 mM
L-glutamine at 37°C in a 5% CO2 incubator. In addition, cells treated with medium
alone or C6/36 supernatant were also included as controls. After 18 h, the cell
suspensions were centrifuged, and the supernatants were analyzed by ELISA for
chemokines and myeloperoxidase (MPO). In some experiments, the cells were
electron microscopy examination.
Enzyme-Linked Immunosorbent Assay (ELISA)
IL -8, MIP-1α, MIP-1β, and MPO were assessed by commercially available ELISA kits (R&D Systems, Minneapolis, MN) according to the manufacturer’s
instructions.
CD11b/CD18 (Mac-1) and TLR4 Staining
Human neutrophils (2 x 104) with or without DV (MOI of 1) or LPS (200 ng/ml)
stimulation were cytospined onto the slides and stained with FITC-labeled mouse
anti-human CD11b monoclonal antibody in a final concentration of 20 µg/ml for 1 h
at RT, and the cells were washed with Tris-buffer and observed by a confocal
spectral microscope. The slides stained with goat anti-human TLR4 antibody (20
µg/ml) for 1 h at 4℃ , were then washed twice and incubated with FITC conjugated
mouse anti-goat IgG antibody (4 µg/ml). After incubation for 30 min at 4 °C in the
dark, the cells were washed with Tris-buffer and observed by a confocal fluorescent
(FACSCalibur, Becton Deckinson, USA).
Electron Microscopy
Neutrophils after incubation with DV were fixed at room temperature for 1 h
using a freshly prepared 3% solution of glutaraldehyde in warm cacodylate buffer.
Fixed cells were resuspended in warm liquid agarose and chilled immediately on ice.
After cutting the agarose into small cubes, cells were postfixed in 2% osmium
tetroxide, dehydrated in a graded series of ethanol and embedded in resin (EMS,
SPUR’S Kit, Washington) according to standard protocols. Ultrathin sections were
cut with ultramicrotome (Reichert-Jung) and contrasted with saturated aqueous
uranyl acetate and lead citrate at room temperature. Specimens were then
observed under a transmission electron microscope (JEOL JEM-1200EX, Tokyo,
Japan) at 100 kV.
Phagocytosis of Neutrophils
The uptake of FITC -carboxylate microspheres (FITC-beads) by neutrophils
MOI of 1 or 5 for 1 h. After pre-incubation, 2 µl of FITC-beads (0.75 µm, 7x106 beads/µl) were added and incubated for 30 min at 37°C. Neutrophils were washed with ice cold 1.6% BSA in PBS to remove free FITC -beads and fixed with 100 µl 4% paraformaldehyde in PBS for 20 min at 4℃ . Some cells were permeabilized with
0.1% saponin in PBS and incubated with biotin labeled monoclonal antibody against
dengue virus E protein at a final dilution of 1:60 for 1 h. The cells were then washed
twice with permeabilization solution and incubated with PE-conjugated streptoavidin
at a final dilution of 1:100 for 30 min at RT in the dark. Fluorescence pattern of
neutrophils was observed by confocal microscopy (Leica TCS SP2, Germany) and
analyzed by dual-laser FACS Calibur (BD Biosciences, Mountain View, CA) using
CellQuest software (BD Biosciences).
Assessment of Neutrophils Viability
Assessment of neutrophil viability was made after incubation neutrophils (2x105)
with LPS (200 ng/ml), DV or UV-DV at MOI of 1 for 18 h. After incubation, cells were
washed and stained with trypan blue or propidium iodide (PI). The number of
cytometry unit. Neutrophils that did not take up trypan blue or PI were counted as
viable. The percentage viability of neutrophils was obtained by dividing the number
of viable cells by the total number of cells. In addition, cell viability was detected
using an XTT kit (Boehringer Mannheim Biochemicals, Indianapolis, IN). Briefly,
cells (2 x 105 cells/well) were incubated with 50 l of the XTT labeling mixture (final
XTT concentration, 0.3 mg/ml). The microtiter plate was further incubated for 2-4 h
at 37℃ in 5% CO2. The spectrophotometrical absorbance of the sample was
measured at 450 nm using a Vmax ELISA reader (Molecule Devices, Menlo Park,
CA).
Neutrophil Apoptosis Assays
The percentage of apoptotic neutrophils were assessed by the terminal
deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling (TUNEL)
assay as well as annexin V stain. Neutrophils (2x105) were incubated with LPS (200
ng/ml), DV or UV-DV at MOI of 1 for 18 h. In addition, some cells were treated with
C6/36 supernatants (C6/36), heat-inactivated DV (H-DV) as negative control. After
for 10 min at room temperature and permeabilized with 70% ethanol at -20℃. The
percentages of apoptotic cells were assessed by TUNEL reaction using ApoAlert
DNA Fragmentation Assay Kit (CLONTECH) according to the manufacture’s
instruction and then analyzed by flow cytometry. In addition, apoptosis was also
assayed by binding annexin-V-FITC to the exposed phosphatidylserine of apoptotic
cells with the Human annexin V Apoptosis Kit (Bender MedSystems) and then
analyzed by flow cytometry.
Detection of Lactate Dehydrogenase
The release of cytoplasmic enzyme lactate dehydrogenase (LDH) of
DV-stimulated neutrophils was measured using a commercial kit (CytoTox Non-
Radioactive Cytotoxicity Assay; Promega). Briefly, Neutrophils (2x105) were
incubated with DV at different MOI as indicated, LPS (200 ng/ml) or without
stimulation at 37℃ for 18 h. The supernatants were collected and assayed for LDH.
Cytotoxicity in experimental samples was measured as % LDH release as
Statistical Analysis
Data are expressed as mean ± SD. The significance of the difference between
the test and the control groups was analyzed using Student’s t-test and the
Results
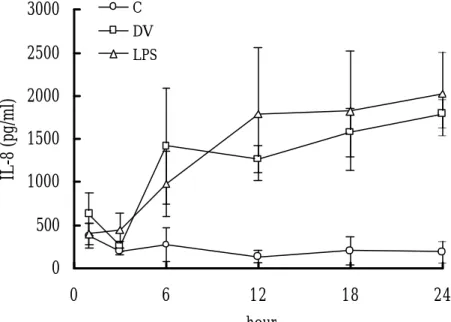
Kinetics of IL-8 Production in Neutrophils Induced by DV Stimulation
Human neutrophils were incubated with DV for increasing periods of time and
IL -8 production was assessed by ELISA. Figure 1 shows the results from one of
these experiments. Freshly isolated neutrophils produced low levels of IL -8. These
levels were not significantly changed by incubation in the medium alone for up to 24
h. However, a time-dependent increase in the secretion of IL -8 was found in both
DV and LPS stimulated neutrophils. Significant level of IL -8 was detected 6 h after
incubation and reached the peak level between 12 h and 24 h, the longest period of
time tested. A dose-dependent IL-8 production was also found when different MOIs
of DV were used to stimulate neutrophils (data not shown).
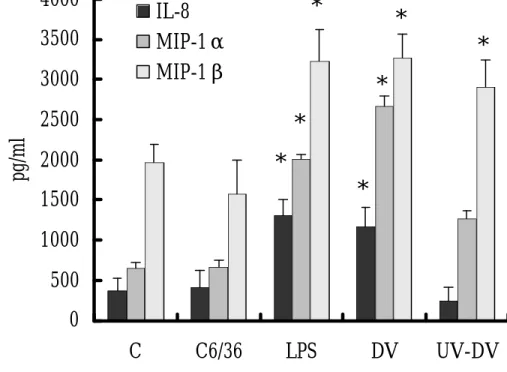
Comparison of IL-8, MIP-1α and MIP-1β Release of Neutrophils Induced by
Live and UV-inactivated DV
The concentrations of IL-8, MIP -1α and MIP -1β in the culture supernatants of neutrophils after incubation with DV or LPS for 18 h are shown in Figure 2.
produced only a small amount of IL -8 and MIP -1α while DV or LPS stimulation significantly increased the release of IL-8 by 2 to 3 fold (1000-1500 pg/ml) and
MIP-1α by 4 to 5 fold (2000-2500 pg/ml). UV inactivation of DV abolished the ability of DV to induce IL-8 and MIP-1α secretion in neutrophils. On the other hand, there was a spontaneous release of MIP-1ß in the control neutrophils without any
stimulation or with C3/36 medium (1500-2000 pg/ml). While DV or LPS stimulation
could further increase the release of MIP -1ß to 3000-3500 pg/ml, UV inactivation of
DV did not significantly reduce its activity to induce MIP -1β release of neutrophils (fig. 2).
Neutrophil Degranulation Induced by Both Live and UV-inactivated DV
Since myeloperoxidase (MPO) is one of the enzymes in the primary granules of
neutrophils, we used MPO as a marker to detect the degranulation of the
neutrophils. After 18 h of stimulation, MPO levels in the culture supernatants were
measured. DV and LPS induced significant amount of MPO release (117 ± 9 ng/ml
and 106± 27 ng/ml, respectively) from neutrophils compared to the un-stimulated
viable DV is required to stimulate neutrophil degranulation, neutrophils were
stimulated with UV -DV. Inactivation of DV with UV radiation did not significantly
abate MPO release from neutrophils (101.5± 1.2 ng/ml), in comparison with viable
DV.
Increase the Expression of CD11b and TLR4 on the Surface of DV-stimulated Neutrophils
To further confirm the activation of neutrophils induced by DV stimulation, the
surface expressions of adhesion molecule CD11b and signal-tranducing molecule
TLR4 of DV-stimulated neutrophils were analyzed by fluorescent antibodies and
confocal fluorescent microscopy. DV and LPS significantly increased the expression
of CD11b and TLR4 on the surface of neutrophils (fig. 3). The expression of CD11b
on neutrophils was further analyzed by flow cytometry. About 2% of the control
neutrophils were CD11b positive before stimulation. LPS, DV and UV-DV
stimulation induced CD11b positive neutrophils to 59.9%, 37.8% and 25.2%,
Infection and Morphological Changes of DV-stimulated Neutrophils
After 18 h of incubation with DV, about 30 .4 % of neutrophils were DV E protein
positive compared to only 2.6% in mock-treated neutrophils as detected by anti-DV
E protein antibody and flow cytometry. However, no viral progeny was detected in
the supernatants or cell lysate of neutrophils by plaque assay. In addition, no NS-1
protein can be detected in these cells using anti-NS-1 antibody (data not shown),
indicating DV did not replicate in neutrophils . When neutrophils were cytospined
onto slides and stained with Liu’s solution or fixed for electron microscopy
examination 18 h after incubation, more vacuoles and less granules were found in
DV-stimulated neutrophils than in C6/36-treated cells (fig. 4).
Phagocytosis and Co-localization of DV and FITC-beads in Neutrophils
To assess the phagocytic activity of neutrophils, FITC-beads were incubated
with neutrophils with or without pre-incubation with DV. FITC -beads were detected
in 29.9% of un-stimulated neutrophils as determined by flow cytometry (fig.5). DV
stimulation of neutrophils increased the percentage of neutrophils with FITC-beads
(fig. 5). About 48% and 62% of neutrophils were positive for FITC -beads in the
presence of DV at MOI of 1 and 5, respectively. The fluorescence intensities of
DV-stimulated cells were also significantly increased, indicating an increase in the
number of FITC-beads in these cells . Using PE-conjugated anti -DV E protein
antibody and confocal microscopy, co-localization of DV E antigen and FITC-beads
was found in DV-stimulated neutrophils as indicated by the appearance of yellow
dots (fig. 6). However, there were a few green and red dots in DV-stimulated
neutrophils also; indicating some of the neutrophils contained only DV or
FITC-beads.
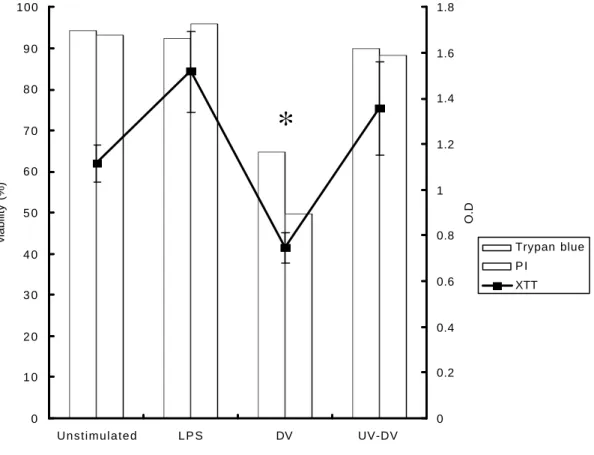
Decrease of Neutrophil Viability and Apoptosis in the Presence of Live but not
UV-inactivated DV
The cell viability of neutrophils after DV stimulation was measured by three
different methods, trypan blue stain, PI staining followed by the flow cytometry
analysis and XTT assay. Both trypan blue and PI stain, which mainly detect the
changes of membrane permeability, showed similar results (fig. 7). No significant
neutrophils (greater than 90%) were still survived after 18 h of incubation in vitro
without any stimulation or stimulated with LPS or UV-DV. However, the survival rate
of neutrophils was reduced to 50-60% in the p resence of live DV (fig. 7). Similar
results were also found in XTT assay, which mainly detect mitochondria enzyme
activity. The survival rate of DV-stimulated neutrophils was significantly decreased
after 18 h of incubation as compared with the controls (fig. 7). To further understand
the mechanism of decreasing the viability of neutrophils in the presence of DV, two
different methods were used to measure the occurrence of apoptosis (annexin V
and TUNEL stains). There was no significant change in the apoptotic cells after 3 h
of incubation in all groups. However, a spontaneous apoptosis of un-stimulated
neutrophils (about 50% by annexin V stain, and 35% by TUNEL) was detected after
incubation for 18 h (fig. 8). Incubation of neutrophils with C6/36, UV-DV or H-DV for
18 h did not change the percentage of apoptotic cells significantly. However,
incubation of neutrophils with LPS or live DV significantly decreased apoptotic cells
to 20-30% (fig. 8). In all groups, annexin V stain seemed to be able to detect more
Increase of Neutrophils Necrosis in the Presence of Live DV
Cellular necrosis was detected by the release of cytoplasmic enzyme LDH. After
12 h of incubation with DV, significant amounts of LDH were released by neutrophils
compared to uninfected cells or LPS-stimulated neutrophils. About 20% and 40% of
Discussion
Neutrophils constitute the first line of host defense against infectious agents
and exert a significant influence on the outcome of infection. In this study, we
demonstrated that a variety of chemokines were released from DV- but not
UV-DV-stimulated neutrophils. Chemokines are important mediators of the
immune system with primary chemotactic properties. By recruiting phagocytes as
well as lymphocytes to infection sites, chemokines connect nonspecific with
specific compartments of the immune system and therefore, play a pivotal role in
developing a rapid, focused, and effective immune response [25]. IL-8 released
by DV-stimulated neutrophils can recruit more neutrophils to the local infected
site and induce neutrophil adherence to vascular endothelium and extravasation
to tissues [6, 20], while MIP -1α and MIP-1β induced by DV-stimulated neutrophils can attract other immune cells such as monocytes, T cells, basophils to the
infected lesion [26, 31, 32]. All these immune cells are potential targets for DV
infection, which in turn can release more inflammatory cytokines to amplify the
elevated levels of IL-8, MIP -1α and MIP-1β are found in dengue patients [23, 30]. In addition, MIP -1α can also inhibit hematopoietic progenitor cell growth, which may account for some of the hematological disorders associated with dengue
illness [22]. DV stimulation of neutrophils also induced MPO release, which is a
marker for the degranulation of neutrophil primary granules. This is consistent
with the results from previous reports which showed that another granular
enzyme, elastase was significantly increased in DHF patients with shock [14].
The activation of neutrophils induced by DV was further supported by the
increased expression of C D11b and TLR4 in DV-stimulated neutrophils. The
increased expression of CD11b/CD18 (Mac-1) integrin on the surface of
DV-activated neutrophils may enhance the binding of neutrophils to ICAM-1
positive endothelium [21, 24, 28]. On the other hand, the significance of TLR4
expression induced in DV-stimulated neutrophils is unclear, except to reflect the
status of neutrophil activation. TLR4 plays a very important role in the signal
transduction induced by LPS and serves as a cell-surface co-receptor for CD14,
leading to LPS-mediated NF-κB activation and subsequent cellular events [4]. In a previous study, it has been shown that LPS can i nhibit DV infection of human
monocytes/macrophages by preventing virus entry via a CD14-dependent
mechanism [3]. Whether TLR4 may involve in the binding and signal transduction
induced by DV requires further study.
In this study we also demonstrated that DV was taken up by neutrophils
through phagocytosis and was presumably degraded inside the cell
compartments. This result is consistent with the previous report which found that
neutrophils did not become infected and replicate DV [33]. Similar phagocytosis
and degradation of DV have been previously reported in other human
phagocytes such as kupffer cells and monocytes [7, 19]. However, the
mechanisms of viral clearance by these phagocytes are unclear; probably both
oxygen-dependent and independent pathways are involved [5]. In striking
contrast to previous reports which indicate the stress of DV infection induced
apoptosis of many different cells [1, 7, 13, 18], we report that DV induced human
neutrophils undergo necrosis. This is based on the following observations that
the survival rate was decreased, while less apoptosis was found in DV-stimulated
neutrophils as compared to un-stimulated neutrophils. In addition, the release of
dose-dependent manner after incubation with DV. Normally, neutrophils are
considered to be short-lived, terminally differentiated cells undergo spontaneous
apoptosis (programmed cell death) in vitro if not appropriately stimulated [9, 27].
Apoptosis, which in contrast to necrosis, maintains the plasma membrane
integrity of neutrophils. This may explain why about 90% of un-stimulated
neutrophils were viable as detected by trypan blue or PI, yet a 35-50%
spontaneous apoptosis were detected in these cells at the same time. Cells
undergo apoptosis can avoid inflammatory reactions due to the release of
tissue-injurious granule contents. On the other hand, release of neutrophils’
formidable arsenal of protease and other toxic intracellular contents into tissues
can create significant damage, prolonging the inflammatory response. Taken
together all these results, we think neutrophils may play an important role in the
clearance of DV at first place. However, excessive activation and necrosis of
neutrophils induced by DV may also contribute to the pathogenic development
Acknowledgments
This work was supported by grants NSC90-2320-B006-058 from National
Science Council and grant NHRI-CN-CL-8901P from National Health Research
References
1. Avirutnan P, Malasit P, Seliger B, Bhakdi S, Husmann M. Dengue virus
infection of human endothelial cells leads to chemokine production,
complement activation, and apoptosis. J Immunol 161:6338-6346;1998.
2. Butthep P, Bunyaratvej A, Bhamarapravati N. Dengue virus and endothelial
cell: a related phenomenon to thrombocytopenia and granulocytopenia in
dengue hemorrhagic fever. Southeast Asian J Trop Med Public Health 24
Suppl 1:246-249;1993.
3. Chen YC, Wang SY, King CC. Bacterial lipopolysaccharide inhibits dengue
virus infection of primary human monocytes/macrophages by blockade of
virus entry via a CD14-dependent mechanism. J Virol 73:2650-2657;1999.
4. Chow JC, Young DW, Golenbock DT, Christ WJ, Gusovsky F. Toll-like
receptor-4 mediates lipopolysaccharide-induced signal transduction. J Biol
Chem 274:10689-10692;1999.
5. Cohen MS. Molecular events in the activation of human neutrophils for
microbial killi ng. Clin Infect Dis 18 Suppl 2:S170-179;1994.
Neutrophil-activating protein 1/interleukin 8 stimulates the binding activity of
the leukocyte adhesion receptor CD11b/CD18 on human neutrophils. J Exp
Med 171:1155-1162;1990.
7. Espina LM, Valero NJ, Hernandez JM, Mosquera JA. Increased apoptosis
and expression of tumor necrosis factor-alpha caused by infection of cultured
human monocytes with dengue virus. Am J Trop Med Hyg 68:48-53;2003.
8. Gubler DJ. Dengue and dengue hemorrhagic fever. Clin Microbiol Rev
11:480-496;1998.
9. Hachiya O, Takeda Y, Miyata H, Watanabe H, Yamashita T, Sendo F.
Inhibition by bacterial lipopolysaccharide of spontaneous and
TNF-alpha-induced human neutrophil apoptosis in vitro. Microbiol Immunol
39:715-723;1995.
10. Halstead SB, O'Rourke EJ, Allison AC. Dengue viruses and mononuclear
phagocytes. II. Identity of blood and tissue leukocytes supporting in vitro
infection. J Exp Med 146:218-229;1977.
11. Henchal EA, Putnak JR. The dengue viruses. Clin Microbiol Rev
12. Huang YH, Lei HY, Liu HS, Lin YS, Liu CC, Yeh TM. Dengue virus infects
human endothelial cells and induces IL -6 and IL-8 production. Am J Trop
Med Hyg 63:71-75;2000.
13. Jan JT, Chen BH, Ma SH, Liu CI, Tsai HP, Wu HC, Jiang SY, Yang KD, Shaio
MF. Potential dengue virus-triggered apoptotic pathway in human
neuroblastoma cells: arachidonic acid, superoxide anion, and NF-kappaB
are sequentially involved. J Virol 74:8680-8691;2000.
14. Juffrie M, va n Der Meer GM, Hack CE, Haasnoot K, Sutaryo, Veerman AJ,
Thijs LG. Inflammatory mediators in dengue virus infection in children:
interleukin-8 and its relationship to neutrophil degranulation. Infect Immun
68:702-707;2000.
15. Kalayanarooj S, Vaughn DW, Nimmannitya S, Green S, Suntayakorn S,
Kunentrasai N, Viramitrachai W, Ratanachu-eke S, Kiatpolpoj S, Innis BL,
Rothman AL, Nisalak A, Ennis FA. Early clinical and laboratory indicators of
acute dengue illness. J Infect Dis 176:313-321;1997.
16. Lei HY, Yeh TM, Liu HS, Lin YS, Chen SH, Liu CC. Immunopathogenesis of
17. Lin YL, Liu CC, Lei HY, Yeh TM, Lin YS, Chen RM, Liu HS. Infection of five
human liver cell lines by dengue-2 virus. J Med Virol 60:425-431;2000.
18. Marianneau P, Cardona A, Edelman L, Deubel V, Despres P. Dengue virus
replication in human hepatoma cells activates NF -kappaB which in turn
induces apoptotic cell death. J Virol 71:3244-3249;1997.
19. Marianneau P, Steffan AM, Royer C, Drouet MT, Jaeck D, Kirn A, Deubel V.
Infection of primary cultures of human Kupffer cells by Dengue virus: no viral
progeny synthesis, but cytokine production is evident. J Virol
73:5201-5206;1999.
20. Matsushima K, Oppenheim JJ. Interleukin 8 and MCAF: novel infla mmatory
cytokines inducible by IL 1 and TNF. Cytokine 1:2-13;1989.
21. Moreland JG, Fuhrman RM, Pruessner JA, Schwartz DA. CD11b and
intercellular adhesion molecule-1 are involved in pulmonary neutrophil
recruitment in lipopolysaccharide-induced airway disease. Am J Respir Cell
Mol Biol 27:474-480;2002.
22. Murgue B, Cassar O, Deparis X, Guigon M, Chungue E. Implication of
haematopoietic progenitor growth by dengue virus. J Gen Virol
79:1889-1893;1998.
23. Raghupathy R, Chaturvedi UC, Al-Sayer H, Elbishbishi EA, Agarwal R,
Nagar R, Kapoor S, Misra A, Mathur A, Nusrat H, Azizieh F, Khan MA,
Mustafa AS. Elevated levels of IL -8 in dengue hemorrhagic fever. J Med Virol
56:280-285;1998.
24. Rochon YP, Kavanagh TJ, Harlan JM. Analysis of integrin (CD11b/CD18)
movement during neutrophil adhesion and migration on endothelial cells. J
Microsc 197:15-24;2000.
25. Rollins BJ. Chemokines. Blood 90:909-928;1997.
26. Schall TJ, Bacon K, Camp RD, Kaspari JW, Goeddel DV. Human
macrophage inflammatory protein alpha (MIP -1 alpha) and MIP-1 beta
chemokines attract distinct populations of lymphocytes. J Exp Med
177:1821-186;1993.
27. Simon HU. Neutrophil apoptosis pathways and their modifications in
inflammation. Immunol Rev 193:101-110;2003.
4):45-53;1993.
29. Smith JA. Neutrophils, host defense, and inflammation: a double-edged
sword. J Leukoc Biol 56:672-686;1994.
30. Spain-Santana TA, Marglin S, Ennis FA, Rothman AL. MIP -1 alpha and
MIP-1 beta induction by dengue virus. J Med Virol 65:324-330;2001.
31. Tanaka Y, Adams DH, Hubscher S, Hirano H, Siebenlist U, Shaw S. T-cell
adhesion induced by proteoglycan-immobilized cytokine MIP-1 beta. Nature
361:79-82;1993.
32. Taub DD, Conlon K, Lloyd AR, Oppenheim JJ, Kelvin DJ. Preferential
migration of activated CD4+ and CD8+ T cells in response to MIP-1 alpha
and MIP-1 beta. Science 260:355-358;1993.
33. Theofilopoulos AN, Brandt WE, Russell PK, Dixon FT. Replication of
dengue -2 virus in cultured human lymphoblastoid cells and subpopulations
Figure legends:
Fig. 1. Kinetics of IL-8 release by neutrophils in response to DV and LPS.
Neutrophils (2x106) were incubated with DV at MOI of 1, LPS (200 ng/ml) or without
stimulation (C) at 37 ℃. The supernatants were collected at indicated times and
assayed for IL -8 by ELISA. The results presented in this figure represent the mean
+ SD of experiments performed in triplicate on neutrophils from three individuals.
Fig. 2. Chemokine release from neutrophils after DV stimulation. The
concentrations of IL-8, MIP -1α and MIP -1β in the culture supernatants of
neutrophils after incubation with LPS, DV or UV inactivated DV (UV-DV) for 18 h
were assayed by ELISA as described in the Materials and Methods. Cells treated
with medium (C) or C6/36 supernatant (C6/36) were also included. Data from three
experiments are shown as mean + SD. *p< 0.05, compared with the corresponding
values from un-stimulated neutrophils (C) or mock DV stock (C6/36).
Neutrophils (2 x 104 cells/well) were incubated with LPS, DV or without any
stimulation (C) for 18 h. Cells were stained with FITC conjugated anti-CD11b or
anti-TLR4 antibodies. The expression of CD11b and TLR4 on the surface of
neutrophils was observed by confocal fluorescent microscopy as described in the
Materials and Methods section.
Fig. 4. Morphological changes of neutrophils after incubation with DV. (A)
Neutrophils (2x106) were incubated with DV or C6/36 supernatant (C6/36) as
indicated at 37 ℃ for 18 h. Cells were cytospined onto slides, stained and observed
under light microscopy at 100X magnification. (B) Transmission electron
micrograph of DV-stimulated neutrophils. Neutrophils which showed numerous
vacuoles (V) also showed fewer granules (G) in the cytoplasm.
Fig. 5. DV stimulation increased phagocytosis of neutrophils. Neutrophils (2x 106)
were pre-incubated with medium (C) or DV at MOI of 1 or 5 for 1 h followed by
incubation with FITC-beads for 30 min at 37℃. Free FITC-beads were washed
the Materials and Methods section.
Fig. 6. Co-localization of DV and phagocytized particles in neutrophils. Neutrophils
(2x 106) were pre-incubated with medium (C) or DV at MOI of 5 (DV) for 1 h
followed by incubation with FITC -beads for 30 min at 37℃. Neutrophils were
washed, fixed and permeabilized after incubation. Intracellular DV particles were
detected using biotin labeled anti-E antibodies and PE-conjugated streptoavidin,
and then visualized under a confocal microscope as described in the Materials and
Methods section.
Fig. 7. Evaluation of cell viability after DV stimulation. Neutrophils (2x 106) were
inoculated with LPS (200 ng/ml), DV or UV-DV at MOI of 1 for 18 h. Trypan blue
up-take and PI stain were assayed using light microscopy and flow cytometry,
respectively and XTT assay was assessed using a microplate reader. The data
represent the mean of results from two different experiments of trypan blue and PI
stain, while the results of XTT assay were from three different experiments and are
Fig. 8. Effects of DV on the spontaneous apoptosis of neutrophils as determined by
annexin V and TUNEL stain. Neutrophils (2x106) were incubated with DV (MOI=1),
LPS (200 ng/ml) or without stimulation (C) at 37 ℃ for 18h. In addition, some cells
were treated with C6/36 supernatants (C6/36), heat or UV-inactivated DV (H-DV or
UV-DV) as indicated. Cells were stained with annexin V or TUNEL and analyzed by
flow cytometry as described in the Materials and Methods section. The data
represent the results from three different experiments and are shown as mean ±
SD. *p < 0.05, compared with un-stimulated neutrophils.
Fig. 9. LDH release of neutrophils induced by DV. Neutrophils (2x105) were
incubated with DV at different MOI as indicated, LPS (200 ng/ml) or without
stimulation (C) at 37 ℃ for 18 h. The supernatants were collected and assayed for
LDH by ELISA. The results presented in this figure represent the mean + SD of
experiments performed in triplicate on neutrophils from three individuals. *p <
Fig. 1 0 500 1000 1500 2000 2500 3000 0 6 12 18 24 hour IL-8 (pg/ml) C DV LPS
Fig. 2
0
500
1000
1500
2000
2500
3000
3500
4000
C
C6/36
LPS
DV
UV-DV
pg/ml
IL-8
MIP-1α
MIP-1β
*
*
*
*
*
*
*
Fig 3
DV LPS C
DV LPS C
CD11b
Fig 4
Fig. 4
Fig. 6
Fig. 7 0 1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 100 Unstimulated L P S DV UV-DV viability (%) 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 O.D Trypan blue P I XTT
*
Fig. 8 0 10 20 30 40 50 60 C C6/36 D V LPS H-DV UV-DV apoptosis % Annexin V TUNEL
*
*
*
*
Fig. 9 0 5 10 15 20 25 30 35 40 45 DV(MOI=1) DV(MOI=2) C LPS LDH release (%)