行政院國家科學委員會專題研究計畫 成果報告
細菌視紫質之蛋白質工程研究及其於生物光電晶片之應用 (第 3 年)
研究成果報告(完整版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2221-E-011-040-MY3
執 行 期 間 : 97 年 08 月 01 日至 98 年 10 月 30 日 執 行 單 位 : 國立臺灣科技大學化學工程系
計 畫 主 持 人 : 陳秀美
計畫參與人員: 碩士班研究生-兼任助理人員:洪國峰 碩士班研究生-兼任助理人員:溫文興 碩士班研究生-兼任助理人員:謝竺君 碩士班研究生-兼任助理人員:王世育 碩士班研究生-兼任助理人員:周珈伃 碩士班研究生-兼任助理人員:余安棣 碩士班研究生-兼任助理人員:賴珮薇 碩士班研究生-兼任助理人員:胡孔政 碩士班研究生-兼任助理人員:陳逸航 碩士班研究生-兼任助理人員:陳欣禹 大專生-兼任助理人員:鄭凱如
處 理 方 式 : 本計畫涉及專利或其他智慧財產權,2 年後可公開查詢
中 華 民 國 99 年 01 月 28 日
行政院國家科學委員會補助專題研究計畫成果報告 行政院國家科學委員會補助專題研究計畫成果報告 行政院國家科學委員會補助專題研究計畫成果報告 行政院國家科學委員會補助專題研究計畫成果報告
細菌視紫質之蛋白質工程研究及其於生物光電晶片之應用 細菌視紫質之蛋白質工程研究及其於生物光電晶片之應用 細菌視紫質之蛋白質工程研究及其於生物光電晶片之應用 細菌視紫質之蛋白質工程研究及其於生物光電晶片之應用
計畫類別: █ 個別型計畫 □ 整合型計畫 計畫編號:NSC 95-2221-E-011-040-MY3
執行期間: 95 年 8 月 1 日 至 98 年 10 月 31 日
計畫主持人:陳秀美
計畫參與人員:溫文興、謝竺君、王世育、洪國峰、周珈伃、余安棣、
賴珮薇、胡孔政、陳逸航、陳欣禹、鄭凱如
成果報告類型(依經費核定清單規定繳交):□精簡報告 █完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列 管計畫及下列情形者外,得立即公開查詢
█涉及專利或其他智慧財產權,□一年█二年後可公開查詢
執行單位:國立台灣科技大學 化學工程學系
中 華 民 國 99 年 1 月 26 日
中文摘要 中文摘要 中文摘要 中文摘要
Halobacterium salinarum 嗜鹽菌紫色細胞膜(purple membrane,PM)內之細菌視紫 質蛋白(bacteriorhodopsin,BR)為一單方向性光驅動質子泵,具有光致變色、瞬態光 電響應、與非線性光學等特殊性能與功用,被視為發展生物光電元件之最佳奈米生物材 料;然而,至今其仍有無法被完全定向固定化與光學中間體之特性未達極臻等課題尚 待突破。本計畫對於 BR 進行基因重組與蛋白質工程之研究,並對於其突變 PM 膜之特 性、功能性與應用性進行研究,以嘗試藉著基因改質來增進其光學特性與穩定性、並同 時改變其膜表面特殊官能基團的種類以簡化定向固定化,滿足其未來商業化的應用需 求。研究共分三年進行,第一年進行 BR D85N、D96N、D85N/D96N 與 G241C 突變質 體的建構,並將基因轉回入 H. salinarum 菌體中以表現突變 BR;第二年則進行各突變 PM 膜之大量與最適化生產研究,並以蛋白質電泳與光譜分析比較其基本特性;第三年 則嘗試進行各突變 PM 膜於 ITO 玻璃上之定向固定化研究,分析所製備晶片之光電特 性。研究結果顯示,D96N 與 G241C 兩個突變種 BR 的 PM 膜產量和原生種菌株相近,
且均可用果糖梯度純化,並可於其胞外側 Lys 169 殘基處進行 biotin 的鍵結修飾;同時 不論是以雙槽液相光電系統、均一方向性單層薄膜晶片系統或製備簡易的高定向性厚膜 晶片系統進行量測均可得到穩定的高光電流應答訊號。二者中,由於 D96N BR 之 M 態 光學中間體的生命週期已被延長,而使質子無法正常由膜外傳送進入膜內,因此 D96N PM 膜晶片的開燈光電流訊號值比原生種 PM 膜晶片的稍高,關燈的電流訊號值則明顯 縮減;但在加入適當的添加劑後,可使其關燈的光電流訊號值回復正常。G241C PM 膜 間(實際上為 BR 分子間)則會形成雙硫鍵,但加入還原劑後可打斷此雙硫鍵,而使其光 電流訊號值回復正常;然而同時也由於 Cys214 殘基所在的位置被其他胺基酸或脂質遮 蔽,奈米金微粒無法有效鍵結上去,以其他修飾性化合物修飾後也是效果有限,因此原 設計建構 G241C 突變種以方便在 PM 膜的胞內側以化學鍵結方式吸附奈米金微粒而達 到多層 PM 膜塗覆之目的並無法達成,目前我們仍持續進行膜胞內側修飾與固定化的探 討。最後,D85N 與 D85N/D96N 兩個突變種 BR 則由於 Asp85→Asn 突變嚴重破壞 BR 的光驅動質子泵功能與 PM 膜的功能,使菌體無法維持正常生長,甚至無法生產 PM 膜,
因此其產量均極低也幾乎無法產生光電流響應。
關鍵詞關鍵詞關鍵詞
關鍵詞:Halobacterium salinarum、細菌視紫質、紫色細胞膜、突變、光電流響應
Abstract
Bacteriorhodopsin (BR), a unidirectional light-driven proton pump protein in the purple membrane (PM) of Halobacterium salinarum, is considered as the most promising nanobiomaterial for developing biophotoelectronic and holography devices due to its distinctive photochromic, photoelectric, and nonlinear optical properties. For BR applications, the achievement of a high degree of PM fabrication orientation in devices and the improvement of the properties of its photo-intermediates remain critical challenges. Therefore, in this study genetic engineering was carried out in the attempts to modulate BR properties for better applications as well as to facilitate the directional fabrication of PM via affinity immobilization. The project was conducted in three years, with the engineering of D85N, D96N, D85N/D96N, and G241C BR mutants in the first year, the overproductions and electrophoretic and spectroscopic analyses of the mutants in the following year, and the directional affinity-immobilization and photoelectrical response analyses of each mutant in the final year. Among the mutants, D96N and G241C BRs exhibited similar production levels to that of the wild-type one, and were readily purified using sucrose-gradient ultracentrifugation and modified by biotin derivatives on Lys169 at the extracellular side. In addition, stable and high photoelectric responses were readily observed from both irradiated mutants in any of the three photoelectric analysis systems set up in this study. The light-on photoelectric response signal of D96N PM was slightly higher than that of the wild-type one, while its light-off response was significantly impeded due to the prolongation of the life-time of the M photo-intermediate state by mutation. The revival of the reduced signal was observed upon the addition of suitable additives in the electrolyte solution. The G241C PM patches tended to aggregate because of the formataion of intermolecular disulfide linkages, which could be readily reduced to restore the photoelectric responses. However, neither nanogold particles nor biotin derivatives was successfully conjugated on G241C PM due to the hindered accessibility of the Cys241 residue by other residues or lipids, failing the original attempt to facilitate multilayer fabrication of PM chips with this mutant. Finally, even though the D85N and D85N/D96N mutants were successfully engineered, the fact that the mutation on Asp85 had completely broken down the light-driven proton pump function of BR led to poor cell growth and consequently almost demolished PM production. This study created valuable portfolios for future application studies on BR.
Key words: Halobacterium salinarum, bacteriorhodopsin, purple membrane, mutation, photoelectric response
目錄 目錄 目錄 目錄
中文摘要 ··· Ⅱ
英文摘要 ··· Ⅲ
目錄 ··· Ⅳ
第一章 前言 ··· 1
第二章 研究目的 ··· 2
第三章 文獻回顧 ··· 3
3-1 Halobacterium halobium ··· 3
3-2 BR 與 PM 基本性質 ··· 4
3-3 BR 的蛋白質序列與基因序列 ··· 7
3-4 氫離子傳遞路徑 ··· 9
3-5 光循環反應 ··· 11
3-6 PM 的固定化與方向性 ··· 14
3-7 PM 之應用 ··· 16
3-8 BR 表現系統 ··· 19
3-9 BR 的突變 ··· 20
3-9-1 D85N 與 D96N 突變 ··· 21
3-9-2 G241C 突變 ··· 22
3-9-3 其他突變研究 ··· 22
第四章 研究方法 ··· 27
4-1 實驗流程 ··· 27
4-1-1 D85N、D96N、D85N/96N 與 G241C 之突變質體建立 ··· 24
4-1-2 嗜鹽菌的轉形 ··· 24
4-1-3 原生種和突變種 PM 膜的純化、蛋白質電泳與光譜量測 ··· 24
4-1-4 G241C PM 膜與 biotin 衍生物或奈米金微粒之結合 ··· 25
4-1-5 原生種和突變種 PM 膜的光電流量測 ··· 25
4-2 實驗材料 ··· 26
4-2-1 菌株 ··· 26
4-2-2 質體 ··· 26
4-2-3 核酸引子設計 ··· 26
4-2-4 酵素 ··· 28
4-2-5 DNA 操作試液套件 ··· 28
4-2-6 標準分子量溶液 ··· 28
4-3 實驗藥品 ··· 28
4-4 實驗設備 ··· 30
第五章 結果與討論 ··· 35
5-1 各突變基因之建構與篩菌 ··· 35
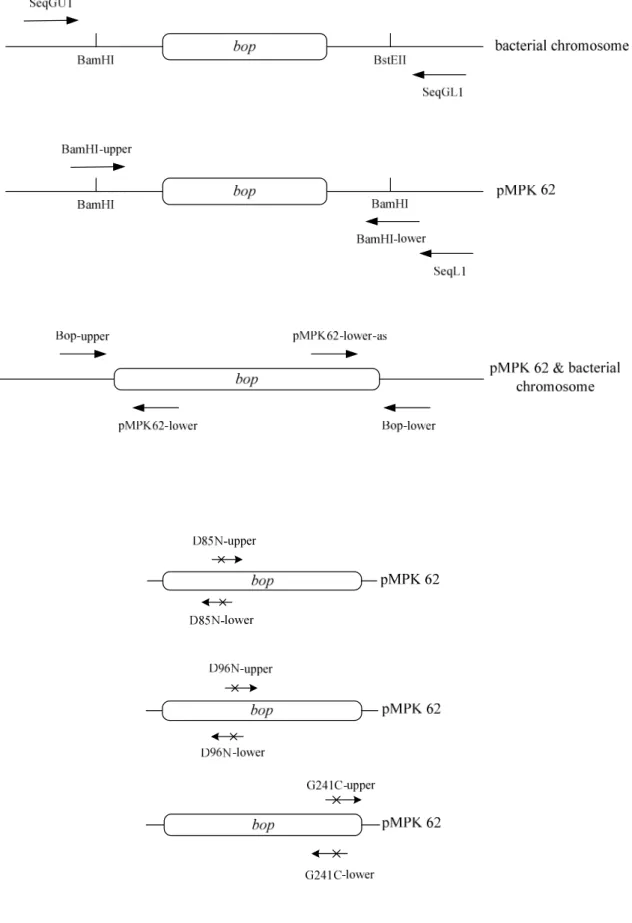
5-1-1 質體 pMPK62 之確認與其轉殖表現 ··· 35
5-1-2 D85N、D96N、D85N/D96N 與 G241C 突變基因質體之建構與篩菌 ···· 36
5-2 嗜鹽菌之轉殖與篩菌 ··· 42
5-3 嗜鹽菌之培養、PM 膜蛋白純化、光譜分析與蛋白質電泳 ··· 44
5-4 同源交換之分析 ··· 44
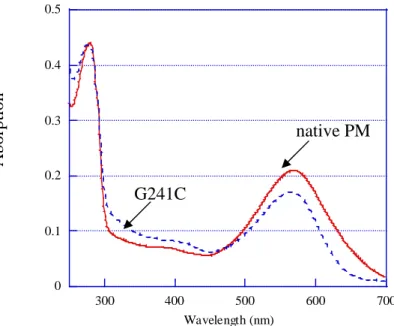
5-5 G241C PM 膜之分析 ··· 57
5-5-1 還原劑對 G241C PM 之影響 ··· 57
5-5-2 以具硫醇基反應性之 biotin 衍生物修飾 G241C PM ··· 57
5-6 各 PM 膜之光電流響應分析 ··· 63
5-6-1 雙槽液相光電系統之光電流響應分析 ··· 63
5-6-2 簡易高方向性厚膜晶片系統之光電流響應分析 ··· 91
5-6-3 均一方向性單層薄膜晶片系統之光電流響應分析 ··· 103
第六章 結論 ··· 119
第七章 參考文獻 ··· 120
第八章 計畫成果自評 ··· 1 2 8 研發成果資料表 ··· 129
第一章 第一章 第一章
第一章 前言 前言 前言 前言
Halobacterium salinarum (原稱Halobacterium halobium) 為一生長在高含鹽份環境中 的嗜鹽菌,其菌體內含有大量的紫色細胞膜 (purple membrane,PM) 而使得菌體外觀顏 色為紫色。Bacteriorhodopsin (BR) 為 PM 膜上唯一光驅動性質子泵的蛋白質,是菌體 處於“低氧”與“照光”環境下所合成的一種深紫色蛋白質。BR 的組成與構造和人類視綱 膜上的visual rhodopsin (VR) 相似,並且僅發現於嗜鹽菌上,因此才得以被人們如此命 名。BR 在吸收光能為能源後,隨即進行一連串的可逆光循環反應機制,將質子由胞內 傳至胞外,產生一膜內外的電位差,此電位差可供菌體細胞膜上的 H+-ATPase 製造合 成 ATP,而使菌體本身可在低氧與低養分環境下生長。
BR 由視黃醛 (retinal) 與視蛋白 (opsin) 以 1:1 之比例所組成,分子量為 26 kDa,
具有 7 個α-helix 與 2 個 β-sheet 的構造,以連續貫穿之方式與三聚體形式二維六角形晶 格排列在 PM 膜上。每個視蛋白由 248 個胺基酸所組成;而視黃醛的碳醯基與位於 Lys216 的ε-amino 基團形一 Schiff base 鍵結。每一分子的 BR 由十個脂質(lipid)所包圍著,而這 些提供了 BR 與 PM 結構的穩定。另外,PM 膜本身的浮力密度 (buoyant density) 為 1.18 g/mL,因此可利用果糖梯進行純化分離。BR 蛋白質具有許多優異性質,包含尺寸極小、
可產生光電訊號、具有獨特的光循環反應與多種光中間體、具有非線性光學特性、可承 受高範圍的溫度與酸鹼值、取材容易等特點,因此可被運用於光儲存體、太陽能電池、
濾片、防偽墨水與感測器等製造,為一極具高附加價值與前瞻性的“綠色”生物性奈米級 材料。
近年來,許多研究學都致力於 BR 的立體結構、質子傳遞路徑、基因重組、突變修 飾、視黃醛衍生物取代的改質與固定化及其性質等研究,其目的主要是增進人們對於 BR 性質的了解與其實用性。在本研究中,我們選定 BR 中的 D85、D96 與 G241 三個 殘基,以定點突變法將原有之胺基酸置換成其他的胺基酸。由於 Asp85 與 Asp96 兩殘基 在 BR 的質子傳遞上分別扮演著質子接受者與提供者的重要角色,具有影響 BR 的吸收 光譜與質子傳遞效率、以及決定 M 中間態的生成與消失速率之地位,因此其 D85N 與 D96N 二突變種可被利用作為日後進一步探討質子傳遞與光循環反應的基礎。其中 D96N 突變預期會造成 BR 光循環 M 中間態的消失延遲,將來可被應用作為製備即時全息記憶 體 (real-time holographic memory) 之材料;D85N 突變則預期會造成光循環 M 中間態的 生成延緩,影響其光驅動質子泵功能。此外,由於 Gly241 殘基位於 PM 膜之 BR 蛋白的 C 末端上,推測對於 BR 的吸收光譜與質子傳遞的特性上並非是重要的胺基酸,因此研 究中進行 Gly241→Cys 的突變,期望利用突變後的 Cys 殘基將 PM 膜以胞內側定向方式 固定在金載體上或與奈米金微粒結合。研究中首先利用基因重組與 SOEing 定點突變技 術建立帶有 D85N、D96N、D85N/D96N 與 G241C 等突變點之質體,再經過一連串的篩 選、膠體純化、定序確認與大量質體抽取等一般實驗步驟後,最後將質體轉殖入嗜鹽菌 的專用宿主菌株中以表現突變膜蛋白,並且將這些具突變膜蛋白之菌株進行大量培養與 突變 PM 膜之純化,並進行特性之研究,並與野生種菌株之 PM 膜相互比較。藉由此 BR 的突變研究不僅可以了解其各胺基酸在 BR 分子之特性與結構上所扮演的角色,並 可藉由基因改質提升其實用性與應用性。
第 第 第
第二 二 二 二章 章 章 章 研究目的 研究目的 研究目的 研究目的
H. salinarum 嗜鹽菌 PM 膜內之 BR 蛋白(bacteriorhodopsin,BR)為一單方向性光 驅動質子泵,具有光致變色、瞬態光電響應、與非線性光學等特殊性能與功用,被視為 發展生物光電元件之最佳奈米生物材料;然而,至今其仍有無法被完全定向固定化與光 學中間體之特性未達極臻等課題尚待突破。因此本計畫對於 BR 進行基因重組與蛋白 質工程之研究,並對於其突變 PM 膜之特性、功能性與應用性進行研究,以嘗試藉著基 因改質來增進其光學特性與穩定性、並同時改變其膜表面特殊官能基團的種類以簡化定 向固定化,滿足其未來商業化的應用需求。本研究具有三個階段性目的,首先建構 BR D85N、D96N、D85N/D96N 與 G241C 共四種突變的 BR 表現質體,並將基因轉回入 H.
salinarum 菌體中以表現突變 BR;其次探討各突變 PM 膜之大量與最適化生產,並以蛋 白質電泳與光譜分析比較其基本特性;最後則嘗試進行各突變 PM 膜於 ITO 玻璃上之定 向固定化研究,並分析所製備晶片之光電特性。此突變研究不僅有助於了解 BR 內主要 胺基酸在其特性與結構上所扮演的角色,釐清 BR 光電流產生之機制,並可藉由基因改 質提升其實用性與應用性。
第 第 第
第三 三 三 三章 章 章 章 文獻回顧 文獻回顧 文獻回顧 文獻回顧
3-1 Halobacterium salinarum
Halobacterium salinarum 為一極端嗜鹽、似 Gram-negative 與兼氧性 (facultative aerobes) 的古細菌,主要生長在富含鹽分 (NaCl:3 ~ 5.2 M) 的環境中。外觀主要顏色 為紫色,為一大小為直徑 0.5 µm、長度 3-10 µm、具鞭毛組織的桿狀菌體。
H. salinarum 之菌體細胞壁以醣蛋白 (glycoprotein) 構成,其本身含有較多的酸性胺 基酸 (acidic amino acids,negative-charged),如 aspartate 與 glutamate。這些酸性殘基提 供了 carboxyl groups 與溶液中的陽離子 (Na+、Mg+2、Ca+2等) 相互吸引作用,在細胞膜 上形成堅固的單一保護層並維持菌體的完整性,稱之為 surface-layer 或 S-layer。當溶液 中的陽離子含量過低太少時,這些 glycoprotein 則產生相互排斥之作用而失去原有結 構,進一步造成溶菌現象 (cell lysis)。菌體細胞膜則主要由 phospholipids、phospholipids 的衍生物與 glycolipids 等組成,特別的是其非極性碳鏈與 glycerol 以 ether 形式鍵結,有 別於 Eubacteria 與 Eukarya 的 ester 形式鍵結。
生長在高濃度鹽分的嗜鹽菌,首先要面對並克服由高濃度鹽分環境所帶來的滲透壓 (osmotic stress) 效應。一般細胞若處在高鹽分溶液 (高張溶液) 中,由於胞內滲透壓遠 小於胞外,造成自身水分的流失並且進一步的萎縮死亡。但對於嗜鹽菌而言,除了上述 文章中菌體的 S-layer 可以提供保護與幫助維持菌體的完整性之外,最主要依賴:(1)運 用 存 在 細 胞 膜 上 的 離 子 泵 浦 來 維 持 胞 內 外 環 境 的 離 子 平 衡 ; (2) 菌 體 自 行 生 產
“compatible solutes”,例如:glycerol、glycine betaine、ectoine 等 (Madigan and Oren, 1999;
Oren, 1999),以維持菌體內的滲透壓。特別的是存在 H. salinarum 菌體內最多的陽離子 是鉀離子 (Matheson et al., 1976;Brock et al., 1994b),而不是鈉離子,並且菌體內的部 分蛋白質與酵素必須依靠鉀離子的幫助,才能維持其活性。
H. salinarum 菌體細胞膜內主要含有兩種色膜:(1)紫色細胞膜 (purple membrane,
PM);(2)紅色細胞膜 (red membrane,RM),分別散佈在細胞膜上並使得菌體外觀帶色。
在 PM 膜上僅含單一光驅動性 (light-driven) 蛋白質-bacteriorhodopsin (BR);而在 RM 膜 上則含有 halorhodopsin (HR)、senory rhodopsin I (SR-I)、senory rhodopsin II (SR-II 或 photorhodopsin,PR) 三種光驅動性蛋白質與其他膜蛋白等。此四種光驅動性蛋白質均 為跨膜蛋白,分子量相近均為約 26 kDa,但各具有不同的光反應功能 (Schäfer et al., 1999),如 bR 為一光驅動性氫離子泵浦 (light-driven proton pump),可將氫離子由胞內傳 送至胞外;HR 為光驅動性氯離子泵浦 (light-driven chloride pump),可將氯離子由胞外 傳送至胞內,維持胞內外氯離子的平衡;SR-I 與 SR-II 本身亦具有氫離子傳遞的功能,
分 別 與 Htr-I 、 Htr-II (homodimeric transducer) 結 合 後 , 可 掌 管 菌 體 的 趨 光 移 動 性 (phototaxis),如 Fig. 3-1 所示,使菌體能夠利用鞭毛移動。
BR 為 PM 膜上之單一膜蛋白,在來源取得與純化方面較其他三種 rhodopsin 蛋白容 易,再加上因為最早被發現、為植物光合作用系統以外的另一組光合成系統、最早被完 成胺基酸與基因序列定序之膜蛋白、具有感光性與氫離子泵浦功能,並且 BR 亦為 G 蛋 白結受器的一種,相似於哺乳類動物負責視覺傳導之蛋白質,因此深受學者專家們的喜 愛與研究。
Fig. 3-1 Four light-driven rhodopsins in H. salinarum (Schäfer et al., 1999).
目前此菌體的培養與 PM 膜的純化大多採用由 Oesterhelt 與 Stoechkenius (1974) 所 提供的方法。其培養基所含的鹽類濃度高達 27 % (w/v,total salts),主要包含 NaCl 與少 量的 MgCl2、KCl 與檸檬酸三鈉鹽,而碳、氮源則由 peptone 所提供。此菌體成長的溫 度最高可達 57 ℃,但在如此高溫環境下,會發生培養基水分蒸發 (塩析) 與溶氧量不足 的問題,因此在液態培養時,一般採用溫度為 37℃、轉速為 200 rpm、培養液與容器體 積比為 1:3 作為培養條件。
照光與通氧量的條件為影響菌體大量合成生產含 BR 之 PM 膜的主要因素 (Shand and Betlach, 1991)。Bacterio-opsin (BO) 為 BR 的前驅物,不含具有 retinal 小分子,其表 現主要受到 Bacterio-opsin activator protein (Bat) 與 Bacterio-opsin-related protein (Brp) 兩 種蛋白質直接與間接的調控,如 Fig. 3-2 所示。在照光環境中,光線可以促進 Brp 的表 現,而 Brp 不但可促進 Bat 的產生亦可幫助 retinal 的合成 (Peck et al., 2001);在低氧環 境下,可以誘發 Bat 的表現與生產,進而促使菌體大量產生 BO。因此在培養菌體時,
大多同時採用照光與降低氧氣供應 (降低搖瓶轉速至 100 ~ 120 rpm 或封口斷氧)。但是 嚴苛地限制氧氣的供應,則會影響 retinal 的合成 (Hartmann et al., 1980)。
Fig. 3-2 (1) Map of the bop gene cluster. (2) Proposed model for regulation of the bop gene cluster (Shand and Betlach, 1991).
3-2 BR 與 與 與 與 PM 基本性質 基本性質 基本性質 基本性質
BR (又名細菌視紅紫質) 為一光驅動性氫離子泵浦的維他命 A 醛復合膜蛋白,於 1971 年由 Oesterhelt 與 Stoeckenius (1971) 發現於嗜鹽菌的 PM 紫膜 (Blaurock and Stoeckenius, 1971) 上。BR 是菌體 H. salinarum 處於“低氧”與“照光”環境下所合成的一種 深色紫色蛋白質,使菌體外觀呈現紫色。在吸收光子後,BR 將光能轉換成電化學能,
迫使氫離子由胞內傳遞至胞外而產生一電位差,此電位差可供細胞膜上的 H+-ATPase 製
造合成 ATP,使菌體在低氧環境下維持生存等多種用途。由於 BR 的組成方式和結構與 人類視綱膜上的 visual rhodopsin (VR) 相似,並且僅發現於嗜鹽菌上,因此被稱為 bacteriorhodopsin。
BR 分成兩部份:retinal (視黃醛,醛基型式的維他命 A) 與 opsin (視蛋白,亦稱為 bacterio-opsin,BO)。其視蛋白部份由 248 個胺基酸所構成,分子量為 26 kDa (Oesterhelt and Stoeckenius, 1971;Bridgen and Walker, 1976),具有 7 個α-helix 與兩個 β-sheet 構造,
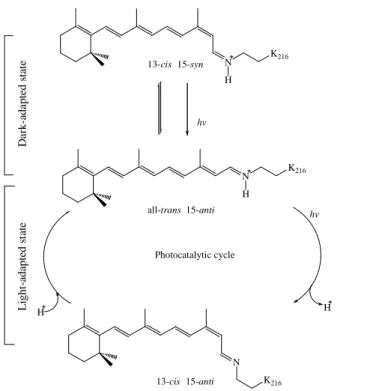
如 Fig. 3-3 所示。蛋白質的 N 端與 C 端分別在菌體細胞膜的兩側;N 端位於胞外;而 C 端則位於胞內 (Gerber et al., 1977)。在解讀上分別以 A 至 G 的 7 個字母位序命名區別 7 個α-helix,彼此以連續貫串的方式,完整地鑲嵌在 PM 膜內,如 Fig. 3-4 所示,其中 helix-B 與 helix-C 之間的 loop 則形成一反平行 (anti-parallel) 的β-sheet。視黃醛為 BR 的發色基 團 (chromophore)。視黃醛的碳醯基與位於 helix-G Lys 216 (Bayley et al., 1981) 之 ε-amino 基團形成一 Schiff base (SB) 鍵結,如 Fig. 3-5 所示,此時視黃醛的吸收值會由 380 nm 增為 568 nm,此象現稱之為 “opsin shift”。並且將 opsin 分成上下兩區域:釋放 氫 離 子 至 胞 外 的 胞 外 通 道 (extracellular pathway, EC) 與 攝 取 氫 離 子 的 胞 內 通 道 (cytoplasmic pathway,CP)。Retinal 受到光線刺激後,在第 13 個與第 14 個碳中間的雙 鍵會由全反式 (all-trans) 轉變為順式 (13-cis 15-trans),並且進行一連串的光循環反應。
當 BR 處在無光環境 (dark-adapted,DA) 中,其 retinal 以 all-trans (BR568或 B 態) 與 13-cis 15-syn (BR548或 D 態) 兩種同分異構物形態以比例大約為 1:2 (Scherrer et al., 1989) 同時存在,此時 BR 沒有進行氫離子的傳遞;反之,BR 置於照光環境 (light-adapted,
LA) 之下,其最大吸收值則增為 568 nm,在 retinal 會收吸光子後,所有的 13-cis 15-syn 瞬間產生異構化形成 all-trans,隨即進入光循環反應與產生氫離子傳遞,如 Fig. 3-6 所 示。
Fig. 3-3 (a) 3-D structure of bacteriorhodpisn. (b) Extracellular side. (c) Cytoplasmic side (Subramaniam and Henderson, 2000;PDB:1FBB).
A B C D E F G
Cytoplasmic side
Extr acellu lar si de
Fig. 3-4 Model of two-dimensional structure of bacteriorhodpisn (Oesterhelt et al., 2000).
(a) (b)
(c)
O
+
Retinal
H2O
N H H2N
K216
K216
protonated Schiff base
Fig. 3-5 Linking of a free retinal with Lys216 through the formation of a protonated Schiff base.
PM 主要由 25 % (w/w) 的膜質與 75 % (w/w) 的 BR 所組成,其脂質雙層厚度約 4.5-5
nm,直徑大小約 1 µm 的 PM 中內含 105的 BR 分子。PM 是由許多個二維六角形結晶晶
格排列構成,每一個六角形結晶晶格含有 3 個 BR 分子 (trimer),如 Fig. 3-7 所示。每 一個 bR 分子大約有十個脂質所包圍環繞,這些脂質可分成兩大類:(1)極性脂質 (polar lipids);(2)中性脂質 (neutral lipids)。在極性脂質方面,包含了 phospholipids (PG:
phosphatidylglycerol、PGS:phosphatidylglycerol sulfate、PGP-Me:phosphatidylglycerol phosphate methyl ester、BPG:bisphosphatidylglycerol) 與 sulfoglycolipids (GlyC:archaeal glycocardiolipin、S-TGD-1:3-sulfate-Galpβ1-6Manpα1-
2Glc pα-1-archaeol);在中性脂質方面,包含了 squalene 與 vitamin MK8兩種 (Corcelli et al., 2000;Krebs and Isenbarger, 2000;Corcelli et al., 2002)。這些脂質提供了 BR 與 PM 結構的穩定,並且使得 PM 具有下列特殊性質:(1)浮力密度 (buoyant density) = 1.18 g/mL,因此可利用果糖梯度分離與純化 (Oesterhelt and Stoechkenius, 1974);(2)可承受 高溫與高範圍酸鹼值的環境。
N K216
H hv
N K216
H 13-cis 15-syn
N 13-cis 15-anti all-trans 15-anti
K216 Photocatalytic cycle
H H
Dark-adapted stateLight-adapted state hv
Fig. 3-6 The isomers of retinal chromophore in dark- adapted and light-adapted bacteriorhosopin (Patzelt et al., 2002).
另外,由 BO 所構成的膜塊稱之為 apomembrane,可分成 whtie membrane (WM,
白膜) 與 brown membrane (BM,灰膜) 兩種。白膜可從突變菌株 (colorless strain、
retinal-deficient strain) 上分離獲得,其 buoyant density 為 1.17 g / mL,分佈在白膜上的 BO 以三聚體亦可形成六角晶格,但是六角晶格的“order”較為鬆散、不緊密 (Usukura et al., 1981;Mukohata et al., 1981);灰膜則是在液態培養基中添加 2 mM 的 nicotine (尼古 丁) 進行培養 (尼古丁可抑制菌體 retinal 之合成與影響 trimer 的形成),其 buoyant density 為 1.14 g / mL 並且 BO 以“分散”的方式分佈在灰膜上 (Sumper et al., 1976;Hiraki et al., 1978;Hwang et al., 1981)。
Fig. 3-7 Hexagonal trimer packing of BR molecules in pruple membrane (Subramaniam et al., 2002)
3-3 BR 的蛋白質序列與基因序列 的蛋白質序列與基因序列 的蛋白質序列與基因序列 的蛋白質序列與基因序列
自從 BR 的許多“特殊”性質被人們發現後,例如:具有 retinal 的復合蛋白、光驅動 性氫離子泵浦 (Oesterhelt and Stoechkenius, 1973)、可產生光電流訊號 (Drachev et al., 1974) 與光循環反應等,部分研究學者為了進一步了解這些“特殊”性質的來龍去脈,進 行了最主要的基本關鍵研究,即揭開 BR 的胺基酸與基因序列。因此,在本節文章中將 主要分別敘述 BR 的胺基酸序列與基因序列。
BR 主要由 248 個胺基酸所組成,結構如前文所敘述可形成 7 個α-helix 與 2 個 β-sheet 構造,其中較具“完整性”的胺基酸序列 (即蛋白質序列) 分別由 Ovchinnikov 等人 (1979) 與 Khorana 等人 (1979) 於 1979 年先後提出發表。事實上,BR 的胺基酸序列被正式提 出發表之前,其部份序列已經直接或間接地被發表 (Bridgen and Walker, 1976;Walker et al., 1979;Gerber et al., 1979)。
根據 Ovchinnikov 等人 (1979) 所整理的研究可知,其主要是先利用化學試劑 (cyanogen bromide,CNBr) 與蛋白水解酵素 (papin、typsin、chymotypsin) 將 BR 分成 多個大小不同的片段,再利用膠體分離純化這些片段,以方便定序工作。而主要的結果 與討論為:(1)BR 具有 247 個胺基酸;(2)部分胺基酸具有穩定結構的完整性與幫助氫離 子傳遞的功能。
Khorana 等人 (1979) 亦採用了化學試劑與蛋白質水解酵素將 BR 分成多個大小不 同的片段,但是在分離純化與定序方面,Khorana 等人 (1979) 則分別使用了兩種方法。
在分離純化蛋白質片段方面,採用了膠體分離與 HPLC 兩種方法,增加蛋白質片段的質 與量;在定序工作方面,則同時以 Edman degradation 與氣相質譜儀 (GC/MS) 兩種方式 相互比較,以增進定序的準確性與可信度。因此,所獲得的 BR 胺基酸序列共有 248 個,
如 Fig. 3-8 所示,包含了 71.8 % 疏水性與 28.2 % 親水性胺基酸,但是,兩組學者所發 表的胺基酸序列間仍有差異,包含了胺基酸總共目數與部分胺基酸 (Leu、Ile、Ser、Ala、
Glu、Gln、 Asn、Asp)的差異,整理如 Table 3-1 所示。造成此差異的原因可能是:(1) 蛋白質片段的純度與定序準確度;(2)菌株的差異性。
Fig. 3-8 The complete amino acid sequence of bacteriorhodopsin (Khorana et al., 1979).
Table 3-1. The difference between Ovchinnikov's and Khorana's protein sequencing results on BR.
Ovchinnikov et al. (1979) Khorana et al. (1979)
Glu 105 Gln 105
Ile 111 Leu 111
Asp 115 Asx 115
Leu 117 Ile 117
Ser 146 Leu 146
Glu 161 Glx 161
Ala 206 Leu 206
Total residues 247 248 (Try-138) Asx: Asp or Asn; Glx: Glu or Gln
隨後,Dunn 等人 (1981) 利用 cDNA (Chang et al., 1981) 為探針 (probe) 與含有 H.
salinarium 染色體片段的 E. coli 進行雜交 (colony filter hybirdation),首度發表了 BR 的 基因列序。其先針對 BR 的第 9~12 個胺基酸設計合適的核酸引子 (primer),與 mRNA 雜交進行反轉錄並產生單股 cDNA (共 74 碼),經過一連串的步驟可獲得雙股 cDNA,再 與質體進行重組,利用 E. coli 大量生產。另外,將 H. salinarium 染色體先以 Pst I 限制 酶酵素處理,與質體進行重組並利用 E. coli 生產,進行雜交工作。最後獲得 1.6 kb 含有 BR 基因 (簡稱 bop) 的片段,如 Fig. 3-9 與 Fig. 3-10 所示。
Fig. 3-9 The 1.6 kb fragment containing bacteriorhodpsin gene (Dunn et al., 1981).
bop 基因總共包含 786 核苷酸,可形成 262 個胺基酸。其中 262 胺基酸可分三部分 為:(1)13 個前導胺基酸 (precursor sequence),位於 BR 的 N 端之前;(2)248 個胺基酸,
可形成 bR;(3)1 個天門冬胺酸 (Asp),位於 BR 的 C 端之後。特別的是這 14 個“額外”
的胺基酸卻不與 bR 同時存在於紫膜中,其功能可能是在 BR 形成之初,幫助 BR 嵌入
脂質中。另外,整個 248 個胺基酸不包含任何 His 與 Cys 胺基酸。與 Ovchinnikov 等人 (1979)、Khorana 等人 (1979) 所發表的胺基酸序列比較,Dunn 等人 (1981) 所得之結果 在胺基酸的目數與序列上與 Khorana 等人 (1979) 所發表的結果較符合一致 (除了 Asp115與 Glu161之外)。
BR 胺基酸列序與基因列序的解開,有助於在 BR 晶體結構與氫離子傳遞方面的探 討,特別是基因列序為一重要的里程碑,除了決解蛋白質列序上的疑慮之外,更開啟日 後運用基因工程技術來進一地研究探討 bR 的吸收光譜、反應中間物、蛋白質與脂質的 相互作用、結構、光電訊號、離子傳遞與應用。
Fig. 3-10 The amino acid sequence and DNA sequence of the bacteriorhodposin (Dunn et al., 1981).
3-4 氫離子傳遞路徑 氫離子傳遞路徑 氫離子傳遞路徑 氫離子傳遞路徑
當 BR 受 到 光 刺 激 ( 即 吸 收 光 子 ) 後 , retinal 隨 即 產 生 光 異 構 化 (photoisomerization),進行一連串的光循環反應,經過 J、K、L、M、N、O 等多種不同 結構形態的變化,並伴隨著氫離子傳遞。BR 的氫離子傳遞可藉由水分子與殘基所共同 形成的氫離子網路,從胞內 (CP 端) 推送至胞外 (EC 端),並且產生一氫離子梯度 (電 位差),可藉助精密儀器來量測電流值或電壓差之大小。在此章節中,先敘述部分胺基 酸在氫離子傳遞上所扮演的角色後,再以 Luecke 等人 (1998;1999b) 所發表的研究結 果進一步說明路徑。
Asp85與 Asp96位於 Schiff base (SB) 兩側 (即 EC 端與 CP 端),分別扮演著氫離子的受 體 (acceptor) 與供體 (donor)。當 retinal 吸收光子後,retinal 與 opsin 會發生“結構改變”,
即 retinal 由全反式 (all-trans) 轉變為順式 (13-cis 15-trans);opsin 則是其 helix 會彎曲變 化以利氫離子傳遞。SB 會首先釋放一氫離子 (deprotonated),此氫離子由 Asp85接收,
最後再由 Asp96獲得一氫離子 (reprotonated),如 Fig. 3-11 所示 (Werner, 2000)。在 SB 的去質子化與再質子化之過程,helix C、 helix F 與 helix G 會產生結構改變。在 BR 分 子的表面上存在著帶電性的胺基酸,這些胺基酸可幫助氫離子的攝取與釋放。在 CP 端 包含了 Asp36、Asp38、Asp102、Asp104、Glu161,這些胺基酸就像觸手一般,幫助 BR 從 水溶液中吸收獲得氫離子 (Checover et al., 1997;Brown et al., 1999),其中 Asp38則是進 入 CP 端之氫離子通道上的第一個入口 (Riesle et al., 1996);而在 EC 端則包含了 Glu9、 Glu74、Glu194、Glu204,這些胺基酸可幫助氫離子釋放至水,其中 Glu204為離開 EC 端之 氫離子通道上的最後一個胺基酸並且推測即為氫離子釋放基 (proton release site,PRS,
即為 XH) (Brown et al., 1995)。
Kimura 等人 (1997) 特別地利用電子顯微鏡描繪存在於 BR 表面上的帶電性胺基 酸,並且根據已知的文獻結果僅提出氫離子傳遞之可能路徑,如 Fig. 3-12(a) 所示,但 對於氫離子傳遞步驟並沒有詳細地說明。目前廣為人們所採用的氫離子傳遞模型係由 Luecke 等人 (1998;1999b) 所發表,如 Fig. 3-12(b)所示。在 pH 值介於 5 至 9 之間的氫 離子傳遞步驟可分為五個重要程序,其依次序為:(1)SB 發生去質子現象而失去一氫離 子,且由 Asp85則接收此氫離子;(2)在 Asp85產生再質子化現象後,另一個“獨立”的氫 離子從氫離子釋放基 (XH,推測為 Glu204) 釋放至水溶液中;(3) Asp96發生去質子現象 而失去一氫離子,並且此氫離子由 SB 接收;(4)Asp96從 CP 端表面獲得一氫離子,即發 生再質子化;(5)Asp85從 SB 所獲得之氫離子釋放至氫離子釋放基上。其反應機構如下 所示 (Balashov, 2000):
O H H Asp SB c hv Asp
O H SBH
t / 2 / 85 ] 85 2
[ − + − + → − + + (1)
+
− +
→ X Hrelease
XH (2)
+
− + −
→
−
+c SB Asp c SBH H
Asp96 96 (3) H
Asp H
Asp96− + + → 96 (4a)
+
+ → −
−SBH t SBH
c (4b) XH
Asp O H SBH t
X H Asp O H SBH
t− + + 2 + 85 + − →[ − +/ 2 / 85−]+ (5)
Fig. 3-11 Molecular mechanism of proton (H+) pumping in bacteriorhodopsin. (Werner, 2000).
Fig. 3-12 (a) Possible proton pathway in bacteriorhdopsin (Kimura et al., 1997). (b) Overall view of bacteriorhodopsin. Shown with the retinal (purple) and residues directly implicated in proton transport. (Luecke et al., 1999b).
其中, t 表示為 all-trans 15-anti,c表示為 13-cis 15-anti。值得特別注意的是發生在 pH 值介於 2 至 5 之間的氫離子傳遞亦可分為五步驟,與 pH 值介於 5 至 9 之間的氫離子 傳遞步驟相似;但其氫離子吸收與釋放的次序以及速率有所變化,亦即包含:(1)SB 發 生去質子現象而失去一氫離子,且由 Asp85則接收此氫離子;(2)Asp96 發生去質子現象 而失去一氫離子,並且此氫離子由 SB 接收;(3)Asp96從 CP 端表面獲得一氫離子,即發 生再質子化;(4)在 Asp85產生再質子化現象後,另一個“獨立”的氫離子從氫離子釋放基 (XH,推測為 Glu204) 釋放至水溶液中;(5)Asp85從 SB 所獲得之氫離子釋放至氫離子釋 放基上 (Fig. 3-13)。在此 pH 值範圍 (2 < pH < 5) 內氫離子是先吸收而後釋放,並且分 別發生在 N→O 與 O→BR568期間,與前述 5 < pH < 9 之範圍時氫離子先被釋收而後吸 收,並且分別發生在 L→M 與 N→O 期間之機制有所不同。而於氫離子釋放至水溶液速 率的差異上,低 pH 值範圍 (2 < pH < 5) 時花費了 3 ms;而高 pH 值範圍(5 < pH < 9) 時 僅耗費 80 µs。
Fig. 3-13 Scheme showing steps in light-driven proton transport by bacteriorhodopsin from the inside the cell to the outside. (Balashov, 2000).
3-5 光循環反應 光循環反應 光循環反應 光循環反應
除了具有氫離子傳遞的功能之外,BR 的光循環反應亦是最為人們所重視與研究探 討的特殊性質。BR 在吸收光子能後,進行了一連串的化學性與物理性的改變,包含了 retinal 的 all-trans 15-anti 與 13-cis 15-anti 二形態間的轉換、氫離子傳遞與蛋白質本體 (BO) 之結構變化,即所謂光學中間體 (photointermediate) 的產生,並且這些中間體均 具有獨特的吸收光譜與生命週期 (lifetime),每一步驟均為快速、具可逆性的光化學反 應。可影響光循環反應的參數包含了壓力、黏度、溫度與溶液成分等 (Vsevolodov,
1998a),這些參數因子會影響各個中間體的生命週期與吸收光譜,而導致了目前各國學 者所提的循環模型之吸收光譜間上之約略差異。
BR 的光學中間體主要包括了 J、K、L、M、N、O等 6 種型態與兩個特殊光學中 間體:P與Q (Váró & Lanyi, 1991;Lanyi, 1997;Haupts et al., 1999),而反應的最終結 果是 BR 分子恢復到基態並完成氫離子傳送,如 Fig. 3-14 與 Table 3-3 所示。在 DA 狀 態下,BR 分子的 retinal 以 13-cis 15-syn 與 all-trans 15-anti 之形態並以 1:2 之比例同時 存在 (如前文 Fig. 3-6 所敘)。在吸收光子後,絕大部份的 13-cis 15-syn 可轉換為 all-trans 15-anti,隨後立即進入了光循環反應並產生氫子傳遞。在 J-state 下,retinal 開始由 all-trans 15-anti 異構化為 13-cis 15-anti,其反應時程極為短暫,約為 500 fs。在 K-state 下,除了 retinal 的結構更加地扭曲以利 L-state 的形成之外,間接地影響活性中心的水分子 W402。在 L-state 下,存在於 CP 端與 EC 端通道上的水分子受到擾動以利氫離子傳遞。
在 L → M 期間,SB 會釋放一氫離子給 Asp85。同時間地,一氫離子從 PRS 上,釋放至 水溶液中。在 M-state (M1→M2)下,BR 的最大吸收光譜仍然為 410 nm,但 BR 分子卻 發生結構上的改變。在 M → N 期間,SB 從 Asp96獲得氫離子,Asp96本身則失去一氫離 子。在 N → O 期間,Asp96從 CP 端表面獲得一氫離子,並且 retinal 由 13-cis 15-anti 恢 復成 all-trans。最後,在 O → BR 期間,Asp85將已獲得之氫離子釋放至 PRS 上,此時 BR 分子恢復至基態 BR568。
A sp 8 5H
Fig. 3-14 Photochemical and thermal conversions of bacteriorhodopsin (Hampp and Silber, 1996)
Table 3-3 Characterization of the intermediates of the BR photocycle (Hampp, 2000a).
name λmax (nm)
ε (M cm-1)
retinal configuration
SB conformation
SB
protonation Tfree (℃)
B 570 63000 all-trans 15-anti yes ---
K 586 52100 13-cis 15-anti yes -190
L 544 48900 13-cis 15-anti yes -100
MI 409 48800 13-cis 15-anti no -30
MII 409 48800 13-cis 15-anti no -10
N 562 46100 13-cis 15-anti yes -60
O 629 61900 all-trans 15-anti yes
D 550 51000 13-cis 15-syn
P 485 39000 9-cis
Q 390 9-cis
另外,BR 分子所處外在環境之 pH 值大小,會影響“氫離子通道”上重要殘基的質子 化與去質子化狀態,進而改變氫離子傳遞的順序與光循環反應路徑。pH 值的影響可區
分成四個分段:(1) pH < 2;(2) 2 < pH < 6;(3) 6 < pH < 9.5;(4) pH > 9.5 來討論,如 Fig.
3-15 所示 (Balashov, 2000)。由於 SB、Asp85、XH 與 Asp96的 pKa值分別為 12.4、2.6、
5.8 與 11,所以在 pH < 2 的極酸環境下,Asp85 被質子化而由 Asp85‐形成 Asp85H,SB 無法產生去質子現象 (即無法將氫離子傳遞給 Asp85),導致了 BR 分子無法形成 M-state,此時 PM 膜會形成 blue membrane (BM) 膜,其最大吸收波長為 603 nm;同時 XH 被抑制而使得 BR 分子不具有氫離子傳遞現象 (即不產生 light-on 與 light-off 的電 流訊號)。在 2 < pH < 6 下,雖然 BR 有完整的光循環反應路徑,但 XH 的氫離子釋放 (解 離) 受到外在環境 pH 值之影響進而被延遲至 O→BR568期間,即氫離子無在 L→M 期間 從 XH 上釋放至水溶液中,並且氫離子傳遞的順序是先吸收而後釋放,導致了所獲得電 流訊號是先發生 light-off 後,再出現 light-on,如 Fig. 3-16a 所示 (Roberston and Lukashev, 1995)。在 6 < pH < 9.5 下,則為前文中所說明,氫離子傳遞的順序是先釋放而後吸收,
導致了所獲得電流訊號是先發生 light-on 後,再出現 light-off,如 Fig. 3-16b 所示。在 pH
> 9.5,XH 呈現去質子化狀態 (即為 X‐),BR 分子的吸收光譜呈現紅位移 (red-shift) 現 象,且其 M-state 的生成快速;而在氫離子傳遞方面,由 Arg82取代原本 XH 釋放氫離 子的功能,並且所獲得的電流訊號會隨著 pH 值的上升而下降;但是若 pH 值大於 11 則 應不產生 light-on 與 light-off 的電流訊號 (Kono et al., 1993;Balashov, 2000)。
Fig. 3-15 Scheme of the photochemical conversion of BR at the different pHs (Balashov, 2000).
Fig. 3-16 Photocurrent versus time for a randomly oriented film of wild-type BR at pH = 3 and pH = 7 (Roberston and Lukashev, 1995).
P-state 與 Q-state 為 BR 的 側光循環反應 (branched-photocycle) 中的光學中間體,
最早係由 Popp 等人 (1993) 所發現的,可藉由紅光誘發生產並以藍光消去。P-state 與 Q-state 的最大吸收波長分別為 490 nm 與 380 nm,將 BR 先以綠光激發並等待 O-state 形成後,再以紅光進行誘發 P-state 的產生,並且 retinal 由 all-trans 轉變成 9-cis 15-anti 如 Fig. 3-17 所示。P-state 本身的生命週期較短暫、不穩定;而 Q-state 在缺乏藍光或處高溫 狀況下,其穩定性高且生命週期可長達 5~20 年。Q-state 在吸收藍光後,retinal 再由 9-cis 轉變成 all-trans 及整個 BR 分子直接恢復至基態。另外,Gillespie 等人 (2002) 再針對 P-state 與 Q-state 進行更精密的研究,結果發現 P490nm 係由 P445nm 與 P525nm 之混合組 成,並且在此文章中提到除了如一般利用紅光誘發之外,尚可利用 blue membrane (BM 或 dIbR,deionized BR) 與 light-induced blue membrane (LIBM,利用 532 nm 波長的雷 射光直接激發) 在吸收光能後直接產生 P-state,如 Fig. 3-18 所示 (Gillespie et al., 2002)。
(a) (b)
在光循環過程中,值得注意的是 M-state 與 Q-state,其它們的吸收波長均與基態 BR568 的吸收位置 568 nm 相距甚遠,在光譜上不產生重疊與干涉,此發現奠定了 BR 在光感 測與光儲存元件之製備與應用上的有利條件。
N K216 H
9-cis 15-anti all-trans 15-anti
N H K216 red-light
Fig. 3-17 The isomers of retinal chromophore between O-state and P-state (Q-state).
Fig. 3-18 Possible photochemical and thermal pathways involving the P and Q states of bacteriorhodopsin. (Gillespie et al., 2002).
3-6 PM 的固定化與方向性 的固定化與方向性 的固定化與方向性 的固定化與方向性
藉由物理性或化學性吸附法可將 PM 固定於基材上,以利於光電流訊號、光學中間 物和表面結構之量測分析或應用設備之製備。其方法包含了 Langmuir-Blodgett (LB) 沈 積吸附 (Weetall and Samuelson, 1998)、電場與磁場的沈降作用 (Keszthelyi, 1982;Dér et al., 1995)、靜電力吸附 (Fisher et al., 1978;Brizzolara and Beard, 1994;Brizzolara, 1995;
He et al., 1998a;He et al., 1998b;Oesterhelt et al., 2000;Chu et al., 2003)、高分子或凝 膠的包覆 (Wu et al., 1993;Beckmann et al., 1997;Fimia et al., 2003;Manoj and Narayan, 2003)、 抗原 -抗 體的 親和性 吸附 (Koyama et al., 1994) 與 交合 連結體 (conjugated crosslinker) (Sharma et al., 2004) 等技術。
在這些琳瑯滿目的固定化技術之中,以 Koyama 等人 (1994) 所發表之技術:運用 抗原-抗體的親和性吸附原理,將 PM 膜固定於 SnO2 電極上,最具重要性。他們將能分 別僅辨識 BR 分子的 N 端或 C 端之兩種單株抗體進行修飾,並且分別固定在表面修飾過 的 SnO2 電極上,以同時比較 PM 膜以紊亂排列、C 端或 N 端貼覆在電極表面上之三種 不同固定化方式下所獲得光電流訊號之差異。其實驗結果如 Fig. 3-19 所示,PM 膜的光 電流訊號大小以 C 端貼覆在電極表面上之情況為最大,其次是以紊亂排列之固定化方式 居中,最後則是以 N 端貼覆在電極表面上之方式為最小。其實驗除了提供一 PM 膜固定 化之方法外,也証明 PM 膜的“方向性”為影響光電流訊號大小之重要因素。He 等人
(1998b) 將帶正電性之高分子 (polydimethyldiallylammonium chloriede,PDAC) 塗佈在 表面帶負電性之基材上,以利用靜電力吸附原理分別使 PM 膜與帶有 D96N 突變膜蛋白 之 PM 吸附上去,並且重覆塗佈與沈積吸附,如 Fig. 3-20(a) 所示。其結果發現開燈 (light-on) 電流訊號會隨著塗佈層數目之增加而呈現正比關係,但超過其臨界值 (層數) 則呈現反比下降關係,如 Fig. 3-20(b) 所示。其實驗除了提供固定多層化之方法外,並 說明電流訊號大小與塗佈層數目的關係。
另外,Lu 等人 (1999) 結合基因工程與 LB 沈積吸附技術,直接將含有 D36C 突變 膜蛋白之 PM 固定在鍍金之石英晶片上,以達方向性與化學性吸附固定化之目的,如 Fig.
3-21(a) 所示。Lu 等人 (1999) 將已吸附 PM 膜之石英晶片分別以水與不同有機溶劑進 行多次清洗以去除單純利用物理性吸附而附著之膜蛋白,並同時利用石英振盪天平檢測 其吸附情形,結果如 Fig. 3-21(b) 所示,PM 與突變膜蛋白 D36C 均可吸附在石英金面 上,先利用水進行 4 次清洗後,兩膜蛋白仍然附著在金面上,隨後再分別以甲醇/水與己 烷/甲醇混合液各進行 3 次與 2 次清洗,發現到 PM 膜蛋白的頻率下降隨著清洗次數增加 而降低,而 D36C 突變膜蛋白則是穩定不變,此結果證明了 D36C 突變膜蛋白可藉由硫 基與晶片表面之 Au 原子形成共價鍵結,而固定於石英晶片上。
Fig. 3-19 Comparsion of the typical photoelectric response from a sandwich-type photocell comprising junctions of SnO2 electrode/PM monolayer film/aqueous gel/Au counter electrode (Koyama et al., 1994).
Fig 3-20 (a) Schematic illustration of PDAC/BR multilayers. (b) Magnitude of the light-on photocurrent versus the number of bilayers of PDAC/BR. (He et al., 1998b).
Fig. 3-21 (a) Scheme of BR D36C (Asp-36 in the loop between helix A and B is replaced by Cys) and the film deposition process. (b) Frequency shifts of QCM after deposition of PM LB films and washing with water or organic solvent (solvent A, methanol:H2O = 2:1;solvent B, hexane:methanol = 4:1) (Lu et al., 1999).
3-7 PM 之應用 之應用 之應用 之應用
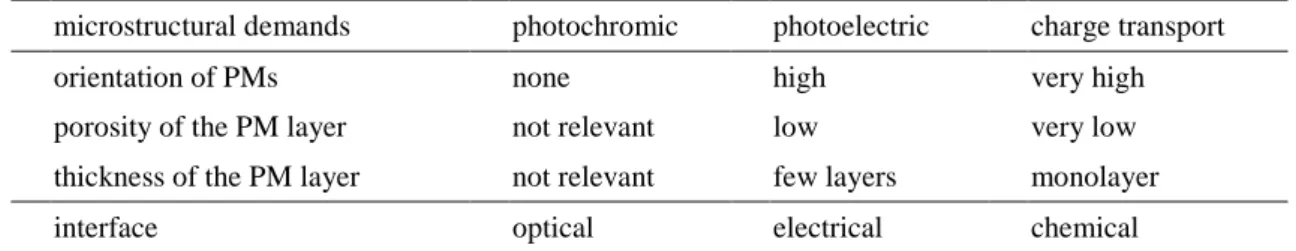
BR 蛋白質具有可產生先光電流訊號 (可將光能轉換成電化學能)、多種且明顯的光 中間體、可承受高溫與高酸鹼值範圍等其他一般生物材料所無法具備的特殊性質。但由 於 BR 本身為膜蛋白之一,因此必需利用脂質來維持其 3D 立體結構,以及二維六角晶 格之 PM 的完整性與活性。此外 BR 另具有其他優點,包含:(1)材料取得容易,直接將 菌體大量培養與純化分離後即可取得,並且對於自然環境不會造成損害和污染,為一“綠 色”材料;(2)可藉由基因工程與突變技術、retinal 基團的不同取代、物理或化學性的修 飾以改變其特性以提昇其應用性;(3)高光靈敏性。這些優越性質再加上其本身尺寸極 小,使得 BR 成為一極具前瞻性的生物性材料之一,在國際市場上的價格相當昂貴,可 被運用在光學訊息處理和光儲存 (二進製光記憶)、生物晶片和生物計算機、全息照相和 存儲、光過濾片、光電池、光開關 (photoswitch) 與感測器之製造等。Hampp (2000a) 曾 對於 BR 的應用做了一番文獻的回顧與探討,除了敘述 BR 許多眾人皆知的基本特點與 原理之外,對於 BR 主要的應用範圍可歸類分成電荷傳遞 (charge transport)、光致變色 (photochromic) 與光電反應 (photoelectric) 三大種類,如 Table 3-4 所示。Table 3-5 為其 在各系統應用上所需達到之條件。
Table 3-4 Technical applications suggested for bacteriorhodopsin (Hampp, 2000a)
charge transport charge transport
ATP generation in reactors information storage
desalination of seawater 2-D storage
conversion of sunlight into electricity 3-D storage
photoelectric holographic storage
ultrafast light detection associative memories
artifical retinas information processing
motion detection optical bistability / light switching
miscellaneous optical filtering
2nd harmonic generation signal conditioning
radiation detection neural networks
biosensor applications spatial light modulators
etc. ……… Phaseconjugation
ppttern recongnition Interfermetry
Table 3-5 Structure demand and interface of BR system based on the phototransport、
photochromic and photoelectric properties of BR (Hampp, 2000a)
microstructural demands photochromic photoelectric charge transport
orientation of PMs none high very high
porosity of the PM layer not relevant low very low
thickness of the PM layer not relevant few layers monolayer
interface optical electrical chemical
光電流裝置 (photovoltaic devices,PD) 是利用 BR 吸收光子後所產生一電位差 (光 能→電能),配合前敘章節中所提到的固定化技術與藉助精密儀器來量測光電流值或光 電壓差之裝置,在電荷傳遞與光電反應的應用範圍上可見到蹤跡。PD 其名稱會隨著研 究者之目的與應用而有所改變,因此亦可被稱之為 photobiosensor、photocell、transducer 或 photodetector 等 (Nicolini et al., 1998;Vsevolodov, 1998b;Nicolini et al., 1999;
Bertoncello et al., 2003;Wang et al., 2004)。這些裝置除了上述基本原理之外,在構造上 通常具有一個共通的特徵:三明治裝置,即以電極/ PM /電極的方式呈現,而在電極與 PM 之間可以利用導電性物質 (如:高分子) 或電解質溶液作為傳導媒介或電極與 PM 直 接接觸。
BR 在光致變色應用範圍上可分成資訊儲存 (information storage) 與資訊程序 (information processing) 兩部分,但以光儲 存體 (optical memory) 與全息式記憶體 (holographic memory) 最為人所知。兩者之間的差別在於光儲存體的儲存方式是以平面 式 0 與 1 之數位形式進行儲存(二維);而全息式記憶體則是以三度立體空間影像之形式 進行儲存 (三維)。BR 在光致變色應用上,主要是利用 Q390 與 M410 二狀態的吸收波長 均與基態 BR568的吸收位置相距甚遠,在光譜上不產生重疊與干涉,以及它們穩定生命 週期的長短不同 (Birge et al., 1999)。全息式記憶體的運作基本原理與全息照像術相同,
先將帶“資訊”光束以光調節器加密,再通過一參考光束,利用了兩光束干涉原理成像 (二 維→三維,即立體成像,並具有明暗對比),直接記錄在以感光材料製造的儲存體上,
可被運用在高容量數位 DVD 光碟或高容量硬碟儲存體之製造。在電腦中,所有資料的 運算與儲存都是以數位形式來進行,即以 0 與 1 來表示 (binary code),而 BR 分子的兩 中間體 BR568與 Q390在光儲存體應用上分別代表 0 與 1,如 Fig. 3-22 所示 (Vsevolodov, 1998c),包含資料寫入、讀取與抹除三部分。在資料寫入部分,先將已塗佈 BR 分子的 基板利用綠光激發後並轉換至 O-state,再利用紅光激發產生 P-state,P-state 則隨即轉換 成 Q-state,而未吸收紅光的 BR 分子則恢復至 BR568,因此在圖形上可形成 0 與 1 的矩 陣排列,即完成資料寫入工作,以符合電腦儲存資料之模式。在資料讀取部分,同樣地 以綠光激發 BR 分子後,再以紅光照射基板,此時先前的 BR568會因吸收紅光後轉換成 Q390, 而 先 前 已 轉 換 成 Q390 之 區 域 則 本 身 不 吸 收 紅 光 並 且 讓 紅 光 透 射 至 偵 測 器 (detector),使偵測器可獲得資料 (即先前所儲存的資料)。最後抹除工作則是直接以藍光 照射基板,讓所有的 Q390恢復至 BR568。因此,藉由綠光、紅光與藍光的交替使用,我 們可以輕易地進行資料寫入、讀取與抹除的工作。Hampp 等人 (2000) 則是將 BR 分子 運用在墨水染劑上,嘗試製作防拷墨水,可運用在防拷貝與反偽造辨識,增加了 BR 的 實用性與商業價值,如 Fig. 3-23 所示。其主要是利用 BR 分子本身光致變性之特性,即 BR568 (紫色) ↔ M410 (黃色) 之變化。首先,他們先在菱形區域外與正方形區域間塗上黃 色顏料以作為參考區域,然後在菱形區域內塗上含有 BR 分子的墨水染劑作為實驗區 域,並將其放置在黑白影印機上進行複印工作。實驗結果如 Fig. 3-23 所示,在經過複印
程序之後發現到不論是原稿或是所獲得複印之文件,其實驗區域之呈色結果均與以黃色 顏料為參考區域之色度相同 (如 Fig. 3-23C 與 Fig. 3-23D 所示),即無法顯示紫色與黃色 的對比;並且原稿在靜置一般時間後即可恢復原有之外觀 (即紫色與黃色的對比)。
此外,由於 BR 的吸收光譜 (工作光譜區) 主要位於了 500 nm ~ 600 nm 之間,並未 包含整個可見光光譜區域 (400 nm ~ 700 nm),因此在感測應用上被限制住。因此,Choi 等人 (1998) 利用黃素 (flavin) 主要可吸收波長 400 nm ~ 450 nm 範圍之光並產生光電 流訊號之特性,而利用 LB 技術將黃素和 BR 結合運用,製造出可以偵測可見光光譜區 域 400 nm ~ 650 nm 的雙頻光接受器 (dual-band photoreceptor)。隨後,Choi 等人 (2001) 利用先前的研究成果更進一步地研究 BR 運用在色彩 (RGB) 影像偵測系統的探討,成 功地初步建立色彩 (RGB) 影像偵測系統,如 Fig. 3-24 所示 (Choi et al., 2001)。在波長 為 400 nm ~ 475 nm 之範圍內,主要是以黃素作為接受器,其所產生的電流訊為負值;
在波長為 475 nm ~ 520 nm,主要是以黃素與 BR 共同作為接受器,因此我們在儀器上的 電流訊可以獲得一合成波之圖形;在波長為 520 nm ~ 650 nm,則是以 BR 為接受器,其 所產生的電流訊為我們所熟悉的圖形。Choi 等人 (2001) 的研究結果可增進人類對於視 覺系統上的了解 (即人類的視網膜上分佈著能解析紅綠藍三種顏色的錐狀細胞,可分別 對不同顏色分布的色光照射產生刺激,並產生電子訊號刺激後方的神經細胞層,使得神 經細胞產生一動作電位傳遞至大腦整合產生視覺),更是除了基因突變方法之外,另一 個增進 BR 應用性之典範。
Fig. 3-22 The sequence of BR optical memory:(1)The first laser 1 impulse (green light) converts the BR molecules in the beam plane to the O state. (2) The second laser 2 impluse (red light) converts the programmed part of O-molecules to the state P. The rest remain in O.
(3)P quickly converts to Q. (4)The read-out occurs at the switch-on of laser 1 (green light) and at the activation of the Q state. (5) Laser 2 (red light) is then turned on, however, in a low power regime. With the help of this weak red light the information, recorded by molecules in O and Q states, is passed onto the diode grating (Vsevolodov, 1998c).
Fig. 3-23 Copyprotection based on bacteriorhodopsin. (Hampp et al., 2000).
Fig. 3-24 A schematic illustration of the principle of color recognition by the proposed biomolecular photoreceptor (Choi et al., 2001).
3-8 BR 表現系統 表現系統 表現系統 表現系統
對於古細菌的基因轉殖系統,Sowers 與 Schreier (1999) 曾做了一番文獻回顧與整理,
其文章包含了古細菌的菌株介紹、各菌株的轉殖方法 (transformation、transduction 與 conjugation)、效率與轉殖的質體等敘述。因此,在本章節中僅針對常用於 H. salinarum 的表現系統與質體作說明。
BR 基因列序的解開 (Dunn et al., 1981)、轉殖技術 (Cline and Doolottle, 1987) 與質 體建立 (Blaseio and Pfeifer, 1990;Krebs et al., 1993) 在 BR 的結構與功能研究歴程上,
扮演著舉足輕重的角色。早期 BR 的研究方式是將經突變的 bop 基因表現在異源 (heterologous) 系統上,利用 Escherichia coli 大量表現與純化 BO,再將 BO 以 all-trans retinal、脂質與清潔劑重新復性 (renaturation) 並形成膠脂粒子 (micelles) (Huang et al., 1981;Oesterhelt, 1982;Gilles-Gonzalez et al., 1986;Braiman et al., 1987),再進行光譜 與氫離子泵浦活性等分析,討論特定胺基酸的影響。Soppa 等人(1989) 則是進行同源 (homologous) 系統研究的學者之一,其利用 X 射線與紫外光直接對 Halobacterium sp.
GRB 菌株照射,以進行 BR 分子突變的研究,其所獲得突變膜蛋白之吸收光譜與上述利 用 E. coli 表現與復性的結果 (Mogi et al., 1988a) 大致符合。
pUBP2 與 pMPK54 (pMPK 系列) 為常應用於可表現 BR 突變蛋白的質體,兩質體均為 shuttle vector (即可同時存在於兩種不同生物體中之質體),其所採用的宿主菌株分別為 H. salinarum L33 與 H. salinarum MPK40。pUBP2 是由 Blaseio 與 Pfeifer (1990) 所建構
的,其由 pHH9 (phage ΦH 的衍生物,其亦包含 H. salinarum 的嵌入序列 ISH27)、抗 Ampicillin 基因、抗 mevinolin 基因與 E. coli origin 所組成的,如 Fig 3-25 所示;而 1.2 kb PstI-BamHI 的 bop 基因可藉由 PstI 與 BamHI 兩限制酶切點插入 pUBP2 中。H.
salinarum L33 是一株不會產生 BO 蛋白質與其他色素物質的菌株 (Wagner et al., 1981),
外觀上則是呈現透明或乳白色。而在 L33 的 bop 基因上具有一段嵌入序列 (insertion sequence,ISH2),導致菌株無法合成 BO,因此被選為可表現原生種 (wild-type) BR 與 突變蛋白的宿主菌株。
Fig 3-25 Restricition map of pUBP2. (Blaseio and Pfeifer, 1990).
pMPK54 是由 Krebs 等人 (1993) 所建構的,其包含了 pGRB1 (存在於 Halobacterium sp. GRB 菌株中的質體)、ISH11 (嵌入序列 ISH1 的衍生物)、抗 Ampicillin 基因、抗 mevinolin 基因與大腸桿菌質體 pAT153;而 pMPK62 是將 pMPK54 插入一段具有 1.6 kb BamHI-BamHI 的 bop 基因片段,如 Fig 3-26 所示。H. salinarum MPK40 是一株外觀呈現 橘 紅 色 的 嗜 鹽 菌 株 , 其 染 色 體 上 的 bop 基 因 已 被 利 用 同 源 交 換 (homologous recombination) 方式破壤並刪除 (knock-out) 大部分基因片段,而使 bop 基因僅剩大約 250 bp。Krebs 等人 (1993) 在建立 H. salinarum 的表現系統與質體時,發現利用 H.
salinarum MPK5 (此菌株之 bop 基因上被插入一段 ISH1 基因,因此亦不會產生 BO) 為 宿主菌株,質體與菌株染色體的 bop 基因先經第一次同源交換後,菌株可獲得完整基因 片段 (包含原生種 BR 基因與突變基因),但再經過二次同源交換並於不含 mevinolin 之 培養基中培養後,菌株染色體中仍然具有含原生種 BR 基因之機率,如 Fig. 3-27 所示。
因此 Krebs 等人 (1993) 為避免原生種 BR 基因所產生的“汙染”,再重新設計一“特殊”
質體,稱之為 pMPK66,不包含 pHH9 基因片段,並且其 bop 基因片段上插入一合成列 序,可利用同源交換的方式來破壤與刪除大部分完整的 bop 基因片段 (約 600 bp 之片 段),最後獲得 H. salinarum MPK40 菌株。除此之外,Krebs 等人 (1993) 進一步地利用 此系統來表現 BR 的突變膜蛋白 (例如:R82A、D85E、D96N、D212N 等等),其結果 發現大部分突變膜蛋白的實驗數據與先前利用 all-trans retinal、脂質與清潔劑重新復性 並形成膠脂粒子的實驗結果相符合,但是唯有突變蛋白 D212N 的光譜產生了紅位移現 象,與先前的藍位移現象不一致。此實驗結果間接地說明表現系統 (同源與異源系統) 的 不同,會導致所表現 BR 膜蛋白之特性結果略有差異。
3-9 BR 的突變 的突變 的突變 的突變
BR 的突變研究除了幫助人們對於其結構功能的了解之外,另一附加的目的就是增 進 BR 之應用性與商業價值 (Wise et al., 2002)。在早期 BR 突變研究之目的是了解突 變對於 BR 氫離子的釋放與吸收、吸收光譜與光循環的影響,並了解部分供胺基酸在 BR 分子上所扮演角色、影響範圍與重要性。這些研究主要是針對可能涉汲氫離子傳遞 與靠近 retinal 週遭的胺基酸來進行突變。而研究方式是將突變 bop 基因以異源系統表
pHH9 mev-r E. coli ori amp-r
pUBP2 12.3 kbp HSPBHK
E
K
K S
S Sm
E
現 (例如:E. coli),並利用 all-trans retinal、脂質與清潔劑形成膠脂粒子並重新復性,討 論特定胺基酸的影響。隨後,Blaseio 和 Pfeifer (1990) 與 Krebs 等人 (1993) 建立了 H.
salinarum 專屬之同源表現系統,但是這些學者卻發現部分突變蛋白,在不同的表現系 統下其所獲得的實驗結果略有所差異或矛盾。此乃因為重新復性的突變蛋白本身缺乏菌 體脂質 (native lipid),造成無法正確地再摺合 (refolding) 與形成三聚體,而通常只以單 體存在 (Brown, 2000)。
Fig 3-26 Construction of mevinolin-resistance plasmids and bop expression vector. (Krebs et al., 1993).
Fig 3-27 Products of homologous recombination in MPK5 transformed with pMPK62 derivatives. (Krebs et al., 1993).
3-9-1 D85N 與與與與 D96N 突變突變突變 突變
D85 與 D96 分別位於 SB 的兩側 (即 D85 位於 EC 端;D96 位於 CP 端),並且在氫離 子傳遞上扮演著不同重要角色。D85 為一重要氫離子接受者,可接受 SB 釋放出的氫離 子,與 D212 可共同穩定 SB 區域水分子 W402,並且同時主宰著 L550 nm→M410 nm的速率。
先前學者將 D85 以 asparagine 取代後,發現可導致:(1)吸收光譜產生紅位移 (10 ~ 50 nm);(2)降低 BR 的氫離子傳遞能力;(3)延遲 M-state 的“生成”;(4)改變 BR 分子的結構,
進而影響 D96 的 pKa值 (Otto et al., 1990;Needleman et al., 1991;Krebs et al., 1993;Turner et al., 1993;Kataoka et al., 1994;Tittor et al., 1995;Dickopf et al., 1995)。反之,D96 與
Sp H A
B B B
N B N B
Sp
pGRB1 ISH11
AmpR MevR
pMPK 62 10.2 kbp
bop B Sp
H
Sp
pGRB1 ISH11
AmpR MevR
pMPK 54 8.6 kbp
N N
A B