國立宜蘭大學自然資源學系碩士論文
Department of Natural Fesources National Ilan University
Master Thesis
和平北溪森林植物社會沿海拔梯度之物種多樣性研究 Study of species diversity along elevation gradients of forest
community in the Heping north river
指導教授:陳子英 博士
Tze-Ying Chen Ph. D.
研究生:洪淑華
Shu-Hua Hung
中華民國九十六年六月
摘要
本文探討和平北溪北坡森林植群沿海拔梯度的植群及物種多樣性之變 化,與植物社會沿海拔梯度物種豐量度類型的改變。
調查上由海拔100 ~ 3400 m,平均每上升 100 m 至少設置一個樣區,共 設置35 個 600 m2樣區,並記錄維管束植物種類129 科 331 屬 582 個分類群。
經TWINSPAN 分析及參考 DCA 結果,可將植物社會區分為 6 個植群帶 10 個植群型,分別為榕楠林帶的茄冬大葉楠型;楠櫧林帶的黃杞長尾尖葉 櫧型;櫟林帶下層的台灣杜鵑型、大葉石櫟猪腳楠型;櫟林帶上層的台灣 扁柏台灣杜鵑型、台灣水青岡型及阿里山灰木白花八角型;鐵杉雲杉林 帶的台灣鐵杉型、台灣二葉松型;冷杉林帶的台灣冷杉型。CCA 分析結果 以土壤含石率、岩石地比例、土壤pH 值、土壤可交換性鈣及鎂與海拔梯度 具有極顯著之相關。物種多樣性在喬木層、草本層、附生植物層及藤本植 物層沿海拔梯度呈左偏之單峰分布;喬木層、灌木層、藤本植物及附生植 物層的物種多樣性在楠櫧林帶最高;草本層則在櫟林帶下層最高。β 多樣 性沿植群帶的轉換程度有逐漸變快的趨勢,但在櫟林帶轉換率變慢。物種 豐量度由低海拔至高海拔的分布類型由對數常態分布、分割線段模式、對 數常態分布、對數級數分布而轉為幾何級數分布,顯示植群帶物種豐量度 的分布類型,由低海拔物種複雜之多樣化環境轉為高海拔環境貧乏之單一 優勢種模式。
關鍵字:和平北溪、清查多樣性、分化多樣性、植群多樣性、植群帶、物 種豐量度
Abstract
An objective that variations of vegetation and species diversity along
altitude gradient were investigated forest community in north aspect of Heping north river. And show that variations of community of species abundances along altitude gradient.35 plots of 600 m2 were surveyed from altitude 100 to 3,400 m, were sampled one at least to rise 100 m along the altitude gradient. The results show that there are 129 families, 331 genera, 582 vascular plants. 6 vegetation zones and 10 vegetation types were classified using TWINSPAN and DCA: Bischofia
javanicaMachilus japonica var. kusanoi type on FicusMachilus zone;
Engelhardia roxburghianaCastanopsis cuspidata var. carlesii type on MachilusCastanopsis zone; Rhododendron formosanum type and Pasania kawakamiiMachilus thunbergii type on Quercus lower zone; Chamaecyparis obtusa var. formosana—Rhododendron formosanum type, Fagus hayatae type, Symplocos arisanensisIllicium anisatum type on Quercus upper zone; Tsuga chinensis var. formosana and Pinus taiwanensis type on TsugaPicea zone;
Abies kawakamii type on Abies zone. Ratio stone of soil, ratio of rock, soil pH,
soil exchangeable Ca, and soil exchangeable Mg are more significant with altitude gradient using CCA. Species diversity of trees, herbaceous, epiphytes, lianas, that show peaked at the left along altitude gradient and vegetation zone.Species diversity of trees, shrubs, lianas, epiphytes were maximum at
MachilusCastanopsis zone, while that of herbaceous was maximum at Quercus
lower zone. β-diversity gradually turn over fast along vegetation zone, while slower in Quercus zone. Rank/abundance plot of Species abundance according to priority show log normal distribution, broken stick model, log normaldistribution, log series distribution, geometric series distribution along altitude gradient. Represent that species complicated at vary environment to become alone speices at poor environment along altitude.
Keywords: Heping north river, inventory diversity, differentiation diversity,
vegetation diversity, vegetation zone, species abundance.
誌謝
何其有幸,從大學到研究所能一直從事自已的興趣,做自己喜歡做的 事,雖然植群調查很辛苦,但是能親臨感受台灣山林的美,為台灣森林進 一份心力,是一直支持我的動力。在與山林互動的期間,讓我體會了謙卑 與尊重,感動與驚喜,這是課堂上所學不到的。
研究所修業期間承蒙 陳子英博士的指導,不論是課業、野外調查上都 給予大力的支持,讓我在就學期間能無後顧之憂,專心所學,在生活上也 時時關心,教導做人處世的道理,讓我成長許多。
感謝台灣大學謝長富博士及屏東科技大學葉慶龍博士撥空細心批閱初 稿,提供寶貴的意見,並遠到而來為學生指正。感謝宜蘭大學蔡呈奇博士 提供土壤化學分析設備上及技術上的幫助,以及初稿的批閱指正,讓論文 更加完善,在此深表致意。
感謝余秋豐技正對論文初成時的指正。宋梧魁、程宗德學長在課業、
野外調查及生活上的照顧,野外調查感謝葉清旺、李智群、林靜香及魏瑞 廷學長姊,林家榮、曾啟政、卓子右、邱宗儀、邱孟韋、李坤益、鄭婉倩、
邱韋達同學,感謝你們無怨無悔的陪我上山下海,爬過懸崖峭壁,和你們 一起上山的日子真的很開心、難忘。以及楊宣惠學姊在蕨類鑑定上,吳欣 玲學姊在行政工作上的幫忙。還有荒野保護協會的廖英杰老師及王俊銘,
武塔的獵人朋友們韋文豪、曹光輝、韋文平、韋文明,有你們的幫助,讓 我更了解和平北溪的人文歷史,重要的是能讓大家平安下山。土壤分析方 面感謝張瑀芳學姊、黃子瑜學妹、許佳雯及楊國祥同學的協助。高職同學 吳美瑤對英文摘要的指正。羅東的舅公、嬸婆對我的照顧,讓我雖遠在宜 蘭也有在家的感覺。還要謝謝我所有的朋友,謝謝你們三不五時的精神喊 話,讓我能充滿活力的迎接每一天。
最後當然要感謝我的家人爸爸、媽媽,對我的信任及支持,讓我能毫
無拘束的做自己想做的事,大哥正龍、大嫂靜儀、大妹淑婷的鼓勵,還有 二妹淑媚文獻搜集的幫助,謹以此論文,獻給我最可愛的家人。
研究生 洪淑華 謹誌
目次
摘要……….…Ⅰ Abstract………Ⅱ 誌謝……….Ⅳ 目次……….Ⅵ 圖目次……….…Ⅷ 表目次……….…Ⅸ
壹、 前言……….…1
貳、 前人研究……….3
一、植群研究………..3
二、物種多樣性理論………..8
三、國內外物種多樣性沿海拔梯度之相關研究..………..……….13
參、研究區域概述……….. 21
一、地理位置……….21
二、地質與氣候……….22
肆、研究方法………...24
一、植物社會之調查及資料收集……….24
二、環境因子之估測……….24
三、資料分析……….…………27
伍、研究流程………...32
陸、結果……….……..33
一、物種組成……….33
二、植物多樣性……….33
三、植物社會之分布序列
...
42四、植物社會變異與環境因子的相關性……….43
五、清查多樣性……….46
六、分化多樣性……….53
七、物種豐量度……….57
八、植物社會結構沿植群帶之變化……….58
柒、討論………...61
一、和平北溪植群多樣性……….61
二、和平北溪植物社會與環境的關係……….64
三、清查多樣性……….65
四、分化多樣性……….68
五、物種豐量度……….69
六、植物社會結構沿海拔梯度之變化……….71
捌、結論………...72
玖、參考文獻………...74
附錄一、和平北溪北坡樣區之植物名錄………...…81
附錄二、和平北溪流域樣區之環境因子……….106
附錄三、和平北溪樣區之土壤化學性質……….107
附錄四、和平北溪喬木層、灌木層、草本層、藤本植物層及附生植物層之 Shannon(H)及均勻度(EH)指數………..…………...108
附錄五、和平北溪樣區之物種豐富度……….109
圖目次
圖1 四種物種豐量度分布之等級-豐富度曲線圖………...13
圖2 和平北溪流域圖……….………21
圖3 太平山地區生態氣候圖………...……….……….23
圖4 和平北溪北坡樣區位置圖……….24
圖5 和平北溪北坡樣區在 DCA 第一、二梯度軸之分布圖………43
圖6 和平北溪樣區及環境因子在 CCA 第一、二軸之分布圖…………..…...46
圖7 和平北溪樣區沿海拔之物種豐富度……….47
圖8 和平北溪樣區所有物種及各生活型物種豐富度沿海拔梯度之變化…..48
圖9 和平北溪樣區各生活型之 Shannon-Wiener 及均勻度指數沿海拔梯度之 變化……….49
圖10 和平北溪樣區不同生活型植物在植群帶內之物種多樣性…………...53
圖11 和平北溪不同生活型沿植群帶之β多樣性………….………...54
圖12 和平北溪植物社會不同生活型在各植群帶之β多樣性………...56
圖13 和平北溪各植群帶之等級-豐富度曲線圖……….57
圖14 植群帶中相對優勢度及胸高斷面積最大之前三科………...59
圖15 最大胸徑值沿植群帶之變化……………….……...60
圖16 物種豐富度與土壤 pH 值、土壤有效性磷、土壤可交換性鈣及鎂之相 關………...67
圖17 和平北溪植物社會沿植群帶之等級-豐富度曲線圖……….70
表目次
表1 和平北溪鄰近流域之植群型一覽表………...…5
表2 物種多樣性及植群多樣性之測計形式及尺度………...9
表3 國內外文獻 α 多樣性沿海拔梯度之變化……...15
表4 國內外文獻 β 多樣性(物種轉換)沿海拔梯度之變化...………..18
表5 太平山氣象站之月平均氣溫與月平均雨量氣候表(1996 年 1 月-2006 年 11 月)………...…22
表6 大濁水雨量站月平均降雨量表(1982-2005 年)………22
表7 和平北溪維管束植物調查結果………...…33
表8 和平北溪植群概要表...………..……34
表9 和平北溪北坡 DCA 三軸之特徵值、梯度軸長及變異量………42
表10 所有環境因子與 CCA 三軸之相關性……….44
表11 主要環境因子與 CCA 三軸及海拔之相關性………..45
表12 和平北溪植群帶之喬木層、灌木層、草本層、藤本植物及附生植物層 之物種多樣性……….51
表13 和平北溪植物社會不同生活型在各植群帶之 DCA 第一軸軸長……55
表14 台灣中部山地(蘇鴻傑,1992)、和平北溪鄰近流域及和平北溪植群帶 之海拔比較………...………..62
壹、 前言
研究物種多樣性是探究生物多樣性(biodiversity)最廣泛也是最方便的 方法,而沿著海拔梯度物種多樣性的研究更是目前國際上生態學者所關注 的問題(蘇鴻傑,2002;唐志堯、方精云,2004;方精云等,2004;朱珣之、
張金屯,2005; Whittaker and Niering, 1975; Su, 1984,1994; Buot and Okitsu, 1998; Sklenár and Jørgensen, 2001; Wang et al., 2002; Bhattari and Vetaas, 2003; Arturo and Lauro, 2005)。台灣地形複雜,氣候多變,地理位置介於熱 帶及溫帶之間,在這小島上已經發現有4000 多種植物之多(謝長富,2005),
也逐漸引起國內重視物種多樣性之研究。一般物種多樣性常受海拔、濕度、
地形、森林結構、干擾等影響,其中又以海拔所造成的影響最大(廖啟政,
1995;蘇鴻傑,2002; Whittaker, 1975; Su, 1994; Bhattari and Vetaas, 2003),
因此瞭解物種多樣性沿海拔梯度的關係是十分重要的。
物種多樣性依測計形式不同又分為清查多樣性(inventory diversity)及分 化多樣性(differentiation diversity);前者在測計一生態單位內之物種豐富 度,是一種計數之清查方式;後者為度量局部生態群落或小單位間物種轉 變的狀況,即度量小單位間之差異程度。依測計形式的不同又分為較被廣 為探討的 α 及 β 多樣性(蘇鴻傑,2004)。
目前生態學者對 α 多樣性沿海拔梯度的分布提出三種類型:(一)物種 多樣性隨海拔升高而減少(蘇鴻傑,2005;Whittaker and Niering, 1975; Su, 1994);(二)物種多樣性隨海拔升高先增加後降低,形為單峰分佈(unimodal distribution) (王國宏,2002;葉清旺,2004;廖家宏,2006; Kessler, 2001; Wang
et al., 2002; Arturo and Lauro, 2005; Bhattarai and Vetaas, 2006);(三)隨海拔升
高多樣性增加,此情形較為少見,多發生在極端環境(Sklenár and Ramsay, 2001)。物種多樣性沿海拔梯度的轉換程度(β多樣性)亦有三種分布類型:(一)
隨海拔升高轉換程度變慢,常發生在極端環境下的高山稀疏草原中(Sklenár and Ramsay, 2001; Wang et al., 2002);(二)沿海拔梯度無規律變化(Kessler, 2000);(三)隨著海拔上升,轉換程度變快。由此可知,不同地區、尺度之 物種多樣性的反應變化也不同。
目前國內雖已有物種多樣性之研究(Su, 1994),但大多的研究區域海拔 範圍都不大(廖啟政,1995;葉清旺,2004;廖家宏,2006;魏瑞廷,2006),
因此所呈現的多樣性變化只是海拔範圍的大小所影響,無法表現整個物種 多樣性沿海拔梯度的變化。而植物社會中不同植群分帶、生活型之物種對 海拔梯度的反應變化也不盡相同(Whittaker and Niering, 1975; Kessler, 2001;
Bhattarai and Vetaas, 2003; Arturo and Lauro, 2005; Hemp, 2006; Kluge et al., 2006)。
和平北溪流域位於宜蘭縣與花蓮縣交界,屬於東部地理氣候區(Su, 1985),海拔由 0-3592 m,本研究以森林社會沿海拔梯度向上來進行取樣調 查,建立物種多樣性沿海拔梯度的分布型態資料,並探討下列幾個議題:(一) 和平北溪北坡天然森林植群型主要的海拔分帶;(二)探討不同生活型物種沿 海拔梯度及森林植群帶之清查多樣性及分化多樣性變化;(三)森林社會沿海 拔梯度之物種豐量度類型的變化。
貳、前人研究
一、植群研究
和平溪流域因地勢險峻,調查困難,至今並無完整且全面性的物種清 查及植群調查研究,僅就和平北溪鄰近地區之植群調查文獻論述,包括廖 秋成(1979)清水山石灰岩地區、林旭宏(1988)南澳闊葉樹保護區、高瑞卿 (1995)於立霧溪流域及葉清旺等(2006)南澳溪流域植群分類之研究,整個植 群帶的海拔劃分,區分之特徵種及優勢種如表1。
整理和平溪流域附近之植群多樣性,以植群帶劃分大致可分為:
(一)冷杉林帶(Abies zone):海拔分布約 3000-3200 m,特徵種及優勢種 均以台灣冷杉(Abies kawakamii)為主。
(二)鐵杉-雲杉林帶(Tsuga-Picea zone):海拔分布約 2000-2950 m,特徵 種為台灣鐵杉(Tsuga chinensis)、台灣雲杉(Picea morrisonicola)、台灣二 葉松(Pinus taiwanensis)、台灣五葉松(Pinus morrisonicola)及高山新木薑 子(Neolitsea acuminatissima),優勢種為台灣鐵杉、台灣雲杉、台灣二 葉 松 、 台 灣 五 葉 松 、 紅 檜(Chamaecyparis formosensis) 、 赤 柯 (Cyclobalanopsis morii)、三斗石櫟(Pasania hancei var. ternaticupula)、
台灣雲杉、高山櫟(Quercus spinosa)、昆欄樹(Trochodendron aralioides)、
狹葉櫟(Cyclobalanopsis stenophylloides)。
(三)櫟林帶上層(upper Quercus zone):海拔分布約 1050-2350 m,特徵 種為假長葉楠(Machilus japonica)、台灣赤楊(Alnus formosana)、台灣水 青 岡(Fagus hayatae) 、 長 葉 木 薑 子 (Litsea acuminata) 、 台 灣 扁 柏 (Chamaecyparis obtusa var. formosana),優勢種為昆欄樹、赤柯、紅檜、
大葉石櫟(Pasania kawakamii)、狹葉櫟、薯豆(Elaeocarpus japonicus)、
高山櫟、三斗石櫟、高山新木薑子、白花八角(Illicium anisatum)。
(四)櫟林帶下層(lower Quercus zone):海拔分布約 800-1900 m,特徵種 為 小 葉 白 筆(Symplocos modesta)、 大 葉 石 櫟 、 短 尾 葉 石 櫟 (Pasania
harlandii)、台灣八角(Illicium arborescens)、油葉石櫟(Pasania konishii)、
台灣水青岡、台灣杜鵑(Rhododendron formosanum)、假長葉楠,優勢種 為猪腳楠(Machilus thunbergii)、黃杞(Engelhardia roxburghiana)、狹葉 櫟、長尾尖葉櫧(Castanopsis cuspidata var. carlesii)、薯豆、大明橘 (Myrsine sequinii) 、 奧 氏 虎 皮 楠 (Daphniphyllum glaucescens subsp.
oldhamii var. oldhamii )、大葉石櫟、錐果櫟(Cyclobalanopsis longinux)、
杜 英(Elaeocarpus sylvestris)、長葉木薑子、毽子櫟(Cyclobalanopsis
sessilifolia) 、 厚 皮 香 (Ternstroemia gymnanthera) 、 鵝 掌 柴 (Schefflera octophylla)。
(五)楠櫧林帶(Machilus-Castanopsis zone):海拔分布約 350-1200 m,特 徵種為青剛櫟(Cyclobalanopsis glauca)、長尾尖葉櫧、大明橘、大頭茶 (Gordonia axillaris)、西施花(Rhododendron leptosanthum)、菱果石櫟 (Pasania synbalanos)、台灣楊桐(Adinandra formosana)、黃杞、厚殼桂 (Cryptocarya chinensis) 、 黃 連 木 (Pistacia chinensis) 、 金 毛 杜 鵑 (Rhododendron oldhamii) 、 三 斗 石 櫟 , 優 勢 種 太 魯 閣 櫟 (Quercus
tarokoensis) 、 栓 皮 櫟 (Quercus variabilis) 、 阿 里 山 千 金 榆 (Carpinus kawakamii) 、 台 灣 肉 桂 (Cinnamomum insularimontanum) 、 九 芎
(Lagerstroemia subcostata)、台灣雅楠(Phoebe formosana)、猪腳楠、鵝 掌柴、薯豆、錐果櫟、樹杞(Ardisia sieboldii)、長葉木薑子。(六)榕楠林帶(Ficus-Machilus zone):海拔分布約 600 m 以下,特徵種為 九 芎 、 澀 葉 榕(Ficus irisana) 、 櫸 (Zelkova serrata) 、 榕 樹 (Ficus
microcarpa) 、 咬 人 狗 (Dendrocnide Meyeniana) 、 菲 律 賓 榕 (Ficus
ampelas)、月橘(Murraya paniculata),優勢種為森氏紅淡比(Cleyera
japonica var. morii)、太魯閣千金榆(Carpinus hebestroma)、三葉山香圓
(Turpinia ternata)、大香葉樹(Lindera megaphylla)、猪母乳(Ficus表1 和平北溪鄰近流域之植群型一覽表
Table 1 Vegetation types nearby Heping north river
植群帶 植物社會 特徵種 優勢種 海拔分佈
冷杉林帶
台灣冷杉林型(高瑞卿,1995)
台灣冷杉 台灣冷杉、台灣鐵杉、巒大花 楸
3000–3200m
鐵杉
- 雲杉林帶
台灣鐵杉林型 台灣二葉松林型
台灣雲杉-高山新木薑子林型 (高瑞卿,1995) 華山松、玉山杜鵑、高山櫟中途簡叢
台灣鐵杉、紅檜、玉山杜鵑簡叢(廖秋成,1979)
台灣雲杉、台灣 鐵杉、台灣二葉 松、華山松、高 山新木薑子
厚葉柃木、尖葉槭、森氏杜鵑、
紅檜、赤柯[森氏櫟]、三斗石 櫟、台灣赤楊、台灣雲杉、台 灣紅榨槭、高山櫟、昆欄樹、
狹葉櫟
2000-2950m
櫟林帶上層 昆欄樹、赤柯簡叢
狹葉櫟、短尾葉石櫟、薯豆、厚皮香簡叢 狹葉櫟、高山櫟、厚皮香中途簡叢(廖秋成,1979) 針闊葉樹混合林型
紅檜-假長葉楠林型
台灣赤楊林型(高瑞卿,1995) 長葉木薑子-紅檜群叢
三斗石櫟-台灣水青岡群叢
台灣杜鵑-台灣扁柏群叢(葉清旺等,2006)
假長葉楠、台灣 赤楊、台灣水青 岡、長葉木薑 子、台灣扁柏、
昆欄樹、森氏櫟[赤柯]、紅檜、
大葉石櫟[大葉柯]、台灣二葉 松、阿里山千金榆、狹葉櫟、
短尾葉石櫟[短尾柯]、薯豆、厚 皮香、高山櫟、三斗石櫟、台 灣杜鵑、高山新木薑子、白花 八角、台灣樹參
1050-2350m
(續下頁)
表1(續)
Table 1 (continued)
植群帶 植物社會 特徵種 優勢種 海拔分佈
櫟林帶下層
狹葉櫟、蘭嵌千金榆、杏葉石櫟簡叢 狹葉櫟、大明橘、短尾葉石櫟簡叢 長尾尖葉櫧、奧氏虎皮楠、猪腳楠簡叢 大葉石櫟、長尾尖葉櫧簡叢(廖秋成,1979) 猪腳楠-小葉白筆林型(高瑞卿,1995) 錐果櫟-長尾尖葉櫧型
短尾葉石櫟-台灣八角亞型
錐果櫟-長尾尖葉櫧亞型(林旭宏,1988) 大葉石櫟-假長葉楠群叢
台灣杜鵑群叢
油葉石櫟-台灣水青岡群叢(葉清旺等,2006)
小葉白筆、大葉 石櫟、短尾葉石 櫟(短尾柯)、台灣 八角(紅花八 角)、油葉石櫟(小 西氏石櫟)、台灣 水青岡、台灣杜 鵑、假長葉楠
猪腳楠[紅楠]、黃杞、狹葉櫟、
長尾尖葉櫧[長尾柯]、薯豆、蘭 嵌千金榆、杏葉石櫟[校力]、大 明橘、奧氏虎皮楠、大葉石櫟[大 葉柯]、錐果櫟、南投木薑子、
杜英、長葉木薑子、毽子櫟、
高山新木薑子、厚皮香、月桂 葉灰木、鵝掌柴
800-1900m
楠櫧林帶 青剛櫟-呂宋莢迷林型 青剛櫟-呂宋莢迷亞型 太魯閣櫟-黃連木亞型 栓皮櫟-金毛杜鵑亞型
青剛櫟-三斗石櫟亞型(高瑞卿,1995) 九芎、台灣雅楠群叢(廖秋成,1979) 厚殼桂-猪腳楠群叢
大明橘-長尾尖葉櫧群叢(葉清旺等,2006)
青剛櫟、呂宋莢 迷、細葉饅頭 果、長尾尖葉 櫧、大明橘、大 頭茶、西施花、
菱果石櫟、台灣 楊桐、黃杞、厚 殼桂、黃連木、
金毛杜鵑、三斗 石櫟
太魯閣櫟、栓皮櫟、阿里山千 金榆、台灣肉桂[土肉桂]、台灣 雅楠、猪腳楠、鵝掌柴、薯豆、
錐果櫟、樹杞、長葉木薑子
350-1200m
(續下頁)
表1(續)
Table 1 (continued)
植群帶 植物社會 特徵種 優勢種 海拔分佈
榕楠林帶 澀葉榕-九芎林型
血桐-構樹林型(高瑞卿,1995)
森氏紅淡比、饅頭果、台灣梣、太魯閣千金榆中 途群叢
九芎、黃肉樹、鵝掌柴簡叢 三葉山香圓、猪母乳簡叢
三葉山香圓、澀葉榕、血桐中途簡叢(廖秋成,
1979)
櫸-大葉楠群叢 榕樹-大葉楠群叢 咬人狗-大葉楠群叢 菲律賓榕-大葉楠群叢
青剛櫟-大葉楠群叢(葉清旺等,2006)
九芎、澀葉榕(糙 葉榕)、櫸、榕樹、
咬人狗、菲律賓 榕、月橘
森氏紅淡比、饅頭果、台灣梣、
太魯閣千金榆、三葉山香圓、
大香葉樹[大葉釣樟]、猪母乳、
黃肉樹[小梗木薑子]、鵝掌柴 [江某]、血桐、茄冬、大葉楠、
台灣雅楠、五掌楠、瓊楠、野 桐、山黃麻、鵝掌柴、樹杞、
石朴
600m 以下
註:[ ]內為原始文獻之植物名稱
fistulosa) 、 黃 肉 樹 (Litsea hypophaea) 、 鵝 掌 柴 、 血 桐 (Macaranga tanarius) 、 茄 冬 (Bischofia javanica) 、 大 葉 楠 (Machilus japonica var.
kusanoi)、台灣雅楠、五掌楠(Neolitsea konishii)、瓊楠(Beilschmiedia erythrophloia)、野桐(Mallotus japonicus)、山黃麻(Trema orientalis)、鵝
掌柴、樹杞。二、物種多樣性理論
物種多樣性依研究尺度不同就會產生不同的物種多樣性型態,因此對 物種多樣性型態的研究應該從不同尺度著手(唐志堯、方精雲,2004);而又 依測計單位及物種分化的差異程度,可分為清查多樣性及分化多樣性(表 2)(蘇鴻傑,2004)。
(一)清查多樣性 1. 物種多樣性
物種多樣性依測計尺度不同有內部α多樣性、α多樣性、γ 多樣性及ε多樣性,本研究針對α及γ多樣性層級,茲敘述如下:
(1)α 多樣性
α 多樣性又稱社會多樣性(community diversity),為某一均 質生育地中,該社會所呈現之植物種數或棲地中物種的聚集程 度,此多樣性的高低亦受取樣樣區面積大小而影響,故取樣時 應大於最小取樣面積。α 多樣性的測量一般分為三種類型:物 種豐富度指數(species richness index)、物種多樣性指數(species diversity index)以及均勻度指數(evenness index)。
a. 物種豐富度指數(species richness index)
物種豐富度即物種的數目,為測量一定面積內之物種種 數,是最簡單、最古老的物種多樣性測度方法。
表2 物種多樣性及植群多樣性之測計形式及尺度(蘇鴻傑,2004) Table 2 The shape and scale of species diversity and vegetation diversity
多樣性形 式
清查多樣性inventory diversity 分化多樣性 differentiation diversity 考慮層級 物種多樣性 Species
diversity
植群多樣性
vegetation diversity
物種及社會(群聚)間 between sp. &
community
測計單位 物種數目 taxa(species) 植物社會數目 syntaxa 物種轉換、樣品間之差 異
測計尺度 1:區塊
內部 α 多樣性、次樣 品多樣性、點多樣性:
均質社會內微棲地樣 品內之種數
測計尺度 2:林分
α 多樣性、棲地內多 樣性:均質社會樣品之 種數
同生群多樣性:社會樣 品內小生境分化之同 生群數目
內部 β 多樣性、格局多 樣性:社會內部分化小 生境間之物種組成變化 程度
測計尺度 3:地景或 區域
γ 多樣性、地景或區 域多樣性:區域地景內 異質社會樣品之總計 物種數
植群型多樣性:區域地 景內異質植物社會基 本單位之數目,如群 叢、群系、優勢型或其 他單位之數目
β 多樣性、棲地間多樣 性:地景或區域內沿環 境梯度或不同社會樣品 間之物種組成變化程度
測計尺度 4:地理區 域
ε多樣性、地理區域多 樣性:廣大地理區域內 涵蓋不同地景之總計 物種數
聚落群多樣性:大地理 區域內異質植物社會 層級分類系統之單位 數目
δ多樣性、地理分化多 樣性:大氣候梯度或地 理區域之間物種組成變 化程度。
註:本研究只針對粗體部分進行探討
物種豐富度僅利用種數及個體數來計算植物社會的多樣 性,並未考慮個體數之分配情形。而物種多樣性指數則必須考 慮樣區大小以及物種分配的均勻度。最普遍及常用的物種多樣 性指數為Simpson 指數及 Shannon-Wiener 指數。
c. 均勻度指數(evenness index)
在一植物社會中,植物個體在不同種間之分佈均勻程度,
稱為該社會之均勻度指數。
(2) γ多樣性
γ多樣性又稱為區域多樣性(regional or gamma diversity),
由組合地區性所有樣區計算而來,亦即某一地理區域α多樣性 的總和(Su, 1994),其測計的方法及意義與α多樣性一樣,唯尺 度較大,因此又視為當地社會多樣性與社會分化數目之乘積,
以整個地區而言,悠久的安定環境及演化歷史、急劇變化的氣 候及地形與水分梯度,以及複雜的母岩與土壤基質,均有助於 提昇γ多樣性。本研究即以植群帶為γ多樣性。
2. 植群多樣性
物種多樣性只是生物層譜的一級而已,但生命現象之複雜性 及生物與環境之整體關聯性也是生態系多樣性所要考慮的,因此,
蘇鴻傑(2004)將物種多樣性觀念提升至生物社會之層級,即植群多 樣性,因為植群多樣性是生態系多樣性具體展現及測計的方法,其 測計單位為植物社會,在地景尺度上,植群型多樣性可顯示於群 叢、群系、優勢型或其他基本單位之數目。依不同測計尺度可分為 同生群多樣性、植群型多樣性及聚落群多樣性,本研究僅針對植群 型多樣性層級論述。
(二)分化多樣性
1. 物種轉換及社會(群聚)間之差異
依不同測計尺度可分為內部β多樣性、β多樣性及 δ 多樣 性,本研究針對β多樣性敘述如下:
(1)β 多樣性
β 多樣性可定義為沿環境梯度不同生育地間物種組成的 相異性或物種沿環境梯度的轉換(turnover)程度(馬克平,1994;
唐志堯、方精雲,2004;Whittaker, 1972),亦可稱為物種替代 速率(species replacement rate)或物種組成變化速率(the rate of change of species-composition) 樣區間物種組成差異愈大,即樣 區間轉換愈快,則 β 多樣性愈大;反之,則 β 多樣性愈小。
β 多樣性的測量方法主要有 Whittaker 的指數(βw)、Cody 的指 數(βc)、Routledge 指數(βR、βI、βE)、Wilson 與 Shmida 指 數(βT)等。其中以 Wilson 與 Shmida 指數(βT)的應用較為廣泛。
以上為 β 多樣性沿著環境梯度的變化,但不同生育地間的相似 性測量則以 Jaccard 指數和 Sørenson 指數的應用較為簡便且應 用最廣。
本研究的目的為沿著海拔梯度的變化,已有確切環境梯 度,且海拔梯度範圍較大,以解釋沿環境梯度轉換程度的βT 指數來表示較理想。βT 是(βw)及 (βc)的結合,能更具體表 現物種沿環境梯度的轉換程度,而不受α多樣性、樣本大小的 影響,因此研究上僅選用βT指數進行計算。
(三)物種豐量度模式(species abundance models)
物種豐量度可以個體數量、生物量、植物覆蓋度、頻度、斷面積 以及生產力等為測度指標(Whittaker, 1972),為了討論方便,多以物種 個體數量作為多度的測度指標。物種豐量度亦有均勻度特性助於分辨
群叢的組成(Magurran, 2004)。
多樣性指數是用一個數量值來表示植群中物種的多樣性,而物種 豐量度是用一組豐量度值及其排列來表示社會中的多樣性。通常以物 種等級-豐量度圖(ranked-abundance plot)表示之,能更全面地反應植群 特徵(張金屯,2004),也是研究植物社會結構最基本的方法之一(彭少 麟等,2003)。等級-豐量度曲線圖常是描述物種豐量度模式最好的表達 方式,陡峭的曲線表示社會有高優勢度,這有可能出現在幾何級數 (geometric series)或對數級數(log series)分佈;而較平緩的曲線表示有高 均勻度,較符合對數常態(log normal)分佈或分割線段模式(broken stick model) (Magurran, 2004),因為沒有一個模式能通用於所有植物社會,
因此物種豐量度的分布模式可以下列 4 種模式的表示效果較好(圖 1):
1.幾何級數分布(geometric series distribution):假設某地區的有效資源 一定,第一優勢種會先佔領有限資源的一定部分;第二優勢種又佔 領剩下的部分資源,依此類推,直到剩下的資源不能再維持一個物 種生存為止。物種豐量度的幾何級數分布多出現在物種貧乏的環境 或植群演替的早期階段,隨著演替的進展或環境條件的改善,物種 豐量度分布則可能轉變為對數級數分布(Magurran, 1988)
2.對數級數分布(log series distribution):在一個或少數幾個環境因子 佔主導地位的植物社會中,物種豐量度分布往往傾向對數級數分 佈,對數級數分布會比幾何級數分布來得均勻,但又不比對數常態 分布均勻。
3.對數常態分布(log normal distribution):生態學家們研究發現,大多 數社會中物種豐量度分布都較符合對數常態分布。對數常態分布本 身是隨機過程的產物,當每一個物種在取樣中的個體數量是隨機決 定而不依賴於它物種時,其物種豐量度常表現為對數常態分布。
4.分割線段模式(broken stick model):物種豐量度的對數常態分布雖 然在自然界中較普遍,但在某些範圍較窄、較均勻的社會中,物種 的相對豐量度往往取決於物種間的競爭關係。在這種情怳下,物種 的相對豐量度往往為分割線段模式。
圖1 四種物種豐量度分布之等級-豐量度曲線圖(摘自 Magurran (1988))
Figure 1 Four species abundance patterns of ranked-abundance curves(abstracted from Magurran (1988))
三、國內外物種多樣性沿海拔梯度之相關研究
生物多樣性沿著環境梯度的變化是生物多樣性研究的重點之一,而海 拔梯度又包含了多種環境因子的梯度效應(唐志堯、方精雲,2004),因此研 究生物多樣性沿海拔梯度變化的規律具有重要意義。而不同生活型植物的 多樣性沿海拔梯度亦呈現不同的變化,普遍來說草本比木本的分布變化又 更複雜。
(一)α多樣性
依山地海拔落差大小分為大尺度(落差超過 1000 m)及小尺度(落差 小於 1000 m)之多樣性,其 α 多樣性沿著海拔梯度主要有三種分布變
化,以不同生活型論述(表 3):
1.即物種多樣性隨海拔升高而減少:
(1)木本層:包括台灣沙里仙溪(Su, 1994)、菲律賓 Pulog 山 (Buot and Okitsu, 1998)、中國秦嶺牛背梁(唐志堯、柯金虎,
2004)、中國長白山北坡(趙淑清等,2004)、中國福建黃崗山 東南坡(鄭成洋等,2004)、中國秦嶺太白山(唐志堯等,2004)、
中國東靈山(任海保等,2005);在小尺度方面則有日本 Tatera 森林保留區(Itow, 1991)。
(2)灌木層:包括中國長白山北坡(趙淑清等,2004)、中國秦嶺太 白山(唐志堯等,2004)、中國東靈山(任海保等,2005)。
(3)草本層:台灣北大武山(廖家宏,2006)。
(4)藤本植物層:喜馬拉雅山區(Bhattarai and Vetaas, 2003)。
(5) 依 科 別 區 分 : 玻 利 維 亞 安 地 斯 山 脈 的 野 牡 丹 科 (Melastomataceae) (Kessler, 2001) 及 爵 床 科 (Acanthaceae) (Kessler, 2000)。
2.物種多樣性隨海拔升高先增加後降低,又稱單峰分布:
(1)所有物種:包括亞利桑那州 Santa Catalina 山區(Whittaker and Niering, 1975)、中國貢嘎山東坡(沈澤昊,2001)、喜馬拉雅 山區(Bhattarai and Vetaas, 2003)、中國秦嶺牛背梁(唐志堯、
柯金虎,2004)、墨西哥內華達山脈(Arturo and Lauro, 2005) 、 中國麗江地區(馮建孟等,2006)、中國天山(胡玉昆等,2007);
在小尺度方面有在加拉巴哥群島的 Isla Santa Cruz(Itow, 1992)。
(2)木本層:包括中國四川峨嵋山(Tang and Ohsawa, 1997) 、沙 烏地阿拉伯(Hegazy et al., 1998)、中國祈連山北坡
表3 國內外文獻 α 多樣性沿海拔梯度之變化
Table 3 Literature review ofαdiversity along the elevation
α 多樣性 國內外文獻
海拔落差
(海拔範圍) 相當氣候帶
海拔升高,
多樣性減少
隨海拔呈單 峰分佈
海拔升高,
多樣性增加
無明顯趨 勢 Su (1994) >1000 m
(1370-3500 m) 暖溫帶-冷溫帶 木本層 草本層
Kluge et al., (2006)
>1000 m
(100-3400 m) 熱帶-冷溫帶 蕨類植物
Hegazy et al., (1998)
>1000 m (0-2500 m)
熱帶-溫帶 木本層
Whittaker and Niering (1975)
>1000 m
(850-2760 m)
亞熱帶-涼溫帶 所有物種
Wang et al.
(2002)
>1000 m (1450-3700 m)
亞熱帶-亞寒帶 木本層
草本層 Kessler (2000) >1000 m
(400-2500 m)
熱帶-溫帶 爵床科 鳳梨科、仙
人掌科、天 南星科、野 牡丹科、蕨 類植物 Bhattarai and
Vetaas (2003)
>1000 m
(100-1500 m)
熱帶-亞熱帶 藤本植物 所有物種
木本層 灌木層
蕨類植物 草本層
Arturo and Lauro (2005)
>1000 m (2800-4000 m)
涼溫帶-亞寒帶 所有物種
木本層 草本層 灌木層 Sklenár and
Ramsay (2001)
>1000 m (3100-4800 m)
冷溫帶-亞寒帶 所有物種
Kessler (2001) >1000 m (220-3950 m)
熱帶-亞寒帶 野牡丹科 爵床科、天
南星科、鳳 梨科、棕櫚 科、蕨類植 物
(續下頁)
表3 (續)
Table 3 (Continued)
α 多樣性 國內外文獻
海拔落差
(海拔範圍) 相當氣候帶
海拔升高,
多樣性減少
隨海拔呈單 峰分佈
海拔升高,
多樣性增加
無明顯趨 勢 Tang and
Ohsawa (1997)
>1000 m (600-3099 m)
亞熱帶-涼溫帶 木本層
沈澤昊等 (2001)
>1000 m (1200-4500 m)
亞熱帶-亞寒帶 所有物種
胡玉昆等 (2007)
>1000 m (2340-4618 m)
溫帶-亞寒帶 所有物種
鄭成洋等 (2004)
>1000 m (450-2158 m)
亞熱帶-溫帶 木本層
唐志堯等 (2004)
>1000 m (1200-3200 m)
暖溫帶-冷溫帶 木本層 灌木層 唐志堯、柯金虎
(2004)
>1000 m (1500-2800 m)
暖溫帶-涼溫帶 木本層 所有物種 草本層
馮建孟等 (2006)
>1000 m (1100-5596 m)
暖溫帶-亞寒帶 所有物種
任海保等 (2005)
>1000 m (1280-2300 m)
暖溫帶-溫帶 木本層
灌木層
草本層
王國宏(2002) >1000 m (1500-3700 m)
暖溫帶-亞寒帶 草本層 灌木層
廖家宏(2006) >1000 m (800-3090 m)
亞熱帶-涼溫帶 草本層 木本層
趙淑清等 (2004)
>1000 m (700-2600 m)
亞熱帶-涼溫帶 木本層 灌木層
草本層
Itow (1992) <1000 m (0-860 m)
亞熱帶 所有物種
Buot and Okitsu (1998)
<1000 m (2000-2700 m)
溫帶 木本層
廖啟政(1995) <1000 m (200-467 m)
亞熱帶 木本層
Itow (1991) <1000 m (140-560 m)
亞熱帶 木本層
(續下頁)
(Wang et al., 2002)、喜馬拉雅山區(Bhattarai and Vetaas, 2003)、墨西哥內華達山脈(Arturo and Lauro, 2005)及北大武 山(廖家宏,2006)。
(3)灌木層:墨西哥內華達山脈(Arturo and Lauro, 2005)
(4)草本層:中國祁連山北坡(王國宏,2002)、中國祈連山北坡 (Wang et al., 2002)、墨西哥內華達山脈(Arturo and Lauro, 2005)、中國東靈山(任海保等,2005)
(5)依科別區分:玻利維亞安地斯山脈的鳳梨科(Bromeliaceae)、
天南星科(Araceae)、蕨類植物(Kessler 2000; Kessler, 2001)、
爵 床 科 、 棕 櫚 科 (Palmae) (Kessler, 2001) 、 仙 人 掌 科 (Cactaceae)、野牡丹科(Kessler, 2000)及哥斯大黎加的蕨類植 物(Kluge et al., 2006)。
3.隨海拔升高多樣性增加,此情形較為少見,多發生在極端環境:
(1)所有物種:包括厄瓜多爾高山(Sklenár and Ramsay, 2001)。
(2)木本層:小尺度方面有南仁山(廖啟政,1995)的研究。
(3)以科別區分:喜馬拉雅山區(Bhattarai and Vetaas, 2003)的蕨 類植物。
(二)β多樣性
β 多樣性沿海拔梯度亦有三種分布變化,以不同生活型論述(表 4):
1.隨海拔升高,物種轉換速率變慢,常發生在極端環境下的高山稀 疏草原中:
(1)所有物種:包括厄瓜多爾高山(Sklenár and Ramsay, 2001)。
(2)木本層:包括中國祈連山北坡(Wang et al., 2002)、中國長白山 北坡(趙淑清等,2004)及中國秦嶺太白山(唐志堯等,2004);
在小尺度方面有加拉巴哥群島的 Isla Santa Cruz(Itow,
表4 國內外文獻 β 多樣性(物種轉換)沿海拔梯度之變化
Table 4 Literature review of βdiversity (species turnover) along the elevation
β 多樣性 國內外文獻
海拔落差
(海拔範圍) 相當氣候帶
海拔升高,物 種轉換速率變 慢
沿海拔梯度, 物種轉換速率 無規律變化
海拔上升,物 種轉換速率變 快
Su (1994) >1000 m
(1370-3500 m)
亞熱帶-冷溫帶 木本層
Wang et al.
(2002)
>1000 m (1450-3700 m)
亞熱帶-亞寒帶 木本層
Kessler (2000) >1000 m (400-2500 m)
熱帶-溫帶 所有分類群
Arturo and Lauro (2005)
>1000 m (2800-4000 m)
涼溫帶-亞寒帶 木本層在低、
高海拔比中海 拔高
Sklenár and Ramsay (2001)
>1000 m (3100-4800 m)
冷溫帶-亞寒帶 所有物種
胡玉昆等 (2007)
>1000 m (2340-4618 m)
溫帶-亞寒帶 所有物種呈單
峰分布 鄭成洋等
(2004)
>1000 m (450-2158 m)
熱帶-溫帶 無明顯趨勢
唐志堯等 (2004)
>1000 m (1200-3200 m)
亞熱帶-冷溫帶 木本層
唐志堯、柯金虎 (2004)
>1000m (1500-2800 m)
暖溫帶-涼溫帶 木本層在低、
高海拔比中海 拔高
趙淑清等 (2004)
>1000 m (700-2600 m)
亞熱帶-涼溫帶 木本層 灌木層 草本層 Itow (1992) <1000 m
(0-860 m)
熱帶-亞熱帶 木本層
Itow (1991) <1000 m (140-560 m)
熱帶 木本層
1992),以及日本 Tatera 森林保留區(Itow, 1991)。
(3)灌木層:有中國長白山北坡(趙淑清等,2004)的研究。
(4)草本層:有中國長白山北坡(趙淑清等,2004)的研究。
2.沿海拔梯度無規律變化:
(1)所有物種:中國天山的結果為單峰分布(胡玉昆等,2007)。
(2)木本層:中國秦嶺牛背梁的結果為海拔兩端高,中間低的 分布情形(唐志堯、柯金虎,2004);中國福建黃崗山的分布 無明顯趨勢(鄭成洋等,2004)。
(3)以科別區分:玻利維亞安地斯山脈的所有分類群(鳳梨科、仙 人掌科、蕨類植物、爵床科、天南星科、野牡丹科)並無明顯 規律(Kessler, 2000)。
3.隨著海拔上升,物種轉換速率變快。
(1)木本層:台灣沙里仙溪的高海拔針葉樹林的β多樣性比中海 拔的闊葉樹林高(Su, 1994);以及墨西哥內華達山脈(Arturo and Lauro, 2005)的結果亦屬此型分布。
(三)物種豐量度
1.幾何級數分布:亞利桑那州 Santa Catalina 山區的高海拔冷杉、松 林(Whittaker and Niering, 1975),以及北大武山高海拔的木本和草 本(廖家宏,2006)皆為此分布。
2.對數級數分布:海南島吊羅山的雨林 (王崢峰等,1999)、北大武 山中低海拔的木本和草本(廖家宏,2006),以及中國四川臥龍研 究指出優勢種明顯較多、普遍種和稀有種發展受約束的植物社會 (石培禮等,2000)屬於此型分布。
3.對數常態分布:亞利桑那州 Santa Catalina 山區的中海拔橡樹林 (Whittaker and Niering, 1975)、北大武山中低海拔的木本和草本
(廖家宏,2006),以及中國四川臥龍研究普遍種較多,優勢種和 稀有種較少的植物社會(石培禮等,2000)屬於此型分布。
4.分割線段模式:中國四川臥龍研究指出所有物種豐量度都相似的 植物社會呈此型分布(石培禮等,2000)。
參、研究區域概述
一、地理位置
和平北溪流域位於宜蘭縣南澳鄉(圖 2),發源於南湖大山北峰(海拔 3592 m),最北為翠峰湖的南邊,與南澳南溪為界;最南為石楠花山與和平 南溪為界,於花蓮縣秀林鄉界與和平南溪匯流成和平溪,因為豐沛的溪水 從南湖大山北峰的崩壁沖蝕下大量泥沙,終年混濁,因此又稱大濁水溪,
河流總長度48.2 km,流域面積 561.06 km2,於和平村出海。重要支流有莫 很溪、布蕭丸溪、拉巴丸溪及次考干溪;重要湖泊有加羅湖群、翠峰湖以 及神祕湖。和平北溪的動、植物資源豐富,因此也是泰雅族人昔日活躍的 地點,著名的比亞毫古道即貫穿和平北溪流域。
圖2 和平北溪流域圖
Figure 2 Range of the Heping north river
二、地質與氣候
和平北溪流域屬於台灣最古老的岩石,由西至東分別屬於始新世畢祿 山層中之蘇澳南的南蘇澳層,主要組成岩層為板岩、千枚岩和變質砂岩,
以及古生代晚期至中生代的大南澳片岩,大南澳片岩又分為片麻岩、混合 岩、黑色片岩、綠色片岩燧岩以及大理岩(何春蓀,2003)。
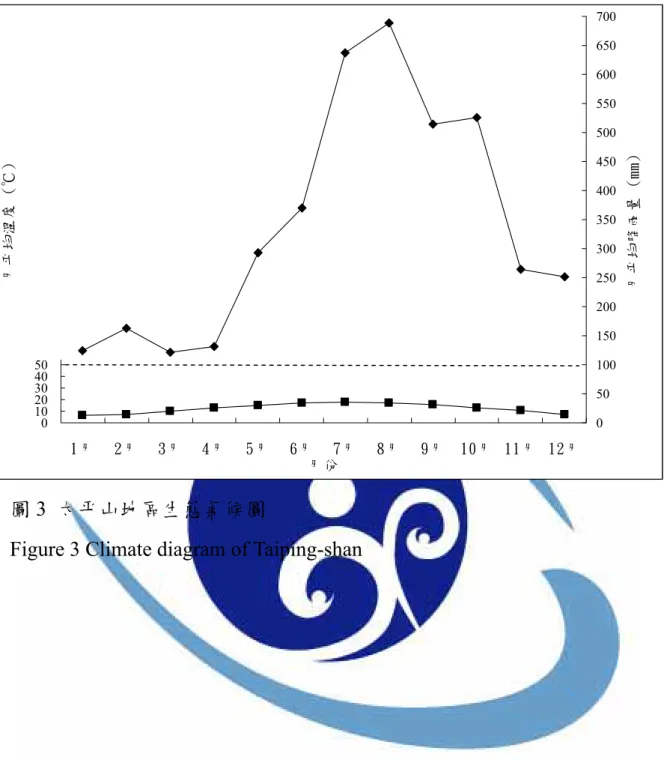
Su (1985)將台灣依雨量及溫度劃分為 11 個地理氣候區,本研究區屬於 東部地理氣候區北段,與恆濕型氣候的東北地理氣候區相鄰,因此為恆濕 型與夏雨型氣候之過渡帶。本研究區範圍由低海拔至高海拔,氣候差異甚 大,因此分別以太平山氣象站(表 5)及大濁水雨量站(表 6)代表中、低海拔之 氣候。太平山年平均降雨量約4402 mm,多集中在 7-10 月,年平均溫度為 12.5 ℃,最高溫在 6-8 月;最低溫為 1 月,偶有降雪,屬於恆濕型氣候(圖 3)。大濁水雨量站位於澳花村和平溪出海口,平均降雨量約 2577 mm,多集 中在5-11 月。
表5 太平山氣象站之月平均氣溫與月平均雨量氣候表(1996 年 1 月-2006 年 11 月)
Table 5 Climate of Taiping-shan weather station(January/1996- November/2006) month 1 2 3 4 5 6 7 8 9 10 11 12 temperature (℃) 6 7 10 13 15 17 18 17 15 13 11 7 mean rainfall (mm) 125 164 121 131 291 370 637 688 514 526 264 252
表6 大濁水雨量站月平均降雨量表(1982-2005 年)
Table 6 Mean rainfall of the Dajuoshuei rainfall station(1982-2005)
month 1 2 3 4 5 6 7 8 9 10 11 12 mean rainfall (mm) 64 94 72 90 203 218 258 284 500 439 239 111
100 2030 4050 6070 8090 100110 120130 140150 160170 180190 200210 220230 240250 260270 280290 300310 320330 340350
1月 2月 3月 4月 5月 6月 7月 8月 9月 10月 11月 12月
月份
月平均溫度 (℃)
0 50 100 150 200 250 300 350 400 450 500 550 600 650 700
月平均降雨量 (mm)
圖3 太平山地區生態氣候圖
Figure 3 Climate diagram of Taiping-shan
肆、研究方法
一、植物社會之調查及資料收集
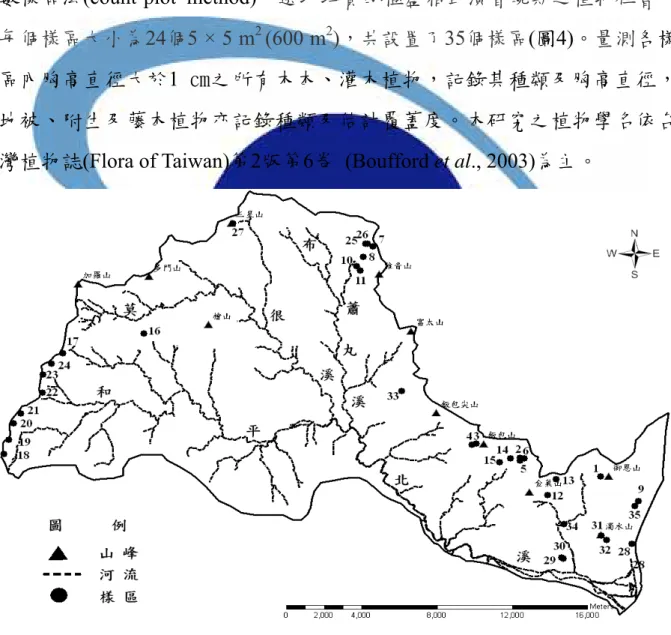
本研究於2006年3月開始蒐集相關資料,包括二萬五千分之一比例尺的 等高線圖,俾開始進行踏勘及調查的工作。為了使海拔之外的環境因子影 響降到最小,樣區取樣只選擇和平北溪北坡在上坡及稜線部分之植物社 會,從海拔100 - 3400 m,平均海拔每上升100 m設置至少一個樣區,採用計 數樣區法(count-plot method),選取均質的極盛相或演替晚期之植物社會。
每個樣區大小為24個5 × 5 m2 (600 m2),共設置了35個樣區(圖4)。量測各樣 區內胸高直徑大於1 ㎝之所有木本、灌木植物,記錄其種類及胸高直徑,
地被、附生及藤本植物亦記錄種類及估計覆蓋度。本研究之植物學名依台 灣植物誌(Flora of Taiwan)第2版第6卷 (Boufford et al., 2003)為主。
圖4 和平北溪北坡之樣區位置圖
Figure 4 Location of the study plots in the Heping north river
植群生態學是瞭解植物分布與環境因子的相關性,因為環境因子對植 群的分佈與結構具有相當的影響,而不同環境層次之因子,將導致不同幅 度的植群變異。因此研究植群生態除了對植物社會加以瞭解外,亦需對生 育地環境因子加以測量,藉由植群分析結果,以瞭解不同植物社會生育地 的生態幅度,及作為各種林型的辦識與分類。因此擬測量或間接評估研究 區域可能影響植物分布之環境因子,分述如下:
(一)海拔高度(altitude)
海拔高度直接影響了溫度梯度,為一高層次的環境因子,可作為 局部地點之氣候評估。本研究以全球衛星定位系統(global positioning system, GPS)配合地圖量測海拔高度值。
(二)坡度(slope)
坡度為生育地的傾斜度,影響土壤含水量及排水,並控制太陽的 入射角度,影響生育地太陽輻射的強度及局部的氣候變化。以傾斜儀 直接量測樣區之坡度數次,取其平均值表示之。
(三)坡向(aspect)
方位係指樣區或生育地最大坡度所面臨之方向(蘇鴻傑,1987a),
不同方位會導致溫度、日照、濕度及土壤水分之差異。以指北針測量 樣區最大坡度所處方向之方位角,並轉為 16 級,以 1-16 表示最乾到最 濕、用以表示相對水分指標值(蘇鴻傑,1987a)。
(四)全天光空域(whole light sky space, WLS)
樣區所在位置上方之空域,經扣除受周圍地形地物遮蔽的部份,
所得之天空比例稱之。量測方式先將 256 點平均分配於圓面積,並實 地量測出樣區周圍 8 個方位遮蔽物各高低轉折點之方位角及高度角 後,再將這些數值分別標於圓面積上,經連結各轉折點,以未遮蔽區 之點數為分子,圓面積上之 256 點數為分母,所得之百分率即為樣區
的全天光空域值。
(五)直射光空域(direct light sky space, DLS)
一年之中,太陽在樣區上空運行軌跡線所夾之天空面積,扣除周 圍地形遮蔽部份,所得之比例稱之。此二軌跡線即為夏至及冬至兩天 之太陽軌跡。此二線中未受遮蔽之點數為分之,全部點數 100 為分母,
所得之百分率為樣區之直射光空域值。
(六)土壤含石率(stoniness of soil)
估測樣區土壤中小石礫(<25 cm)之含量,以百分比表示之。
(七)岩石地比例(proportion of rock)
估測樣區中大石礫(>25 cm)所占之比例,以百分比表示之。
(八)地表裸露度(uncovered ratio of herbs layer)
樣區中地表無植被覆蓋的比例,以百分比表示之。
(九)土壤化學性質分析
1.土壤反應(pH 值)(McLean, 1982):
土壤 pH 值常隨著雨量增加而降低,亦影響土壤礦物質及有 機物之化學性質,而各種礦物質養分之溶解度與 pH 值有關,而 植物對養分之吸收效率,亦受其影響。土壤與蒸餾水依 1:1 的比 例充分攪拌後,靜置一小時後以玻璃電極測定。
2.土壤有機碳(organic carbon)(Nelson and Sommer, 1982):
秤取 0.5 g 風乾土、放在 500 mL 錐形瓶中。加入 10 mL 1N K2Cr2O7,搖勻,再迅速加入20 mL 濃硫酸,搖勻後靜置 30 分鐘,
再加入 200 mL 蒸餾水和 10 mL 85%之磷酸,放冷,加入 30 滴二 苯胺指示劑後,在電動攪拌器上攪動,以 Fe2+滴定之。其顏色變 化:暗褐色→濁藍色→鮮明藍色→綠色(滴定終點)。
3.有效性磷(available phosphorus)(Nelson and Sommer, 1982):
取 1 g 風乾土於試管中,加入 10 mL 萃取液(0.03N NH4F + 0.025N HCl),震盪 40 秒後過濾。取 5 mL 濾液、加入 5 滴鉬酸銨 ((NH4)6Mo7O244H2O)溶液及 5 滴還原劑,充分搖晃後靜置 30 分 鐘,用光電比色計在650 nm 波長下測其透光度。
4.有效性鉀(available potassium)(Helmke and Sparks, 1996):
取 5 g 風乾土,加入 20 mL 抽出液(0.05N HCl + 0.025N H2SO4),搖盪 5 分鐘後過濾,以原子吸光儀(AAS)測定鉀含量。
5.可交換性鹽基(exchangeable bases)(Thomas, 1982):
秤取 10 g 風乾土,加入 100 mL 1M 醋酸銨(pH 7.0)搖盪 1 小 時淋洗,所交換出來的陽離子用原子吸光儀(AAS)測定可交換性 鉀、鈉、鈣以及鎂等含量,並以cmol(+)/kg soil 表示之。
三 、 資 料 分 析 (一 )資 料 統 計
植物社會分類之原始資料分為木本(DBH ≧ 1)及草本層,將其介 量轉化為重要值指數(important value index,IVI),木本層的重要值 指數即為相對密度(relative density)、相對頻度(relative frequency)、相 對優勢度(relative dominance)的總合,此重要值指數最高為300 %;草 本層的重要值指數則為相對覆蓋度及相對頻度的總合,為200 %,並 全換算為以100 %為基礎的重要值指數。將各樣區重要值指數及環境 因子資料,利用Excel軟體整理成原始矩陣資料,以供電腦軟體分析使 用。
(二 )植 群 多 樣 性 分 析
植群多樣性的意義不止在清查植物社會的數目,與物種多樣性及 分化多樣性間更有密不可分的關係。本研究將物種多樣性的α多樣性 尺度提升至γ多樣性時,則必須先進行植群分析,以確切且具體劃分
植 群 林 帶 。 植 群 生 態 多 變 數 分 析 方 法 依 研 究 方 法 可 分 為 分 類 法 (classification)及分布序列法(ordination)。所謂分類法即將組成植物相 似的樣區合併成植群型(vegetation type),並與環境因子共同分析,尋 求可能影響植群分化的環境因子。而分布序列法是將樣區或植物依相 關性排列在具有影響力的梯度軸上,並加入環境因子,以尋求植群變 異與環境梯度的相關性。而植物社會之分類及單位間之差異,也表現 於物種之轉換,故β多樣性也可以用到植群多樣性,植群多樣性的分 類法旨在定義植群型之範圍及數目,但以植群型之數目來表示多樣性 會因為人為的技術性而有所差異,因此以環境及組成最大變異梯度上 之物種轉換比例(即β多樣性),例如分布序列法中的降趨對應分析法 (detrended crrespondence analysis, DCA)來度量植群多樣性,植群分析 以分類法及分布序列法相輔相成,可達成區分植群型與顯示變異程度 之雙重標準。
植物社會分類法採用雙向指標種分析(two-way indicator species analysis, TWINSPAN),依指標種在樣區中出現的忠誠度將樣區劃分植 群型(Hill, 1979)。
在分布序列方面,以 PC-ORD 4.0 程式中的典型對應分析法 (canonical correspondence analysis, CCA)分析,CCA 是間接梯度分析 的一種,結合了物種資料與環境因子同時進行迴歸及相關測試,並計 算各梯度軸能夠解釋物種與環境的變異量,可說明樣區與環境變數的 關係,選擇具有顯著相關的環境因子,進行樣區與環境因子間相關性 之探討。
再以降趨對應分析法(detrended crrespondence analysis, DCA)進行 分析,是直接梯度分布的一種,即將植物或樣區排列在已知影響的環 境梯度上(蘇鴻傑,1987b,1996)。其軸長為物種的標準偏差單位
(standard deviation, SD),若物種組成的轉變較快,則二端的樣點共有 的物種種類較少,因此軸長又可視為β多樣性單位,軸長度數值較 高,代表β多樣性高,若差距在 4 SD 以上,代表兩樣點之間沒有共 同的種類(Jr. Gauch, 1982; 劉和義,2004)。
(三)物種多樣性分析
1.清查多樣性
一般以直接量測樣區或區域內物種種數之多寡為主,為許多 方法中最簡單且易於使用之指數。在比較時要考慮植物社會單位 之大小,一般依營養級、生態地位或生活型(life form)分開統 計(劉棠瑞、蘇鴻傑,1983)。本研究參考方精云等(2004)、魏瑞 廷(2006)、Whittaker and Niering(1975)、Patricia et al.(2002)、Wang
et al.(2002)、Bhattarai and Vetaas(2003)、Arturo and Lauro (2005)
及Hemp(2006)之研究,依林分垂直結構將每個樣區劃分為喬木層 (DBH ≧ 5 cm)、灌木層(DBH < 5 cm)、草本層、附生植物層以及 藤本植物層五種生活型,分別討論其物種多樣性沿海拔之變化。本研究橫跨了數個植群帶,因此可將植群帶視為γ多樣性的區域 單位,以進行物種多樣性探討。
(1)物種豐富度指數
此指數即為測定範圍內的物種數量,是為最簡易多樣性指 數,然樣區大小會影響物種的豐富度。本研究採用600 m2樣區 面積內之物種數表示之。
(2)Shannon-Wiener 指數(H)
此指數為假設在植物社會中隨機抽取一個體,該個體屬於 那個物種是隨機的,且物種數愈多,其不定性也愈大。此指數 受種數與個體數影響,即種數愈多,其多樣性愈高。因可能率
之指數為負值,故前面再加負號,以轉為正值。
H = –∑(ni/N)ln(ni/N)= –∑PilnPi
ni 為第 i 種之介量;N 為所有物種介量之總和。
(3)均勻度指數(EH)
為植物社會中物種個體在不同種間分布的均勻程度。多樣 性高的植物社會,也可能出現每一物種具有相同豐富度的情 形,因此,均勻度指數以 Shannon 指數為主軸,將其值限於 1 與 0 之間。
EH=H/lnS
H 為 Shannon 多樣性指數;S 為物種數。
(4)物種豐量度曲線(rank/abundance plot)
不同的研究者對於物種豐量度會使用不同的方法,其中較 通用且較有效用的方法是等級-豐量度圖,即物種豐量度曲線 (Magurran, 2004)。本研究物種依豐量度從多到最少連續排列於 水平軸(x 軸),豐量度則以株數的對數(log)表示(y 軸),如此,
易於在同一曲線圖呈現跨度數個等級的物種豐量度。並依不同 植群型、植群帶分別探討其差異。等級-豐量度圖的優點是能將 物種豐量度清楚顯明的表示出來,甚至種量較少的物種亦能清 楚的表達其豐量度,此外,還能強調社會中均勻度的差異。
2.分化多樣性 (1)β 多樣性
β 多樣性為沿著某一環境梯度物種轉的程度或速率。β 多 樣性還反應了不同植群間物種組成的差異,不同植群或某環境 梯度上不同點之間的共有種愈少,β 多樣性愈大。
a. Wilson 與 Shmida 指數(βT)
此指數具有物種在梯度上的獲得與消失之物種轉換的 概念,即量測物種在環境梯度上兩相鄰樣區間的物種轉換程 度(Magurran, 1988)。
βT =[ g(H)+l(H) ] /2a
g(H)爲沿環境梯度所增加的物種數目;l(H)為沿環境 梯度所失去的物種數目;a 為相鄰兩樣區之平均種數。
伍、研究流程
綜合分析、討論
與國內外研究結果比較及應證 確定研究動機、目的與範圍
(探討和平北溪物種多樣性沿海拔梯度之變化)
蒐集前人研究文獻
蒐集研究區域環境相關資料 研究地區勘察及路線設定 文獻整理回顧(多樣性沿
海拔變化之理論及國內 外研究趨勢)
樣區選擇與取樣方法
野外調查:植物介量資料蒐集 環境因子估測 土壤取樣
製作原始矩陣表 及環境因子矩陣
植群多樣性及與環境 間之相關性
(TWINSPAN、DCA、CCA)
清查多樣性 分化多樣性 物種豐量度 植物社會結構之變化
初步結果
結 論 土壤化學性質分析
陸、結果
一、物種組成
本研究共調查 35 個樣區,記錄蕨類植物 128 種、裸子植物 8 種、雙子 葉植物365 種、單子葉植物 81 種,共計維管束植物種類 129 科 331 屬 582 種(表 7)。種類豐富度最高的前 5 科為蘭科(Orchidaceae)32 種、薔薇科 (Rosaceae)26 種、鱗毛蕨科(Dryopteridaceae)24 種、水龍骨科(Polypodiaceae) 22 種及茜草科(Rubiaceae)17 種。
表7 和平北溪維管束植物調查結果
Table 7 Vascular plants recorded in the Heping north river, with numbers of families, genera and species
分類群 科 屬 種
蕨類植物 27 56 128
裸子植物 3 7 8
雙子葉植物 87 218 365 單子葉植物 12 50 81
合計 129 331 582
二、植群多樣性
本研究區域為和平北溪北坡之上坡、稜線地形的植物社會,以雙向指 標種分析(TWINSPAN)依特徵種在樣區中出現的忠誠度將植物社會分群,並 參考特徵種在野外的分布,略加調整並整理為概要表,共分為 6 個植群型 10 個植物社會(表 8)。各植群型命名原則以特徵種在前,優勢種在後的方式 命名(劉棠瑞、蘇鴻傑,1983)。將 10 個植物社會由低至高海拔分述如下。
Ι. 榕楠林帶(FicusMachilus zone):林帶之海拔分布 0-630 m。
A. 茄 冬 大 葉 楠 型 (Bischofia javanica Machilus japonica var.
kusanoi type):
表8 和平北溪植群概要表
Table 8 Synoptic table of the clusters based on TWINSPAN classification of 35 plots of the Heping river basin
植群帶 Ⅰ Ⅱ Ⅲ Ⅳ Ⅴ Ⅵ
植群型 A B C D E F G H I J
2 2 3 3 3 3 3 3 1 1 1 1 1 1 1 1 2 2 2 2 2 1 2 2 2 1 樣區
8 9 0 4 5 9 2 1 3 1 5 4 6 3 6 4 0 2 5 3 2 8 1 7 7 5 6 3 4 7 9 0 1 2 8
海拔 170 598 630 220 522 408 917 812 1100 1227 1309 1410 1550 1450 1738 1640 1500 1462 1520 1755 1690 1715 1617 1800 2306 1837 1878 2507 2522 2355 3119 2944 2857 2733 3372
榕樹 7 6
茄冬 5 3 4 4
澀葉榕 8 6 7
薄葉嘉賜木 2 3 2 1
粗糠柴 3 1 3 2
瓊楠 4 1 4 1
五掌楠 5 7 3 2 1
大葉楠 4 6 7 5 2 5 4 4
台灣糊樗 2 2 4 2 4
茜草樹 3 3 1
水金京 1 2 5 5
披針葉饅頭
果 3 1 3 1
細葉饅頭果 2 3 1
頷垂豆 4 4 2
白匏子 4 5 6 4
台灣山桂花 3 1 3 2 2
山紅柿 2 4 3 3 1
小葉樹杞 3 5 1 1
黃杞 4 4 4 4 4 5 6 4 4 1
狗骨仔 3 4 5 2 4
短尾葉石櫟 3 1 1 1 3 2 1
柏拉木 3 4 5 5 1
奧氏虎皮楠 5 5 1 1 1 1 3 1 2 1 3
杜英 1 1 4 4 3 2 1 3 1 4 1
小花鼠刺 1 2 4 1 3 1 1 2 1 2
烏皮茶 4 2 5 5
大葉石櫟 1 1 3 3 4 4 4 5 2
油葉石櫟 2 6 5 4 1
月桂葉灰木 5 3 3 4 2 3
薯豆 1 3 3 5 5 3 2 6 6 3
厚皮香 2 1 1 1 1 4 4 1 1 1
太平紅淡比 1 3 1 4 5 6 1
變葉新木薑
子 2 2 1 4 4 4 1 1
小葉赤楠 1 2 5 5
南嶺灰木 1 2 3
台灣杜鵑 8 8 7
台灣扁柏 7 7 1 1
石斑木 3 2