國立臺灣大學生物資源暨農學院農業化學研究所 碩士論文
Graduate Institute of Agricultural Chemistry
College of Bioresources and Agriculture National Taiwan University
Master Thesis
大腸桿菌熱休克蛋白酶 ClpYQ 之基質 SulA 被辨識區域特性之研究
The recognition region in SulA by ClpYQ protease from Escherichia coli
翁于婷 Yu-Ting Weng
指導教授:吳蕙芬 博士 Advisor: Whei-Fen Wu, Ph.D.
中華民國九十八年六月
June, 2009
口試委員會 口試委員會 口試委員會
口試委員會審定書 審定書 審定書 審定書
ii
誌謝 誌謝 誌謝 誌謝
來到這個實驗室,前後斷斷續續地過了四個年頭,從剛開始的迷惘生疏,到 今天能夠順利完成論文,真的要感謝許許多多師長及朋友們的幫助與陪伴,使我 在這段時間裡,可以不畏艱難充滿勇氣的向前邁進。
首先要感謝帶領我進入研究領域的指導老師 吳蕙芬教授,老師對我的悉心指 導及訓練,使我學習到身為一個研究者應具有的基本能力及素養,而老師於研究 上專注嚴謹的實驗態度與邏輯思考,更是我學習效法的對象,衷心的謝謝老師這 段時間來的指導與照顧。
感謝 楊啟伸教授、蔡孟勳教授、蘇南維教授、徐駿森教授擔任我的口試委員,
撥空審閱我的論文初稿,細心的逐字釜正,並提出許多精闢的見解及寶貴的意見,
提醒我用不同的思考角度看待結果,使論文內容可以更趨完善,也謝謝老師們對 我的鼓勵,讓我有信心面對更多的挑戰。
此外,感謝實驗室裡與我共度這段時光的夥伴們,親愛的大美女湘芸學姐、
美食達人汎擎學長、細心可靠的鈞暘學長、常讓我搭便車的 Asonn、帥氣又搞笑的 宣哈、總有獨道見解的岳隆、活潑充滿朝氣的郁君、可愛貼心的玉鳳,已經畢業 的學長姐們及各自向外發展的學弟們;還有對面常讓我去串門子的朋友們及樓上 的小祕書,因為有你們使實驗室的生活增加更多的樂趣與歡笑。
最後要感謝親愛的爸媽,因為你們的支持,並給我無限的自由和絕對的信任,
讓我能走自己的路。還有最愛的慶,謝謝你陪我走過每一個歡樂與傷心的時刻,
有你的陪伴,讓我不再感到寂寞。
摘要 摘要 摘要 摘要
大腸桿菌中 ClpYQ 蛋白酶為一種 ATP 依賴蛋白酶,由具有 ATPase 及 unfoldase 活性的 ClpY,與具有 peptidase 活性的 ClpQ 所構成的雙單元體。這類蛋白酶在細 胞中,可降解構形錯誤或是具危害性的蛋白質,以維持細胞正常生理作用,避免 細胞受到危害。在 ClpYQ 蛋白酶中,ClpY 會負責辨識基質,並水解 ATP 作為能 量來源,將基質結構打開並傳送至 ClpQ 的活性區,以進行降解作用。然而,關於 ClpYQ 蛋白酶是如何選擇辨識基質,及後續降解作用的詳細機制,目前仍不清楚。
SulA 是一個細胞分裂的抑制物,當細胞暴露在逆境下時會產生 SOS 反應,誘導大 量 SulA 蛋白表現,以避免受損的 DNA 傳到子代細胞。目前已知可分解 SulA 的蛋 白酶為 Lon 及 ClpYQ,其中又以 Lon 為主要負責分解的蛋白酶。之前有研究指出,
Lon 可以藉由辨認 SulA 之 C-端末 8 個胺基酸,與之結合並將其降解,但是 ClpYQ 卻不能。對於 ATP 依賴蛋白酶來說,為避免不必要的降解,如何選擇辨認需要降 解的基質是非常重要的。本實驗中,為確認 ClpYQ 蛋白酶辨認 SulA 蛋白的區域,
建構 SulA 之 C-端不同大小片段缺失的突變蛋白,以酵母菌雙雜交系統測試,各個 SulA 突變蛋白與 ClpY 之間交互作用的情形,發現 ClpY 辨識的區域可能位於 C- 端的高疏水性片段,C-端第 20 - 30 個胺基酸。於此區域內再建構點突變蛋白,分 析不同性質的胺基酸對於 ClpY 交互作用的影響,結果顯示當點突變取代為親水性 胺基酸時,會降低 SulA 蛋白與 ClpY 之間的交互作用。之後測試各個 SulA 突變蛋 白的活性表現,及被 ClpYQ 蛋白酶降解之情形,結果顯示 SulA 之 C-端第 20 - 45 個胺基酸的區域,對於其抑制細胞分裂的活性表現是重要的,且對於 ClpYQ 蛋白 酶的降解作用也會造成影響,因此 ClpYQ 蛋白酶應可藉由 SulA 蛋白之活性表現 與否,來辨別其是否需要降解。
關鍵詷:ATP 依賴蛋白酶、ClpYQ、Lon、SulA、蛋白質辨識
iv
Abstract
ClpYQ is an ATP-dependent protease from Escherichia coli and a two component
complex composed of ClpY, which is an ATPase and unfoldase, and ClpQ peptidase.
Degradation of denatured or damaged proteins by this proteases helps protect the
normal cell growth from the harmful effects of these proteins. The ClpY is thought to
recognize protein substrates, denature them, and translocate the unfolded polypeptide
into the catalytic cavity of the ClpQ for degradation. However, little information is
available on the recognition of substrates for ClpYQ and on the mechanism by which
they were selected, unfolded, and translocated by ClpY to the interior of the ClpQ. SulA,
induced in the SOS response, is a cell division inhibitor and prevents the distribution of
damaged DNA into daughter cells during DNA repair processes. SulA can be degraded
by ATP-dependent proteases such as Lon and ClpYQ, and the degradation in vivo seems
to be predominantly by Lon, while ClpYQ appears to act as a backup for Lon. It was
reported previously that the region of C-terminal eight amino acid residues of SulA was
essential for interaction with Lon but not with ClpYQ. To avoid unnecessary
degradation of cellular proteins, substrate selection by ATP-dependent proteases is
tightly regulated; therefore, it is interesting to investigate the recognition region of SulA
by ClpYQ. In this study, the deletion mutants of SulA with regard to C-terminus were
constructed and the interaction with ClpY was analyzed in yeast two-hybrid system. The
results showed that ClpY recognized the hydrophobic region of SulA, C20 - 30 aa, and
the recognition was also likely to rely on hydrophobic interaction following the
observation that the binding activity decreased if the substituted residue was polar. The
C-terminal region of SulA, C20 - 45 aa, seemed to be important for its activity with an
inhibition of cell division, and the region is necessary for the degradation by ClpYQ.
Therefore, ClpYQ protease would be able to distinguish whether SulA is to be degraded
by the activity of inhibition.
Keywords: ATP-dependent protease, ClpYQ, Lon, SulA, Protein recognition
vi
目錄 目錄 目錄 目錄
口試委員會審定書 口試委員會審定書 口試委員會審定書
口試委員會審定書 ... i
誌謝誌謝 誌謝誌謝 ... ii
摘要 摘要 摘要 摘要 ... iii
Abstract ... iv
目錄目錄 目錄目錄 ... vi
表目錄 表目錄 表目錄 表目錄 ... viii
圖目錄圖目錄 圖目錄圖目錄 ... ix
附圖目錄 附圖目錄 附圖目錄 附圖目錄 ... x
壹 壹 壹 壹、 、 、 、前言 前言 前言 前言 ... 1
一 一 一 一、、、、ATP 依賴蛋白酶依賴蛋白酶依賴蛋白酶依賴蛋白酶 ... 1
二二 二二、、、、ClpYQ 蛋白酶蛋白酶蛋白酶 ... 2 蛋白酶 三 三 三 三、、、、ClpYQ 複合體結構複合體結構複合體結構 ... 4 複合體結構 四四 四四、、、、ClpYQ 蛋白酶之基質蛋白酶之基質蛋白酶之基質 ... 5 蛋白酶之基質 五 五 五 五、、、、細胞分裂抑制物細胞分裂抑制物細胞分裂抑制物細胞分裂抑制物 SulA ... 7
六 六 六 六、、、、蛋白酶對基質蛋白酶對基質蛋白酶對基質蛋白酶對基質 SulA 辨認之相關研究辨認之相關研究辨認之相關研究辨認之相關研究 ... 9
七七 七七、、、、研究動機與目的研究動機與目的研究動機與目的研究動機與目的 ... 12
貳 貳 貳 貳、 、 、 、材料與方法 材料與方法 材料與方法 材料與方法 ... 13
一 一 一 一、、、、實驗材料實驗材料實驗材料實驗材料 ... 13
(一一一一) 菌株與質體菌株與質體菌株與質體菌株與質體 ... 13
(二二二二) 藥品與試劑藥品與試劑藥品與試劑藥品與試劑 ... 14
(三三三三) 器材設備器材設備器材設備器材設備 ... 15
(四四四四) 分析軟體分析軟體分析軟體分析軟體 ... 15
二 二 二 二、、、、實驗方法實驗方法實驗方法實驗方法 ... 16
(一一一一) 一般實驗方法一般實驗方法一般實驗方法一般實驗方法 ... 16
(二二二二) 菌體的建構菌體的建構菌體的建構菌體的建構 ... 20
(三三三三) 突變基因建構突變基因建構突變基因建構突變基因建構 ... 25
(四四四四) 酵母菌雙雜交系統分析酵母菌雙雜交系統分析酵母菌雙雜交系統分析酵母菌雙雜交系統分析 ... 28
(五五五五) 大腸桿菌選殖基因表現系統大腸桿菌選殖基因表現系統大腸桿菌選殖基因表現系統大腸桿菌選殖基因表現系統 ... 33
(六六六六) 西方墨點分析西方墨點分析西方墨點分析西方墨點分析 (Western Blotting) ... 35
參 參
參 參、 、 、 、實驗結果 實驗結果 實驗結果 實驗結果 ... 41
一 一 一
一、、、、確認確認確認確認 SulA 蛋白被蛋白被蛋白被蛋白被 ClpY 辨識之區域辨識之區域辨識之區域辨識之區域 ... 41 (一一一一) 建構建構建構建構 SulA 蛋白蛋白蛋白蛋白 C-端不同大小片段缺失之突變端不同大小片段缺失之突變端不同大小片段缺失之突變 ... 41 端不同大小片段缺失之突變 (二二二二) 以酵母菌雙雜交系統測試各個以酵母菌雙雜交系統測試各個以酵母菌雙雜交系統測試各個以酵母菌雙雜交系統測試各個 SulA 突變蛋白與突變蛋白與突變蛋白與 ClpY 之交互作用突變蛋白與 之交互作用之交互作用 ... 41 之交互作用 二
二 二
二、、、、SulA 蛋白被辨識區域之特性蛋白被辨識區域之特性蛋白被辨識區域之特性 ... 43 蛋白被辨識區域之特性 (一一一一) SulA 蛋白蛋白蛋白蛋白 C-端序列特性分析端序列特性分析端序列特性分析 ... 43 端序列特性分析 (二二二二) 於於於於 SulA 蛋白被辨識區域中建構不同胺基酸性質之點突變蛋白被辨識區域中建構不同胺基酸性質之點突變蛋白被辨識區域中建構不同胺基酸性質之點突變蛋白被辨識區域中建構不同胺基酸性質之點突變 ... 43 (三三三三) 利用酵母菌雙雜交系統測試利用酵母菌雙雜交系統測試利用酵母菌雙雜交系統測試利用酵母菌雙雜交系統測試 SulA 點突變蛋白與點突變蛋白與點突變蛋白與點突變蛋白與 ClpY 之交互作用之交互作用之交互作用 ... 44 之交互作用 三
三 三
三、、、、不同蛋白酶對於不同蛋白酶對於不同蛋白酶對於不同蛋白酶對於 SulA 蛋白辨識區域之差異蛋白辨識區域之差異蛋白辨識區域之差異蛋白辨識區域之差異 ... 45 (一一一一) Lon 蛋白酶與蛋白酶與蛋白酶與蛋白酶與 SulA 缺失突變蛋白間之交互作用缺失突變蛋白間之交互作用缺失突變蛋白間之交互作用缺失突變蛋白間之交互作用 ... 45 (二二二二) ClpY I domain 上兩個上兩個上兩個上兩個 loop 對於對於對於對於 SulA 蛋白辨識之影響蛋白辨識之影響蛋白辨識之影響蛋白辨識之影響 ... 46 四
四 四
四、、、、SulA 突變蛋白之活性表現及受突變蛋白之活性表現及受突變蛋白之活性表現及受 ClpYQ 蛋白酶降突變蛋白之活性表現及受 蛋白酶降蛋白酶降解之現象蛋白酶降解之現象解之現象解之現象 ... 47 (一一一一) C-端端端端末末末末 20 個胺基酸缺失之個胺基酸缺失之個胺基酸缺失之 SulA 突變蛋白個胺基酸缺失之 突變蛋白突變蛋白 ... 48 突變蛋白 (二二二二) C-端二級結構部份缺失之端二級結構部份缺失之端二級結構部份缺失之端二級結構部份缺失之 SulA 突變蛋白突變蛋白突變蛋白 ... 50 突變蛋白 (三三三三) SulA 不同性質胺基酸之點突變蛋白不同性質胺基酸之點突變蛋白不同性質胺基酸之點突變蛋白不同性質胺基酸之點突變蛋白 ... 51
肆 肆 肆
肆、 、 、 、討論 討論 討論 討論 ... 53
一一
一一、、、、SulA 蛋白被蛋白被蛋白被 ClpY 辨識之區域及其特性蛋白被 辨識之區域及其特性辨識之區域及其特性... 53 辨識之區域及其特性 二
二 二
二、、、、不同蛋白酶對於辨認相同基質之差異不同蛋白酶對於辨認相同基質之差異不同蛋白酶對於辨認相同基質之差異不同蛋白酶對於辨認相同基質之差異 ... 54 三三
三三、、、、SulA 突變蛋白之活性表現與突變蛋白之活性表現與突變蛋白之活性表現與 FtsZ 的相關性突變蛋白之活性表現與 的相關性的相關性 ... 55 的相關性 四
四 四
四、、、、ClpYQ 蛋白酶對於蛋白酶對於蛋白酶對於 SulA 突變蛋白之降解情形蛋白酶對於 突變蛋白之降解情形突變蛋白之降解情形 ... 57 突變蛋白之降解情形 五
五 五
五、、、、質體質體質體質體 SulA 的表現會誘導染色體之的表現會誘導染色體之的表現會誘導染色體之的表現會誘導染色體之 SulA 表現表現表現表現 ... 58
伍 伍 伍
伍、 、 、 、結論 結論 結論 結論 ... 60 陸
陸 陸
陸、 、 、 、參考文獻 參考文獻 參考文獻 參考文獻 ... 61
viii
表目錄 表目錄 表目錄 表目錄
表一、本論文所使用的菌株與噬菌體 ... 67
表二、本論文所使用的質體 ... 68
表三、本論文所使用的引子對 ... 69
表四、在酵母菌雙雜交系統中 SulA 突變蛋白與 ClpY 之交互作用情形 ... 71
表五、SulA 突變蛋白之活性測試及受 ClpYQ 蛋白酶降解之情形 ... 72
圖目錄 圖目錄 圖目錄 圖目錄
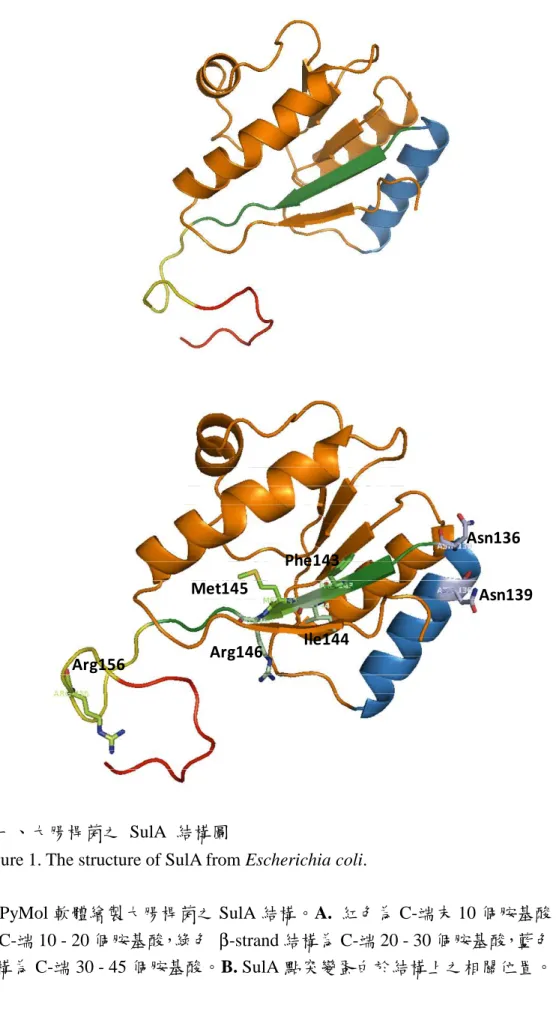
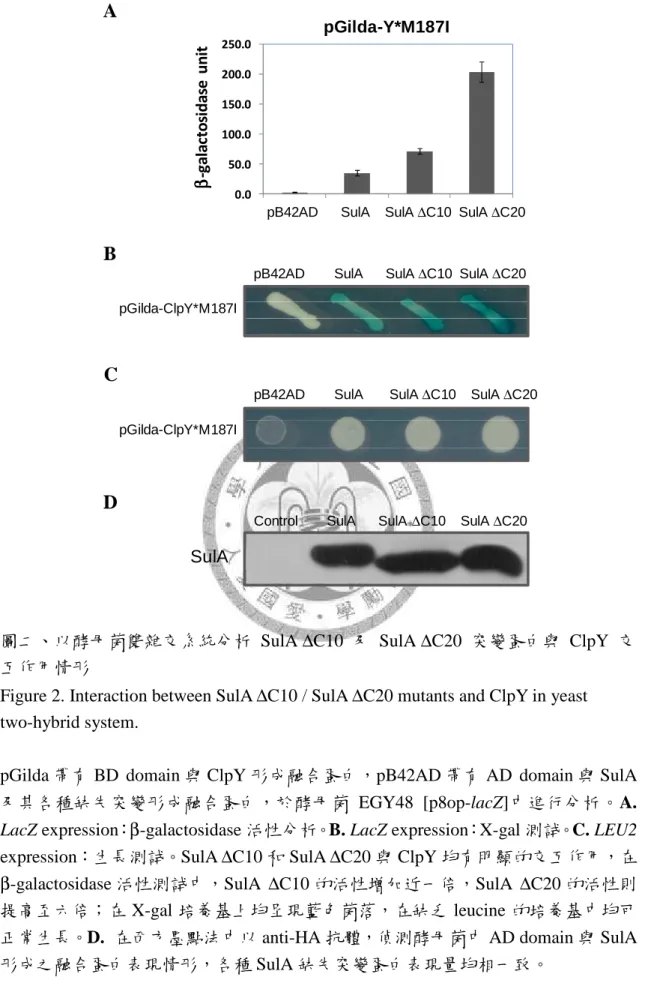
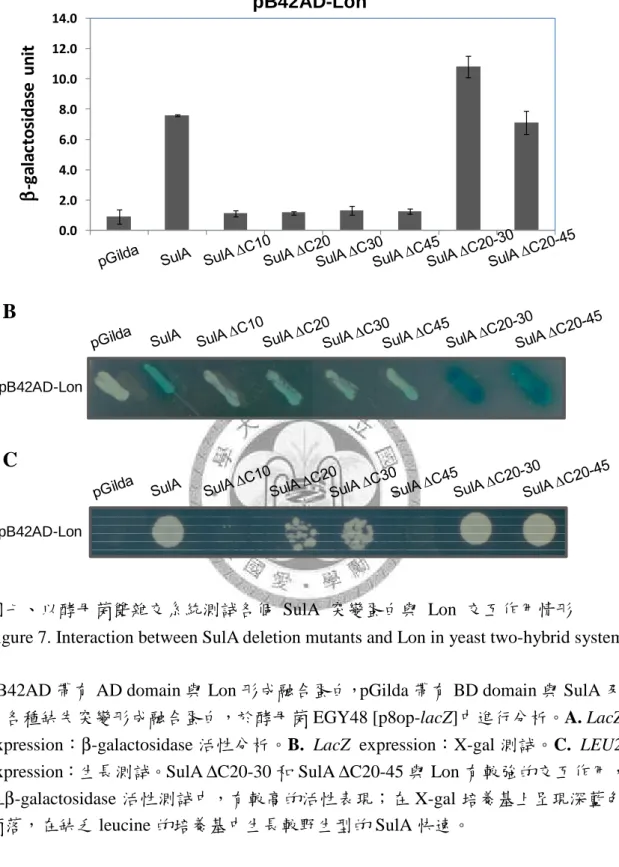
圖一、大腸桿菌之 SulA 結構圖 ... 73 圖二、以酵母菌雙雜交系統分析 SulA ∆C10 及 SulA ∆C20 突變蛋白與 ClpY 交互
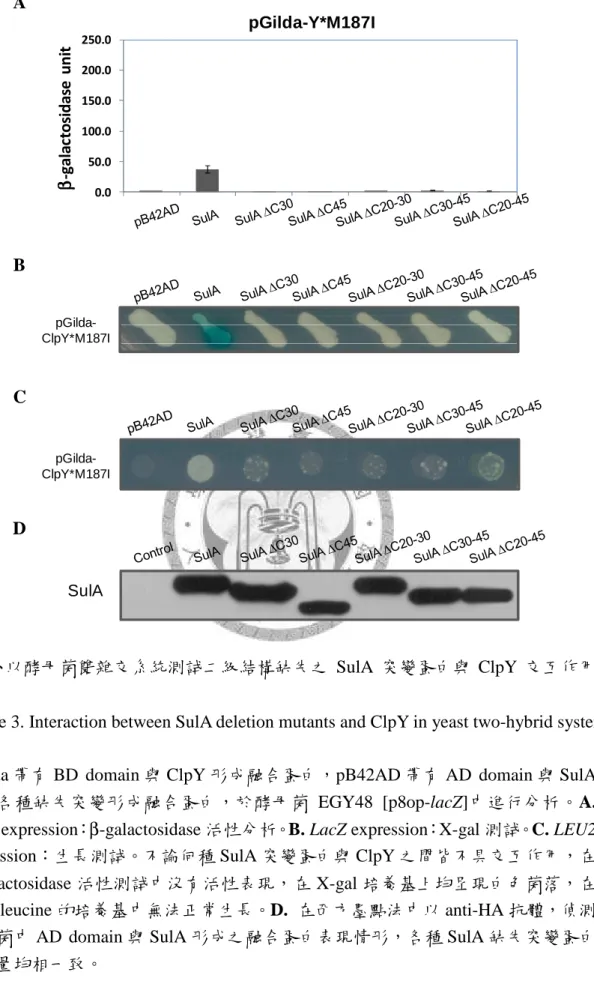
作用情形 ... 74 圖三、以酵母菌雙雜交系統測試二級結構缺失之 SulA 突變蛋白與 ClpY 交互作
用情形 ... 75 圖四、大腸桿菌 SulA 之 C 末端序列分析 ... 76 圖五、大腸桿菌與四種腸道菌微生物之 SulA 相似序列的比對 ... 77 圖六、以酵母菌雙雜交系統測試各個 SulA 點突變蛋白與 ClpY 交互作用情形 78 圖七、以酵母菌雙雜交系統測試各個 SulA 突變蛋白與 Lon 交互作用情形 ... 79 圖八、以酵母菌雙雜交系統測試各個 SulA 突變蛋白與 ClpY 之 loop 缺失突變蛋
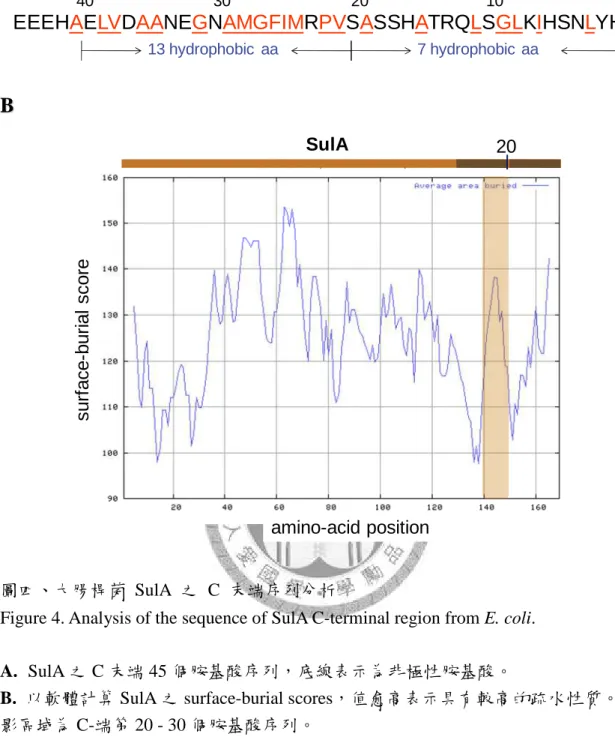
白交互作用情形 ... 80 圖九、AC3112 菌株及 sulA- 菌株 YT10010 之 MMS 生長測試 ... 81 圖十、SulA ∆C10 和 SulA ∆C20 突變蛋白活性表現及受 ClpYQ 蛋白酶降解之情形 .. 82 圖十一、二級結構缺失之 SulA 突變蛋白活性表現及受 ClpYQ 蛋白酶降解之情形 ... 83 圖十二、SulA 點突變蛋白之活性表現及受 ClpYQ 蛋白酶降解之情形... 84
x
附圖目錄 附圖目錄 附圖目錄 附圖目錄
附圖一、大腸桿菌 ClpYQ 結構圖 ... 85
附圖二、SOS 反應 ... 86
附圖三、大腸桿菌及綠膿桿菌之 SulA 胺基酸序列比對 ... 87
附圖四、綠膿桿菌之 SulA 結構圖 ... 88
附圖五、酵母菌雙雜交系統之報導基因表現示意圖 ... 89
壹 壹 壹
壹、 、 、前 、 前 前言 前 言 言 言
一 一 一
一、 、 、 、ATP 依賴蛋白酶 依賴蛋白酶 依賴蛋白酶 依賴蛋白酶
蛋白質品質控制系統 蛋白質品質控制系統 蛋白質品質控制系統
蛋白質品質控制系統 (Protein quality-control systems)
生物體正常的生長代謝過程中,均需要藉由蛋白質來進行各種生理作用,因 此產生正確且具有功能的蛋白質,是維持生命現象所必需的。因此在細胞中具有 一蛋白質品質控制系統 (Protein quality-control systems),負責維持蛋白質的正常活 性及功能,以避免細胞受到危害。此系統包含幾個重要的角色:ATP 依賴蛋白酶 (ATP-dependent proteases)、伴隨蛋白質 (chaperones)、熱休克蛋白質 (heat shock proteins),及一些調控分子等;這些系統網絡可幫助新生成蛋白質折疊成正確構 形、拆解 (disaggregation) 錯誤折疊蛋白質之聚合物 (aggregate),並幫助其重新折 疊恢復正確構形,或是降解 (degradation) 不能修復而具有危害性的蛋白質,以維 持細胞正常的生長活動 (Sauer et al., 2004, Bukau et al., 2006)。
ATP 依賴蛋白酶依賴蛋白酶依賴蛋白酶依賴蛋白酶 (ATP-dependent proteases)
ATP 依賴蛋白酶 (ATP-dependent proteases) 為一調控蛋白質品質的系統,這類 蛋白酶廣泛存在於真核細胞、細菌及古細菌中。當細菌的生長受到環境壓力時,
如:溫度驟變,易導致菌體內不正常蛋白質的生成累積而造成危害,此時 ATP 依 賴蛋白酶可藉由水解 ATP 作為能量來源,將不必要的或具有危害性的蛋白質降 解,幫助細菌因應外在環境的變化,於不利生長的環境中存活 (Wickner et al., 1999, Zwickl et al., 2000)。
在原核生物中主要的 ATP 依賴蛋白酶有 Lon、ClpAP、ClpXP、ClpYQ (HslUV) 及 FtsH。Lon 和 FtsH 均是 single-chain proteases (Gottesman, 2003),Lon 位於細胞 質內,為最主要的蛋白酶,FtsH 附著於細胞膜上,主要負責膜蛋白質的降解;而 其他 ClpAP、ClpXP 及 ClpYQ 則是位於細胞質內,由兩種蛋白質組成的雙單元體
2
(two component system) 蛋白酶 (Katayama-Fujimura et al., 1987, Gottesman et al., 1998, Chuang et al., 1993)。
此類蛋白酶通常會形成桶狀的聚合體,其中央具有一狹窄孔洞,由 X-ray 結晶 結構的分析發現,孔洞直徑大小約略為 10 Å 左右,這樣的寬度不足以讓正常構形 的蛋白質進入 (Bochtler et al., 2000, Guo et al., 2002);然而,蛋白酶的活性區卻是 位於桶狀結構之中,因此必須設法將基質傳遞至中央,才能進行蛋白質的水解作 用 (Bochtler et al., 1997, Wang et al., 1997)。直至目前為止眾多的研究顯示,認為這 類蛋白酶會先辨認基質蛋白,並將其結構打開,才能穿越中央孔洞傳送至活性區 進行降解 (Sauer et al., 2004, Zhang et al., 2002, Matouschek, 2003)。
二 二 二
二、 、 、 、ClpYQ 蛋白酶 蛋白酶 蛋白酶 蛋白酶
熱休克蛋白 熱休克蛋白 熱休克蛋白
熱休克蛋白 (Heat shock proteins)
ClpYQ 蛋白酶屬於熱休克蛋白的一種,最早於 1993 年被提出,在大腸桿菌中 以熱休克選殖的方法發現,位於大腸桿菌基因組 (genome) 之 88.9 分鐘處,約 2 kb 左右的操縱組 (operon),clpQ 在前而 clpY 在後,組成 clpQY operon;其會受到熱 逆境而誘導表現,clpQY mRNA 表現量相較於未誘導時增加約 1.5 倍 (Chuang et al., 1993)。在 clpQY operon 的上游序列中,具有一高度保守性的啟動子 (promoter) 序 列,此序列與 dnaK 和 groE 等可被 σ32辨認的啟動子序列非常相似,而在大量誘 導 σ32 表現的實驗中,ClpYQ 蛋白酶的表現量亦會隨之增加約十倍,因此認為 clpQY 為一熱休克基因 (heat shock gene),又稱為 hslVU (heat shock locus) (Rohrwild et al., 1996)。
clpQY operon 可轉譯出兩個蛋白質,分別為 19.1 kDa 的 ClpQ 與 49.6 kDa 的 ClpY。經由胺基酸序列的比對發現,ClpQ 的序列與真核生物 20S proteasome 的 β-type 單體具有 18%的相似性,實驗結果也顯示 ClpQ 與 20S proteasome 同樣屬於
threonine protease (Yoo et al., 1996, Missiakas et al., 1996);而 ClpY 在靠近 N 端部份 有一個 ATP/GTP-binding motif 的保守性序列 (GXXGXGKT),若在此序列造成突 變,則 ClpY 會喪失水解 ATP 之能力,並且無法與 ClpQ 協同降解短鏈胜肽 (Shin et al., 1996)。實驗中若以 ADP、AMP 或其他 ATP 之類似物取代 ATP 時,ClpYQ 便 無法分解疏水性基質 Z-Gly-Gly-Leu-AMC,因此認為 ClpYQ 為 ATP 依賴蛋白酶的 一種,可水解 ATP 並降解蛋白質 (Rohrwild et al., 1996)。
Clp ATPase Family
在胺基酸序列比對分析的結果中顯示,ClpY 與另一 ATP 依賴蛋白酶 ClpXP 複合體中的 ClpX,具有 50%的胺基酸相同度 (identity);且 ClpY 之 ATP/GTP-binding motif 保守性序列,亦與 ClpAP 蛋白酶中的 ClpA 序列相同,因此將 ClpY 歸為 Clp ATPase family 的一員;這類蛋白酶在 in vitro 下,能促使協同的 peptidase 蛋白酶分 解酪蛋白 (casein),故以此能力 caseinolytic protease (Clp) 來命名 (Schirmer et al., 1996)。
Clp ATPase family 是屬於 AAA+ (ATPases associated with diverse cellular activities) superfamily 中的一群,均具有保守性的 ATP binding domain,約 200 - 250 個胺基酸左右,負責 ATP 的結合與水解 (Neuwald et al., 1999, Lupas & Martin, 2002)。而依據保守性區域的個數可將 Clp ATPase family 的蛋白酶分為兩類,第一 類如:ClpA、ClpB 和 ClpC,為具有兩個保守性 domain 的蛋白酶,AAA-1 (D1) 和 AAA-2 (D2) 之間會由一段序列隔開,在不同菌種中相隔的距離不同;第二類如:
ClpX 和 ClpY,為僅具有一個保守性 domain 的蛋白酶,且其序列上是與 AAA-2 (D2) 較具同源相似性 (Schirmer et al., 1996)。
雙單元體雙單元體
雙單元體雙單元體 (Two component system)
ClpYQ 蛋白酶為雙單元體 (two component system) 的蛋白酶,如同 Clp family 中 的 ClpAP 和 ClpXP , 包 含 兩 個 不 同 功 能 的 單 元 體 peptidase 及 ATPase
4
(Katayama-Fujimura et al., 1987, Gottesman et al., 1993);其中 ClpP 為具有 peptidase 功能的單元體,而 ClpA 與 ClpX 相似,同樣是具有 ATPase 功能之單元體。此外 ClpA 及 ClpX 本身亦具有 chaperone 功能,且 ClpA 與 Clp family 中另一成員 ClpB 具有 80%的相似度,ClpB 同樣有 chaperone 的功能 (Gottesman, 1996)。
在 ClpYQ 雙單元體蛋白酶中,ClpQ 扮演著 peptidase 的角色,具有降解基質 的活性。ClpQ 是由 176 個胺基酸構成,在蛋白酶性質的分類上,ClpQ 的活性中心 為 threonine residue,與真核生物 20S proteasome 同樣屬於 threonine protease,但與 ClpP 的所屬的 serine protease 不同 (Missiakas et al., 1996)。
ClpY 則與 ClpA、ClpX 相似,帶有一個保守性的 ATP binding domain,在 ClpYQ 雙單元體蛋白酶中,主要扮演 ATPase 及 unfoldase 的角色,具水解 ATP 的活性,
其本身也有 chaperone 的功能。ClpY 是由 443 個胺基酸構成,主要功能是以水解 ATP 作為能量來源,負責基質的辨認、結合,並將基質結構打開,再經由孔洞傳 送入 ClpQ 的活性中心進行降解 (Sauer et al., 2004)。
三 三
三 三、 、 、 、ClpYQ 複合體 複合體 複合體 複合體結構 結構 結構 結構
最早以生化方法純化 ClpYQ 時,發現其分子量約為 650 - 700 kDa,遠大於單 體的 ClpY 或 ClpQ,顯示 ClpYQ 均是以聚合蛋白的形式存在,並相互結合成一複 合體 (Rohrwild et al., 1996)。從分子量的倍數差異推測,ClpQ 聚合蛋白約是由 12 - 14 個單體組成,ClpY 聚合蛋白約是由 6 - 7 個單體所組成的 (Kessel et al., 1996)。
之後透過電子顯微鏡的觀察 (Rohrwild et al., 1997) 及 X-ray 結晶繞射的分析 (Bochtler et al., 1997, Bochtler et al., 2000),確認 ClpQ 及 ClpY 均是以六個單體排列 成環狀的形式存在,其中 ClpQ 的直徑約 10 - 11 nm,ClpY 則稍微大些,直徑約 13 nm。整個 ClpYQ 複合體是由四個平行排列的圓環形成一桶狀結構,其中兩個是 ClpQ 的六元環,位於中央作為蛋白酶的催化中心,另兩個則是 ClpY 的六元環,
以相反的方向分別接在兩個 ClpQ 的外側,組成 YQQY 形式的複合體 (附圖一)。
在結晶結構的分析中,Bochtler 等人將 ClpY 蛋白質分為三個區域 (附圖一),
分別是 N domain:N-terminal domain (S2-K109;I244-L332),I domain:Intermediate domain (M110-A243),以及 C domain:C-terminal domain (Q333-L443) (Bochtler et al., 2000)。其中 N domain 帶有 ATP 可結合的區域 (ATP binding site),I domain 位於遠 離 ClpQ 的一側,負責基質的辨識與結合作用 (Wang et al., 2001a, Ramachandran et al., 2002);C domain 則在靠近 ClpQ 的一側,負責 ClpY 與 ClpQ 分子間的聚合作 用,且其尾端 8 - 10 左右的胺基酸對於活化 ClpQ 具有重要性 (Seong et al., 2002, Lee et al., 2003)。
ClpYQ 蛋白酶與其他 ATP 依賴蛋白酶相似,複合體由四個相互堆疊的圓環構 成桶狀的聚合體,將水解蛋白質的活性區包埋於中央的空腔內,兩邊的出入口則 是狹窄的孔洞,孔徑約 10Å 左右,以確保構形正確的蛋白質 (folded proteins) 無法 進入水解中心;因此兩側的 ClpY 必須先將辨認的目標基質打開構形 (unfolding),
以單一條或二條胜肽鏈的形式,才能通過孔洞送入 ClpQ 的活性區進行降解作用 (Zolkiewski, 2006)。此外,也有研究認為當 ClpY 之 I domain 尚未與基質辨識結合 時,其孔洞是呈現關閉的,為親水性;而當 I domain 辨認基質後,孔洞才會打開 並轉為親脂性,以便將基質送入 (Wang et al., 2001b)。
四 四
四 四、 、 、 、ClpYQ 蛋白酶 蛋白酶 蛋白酶 蛋白酶之 之 之基質 之 基質 基質 基質
變異蛋白質 變異蛋白質 變異蛋白質
變異蛋白質 (Abnormal proteins)
ClpYQ 蛋白酶在生物體中可降解的基質主要有兩大類,一類是變異的蛋白質 (abnormal protein),包括構形錯誤 (misfolded) 的蛋白質,或是在逆境下不正常累 積的蛋白質。在實驗中過量表現 ClpYQ 蛋白酶,可以抵抗細胞中累積的有害蛋白 質毒素如嘌呤黴素 (puromycin) 及伴刀豆蛋白 A (concanavalin A) (Missiakas et al.,
6
1996)。這類基質的降解作用是為維持細胞內蛋白質的平衡穩定及正常的活性功 能,以避免細胞受到危害,也就是先前提到的蛋白質品質控制系統 (Protein quality-control systems)。
另一類的基質則是些生理上的物質 (physiological substrates),如 SulA、RcsA、
RpoH 及 TraJ 等,這類物質通常為短暫存在的調控蛋白,當其作用完後就必須快速 被降解,以維持細胞的正常生長。其中 SulA 及 RcsA 亦是 Lon 蛋白酶的基質 (Mizusawa & Gottesman, 1983, Gottesman & Maurizi, 1992),RpoH 亦為 FtsH 蛋白酶 的基質 (Yura et al., 1993)。
SulA (Suppressor of ultraviolet sensitivity A)
SulA 是基因 sulA (sfiA) 轉譯出之蛋白質,在 SOS 反應中會被大量誘導表現,
為一細胞分裂的抑制物,可與細胞分裂蛋白 FtsZ 結合形成複合物,達到抑制分裂 的效果,導致細胞死亡 (Bi & Lutkenhaus, 1990)。最早,發現 SulA 可以被 Lon 蛋 白酶所分解,為 Lon 的基質之一 (Mizusawa & Gottesman, 1983);於 1997 年時在 lon- 突變株中發現,大量表現 ClpYQ 蛋白酶可以彌補 Lon 蛋白酶分解 SulA 的功 能,使原本因缺乏 Lon 蛋白酶無法分裂的絲狀細胞,恢復成正常分裂的細胞,因 此推測 SulA 亦為 ClpYQ 蛋白酶的基質 (Khattar, 1997, Kanemori et al., 1999a)。
RcsA (Regulator of capsule synthesis A)
RcsA 為一胞外莢膜生成基因 cps (capsule synthesis) 的調控蛋白,是一種轉錄 活化物 (transcriptional activator),能夠正向調控基因表現量的增加,促使筴膜生 成,而莢膜一般被認為與細菌的致病力及於宿主外存活能力有關 (Torres-Cabassa
& Gottesman, 1987, Stout et al., 1991)。最早於 1985 年時,Gottesman 等人提出 Lon 蛋白酶可間接調控莢膜生成,後續的研究也證實可調控莢膜生成的 RcsA 為 Lon 蛋白酶的基質之一 (Gottesman et al., 1985, Gottesman & Maurizi, 1992)。之後陸續 研究發現在 lon- 突變株中,RcsA 仍會被降解,而 clpQY operon 缺失的突變株則會
呈現輕微的類黏蛋白現象 (mucoid) (Missiakas et al., 1996);於 1999 年 Wu 等人藉 由 cpsB::lacZ 融合報導基因,在 lon- 突變株中大量表現 ClpYQ 蛋白酶,可使得 cpsB::lacZ 表現量大幅降低,顯示 ClpYQ 蛋白酶同樣具有分解 RcsA 的能力,可能 是扮演 Lon 蛋白酶的輔助角色。因此 RcsA 同樣是 ClpYQ 與 Lon 蛋白酶的基質 (Wu et al., 1999, Kuo et al., 2004)。
RpoH (Sigma factor, σσσσ32)
RpoH 為一 sigma factor (σ32),可啟動熱休克基因的表現,為一熱休克基因轉 錄因子。在正常溫度下,RpoH 非常不穩定半衰期約 1 分鐘,然而於高溫或逆境下 時,RpoH 因穩定度提高而大幅增加,可進一步啟動熱休克基因表現。在一些蛋白 酶的突變株中(lon-、clpP-、ftsH-),若將 clpQY 基因去除,會使得 RpoH 的穩定度 大幅提高,而增加熱休克蛋白的合成,並對高溫敏感;然而,當大量表現 ClpYQ 蛋白酶後,RpoH 的量明顯減少並有抑制熱休克反應的現象,因此推測其為 ClpYQ 蛋白酶之基質 (Kanemori et al., 1997, Kanemori et al., 1999b)。
TraJ (F regulatory protein)
TraJ 為 F plasmid conjugation 過程中的一個活化物,對於細菌間的接合作用具 有正向調控 (Frost et al., 1994)。最近的研究顯示,在實驗中大量誘導外膜脂蛋白 (outer membrane lipoprotein, nlpE) 生成,會導致 Cpx (conjugative plasmid expression) 壓力反應 (Cpx stress response),而促使 ClpYQ 蛋白酶降解 TraJ (Lau-Wong et al., 2008)。
五 五
五 五、 、 、 、細胞分裂抑制物 細胞分裂抑制物 細胞分裂抑制物 細胞分裂抑制物 SulA
SOS 反應誘導反應誘導反應誘導反應誘導 SulA 表現表現表現 表現
當大腸桿菌 (Escherichia coli) 細胞受到環境壓力,例加熱或冷的溫度驟變、
pH 值改變、氧化自由基壓力、化學藥劑或是 UV 輻射等,會導致菌體內蛋白質不
8
正常的生成累積,或是干擾 DNA 複製造成 DNA 損傷,此時細胞會啟動一連串自 我保護的機制,以維持細胞的存活,如熱休克反應 (Heat shock response) 或 SOS 反應 (SOS response)。
SOS 反應 (附圖二) 會透過 LexA 的自我水解機制 (Fernandez De Henestrosa et al., 2000),誘導將近 30 個基因的表現,大部份是與 DNA 修復相關的蛋白質或酵 素 (Walker, 1996)。其中一個受 SOS 反應調控的基因 sulA (sfiA),為細胞分裂的抑 制物,其在 SOS 反應中會大量的生成累積,可促使細胞停止分裂,避免在 DNA 修復過程中,將受損的 DNA 傳到子代細胞中 (Huisman & D'Ari, 1981, Gottesman et al., 1981)。
SulA 抑制細胞分裂抑制細胞分裂抑制細胞分裂抑制細胞分裂
蛋白質 SulA 是由 169 個胺基酸組成,分子量為 18 kDa,其在各種腸道菌中具 有高度的保守性 (Freudl et al., 1987);當 DNA 受到損傷時,SulA 會被誘導大量生 成,直接和細胞分裂的必需蛋白質 FtsZ 結合 (Schoemaker et al., 1984, Higashitani et al., 1995)。原核細胞進行細胞分裂的過程中,FtsZ 會受到 GTP 水解的調控,於細 胞中央逐漸聚合成環狀構造 FtsZ-ring,形成細胞隔板,以達成細胞分裂(Bi &
Lutkenhaus, 1991);然而,當 SulA 和 FtsZ 結合成複合物,則會抑制 FtsZ 的聚合,
無法形成環狀構造,導致細胞分裂停止 (Mukherjee et al., 1998)。
直到 DNA 修復完成,SOS 反應結束後,細胞才會再次進行細胞分裂。此時有 兩個系統分別調控細胞分裂的重新啟動:其一是藉由 SOS 反應的終止,停止 SulA 生成,屬於轉錄層次上的調控 (Gottesman et al., 1981, Mizusawa & Gottesman, 1983);另一則是轉譯後調控,透過 ATP 依賴蛋白酶,如:Lon 及 ClpYQ (HslUV),
盡快將剩餘的 SulA 分解,以恢復細胞正常的生長活動 (Seong et al., 1999, Wu et al., 1999)。
SulA 之結之結之結之結晶結構晶結構晶結構 晶結構
SulA 於 2003 年在綠膿桿菌 (Pseudomonas aeruginosa) 中被解出結晶結構,主 要是由五個平行排列的 β-strands 與四個環繞在兩側的 α-helices 所組成 (圖一),
在序列上 α-helices 和 β-strands 是相互交錯排列的 S1-H1-S2-H2-S3-H3-S4-H4-S5 (Cordell et al., 2003)。然而,其中 SulA 的 N 端約 40 個胺基酸,在結晶結構中無法 看到明確的電子密度圖 (附圖三),可能是因為此區域較具彈性而不易解出結構,
且其在序列比對上亦較不具保守性 (圖五) (Freudl et al., 1987);在酵母菌雙雜交系 統中,即使去除 SulA 之 N 端 30 個胺基酸,仍不影響其與 FtsZ 的結合作用 (Huang et al., 1996)。
此外,結晶結構也顯示 SulA 是以二聚體 (dimer) 的形式存在 (附圖四),由單 體的第一個 α-helices (H1) 和 β-strands (S1) 部份,與另一單體的相對位置結合,
其中 SulA 的 β-strands 相互平行排列,形成一橫越二聚體的 β-sheet 平板。同時作 者也發現,SulA 均是以此二聚體的形式,由外側兩端的 S3 和 H3 部份,分別與 FtsZ 的 T7 loop 結合,形成 Z:A:A:Z 的長形複合物,以阻斷 FtsZ 的聚合作用,達到抑 制細胞分裂 (Cordell et al., 2003)。
六 六 六
六、 、 、 、蛋白酶對基質 蛋白酶對基質 蛋白酶對基質 蛋白酶對基質 SulA 辨認 辨認 辨認之相關研究 辨認 之相關研究 之相關研究 之相關研究
一般蛋白酶 一般蛋白酶 一般蛋白酶
一般蛋白酶對於基質的對於基質的對於基質的對於基質的辨認辨認辨認辨認
在蛋白質品質調控系統中,為避免不必要的蛋白質降解,對於被降解基質的 選擇都有嚴密的調控,如 ATP 依賴蛋白酶便擁有複雜的桶狀結構,將蛋白水解的 活性區包埋於中央,利用立體空間的阻隔減少不慎地與蛋白質結合,唯有先被辨 認過蛋白質,才能穿越狹窄通道被傳送到達水解活性區。在 ATP 依賴蛋白酶中,
是由 ATPase 的部份負責基質辨認的工作,並利用水解 ATP 作為能量來源,將蛋白 質結構打開,再傳送基質至 peptidase 蛋白水解的活性區。
10
因此,蛋白酶如何選擇辨認基質的機制便顯得非常重要,目前眾多研究指出,
ATP 依賴蛋白酶可能是藉由辨認目標基質的 N-端或 C-端之特定序列進行結合,而 這個結合作用是將基質降解的重要起始步驟。不同的蛋白酶之間也有著不同的辨 識特徵,如 ClpAP 和 ClpXP 蛋白酶可以辨認 C-端帶有 ssrA tag 的蛋白質,而進行 降解作用,但 ClpYQ 與 Lon 卻不能 (Gottesman et al., 1998)。FtsH 蛋白酶可降解 C-端帶有非極性胺基酸序列的蛋白質 (Herman et al., 1998);最近的研究則指出,
Lon 蛋白酶可以辨認一段 β20 的序列,其序列中含有較多的芳香族胺基酸 (Gur &
Sauer, 2008)。
ClpYQ 與與與 Lon 對於與 對於對於 SulA 的辨認對於 的辨認的辨認的辨認
SulA 是 ClpYQ 蛋白酶可降解基質第二類 (physiological substrates) 中最重要 的一個。在環境壓力下,SulA 蛋白質的表現會抑制細胞分裂,避免將受損的 DNA 傳遞到子代細胞,當 DNA 修復完成後,就必須將其快速降解以恢復細胞的正常分 裂,因此蛋白酶對於 SulA 的辨認是非常重要的。目前已知 SulA 為 Lon 及 ClpYQ 蛋白酶的基質,其中又以 Lon 為主要負責的蛋白酶,可快速降解 SulA 蛋白,當有 Lon 蛋白酶時,SulA 的半衰期僅 1.2 分鐘,而在 lon- 突變株中,SulA 的半衰期可 達 19 分鐘之久 (Mizusawa & Gottesman, 1983)。而 ClpYQ 蛋白酶似乎扮演著輔助 的角色,因為實驗顯示,在大腸桿菌 lon- 突變株中,誘導 SOS 反應會有明顯 SulA 蛋白的累積,造成細胞分裂被抑制;當大量誘導 ClpYQ 蛋白酶表現後,則可降低 SulA 蛋白的累積,恢復細胞分裂 (Goldberg, 1992)。
目前有研究指出,當 SulA 去除 C-端末 8 個胺基酸 (SA8) 後,在 in vivo 下相 較於正常的 SulA 蛋白,累積量變多且較穩定,而在 in vitro 下,其與 MBP 之融合 蛋白無法被 Lon 蛋白酶所降解,推測可能是因為去除 C-端 8 個胺基酸後的 SulA (SulAC161),無法與 Lon 進行結合而影響降解作用。若將此 8 個胺基酸 (SA8) 與 Lon 蛋白酶反應,發現其能夠直接與 SA8 結合,並抑制 Lon 蛋白酶對於正常 SulA
的降解,因此認為 Lon 蛋白酶可以藉由辨認 SulA 之 C-端末 8 個胺基酸,來進行結 合以及後續的降解作用 (Ishii et al., 2000)。
之後的研究進一步指出,SulA 之 C-端末 8 個胺基酸 KIHSNLYH 中,又以 Histidine 佔有較重要的角色,如果只將最後一個胺基酸置換成 Aranine,則 SulA 蛋白會大量累積並提高穩定度,與 SulA 去除末 8 個胺基酸 (SulAC161) 的結果相 似;然而,若將 SulAC161 末端再加上一個 Histidine 胺基酸 (SulAC161+H),會使 SulA 變得不穩定且快速被 Lon 蛋白酶降解 (Ishii & Amano, 2001)。
然而,有研究利用 SulA 之末端 11 個胺基酸 (CT11) 接於 GFP 的 C-端,作為 ClpYQ 蛋白酶辨膱的 tag,但卻無法使 GFP 被降解 (Kwon et al., 2004)。而在 2003 年,Lee 等人的研究指出,ClpY 與 SulA*M89I 之間有很強的結合力,故以 SulA*M89I 作測試,當 SulA 去除 N 端 43 個胺基酸後,仍可與 ClpY 有交互作用;但 SulA 去 除 C-端 40 個胺基酸後,則與 ClpY 沒有交互作用的情形,因此認為 ClpYQ 蛋白酶 可能是藉由 C-端序列來辨識 SulA 蛋白質 (Lee et al., 2003)。
在 ClpY 的 I domain 上具有兩個 loop,分別是 loop1:137 - 150 a.a. 和 loop2:
175 - 209 a.a.,有研究認為 ClpY 是藉由這兩個 loop 來進行基質的辨識與結合,以 酵母菌雙雜交系統測試的實驗結果中,若將兩個 loop 都去除,則 ClpY 無法與基 質 SulA 結合,但若只去除 loop1 或只去除 loop2 時,ClpY 仍然可與基質 SulA 結 合,顯示 ClpY 之 I domain 上的兩個 loop 均具有基質辨識與結合的功能。其中又 以 loop2 較為重要,因為去除 loop2 之後的 ClpY 便無法降解基質,而只去除 loop1 的 ClpY 仍可正常降解,顯示 loop2 可能還參與後續基質結構打開及傳送的過程 (Lien et al., 2009)。
12
七 七 七
七、 、 、 、研究動機與目的 研究動機與目的 研究動機與目的 研究動機與目的
對 ATP 依賴蛋白酶來說,如何選擇辨認需要降解的基質,是非常重要的,可 能是藉由某段本來包埋於結構中的短鏈胜肽,或是某些特定的胺基酸序列,來進 行辨識結合的作用;然而,關於蛋白酶是如何辨認的詳細機制目前仍不明楚。但 大部份的 ATP 依賴蛋白酶應該會使用相似的辨識機制,因此瞭解蛋白質基質最初 是如何被蛋白酶辨認並結合,應有助於瞭解後續蛋白酶將基質結構打開及降解的 作用。相關的蛋白酶如 ClpXP、ClpAP,對於基質的辨認結合已有廣泛的研究,然 而對於 ClpYQ 蛋白酶是如何選擇特定的基質,及其詳細的降解機制,相關的資訊 卻較少。
因此為找出 ClpYQ 蛋白酶是如何辨認基質的機制,我們以 SulA 作為基質,
建構 C-端不同大小片段缺失的突變,利用酵母菌雙雜交系統,測試各種 SulA 的突 變蛋白與 ClpYQ 蛋白酶結合之情形,縮小可能辨識的範圍,再建構單一胺基酸的 點突變,分析不同性質的胺基酸對於 ClpYQ 蛋白酶結合的影響,並測試各種突變 的 SulA 蛋白活性表現,以及被 ClpYQ 蛋白酶降解之情形。希望藉此找出 ClpYQ 蛋白酶是如何選擇它的基質 SulA,包括其辨認的區域特性,以及對於後續降解作 用的影響。
貳 貳 貳
貳、 、 、 、材料與方法 材料與方法 材料與方法 材料與方法
一 一 一
一、 、 、 、實驗材料 實驗材料 實驗材料 實驗材料
(一 一 一) 菌株與質體 一 菌株與質體 菌株與質體 菌株與質體
本研究使用到的菌株與質體如表一、表二所示。
1. 選殖基因表現系統選殖基因表現系統選殖基因表現系統選殖基因表現系統
選殖基因表現的宿主細胞為 lon- clpY- clpQ- 之大腸桿菌菌株 AC3112,並以 pBAD24、pBAD33 及 pTH18kr 作為選殖基因表現的載體。其中 pBAD24 和 pBAD33 具有 ara operon 的啟動子 PBAD及調控基因 araC,故可利用阿拉伯糖 (L-arabinose) 誘導或以葡萄糖 (D-glucose) 抑制目標基因的表現 (Guzman et al., 1995)。而 pTH18kr 為 low copy 數的質體,帶有啟動子 Plac,可以 IPTG (Isopropyl β-D-1-thiogalactopyranoside) 誘導下游基因表現 (Hashimoto-Gotoh et al., 2000)。
2. 酵母菌雙雜交系統酵母菌雙雜交系統酵母菌雙雜交系統酵母菌雙雜交系統
本實驗是使用 LexA Two-hybrid System,此系統的宿主細胞為帶有報導質體 p8op-lacZ 的酵母菌 EGY48,其染色體的 LEU2 基因上游接有六個 LexA 操縱子 (operator) 序列,可作為報導基因,於 leucine 缺乏的培養基上進行生長測試;報 導質體則是在 lacZ 前方接上八個 LexA 操縱子序列,利用轉譯出的β-galactosidase 直接進行活性分析,或是以含有 X-gal 的培養基測試報導基因的表現。此系統中 的兩個重要載體 pB42AD 及 pGilda 質體,可分別表現出帶有 AD (activation domain) 或 BD (DNA binding domain) 與選殖基因產物之融合蛋白,當兩蛋白質 間有交互作用時,即會啟動上述報導基因表現 (附圖五)。
pB42AD (Clontech,原名 pJG4-5) (Gyuris et al., 1993),大小 6.45 kb,為一 shuttle vector 可適用於大腸桿菌與酵母菌系統中。pB42AD 之 MCS 上游可轉譯出
14
一段聚胜肽 B42 及 HA (hemagglutinin) epitope tag,其中 B42 具有酵母菌轉錄活 化 (transcription activator) 的功能,可作為 activation domain (Ma & Ptashne, 1987)。其調節性啟動子 PGAL可受到半乳糖 (galactose) 及植物蜜糖 (raffinose) 的 誘導,或是葡萄糖的抑制,以調控融合蛋白的基因表現。pB42AD 可藉由 Ampr 在大腸桿菌或 TRP1 在酵母菌中進行篩選。
pGilda (Clontech) (Gimeno et al., 1995),大小 6.57 kb,亦為大腸桿菌與酵母 菌系統中的 shuttle vector。pGilda 之 MCS 上游為大腸桿菌的 lexA,LexA 為 E. coli 中 SOS 反應的抑制物,可辨認結合 LexA 操縱子序列,在酵母菌雙雜交系統中作 為 DNA binding domain。其同樣以調節性啟動子 PGAL來調控融合蛋白的基因表 現,可藉由 Ampr在大腸桿菌或 HIS3 在酵母菌中進行篩選 (Ebina et al., 1983)。
(二 二 二) 藥品與試劑 二 藥品與試劑 藥品與試劑 藥品與試劑
1. 一般藥品及相關酵素一般藥品及相關酵素一般藥品及相關酵素一般藥品及相關酵素
一般藥品購自 Sigma (St. Louis, U.S.A.)、J.T. Baker (Phillipsburg, U.S.A)、和光純 藥工業株式會社 (Osaka, Japan) 及生工公司 (台北,台灣)。
一般限制酶、接合酶購自 TaKaRa (Kusarsu, Japan)。
PCR 反應套組購自生工、萬造公司 (台北,台灣)。
DNA 純化套組購自 Viogene (台北,台灣)。
2. 培養基培養基培養基培養基
(1) LB (Luria-Bertani) Broth
購自 BD Biosciences (U.S.A.),用於一般大腸桿菌培養,並加入適量抗生 素以篩選轉形株。抗生素 stock 濃度分別為 Ampicillin (100 µg/ml)、Kanamycin (25 µg/ml)、Chloramphenicol (34 µg/ml);使用時以 1:1000 倍率稀釋使用。
本論文中,一般大腸桿菌培養於 37 oC,菌株 AC3112 則培養於 30 oC。
(2) SD (synthetic dropout) medium
Yeast Nitrogen Base w/o Amino Acid (Difco, BD) Minimal SD Base Gal/Raf (Clontech)
一種以 glucose 作為碳素源的營養限制培養基 (minimum medium),藉由 調 配 缺乏 某些 特 定胺基 酸 的 SD medium ,作為選擇性培養基 (selective medium),可用以篩選轉形株。在 LexA 系統中,一般培養是使用 glucose 作 為碳素源,當需要誘導質體表現所建構的 LexA 及 B42 融合蛋白時,則以 galactose 和 raffinose 作為碳素源,以進行報導基因測試。本論文中酵母菌均 培養於 30oC。
3. 實驗相關核酸引子實驗相關核酸引子實驗相關核酸引子實驗相關核酸引子
參照表三。PCR 引子由基龍米克斯、富聯公司 (台北,台灣) 所合成。
(三 三 三) 器材設備 三 器材設備 器材設備 器材設備
微量離心機 (Kubota KM-15200,雙鷹公司,台北,台灣) 冷凍超高速離心機 (CF 15D2, Hitachi, Japan)
PCR 反應槽 (T3 Thermocycler, Biometra, Germany) DNA 電泳槽 (伯昂公司,台北,台灣)
蛋白質電泳槽 (Amersham, CA, U.S.A.) 蛋白質轉印槽 (Bio-Rad, CA, U.S.A.)
分光光度計 (Ultraspec 2000, Pharmacia, H.K.)
全波長掃描式微盤分析儀 (SpectraMax 340PC384, CA, U.S.A.)
(四 四 四) 分析軟體 四 分析軟體 分析軟體 分析軟體
BioEdit Sequence Alignment Editor (BioEdit) v7.0.9.0
Hydrophobicity calculations:於 http://www.expasy.org/tools/protscale.html 上輸入序 列,選擇“average area buried”,以預測軟體計算蛋白質之 Surface-burial score。
16
二 二 二
二、 、 、 、實驗方法 實驗方法 實驗方法 實驗方法
(一 一 一) 一般 一 一般 一般 一般實驗方法 實驗方法 實驗方法 實驗方法
1. 相關套組使用相關套組使用相關套組使用相關套組使用
(1) Mini-MTM Plasmid DNA Extraction kit (Viogene,富聯生物科技公司) 此套組用以小量萃取細菌中質體,詳細實驗步驟參考使用手冊。
質體 pTH18kr 以三倍量體積的液態培養基培養。
(2) QIAGENR Plasmid Midi kit (Qiagene,購自台灣諾貝爾生物公司) 此套組用以大量萃取細菌中質體,詳細實驗步驟參考使用手冊。
(3) Gel-MTM Gel Extraction kit (Viogene,富聯生物科技公司)
此套組用以回收電泳膠體中之 DNA,詳細實驗步驟參考使用手冊。
(4) MontageR PCR Centrifugal Filter Devices (Montage,購自伯森生物科技公司) 此套組用以純化 PCR 產物,詳細實驗步驟參考使用手冊。
2. 勝任細胞製備勝任細胞製備勝任細胞製備勝任細胞製備
(1) 氯化鈣法氯化鈣法氯化鈣法氯化鈣法 (一般情況使用一般情況使用一般情況使用) 一般情況使用
1. 於 37oC 培養箱中隔夜培養菌體 (12 - 16 小時)。
2. 取 1 ml 菌液接種於 100 ml LB 培養液,重新震盪培養菌體至 OD600吸光 值為 0.2 - 0.8。
3. 將菌液平均分裝至兩支離心管,冰浴 10 分鐘。自此步驟起均保持低溫操 作。
4. 以 6000 rpm 在 4oC 下,離心 10 分鐘,移除上清液。
5. 每管以 1 - 2 ml 冰的 0.1 M CaCl2 懸浮菌體,再加入 3 - 5 ml 冰 0.1 M CaCl2
混勻,冰浴 30 分鐘。
6. 以 4000 rpm 在 4oC 下,離心 10 分鐘,移除上清液。
7. 重複步驟 5 和 6 一次。
8. 每管加入 1 ml 冰 0.1 M CaCl2 / 15% glycerol,懸浮菌體。
9. 每 100 µl 菌液分裝至菌種保存管中,儲存於 - 70oC。
(2) 電穿孔法電穿孔法電穿孔法電穿孔法 (用於電穿孔實驗用於電穿孔實驗用於電穿孔實驗) 用於電穿孔實驗
1. 於 37oC 培養箱中隔夜培養菌體 (12 - 16 小時)。
2. 取 1 ml 菌液接種於 100 ml LB 培養液,重新震盪培養菌體至 OD600吸光值 為 0.4 - 0.6。
3. 將菌液平均分裝至兩支離心管。自此步驟起均保持低溫操作。
4. 以 4000 rpm 在 4oC 下,離心 15 分鐘,移除上清液。
5. 加入 100 ml 滅菌的 10% glycerol 清洗菌體,以 4000 rpm 在 4oC 下,離心 15 分鐘,移除上清液。
6. 加入 50 ml 滅菌的 10% glycerol 清洗菌體,以 4000 rpm 在 4oC 下,離心 15 分鐘,移除上清液。
7. 加入 2 ml 滅菌的 10% glycerol 清洗菌體,以 4000 rpm 在 4oC 下,離心 15 分鐘,移除上清液。
8. 加入 200 µl 滅菌的 10% glycerol 混勻,即製成勝任細胞,每 50 µl 分裝成 一管。(冷凍過的勝任細胞轉形效率較低。)
3. 轉形作用轉形作用轉形作用轉形作用
(1) E. coli Heat shock transformation (Sambrook, 1989)
1. 取 50 µl 勝任細胞與 20 µl 質體接合反應 (ligation) 的產物,或 5 µl 小量製 備的質體混合,冰浴 50 分鐘。
2. 置於 42oC 下,進行熱休克反應 (heat shock) 90 秒後,迅速放入冰中。
3. 加入 1 ml LB 液態培養基,在 37oC 培養箱中震盪培養 1 小時。
4. 以 6000 - 8000 rpm 離心四分鐘,移除上清液,留下約 50 µl 懸浮菌體。
5. 塗於含適當抗生素的培養基上,置於 37oC 培養箱培養隔夜。
18
(2) E. coli Electropotation transformation
1. 取 2 µg 的質體 (約 0.5 µl 小量製備的質體) 加入事先準備的 50 µl 電穿孔 勝任細胞中混勻。
2. 將混勻的勝任細胞放入 P/N 620 cuvette 中,以 2500 V 電壓進行電擊穿孔 反應。
3. 電擊結束後,迅速加入 1 ml SOC 培養液,在 37oC 培養箱中震盪培養 1 小 時。
4. 以 6000 - 8000 rpm 離心四分鐘,移除上清液,留下約 50 µl 懸浮菌體。
5. 塗於含適當抗生素的培養基上,置於 37oC 培養箱培養隔夜。
SOC medium 4oC 保存
Tryptone 20.0 g
Yeast extract 5.0 g
NaCl 0.5 g
1M KCl 2.5 ml
調整 pH 至 7.0,加水至總體積為 1 升,滅菌後再加入 1000 ml
1M Glucose 20.0 ml
2M MgCl2 5.0 ml
(3) TSS-Transformation
1. 取 30 µl 隔夜培養菌液,加入 3 ml LB 培養液中,震盪培養菌體至 OD600
吸光值為 0.6 - 0.8。
2. 將菌液以 3000 rpm,離心 10 分鐘,移除上清液。
3. 以 0.3 ml 2X TSS 溶液將菌體懸浮,加入 5 µl 質體混勻後,冰浴 50 分鐘。
4. 置於 37oC 下,進行熱休克反應 (heat shock) 2 分鐘後,迅速放入冰中。
5. 加入 0.5 ml LB 液態培養基,在 30oC 培養箱中震盪培養 1 小時。
6. 以 6000 - 8000 rpm 離心四分鐘,移除上清液,留下約 50 µl 懸浮菌體。
7. 塗於含適當抗生素的培養基上,置於 30oC 培養箱培養隔夜。
2X TSS (Transformation and storage solution) 4oC 保存 PEG 8000 (Polyethylene glycol 8000) 20.0 g
1 M MgSO4 4.0 ml
加 LB 至總體積 190 ml,滅菌後再加入 190 ml DMSO (Dimethyl sulfoxide) 10.0 ml
(4) Yeast transformation
1. 挑選單一酵母菌菌落接種至 5 ml 選擇性培養液中 (SD medium),於 30oC 培養箱隔夜培養約 12 - 16 小時,OD600 = 0.4 - 0.8,以 3000 rpm 離心 10 分 鐘,移除上清液,收集菌體。
2. 用 2.5 ml 無菌水清洗菌體,再以 3000 rpm 離心 10 分鐘,移除上清液 3. 加入 100 µl 的 100 mM LiOAc 懸浮菌體,混勻後移至微量離心管。以 14000
rpm 離心 5 秒,移除上清液。
4. 再加入 40 µl 的 100 mM LiOAc 懸浮菌體,以 14000 rpm 離心 5 秒,移除 上清液。酵母菌勝任細胞製備完成。
5. 先以 240 µl 的 50% (w/v) PEG 4000 將酵母菌勝任細胞懸浮,再加入 36 µl 的 1 M LiOAc、25 µl 的 salmon sperm carrier DNA 及 50 µl 欲送入的質體 (以 5 µl 小量製備的質體稀釋 10 倍),劇烈震盪 1 分鐘混合均勻。
* 其中 salmon sperm carrier DNA 需先以 99oC 處理 10 分鐘打開雙股,再迅 速置於冰上待用。
6. 於 30oC 培養箱中震盪培養 30 分鐘,再移至 42oC 水浴槽進行熱休克反應 (heat shock) 20 分鐘。
7. 以 14000 rpm 離心 15 秒,移除上清液。以 100 µl 無菌水懸浮菌體,塗抹 於適當篩選的 SD 培養基,於 30oC 培養箱培養 2 天。
4. 瓊脂膠體電泳分析瓊脂膠體電泳分析瓊脂膠體電泳分析瓊脂膠體電泳分析
(1) 限制酶截切反應限制酶截切反應限制酶截切反應限制酶截切反應 (Digestion)
取適量質體,加入選用的限制酶及其配合之緩衝液,反應溫度視限制酶
20
種類決定,反應時間約 60 - 70 分鐘。反應後將溶液加入適量的追蹤染劑,進 行 DNA 瓊脂膠體電泳。
(2) DNA 瓊脂膠體瓊脂膠體瓊脂膠體瓊脂膠體 (agarose gel) 電泳電泳電泳電泳
用 1× TAE 緩衝液配製 0.8 - 1.0% 的瓊脂膠體,以 100 伏特之電壓進行 電泳,反應時間 25 - 35 分鐘後;利用 Ethidium Bromide (EtBr) 進行外染,經 退染後再以紫外光觀察 DNA 產物。
(3) 回收電泳膠體之回收電泳膠體之回收電泳膠體之回收電泳膠體之 DNA
進行 DNA 瓊脂膠體電泳,並經 EtBr 染色及退染後,使用長波長之紫外 光源 (365 nm),觀察 DNA 產物片段大小,並挖取欲回收之片段位置,再使 用 Viogene Gel-M Gel Extraction kit 回收電泳膠體之 DNA。
(二 二 二) 菌體的建構 二 菌體的建構 菌體的建構 菌體的建構
1. 噬菌體實驗方法噬菌體實驗方法噬菌體實驗方法噬菌體實驗方法
(1) 製備噬菌體製備噬菌體製備噬菌體製備噬菌體 (Preparation of phage plate lysate)
1. 取隔夜培養菌液 0.2 ml,加入 2 - 3 個 plaques,於 37oC 靜置培養 15 分鐘。
2. 將 3 ml 的 R-Top Agar 隔水加熱溶解,待其降溫後將步驟 1 之菌液加入。
3. 混合均勻後倒在 LA 平板培養基上,搖晃使其攤平,待乾。
4. 於 37oC 培養箱中倒置培養 5 - 8 小時,待培養基上長出許多 plaques 後,
即可收集噬菌體 lysate。
5. 加 5 ml TM buffer 於培養基內,靜置於 4oC 數小時或隔夜後,將培養基內 的溶液吸取到試管內。
6. 加入 0.4 ml 的氯仿 (chloroform),於 4oC 下靜置 30 分鐘後,以 3000 rpm 離心 10 分鐘,轉移上清液至乾淨試管。此步驟重複二次。
7. 取得之噬菌體 lysate 保存於 4oC。
R-Top Agar
Bacto tryptone 10 g
Bacto yeast extract 1 g
Difco agar 8 g
NaCl 8 g
加水至總體積為 1000 ml,滅菌後再加入 1000 ml
1 M CaCl2 2 ml
20% Glucose 5 ml
Tris-magnesium (TM) buffer
1M Tris-HCl (pH 7.5) 20 ml
MgSO4 0.4 g
加水至總體積為 400 ml,滅菌後 4oC 保存 400 ml
(2) P1 lysate 的製備的製備的製備的製備
以 P1 噬菌體感染染色體上帶有某特定基因之細菌細胞,再收集噬菌體之 lysate,用以進行後續的 P1 轉導作用 (P1 transduction)。
1. 將 E. coli donor strain 接種至 5 ml LB 培養液中,震盪培養於生長溫度 (依 菌株特性決定生長溫度 37oC 或 30oC)。
2. 將隔夜培養菌液以 1:100 接種於 5 ml LB 培養液中,其內含有 50 µl 的 20%
Glucose (終濃度 0.2%),及 25 µl 的 1M CaCl2 (終濃度 5 mM),於適合之溫 度震盪培養 30 - 45 分鐘。(鈣離子為病毒吸附所必須。)
3. 加入 100 µl 新製備的 P1 vir stock (109 - 1010 pfu/ml),繼續震盪培養約 3 小 時,直到菌體細胞呈現溶解 (lysis)。
4. 加入 200 µl 氯仿,連續搖動數分鐘使其充分混合,以確保培養液中無殘留 細菌,置於冰上 20 分鐘。
5. 以 3500 rpm 離心 10 分鐘後,轉移上清液至新試管,再重複步驟 4 一次。
6. 將上清液轉移至新試管後,標記 lysate 種類 (如:P1vir/sulA-),置於 4oC 保存。(此 lysate 可於 4oC 下保存數年,但氯仿可能會減低病毒存活率。)
22
(3) P1 轉導作用轉導作用轉導作用轉導作用 (P1 transduction)
以帶有特定基因之 P1 lysate 感染欲接受的菌體細胞,將此特定基因轉移 至目標菌體中。
1. 將 E. coli recipient strain 接種至 5 ml LB 培養液中,震盪培養於生長溫度。
2. 取隔夜培養菌液 1.6 ml 於微量離心管中,以 14000 rpm 離心 2 分鐘,移除 上清液。
3. 加入 0.8 ml 的 P1 salts solution 打散細胞,每 400 µl 分裝至新微量離心管。
4. 混入 100 µl 欲轉導的 lysate,於生長溫度中靜置 30 分鐘,使噬菌體吸附 於細胞。
5. 添加 1 ml LB 及 200 µl 的 1M sodium citrate,於生長溫度中繼續震盪培養 1 小時。(sodium citrate 可將鈣離子嵌合,避免剩餘病毒再次感染。) 6. 以 14000 rpm 離心 2 分鐘,移除上清液,加入 100 µl LB 混合,塗抹於含
適當抗生素的培養基上,培養於生長溫度中 1 - 2 天。
7. 所得的菌落以劃線法純化出單一菌落二次,避免殘存的 P1 噬菌體感染。
8. 依據菌體的基因型及表現型確認轉導到正確的基因。可利用 colony PCR 方法確認轉導基因是否於正確位置上。
P1 salts solution
CaCl2 (終濃度 10 mM) 0.44 g
MgSO4 (終濃度 5 mM) 0.24 g
加水至總體積為 400 ml,滅菌後常溫保存 400 ml
2. 建構建構建構建構 sulA- 突變株突變株突變株突變株
從日本 National BioResource Project E. coli strain 取得的大腸桿菌 sulA-菌株 (Keio Collection JW0941),其染色體上 sulA 基因已被去除,並帶有 kanamycin 抗 藥性基因。以 AC3112 為原始菌株,利用 P1 轉導作用的方式,建構 AC3112 菌株 之 sulA-突變株。再利用 FLP 重組酶 (recombinase),去除 kanamycin 抗藥性片段。
1. 先以 P1 噬菌體感染 JW0941,收集 lysate,再利用 P1 轉導作用將此片段 轉導至 AC3112 菌株,以含有 Kan 的培養基進行篩選,並利用 colony PCR 方式確認抗藥性基因片段是否於正確位置上。
2. 將質體 pCP20 以 TSS transformation 方式轉形至此菌株中,以含有 Amp 的培養基進行篩選,於 30oC 培養箱中培養隔夜。
* 質體 pCP20 為一 FLP-recombination plasmid,可辨認 FRT 位置進行重組反 應 (FLP-mediated site-specific recombination) , 其 對 溫 度 敏 感 並 帶 有 ampicillin 抗性 (Cherepanov & Wackernagel, 1995)。
3. 將所得菌落接種於 3 ml LB 培養液中,於 30oC 培養箱中繼代培養 2 - 3 次,
去除 kanamycin 抗藥性片段。
4. 塗於 LA 培養基上,於 37oC 培養箱中培養隔夜,去除質體 pCP20。
5. 挑選單一菌落,以含有 Amp 及 Kan 的培養基於 37oC 培養箱中培養隔夜,
測試質體 pCP20 及 kanamycin 抗藥性片段是否均掉落。
3. 建構建構建構建構 genomic sulA + 3xFLAG tag 之菌株之菌株之菌株之菌株
以 PCR 增幅反應產生兩側具有與目標基因末端同質性 (homologous) 的 3xFLAG tag 及抗藥性基因片段,將此片段以電穿孔方法送入細胞中,再誘導可表 現重組酶 (recombinase) 的質體,利用同質性交換 (homologous recombination) 進行重組反應,將此片段插入目標基因後方;利用溫度、抗性篩選 genomic sulA + 3xFLAG tag 之菌株 (Uzzau et al., 2001)。
* 質體 pKD46:帶有λ 噬菌體之 Red 重組酶且對熱敏感 (thermal-sensitive) 的 質體,可受誘導表現進行同質性交換 (Datsenko & Wanner, 2000)。
* 質體 pSUB11:具有 3xFLAG 及 kanamycin 抗藥性基因。
* PCR 引子設計:見表三。
24
Preparation of linear DNA
Template : pSUB11 2.0 µl
10X Ex Taq buffer 5.0 µl
dNTP (2.5 mM) 4.0 µl
Forward primer (5 µM) (sulA-3xFLAG_Fw) 1.0 µl Reversed primer (5 µM) (sulA-3xFLAG_Rv) 1.0 µl
TaKaRa Ex Taq 0.5 µl
ddH2O 36.5 µl
Total 50.0 µl
PCR 反應條件
1. Denaturation 94oC: 3 min 2. Denaturation 94oC: 30 s
3. Annealing 50oC: 30 s 30 cycles 4. Extension 72oC: 2 min
5. Extension 72oC: 2 min 6. Stop 4oC: stop
1. 以設計的引子經由 PCR 增幅反應產生帶有 3xFLAG 及抗藥性基因之片 段,再利用 DNA 瓊脂膠體電泳進行分析,回收目標大小分子量 (約 1.6 kb) 的 DNA 片段。
2. 定量 DNA 濃度達 50 - 400 ng。
* 基因片段準備完成。
3. 於含有 0.02% arabinose 及 Amp 之 LB 培養液中,在 30oC 培養箱隔夜培養 帶有質體 pKD46 之菌株 BW25113。
4. 取 0.4 ml 菌液接種於含有 0.02% arabinose 及 Amp 之 20 ml LB 培養液,重 新震盪培養至 OD600吸光值為 0.4 - 0.6。
5. 將菌液平均分裝至兩支離心管。自此步驟起均保持低溫操作。
6. 以 5500 rpm 在 4oC 下,離心 8 分鐘,移除上清液。
7. 加入 2 ml 冰無菌水懸浮菌體,分裝至兩支微量離心管中。
8. 以 12000 rpm 在 4oC 下,離心 20 秒,移除上清液。再加入 1 ml 冰無菌水。
重複此步驟三次。
* 電穿孔勝任細胞製備完成。
9. 將步驟 2 之 DNA 約 7 µl (50 - 400 ng) 加入 50 µl 製備好的電穿孔勝任細胞 中。
10. 混勻後的勝任細胞移置 P/N 620 cuvette 中,以 2500 V 之電壓進行電擊穿 孔反應。
11. 電擊結束後,迅速加入 1 ml SOC 培養液,在 37oC 培養箱中震盪培 1 - 2 小時。
12. 以 6000 - 8000 rpm 離心四分鐘,移除上清液,留下約 50 µl 懸浮菌體。
13. 塗於含有 Kan 的培養基上進行篩選,置於 37oC 培養箱培養隔夜。
14. 挑選單一菌落,利用 colony PCR 確認 3xFLAG 及 kanamycin 抗藥性基因 片段插入目標基因後方。
(三 三 三) 突變基因建構 三 突變基因建構 突變基因建構 突變基因建構
利用 PCR based site-directed mutagenesis 的方法,建構目標基因的 deletion mutants 或是 point mutants。將此選殖的基因片段接合至質體 pGilda 上,先轉形至 E. coli XL1-Blue 中進行初步篩選,再萃取出質體經定序確認建構的突變序列正 確。之後利用限制酶的切位將選殖的基因插入適當的質體中。
1. 選殖基因片段製備選殖基因片段製備選殖基因片段製備選殖基因片段製備
以質體 pB42AD-sulA 作為模板,分別設計帶有限制酶切位以及突變序列的引 子 (見表三),利用 PCR 增幅反應產生突變點的前後兩片段,以此做為模板再進 行第二次的 PCR 增幅反應,建構出帶有突變的基因片段。
1. 第一次 PCR 增幅反應
Template : pB42AD-sulA 2.0 µl
10X Taq buffer 5.0 µl
dNTP (10 mM) 1.0 µl
26
Forward primer (5 µM) (5') 0.5 µl Reversed primer (5 µM) (3') 0.5 µl
Taq DNA polymerase 0.5 µl
ddH2O 40.5 µl
Total 50.0 µl
PCR 反應條件
1. Denaturation 95oC: 10 min 2. Denaturation 95oC: 1 min 30 s
3. Annealing 48~65oC: 1 min 30 cycles 4. Extension 72oC: 1 min 30 s
5. Extension 72oC: 10 min 6. Stop 4oC: stop
* Annealing 溫度依照引子的不同而稍作改變,為 Tm 值減 5 度。
2. 將 PCR 產物經瓊脂膠體電泳分析,切下正確大小分子量的片段,利用 Viogene Gel-M Gel Extraction kit 回收純化,作為第二次 PCR 反應的模板。
3. 第二次 PCR 增幅反應
Template 1 1.0 µl
Template 2 1.0 µl
10X Pfu buffer 5.0 µl
dNTP (10 mM) 1.0 µl
Forward primer (5 µM) (5' sulA EcoRI) 0.5 µl Reversed primer (5 µM) (3' sulA BamHI) 0.5 µl
Pfu DNA polymerase 0.5 µl
ddH2O 40.5 µl
Total 50.0 µl
PCR 反應條件
1. Denaturation 95oC: 10 min 2. Denaturation 95oC: 1 min 30 s
3. Annealing 48~65oC: 1 min 30 cycles 4. Extension 72oC: 1 min 30 s
5. Extension 72oC: 10 min 6. Stop 4oC: stop
* Annealing 溫度依照引子的不同而稍作改變,為 Tm 值減 5 度。