國立臺東大學生命科學系 碩士論文
Department of Life Science National Taitung University Master Thesis
指導教授:李俊霖 博士 Advisor: Chun-Lin Lee, Ph.D.
探討副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶 及其功效成分改善阿茲海默症之研究
Study on the improvement of Alzheimer’s disease by
Lactobacillus paracasei subsp. paracasei NTU 101 fermanted soy-milk and the functional compounds
研究生:楊宗諺 撰 Student: Zong-Yang Young
中華民國 105 年 7 月
July 2016
國立臺東大學生命科學系 碩士論文
Department of Life Science National Taitung University Master Thesis
指導教授:李俊霖 博士 Advisor: Chun-Lin Lee, Ph.D.
探討副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶 及其功效成分改善阿茲海默症之研究
Study on the improvement of Alzheimer’s disease by
Lactobacillus paracasei subsp. paracasei NTU 101 fermanted soy-milk and the functional compounds
研究生:楊宗諺 撰 Student: Zong-Yang Young
中華民國 105 年 7 月
July 2016
謝辭
首先感謝在這七百多個日子裡,不斷教導我們的指導教授李俊霖老師,若不 是老師花費龐大時間與精力的指導,學生則不可能如此順利且迅速的完成此論文,
從最初的實驗設計、題目訂定、手術操作,到後續的樣品分析、口頭報告,都多 虧老師盡心盡力的教導,老師平時對我們實驗的要求,使我們精進培養出良好的 研究態度,這難能可貴的傳授使我們往後的人生受用無窮。
再者感謝潘子明老師花費龐大的時間審閱論文,並給予了我們須多寶貴的建 議,也提點了我們許多的錯誤,使本篇論文得以盡善盡美。感謝蔡宗佑老師在口 頭報告的指導和建議也使我們能發現自己的不足,以此為戒做的更好,感謝林志 輝老師在論文修訂上費盡苦心,許多學生容易忽略的小地方以及邏輯上的小缺失 都能提點我們,感謝各位老師的全心投注,若沒有各位老師的教導,本篇論文也 無法順利完成。
同時也感謝實驗室的學長學姐、同學以及學弟學妹的大力幫助,感謝黃鈞鼎 學長、洪宇平學長、賴昭如學姊以及石曉凡同學,陳瑜庭學妹、王奕婷學妹、柯 柏鈞學弟、佘文良學弟、吳彥忠學弟、何儀君學妹,動物實驗需要人手,既冗長 又繁瑣,如果沒有各位的幫忙,單單自己是無法完成實驗的,感謝各位能抽出自 己的時間不嫌麻煩的協助我,你們的付出這段文字是不足以表達的,但感謝之意 我會一直記在心底。
最後感謝我的父母,在我人生的求學階段全心全力的支持我,在我失落停滯 時鼓勵我,得意衝動時點醒我,讓我能專注在自己所選擇所喜愛的事物上,在此 與您們分享我的喜悅。
I
縮寫表 (Abbreviations)
ACh (acetylcholine) 乙醯膽鹼
AChE (acetylcholinesterase) 乙醯膽鹼酯酶
AChEI (acetylcholinesterase inhibitor) 乙醯膽鹼酯酶抑制劑
ACID (APP intracellular domain)
Al (Aluminum) 鋁
Apo E4 (apolipoprotein E4) 脂質轉運蛋白 E4
APP (amyloid precursor protein) 類澱粉樣前驅蛋白
sAPPα (soluble amyloid precursor protein α-secretase cleaved fragment) 水溶性 類澱粉樣前驅蛋白 α- 分泌酶剪切片段
sAPPβ (soluble amyloid precursor protein β-secretase cleaved fragment) 水溶性 類澱粉樣前驅蛋白 β- 分泌酶剪切片段
Aβ (amyloid β-protein) 類澱粉樣蛋白
Aβ40 (amyloid β-protein (1-40) 類澱粉樣蛋白 1-40
BACE (beta-site cleaving enzyme, β-secretase) β-切位剪切酶
CAT (catalase) 過氧化氫酶
COX-2 (cyclooxygenase-2) 環氧化酶
CTF 83 (carboxy-terminal fragment 83)
CTF 99 (carboxy-terminal fragment 99)
DPPH (α,α-diphenyl β-picrylhydrazyl) 1,1-二苯基苦味肼基自由基
ERK 1/2 (extracellular signal-regulated-kinases 1/2) 細胞外信號調節酶
FDA (Food and Drug Administration) 美國食品藥物管理局
GSH (glutathione) 穀胱甘肽 HDL-C (high density lipoprotein cholesterol) 高密度脂蛋白膽固醇
HRT (hormone replacement therapy) 荷爾蒙補充療法
IL1-β (interlukin-1β) 介白素-1β
IL-6 (interlukin-6) 介白素-6
IL-8 (interlukin-8) 介白素-8
II
IL-10 (interlukin-10) 介白素-10
iNOS (inducible nitric oxide synthase) 誘導型一氧化氮合成酶
LDL-C (low density lipoprotein cholesterol) 低密度脂蛋白膽固醇
mRNA (messenger ribonucleic acid) 信息核糖核酸
MTT (methylthiazolyl tetrazolium)
NFTs (neurofibrillary tangles) 神經纖維纏結
NF-κB (nuclear factor κ-light-chain-enhancer of activated B cells) 制核因子
NGF (nerve growth factor) 神經生長因子
NMDA (N-methyl-D-aspartate receptor) N-甲基-D-天門冬胺酸受體
NO (nitric oxide):一氧化氮
NSCs (neural stem cells) 神經幹細胞
NTU 101 (Lactobacillus paracasei subsp. paracasei NTU 101) 副乾酪乳酸桿菌 副乾酪亞種 NTU 101
OGD (oxygen-glucose deprivation) 缺血性氧糖剝奪症
PI3K (phosphoinositide-3-kinase) 磷酸肌醇 3- 激酶
PPAR-α (peroxisome proliferator-activated receptor-α)
PPAR-γ (peroxisome proliferator-activated receptor-γ)
PSEN (presenilin) 早老素
RAGE (receptor for advanced glycation endproducts)
RIP (receptor-interacting protein) 接受器互動蛋白
ROS (reactive oxygen species) 活性氧原子
SOD (superoxide dismutase) 超氧化物歧化酶
TAS (total antioxidant status) 總抗氧化能力
TBARS (thiobarbituric acid reactive substances) 脂蛋白脂質過氧化分析法
TLR-2 (Toll-like receptor 2)
TLR-4 (Toll-like receptor 4)
TMP (1,1,3,3-tetramethoxypropane) 1,1,3,3- 四乙氧基丙烷
TNF-α (tumor necrosisfactor-α) 腫瘤壞死因子-α
III
TRAF 2 (TNF receptor- associated factor 2) 腫瘤壞死因子接受器因子 2
TRADD (TNF receptor-associated death domain protein) 導腫瘤壞死因子接受 器之相關死亡蛋白
VEGF (vascular endothelial growth factor) 血管內皮生長因子
V-LDL (very low density lipoprotein) 非常低密度脂蛋白
IV
中文摘要
阿茲海默症 (Alzheimer’s disease, AD) 是為最常見腦神經退化性疾病,病徵 出現於大腦皮質 (crotex) 和海馬迴 (hippocampus)。類澱粉樣蛋白質 (amyloid β- peptide, Aβ) 被證實會於腦部中沉積造成氧化壓力與發炎反應造成記憶學習能力 的障礙。乳酸菌發酵能產生有更高生理活性的去醣基異黃酮,具有清除自由基、
提升總抗氧化力與抑制氧化發炎反應的效果。本研究以 Lactobacillus paracasei subsp. paracasei NTU 101 乳酸菌株發酵豆奶和其萃取物作為試驗物質,以腦部 連續 28 天輸注 Aβ40 建立 AD 模式大鼠探討 NTU 101 發酵豆奶與未發酵豆 奶對 AD 之預防改善效果。以 Morris 參考記憶試驗與空間探測試驗中顯示 NTU 101 發酵豆奶酒精萃取物組 (ethanol extracts of NTU 101-fermented soy milk group, L-SmE group) 有改善記憶學習能力。在功效成分比較試驗中的參考記憶 試驗與空間探測試驗中顯示 genistein 效果最好其次為 daidzein 與 equol。
Genistein 可顯著降低 cortex 中 TBARS 含量。前發炎因子蛋白質的表現試驗中,
NTU 101 發酵豆奶的改善效果優於未發酵豆奶。純物質組方面則均有顯著改善 效果。AD 相關因子試驗與免疫組織染色之結果可以看出,Aβ 組多量的 AD 相 關因子表現 NTU 101 發酵豆奶有改善之成效。Astrocyte 及 microglia 活化因子 結果顯示,NTU 101 發酵豆奶在大腦皮質有極顯著的改善 RAGE 與 TLR-2 蛋 白表現的效果,但是在海馬迴組織中卻沒有此趨勢。純物質餵食組都具有極顯著 的降低 RAGE 蛋白表現的效果,TLR-2 蛋白表現在皮質是 Gse 與 E 組效果 最佳,海馬迴組織中則是 Dze 組有最好的效果。上述實驗可以看出腦部輸注 Aβ40 會造成 astrocyte 及 microglia 活化因子的增量引起前發炎因子與相關因 子的大量表現,導致記憶學習能力失調。而發酵豆奶的乙醇萃取物可以有效降低 Astrocyte 及 microglia 活化因子、前發炎因子與相關因子來改善記憶學習能力,
原因來自於萃取物中含有大量去醣基異黃酮 genistein 與 equol 的功效成分,又 以 genistein 影響為最佳。
關鍵字:阿茲海默症、類澱粉樣蛋白、乳酸菌發酵豆奶、去醣基異黃酮
V
Abstract
Alzheimer’s disease, AD is one of the most common degenerative brain disease.
Research had be certain amyloid β-peptide, Aβ deposit in brain will cause oxidative stress and inflammation which result in learning ability and memory disorders. The aglycone isoflavones produced by fermented one has higher physiological activity than unfermented one. Due to these advantages in these benefits, our goal is to test whether Lactobacillus paracasei subsp. paracasei NTU 101 fermanted soy-milk will have effects on preventing human from AD or not by conducting experiments on NTU 101- fermentated soy milk and its’ extractions. To do these experiment, we continuously injected Aβ40 through out 28 days to built AD pattern with rats and observe the effect of NTU 101 fermanted soy-milk and soy milk on AD prevention. The Morris water maze test result shows that NTU 101-fermented soy milk (L- has better learning ability.
Genistein is the best, followed daidzein and equol comes second and third. Genistein can significantly lower the reading of TBARS in cortex. The test result shows NTU 101 fermented soy-milk has significant improvement. The results of immunohistochemistry staining can realize, the Aβ40 protein deposition Aβ group slices on a significant excess than Vh group. In the result of astrocyte and microglia, it shows that NTU 101 fermented soy-milk decrease TLR-2 and RAGE protein. Form the result above, Aβ40 infusion in brain for 28 days continuouse could cause astrocyte and microglia activator incremental, performance attracted a large number of pro-inflammatory factors and the related factors of AD, which lead to memory learning ability disordered. The ethanol extract of fermented soy milk can reduce astrocyte and microglia-activating factor, pro- inflammatory factors and the related factors of AD to improve memory learning ability.
Reason comes from the extract containing a large amount of effective ingredients, the aglycone isoflavones genistein and equol. And the best improving effect comes from the impact of genistein.
Keyword: Alzheimer’s disease, amyloid β-peptide, Lactobacillus fermanted soy-milk, aglycone isoflavones
VI 目錄
縮寫表---I 中文摘要---IV 英文摘要---V 目錄---VI 圖目錄---XI 表目錄---XIII
前言---01
第一章、 文獻回顧---04
第一節、 阿茲海默症之簡介---04
一、 阿茲海默症之病徵---04
二、 阿茲海默症之形成機制---06
三、 阿茲海默症之治療方式---15
第二節、 副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶簡介---18
一、 益生菌及乳酸菌之簡介---18
二、 副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶之簡介--- ---19
第三節、 乳酸菌發酵豆奶與阿茲海默症之預防潛力---24
一、 去醣基異黃酮與雌性激素---24
二、 去醣基異黃酮抑制類澱粉樣蛋白質剪切路徑---25
三、 去醣基異黃酮活化神經傳導路徑---26
四、 去醣基異黃酮對氧化壓力與發炎反應之抑制效果---26
五、 去醣基異黃酮對 astrocyte 及 microglia 活化之抑制效果--- ---27
第二章、 研究動機與目的---29
第一節、 研究動機與目的---29
第二節、 研究架構---30
VII
第三章、 實驗材料與方法---32
第一節、 實驗材料---32
一、 實驗藥劑---32
二、 實驗儀器---34
第二節、 實驗方法---36
一、 試驗物質製備---36
二、 動物行為試驗---36
三、 生理生化分析---45
四、 統計分析---48
第四章、 結果---49
第一節、 NTU 101 乳酸菌發酵豆奶與未發酵豆奶對預防阿茲海默症的效 果---49
一、Morris 水迷宮試驗分析其記憶行為能力之影響---49
二、脂質過氧化物 TBARS 含量之影響---52
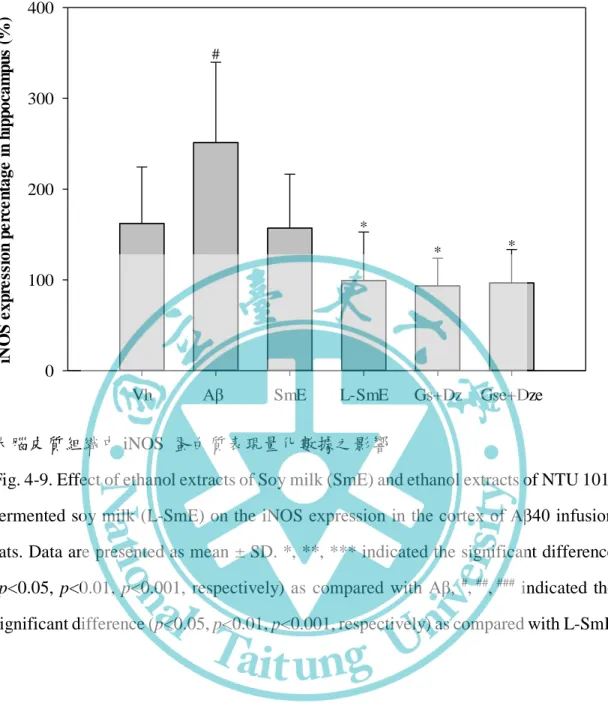
三、前發炎因子 NF-κB、iNOS 與 COX-2 蛋白表現之影響---55
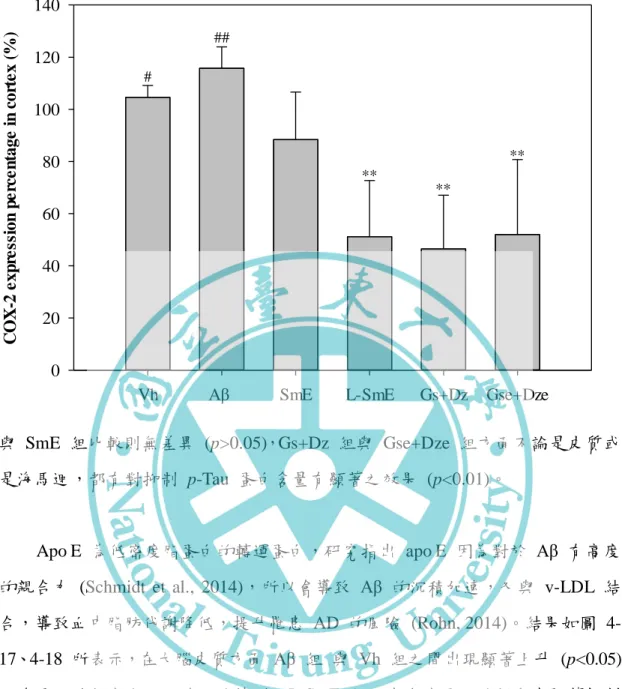
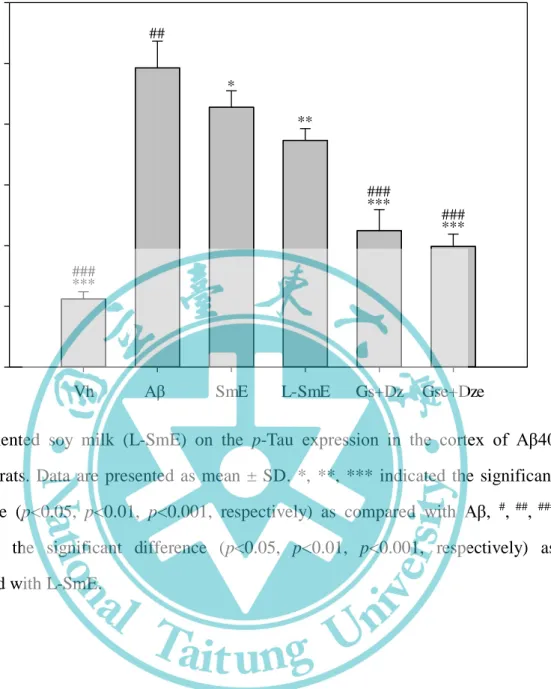
四、阿茲海默症相關因子 apo E 與 p-Tau 的蛋白表現---62
五、阿茲海默症相關因子 Aβ40 在組織間蛋白沉積量之表現---- ---71
六、Astrocyte 及 microglia 活化因子 TLR-2 與 RAGE 之蛋白 表現---71
第二節、 NTU 101 乳酸菌發酵豆奶中預防阿茲海默症主要功效成分之探 討---75
一、Morris 水迷宮試驗分析其記憶行為能力之影響---75
二、脂質過氧化物 TBARS 含量之影響---79
三、前發炎因子 NF-κB、TNF-α、IL-1β 與 IL-6 蛋白表現之影 響---79
四、阿茲海默症相關因子 Aβ40 在組織間蛋白沉積量之表現---- ---92
VIII
五、Astrocyte 及 microglia 活化因子 TLR-2 與 RAGE 之蛋白
表現---92
第五章、 討論---101
第一節、 乳酸菌發酵豆奶對腦部輸注 Aβ40 誘發阿茲海默症大鼠記憶學 習能力之影響---101
一、發酵豆奶與未發酵豆奶之比較---101
二、純物質 genistein、daidzein 和 equol 之比較---102
第二節、 乳酸菌發酵豆奶對腦部輸注 Aβ40 誘發阿茲海默症大鼠之氧化 壓力之改善效果---103
一、發酵豆奶與未發酵豆奶之比較---104
二、純物質 genistein、daidzein 和 equol 之比較---104
第三節、 乳酸菌發酵豆奶對腦部輸注 Aβ40 誘發阿茲海默症大鼠之前發 炎因子之改善效果---105
一、發酵豆奶與未發酵豆奶之比較---105
二、純物質 genistein、daidzein 和 equol 之比較---106
第四節、 乳酸菌發酵豆奶對腦部輸注 Aβ40 誘發阿茲海默症大鼠之 AD 相關因子之改善效果---106
一、發酵豆奶與未發酵豆奶之比較---106
二、純物質 genistein、daidzein 和 equol 之比較---107
第五節、 乳 酸 菌 發 酵 豆 奶 對 腦 部 輸 注 Aβ40 誘 發 阿 茲 海 默 症 大 鼠 之 Astrocyte 及 microglia 活化因子之改善效果---108
一、發酵豆奶與未發酵豆奶之比較---108
二、純物質 genistein、daidzein 和 equol 之比較---108
第六章、 結論---113
第一節、 抑制氧化發炎能力---113
第二節、 清除 AD 相關因子能力---113
第三節、 改善記憶學習能力---113
第七章、 參考文獻---117
IX 圖目錄
圖 1-1、類澱粉樣前驅蛋白質兩種路徑之酶切---08
圖 1-2、Tau 蛋白之結構與功能---10
圖 1-3、Aβ 誘發之氧化壓力反應路徑機制圖---12
圖 1-4、發炎反應路徑示意圖---14
圖 1-5、異黃酮 genistin、genistein、daidzin、daidzein 與 equol 的結構式---21
圖 2-1、本研究之研究架構---31
圖 3-1、ALZET 腦部輸注幫浦---41
圖 3-1、試驗物質改善阿茲海默症大鼠之記憶學習試驗日程表---42
圖 3-2、水迷宮裝置與分區域示意圖---44
圖 4-1、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠於 參考記憶試驗中找尋平台時間記憶學習能力---50
圖 4-2、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠於 probe test 試驗中於目標象限之游泳時間記憶學習能力之影響---51
圖 4-3、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠於 工作記憶試驗中記憶學習能力之影響---53
圖 4-4、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之 大腦皮質與海馬迴中 TBARS 含量之影響---54
圖 4-5、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之 大腦皮質組織中前發炎因子蛋白 Western blotting 圖---56
圖 4-6、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之 海馬迴組織中前發炎因子蛋白 Western blotting 圖---57
圖 4-7、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之 大腦皮質組織中 NF-κB 蛋白質表現量化數據之影響---58
圖 4-8、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之 海馬迴組織中 NF-κB 蛋白質表現量化數據之影響---59 圖 4-9、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠之
X
大腦皮質組織中 iNOS 蛋白質表現量化數據之影響---60 圖 4-10、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中 iNOS 蛋白質表現量化數據之影響---61 圖 4-11、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之大腦皮質組織中 COX-2 蛋白質表現量化數據之影響---63 圖 4-12、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中 COX-2 蛋白質表現量化數據之影響---64 圖 4-13、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之大腦皮質組織阿茲海默症中相關因子 Western blotting 圖---65 圖 4-14、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中阿茲海默症中相關因子 Western blotting 圖---66 圖 4-15、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之大腦皮質組織中 p-Tau 蛋白質表現量化數據之影響---67 圖 4-16、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中 p-Tau 蛋白質表現量化數據之影響---68 圖 4-17、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之大腦皮質組織中 apo E 蛋白質表現量化數據之影響---69 圖 4-18、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中 apo E 蛋白質表現量化數據之影響---70 圖 4-19、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對於腦部輸注 Aβ40 大 鼠之大腦皮質組織中 Aβ40 蛋白沉積量之影響 (100X)---72 圖 4-20、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之大腦皮質中抑制 astrocyte 及 microglia 活化因子蛋白 Western blotting 圖---- ---73 圖 4-21、豆奶乙醇萃取物與乳酸菌發酵豆奶乙醇萃取物對腦部輸注 Aβ40 大鼠 之海馬迴組織中抑制 astrocyte 及 microglia 活化因子蛋白 Western blotting 圖- ---74 圖 4-22、Genistein、deidzein 和 equol 對腦部輸注 Aβ40 大鼠於參考記憶試驗
XI
中找尋平台時間記憶學習能力---76 圖 4-23、Genistein、deidzein 和 equol 對腦部輸注 Aβ40 大鼠於 probe test 試驗 中於目標象限之游泳時間記憶學習能力之影響---77 圖 4-24、Genistein、deidzein 和 equol 對腦部輸注 Aβ40 大鼠於工作記憶試驗 中記憶學習能力之影響---78 圖 4-25、Genistein、deidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質與海 馬迴中 TBARS 含量之影響---80 圖 4-26、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中阿茲海默症前發炎因子 Western blotting 圖---81 圖 4-27、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 前發炎因子 Western blotting 圖---82 圖 4-28、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 NF-κB 蛋白質表現量化數據之影響---83 圖 4-29、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 NF-κB 蛋白質表現量化數據之影響---84 圖 4-30、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 TNF-α 蛋白質表現量化數據之影響---86 圖 4-31、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 TNF-α 蛋白質表現量化數據之影響---87 圖 4-32、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 IL-1β 蛋白質表現量化數據之影響---88 圖 4-33、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 IL-1β 蛋白質表現量化數據之影響---89 圖 4-34、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 IL-6 蛋白質表現量化數據之影響---90 圖 4-35、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 IL-6 蛋白質表現量化數據之影響---91 圖 4-36、Genistein、daidzein 和 equol 對於腦部輸注 Aβ40 大鼠之大腦皮質組
XII
織中 Aβ40 蛋白沉積量之影響 (100X)---93
圖 4-37、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中抑制 astrocyte 及 microglia 活化因子蛋白 Western blotting 圖---94
圖 4-38、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 抑制 astrocyte 及 microglia 活化因子蛋白 Western blotting 圖---95
圖 4-39、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 RAGE 蛋白質表現量化數據之影響---96
圖 4-40、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 RAGE 蛋白質表現量化數據之影響---97
圖 4-41、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之大腦皮質組織 中 TLR-2 蛋白質表現量化數據之影響---98
圖 4-42、Genistein、daidzein 和 equol 對腦部輸注 Aβ40 大鼠之海馬迴組織中 TLR-2 蛋白質表現量化數據之影響---99
圖 5-1 發酵豆奶抑制 AD 機制圖---110
圖 5-2 Genistein 抑制 AD 機制圖---111
圖 5-3 Equol 抑制 AD 機制圖---112
XIII
表目錄
表一、L-SmE 與 SmE 改善 Aβ40 腦部輸注之阿茲海默症大鼠記憶學習能力試 驗之動物分組---38 表二、Genistein、deidzein 和 equol 改善 Aβ40 腦部輸注之阿茲海默症大鼠記憶 學習能力試驗之動物分組---39 表三、發酵豆奶與未發酵豆奶對於阿茲海默症預防之效果---115 表四、Genistein、daidzein 與 equol 對於阿茲海默症預防之效果---116
1
前言
2015 年 3 月根據世界衛生組織 (World Health Organization, WHO) 指出全 球有 4750 萬名失智症 (Dementia) 患者,阿茲海默症 (Alzheimer’s disease, AD) 近年來逐漸成為全球不容忽視的疾病 (Alzheimer’s Disease International, 2015),
AD 會逐漸造成記憶損害和學習障礙,進而影響日常活動和與他人溝通,最終影 響長期記憶能力 (Jansen et al., 2015)。
影響 AD 的危險因子包括乙醯膽鹼 (acetylcholine, ACh) 和乙醯膽鹼酯酶 (acetylcholinesterase, AChE) 的 形 成 與 代 謝 異 常 (Amat-Ur-Rasool and Ahmed, 2015);微管關聯蛋白 (tau) 的不正常磷酸化 (Yang et al., 2015);類澱粉樣蛋白質 (amyloid β-peptide, Aβ) 導致神經原纖維纏結 (neurofibrillary tangles, NBT) 和老 年斑塊 (senile plaque) 的沉積 (Roberts et al., 2012);apolipoprotein E (apo E) 加 速 Aβ 的沉積 (Monsell et al., 2015; Schmidt et al., 2014);鋁離子 (aluminum, Al) 在腦部的大量推積 (Thomas et al., 2015)。
AD 也 會 造 成 氧 化 與發 炎 反 應, 神經 膠 細 胞 (astrocyte) 及 微小 膠細胞 (microglia) 會釋出過氧化物攻擊神經細胞造成其死亡 (Dumont and Beal, 2011)。
自由基的產生使粒腺體 (mitochondria) 出現功能障礙最後導致細胞死亡 (Sinha et al., 2013)。影響發炎物質如 tumor necrosis factor-α (TNF-α)、nuclear factor κ- light-chain-enhancer of activated B cells (NF-κB) 或是 interleukin-1β (IL-1β) 等。
TNF-α 會活化 NF-κB 轉譯出 cyclooxygenase-2 (COX-2) 與 inducible nitric oxide synthase (iNOS) (Lin et al., 2013),COX-2 能擴張血管增加血液流量並活化巨噬細 胞 (macrophages) 充斥於發炎組織 (Medeiros et al., 2010)。iNOS 誘發產生大量 的 nitric oxide (NO) 容易與氧結合產生大量自由基攻擊細胞組織而造成損傷 (Song et al., 2014)。IL-1β 是發炎和免疫反應主要調節細胞因子,透過誘導 COX- 2 的產生引起發炎反應 (Kitazawa et al., 2008)。
AD 目前治療方式為,調控膽鹼神經系統 (cholinergic) 的 AChE 抑制劑,
和 N-methyl-D-aspartate receptor (NMDA) 受體拮抗劑,阻止 AD 病程的方式包 含調控 Aβ 蛋白質剪切、抑制 microglia 與 astrocyte 的活化、抑制 Aβ 造成之
2 氧化發炎和穩定鈣離子的流入等等。
副 乾 酪 乳 酸 桿 菌 副 乾 酪 亞 種 NTU 101 (Lactobacillus paracasei subsp.
paracasei NTU 101) 菌株為國立臺灣大學潘子明教授研究室,自嬰兒糞便中分離 之乳酸菌菌株 (Lin et al., 2004)。在發酵豆奶的過程中會將含醣基大豆異黃酮 (glucoside isoflavones) 代謝成具有更高生理活性的物質不含醣基大豆異黃酮 (aglycone isoflavones),更容易讓人體消化吸收。
副乾酪乳酸桿菌 NTU 101 有減少肝臟中與血液中的膽固醇之功效 (Chiu et al. 2006),增加 SOD 和降低脂質過氧化指數,防止血脂引起的氧化損傷與動脈 粥狀硬化 (Tsai et al., 2009)。NTU 101 可提升的樹突細胞 (dendritic cell) 的抗原 呈現能力,加強自然殺手細胞 (natural killer cell) 的胞殺作用 (Tsai et al., 2008),
和 調 節 interleukin-10 (IL-10) 與 interleukin-12 (IL-12) 達 到 提 升 免 疫 能 力 (Chiang and Pan 2012) 等。
大豆異黃酮本身也具有許多的功效,genistein 能與雌性激素之受體結合而 同時也能與之拮抗受體結合 (Bagheri et al., 2011),改善荷爾蒙的相關症狀 (Bang et al. 2004)。通過刺激 ERK1/2 和 PI3K 信號調節軸突刺激其生長,使已 1- methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) 誘發神經變性的小鼠,達到保 護其多巴胺細胞效果 (Zhao et al., 2015)。透過改善突 synaptic plasticity 和增加 cholinergic 神經系統,達到改善學習和記憶 (Wei et al., 2015)。
Aglycone isoflavones 當中的 genistein 由於其作用與構型類似雌性激素,固 有植物的雌性激素之稱,有神經退化性的疾病具有適性相當良好的改善效果,所 以被拿來作荷爾蒙補充療法 (hormone replacement therapy, HRT) 的替代品,而又 不會造成細胞無限制的增生導致罹患癌症的風險增高 (National Institutes of Health, 2005)。Genistein 具有神經保護功能對於神經母細胞瘤 (Neuroblastoma, NB) 的研究,結果顯示 genistein 能有效降低 P53 與 CHD5 等致癌基因的表現,
並抑制其血管的增生來抑制神經瘤的產生 (Li et al., 2012)。Daidzein 也具有神經 保護功能,daidzein 有促進軸突生長的功能,並能使先天性缺乏脂質載體蛋白 (apo E) 的小鼠刺激產生 apo E 之功效,所以對於脂質大量堆積所造成的中風 (stroke) 具有改善的功效,並且說明長期的食用 daidzein 對於慢性的神經疾病有
3 保護作用 (Kim et al., 2015)。
還有如抑制 Aβ 之剪切路徑,使 APP 無法經由 amyloidogenic 路徑剪切成 Aβ,阻止其產生類澱粉斑塊產生堆積 (Ahmad et al., 2015);活化神經傳導路徑,
提升 ACh 的濃度或降低 AChE 的含量,阻止 NMDA 受體打開離子通道,使 神經元部會受到過多鈣離子傷害 (Bagheri et al., 2012);也能減緩 astrocyte 及 microglia 釋出的氧化發炎物質,提升 superoxide dismutase (SOD)、catalase (CAT) 等抗氧化酵素,或降低 TNF-α 和 IL-1β 等前發炎因子的表現,以達到預防或抑 制 AD 的功效 (Bagheri et al., 2011)。
4
第一章、文獻回顧
第一節、阿茲海默症之簡介
阿茲海默症 (Alzheimer’s disease, AD) 於 1906 年,德國的神經病理學家 Alois Alzheimer 發現並首次記錄了阿茲海默症患者腦部的微觀變化,發病的病 患通常會出現記憶能力衰退、心智混亂退化最後喪失行為能力等症狀,同時於病 患生前腦部中發現有班塊 (plaques) 和糾結 (tangles) 等特徵 (Alzheimer, 1907)。
隨著人類平均壽命的延長,慢性疾病的發生頻率也越來越普及,根據世界衛 生組織 (World Health Organization, WHO) 於 2015 年 3 月時指出全球有 4750 萬名失智症 (dementia) 患者,預計這個數字到 2050 年將增長到 1.315 億 (Alzheimer’s Disease International, 2015),台灣衛生福利部公告民國 103 年底罹 患 dementia 人口統計資料估算:台灣 65 歲以上老年人口罹患 dementia 佔 8.09%,約有 227,137 人並且有逐年成長的趨勢,預計於民國 150 年 dementia 人口將達 93 萬人而其中又以 AD 的病患占多數。
AD 近年來逐漸成為全球不容忽視的疾病,然而高油脂、高鹽、高糖的飲食 所引發的現代流行病氾濫又近一步增長了 AD 的發生 (Alzheimer’s Disease International, 2015),AD 儼然成為最常見腦神經退化性疾病 (Zhang et al., 2015),
在 AD 的病患發病的過程中會逐漸的造成記憶損害和學習障礙,進而影響日常 活動和與他人溝通之能力,病患腦部會首先發現神經元功能障礙;皮質和海馬迴 的退化,最終影響長期記憶能力 (Jansen et al., 2015)。
一、阿茲海默症之病徵 (一) 阿茲海默症之形成原因 1. 原發性阿茲海默症
原發性阿茲海默症往往跟基因遺傳有著密不可分的關係,過去研究顯示早老 素 (presenilin, PSEN) 基因和 (apolipoprotein E, apo E) 基因是 AD 的重要因素 (Heilig et al., 2010; Monsell et al., 2015),而早老素基因轉譯出來的兩個蛋白 PSEN I 和 PSEN II 能組合成與 Aβ 剪切相關之酵素 γ-secretase 會加速 Aβ 的形成
5
(Heilig et al., 2010),而 apo E 基因轉譯的四個蛋白中,其中 apo E4 蛋白對於 Aβ 有高度的親和性,能加速其沉積 (Hyman and Holtzman, 2015)。
2. 偶發性阿茲海默症
相較於原發性阿茲海默症,大部分阿茲海默症病患並沒有體染色體遺傳疾病,
這些病徵則稱為偶發性阿茲海默症。而 AD 的發生複雜而多元,因此產生了許 多假說,包含乙醯膽鹼 (acetylcholine, ACh) 形成異常或含量不足與乙醯膽鹼酯 酶 (acetylcholinesterase, AChE) 的過量表現與代謝異常,導致神經傳遞訊號中斷 (Amat-Ur-Rasool and Ahmed, 2015);微管關聯蛋白 (tau) 的不正常磷酸化,導致 微管 (microtube) 瓦解,因而神經傳遞物質無法順利傳遞與降解 (Yang et al., 2015);以及最被學界所接受的類澱粉樣蛋白質 (amyloid β-peptide, Aβ) 假說,許 多研究表明 Aβ 代謝異常與沉積導致神經原纖維纏結 (neurofibrillary tangles, NBT) 和老年斑塊 (senile plaque) 的沉積是影響 AD 發病的主要原因 (Roberts et al. 2012);另外 apo E4 與 Aβ 的高度親和力,加速了 Aβ 的沉積,導致老年 斑塊的加速形成 (Monsell et al., 2015; Schmidt et al., 2014) 亦或是與極低密度脂 蛋白 (very-low-density lipoprotein, V-LDL) 結合,而降低血中脂肪代謝,提升罹 患 AD 風險 (Divekar et al., 2014);最後鋁離子 (aluminum, Al) 近代被研究出具 有穿透血腦障壁 (blood-brain barrier) 之能力,在腦部的大量推積也被認為是引 發或加重 AD 的原因之一 (Thomas et al., 2015)。
(二) 阿茲海默症之臨床病徵 1. 阿茲海默症之臨床病徵
台灣失智症協會將 dementia 分成兩個種類,退化性失智症和血管性失智症,
退化性失智症是一種不正常的老化現象,腦部出現退化的現現,包含額頁、顳頁 型失智症 (Frontotemporal lobe degeneration) 額頁及顳頁的退化,造成人格變化 和行為控制力的喪失 (Luzzi et al., 2015);路易氏體失智症 (Dementia with Lewy Bodies) 則 是 因 為 路 易 氏 體 (Lewy Bodies) 的 退 化 而 造 成 認 知 功 能 的 障 礙 (Raggi et al., 2015);血管性失智症 (Vascular Dementia) 是因為腦血管的病變,造
6
成腦部血液循環不良,導致腦細胞死亡、智力減退等現象 (Ishii and Iadecola, 2015)。
AD 是退化性與血管性並存的一種病例,從發病後觀察到的臨床病徵與時間 分成了三個時期 (湯,2008)。
(1). 初期,病患可能會開始出現記憶力減退、暫時性語言表達困難、對時間產生 混淆的現象。
(2). 中期,變得健忘、反覆詢問同樣的問題,生理需求開始依賴他人的協助,無 法單獨生活,語言表達越來越困難,出現遊走的、失去方向感的問題。
(3). 晚期,病患已經完全喪失認知與記憶能力,已無法獨立生活,需要仰賴他人 照料。
2. 阿茲海默症之組織病理
另外從微觀的方面來說,AD 患者的腦部會出現 NBT 和 senile plaque 的沉 積等特殊的病理特徵 (Hohsfield and Humpel, 2015),NBT 形成是由 tau 蛋白過 度磷酸化,導致 microtube 瓦解、纖維聚集成團,導致神經傳遞物質無法傳遞 (Bloom, 2014),而 senile plaque 主要成因是因為不可溶 Aβ 在細胞外大量沉積,
而誘使神經細胞中星狀細胞 (astrocyte) 及微小膠細胞 (microglia) 釋放發炎物 質,最後免疫複合物和 Aβ 於神經纖維外集結成團,阻絕神經傳遞物質的傳遞是 加重 AD 的原因之一 (Hohsfield and Humpel, 2015)。
二、 阿茲海默症之形成機制
(一) 阿茲海默症之形成相關因子
1. 類澱粉樣前驅蛋白質 (amyloid precursor protein, APP)
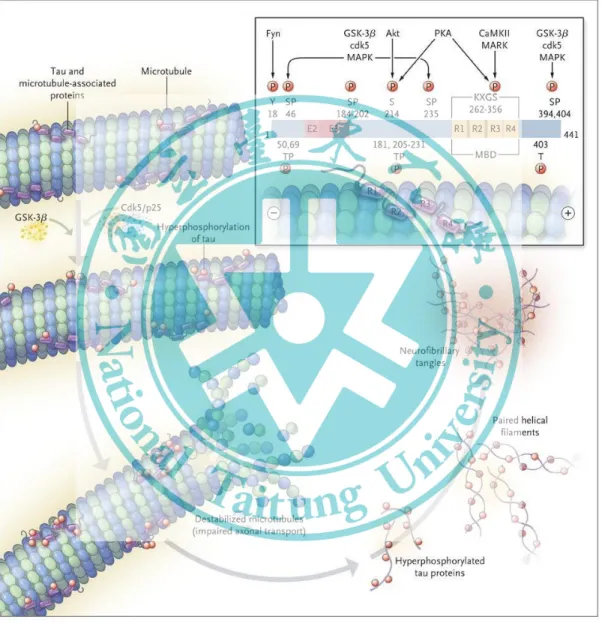
APP 蛋白為一種跨膜蛋白,常見於神經元突觸上,通常由高基氏體 (golgi apparatus) 分泌至細胞膜上,一般 APP 經兩種途徑被水解 non-amyloidogenic 與 amyloidogenic ,non-amyloidogenic 路徑會經 α-secretase 裁切 APP 產 生 secreted amyloid precursor α (sAPP-α) 和 含 C 端 83 個 氨 基 酸 的 carboxy- terminal fragment 83 (CTF 83) , 再 由 γ-secretase 剪 切 則 會 形 成 p3 與 APP intracellular domain (ACID)。
7
Amyloidogenic 路徑則 會 經 β-secretase 裁 切 APP 產生 secreted amyloid precursor β (sAPPβ) 與 carboxy-terminal fragment 99 (CTF 99),CTF 99 再經由 γ- secretase 剪切成為 Aβ 與 ACID (圖 1-1)。
APP 蛋白裁切成的 sAPP-α 和 sAPP-β 其實兩個片段差異並不大,僅於 C 端少了 16 個胺基酸片段都有保護生長因子的特性其功能,包含減少神經元損傷 和穩定細胞靜止的膜電位,只是相較於sAPPα,sAPPβ 通常表現出較低的保護效 果 (Vella and Cappai 2012),但過去研究指出 sAPPβ 能增加胚胎幹細胞與神經元 幹細胞的生長能力 (Baratchi et al., 2012)。
而過去研究證實由 γ-secretase 剪切後殘存的 ACID 片段具有使神經元再生 的作用 (Li et al., 2015),而在缺乏 ACID 的小鼠上也可以觀察到突觸可塑性 (synaptic plasticity) 下降;學習和記憶的損傷的情況 (Klevanski et al., 2015)。
2. 類澱粉樣蛋白質 (amyloid β-peptide, Aβ)
CTF 99 經 γ-secretase 切割後產生的片段,其中 90% 為 Aβ40,約 10% 為 Aβ42,雖然 Aβ42 一般在人體中有更高的毒性但比例相對較少 (Igarashi et al.
2015),一般在人體血漿及腦脊髓液中就可偵測到少量之 Aβ,為一種正常的代謝 現象,然而在 AD 患者的腦脊髓液則表現較高量之 Aβ 其清除能力也較正常人 低 (Sun et al., 2015)。可溶的 Aβ 較不具毒性,必須經聚合形成 Aβ fibrils 後才 經由破壞細胞內離子平衡與引發氧化發炎毒殺神經細胞。由上述可知 Aβ 代謝 異常、沉積將導致 NBT 和 senile plaque 的沉積 (Lloret et al., 2015),而其過程 會誘使 astrocyte 及 microglia 釋放發炎物質例如 tumor necrosis factor-α (TNF- α) 、 nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) 或 是 interleukin-1β (IL-1β) 等 (Dal Pra et al., 2015)。
3. 過度磷酸化微管關聯接蛋白 (p-tau)
神經纖維纏結 (neurofibrillary tangles, NBT) 的成因可能為微管連接蛋白,
tau 蛋白過度磷酸化。正常 tau 蛋白為一種多功能的蛋白質,一般在體內有穩定
8 圖 1-1、類澱粉樣前驅蛋白質兩種路徑之酶切
Fig. 1-1. Proteolytic processing of amyloid precusor protein (APP) by two pathway (David et al., 2012).
9
微管 (microtube) 或與 microtube 相似的作用,當 tau 蛋白於正常狀態下磷酸化 時,本身會形成 microtube 或其部分 (Qu et al., 2011),但是 tau 蛋白的異常磷 酸化時會形成雙股螺旋纖維,與 Aβ 及 apo E4 組成 NBT (Dai et al., 2015)。其 不正常磷酸化導致 microtube 組裝能力降低甚至瓦解,繼而損害軸突運輸 (axoplasmic transport),導致神經傳遞物質無法順利降解,最終使神經元細胞退化 死亡 (Wu et al., 2015) (圖 1-2)。
4. 脂質轉運蛋白 E4 基因 (apolipoprotein E4, apo E4)
位於第 19 對染色體上,基因型有 apo E2、apo E3、apo E4 等 3 種為血清 中主要脂質轉運蛋白之一,轉譯出的蛋白 apo E2、apo E3 為轉運高密度脂蛋白 的蛋白,apo E4 則為低密度脂蛋白的轉運蛋白。而有研究指出 apo E4 因為對於 Aβ 有高度的親合力 (Schmidt et al., 2014),所以會導致 Aβ 的沉積加速,又與 v-LDLs 結合,導致血中脂肪代謝降低,提升罹患 AD 的風險 (Rohn, 2014)。
5. 早老素 (presenilin, PSEN)
PSEN 是一種橫跨細胞膜的蛋白質酵素,是影響大腦內 Aβ 異常沉積的原 因之一,由於在第 21 對染色體上 APP 基因突變或是位在第 14 對染色體上的 老化素 I (presenilin I, PSEN I) 基因突變,或位於第 1 對染色體上老化素 II (presenilin II, PSEN II) 基因產生突變,二者轉譯出來的兩個蛋白 PSEN I 和 PSEN II 能組合成與 Aβ 剪切相關的酵素 γ-secretase,使得 Aβ 的生合成增加 (Wang et al., 2011)。
6. 鋁離子 (aluminum, Al)
許多研究顯示環境當中的 Al 與 AD 的發病有一定的關聯性,早期因為懷 疑 Al 無法通過血腦障壁 (blood-brain barrier) 而把其危險性屏除,但近代被研 究出 AD 患者腦部中發現有 Al 的大量推積的情況,打破無法通過 blood-brain barrier 一說,又過去研究表明 AD 患者腦脊髓液和血清中 Al 的含量有顯著的 上升,特別是於血清中 (Virk and Eslick, 2015),因此 Al 也逐漸被認為是引發或
10 圖 1-2、Tau 蛋白之結構與功能
Fig. 1-2. Tau protein structure and function. (Querfurth and Laferla, 2010).
11 加重 AD 的相關原因之一。
(二) 阿茲海默症誘導之氧化壓力與發炎反應 1. 氧化壓力
大部分老年失智病因的中間產物如:Aβ 沉澱、鋁中毒、免疫作用等,常造 成蛋白質變性、脂質過氧化物、DNA、mitochondria 失去功能使神經老化病變而 失去作用,造成老年失智的病症。
Aβ 具有誘導氧化壓力的危險性,包括過氧化物和脂質過氧化物在神經元中 的增加;astrocyte 及 microglia 也會因 Aβ fibrils 的結合而釋出過氧化物質,如 一氧化氮 (nitric oxide, NO)、超氧陰離子 (O2-) 等自由基攻擊神經細胞造成其死 亡 (Dumont and Beal, 2011) (圖 1-3)。Aβ 也能直接於 mitochondria 誘導自由基 的產生,使的 mitochondria 出現功能障礙最後導致細胞死亡 (Sinha et al., 2013)。
Aβ 所造成過多的自由基也會影響不飽和脂肪酸,如細胞膜的雙層磷脂質構造,
不飽和脂肪酸受自由基攻擊後形成脂質過氧化物,以自催化特性產生更多的自由 基,蛋白質和 DNA 被自由基攻擊,導致細胞內的高度損傷 (Zhu et al., 2007)。
2. 發炎反應
Astrocyte 及 microglia 的活化會釋出致發炎物質,所以此二細胞的活化與 否,也影響著發炎反應的強弱,astrocyte 及 microglia 會被 receptor for advanced glycation endproducts (RAGE) 活化,RAGE 為一種跨膜受體的免疫球蛋白可與 醣基化終點產物 advanced glycation endproducts (AGEs) 或 Aβ 結合 (Walker et al., 2015),進而產生 NF-κB 進入細胞核中,接著藉由回饋作用產生大量的 NF- κB 引起發炎反應 (Ben Haim et al., 2015),另外 toil-like receptor 2 (TLR-2) 常被 發現於 astrocyte 及 microglia 的細胞表面,主要功能於辨識抗原以及引發先天 性免疫反應,在與抗原結合後會釋出 IL-6、IL-1β 與 NF-κB 等發炎物質 (Ebert et al., 2015)。
Astrocyte 及 microglia 釋出致發炎物質如先前所提及 TNF-α、NF-κB 或是
12 圖 1-3、Aβ 誘發之氧化壓力反應路徑機制圖
Fig. 1-3. The mechanism of Aβ indused oxidative stress pathway (Pan et al., 2010).
13
IL-1β 等,攻擊神經細胞造成其變性、死亡,形成老年失智 (Zhou et al., 2014a)。
腦部受到的損傷 (頭部外傷、腦缺血、基因因素、病毒感染等造成神經病變) 皆 會引起發炎反應 (Sardi et al., 2011)。調控發炎反應的路徑眾多,其中 TNF-α 與 腫瘤壞死因子接受器 (TNF receptor) 結合後會誘導腫瘤壞死因子接受器之相關 死亡蛋白 (TNF receptor-associated death domain protein, TRADD)、接受器互動蛋 白 (receptor-interacting protein, RIP) 以 及 腫瘤 壞死 因子 接 受器 因子 2 (TNF receptor- associated factor 2, TRAF 2) 活化,刺激 1κB kinase 釋放 p50 與 p65 並聚合成 NF-κB (Richman et al., 2013)。
釋出的 NF-κB 便進入細胞核與 promoter 結合進而活化相關基因,接著轉 譯出發炎反應蛋白 cyclooxygenase-2 (COX-2) 與 inducible nitric oxide synthase (iNOS) (Lin et al., 2013),COX-2 其功能在於刺激血管擴張素與組織胺以擴張血 管增加血液流量並活化巨噬細胞 (macrophages) 或其他免疫細胞充斥於發炎組 織 (Medeiros et al., 2010)。
iNOS 為一重要的細胞信號分子,在 NF-κB 活化相關基因後轉譯出 iNOS (Cui et al., 2012),而 iNOS 不需要經過鈣離子和鈣調蛋白 (calmodulin) 的催化,
本身就可以誘發產生大量的 NO,其作用在於刺激血管的平滑肌放鬆而擴張血管;
也作為神經傳遞因子,與學習及記憶有關;或是造成被腫瘤、寄生蟲等外來因素 影響之細胞停止生長、分裂或直接壞死。但 NO 本身也為極不穩定之分子容易 與氧結合而產生大量自由基,所以大量的 iNOS 產生大量的 NO,便會引起大量 自由基攻擊細胞組織,而造成損傷 (Song et al., 2014) (圖 1-4)。
另外當 macrophages 受到 TNF-α 所刺激便會釋出 IL-1β,IL-1β 是發炎和 免疫反應的主要調節細胞因子,參與產生急性期蛋白的反應,如低血壓、發燒與 血小板凝集。IL-1β 可以誘導不同類型的細胞的凋亡 (Goncalves et al., 2014),IL- 1β 也是重要的發炎反應訊號因子,可以透過誘導 COX-2 的產生引起發炎反應 (Kitazawa et al., 2008),IL-1β 與 macrophages 也能產生 interleukin-6 (IL-6),IL- 6 屬多功能之細胞激素其功能包括誘導急性期蛋白質之生合成或刺激 T 細胞的 活化與刺激 B 細胞產生抗體 (Pabois et al., 2014)。
14 圖 1-4、發炎反應路徑示意圖
Fig. 1-4. The mechanism of inflammatory pathway (Derek et al., 2011).
15 一、阿茲海默症之治療方式
治療 AD 的藥物目前市面上只有兩種類型,調控膽鹼神經系統的 AChE 抑 制劑,和 NMDA 受體的拮抗劑,但從過去研究所知 AD 的治療方式有許多有 效和直接的方法,如調控 Aβ 蛋白質剪切、調控神經修復生長因子、抑制神經系 統的氧化發炎和穩定鈣離子的流入等等。
(一) 調控膽鹼神經系統
ACh 與 AChE 影響傳遞訊息。AD 的患者腦部 ACh 的含量下降,最直接 的治療法為提升 ACh 的濃度或降低 AChE 的含量,因此使用 AChE 抑制劑,
可以提昇突觸間 ACh 的濃度,有助於增進 cholinergic 神經的作用。抑制 AChE 的作用不僅增加了 ACh 的量,同時減少類澱粉樣斑塊的產生,因此這類藥物也 具有減緩及改善 AD 認知缺失的功能 (Xiao et al., 2000)。
市面上常見上市藥物為 acetylcholinesterase inhibitor (AChEI):抑制分解酵素 的作用,以增加 ACh 之濃度,是唯一通過美國食品藥物管理局 (Food and Drug Administration, FDA) 核准使用之藥物 (Shaywitz and Shaywitz, 2000)。
(二) 調控 NMDA 受體及穩定鈣離子
NMDA 受體主要作用為麩氨酸 (glutamate) 之受體,而 glutamate 為重要之 神經傳遞訊號,但過量的話會造成鈣離子濃度不斷上升 (Hull et al., 2015),而阿 茲海默症神經元損壞與 Ca2+ 的調節變化有關,大量的 Ca2+ 內流藉由改變胞內 pH 值、活化蛋白激酶破壞胞膜、干擾 mitochondria 使細胞能量缺乏導致細胞凋 亡,造成神經損傷 (Apostolova et al., 2015)。
所以使用的治療方法有兩種為將其受體阻斷使離子通道關閉,或是 給予 Ca2+ 穩定劑或阻斷 Ca2+ 內流入阿茲海默症患者大腦神經元細胞 (Gan and Silverman, 2015)。Memantine 為市面上 NMDA 受體之拮抗劑,用於治療中度或 重度 AD (Bird, 1993)。
16 (三) 調控類澱粉樣蛋白質剪切
最被學界所接受的 Aβ 堆積假說中,許多研究表明 Aβ 代謝異常與沉積導 致 AD 發病的主要原因,反之若能抑制 Aβ 之產生則能抑制或降低其發生。如 許多研究表明,大豆異黃酮中的 genistein 具有抑制 APP 剪切成 Aβ 之能力,
而是將路徑導向 non-amyloidogenic 的剪切路徑 (Kim et al., 2002),也可以影響 γ-secretase 剪切的片段,達到抑制 Aβ 產生的目的 (Chong et al., 2001)。而另一 個大豆異黃酮素 daidzein 則是抑制 β-secretase 的活性,達到抑制 Aβ 產生 (Ahmad et al., 2015)。
可惜的是至今市面上並沒有能調控其剪切的藥物,而此方面的藥物或許是正 被迫切需要直接能治療 AD 的方式。
(四) 調控神經修復生長因子
神經生長因子是神經細胞生存和分化所必需的物質,過去研究顯示,神經幹 細胞 (neural stem cells, NSCs) 能自我修復,分化出新的細胞成為不同的神經細 胞。當移植到患病的大腦,神經幹細胞能迅速修復和分化後頂替受傷的組織 (Lee et al. 2015)。
另外過去研究指出血管內皮生長因子 (vascular endothelial growth factor, VEGF) 會促進軸突生長及細胞存活,VEGF 能引起培養的神經細胞或神經突觸 數目上及長度上的增加,VEGF 可以改變急性和慢性的神經退化過程。此作用可 能是經由 VEGF 對血管、神經細胞、神經膠細胞的作用,刺激大腦的神經細胞 生成,而可以抑制 AD 帶來的腦部神經元損傷 (Kalaria, 2002)。
(五) 抑制氧化發炎
Aβ 對於神經細胞所造成的氧化性傷害在 AD 中扮演重要的角色。過去研 究指出大豆異黃酮 genistein 對於 Aβ 在腦部所引起的發炎病症有保護的效果,
能有效降低 TNF-α 和 IL-1β 等前發炎因子的表現,或是調節 NF-κB 的訊號傳 遞,抑制發炎病症的發生 (Ma et al., 2015a),也能經由阻斷 microglia 上的 toll- like receptor 4 (TLR-4) 阻斷 NF-κB 的訊號傳遞達到抑制氧化發炎的效果 (Zhou
17
et al., 2014b),而對於氧化性傷害 genistein 能有效減少活性氧化物質 (reactive oxygen species, ROS),提升超氧化物歧化酶 (superoxide dismutase, SOD)、過氧化 氫酶 (catalase, CAT) 和穀胱甘肽 (glutathione, GSH) 等抗氧化酵素的含量。
大豆異黃酮 daidzein 也能清除 (diphenyl-1-picrylhydrazyl, DPPH) 自由基 以減輕氧化損傷 (Ahmad et al., 2015),對於腦部的發炎反應 daidzein 被研究出 也能降低 IL-1β 和 interleukin-6 (IL-6) 的蛋白,及 mRNA 表現量。
18
第二節、 副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶簡介 一、 益生菌及乳酸菌之簡介
(一)益生菌之簡介
益生菌 (probiotics),最早由 Lilly 及 Stillwell 於 1965 提出,原意是指一 原生動物產生的物質可刺激另一原生動物生長 (Lilly and Stillwell, 1965),而後 WHO 於 2001 將其定義是:凡應用至人類或其他動物,藉由改善腸內微生物相 平衡、益於宿主之活菌,可能作用為:改善腸道內菌群的平衡、增強免疫系統以 及促進營養之消化與生化作用,只要能增加其營養價值,無論是單一或混合菌株 皆可視為 probiotics 。
(二)乳酸菌之簡介
乳酸菌為兼性厭氧菌 (facultative anaerobic bacteria),乳革蘭氏陽性 (Gram positive),通常外型分為兩個種類:球菌與桿菌,並不會形成有性孢子,其定義 為能夠代謝碳水化合物,產生 50% 以上乳酸之細菌,而乳酸菌為習慣用語,並 不是分類學上正式用語 (Landete, 2016)。
1. 乳酸菌的分類
乳酸菌於分類學上已 16S rRNA 基因序列分析鑑定後,分出 15 個菌屬,
Leuconostoc 、 Streptococcus 、 Pediococcus 、 Enterococcus 、 Lactococcus 、 Carnobacterium、Vagococcus、Tetragenococcus、Atopobium、Weissella、Oenococcus、
Lactobacillus、Propionibacterium、Lactosphaera (Makarova et al., 2006)。
2. 乳酸菌的發酵轉換之益處
乳酸菌可將碳水化合物代謝成乳酸,亦會產生其他如醋酸、乙醇及二氧化碳 等多種產物,而加入食品經過發酵轉換後,會帶來不同的營養價值,乳酸菌對於 乳品中的乳糖可進行發酵,產生乳糖分解酶,故能夠改善患有先天性腸黏膜 β 半 乳 糖 苷 酶 (β-galactosidase) 缺 乏 症 所 造 成 的 乳 糖 代 謝 障 礙 (Saraniya and Jeevaratnam, 2015);乳酸菌可產生 diacetyl、acetoin、acetylaldehyde 等香味成分,
19
常被使用於乳製品中;而乳酸菌具有蛋白質分解酵素、有助於蛋白質之分解與代 謝,因而可提高食品中之營養價值;在保健功效方面乳酸菌可以藉由增加糞便中 的 β-galactosidase 活性,減少尿素酵素 (urease) 活性,改善大腸急躁症候群臨 床症狀之效果 (Galat et al., 2016);乳酸菌可活化免疫反應,因為細胞壁組成含有 胜肽聚醣 (peptidoglycan),可刺激單核球 (monocyte) 產生 TNF-α 與 IL-6 並誘 導 macrophages 形態的改變增加對抗原的吞噬能力 (Feng et al., 2015b)。
(三) 副乾酪乳酸桿菌副乾酪亞種 NTU 101 之簡介
副 乾 酪 乳 酸 桿 菌 副 乾 酪 亞 種 NTU 101 (Lactobacillus paracasei subsp.
paracasei NTU 101) 菌株為國立臺灣大學潘子明教授研究室自嬰兒糞便中分離 或得知乳酸菌菌株,自然環境下可以耐胃酸和膽汁,是目前為止唯一具多重功效 的單一菌株 (Lin et al., 2004)。NTU 101 有效地減少在肝臟中與血液中的膽固醇 (Chiu et al. 2006),另外藉由增加 SOD 和降低脂質過氧化指數,可有效地防止和 延緩高血脂引起的氧化損傷與動脈粥狀硬化 (Tsai et al., 2009)。NTU 101 可提升 的樹突細胞 (dendritic cell) 的抗原呈現能力,導致自然殺手細胞 (natural killer cell) 的胞殺作用加強 (Tsai et al., 2008),亦或是增加 macrophages 與脾臟淋巴細 胞 (splenocytes) 的分裂能力,和調節 interleukin-10 (IL-10) 與 interleukin-12 (IL- 12) 的能力,達到提升免疫與調節發炎的能力 (Chiang and Pan, 2012),NTU 101 對於幽門 (pylorus) 經加入鹽酸的乙醇 (acidified ethanol) 造成的胃損傷動物,能 提高其 SOD 的活性和前列腺素 E2 (prostaglandin E2, PGE2) 的合成,具有防止 胃損傷的效果 (Liu et al., 2009)。
二、 副乾酪乳酸桿菌副乾酪亞種 NTU 101 發酵豆奶之簡介
豆奶起源於中國,在營養價值方面含有大量的含醣基大豆異黃酮 (glucoside isoflavones) 主要包含 6 種:daidzin、genistin、glycitin、6” -O-acetyldaidzin、6”
-O-acetylgenistein、6” -O-acetylglycitein,而乳酸菌發酵的過程中,會將 glucoside isoflavones 代謝成具有更高 生理活 性的物 質不含醣基大豆 異黃酮 (aglycone isoflavones),能更容易讓人體消化吸收,主要包含 4 種:daidzein、genistein、
20
glycitein,以及 daidzein 的二次代謝產物 equol (Hati et al., 2015) 其結構如圖 1- 5 示。
(一) 副乾酪乳酸桿菌 NTU 101 發酵豆奶之功效
豆 奶 在經過 Lactobacillus paracasei subsp. paracasei NTU 101 發酵後,
glucoside isoflavones 會被轉換成具有更高生理活性的物質 aglycone isoflavones。
過去研究發現,使用 NTU 101 副乾酪乳酸桿菌所發酵的豆奶,具有免疫調 節與最大抗菌能力 (Lin and Pan, 2015)。其水萃取物能使餵食高油脂飲食之大鼠 降低主動脈周遭脂質斑塊形成,降低脂肪細胞的橫截面面積,具有抑制脂肪生成 與調控脂肪細胞的能力 (Cheng et al., 2015)。發酵的豆奶的凍乾粉末作為膳食補 充劑可以減輕因老化所誘導的骨質損失,並可能降低骨質疏鬆的危險 (Chiang and Pan, 2011)。
另外 NTU 101 副乾酪乳酸桿菌所發酵的豆奶與苦瓜能延緩高油脂血症帶 來的氧化發炎反應並顯著降低低密度脂蛋白膽固醇 (low density lipoprotein cholesterol, LDL-C) 與提高高密度脂蛋白膽固醇 (HDL-C) 的比值。也能增加 SOD 和血液的總抗氧化能力 (total antioxidant status, TAS) 的活性和降低脂質過 氧化所造成的氧化損傷,降低動脈粥狀硬化的可能性。
(二)大豆異黃酮之功效 1. 雌性激素替代治療
荷爾蒙補充療法 (hormone replacement therapy, HRT) 是醫學上幫助年長女 性緩和更年期症狀的方法,但是一直有造成罹患癌症的風險增高的疑慮,美國國 家衛生院 (National Institutes of Health, NIH) 在 2002 年的重要研究顯示,0.06%
採用 HRT 的婦女一年後在減少結腸癌以及骨質疏鬆的機率,但卻增加 0.08%
中風、肺臟血管出現血塊和乳癌的機率 (National Institutes of Health, 2002)。更在 2005 年 NIH 研究出,使用 HRT 的女性病患中有罹患乳癌的風險會升高 26%
(National Institutes of Health, 2005)。
21
圖 1-5、異黃酮 genistin、genistein、daidzin、daidzein 和 equol 的結構式 Fig. 1-5. The structural formula of the isoflavones: genistin、genistein、daidzin、
daidzein and equol. (Michio, 2011)
daidzin genistin
daidzein genistein
equol
22
大豆異黃酮中的 genistein 構型因為近似於動情激素,固有植物性雌激素,
能與雌性激素之受體結合而同時也能與之拮抗受體結合,如婦女的女性荷爾蒙太 多,異黃酮素就會與雌激素競爭,減少雌激素對乳房和子宮內膜的刺激,因而減 少這兩器官發生癌症的機會 (Bagheri et al., 2011)。過去研究指出 genistein 能改 善女性荷爾蒙的相關症狀,改善停經後潮熱、心悸、記憶衰退與骨質流失等症狀 (Bang et al., 2004)。
2. 神經保護及修復生長
神 經 生長 因子 (nerve growth factor, NGF) 通 過刺激細胞外信 號調節酶 (extracellular signal-regulated-kinases 1/2, ERK 1/2) 和 磷 酸 肌 醇 3- 激 酶 (phosphoinositide-3-kinase, PI3K) 調節神經元的分裂與分化,葛根中發現 C- 端 有醣基化的異黃酮素稱葛根素,可以通過刺 ERK 1/2 和 PI3K 信號調節軸突刺 激其生長,使已 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) 誘發神經變 性的小鼠,達到保護其多巴胺細胞與改善行為障礙的效果 (Zhao et al., 2015)。
有過去研究顯示,大豆異黃酮可以保護神經元母細胞,以保護 6- 羥基多巴 胺 (6-hydroxydopamine) 誘發細胞損傷的神經元 (Hiep et al., 2015),而另外也有 文 獻 指出 大 豆異黃酮 可以透 過改善 突觸可塑性 (synaptic plasticity) 和增 加 cholinergic 神經系統的活性等神經保護及修復生長,達到改善學習和記憶、神經 元變性和海馬迴凋亡的情況 (Wei et al., 2015)。紅蜂膠 (red propolis) 當中的異黃 酮素也能修復坐骨神經 (sciatic nerve injury) 的損傷,其原因不只是抑制了周圍 膠細胞所引起的發炎反應,同時也促進了神經細胞的加速生長,加速 sciatic nerve injury 損傷後的功能恢復 (Barbosa et al., 2015)。
3. 調節血脂
大豆異黃酮素近年來被研究出可降低血管中 LDL 並增加 HDL,減少血脂 沈積在血管壁的機會,因此對血管具有保護作用,此功效也被運用在保健食品之 上,過去研究顯示異黃酮和植物固醇 (phytosterols) 通過提高膽酸 (bile acids) 的 清除,減少腸內膽固醇的吸收可以發揮降低小鼠血液中膽固醇作用 (Feng et al.,
23
2015a)。另外,對於高血脂症的大鼠,大豆異黃酮素也能透過降低 LDL-C 並且 增高 HDL-C 的含量,同時增加睪固酮 (testosterone) 的濃度降低腎上腺皮質素 (cortisol) 的分泌,達到降低血液中膽固醇作用 (Rababah et al., 2015)。
4. 預防骨質疏鬆症
大豆異黃酮可協助體內鈣質保留以預防骨質疏鬆,在日本的女性,因為較經 常食用大豆製品而攝取較多異黃酮素,以至於在停經後不會大量流失鈣質預防了 骨質疏鬆 (Greendale et al., 2015)。經過去研究顯示,其能提高葡萄糖胺聚醣 (glycosaminoglycans) 在去除卵巢的大鼠體內,能增加其骨質質量和膠原纖維 I (type I collagen fibers) (Santos et al., 2014)。
5. 抗氧化發炎
大豆異黃酮之抗氧化能力在市面上已經有許多的保健應用,過去研究出大豆 異黃酮防止脂肪肝病 (fatty liver disease) 的能力,是由於透過調節脂肪酸 β (fatty acid β) 的氧化形成脂質,同時也能阻斷 peroxisome proliferator-activated receptor- α (PPAR-α) 誘發的脂肪酸氧化;還有抑制醛醣還原酶 (aldose reductase),使血糖 不會被還原成醛醣,近一步造成細胞膜通透性的損傷;減弱 Cytochrome P450 2E1 (CYP2E-1) 誘發的氧化反應,減少肝臟脂肪變性 (Qiu and Chen, 2015)。而其抗 氧化能力對於預防阿茲海默症扮演重要之角色,在 D-galactose 誘發形成之老化 小鼠試驗中顯示。大豆異黃酮能抑制模式動物所引起的氧化壓力與相關老化因子 之表現 量。 D-galactose 顯著提 高腦 部 Aβ 、presenilin-I 與 beta-site amyloid precursor protein cleaving enzyme-1 (BACE-1) 之表現量。而大豆異黃酮則能藉由 氧化壓力的改善而使這些阿茲海默症相關危險因子之表現下降。
24
第三節、 乳酸菌發酵豆奶對阿茲海默症之預防潛力
豆奶在經過 NTU 101 發酵的過程當中,glucoside isoflavones 會被轉換成具 有更高生理活性、更容易被生物體所吸收的物質 aglycone isoflavones,其中的主 要功效成分為 genistein、daidzein 與 daidzein 的次級代謝物 equol。而這些被轉 化的大豆異黃酮已經於過去的許多研究中,看出具有預防 AD 之潛能 (Gleason et al., 2015)。如抑制 Aβ 之剪切路徑,阻止其產生類澱粉斑塊產生堆積;活化神 經傳導路徑,提升 ACh 的濃度或降低 AChE 的含量,阻止 NMDA 受體打開 離子通道,使神經元不會受到過多鈣離子傷害;也能減緩 astrocyte 及 microglia 釋出的氧化發炎物質,提升 SOD、CAT 等抗氧化酵素,或降低 TNF-α 和 IL-1β 等前發炎因子的表現,以達到預防或抑制 AD 的功效。
一、去醣基異黃酮與雌性激素
Aglycone isoflavones 當中的 genistein 由於其作用與構型類似動情激素,固 有植物的雌性激素之稱,如其名 genistein 具有與雌性激素相似的功效,如使細 胞增生並且分化的功效,對於神經退化性的疾病具有適性相當良好的改善效果,
所以被拿來作荷爾蒙補充療法 (HRT) 的替代品,而又不會造成細胞無限制的增 生導致罹患癌症的風險增高 (National Institutes of Health, 2005)。
Genistein 具有神經保護功能,在 2016 年 Aoyama 的研究中發現 genistein 的攝取能夠減緩肌肉萎縮的發生,因為肌肉萎縮的症狀與雌性激素的減少導致神 經衝動減少所致,而在切除坐骨神經而引起肌肉萎縮的雄性 Wistar 大鼠餵食過 genistein 後,達到了減緩萎縮的效果 (Aoyama et al., 2016)。而對於肌肉的研究 也有在 2012 年 Valeri 所作之研究,genistein 有如同雌性激素的效果能治療神 經性的膀胱肌肉鬆弛,在餵食過 genistein 後對於大鼠之逼尿肌 (detrusor smooth muscle) 具有刺激反應與清除 MDA 以減緩氧化損傷的功效 (Valeri et al., 2012)。
另外也有對於神經母細胞瘤 (Neuroblastoma, NB) 的研究,結果顯示 genistein 能有效降低 P53 與 CHD5 等致癌基因的表現,並抑制其血管的增生來抑制神 經瘤的產生 (Li et al., 2012)。
Daidzein 也具有神經保護功能,在 2015 年 Kim 的研究中發現 daidzein
25
有促進軸突生長的功能,並能使先天性缺乏脂質載體蛋白 (apo E) 的小鼠刺激產 生 apo E 之功效,所以對於脂質大量堆積所造成的中風 (stroke) 具有改善的功 效,並且說明長期的食用 daidzein 對於慢性的神經疾病有保護作用 (Kim et al., 2015)。
二、去醣基異黃酮抑制類澱粉樣蛋白質剪切路徑
Aglycone isoflavones 當中的 genistein,過去研究指出使卵巢切除的小鼠因 為失去製造雌性激素的能力,往往會導致細胞的代謝失調,在 AD 中也會使 Aβ 不易代謝與死亡的神經細胞無法順利增生等增加 AD 嚴重性的症狀出現,而在 餵 食過 genistein 後發現 genistein 能抑制 β-secretase 之 mRNA 含量 而使 APP 無法剪切成 Aβ,使卵巢切除而缺乏雌性激素的小鼠達到預防 AD 的功效 (Li et al., 2013)。同樣在 2015 年 Ahmad 的研究中顯示,在 Aβ42 的體外細胞 實 驗 當中 看出 genistein 與 daidzein 能有效降低 β-secretase 之活性與清除 DPPH 自由基之能力 (Ahmad et al., 2015)。
另外當腦部缺血時, glutamate 會被大量的產生雖然為重要之神經傳遞訊號,
但過量的話會造成鈣離子濃度不斷上升,過量的鈣離子會破壞細胞膜電位導致細 胞死亡,也會大量活化 APP,而 APP 的大量表現會導致下游的代謝路徑加速,
破壞細胞所造成的自由基會引發 NO 的大量產生,大量的 NO 會催化進入 amyloidogenic 路徑產生 Aβ,而 genistein 能降低 NO 的表現並也能增加 α- secretase 同 時 減 少 β-secretase 與 γ-secretase 的 表 現 , 使 APP 無 法 經 由 amyloidogenic 路 徑 剪 切 成 Aβ , 也 能 增 加 胰 島 素 降 解 酶 (insulin-degrading enzyme) 加速 Aβ 的降解 (McCarty, 2006)。Genistein 在體外細胞試驗中,可以 增加 α-secretase 的表現量已達到減少 Aβ(25-35) 含量之能力,已到神經保護之 能力 (Liao et al., 2013),而在基因缺陷的小鼠注射 genistein 治療的試驗中,注 射 genistein 的組別與注射 17β- 雌性激素 (17 beta-estradiol, E2) 的組別,都有 降低腦部澱粉樣蛋白含量與增加 Aβ 之清除率的能力 (Li et al. 2013),
而白皮杉醇 (piceatannol) 與茶樹 (camellia sinensis) 之萃取物在 Y 自迷宮 的行為試驗之中,能有效改善 AD 大鼠的記憶行為能力,而之後分析其萃取物
26
之成分,主要功效成分就含異黃酮素 daidzein (Hassaan et al., 2014)。Daidzein 也 能抑制 β-secretase 的活性,達到抑制 Aβ 產生 (Ahmad et al., 2015)。
三、去醣基異黃酮活化神經傳導路徑
Genistein 透過了抑制細胞中鈣離子的濃度,使 Aβ(25-35) 造成的細胞損傷 和神經細胞凋亡減輕 (Zeng et al., 2004),提升 ACh 的濃度以及降低 AChE mRNA 的表現量 (Xi et al., 2011) 並增加海馬迴中 neuronal nitric oxide synthase (nNOS) 的含量,達到增加神經傳遞因子的濃度 (Bagheri et al., 2012)。以 (3- Nitropropionic acid, 3-NPA) 誘發切除卵巢大鼠的記憶障礙,在 genistein 處理過 後能使大鼠在被動迴避行為試驗得到改善,並發現 AChE 的活性下降、前發炎 因子 iNOS 與 COX-2 的表現量下降,達到神經保護和增強記憶能力的效果 (Menze et al., 2015)。
四、去醣基異黃酮對氧化壓力與發炎反應之抑制效果
Genistein 急劇減少 PC12 細胞群中過多的 ROS,並增加了 GSH 的表現量,
減弱 Aβ(25-35) 細胞毒性誘發的細胞凋亡 (Ma et al., 2010),其與雌性激素相似 的結構可與雌性激素受體結合,增加 SOD 的活性並減少 MDA 與 NO 等氧化 因子的影響 (Bagheri et al., 2011; Ma et al., 2013)。大豆異黃酮中的植物性雌激素 genistein 於過去多項研究中顯示具有預防阿茲海默症之效果,Aβ40 腦部輸注之 之大鼠模式中證實,餵食 genistein 能改善記憶學習能力,此外其對於 Aβ之腦 部氧化壓力具有改善效果,包含抑制 Aβ 於腦部引起的 malondialdehyde (MDA) 上升與 SOD 下降,進而改善短期的空間記憶能力。genistein 具有預防阿茲海默 症之效果,Aβ40 腦部輸注大鼠模式中證實,餵食 genistein 能改善記憶學習能 力,對於 Aβ 之腦部氧化壓力具有改善效果,抑制 Aβ 於腦部引起的 MDA 上 升與 SOD 下降,進而改善短期的空間記憶能力。Genistein 因為本身的強抗氧 化抗發炎能力,在過去研究中看出具有抑制非酒精性脂肪肝 (Nonalcoholic steatohepatitis) 之功效,研究成果顯示 6 週後再餵食 genistein 的高油紙飲食組 SD 大鼠,肝臟中的 MDA 脂質過氧化指標有顯著的下降,以及當中的前發炎因
27
子 TNF-α 與 (peroxisome proliferator-activated receptor-γ, PPAR-γ) 都有顯著的差 異,而結論是 genistein 因為抑制氧化壓力與發炎反應,而達到治療 Nonalcoholic steatohepatitis 之功效 (Susutlertpanya et al., 2015)。在 2015 年 Ganai 的研究中 發現,genistein 可以用來治療 d-Galactosamine 誘發的猛爆性肝衰竭 (fulminant hepatic failure) Wistar 大鼠,透過抑制 iNOS 與 COX-2 來降低 NO 與前列腺素 (PGE-2) 等前發炎因子表現,以及抑制 NF-κB 進入細胞核而降低 TNF-α、IL-1β 等前發炎因子的表現量,達到治療肝炎的功效 (Ganai et al., 2015)。另外 genistein 通過抑制氧化壓力與減少自由基的釋放,還有抑制 NF-κB、IL-6 與細胞黏附因 子 (intercellular adhesion molecule 1, ICAM-1) 的活化達到抑制內皮細胞的發炎 損傷,避免發炎的細胞和血小板黏合 (Han et al., 2015)。
Daidzein 預防神經元損傷可以經由抑制 PPAR-γ 的受體,使前發炎因子無 法再成發炎反應,減少大鼠皮層神經元因為缺血性氧糖剝奪症 (oxygen-glucose deprivation, OGD) 所造成的細胞死亡 (Hurtado et al., 2012)。Daidzein 的抗氧化 能力也被運用在肺損傷而引發敗血症的治療上,以盲腸結紮穿孔 (cecal ligation and puncture) 的方 式引 發小 鼠肺 臟損 傷 而誘 發敗 血症 作為 模式 動物 ,並 以 daidzein 皮下注射的方式進行治療,結果發現 daidzein 的治療使腹膜液、血液和 肺臟中細菌數降低,降低的血液中 TNF-α 的表現,導致發炎物質 NO 的含量下 降,結論是 daidzein 治療 7 天後使肺損傷而引發敗血症的小鼠增加存活時間,
原因是來自於降低細菌數與降低發炎物質的含量導致發炎反應降低 (Parida et al., 2015)。
五、去醣基異黃酮對 astrocyte 及 microglia 活化之抑制效果
Genistein 為大都異黃酮中含量比例最高的成分,在 2015 年 Ma 的研究中 可以發現,在被 Aβ(25-35) 處理過的 C6 神經膠質細胞 (C6 glial cells) 經過 genistein 共培養 2 個小時後,在 (methylthiazolyl tetrazolium, MTT) 處理的細胞 存活率試驗中發現 genistein 共培養 2 個小時的組別,會有顯著的提升細胞存 活率,並且發現 genistein 可以抑制 TLR-4 的含量,導致下游的發炎反應受到 抑制,包括 TNF-α、IL-1β 與 NF-κB 等前發炎因子的表現量下降,而這可能是