台灣中部蓮華池森林樹種群聚與地形、
水分有效性及樹種耐旱性的關係
郭耀綸1,3) 張勵婉2) 林易養1) 余尚鈺1)
摘 要
台灣中部蓮華池森林在山坡不同位置的生育地有不同的樹種群聚。本研究探討該森林樹種群聚 與地形、水分有效性及樹種耐旱性間的關係。試驗樹種包括分布在山脊、坡面、溪谷三處生育地的52 樹種。本研究藉土壤含水率及植株黎明前葉部水勢(Ψpd),指示各生育地的水分有效性。藉各樹種的膨 壓喪失點葉部水勢(πtlp),比較樹種間及不同樹種群聚間葉部耐旱性的差異。結果發現,在乾季期間山 脊生育地的土壤含水率及植株的Ψpd,均顯著低於溪谷生育地。供試所有樹種乾季時πtlp在-2.98~-2.03 MPa範圍,平均為-2.49±0.03 MPa,以大丁黃、南投石櫟、臺灣紅豆樹有最高的葉部耐旱性。山脊、
坡面、溪谷三處生育地的樹種群聚,乾季期間的平均πtlp分別為-2.66±0.05、-2.50±0.07、-2.25±0.07 MPa,以山脊群聚樹種的葉部耐旱性顯著最高。依據πtlp區分樹種耐旱性等級,供試樹種中有29種歸類 為「耐旱」等級,另23種則屬於相對較不耐旱的「中等耐旱」等級。乾季期間大多數樹種的πtlp顯著低 於雨季時,顯示葉部耐旱性呈現季節性馴化。此外,同一樹種沿著水分有效性梯度,植株葉部耐旱性 有明顯的空間馴化,生長在山脊植株的πtlp顯著低於溪谷同種的植株。乾季期間生長在山脊18樹種的πtlp
與Ψpd呈顯著負相關,顯示生理耐旱性較高的樹種,形態上可能也具根深性。本研究發現,生育地水分 有效性及樹種葉部耐旱性,是影響蓮華池森林樹種群聚於生育地空間分化眾多機制中重要的兩項。
關鍵詞:生育地水分有效性、葉部耐旱性、膨壓喪失點葉部水勢、黎明前葉部水勢、季節性馴化。
郭耀綸、張勵婉、林易養、余尚鈺。2020。台灣中部蓮華池森林樹種群聚與地形、水分有效性及樹種 耐旱性的關係。台灣林業科學35(2):123-42。
1) 國立屏東科技大學森林系,91201屏東縣內埔鄉學府路1號 Department of Forestry, National Pingtung Univ. of Science and Technology, 1 Xuehfu Rd., Neipu Township, Pingtung 91201, Taiwan.
2) 林 業 試 驗 所 森 林 保 護 組 ,10066台北市南海路53號 Forest Protection Division, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
3) 通訊作者 Corresponding author, e-mail:[email protected]
2020年3月送審 2020年6月通過 Received March 2020, Accepted June 2020.
Research paper
Relationships of Tree Species Associations with the Topography, Water Availability, and Species Drought
Tolerance in the Lienhuachih Forest of Central Taiwan
Yau-Lun Kuo,1,3) Li-Wan Chang,2) Yi-Yang Lin,1) Shang-Yu Yu1)
【Summary】
Different species associations are situated in habitats at various slope positions in the Lien- huachih forest of central Taiwan. This research investigated relationships of tree species associa- tions with the topography, water availability, and species drought tolerance of this forest. In total, 52 species distributed in ridge, slope, and valley habitats were studied. The soil water content and predawn leaf water potential (Ψpd) were adopted to indicate the water availability of each habi- tat. The leaf water potential at the turgor loss point (πtlp) was used to compare differences in leaf drought tolerance among species and species associations. Results showed that the soil water con- tent and Ψpd of species in the ridge habitat were significantly lower than those in the valley habitat.
Values of the πtlp of all species during the dry season ranged -2.98~-2.03 MPa, with a mean πtlp of -2.49±0.03 MPa. Among the studied species, Euonymus laxiflorus, Lithocarpus nantoensis, and Ormosia formosana had the highest leaf drought tolerance. Species associations of ridge, slope, and valley habitats showed mean πtlp values of -2.66±0.05, -2.50±0.07, and -2.25±0.07 MPa, respectively, with the species association of the ridge habitat having the highest leaf drought toler- ance. After adopting values of the πtlp to categorize drought tolerance classes of the 52 tested spe- cies in the Lienhuachih forest, 29 species were classified into the ‘drought-tolerant’ category, and the other 23 species were ‘mid-drought tolerant’ with relatively less tolerance. The πtlp of every species was significantly lower during the dry season than the rainy season, indicating seasonal ac- climation of the leaf drought tolerance. In addition, there was also significant spatial acclimation in the leaf drought tolerance along the water availability gradient, i.e., πtlp values of individuals growing in the ridge habitat were all significantly lower than those of individuals of the same spe- cies growing in the valley habitat. During the dry season, the Ψpd and πtlp of 18 species growing in the ridge habitat appeared to be negatively correlated, indicating that species with high physiologi- cal drought tolerance might also have deeper root systems. This research found that habitat water availability and leaf drought tolerance are two important mechanisms, among others, driving habi- tat divergence of tree species associations in the Lienhuachih forest.
Key words: habitat water availability, leaf drought tolerance, leaf water potential at turgor loss point, predawn leaf water potential, seasonal acclimation.
Kuo YL, Chang LW, Lin YY, Yu SY. 2020. Relationships of tree species associations with the to- pography, water availability, and species drought tolerance in the Lienhuachih forest of cen- tral Taiwan. Taiwan J For Sci 35(2):123-42.
緒 言
地 形(topography)是地表高低起伏的狀 態。在山地地形,由下坡到上坡山脊不同坡面 位 置 , 林 地 能 保 有 的 水 分 含 量 通 常 會 減 少 , 致土壤水分有效性(soil water availability)呈現 梯度變化(Becker et al. 1988, Daws et al. 2002, Comita and Engelbrecht 2009)。因地形引起的 水分有效性變異,常會影響樹種的空間分布,
因而在水分有效性差異大的生育地有不同樹種 群聚(tree species association) (Daws et al. 2002, Engelbrecht et al. 2007, Bartlett et al. 2016b)。
縱 使 在 相 對 潮 濕 的 森 林 生 態 系 , 樹 種 群 聚 也 會 受 到 地 形 變 異 造 成 水 分 有 效 性 改 變 的 影 響 (Markesteijn and Poorter 2009)。另一方面,
許多研究發現樹種的生理耐旱性(physiological drought tolerance),是決定樹種在不同水分 有效性梯度分布的重要機制(Tyree et al. 2003, Comita and Engelbrecht 2009, Markesteijn et al.
2011, Fletcher et al. 2018, Kuo et al. 2018)。
無 論 大 尺 度 或 局 部 地 區 小 尺 度 , 生 育 地 水 分 有 效 性 與 樹 種 耐 旱 性 常 具 顯 著 負 相 關 , 生 長 在 缺 水 環 境 的 樹 種 多 具 有 較 高 的 生 理 耐 旱 性 (Markesteijn and Poorter 2009, Bartlett et al.
2012, Zhu et al. 2018)。因此,樹種的耐旱性 與生育地水分有效性,是樹種生育地群聚的強 烈驅動力(Baltzer et al. 2008, Markesteijn and Poorter 2009, Bartlett et al. 2016b)。在臺灣亞 熱帶山區樹種群聚的空間分布樣式,應也會受 到地形、水分有效性及樹種耐旱性三者共同的 影響。
林 木 葉 部 水 勢 可 反 應 該 植 株 根 系 分 布 範 圍的土壤水分狀況,且在清晨未進行蒸散作用 前,植株水勢與土壤水勢可達平衡狀態(Kramer 1983),兩者具顯著正相關,因此黎明前葉部 水勢(Ψpd)可反應土壤水分有效性程度,可當做 指示生育地水分有效性良好的指標(Kuo 1994, Stahl et al. 2013, Zhou et al. 2014, Mitchell and O’Grady 2015)。在水分有效性低的季節,林木 的Ψpd會降低(Kuo 1994, Zhou et al. 2014, Kuo et al. 2017);同一季節生長在水分有效性低之生育 地的植株,Ψpd也會顯著較低(Kuo et al. 2018)。
此外,在水分有效性類似的同一處生育地,具 深根性的樹種(或植株)因可獲取較深層土壤水 分,其Ψpd會高於根系較淺的樹種(或植株) (Kuo 1994, Fallon and Cavender-Bares 2018)。因此,
在同一處生育地可藉Ψpd判斷樹種根系分布的深 淺(Kuo et al. 2017)。
耐 旱 性 為 植 物 的 遺 傳 性 狀 , 不 同 樹 種 對 缺 水 逆 境 的 敏 感 性 或 忍 受 性 的 差 異 很 大(Lenz et al. 2006, Niinemets and Valladares 2006, Engelbrecht et al. 2007)。樹種的耐旱能力可藉 由延遲脫水(dehydration postponement)機制或 忍受脫水(dehydration tolerance)機制,其中一 項或兩者共同作用而達成(Kuo 1994, McDowell et al. 2008, Comita and Engelbrecht 2014, Kuo et al. 2017)。例如有些樹種在形態上具有深根性 或具良好的儲水能力,在生理上氣孔具有良好 的控制水分散失能力,或具落葉性。前兩項為 開源(增加水分獲取)的性狀,後兩項則為節流 (節約用水)的性狀。上述開源與節流的能力可歸 類在植物的延遲脫水機制(Kramer 1983, Volaire 2018)。另一方面,當生育地土壤水分持續喪 失,缺水逆境較嚴重時,有些樹種葉部細胞在 本質上即具忍受脫水能力,或導管仍能維持水 分傳導,因此生理上耐旱能力高(Tyree et al.
2003, Maréchux et al. 2018)。有些樹種可藉由滲 透壓調節(osmotic adjustment)方式降低葉部滲 透勢,因而降低膨壓喪失點葉部水勢(leaf water potential at the turgor loss point, πtlp),可讓葉片 在較低水勢條件下仍能維持細胞膨壓,因而維 持正常生理活動(Baltzer et al. 2008, Bartlett et al. 2012)。上述各項耐旱性狀可歸類在忍受脫水 機制(Kramer 1983, Delzon 2015, Volaire 2018)。
忍受脫水機制即為植物在遺傳上具有的生理耐 旱性(Poorter and Markesteijn 2008)。
膨 壓 喪 失 點 葉 部 水 勢(πtlp)常被用來當做 植 物 葉 部 耐 旱 性 的 指 標 , 因 為 此 性 狀 界 定 了 植 物 有 能 力 控 制 生 長 及 氣 體 交 換 的 水 勢 範 圍 (Mitchell and O’Grady 2015, Blackman 2018),
可用來量化物種生理耐旱性(Lenz et al. 2006, Blackman et al. 2010, Bartlett et al. 2012, Maréchaux et al. 2015)。因為πtlp代表葉部缺水
及土壤乾燥所引發的葉片凋萎,此參數被認為 是植物的高階性狀(higher-level trait) (Sack et al.
2003, Lenz et al. 2006)。乾季期間有些樹種具滲 透壓調節能力,πtlp可顯著較雨季期間降低,此 即為πtlp的季節性調節或馴化(Bartlett et al. 2012, Maréchaux et al. 2015)。研究顯示全球236種木 本植物πtlp分布範圍在-0.75~-4.59 MPa (Bartlett et al. 2016a, appendix);中國9種林型共389種 木本植物的πtlp在-0.68~-4.16 MPa (Zhu et al.
2018, appendix);澳洲174樹種πtlp的分布範圍 在-1.16~-4.72 MPa (Mitchell and O’Grady 2015, appendix);南美洲法屬蓋亞納熱帶雨林71樹 種的πtlp在-1.04~-3.20 MPa (Maréchaux et al.
2015);中國西雙版那熱帶林43樹種πtlp的分布 範圍為-1.17~-2.36 MPa (Bartlett et al. 2016b)。
上述報導顯示不同生態系間或同一生態系內不 同樹種間,葉部耐旱性的變異都很大。因為πtlp 數值可指示不同樹種葉部耐旱性的差異,而葉 部耐旱性與全株耐旱性也具密切關係(Bartlett et al. 2012, Maréchux et al. 2018),或許可藉πtlp數 值區別樹種的耐旱性等級。然而,目前學界尚 無藉πtlp數值區別樹種耐旱性等級的報導。
台 灣 中 部 蓮 華 池 森 林 動 態 樣 區 的 植 群 可 分 為 四 種 類 型 , 第 一 型 南 投 石 櫟– 茜 草 樹 型 (Lithocarpus nantoensis-Randia cochinchinensis type),分布在山脊及最高海拔;第二型白匏 子–黃杞型(Mallotus paniculatus-Engelhardtia roxburghiana type),分布在中上坡(upper slope);第三型山紅柿–厚殼桂型(Diospyros morrisiana-Cryptocarya chinensis type),
分 布 在 下 坡 及 溪 邊 ; 第 四 型 大 葉 楠– 山 龍 眼 型(Machilus japonica var. kusanoi-Helicia formosana type),分布在樣區西側溪邊(Chang et al. 2012)。 Chang et al. (2012)指出這四類 植 群 型 的 樹 種 生 育 地 群 聚 及 棲 位 分 化( n i c h e divergence),與該處微地形的變異極具相關 性,且以距山脊距離、距溪流距離、凹凸度此 三項微地形因子影響最大。上述三項微地形因 子均與水分有效性息息相關。因此水分有效性 的差異可能是蓮華池森林不同類型的植群,群 聚在不同微地形生育地的重要趨動力。近年來
因極端氣候造成的乾旱事件越趨頻繁,嚴重性 也更加劇;全球變遷型的乾旱(global change- type drought)引起的大面積樹木死亡令初級生產 力減損(Choat et al. 2018),也可能因較不耐旱 樹種死亡而改變植群組成結構。因此,對台灣 原生樹種耐旱能力與耐旱機制的瞭解,為急切 需要的基礎生態生理學知識。若能得知蓮華池 森林在不同地形的植群間,組成樹種之生理耐 旱性本質及其差異程度,則可增加我們對此森 林樹種空間分布機制的瞭解。
本研究目的為探討蓮華池森林樹種群聚與 地形、水分有效性及樹種耐旱性間的關係。此 外,本研究也想瞭解蓮華池森林樹種的耐旱性 等級,以及如何應付乾季期間的缺水逆境。針 對此目的,本研究提出下列幾項問題:(1)蓮華 池森林不同坡位的生育地,乾季期間土壤水分 有效性是否具顯著差異?(2)此森林群聚在水分 有效性較低之生育地的樹種,其葉部耐旱性是 否會顯著高於群聚在水分有效性較高之處的樹 種?(3)在乾、雨兩季節,供試樹種的葉部耐旱 性是否有季節性馴化?(4)同一樹種分布在不同 生育地的植株,其葉部耐旱性是否會沿著水分 有效性梯度而有馴化表現?(5)生長在水分有效 性最低之生育地的樹種,在乾季期間是否能藉 由延遲脫水機制,提高應付缺水逆境的能力?
材 料 與 方 法
試驗地概述
試驗地位於南投縣魚池鄉,林業試驗所蓮 華池研究中心所轄的25 ha森林動態樣區,以及 該樣區範圍外較低海拔的森林。該樣區最高海 拔為845 m,坡度平均為35.3°,具有山谷、峭 壁及稜脊地形(Chang et al. 2012)。該中心氣象 站1928~2016年之記錄顯示此處年平均降雨量 為2409 mm,雨量集中在4~9月(85.8%),10月 至翌年3月為乾季,且乾季前4個月之各月平均 雨量都低於50 mm (Lu et al. 2018)。根據林業 試驗所集水區經營組提供之資料,蓮華池研究 中心近90年之10月至翌年2月期間,累積雨量低 於150 mm的只有8個年度,其中低於50 mm的
只有1939~1940及2018~2019兩個年度,而本研 究調查期間2018年10月至2019年2月累積雨量 僅11 mm,且由2018年9月27日起至隔年1月3日 才降下1.0 mm雨量,該期間連續無降雨日高達 98天,已超過該地區之「極端乾旱」門檻值(65 天連續無降雨) (Lu et al. 2018),為歷年少見的 極端乾旱期。蓮華池森林動態樣區植群屬於亞 熱帶常綠闊葉樹林,有144種木本植物,以茜 草科有最多株數(20.86%),殼斗科有最大胸高 斷面積(26.47%),而以樟科物種數最多(14種) (Chang et al. 2012)。
試驗樹種
蓮 華 池 森 林 動 態 樣 區 的 樹 種 群 聚 可 區 分 為 四 類 植 群 型 , 在 地 形 的 空 間 分 布 上 第 一 型 位於山脊,第二型位於上坡及中坡,第三型在 下坡及溪邊,第四型在西側溪邊(Chang et al.
2012)。第三與第四植群型分布的地形相同,主 要是方位不同,因此本研究將此兩型合併。上 述三類植群型的生育地分別稱為山脊、坡面、
溪谷生育地。就海拔高度而言,蓮華池森林海 拔780 m至 845 m平坦稜線處為山脊生育地,海 拔720~780 m屬坡面生育地,海拔660~720 m為 溪谷生育地。本研究取樣路線由海拔665 m的登 山口沿著登山步道至稜線,在三處生育地共選 取52樹種供葉部耐旱性測定。根據各供試樹種 在動態樣區山脊、坡面、溪谷三處生育地的株 數及胸徑所計算的重要值(importance value),
顯示有10, 20, 9樹種的族群分別以山脊、坡面、
溪谷為專一生育地(重要值較另兩處高出15%以 上) (Table 1)。然而,族群在山脊與坡面兩處生 育地重要值都大於40%或相差少於10%的有7樹 種,為山脊與坡面共域(sympatric)樹種;族群 在坡面與溪谷兩處生育地的重要值都大於40%
的有6樹種,為坡面與溪谷共域樹種(Table 1)。
此52樹種佔蓮華池森林動態樣區144種木本植 物總數的36.1%,但已包含此區主要的組成樹 種,且此52樹種在動態樣區合計的重要值已高 達82.66% (Chang et al. 2012)。供試樹種中以殼 斗科(10種)及樟科(9種)物種數較多。供試各樹 種的學名依據2017台灣維管束植物紅皮書名錄
(Editorial Committee of the Red List of Taiwan Plants 2017)所示。
不同坡位生育地土壤含水率測定
於2 0 1 9 年 2 月 2 6 日 乾 旱 嚴 重 期 , 以 及 同 年5月與9月雨季期間,在蓮華池森林山脊(海 拔 高820~830 m)、坡面(730~740 m)及溪谷 (690~7000 m)三處生育地,每處選取相距4~6 m 的20處測點,移除土壤上方的腐植質後,使用 12 cm長的探針測定該深度範圍的土壤體積含水 率。使用的儀器為土壤水分測定計(TDR100 soil moisture meter, Spectrum Technologies, Aurora, IL, USA)。
黎明前葉部水勢測定
本 研 究 以 兩 項 試 驗 測 定 不 同 生 育 地 植 株 的 黎 明 前 葉 部 水 勢(Ѱpd)。第一項試驗選定山 脊 及 溪 谷 都 有 分 布 的8 種 樹 種 的 稚 樹 , 於 乾 季 測 定 其Ѱp d, 藉 植 株 的Ѱp d指 示 不 同 坡 位 生 育 地 土 壤 水 分 有 效 性 的 差 異 。 選 定 稚 樹 而 非 成 樹 的 原 因 是 稚 樹 植 株 小 , 根 系 分 布 可 能 較 淺 , 可 反 應 較 淺 層 土 壤 的 水 分 狀 態 。 選 定 的 樹 種 包 括 小 葉 樹 杞(Ardisia quinquegona)、
柏 拉 木( B l a s t u s c o c h i n c h i n e n s i s ) 、 香 桂 (Cinnamomum subavenium)、厚殼桂、台灣 八 角(Illicium arborescens)、變葉新木薑子 (Neolitsea aciculata var. variabillima)、台灣紅 豆樹(Ormosia formosana)、九節木(Psychotria rubra)等8樹種。每一樹種分別在山脊及溪谷 兩處生育地各標定5株樹高80~190 cm的稚樹,
只 有 台 灣 八 角 在 溪 谷 生 育 地 的 稚 樹 較 小(樹高 30~80 cm)。於2019年1月21日乾旱期間測定稚 樹的Ѱpd (測定日前116日累積雨量僅5.5 mm)。
測定日當天早上05:00黎明前採集兩生育地8樹 種稚樹植株上方各一枝帶葉枝條,裝入夾鏈袋 再 置 於 保 冷 袋 內 。 攜 回 蓮 華 池 研 究 中 心 後 測 定同一枝條2~3個葉片的Ѱpd。使用儀器為植物 水勢計(pressure chamber, Model 1505D, PMS Instrument, Albany, OR, USA)。將各單株2~3個 Ѱpd測值平均,得該單株Ѱpd數值,再以各樹種5 單株的Ѱpd平均值代表該樹種當天在山脊或溪谷
Table 1. List of species, family, importance value at each habitat, and major habitat associations of the 52 tested species in the Lienhuachih Forest Dynamics Plot
Species Family Importance value (%) Major
Ridge Slope Valley habitat
Podocarpus nakaii (桃實百日青) Podocarpaceae 42.2 52.4 5.4 R, S1) Ardisia quinquegona (小葉樹杞) Myrsinaceae 23.3 49.2 27.5 S Ardisia sieboldii (樹杞) Myrsinaceae 0.4 14.6 85.0 V Castanopsis cuspidata var. carlesii (長尾尖葉櫧) Fagaceae 42.9 49.8 7.3 R, S
Castanopsis fargesii (火燒栲) Fagaceae 33.7 55.2 11.1 S
Castanopsis kawakamii (大葉苦櫧) Fagaceae 25.9 56.6 17.5 S
Castanopsis uraiana (烏來柯) Fagaceae 24.6 52.6 22.8 S
Cinnamomum osmophloeum (土肉桂) Lauraceae 0.0 2.9 97.1 V Cinnamomum subavenium (香桂) Lauraceae 35.4 50.9 13.7 S Cryptocarya chinensis (厚殼桂) Lauraceae 4.9 46.0 49.1 S, V Diospyros morrisiana (山紅柿) Ebenaceae 25.1 51.7 23.2 S Distyliopsis dunnii (尖葉水絲梨) Hamamelidaceae 42.7 46.7 10.6 R, S Elaeocarpus japonicus (薯豆) Elaeocarpaceae 60.0 39.0 1.0 R Elaeocarpus sylvestris (杜英) Elaeocarpaceae 29.8 46.8 23.4 S Engelhardia roxburghiana (黃杞) Juglandaceae 20.2 62.9 16.9 S Euonymus laxiflorus (大丁黃) Celastraceae 59.4 38.8 1.8 R Eurya loquaiana (細枝柃木) Pentaphylaceae 1.2 22.3 76.5 V Eustigma oblongifolium (秀柱花) Hamamelidaceae 25.8 60.2 14.0 S
Ficus fistulosa (水同木) Moraceae 0.0 1.8 99.2 V
Gordonia axillaris (大頭茶) Theaceae 52.1 39.5 8.3 R
Helicia cochinchinensis (紅葉樹) Proteaceae 9.0 74.3 16.7 S
Helicia formosana (山龍眼) Proteaceae 0.1 10.7 89.2 V
Helicia rengetiensis (蓮華池山龍眼) Proteaceae 49.1 49.8 1.1 R, S
Ilex ficoidea (臺灣糊樗) Aquifoliaceae 13.8 58.1 28.1 S
Ilex goshiensis (圓葉冬青) Aquifoliaceae 67.9 31.8 0.3 R
Lithocarpus amygdalifolius (杏葉石櫟) Fagaceae 16.2 57.4 26.4 S Lithocarpus harlandii (短尾葉石櫟) Fagaceae 24.7 52.2 23.1 S Lithocarpus konishii (小西氏石櫟) Fagaceae 34.6 53.7 11.7 S Lithocarpus nantoensis (南投石櫟) Fagaceae 80.2 19.6 0.4 R Lithocarpus synbalanos (菱果石櫟) Fagaceae 60.0 39.0 1.0 R Litsea acuminata (長葉木薑子) Lauraceae 7.9 46.5 45.6 S, V Machilus japonica var. kusanoi (大葉楠) Lauraceae 0.0 3.0 97.0 V
Machilus thunbergii (紅楠) Lauraceae 16.5 65.7 17.8 S
Machilus zuihoensis (香楠) Lauraceae 2.0 20.0 78.0 V
Melicope pteleifolia (三腳虌) Rutaceae 12.1 44.8 43.1 S, V Meliosma squamulata (綠樟) Sabiaceae 46.6 50.5 2.9 R, S Neolitsea aciculata var. variabillima (變葉新木薑子) Lauraceae 43.6 48.0 8.4 R, S
Neolitsea konishii (五掌楠) Lauraceae 0.0 1.2 98.8 V
Ormosia formosana (臺灣紅豆樹) Leguminosae 39.6 49.2 11.2 R, S
Quercus pachyloma (捲斗櫟) Fagaceae 52.9 39.7 7.4 R
生育地的Ѱpd數值。
第 二 項 試 驗 選 定 已 在 山 脊 建 立 的18樹種 的 成 樹 , 於2019年2月27日測定植株的Ѱpd, 供 比 較 各 樹 種 於 乾 季 期 間 獲 取 水 分 能 力 的 差 異 。 該 測 定 日 之 前 的1 5 3 日 累 積 雨 量 僅 11 . 0 mm。供試樹種包括14種原本即以山脊為族群 主 要 分 布 地 的 樹 種 , 以 及4種族群主要群聚在 坡 面 , 但 也 有 植 株 分 布 在 山 脊 的 樹 種 , 分 別 為烏來柯(Castanopsis uraiana)、小西氏石櫟 (Lithocarpus konishii)、火燒栲(Castanopsis fargesii)及狗骨仔(Tricalysia dubia)。供試 18樹種在山脊稜線(海拔820~830 m)各選取 形 體 大 小 接 近 的3 株 成 樹 供 Ѱp d測 定 。 供 試 樹 種 植 株 胸 徑 大 多 在2~10 cm範圍,樹高多在 3 ~ 8 m 範 圍 , 只 有 南 投 石 櫟 植 株 形 體 較 大 , 胸 徑 為2 1 ~ 5 7 c m , 樹 高 11 ~ 1 5 m , 而 薯 豆 (Elaeocarpus japonicus)有2株樹高達12 m。測 定 當 天 於05:00黎明前,採集樣木樹冠外圍的 一段帶葉枝條,放入夾鏈袋再置於保冷袋攜回 蓮華池研究中心。以植物水勢計(PMS, model 1505D)測定各枝條3個葉片的Ѱpd,平均後代表 該單株的Ѱpd,而以同一樹種3個單株Ѱpd的平均 值代表該樹種的Ѱpd。
不同生育地樹種葉部耐旱性測定
在蓮華池森林現地於2016年至2019年,四 年期間於雨季及乾季分年度採取供試52樹種的
葉片進行葉部耐旱性測定。雨季的採樣月份為 4~8月,乾季則於11~3月間採樣。試驗樹種多 在其族群主要生育地採取3株成樹枝葉,但在坡 面生育地採集路線未發現小西氏石櫟、烏來柯 及大葉苦櫧(Castanopsis kawakamii)植株,故此 三樹種是在山脊生育地採樣;因相同狀況,菱 果石櫟(Lithocarpus synbalanos)是在坡面生育 地採樣,杏葉石櫟(Lithocarpus amygdalifolius) 是在溪谷生育地採樣。植株有兩個優勢生育地 的樹種,原則上是在海拔較高處採樣,但因採 樣 時 的 可 及 性 , 變 葉 新 木 薑 子 、 紅 皮(Styrax suberifolius)、水金京(Wendlandia formosana)及 三腳虌(Melicope pteleifolia)是在較低海拔的生 育地採樣。
採 樣 時 以 高 枝 剪 剪 下 樣 木 樹 冠 外 圍 日 照 較 充 足 的 一 段 帶 葉 枝 條 , 置 於 保 冷 袋 。 當 天 帶 回 屏 科 大 實 驗 室 放 入 水 桶 內 , 重 新 剪 除 枝 條 末 端2 cm後,罩以黑色塑膠布過夜。第二 天 清 晨07:00從所採的枝條各選取健康的成熟 葉4片(若葉柄過短則用帶葉小枝),將葉表擦 乾後稱取初始鮮重,立即以植物水勢計(PMS, Model 1505D)或(Model 3005, Soilmoisture Equipment, Santa Barbara, CA, USA)測定葉片 初始水勢,若高於-0.10 MPa則視為已達飽水狀 態,可將其置於桌上自由失水,隔半小時重複 進行葉鮮重及水勢(Ψli)測定。測完4回後,延長 至每小時測定一回,直到有兩回水勢低於-3.0 con’t
Randia cochinchinensis (茜草樹) Rubiaceae 57.8 38.5 3.7 R Rhododendron leptosanthum (西施花) Eyicaceae 72.3 11.6 16.1 R
Psychotria rubra (九節木) Rubiaceae 23.2 51.0 25.8 S
Pyrenaria shinkoensis (烏皮茶) Theaceae 31.3 48.6 20.1 S Saurauia tristyla var. oldhamii (水冬瓜) Actinidiaceae 0.0 0.2 99.8 V Schefflera octophylla (江某) Araliaceae 7.8 41.5 50.7 S, V
Schima superba (木荷) Theaceae 35.1 51.6 13.3 S
Styrax suberifolius (紅皮) Styracaceae 7.2 49.7 43.1 S, V Symplocos theophrastifolia (山豬肝) Symplocaceae 13.1 54.8 32.1 S
Syzygium buxifolium (小葉赤楠) Myrtaceae 75.6 23.6 6.8 R
Tricalysia dubia (狗骨仔) Rubiaceae 28.4 48.6 23.0 S
Wendlandia formosana (水金京) Rubiaceae 1.7 44.9 53.4 S, V
1) R, ridge habitat; S, slope habitat; V, valley habitat.
MPa。完成後將葉片以70℃烘乾至恆重,可得 各 葉 片 乾 重 。 由 失 水 各 階 段 的 葉 鮮 重 減 去 乾 重,即可得葉片水重,再由各階段的葉片水重 除以初始水重,可計算該樣本葉片各失水階段 的相對含水率(relative water content, RWC);
以100減去各階段RWC的數值(100 - RWC)當作 橫軸,各階段的葉片水勢倒數(1/Ψli)當縱軸,
繪製散佈圖,此即為壓力-體積曲線(P-V curve) (Sobrado 2015)。在葉片膨壓(ΨP)未喪失前,樣 點在該散佈圖呈二次曲線下降;當葉片水勢達 膨壓喪失點(πtlp)之後,樣點即呈直線下降。選 取P-V curve呈直線下降的所有樣點,藉下列公 式可求得各樣本葉片的飽水滲透壓(π0),並由 計算所得數據找出各樣本葉片的πtlp (Sack and Pasquet-Kor 2011)。
Slope = -(SD (-1/Ψl)i) / (SD (100 - RWC)i) ... (1) Intercept = (Avg (-1/Ψl)i) – (Slope×Avg (100 - RWC)i) ... (2) Ψ0i = -1/(Intercept+Slope×(100 - RWC)i) .... (3) ΨPi = Ψli - Ψ0i ...(4) 上述公式中SD為呈直線下降各樣點之(-1/
Ψl)或(100 - RWC)數值的標準差;Avg為各 樣點之(-1/Ψl)或(100 - RWC)數值的算術平均 數;Ψ0i為葉片各失水階段的滲透壓;ΨPi為葉 片各失水階段的膨壓。先以公式(1)計算所選取 樣 點 直 線 迴 歸 式 的 斜 率(slope),再以公式(2) 計算所選取樣點在y軸的截點(intercept),此截 點即為飽水滲透壓的倒數(1/π0)。利用slope和 intercept依公式(3)可算出該葉片各失水階段的 Ψ0i,再以公式(4)算出葉片各失水階段的ΨPi。 因為Ψl = Ψ0+ ΨP,膨壓喪失時的葉部水勢應等 於其滲透壓(當ΨP = 0, Ψl = Ψ0 = πtlp),因此可 由葉片ΨPi最接近0之時所對應的Ψ0i數值,來指 示該葉片的πtlp。
每個樹種由至少12筆的π0及πtlp中選取負值 較大,且變異係數(CV)在5%以下的5筆數值平 均,分別代表該樹種的飽水滲透壓及膨壓喪失 點葉部水勢。計算同一樹種雨季及乾季π0或πtlp
的差值,可求得該樹種兩季節間π0或πtlp的調節 (馴化)程度(Bartlett et al. 2012)。
同一樹種在不同生育地植株葉部耐旱性測定 為 比 較 同 一 樹 種 生 長 在 不 同 坡 位 生 育 地 植 株 的 葉 部 耐 旱 性 , 是 否 會 隨 著 生 育 地 水 分 有 效 性 梯 度 而 產 生 馴 化 , 本 研 究 選 取 桃 實 百 日 青(Podocarpus nakaii)、火燒柯、短尾葉 石櫟(Lithocarpus harlandii)、綠樟(Meliosma s q u a m u l a t a ) 、 九 節 木 、 江 某 ( S c h e f f l e r a octophylla)等6樹種,分別生長在山脊、坡面、
溪 谷 三 處 生 育 地 各3株成樹,於12月至2月乾 季期間採取樹冠外圍日照較充足的一段帶葉枝 條,藉上述P-V curve測定方法計算各樹種在各 生育地12個葉片的πtlp。
統計分析
山脊、坡面、溪谷三處生育地土壤含水率 的比較,以及同一樹種生長在三處生育地植株 葉部耐旱性的比較,都先以單因子變異數分析 (ANOVA)比較,再以Scheffé’s test進行事後檢 定,瞭解何者間具顯著差異。同一樹種稚樹在 山 脊 與 溪 谷 兩 處 植 株Ѱpd的 差 異 顯 著 性 , 是 以 t-test 進行分析。生長在山脊18樹種於乾旱嚴重 期間Ѱpd差異顯著性的比較,也是先以ANOVA 進行分析,再以Scheffé’s test進行事後檢定。
在比較三處生育地樹種群聚葉部耐旱性時,各 生育地樹種的選取較為複雜,因有7樹種在山脊 與坡面生育地均有類似的重要值(山脊與坡面共 域),而有6樹種在坡面與溪谷生育地共域(Table 1)。因此在比較不同生育地樹種群聚間葉部耐 旱性的差異時,山脊生育地的樹種樣本數為該 處專一10種,加上山脊與坡面共域7種,共計17 種。坡面生育地樹種樣本數為該處專一20種,
坡 面 與 山 脊 共 域7種,加上坡面與溪谷共域6 種,共計33種。溪谷生育地的樹種樣本數為該 處專一9種,加上溪谷與坡面共域6種,共計15 種。此外,族群在山脊與坡面共域的桃實百日 青及綠樟,在乾季時測過三處生育地植株的πtlp 及π0,在分析時兩處生育地是以各自生育地植 株的πtlp及π0數值為樣本。同樣的,江某在坡面 與溪谷兩處以各自植株的πtlp及π0數值為樣本。
在進行同一季節三處生育地群聚樹種間πtlp或π0
差異顯著性比較時,也是以單因子變異數分析
(ANOVA) 進行比較,再以Scheffé’s test做事後 檢定。各樹種或合併所有樹種乾、雨兩季間πtlp
或π0的差異顯著性比較,是以t-test進行檢定。
以迴歸分析法檢定所有52樹種同一季節πtlp及π0
是否具顯著相關,以及πtlp或π0在兩季節間是否 具顯著相關。乾旱嚴重期山脊18樹種Ѱpd與πtlp是 否具顯著相關,也是以迴歸分析法進行檢定。
結 果
不同生育地水分有效性比較
蓮華池森林於2019年2月乾旱嚴重期間,
山脊、坡面、溪谷三處生育地由地表至12 cm深 的土壤體積含水率分別為3.3、6.6及10.0%,三 處均具顯著差異,以山脊生育地的土壤含水率 最低,溪谷生育地最高(Fig. 1)。雨季期間於5月 及9月測定時,三處生育地的土壤含水率均較乾 季時提高,但仍以山脊生育地的土壤含水率顯 著最低(Fig.1)。
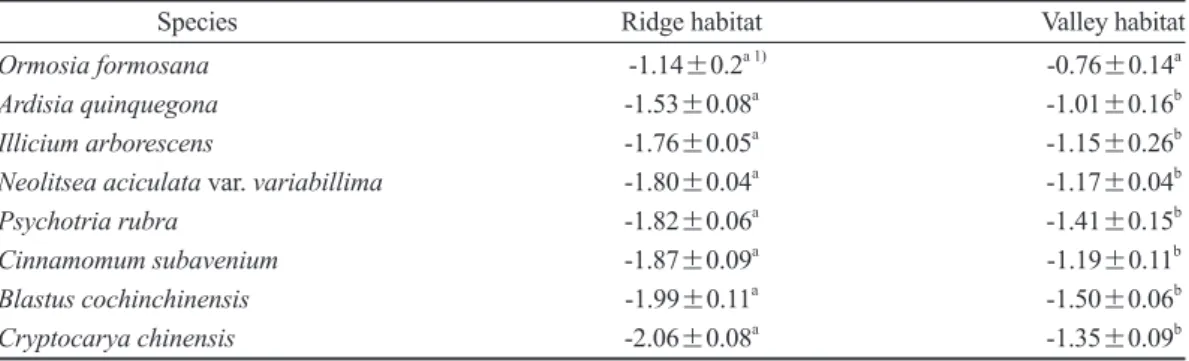
於2019年1月乾季期間,測定蓮華池森林 於山脊與溪谷兩處生育地均有稚樹建立的8種樹 種的黎明前葉部水勢(Ѱpd),發現除了台灣紅豆 樹之外,其餘7樹種生長在山脊之稚樹的πtlp均 顯著低於同種生長在溪谷的植株(Table 2),且 山脊植株的Ѱpd都低於-1.50 MPa,厚殼桂甚至低 至-2.06±0.08 MPa,顯現出已遭嚴重缺水逆境。
然而,台灣紅豆樹生長在溪谷生育地稚樹的Ѱpd
高達 -0.76 MPa,而其它7樹種之Ѱpd則在-1.00~- 1.50 MPa範圍(Table 2),溪谷生育地的植株能獲 取的水分較山脊生育地多。由土壤含水率及植株 水分狀態兩項測定結果,顯示蓮華池森林在乾季 期間最高海拔之山脊生育地的土壤水分有效性,
會顯著低於坡面及溪谷生育地。
同一樹種葉部耐旱性隨水分有效性梯度的馴化 比較同一樹種生長在山脊、坡面、溪谷三 處生育地植株πtlp的差異,發現供試6樹種生長在 山脊生育地植株於乾季期間的πtlp,均顯著低於 同種生長在溪谷生育地的植株(Table 3)。除了 九節木之外,有5樹種生長在山脊植株的πtlp也都 顯著低於同種生長在坡面的植株。火燒栲生長 在坡面生育地植株的πtlp也會顯著低於溪谷處的 植株。上述結果顯示隨著生育地水分有效性的 改變,同一樹種不同植株的葉部耐旱性會有顯 著的空間性馴化,在水分有效性最低之山脊生 育地,植株的葉部耐旱性會顯著提高。
不同生育地樹種群聚葉部耐旱性比較
本研究以供試樹種乾季期間的πtlp當做該樹 種葉部耐旱性的生理指標。結果發現供試52樹 種乾季期間πtlp的分布範圍在-2.98~-2.03 MPa之 間(Table 4),平均為-2.49±0.03 MPa。山脊生
Fig. 1. Comparisons of soil water contents among different habitats during the dry and rainy seasons in the Lienhuachih forest. Different letters among habitats in each sampling month denote a significant difference by Scheffé's test at p < 0.05.
育地專一的大丁黃(Euonymus laxiflorus) πtlp低 至-2.98 MPa,而南投石櫟及台灣紅豆樹的πtlp
都是-2.87 MPa,此3樹種葉部耐旱性都極高。
供試樹種πtlp在-2.79~-2.60 MPa另有11樹種,包 括桃實百日青、杏葉石櫟、小葉赤楠(Syzygium buxifolium)等,這些樹種的葉部耐旱性也很高 (Table 4)。供試樹種中πtlp在-2.59~-2.50 MPa者 計有15種;πtlp在-2.49~-2.20 MPa者計有17種;
πtlp高於-2.20 MPa者有6種,以溪谷生育地專一 的山龍眼、五掌楠(Neolitsea konishii)、水同木 (Ficus fistulosa)及樹杞(Ardisia sieboldii)的葉 部耐旱性最低(Table 4)。供試52樹種中有46種 於乾、雨兩季間πtlp具顯著差異,季節性馴化明 顯;但只有34樹種在乾、雨兩季間π0具顯著差 異(Table 4)。
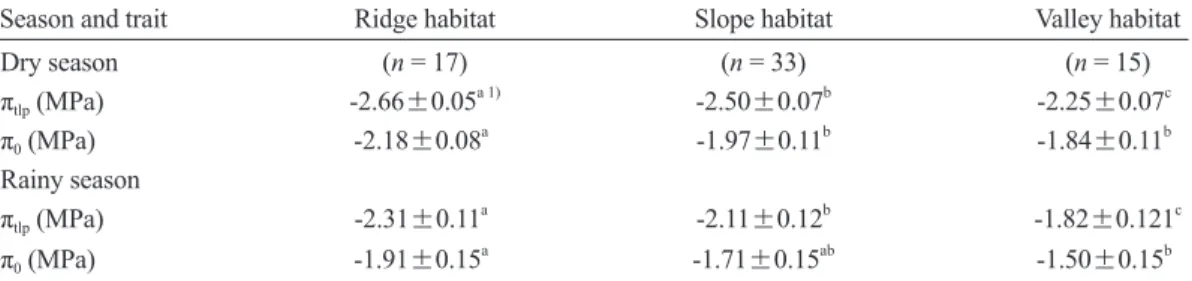
比 較 蓮 華 池 森 林 在 不 同 生 育 地 樹 種 群 聚 間πtlp及π0的差異,發現乾季期間群聚在山脊、
坡 面 、 溪 谷 三 處 生 育 地 樹 種πt l p的 平 均 值 分 別為-2.66±0.05、-2.50±0.07、-2.25±0.07 MPa,三樹種群聚間均具顯著差異,以山脊樹 種πtlp顯著最低(Table 5),亦即群聚在山脊樹種 之葉部耐旱性顯著最高。乾季期間群聚在山脊 生育地樹種的π0會顯著低於群聚在坡面及溪谷 兩生育地的樹種(Table 5)。在雨季期間三處生 育地樹種群聚間的πtlp也均具顯著差異,同樣是 以群聚在山脊樹種的πtlp顯著最低(Table 5)。此 外,合併所有樹種乾季期間的πtlp均顯著低於雨 季時(Fig. 2),平均降低0.39±0.03 MPa,顯示 蓮華池森林樹種的πtlp也具有季節性馴化現象。
在雨季期間群聚在山脊樹種的π0顯著低於群聚 在溪谷的樹種(Table 5)。合併所有樹種,乾季 期間的π0較雨季期間也顯著降低(Fig. 2),平均 降低0.28±0.03 MPa。本研究也發現蓮華池森 林供試52樹種無論在乾季或在雨季,同一季節 Table 2. Comparisons of the predawn leaf water potential (MPa) between saplings of the same species growing in the ridge and valley habitats during a severe drought period in the Lienhuachih forest (mean±standard error, n = 5)
Species Ridge habitat Valley habitat
Ormosia formosana -1.14±0.2a 1) -0.76±0.14a
Ardisia quinquegona -1.53±0.08a -1.01±0.16b
Illicium arborescens -1.76±0.05a -1.15±0.26b
Neolitsea aciculata var. variabillima -1.80±0.04a -1.17±0.04b
Psychotria rubra -1.82±0.06a -1.41±0.15b
Cinnamomum subavenium -1.87±0.09a -1.19±0.11b
Blastus cochinchinensis -1.99±0.11a -1.50±0.06b
Cryptocarya chinensis -2.06±0.08a -1.35±0.09b
1) Different letters between ridge and valley habitats denote a significant difference according to a t-test at p < 0.05.
Table 3. Comparisons of the leaf water potential at the turgor loss point (MPa) of the same species growing in different habitats during a severe drought period in the Lienhuachih forest (mean±standard error, n = 5)
Species Ridge habitat Slope habitat Valley habitat Podocarpus nakaii -2.79±0.02a 1) -2.49±0.01b -2.41±0.03b Castanopsis fargesii -2.79±0.02a -2.58±0.01b -2.45±0.01c Meliosma squamulata -2.66±0.04a -2.49±0.01b -2.40±0.02b Lithocarpus harlandii -2.65±0.04a -2.49±0.01b -2.45±0.03b Schefflera octophylla -2.47±0.03a -2.21±0.03b -2.19±0.01b
Psychotria rubra -2.33±0.02a -2.35±0.03a -2.22±0.01b
1) Different letters among habitats denote a significant difference according to Scheffé’s test at p < 0.05.
Table 4. Major habitat association (Hab), leaf water potential at the turgor loss point (πtlp), and osmotic potential at saturation (π0) during the dry and the rainy seasons for the 52 tested species in the Lienhuachih forest (mean±standard error, n = 5)
Species Hab πtlp (MPa) π0 (MPa)
Dry Rainy Dry Rainy
Euonymus laxiflorus R1) -2.98±0.10 -2.21±0.04*2) -2.34±0.04 -1.95±0.03*
Lithocarpus nantoensis R -2.87±0.05 -2.55±0.05* -2.37±0.09 -2.23±0.04 Ormosia formosana R, S -2.87±0.01 -2.81±0.04 -2.62±0.02 -2.54±0.09 Podocarpus nakaii R, S -2.79±0.02 -2.34±0.08* -2.10±0.06 -1.80±0.07*
Lithocarpus amygdalifolius S -2.78±0.02 -2.39±0.06* -2.06±0.07 -2.01±0.06 Syzygium buxifolium R -2.77±0.02 -2.25±0.08* -2.35±0.10 -1.81±0.04*
Lithocarpus konishii S -2.74±0.04 -2.33±0.08* -2.00±0.11 -1.80±0.03 Castanopsis uraiana S -2.71±0.02 -2.50±0.04* -2.09±0.02 -1.75±0.21 Gordonia axillaris R -2.69±0.03 -2.24±0.04* -2.10±0.02 -1.72±0.06*
Meliosma squamulata R, S -2.66±0.04 -2.23±0.06* -2.10±0.04 -1.96±0.04*
Randia cochinchinensis R -2.65±0.02 -2.58±0.07 -2.05±0.10 -2.02±0.03 Eustigma oblongifolium S -2.63±0.04 -2.10±0.09* -2.03±0.03 -1.58±0.06*
Quercus pachyloma R -2.60±0.04 -2.59±0.04 -2.28±0.07 -2.27±0.04 Ilex goshiensis R -2.60±0.03 -1.97±0.04* -2.21±0.03 -1.52±0.05*
Castanopsis cuspidata var. carlesii R, S -2.59±0.02 -2.12±0.04* -1.91±0.07 -1.65±0.04*
Tricalysia dubia S -2.59±0.03 -2.22±0.03* -2.02±0.04 -1.95±0.04 Pyrenaria shinkoensis S -2.59±0.01 -2.11±0.07* -2.15±0.02 -1.69±0.04*
Rhododendron leptosanthum R -2.58±0.03 -1.94±0.06* -1.87±0.02 -1.41±0.04*
Castanopsis fargesii S -2.58±0.01 -2.31±0.09* -2.12±0.04 -1.96±0.06*
Castanopsis kawakamii S -2.58±0.03 -2.24±0.09* -1.90±0.03 -1.56±0.05*
Engelhardia roxburghiana S -2.58±0.04 -2.06±0.07* -2.20±0.03 -1.65±0.04*
Neolitsea aciculata var. variabillima R, S -2.57±0.02 -2.44±0.04* -2.11±0.04 -2.07±0.04 Schima superba S -2.57±0.03 -2.09±0.03* -2.05±0.06 -1.56±0.02*
Elaeocarpus sylvestris S -2.57±0.02 -2.09±0.06* -2.15±0.04 -1.75±0.03*
Elaeocarpus japonicus R -2.56±0.03 -2.52±0.05 -2.18±0.04 -2.13±0.04 Lithocarpus synbalanos R -2.55±0.04 -2.42±0.03* -2.06±0.04 -2.04±0.02 Distyliopsis dunnii R, S -2.55±0.04 -2.11±0.06* -2.07±0.04 -1.68±0.08*
Machilus thunbergii S -2.54±0.03 -1.99±0.02* -2.35±0.02 -1.83±0.04*
Ilex ficoidea S -2.51±0.02 -2.05±0.07* -1.76±0.04 -1.67±0.04 Lithocarpus harlandii S -2.49±0.01 -2.26±0.02* -1.80±0.03 -1.77±0.09 Cinnamomum subavenium S -2.46±0.01 -2.34±0.04* -2.15±0.04 -2.11±0.06 Litsea acuminata S, V -2.46±0.04 -2.02±0.04* -2.15±0.06 -1.75±0.04*
Styrax suberifolius S, V -2.45±0.02 -1.77±0.05* -1.83±0.03 -1.50±0.02*
Helicia cochinchinensis S -2.43±0.02 -2.22±0.05* -1.95±0.04 -1.89±0.02 Cinnamomum osmophloeum V -2.42±0.04 -1.81±0.03* -1.96±0.03 -1.62±0.02*
Helicia rengetiensis R, S -2.41±0.02 -1.94±0.06* -1.93±0.06 -1.72±0.07*
Diospyros morrisiana S -2.41±0.02 -2.04±0.07* -2.17±0.04 -1.73±0.03*
Machilus japonica var. kusanoi V -2.41±0.03 -2.36±0.01 -2.05±0.03 -2.00±0.01 Cryptocarya chinensis S, V -2.37±0.09 -1.87±0.04* -2.11±0.07 -1.72±0.07*
con’t
Psychotria rubra S -2.35±0.03 -1.86±0.04* -2.03±0.08 -1.63±0.05*
Wendlandia formosana S, V -2.29±0.05 -1.77±0.04* -1.75±0.06 -1.42±0.03*
Eurya loquaiana V -2.27±0.03 -1.69±0.05* -1.84±0.01 -1.22±0.05*
Machilus zuihoensis V -2.25±0.03 -1.70±0.04* -2.03±0.08 -1.39±0.04*
Ardisia quinquegona S -2.23±0.06 -2.12±0.04 -2.02±0.07 -1.97±0.06 Saurauia tristyla var. oldhamii V -2.23±0.02 -1.51±0.04* -1.79±0.03 -1.16±0.02*
Schefflera octophylla S, V -2.21±0.03 -2.01±0.01* -1.79±0.09 -1.47±0.04*
Melicope pteleifolia S, V -2.17±0.04 -1.59±0.08* -1.75±0.05 -1.39±0.06*
Symplocos theophrastifolia S -2.13±0.06 -1.67±0.03* -1.83±0.05 -1.48±0.01*
Helicia formosana V -2.09±0.08 -1.61±0.03* -1.56±0.03 -1.47±0.03*
Neolitsea konishii V -2.08±0.02 -1.88±0.09* -1.88±0.04 -1.53±0.08*
Ficus fistulosa V -2.07±0.01 -2.00±0.03* -1.70±0.08 -1.65±0.02 Ardisia sieboldii V -2.03±0.04 -1.96±0.08 -1.72±0.03 -1.46±0.08*
1) R, ridge habitat; S, slope habitat; V, valley habitat.
2) * Indicates a significant difference between seasons (t-test, p < 0.05).
各樹種的πtlp與π0間均具極顯著正相關(Fig. 3a, b) (p < 0.001),且兩季節間的πtlp或π0也均具極顯著 正相關(Fig. 3c, d) (p < 0.001)。
乾旱嚴重時期山脊18樹種成樹的黎明前葉部 水勢
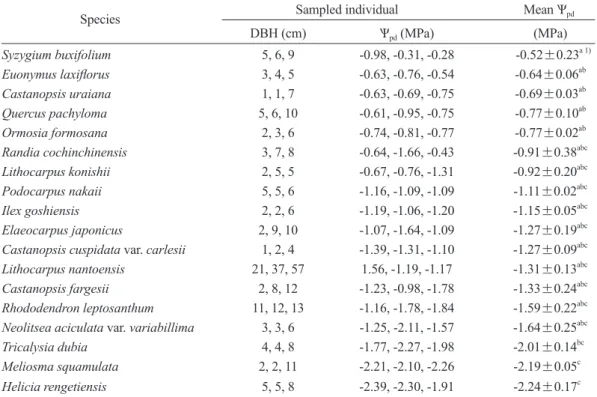
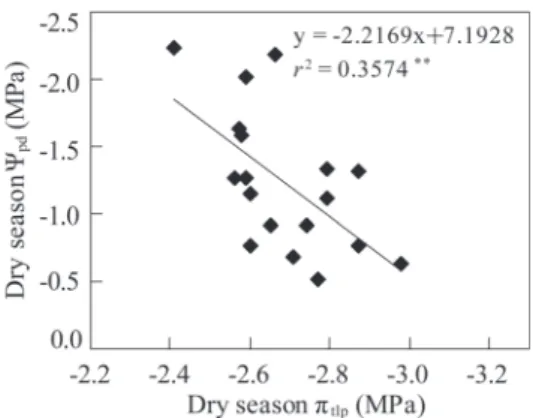
蓮華池山脊稜線18樹種的成樹,在2019年 2月乾季最嚴重期間測得黎明前葉部水勢(Ѱpd) 的分布範圍在-0.52~-2.24 MPa之間(Table 6)。
供 試 樹 種 間Ѱpd數 值 差 異 雖 很 大 , 但 因 為 大 多 數樹種同種3植株間的Ѱpd差異也很大,因此這 18種供試樹種中有15種的Ѱpd平均值在統計上 不具顯著差異(Table 6)。小葉赤楠、大丁黃、
烏來柯、捲斗櫟(Quercus pachyloma)及台灣紅 豆樹,此5樹種在經歷5個月極度乾旱後僅遭輕 度缺水逆境,Ѱpd平均值仍可保持在-0.52~-0.77 MPa,且各單株的Ѱpd均 高 於-1.00 MPa。此 外,上述5樹種的Ѱpd顯著高於狗骨仔、綠樟及 蓮華池山龍眼(Helicia rengetiensis)。後3樹種的 Ѱpd平均值均低於-2.00 MPa,顯示已遭嚴重缺水 逆境。進一步分析這18樹種Ѱpd與乾季πtlp間之關 係,發現此兩生理參數間具極顯著負相關(r2 = 0.36, p < 0.01) (Fig. 4)。此結果顯示分布在該山 脊生育地葉部耐旱性較高的樹種,在乾旱嚴重 期間植株仍可維持較高的水分狀態。
討 論
不同生育地的水分有效性
蓮 華 池 森 林 在 海 拔8 2 0 ~ 8 3 0 m 山 脊 生 育 地 , 乾 季 時 的 土 壤 含 水 率 顯 著 低 於 坡 面 (730~740 m)及溪谷生育地(690~700 m),顯示 不同海拔高的生育地有差異明顯的水分有效性 梯度。在山地地形,山脊上坡的土壤水分較下 Fig. 2. Seasonal comparisons of the leaf
water potential at the turgor loss point (πtlp) and osmotic potential at saturation (π0) for all 52 tested species combined. Different letters between seasons denote a significant difference by a t-test at p < 0.001.
Fig. 3. Relationships between the leaf water potential at the turgor loss point (πtlp) and the osmotic potential at saturation (π0) in the dry (a) and rainy seasons (b), πtlp between the 2 seasons (c), and π0 between the 2 seasons (d) for all 52 tested species. *** p < 0.001.
坡溪谷少的原因,一方面是降雨時山脊的水分 因重力快速滲流至中、下坡處,山脊上坡土壤 能保有的水分較少;另一方面是因山脊上坡較 常暴露在氣流中,水分由地表喪失的程度較下 坡高,因此山脊上坡的土壤水分有效性通常較
下坡低(Becker et al. 1988)。例如巴拿馬半落葉 熱帶林,在山脊生育地的土壤水分有效性顯著 低於坡面,且在每年的乾季,山脊生育地乾旱 的日數會多於坡面(Daws et al. 2002)。另一方 面,因為植株的黎明前葉部水勢(Ѱpd)可指示生 Table 5. Comparisons of the leaf water potential at the turgor loss point (πtlp) and osmotic potential at saturation (π0) during the dry and rainy seasons among different habitat associations in the Lienhuachih forest (mean±standard error)
Season and trait Ridge habitat Slope habitat Valley habitat Dry season (n = 17) (n = 33) (n = 15) πtlp (MPa) -2.66±0.05a 1) -2.50±0.07b -2.25±0.07c π0 (MPa) -2.18±0.08a -1.97±0.11b -1.84±0.11b Rainy season
πtlp (MPa) -2.31±0.11a -2.11±0.12b -1.82±0.121c π0 (MPa) -1.91±0.15a -1.71±0.15ab -1.50±0.15b
1) Different letters for each trait among the habitats denote a significant difference according to Scheffé’s test at p
< 0.001.
育地的土壤水分有效性(Stahl et al. 2013, Zhou et al. 2014, Mitchell and O’Grady 2015),而本 研究於乾季期間測得相同樹種生長在山脊稚樹 的植株,其Ѱpd會顯著低於溪谷生育地同種稚樹 的植株,這也顯示海拔較高之山脊生育地的水 分有效性,顯著低於海拔較低處的生育地。臺 灣紅豆樹生長在山脊稚樹的Ѱpd與生長在溪谷的 稚樹並無顯著差異(-1.14 vs. -0.76 MPa),且本 樹種兩處稚樹的Ѱpd都高於另7樹種的稚樹(Table 2)。生長在相同生育地某樹種植株若有顯著較 高 的 水 分 狀 態 , 很 可 能 是 該 樹 種 有 分 布 較 深 的根系,可吸收較深層的土壤水分(Kuo 1994, Fallon and Cavender-Bares 2018)。因此,臺灣 紅豆樹稚樹的根系深度可能較其它供試樹種的 稚樹深。此外,本研究曾測定過臺灣紅豆樹與 變葉新木薑子此兩樹種生長在山脊相同生育地 之稚樹與成樹的Ѱpd,該兩樹種稚樹的Ѱpd分別 為-1.14及-1.80 MPa (Table 2),成樹的Ѱpd則分
別為-0.77及-1.64 MPa (Table 6),成樹在乾旱更 嚴重時仍比稚樹有較高的Ѱpd,顯示成樹根系可 能較深,吸收土壤水分的能力較稚樹高。在比 較山脊與溪谷兩處生育地水分有效性時,若用 成樹而非稚樹當樣木,或許仍可呈現山脊之水 分有效性顯著低於溪谷,但兩處水分差異的程 度可能比用稚樹當樣木低。
樹種葉部耐旱性等級區分
樹 種 的 耐 旱 性 是 由 形 態 、 生 理 及 生 化 等 複雜的性狀所構成(Farooq et al. 2009),而木 本植物的耐旱機制又有延遲脫水及忍受脫水兩 機制,單獨或共同作用(Kramer 1983, Volaire 2018)。Niinemets and Valladares (2006)曾藉 由樹種分布處的生育地特性及樹種的生理潛力 兩項性狀,去量化北半球溫帶樹種的耐旱性分 數,並將樹種耐旱性區分為五等級。因為不同 樹 種 耐 旱 的 機 制 及 對 缺 水 逆 境 的 反 應 極 為 複
Table 6. Diameter at breast height (DBH) and predawn leaf water potential (Ѱpd) of each sampled individual, and interspecific comparisons of mean Ѱpd values among the 18 tree species growing in the ridge habitat of the Lienhuachih forest (mean±standard error, n = 3)
Species Sampled individual Mean Ѱpd
DBH (cm) Ѱpd (MPa) (MPa)
Syzygium buxifolium 5, 6, 9 -0.98, -0.31, -0.28 -0.52±0.23a 1) Euonymus laxiflorus 3, 4, 5 -0.63, -0.76, -0.54 -0.64±0.06ab Castanopsis uraiana 1, 1, 7 -0.63, -0.69, -0.75 -0.69±0.03ab Quercus pachyloma 5, 6, 10 -0.61, -0.95, -0.75 -0.77±0.10ab Ormosia formosana 2, 3, 6 -0.74, -0.81, -0.77 -0.77±0.02ab Randia cochinchinensis 3, 7, 8 -0.64, -1.66, -0.43 -0.91±0.38abc Lithocarpus konishii 2, 5, 5 -0.67, -0.76, -1.31 -0.92±0.20abc Podocarpus nakaii 5, 5, 6 -1.16, -1.09, -1.09 -1.11±0.02abc Ilex goshiensis 2, 2, 6 -1.19, -1.06, -1.20 -1.15±0.05abc Elaeocarpus japonicus 2, 9, 10 -1.07, -1.64, -1.09 -1.27±0.19abc Castanopsis cuspidata var. carlesii 1, 2, 4 -1.39, -1.31, -1.10 -1.27±0.09abc Lithocarpus nantoensis 21, 37, 57 1.56, -1.19, -1.17 -1.31±0.13abc Castanopsis fargesii 2, 8, 12 -1.23, -0.98, -1.78 -1.33±0.24abc Rhododendron leptosanthum 11, 12, 13 -1.16, -1.78, -1.84 -1.59±0.22abc Neolitsea aciculata var. variabillima 3, 3, 6 -1.25, -2.11, -1.57 -1.64±0.25abc Tricalysia dubia 4, 4, 8 -1.77, -2.27, -1.98 -2.01±0.14bc Meliosma squamulata 2, 2, 11 -2.21, -2.10, -2.26 -2.19±0.05c Helicia rengetiensis 5, 5, 8 -2.39, -2.30, -1.91 -2.24±0.17c
1) Different letters among the species denote a significant difference according to Scheffé’s test at p < 0.05.