行政院國家科學委員會補助專題研究計畫成果報告
Halobacterium halobium 紫色細胞膜與 bacteriorhodopsin 蛋白質於生物光電晶片之研究 (3/3)
計畫類別:█ 個別型計畫 □ 整合型計畫
計畫編號:NSC 94- 2214-E - 011 - 003-
執行期間: 94 年 8 月 1 日至 95 年 7 月 31 日
計畫主持人:陳秀美
計畫參與人員:謝謹謙、謝竺君、謝育卿、王世育
成果報告類型(依經費核定清單規定繳交):□精簡報告 █完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計 畫、列管計畫及下列情形者外,得立即公開查詢
█涉及專利或其他智慧財產權,□一年█二年後可公開查詢
執行單位:國立台灣科技大學 化學工程學系
中 華 民 國 95 年 10 月 20 日
II
中文摘要
細菌視紫質(bacteriorhodopsin,bR) 為一個具有光驅動質子唧筒功能的蛋白 質,存在於 Halobacterium halobium 的紫色細胞膜 (purple membrane,PM)內。本
文主要的研究目的在於建立一個電化學光電系統,以量測PM 溶液受光激發後所引
起的電流變化。實驗中先由基本的比色管光電化學系統進行探討,以尋求系統的最 適化操作條件與參數;而後再將此條件應用到 PDMS 陣列系統,並比較此二系統
之差異。由初步的比色管實驗,發現工作電極以ITO 玻璃、逆電極以白金棒、導電
介質為銅片時最為適當,其光電流訊號值在微安培 (μA) 的範圍內;而將相同的操
作條件應用到PDMS 陣列系統也同樣可行。文中並就光電流訊號產生的機制進行探
討。對比色管系統進一步探討,發現對系統最適合的PM 酸鹼值為 6;而電解液之
最適值則是 7;且在不同酸鹼值下 PM 細胞膜的外觀有所變化。同時在固定 PM 溶 液量下, PM 的濃度與燈源瓦數不會影響光電流訊號的大小;照光頻率越高,光電 流訊號值會越小;而藍色與黃色濾片所測得之光電流訊號會較綠色及紅色濾片為 大。以比色管與 PDMS 陣列系統做比較可以發現,PM 體積的大小以及電極與銅片 間的距離皆會影響光電流訊號的大小;且在相近的 PM 用量下由於系統尺寸的縮 小,PDMS 陣列系統可較比色管系統測得較大的光電流訊號值。此光電化學系統的 建立對於 bR 感光材料的應用上,樹立相當程度的研發基礎。
關鍵詞:細菌視紫質、紫色細胞膜、電化學光電
Abstract
Bacteriorhodopsin (bR) is a protein of light-driven proton pump located in the purple membrane (PM) of Halobacterium halobium. In this study, two photo-electrochemical systems were set up to measure the photocurrent produced from illuminated PM. A cuvette system was first used to study the optimal operation condition including the intermediate, the pH values of PM and electrolyte solution, PM concentration, and the incident light intensity, which was subsequently applied to a PDMS array system. The cuvette study showed the highest photocurrent, which was in a range of microamperes, was achieved when ITO glass as the working electrode, a Pt wire as the counter electrode, and a Cu film as the intermediate were used. The production mechanism of the photocurrent was proposed according to its signal profile. The photocurrent of the PDMS array system was also obtained when the same electrodes/intermediate setup as that of the cuvette system was built. The studies on the cuvette system also revealed an optimal operation pH value of PM as 6, while the light intensity and the PM concentration did not have any effect on photocurrents if the same PM amount was used. The comparison between the cuvette and PDMS systems suggested that the distances between the electrodes and the Cu intermediate film determined the photocurrent values, yielding a higher signal in the latter system than in the former one if the same PM amount was used. This study provides valuable informations for the future photoelectric applications of the photosensitive bR.
Key words: bacteriorhodopsin, purple membrane, photo-electrochemical
IV
目錄
中文摘要··· Ⅱ
英文摘要··· Ⅲ
目錄··· Ⅳ
第一章 前言··· 1
第二章 研究目的··· 2
第三章 文獻回顧··· 3
3-1 Halobacterium halobium··· 3
3-2 Bacteriorhodopsin (bR)··· 6
3-3 PM 的光電流訊號··· 18
3-4 PDMS (poly-dimethylsiloxane) ··· 25
第四章 研究方法··· 27
4-1 實驗藥品 ··· 27
4-2 實驗設備 ··· 28
4-3 實驗流程 ··· 30
4-4 實驗步驟 ··· 31
第五章 結果與討論··· 38
5-1 菌種培養及觀察··· 38
5-2 PM 純化程序及其特性 ··· 39
5-3 光電化學系統··· 41
第六章 結論··· 92
第七章 參考文獻··· 94
第八章 計畫成果自評………101
研發成果資料表………..102
第一章 前言
Halobacterium halobium 為一可以生存在鹽濃度高於 12 % 環境下的嗜鹽 菌體,在飽和鹽濃度的環境下也能生存,所以在鹽湖、鹽田或死海等高鹽分的地方,
都能發現它的蹤跡。H. halobium 是兩端都有著鞭毛的桿菌,其細胞膜主要由蛋白質 及少數的碳水化合物所構成,細胞膜是呈暗紅色的,可將其進一步分離成紅色細胞膜 (red membrane,RM)以及細胞只有在低氧環境下生長時才會形成的紫色細胞膜(purple membrane,PM)。H. halobium 在有充足氧氣的環境下,絕大部分在生長時都是形成 RM;而在缺氧及光照的環境下生長時,則是可以形成含量一半以上的 PM。
此菌特別的地方就在於,當環境為缺氧的狀態,而菌體無法行正常的新陳代謝 時,它可以將光能轉換成化學能,形成一個橫跨細胞膜的氫離子濃度梯度,然後促進 ATP 的合成,以提供細胞正常活動時所需之能量(Henderson, 1977)。此能力的來源就 在於 H. halobium 的 PM 紫色細胞膜中,存在一據有光驅動質子唧筒功能的蛋白質,
bacteriorhodopsin (bR)。bR 由於有著一個發色基(chromophore) ─ retinal 的存在,所 以據有光驅動質子唧筒(light-driven proton pump)的功能。bR 的存在,對於無法行呼 吸作用時的 H. halobium 來說,是一個可以緊急驅動,而使得菌體可以繼續生存下去 的一種生命維持裝置。
由於 bR 有此可以將光能轉換成化學能的光驅動質子唧筒能力,且其蛋白質結 構穩定,有極高的熱穩定性,不容易失去活性,不具污染性,所以開始受到奈米生物 材料應用上的重視。bR 對光的敏感性,及其特殊的光循環機制,亦使得 bR 成為新 興感光材料上重要的一環。
本文主要的研究目的在於建立一個光電化學系統,可以直接量測PM溶液受光激 發後所引起的電流變化;其次則是將 bR與 PDMS (poly-dimethylsiloxane)結合,以縮 小光電化學系統的尺寸。PDMS 為一種具有高透光性的高分子材料。它有價格便宜,
方便且可快速製作,且具有非常高的透光性 (能穿透波長大約在 190-700 nm 間的光) (Jo et al., 2000)的優點;此外,具有著非常高的生化及化學穩定性,其穩定溫度可以 高達 250℃,所以非常適合做為各種微小裝置的基材(Armani et al., 1999),故選擇其 為光電化學系統的材料。實驗中將進一步的找出適合PM 溶液之酸鹼值環境,且探討 PM 濃度、燈源瓦數、照光頻率與不同顏色光源對光電流訊號的影響。
第二章 研究目的
H. halobium S-9 嗜鹽菌 PM 細胞膜內的 bR 蛋白質具有光驅動質子唧筒功能,
可廣泛應用於光感測器、影像偵測器等舉凡有關將光能轉換成電能的領域,更由於其 特殊的光循環機制而可以更延伸應用於生物記憶體及影像辨色系統等。
為了能應用 bR 做為生物材料,了解其基本性質就顯得格外重要。本文主要的研 究目的在於建立一個光電化學系統,可以直接量測PM 紫色細胞膜溶液受光激發後所 引起的光電流變化,由於細胞膜無須固定化,因而可以避免改變甚至破壞 bR 蛋白 質的結構與特性,提升系統的穩定與重複使用性。
由於所純化的 PM 細胞膜為溶液狀,所以本研究在初步設計光電化學系統時,
是以電化學系統為雛型。逆電極選擇穩定性最高的白金,工作電極的考量則因為不能 阻礙到光照,所以我們選擇透明且導電的 ITO 玻璃來做為系統的工作電極。
實驗中將先由基本的比色管光電化學系統進行探討,以尋求系統的最適化操作條 件。實驗中最重要的第一步為先找出適合光電化學系統的中間導電介質,其角色類似 電化學電池中的鹽橋,作用為使整個系統形成一個完整的迴路。建立出初步的光電化 學系統後,則進一步地探討適合系統的 PM 溶液與電解質之酸鹼值,找出適合的操 作環境,其次探討 PM 濃度、燈源瓦數、照光頻率與光源顏色對光電訊號的影響。
最後,建構與比色管系統原理相同的 PDMS 陣列系統以將系統微小化,並再進一步 比較其兩者之優缺點。
第三章 文獻回顧
3-1 Halobacterium halobium
3-1-1 菌體介紹
Halobacterium halobium 為一可以生存在鹽濃度高於 12 % 環境下的菌體,甚至 在飽和鹽濃度的環境下也能生存的非常好。其最適合生長的環境是在鹽濃度為 25 % (wt/vol)的環境中,所以在鹽湖、鹽田或死海等高鹽分的地方,都能發現它的蹤跡。
H. halobium 是兩端都有著鞭毛的桿菌 (Fig. 3-1),其細胞膜主要由蛋白質及少數 的碳水化合物所構成。細胞膜是呈暗紅色的,可將其進一步分離成紅色細胞膜(red membrane,RM)以及細胞只有在低氧環境下生長時才會形成的紫色細胞膜(purple membrane,PM)。H. halobium 在有充足氧氣的環境下,絕大部分在生長時都是形成 RM;而在缺氧及光照的環境下生長時,則是可以形成含量一半以上的 PM。因此我 們不只可以在菌體生長時,製造低氧的環境來促使細胞合成 PM,更可額外提供光 源,來增加 PM 在每個細胞上的含量。
此菌特別的地方就在於,當環境為缺氧的狀態、而無法行正常的新陳代謝時,它 可以將光能轉換成化學能,形成一個橫跨細胞膜的氫離子濃度梯度,然後促進 ATP 的合成,以提供細胞正常活動時所需之能量(Henderson, 1977)。
3-1-2 紫色細胞膜 ( PM )
H. halobium 的 PM 紫色細胞膜中,存在一具有光驅動質子唧筒功能的蛋白質,
bacteriorhodopsin (bR)。bR 由於有著一個發色基(chromophore) ─ retinal 的存在,所 以具有此特殊功能。
由化學分析可以知道,PM 中的成分為 75 % 的蛋白質與 25 % 的脂質(lipid)。
而脂質的成分有 phosphatidyl glycerophosphate (52 %)、 phosphatidyl glycerol (4 %)、
largely squalenes (9 %) 、photphatidylglycerosulfate (5 %)、 glycolipid sulfate (10 %)、
triglycosyl diether (19 %) 與其他一些 glycolipid。而 RM 中的脂質成分則與 PM 的有 著些許的不同,我們將 PM 做蛋白質電泳的分析,發現只有單一個明顯的色帶,這 表示在 PM 中只存在著一種蛋白質,而且其分子量估算大約為 26,000 Da,此即 bR 蛋白質。而在 RM 中則包含了 25 種以上的蛋白質。由此可以推測,RM 上的蛋白 質包辦了細胞需正常活動的所有機能,而 PM 則只有在缺氧且照光時,啟動光驅動 質子唧筒這唯一的功能(Henderson, 1977)。
Fig. 3-1 H. halobium is a rod-shaped bacterium normally flagellated both ends. The white is the red membrane, and the gray is the purple membrane.
(http://omega.ilce.edu.mx:3000/sites/ciencia/volumen2/ciencia3/092/img)
bR 是由嗜鹽菌體 H. halobium 在低氧環境下所合成的,它是以二維六角晶格結 構排列在菌體的細胞膜上(Fig. 3-2)。每一個六角晶格是由三個蛋白質所形成的三聚體 (trimer),而蛋白質之間則為可以穩定結構的脂質。這些脂質對於 bR 來說是非常重 要的,除了穩定結構之外,在 bR 推出質子時還有調節 pKa 值的功能(Hendler and Dracheva, 2001)。
在 Henderson (1977) 的文獻回顧中有提到,Blaurock 與 Stoeckenius (1971)是最 先觀察到 PM 中的蛋白質與一些脂質為規則性排列的人。他們以 X-ray 觀察在雲母 基材上乾燥的 PM,發現在平行於細胞膜的平面上,有六角形排列的物質,且是遍及 整個 PM 的表面,而膜的厚度大約只有 49 Å (Henderson, 1977; Lanyi and Luecke, 2001)。Qutub 等人在 2004 年時也用 lipidic cubic phase 的方式,讓 bR 在他們所設 計的 Cryoloop 上呈現六角晶格狀(Fig. 3-3),他們且更進一步地以 AFM (atomic force microscopy)分析探討 bR 結晶的情形。而 Nollert 在 2004 年曾針對 lipidic cubic phase 法於膜蛋白結晶的應用上做詳盡的說明及回顧(Fig. 3-4)。
Fig. 3-2 Arrangement of bR and lipids in 3D crystals grown in the cubic lipid phase (Lanyi and Luecke, 2001).
Fig. 3-3 An optical microscopy image of a hexagonal bR crystal grown in monoolein cubic phase (Qutub et al., 2004).
Fig. 3-4Growth of bR crystals within a monoolein-based lipidic cubic phase matrix in a glass vial. (A) The 10 µL crystallization experiment was set up in a glass vial. (B) bR microcrystals formed within 1 month. (C) Schematic close-up of purple bR crystals with sizes up to ca. 100 µm along their longest dimension (Nollert, 2004).
3-2 Bacteriorhodopsin (bR)
3-2-1 bR 之結構
bR 的存在,對於無法行呼吸作用時的 H. halobium 來說,是一個可以緊急驅動 而使得菌體可以繼續生存下去的一種光驅動質子唧筒(light-driven proton pump)。它可 以將光能轉換成質子梯度來啟動一連串的反應,以利菌體的生存;而由吸收光譜可以 知道 bR 的最大吸收波長大約在 568 nm。
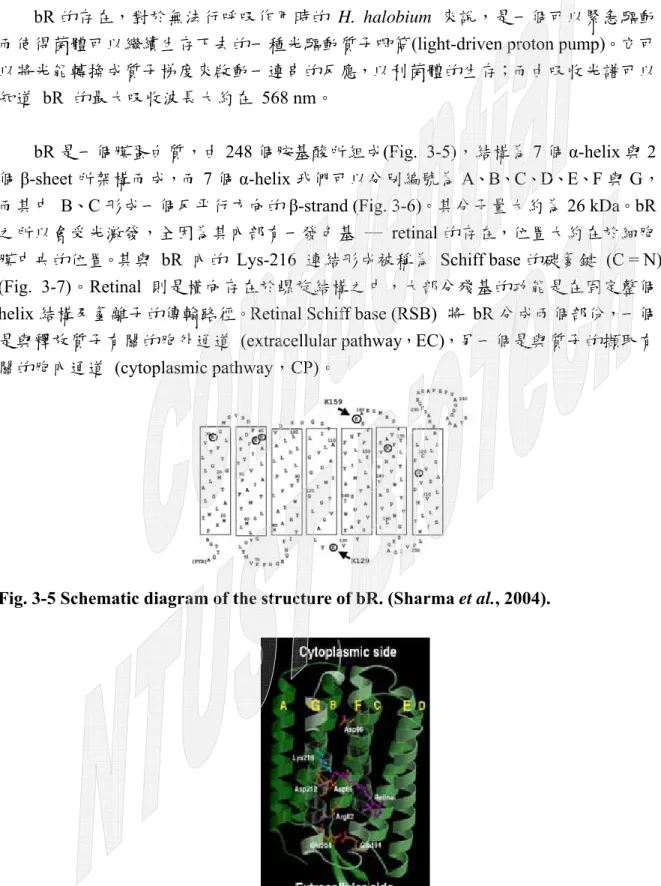
bR 是一個膜蛋白質,由 248 個胺基酸所組成(Fig. 3-5),結構為 7 個 α-helix 與 2 個β-sheet 所架構而成,而 7 個 α-helix 我們可以分別編號為 A、B、C、D、E、F 與 G,
而其中 B、C 形成一個反平行方向的 β-strand (Fig. 3-6)。其分子量大約為 26 kDa。bR 之所以會受光激發,全因為其內部有一發色基 ─ retinal 的存在,位置大約在於細胞 膜中央的位置。其與 bR 內的 Lys-216 連結形成被稱為 Schiff base 的碳氮鍵 (C = N) (Fig. 3-7)。Retinal 則是橫向存在於螺旋結構之中,大部分殘基的功能是在固定整個 helix 結構及氫離子的傳輸路徑。Retinal Schiff base (RSB) 將 bR 分成兩個部份,一個 是與釋放質子有關的胞外通道 (extracellular pathway,EC),另一個是與質子的擷取有 關的胞內通道 (cytoplasmic pathway,CP)。
Fig. 3-5 Schematic diagram of the structure of bR. (Sharma et al., 2004).
在retinal 朝向胞外的那端,有著許多極性的殘基(如:Asp-36、Asp-102、Asp-104、
Lys-159、Glu-161、Arg-164、Glu-166、Arg-227),他們扮演著讓 Schiff base 去質子化 及釋放質子到細胞膜表面的角色,比較重要的有 Asp-85、Arg-82 及較靠近的 Glu-194 及Glu-204。而在朝胞內的那端,卻沒有發現可以將 retinal 再質子化的結構分子。在 高解析的結構圖(Fig. 3-8)上我們可以發現,有一個水分子 Wat-402 將質子提供者 Schiff base,與質子接受者 Asp-85 區隔開來,在胞外的區域包含 7 個水分子,與一些有極 性的側鏈,有Asp-85,Asp-212,Thr-89,Trp-86,Tyr-83,Tyr-57,Tyr185,Arg-82,
Glu-194,Glu-204 與 Ser-193,而形成一個三維的氫鍵鍵結結構,連結 Schiff base 與 細胞膜表面(Lanyi, 2004)。
Fig. 3-7 X-ray crystallographic structures of the Schiff base region in bR (Kandori, 2004).
Fig. 3-8 Structure of bR (Lanyi, 2004).
3-2-2 bR 的光循環機制
bR 在受光激發後,其結構會變化成一連串的中間產物,至今能確定且較穩定的 激發態包括:K、L、M、N 及 O State (Fig. 3-9)。
Fig. 3-9 The photocycle of bR (Heberle et al., 2000).
其實從十七世紀,bR 被發現後至今,其光循環機制仍然是眾說紛紜,如光循環 是單一循環,或是多循環;是可逆方向性的,還是無方性的,到現在都還是受爭議的 議題。既然沒有一個可完美解釋的光循環機制,現在大部分都以單一且可逆的光循環 機制,及已經非常確定結構的各個基本中間介質,來當作基本的機制 (Lanyi, 2004)。
Fig. 3-10 Photocycle scheme of bR with proton release (Lanyi, 2004).
Fig. 3-10 為單一且可逆的光循環機制,最主要的中間介質為 M 及 N state。M state共分三種,首先是因 L state 的 RSB 去質子化而形成的 M1。在 M1 中 RSB 因 質子化而與胞外環境隔絕,與胞內連接,形成 M2。而後bR推出質子到胞外,形成 M2´,Schiff base 因 Asp-96 的水解而再質子化,形成 N state。N state 再因 Asp-96 從胞內得到質子而再質子化,而形成 N´ state (Lanyi, 2004; Heberle, 2000)。
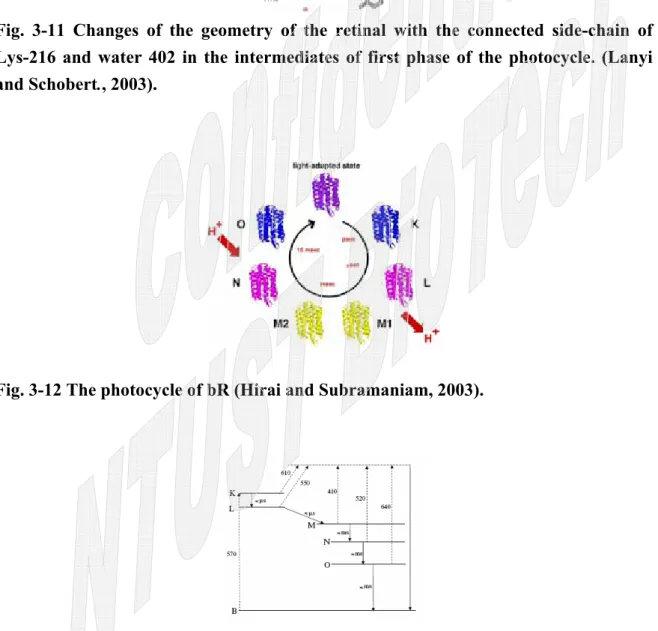
Fig. 3-11 Changes of the geometry of the retinal with the connected side-chain of Lys-216 and water 402 in the intermediates of first phase of the photocycle. (Lanyi and Schobert., 2003).
Fig. 3-12 The photocycle of bR (Hirai and Subramaniam, 2003).
Fig. 3-13 Simplified level diagram representing the photochemical cycle of bR molecule (Singh and Roy, 2003).
由於各個中間分子的結構不同,使得其顏色也會各有所變化,自然其吸收波長也 就大不相同(Fig. 3-11、Fig. 3-12、Fig. 3-13) (Hirai and Subramaniam, 2003)。
在光循環機制上還存在著其他分支的中間激發態存在,如不同波長的光源會激發 bR 形成不同的中間象態,例如若以紅光照射 bR,則bR會由 all-trans 的 O state 變 成另一個不在主光循環機制中的 9-cis P state (Fig. 3-14)。
Zhang 等人在 2004 年時,利用 150 W 的鹵素燈透過紅色濾片(λ = 680 nm)照射 bR 薄膜,利用量測照射紅光後的 bR吸收光譜變化,由原本的最大吸收波長 570 nm 變成 488 nm,來證明 P state 的形成。此外,他們更利用化學方法修飾的 bR 薄膜,
來增加且加速 P state的產生;其將 bR 與 C8H20ClN (tetraethylammonium chloride) 以 一定比例混合之後,再加入已溶在檸檬酸中的凝膠溶液,最後將混有 bR 的凝膠溶 液塗覆在親水性的石英基材上乾燥成膜。由 FTIR 分析可發現如此的化學修飾並沒 有破壞 bR 的基本二維結構。
Fig. 3-14 The photocycle of bR with a branching reaction (Zhang et al., 2004).
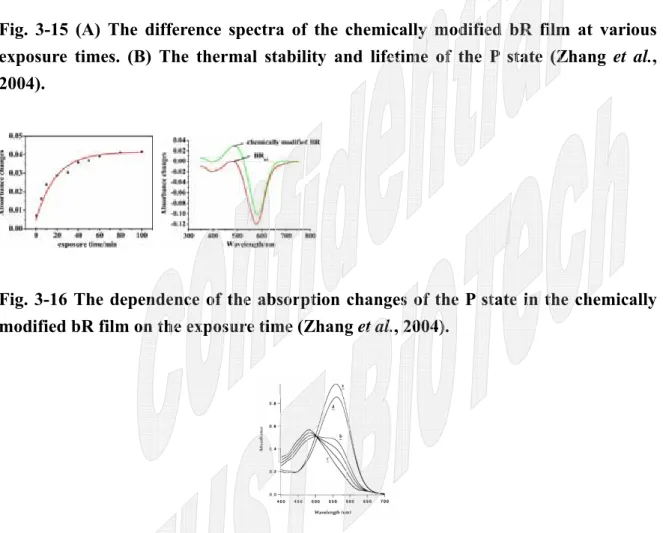
他們也證明了隨著照光時間的增長,P state 的量會持續增加,直到整個系統達到光化 學平衡。而且 P state 是非常穩定的物質,在照光過後的 30 天後仍然非常穩定地存 在著(Fig. 3-15、Fig. 3-16)。
Boyer 等 人 在 1995 年 時 則 是 利 用 添 加 Enflurane (trademark Ethrane; 2 chloro-1,1,2 trifluoroethyl difluoromethyl ether) 得到 bR 的 P state,而且由光譜可以證 明只要將此添加物移走,bR 仍然可以回到原始的基態(Fig. 3-17)。Enflurane 影響 bR 結構的原因,主要是影響 Schiff base 的去質子化,其量越多,bR 中 all-trans retinal 的比例會越高,即 bR 在 560 nm 的吸收峰會越來越小,而於 480 nm 的吸收峰則 會越來越大 (Nishimura et al., 1985)。
Fig. 3-15 (A) The difference spectra of the chemically modified bR film at various exposure times. (B) The thermal stability and lifetime of the P state (Zhang et al., 2004).
Fig. 3-16 The dependence of the absorption changes of the P state in the chemically modified bR film on the exposure time (Zhang et al., 2004).
Fig. 3-17 Reversible transformation of bR absorption spectrum in purple membrane films in the presence of Enflurane (Boyer et al., 1995).
3-2-3 氫離子之傳輸路徑
bR 受光激發後所產生質子的傳遞是由於 retinal 形成異構物所產生的。在光循 環過程中,bR 須經過一連串中間介質的改變,然後回到基態。
bR568 ⎯⎯→hv K590 ⇔ L550 ⇔ M1410 ⇔ M2410 ⇔ N560 ⇔ O610 ⎯⎯→ bR568
在 bR 被激發成 K state 時,其中至少儲存了 11.6 kcal/mol 的能量,以驅動接 下來光循環中一連串的等溫反應。這些反應都牽涉了氫離子的傳輸,而氫離子傳輸大 約分成五大步驟,其中兩個步驟是有關於將氫離子推出胞外,及從胞內吸取氫離子有 關,其餘的步驟則是氫離子在細胞膜內的傳遞 (Fig. 3-18) (Balashov, 2000)。
Fig. 3-18 Scheme showing steps in light-driven proton transport by bR from the inside the cell to the outside (Balashov, 2000).
當 bR 在基態時,Schiff base是質子化的 (C = NH+),受光激發後會使 Schiff base 去質子化,繼而 bR 會推出質子到胞外。Schiff base 的再質子化則是從胞內擷取質 子。而在 bR 內有兩個半通道 (half-channel),以連接胞內環境與Schiff base,Schiff base 與胞外環境。連接胞外的通道是較寬的;連接胞內的則較窄(Fig. 3-19)。
除了 Schiff base 之外,跟氫離子傳輸有關的殘基,就屬 Asp-85 最為重要了,
此胺基酸的位置大約在 Schiff base 往胞外方向大約 4.2 Å 的地方。在 Schiff base 去 質子化的時候,Asp-85則是扮演質子接收者的角色。再來則是 Asp-96,其位置在 Schiff base 朝胞內約 11 Å 的位置,大約是 Schiff base 與 胞內表面距離的一半,在 bR 為基態時其為質子化的狀態,等 bR 受光激發,則會去質子化而提供質子給 Schiff base。簡單來說 Schiff base 從 Asp-96 得到質子,而提供質子給 Asp-85,
Asp-96 從胞內得到質子,Asp-85 則推出質子到胞外。驅動質子傳遞的能量主要來自 於受光激發時,retinal 由 all-trans 轉為 13-cis 時所儲存的能量(Balashov, 2000)。
當 bR 在 5 < pH < 9 時, retinal 受光激發變為 K state,C13 = C14 幾乎全部轉 成 13-cis 的形式,接著轉變成 L state。在 L state 時,Schiff base 準備釋放出質子 給 Asp-85,由 L state 轉變成 M state 時,bR 內傳遞質子的通道會打開(Fig. 3-20),
Schiff base 會推出質子給 Asp-85。為了要使 pKa 值較高的 Schiff base 能提供質子 給 pKa 值較低的 Asp-85,必須要有額外的能量的存在,此能量則來自於 bR 的基 態受光激發,變為 K state 時,retinal 從all-trans 轉為 13-cis時所產生的 (Fig.
時負責穩定結構的水分子內之氫鍵打斷(Balashov, 2000; Hirai and Subramaniam, 2003)。
在幾乎同一個時間中,接近細胞膜表面的 PRC (proton release complex,包含 Arg-82,glutamic acid residues,Glu-204,Glu194及一些關鍵的水分子的部分)會推出 質子到胞外,而 PRC 的 pKa 大約從 9.5 降低為 5.8。
接著在 M state 變為 N state 時,Asp-96 會提供一個質子給 Schiff base。在基 態時,Asp-96 的 pKa 值是很高的,甚至大於 11。在 M state 變為 N state 時, Schiff base 與 Asp-96 會連接在一起,以降低 Asp-96 的氫離子親和性,而能傳遞質子給 Schiff base。
當 N state 變為 O state 時,Asp-96 會再質子化,而 bR 會從胞內擷取質子。
因 Asp-96 的再質子化,而與胞內接觸之表面積增加,使 pKa 降低到 7.2 – 7.5 之 間。bR 在擷取質子的同時,retinal 也從13-cis 的 N state 轉為 all-trans
Fig. 3-19 Structural model of bR based on a resolution of 2.3 Å (Heberle, 2000).
(a) (b)
Fig. 3-20 Schematic representation of the bR transport mechanism. The extracellular medium is in the downward direction. (a) The unilluminated state and (b) the M photointermediate (Lanyi and Luecke, 2001).
Fig. 3-21 The structures of all-trans 15-anti, and 13-cis 15-anti states of retinal (Hirai and Subramaniam, 2003).
的 O state。最後則是 Asp-85 提供質子給 PRC (proton release complex),bR 則從 O state 回 到 基 態 (Balashov, 2000; Lanyi, 2004; Lanyi and Luecke, 2001; Hirai and Subramaniam, 2003; Kalaidzidis et al., 2001; Bondar et al., 2004)。
所以 bR 受光激發後,氫離子的傳輸機制,可以簡單的列成下列五個步驟
[t-SBH+/H2O/Asp-85-] + hν → c-SB + Asp-85H + H2O (1)
XH → X- + H+ (release) (2)
Asp-96H + c-SB → Asp-96- + c-SBH+ (3)
Asp-96- + H+ (uptake) → Asp-96H (4a)
c-SBH+ → t-SBH+ (4b)
t-SBH+ + H2O + Asp-85- + X- → [t-SBH+/H2O/Asp-85-] + XH (5)
在上列的式子中 SB 代表 Schiff base,t 與 c 代表 retinal 為 all-trans 或 13-cis,X 代表 PRC (proton release complex),中括號則代表被水分子穩定的離子對 (Balashov, 2000),上列氫離子的傳遞步驟可參考 Fig. 3-22。
在 2 < pH < 5 時,則釋放質子到胞外及從胞內擷取質子的順序會顛倒,即 bR 會先從胞內吸收質子,再推出質子到胞外。而在不同的酸鹼值之下,bR 的光循環機
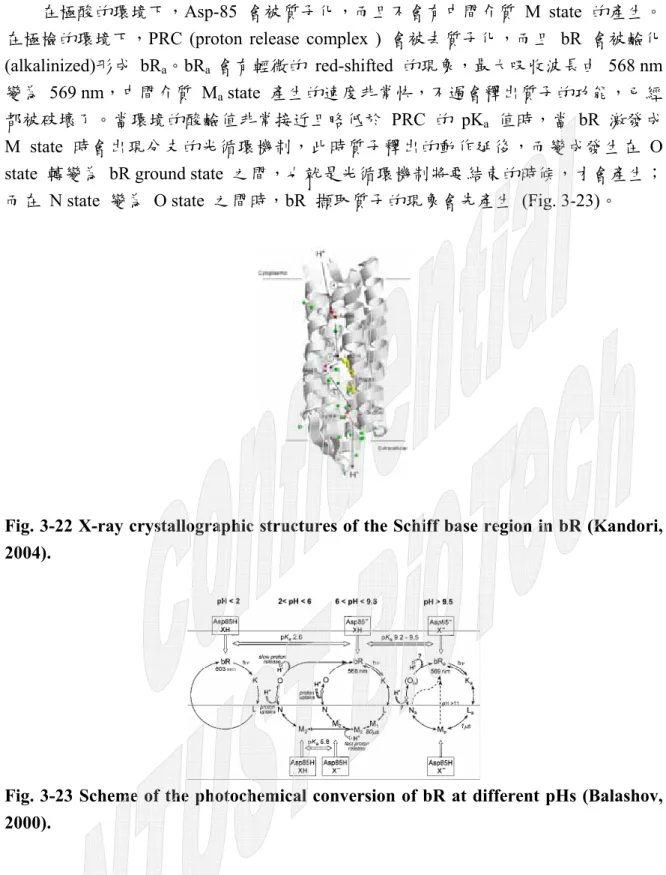
在極酸的環境下,Asp-85 會被質子化,而且不會有中間介質 M state 的產生。
在極檢的環境下,PRC (proton release complex ) 會被去質子化,而且 bR 會被鹼化 (alkalinized)形成 bRa。bRa 會有輕微的 red-shifted 的現象,最大吸收波長由 568 nm 變為 569 nm,中間介質 Ma state 產生的速度非常快,不過會釋出質子的功能,已經 都被破壞了。當環境的酸鹼值非常接近且略低於 PRC 的 pKa 值時,當 bR 激發成 M state 時會出現分支的光循環機制,此時質子釋出的動作延後,而變成發生在 O state 轉變為 bR ground state 之間,也就是光循環機制將要結束的時候,才會產生;
而在 N state 變為 O state 之間時,bR 擷取質子的現象會先產生 (Fig. 3-23)。
Fig. 3-22 X-ray crystallographic structures of the Schiff base region in bR (Kandori, 2004).
Fig. 3-23 Scheme of the photochemical conversion of bR at different pHs (Balashov, 2000).
在 bR 由 L state 變為 M state 時,質子是如何在 Schiff base 與 Asp-85 之間 傳遞的,到現在其真正確實的機制仍然是未知的。其中一個可能性就是存在於 Schiff base 與 Asp-85 之間的水分子,Wat-402。在 bR 為基態時,Wat-402 從 Schiff base 得到氫鍵,而提供氫鍵給 Asp-85 與 Asp-212。而在 L state 時,Wat-402 會因 Schiff base 與 Asp-85 而帶有相當強的極性,這會促進水分子的解離,而提供質子給 Asp-85,另一邊的氫氧鍵則給 Schiff base,以形成新的水分子,將此兩邊的極性恢復
成中性。另一種可能性則是 Schiff base 自己本身解離,提供質子給 Wat-402 或其他 有與 Schiff base 作用的水分子,而水分子則提供質子給 Asp-85。第一種情形就跟 hR (halorhodopsin) 的光循環機制中,帶負電的氯離子與 Schiff base 間的作用類似。這 些機制都是建立於假設在 bR 於 ground state 時,連結 Schiff base 與 Asp-85 的水 分子,Wat-402,在 L state 時還存在著(Balashov, 2000; Heberle, 2000; Kandori, 2004)。
在 bR 轉變為 K state 時,Wat-402 與 Schiff base 跟 Asp-85 間的氫鍵會被打 斷,這個結構幾何與氫鍵的改變,是可以破壞 Schiff base 與 Asp-85 間的離子平衡,
造成兩者間有質子傳輸的關鍵步驟。目前可由 FTIR 光譜觀察到,在光循環機制中,
bR 中會有水分子因為受光激發而改變鍵結結構。在 L state 時,retinal 改變結構形 成異構物,會破壞至少兩個水分子的結構,其中一個就在 Asp-85 的附近,這些水分 子在 L state 時穩定離子間的平衡,和 L 變成 M state 時,Schiff base 與 Asp-85 間 的質子傳輸都有關。在 Balashov (2000) 的文獻中有提到, Maeda 等人在 2000 年 時,利用 FTIR 的研究證明 bR 在 M 轉變成 M´ state 時,會有二到三個水分子在 retinal 改變結構時,有受到擾動的現象,其中一個位置靠近於 Phe-219。
在 M state 中包含了兩至三種不同的型態,在第一個象態中,只有一小部份的 Schiff base 被去質子化,緊接著是其他兩種不同的象態的出現(Betancourt and Glaeser, 2000) 。 這 些 改 變 使 得 原 本 在 L state 中 帶 有 極 性 的 離 子 對 (protonated Schiff base/Asp-85-) ,在 M state 中變成中性不帶電的狀態 (Schiff base/Asp-85H)。在這些 結構轉換的過程中,質子也同時被釋放到胞外了。有一些方式可以使 M state 存在的 時間變長,例如將 bR 做突變、化學修飾或是將其與特定的溶劑混在一起,都可以 達到某種程度的效果(Weetall and Druzhko, 2003; Dyukova et al., 1997),這對於要將 bR 應用在生化記憶體方面有著一定的幫助。而藍光去可以使 M state 直接變回 ground state 而縮短整個光循環的時間 (Fig. 3-24) (Lewis et al., 1997)。
在 M state 之後,進行一連串 retinal 結構的改變,最後恢復到 all-trans,讓 Asp-85 的質子回到 Schiff base 中,恢復 Schiff base/Asp-85 的離子對,而完成整個 光循環機制(Lanyi, 2004)。而由於 bR 結構上的改變,也可以觀察到 bR 受激發時 PM 表面結構的改變,如 Prsike 等人在 2000 年時,利用 AFM 觀察到照光的 PM,
其表面所排列的 bR 三聚體六角晶格,會比不照光者來的較小一些,他們稱此現象 為更加凝聚的 bR 三聚體結構。
3-2-4 bR 在感光材料上的應用
由於 bR 對光的敏感性,及其特殊的光循環機制,使得 bR 成為新興感光材料 上重要的一環。如攝影的感光材料 (Hampp et al.,1990; Chan et al., 2004; Juchem and Hampp, 2000; Yang and Wang, 1998)、光開關、光感測器、光學記憶體(Chen et al., 1995;
Marcy et al., 2000; Tokes et al., 2000; Wise et al., 2002; Birge et al., 1994; Stuart et al., 1996)及彩色影像偵測器(color image detection) 等等 (Choi et al., 2001; Frydrych et al., 2000)。
Fig. 3-25 A biomolecular photoreceptor (Choi et al., 2001).
Fig. 3-25 為 Choi 等人在 2001 年利用 bR 所做出來的彩色影像偵測器,他們 利用 bR 感光的特性,加上利用光源透過不同顏色的濾片,用電腦去擷取 bR 受光 時所產生的訊號,進一步的經過電腦的計算跟轉換後,將相對的受光顏色顯示在顯示 器上 (Fig. 3-26),而成功的作出一個以 bR 為基礎的影像偵測器。
Fig. 3-26 The overall process of color image interpretation and visualization. (Choi et al., 2001).
3-3 PM 的光電流訊號
3-3-1 PM 光電訊號流
為了測量 PM 因光激發所產生的光電流訊號,必須先設計出適合的系統。因為 所要量測的對象為 bR 所釋放出的質子,所以目前大多數都是以光電化學系統 (photo-electrochemical system)的形式,來量測其光電流訊號;此外,也有設計成光電 積分迴路 (opto-electronic integrated circuit)的形式,與半導體結合來量測 (Xu et al., 2004)。目前 bR 的光電系統可以做到微米尺寸的大小(Haronian and Lewis, 1992;
Fukuzawa et al., 1996)。
光電流訊號最典型的形式就是當 bR 受光時,因為放出質子,所以系統會有一 個正向電流的訊號;接著由於整個光電系統要維持電中性,所以訊號會下降;而不照 光後,bR 吸收質子,則系統會有一個負向的訊號;接著再因系統要維持電中性,而 再把訊號拉回平衡的位置,如 Fig. 3-27 的上圖所示,而下圖則為 bR 的全吸收光譜 圖,其最大吸收波長在 560 nm 左右。
Fig. 3-27 Characteristic pattern and action spectrum of the photoresponse of the native bacteriorhodopsin film (Boyer et al., 1995).
3-3-2 影響光電流訊號的因子 3-3-2-1 電極的選擇
在工作電極 (working electrode)的選擇上有許多種,目前大多數被選擇當作工作 電極的材質不外乎有 ITO (Indium Tin Oxide-銦錫氧化合物) 玻璃、金(Au)電極、白金 (Pt)電極及氧化錫(SnO2)等。但因各種導電物質的性質不同,所以量測出來的訊號也 會有所不同。如用Au 電極與 SnO2電極就會有所不同,Au 電極必須有額外施加電壓,
才能測得與完全沒有額外電壓的SnO2電極一樣的訊號(Saga et al., 1999)。而固定 bR
作工作電極的材料,都是不錯的選擇。
3-3-2-2 固定化方式
利用不同的方式將 bR 或 PM 固定在工作電極上也是影響光電流訊號的主因 之一。如用不同的高分子固定化,所得之光電訊號也會有所差異。Chu 等人在 2003 年時,是先用一層高分子( PAH 或 PDAC )鍍在已將表面負電性化的 ITO 玻璃上,
再將 PM 沉積在高分子上,而後可在 PM 上繼續鍍上高分子,重複這個步驟可以達 到多層的效果 (layer-by-layer)。由他們所用的兩種高分子物質的研究結果可以發現,
PAH (poly-allylamine hydrochloride)與 PDAC (poly-dially dimethyl ammonium chloride) 雖然是用相同的固定方法,不過所得到的光電訊號卻有所不同,利用 PAH 固定化所 得的訊號值,會比用 PDAC 者高出大約 30 % (Fig. 3-28,Fig. 3-29)。此外可用以包 埋 bR 的高分子還有 PVA (polyvnylalcohol) (Fig. 3-30),不過其固定化方式為先將高 分子材料直接與 bR 混合均勻,再將整個混合物固定在光電裝置上 (Lensu et al., 2004),但研究中只有探討受光時的正向光電流訊號
Fig. 3-28 A photoelectric response of the (PAH/PM)6 multilayer films (Chu et al., 2003).
Fig. 3-29 A photoelectric response of (PDAC/PM)6 multilayer films (Chu et al., 2003).
Fig. 3-30 bR-in-polyvinylalcohol element structure (Lensu et al., 2004).
Fig. 3-31 Schematic description of the purple membrane adsorbed Lumirror system (Muneyuki et al., 1998).
而 Muneyuki 等人在 1998 年將 PM 直接沉積在一個 Lumirror 高分子的薄膜 ( 900 nm 厚之 polyester 膜)上,並以雙電極的系統來測量光電流訊號。他們用此系 統來探討環境的酸鹼值對光電流訊號的影響 (Fig. 3-31)。另外,也有學者使用夾層的 方式,如之前提過的高分子夾層,或是直接用兩層導電物質將 bR 、 PM 或其與高 分子物質的混合物固定在中間 (Fig. 3-32) (Xu et al., 2003; Manoj and Narayan, 2004;
Hong, 1997; Yao et al., 2003)。
其實不論是何種夾層的方式,其最終目的都是為了要能測得 bR 的光電流訊 號,若不要刻意的將 bR 作不同的性質改變,這幾種固定化方式的差別,只在於所 得的訊號值大小及訊號峰值的寬度的不同,而這只與每個光電訊號裝置的靈敏度有 關,而不是 bR 本身所造成的差異性。
Min 等人在 1998 年比較兩種不同的 bR 固定化方法,一種是普遍被採用的 bR Langmuir-Blodgett (LB) film 另一種則是電泳沉降 (electrophoretic sedimentation,EPS) 法。他們比較兩種固定方式對光電訊號之影響,發現在相同的條件下,EPS 法所得 到的光電流訊號值會比 LB film 來的大,由此可知固定化的方式對於光電流訊號有 著一定程度的影響。
Choi 等人在 2002 年,以自排性單分子膜 (self-assembled monolayers,SAMs) 的方式來固定 bR,他們先將 11-mercaptoundecanoic acid (11-MUA)用 SAMs 固定在 金的基材上,再用 poly-L-lysine (PL) 來當作 11-MUA 與 bR 之間連結的架橋 (Fig.
3-33)。而在此研究中所用以測量光電流的系統也是設計成電化學的形式,工作電極 為有固定 bR 的金板,逆電極則選擇 ITO 玻璃,電解質是用 0.1 M KCl,他們則利
不論是直接固定 bR 或是固定 PM ,如何使其穩定地固定在基材上,對於 bR 未來的應用上,不論是光感測器或是生化電腦記憶體等,固定化都是一個非常重要的 研究課題。
Fig. 3-32 A motion-sensitive sensor array based on the photoelectric effect of bR.
(Hong, 1997).
Fig. 3-33 A schematic illustration of the bR thin films adsorbed onto the modified SAMs (Choi et al., 2002).
3-3-2-3 酸鹼值與溫度效應
環境的酸鹼值對 bR 的特性來說是非常重要的,因為不同的酸鹼值會對 bR 釋 放質子的動作有所影響,而且有可能會進一步改變 bR 的結構而改變光循環的機 制。如之前所提過的,在極酸的環境下,Asp-85 會被質子化,而不會有中間介質 M state 的產生。在極檢的環境下,PRC 會被去質子化,而且 bR 會被鹼化形成 bRa, 其中間介質 Ma state 產生的速度非常快,不過會釋出質子的功能,已經都被破壞了。
當環境的酸鹼值非常接近且略低於 PRC 的 pKa 值時,在 bR 激發成 M state 時會 出現分支的光循環機制,此時質子釋出的動作延後,而變成在 O state 轉變為 bR ground state 之間,也就是光循環機制將要結束的時候,才會產生;而在 N state 變 為 O state 之間時,bR 擷取質子的現象會先產生。Saga 等人在 1998 年也探討酸鹼 值對 bR 的影響 (Fig. 3-34,Fig. 3-35),其研究方法雖然與 Balashov 在 2000 年時 的研究有著些許的不同,不過都同樣證明了酸鹼值對於 bR 有著相當程度的影響。
(Balashov, 2000; Muneyuki et al., 1998; Choi et al., 2002; Wang et al., 1999; Saga et al.,
1998)。bR 在中性與酸性環境下推出與吸收質子的順序相反,這會對光電系統所得到 光電流訊號的正負值有影響,也就是若光電系統在 bR 為中性環境下得到一個正值 的開燈訊號,則酸性環境下所得到之開燈訊號就必定為負值,這對於沒有參考電極的 雙電極光電系統在光電流訊號機制的判別上非常重要。
Fig. 3-34 Simplified bR photocycles at neutral, low and high pH. (Saga et al., 1998).
Fig. 3-35 pH-Dependence of peak photocurrent of bR (Saga et al., 1998).
至於溫度則是會影響各個激發態中間介質存在的時間,如 M state 或 N state 等,繼而影響光電訊號值。其最明顯的的不同就在於所得光電訊號波峰的寬度,會隨 著溫度上升而更顯得細長尖銳,而訊號值的大小也會有所不同 (Wang et al., 1999)。
Fig. 3-36 The temperature dependence of differential photocurrent of bR-LB films:
(Wang et al., 1999).
3-3-2-4 光源強度之效應
因bR 對於光有很高的敏感度,所以照射的光源強弱,也會影響光電流訊號值的 大小,如 Fig. 3-37 所示,照射 bR 的光源越強,其光電流訊號值越大 (Choi et al., 2002; Miyasaka et al., 1992)。
Fig. 3-37 Photocurrent the prepared bR thin films according to the various incident light intensities (Choi et al., 2002).
3-3-2-5 照光頻率的影響
在光電流訊號的產生機制中,一開始的上升訊號為 bR 照光推出質子,而造成 工作電極與逆電極之間的有電位差所測得的電流值,緊接著下降回復到基線的訊號,
則為系統為維持電中性而造成離子移動的訊號;相同的,關燈時下降的訊號為不照光 時 bR 吸收質子所造成的訊號,上升回復到基線的也是系統為維持電中性而造成離 子移動的訊號。
bR 受光推出質子,而光電系統去反應出相對的電流訊號也是需要時間的。也就 是說 bR 照光的時間要夠長,光電系統才能完全的反應出相對的光電訊號,另一方 面,足夠的時間也才能夠激發所有的 bR 蛋白質。所以當照光時間太短時會造成光 電系統來不及完全的反應出 bR 推出質子所造成的電流,或是系統中的 bR 還沒完 全被激發,馬上就因關燈的動作 bR 開始吸收質子,系統開始反映出下降的訊號,
自然而然所得的光電流訊號值就會比照光時間足夠者小。所以當照光頻率越快時,也 就是 bR 照光的時間越短,所得到的光電流訊號會相對的越小 (Fig. 3-38)。
Fig. 3-38 The characteristic of differential photocurrent from 10-layer bR-LB films under different light stimulus frequencie (Wang et al., 1999).
3-3-2-6 添加劑
在 bR 的環境中添加額外的物質也會影響其光電流訊號,如添加金屬離子會影 響環境的pKa 值,所以也會影響 bR 質子唧筒作用的情形(Saga et al., 2002; Shibata et al., 2002; Saga et al., 2001; Drachev et al., 1976),而且 bR 固定在 PM 中時有跟某些 二價的金屬離子(如 Ca2+ 與 Mg2+)鍵結,如果將這些金屬離子移走,則原本吸收波長 為 568 nm 的紫色細胞膜(PM),會變成吸收波長為 605 nm、 且沒有質子唧筒功能的 藍色細胞膜 (blue membrane,BM) (Shibata et al., 2002; Tan et al., 1996),自然沒有光 電訊號的產生。
某 些 添 加 劑 則 是 會 增 加 光 電 流 訊 號 的 強 度 , 如 FCCP (carbonyl cyanide-p-trifluormethoxy phenyl-hydrazone),它是一個良好的質子載體,所以當 bR 推 出質子時,有 FCCP 存在的話,會加速從胞內推出質子到胞外的速度(Li et al., 1995)。
前面所述的一些因素都只是在 bR 研究上比較常見的,除此之外,影響其光電 訊號的因子還有很多。例如 bR 所處環境的溼度,也是原因之一,因為水分對於要 進行脫附氫離子的 bR 非常重要,所以環境的溼度也是影響因素之一(Crittenden et al., 2003)。
Min 等人在 2001 年進行了一個雖然基本但是非常有趣的實驗。他們所設計的 光電系統為一個九宮格形狀的 bR 膜,也就是 9 個 bR 膜在測光電流時是一體的,可 是照光時卻是能各自獨立分開利用遮蔽物來控制哪個 bR 膜照光與否 (Fig. 3-39)。他 們在量測各個不同位置的光電流訊號後,發現光電流訊號之間有加成性。例如我們將 九個地方的 bR 分別編號為 1 ~ 9。第一次只測 1 跟 3 一起照光的光電訊號,第二次 只做2 照光的訊號,第三次則作 1、2、3 一起照光的訊號,最後會發現第一次跟第二 次實驗所得光電流訊號值的和大約等於第三次實驗所得之光電流訊號值 (Min et al., 2001),由此更可證明只有受光的 bR 才會有推出質子的動作,而且利用此基礎,對 於控制 bR 的光電流訊號值大小也非常有幫助。
Fig. 3-39 Schematic illustration of bR-based photoreceptor for image extraction (Min et al., 2001).
3-4 PDMS (poly-dimethylsiloxane)
3-4-1 PDMS 材料簡介
PDMS (poly-dimethylsiloxane) 為一種具有高透光性(能穿透波長大約在 190-700 nm 間的光)的高分子材料 (Fig. 3-40),在微流晶片的領域中,尤其是製作上非常受到 重視。它具有價格便宜,方便且可快速製作 (Jo et al., 2000) 的優點;此外,其具有 著非常高的生化及化學穩定性,穩定溫度可以高達 250℃,所以非常適合做為各種微 小裝置的基材(Armani et al., 1999)。
Fig. 3-40 Structure of Polydimethylsiloxane ( http://lifesci3.arc.nasa.gov/SpaceSettlement/spaceres )
PDMS 在被製為成品之前是為液態的,在加入 curing 試劑之後才會完全變成固 態的彈性體,所以我們可以搭配任意的模組設計,來使所製得的 PDMS 形狀及尺寸 符合我們的需求。Fig. 3-41 為一個用 PDMS 製作微裝置典型的例子,簡單的來說,
就是先以曝光顯影及蝕刻的技術來製作我們的模組,而後再將 PDMS 成形前的溶液 倒入模組中,等成形後就是我們所要的 PDMS 裝置。
Fig. 3-41 Schematic representation of the steps involved in the fabrication process of the PDMS-glass hybrid microchip (Hong et al., 2000).
3-4-2 PDMS 之應用
由於 PDMS 具有容易製作成微小尺寸的優點,所以在應用上非常地廣,如微電 泳晶片(Hong et al., 1999)、微流晶片(Zeringue et al., 1999; Jo et al., 2000)、免疫感測晶 片(Choi et al., 1997)、PCR 晶片 (Fig. 3-42) (Hong et al., 2000; Stern et al., 2002)等等。
而不論何種裝置,為了操作的便利性及節省成本的考量,裝置的尺寸都是越小越好,
PDMS 由於其種種的優點,目前自然成為了在製作微小系統上不可或缺的一員。
Fig. 3-42 Layout of PDMS-glass hybrid microchip integrating PCR and capillary gel electrophoresis (Hong et al., 2000).
第四章 研究方法
4-1 實驗藥品
(1) Agar,瓊脂:Detroit,金暐代理
(2) Antifoam,polyproylene glycol,H[OCH(CH)3CH2]nOH,消泡劑:Nacalai Tesgue Inc., Japan,鼎好代理
(3) Bacteriological peptone,消化蛋白質:Oxoid Ltd., Basingstoke England,金暐代理 (4) Deoxyribonuclease-I,Dnase,DNA 分解酵素:Sigma,六和代理,酵素活性為
2000 unit/mg
(5) Dextrose,C6H12O6,葡萄糖:J. T. Baker (6) D. I. Water,H2O,去離子水:8 樓實驗室公用 (7) Ethanol,C2H5OH,乙醇:國宜代理
(8) Halobacterium halobium–S9:實驗生產培養而來。由德國 Max-Plank Institute 的 Dieter Oesterhelt 教授提供 H. halobium –S9 菌種
(9) Hydrochloric acid,HCl,鹽酸:J. T. Baker
(10) Idium Tin Oxide glass,ITO 導電玻璃:正隆公司提供,電阻值為 10 Ω,ITO 鍍膜厚度值為1500 Å
(11) Isopropanol,C3H7OH,異丙醇:J. T. Baker
(12) Magnesium sulfate,MgSO4‧7H2O,硫酸鎂:Sigma (13) Nitric acid,HNO3,硝酸:J. T. Baker
(14) Polydimethylsiloxane,PDMS,聚二甲基丙烯酸甲酯:DOW CORNING,喬越 公司
(15) Potassium chloride,KCl,氯化鉀:J. T. Baker (16) Potassium hydroxide,KOH,氫氧化鉀:ACROS
(17) Potassium phosphate, dibasic,K2HPO4,磷酸氫二鉀:J. T. Baker (18) Potassium phosphate, monobasic,KH2PO4,磷酸氫化鉀:J. T. Baker (19) Purple membrane,PM 細胞膜:實驗生產純化而來
(20) Sodium chloride,NaCl,氯化鈉:J. T. Baker
(21) Sodium hydroxide,NaOH,氫氧化鈉:Mallinckrodt
(22) Sodium phosphate, dibasic,Na2HPO4,磷酸氫二鈉:J. T. Baker
(23) Sodium phosphate, monobasic,NaH2PO4,磷酸氫化納:J. T. Baker (24) Sucrose,C12H22O11,果糖:Sigma
(25) Trisodium citrate,C6H5O7Na3‧2H2O,檸檬酸鹽:Sigma (26) 銅膠帶:皓睿科技有限公司
(27) 抗酸鹼膠帶:友俊國際股份有限公司 (28) 矽膠:DOW CORNING,喬越公司 (29) 模型改造版:TAMIYA,萬品館模型專賣 (30) 模型膠:TAMIYA,萬品館模型專賣 (31) 鋁膠帶:皓睿科技有限公司
4-2 實驗設備
Fig. 4-1 為光電系統設備的架設圖,主要元件有氙燈、濾片、光遮蔽物,光電化學系 統,微電流儀及電腦。我們所使用的微電流儀為雙電極的形式,其餘的設備,包括養 菌發酵槽的列表如下:
(1) 微生物菌體發酵槽:B. Braun Biotech Internatioonal,型號為 Biostat B,尚偉公司 代理
(2) 循環水控制水槽:EYELA COOL ACE,型號為 CA-1200,尚偉公司代理
(3) 灌注式管柱層析蛋白質純化系統 (Perfusion Chromatography Workstation) : ABI Applied Biosystems,型號為 Bio CAD 700E,展陽公司代理
(4) 高速離心機:Beckman,型號為 Avanti J-25,貝克庫爾特公司代理
(5) 超高速離心機:Beckman,型號為 OPTIMA XL-90K,貝克庫爾特公司代理 (6) 高速離心機所用轉子:Beckman,型號為 JA 25.5 (fixed-angle rotor , 8 × 50 ml),貝
克庫爾特公司代理
(7) 高速離心機所用轉子:Beckman,型號為 JA 10 (fixed-angle rotor , 6 × 500 ml),貝 克庫爾特公司代理
(8) 超高速離心機所用轉子:Beckman,型號為 SW 28 (6x swinging-buckets rotor),貝 克庫爾特公司代理
(9) 超高速離心機所用離心管:Beckman,型號為 Beckman Ultra-clear tube,產品貨號 為344058,管長 25 × 89 mm,容量為 38.5 ml,貝克庫爾特公司代理
品,型號為Jasco V-530,全拓公司代理
(11) pH 值測定儀:JENCO Electronics. LTD.,型號為 Microcomputer pH-Visio 6071,伯昂興業股份有限公司代理
(12) 透析袋:SPECTRA/Pro.,產品貨號為 132706,直徑為 29 mm,MWCO 值為 12K-14 K,康谷公司代理
(13) 透析夾:SPECTRUM MEDICAL Industries,產品貨號為 Closure 132736,夾 長為4 cm,康谷公司代理。
(14) 磁石加熱攪拌器:Corning。
(15) 超音波震盪器:NEY Industry,型號為 ULTRA sonic 19B
(16) 氙燈激光源組:Model A-1010B,Photon Technology International (17) 光源遮斷器:Model 300C,Scitec Instruments
(18) 濾光片(Blue、Green、Red、Yellow) :Model 079-0010、0020、0030、0040,OptoSigma (19) 逆電極:白金棒,國宜代理
(20) 微電流儀:Model 6485,Kiethley
(21) 微量電子天秤:Model AG104,Mettler Toledo (22) 恆溫振盪培養器:Model LM-570,COCONO
(23) 桌上型微量離心機(Microcentrifuge) :Model UFO2100,Pantech (24) 微量離心機:Model 5410,Eppendorf
4
Kiethley-6485 picoammeter
Counter electrode Working
electrode
Xenon light Filter Light gate Photo-electrochemical system Computer
Fig. 4-1 光電流訊號裝置圖
4-3 實驗流程
實驗流程圖如 Fig. 4-2 所示,其中包括 H. halobium-S9 菌體的培養、紫色 細胞膜的純化及光電化學系統的建立。
在菌體發酵培養時,利用 UV/Vis 光譜測量菌液之 OD560 與 OD660 的數 據,以了解菌體的生長及紫色細胞膜合成的情形。
本文研究的重點主要在於光電化學系統的建立,建立出能直接量測所得到 紫色細胞膜溶液的光電訊號。所以實驗中包括先利用初步的比色管系統 (cuvette system) 找出系統最適化條件,如最適合的中間介質等,再用同樣的操作條件去 建立尺寸較小的 PDMS 陣列系統,看是否也能測得紫色細胞膜溶液的光電訊 號。
本研究也利用所建立出的光電化學系統,初步的探討影響光電訊號的因 子,如燈源強弱、紫色細胞膜濃度、紫色細胞膜酸鹼值、電解質的酸鹼度、照光 頻率、光源顏色及比色管系統與 PDMS 陣列系統的比較。
H. halobium-S9 菌體搖瓶培養
H. halobium-S9 發酵培養
紫色細胞膜純化
比色管及 PDMS 陣列光電化 學系統之建立
影響光電流訊號之因子探討
中間介質 的影響
燈源瓦數 的影響
紫色細胞膜酸鹼 值及濃度的影響
電解質酸鹼 值之影響
比色管與 PDMS 陣列系統之比較 照光頻率
的影響
光源顏色 的影響
4-4 實驗步驟
4-4-1 H. halobium-S9 菌株培養 發酵培養
(1) 發酵系統外圍架設日光燈管,並依發酵槽標準操作程序進行前置作業。
(2) 由濃度 2 N、體積各為 500 mL 的 NaOH 及 HCl 所組成之酸鹼液,藉系統 pH 電極之檢測自動控制發酵槽槽體內之pH 值。
(3) 於配製好之 2 公升培養基中加入適量消泡劑,及 40 g 之 glucose 做為碳源來 源,每一公升約添加5 滴消泡劑,以避免發酵時有過多氣泡產生,造成氣泡 破裂時所產生的剪力傷害到菌體。
(4) 進行槽體與饋料瓶之滅菌操作,並調控好循環水系統確定系統中含有充分 的控溫循環水可用。
(5) 空氣壓縮機為桌上型小型壓縮機,直接接上管線至發酵槽。
(6) 槽體滅菌後接上相關管路及循環系統裝置與偵測器等外接元件,連結控制 器平面與發酵槽相關管路元件後,即進行氧氣電極極化程序。
(7) 至氧氣電極極化 6 小時後,確定槽體溫度下降後即進行發酵槽系統參數設 定。
(8) 設定溫度為 37 ℃,槽體葉片攪拌速率為 40 rpm,酸鹼 pH 質以 2N 的 NaOH 與HCl 之饋料控制,維持系統 pH 值在 7-7.2 範圍內,通氧量則為壓縮機的 最小值。
(9) 設定好系統控制參數後,即進行種菌動作。將 200 mL 成熟菌液經種菌口送 入槽體培養基中。
(10) 每日經取樣瓶進行取樣動作,藉 UV/Vis 光譜分析儀紀錄菌液 OD560、OD580、 OD660等數值。
4-4-2 PM 細胞膜純化
(1) 將發酵後之 2.2 公升成熟菌液,利用 10,000 rpm 轉速於 4 ℃環境下離心 30 分鐘,以進行收菌動作。
(2) 使用 BS 鹽水溶液清洗離心後之菌體顆粒,並以 10,000 rpm 轉速於 4℃環境 下離心30 分鐘進行清洗收菌,重複上述清洗與離心之程序 3 次,且需注意
所使用之離心管的數量應逐次減少以利菌體收集。
(3) 加入 40 mL 的 BS 鹽水溶液溶解清洗完畢之菌體顆粒,置入孔徑大小為 13 K 的透析膜中,並添加1500 Units 的 DNase 於上述溶液中,將透析膜使用透 析夾做封口動作後,即置入外在環境為 5 公升之去離子水溶液中,於室溫 下進行透析程序約16 個小時。
(4) 隔日即將透析袋中的紫色溶液,利用超高速離心機進行收集,設定轉速為 12,500 rpm 溫度控制為 4 ℃環境下,離心 30 分鐘後可得紫色外觀之顆粒。
(5) 使用 0.1 M NaCl 鹽水溶液進行清洗程序,設定離心機轉速為 12,500 rpm,
在4 ℃環境下離心 30 分鐘,收集外觀為紫色的沉澱物。
(6) 以去離子水清洗所得紫色沉澱物,並設定超高速離心機轉速為 12,500 rpm,
在4 ℃環境下離心 30 分鐘﹔重複此步驟至上清液為清澈不帶色為止,並需 注意要逐次減少離心管使用數量,以減少沉澱物附著於管壁造成收集時有 所損失。
(7) 將經上述步驟所得之紫色沉澱物溶於 18 mL 的去離子水溶液中。
(8) 製作果糖梯度,平均每管離心管中置入 2 mL 濃度為 60 %的果糖濃度於底 部,於其上製造出30 mL 容積之 30 %-50 %的果糖濃度梯度,再將(7)步驟 中的紫色溶液每管分配6 mL 至裝有果糖濃度梯度之離心管的最上層。
(9) 進行超高速離心操作,將上述裝有樣品與果糖梯度之離心管,先以天平準 確量測各管重量相同至小數第 3 位,並小心盡量減少搖晃離心管,將其置 入SW 28 轉子中設定轉數為 12,500 rpm,環境溫度為 4 ℃下離心 18 小時。
(10) 於超高速離心程序後,若無誤即可明顯觀測到離心管中 PM 與 RM 的分 層,此時利用微量吸管吸取出PM 分布帶。
(11) 使用去離子水進行 PM 與殘餘果糖的清洗程序,為避免 PM 樣品有過多果 糖的殘餘,仍需以去離子水加以清洗並以超高速離心機進行沉澱物的收 集,設定轉速為12,500 rpm,溫度為 4 ℃環境下離心 45 分鐘。
(12) 重複(11)步驟 3 次。
(13) 將收集所得知 PM 樣品溶於適量的去離子水中保存,並將其分裝成 1 mL 來保存。短期保存而言儲放溫度為0 ℃ - 4 ℃環境,若長期儲放則以-20 ℃ 環境為佳。
4-4-3 光電化學系統的建立
(1) 將兩個全量比色管的一個完全透明面切除。
(2) 在兩個比色管缺面的部分插入中間介質,用矽膠黏合比色管與中間介質,
如 Fig. 4-3 所示。
(1) 將組合好的裝置放入乾燥箱,等待一天完全黏合後,就可架設光電化學系統 來測量紫色細胞膜之光電訊號。
(a) (b)
cuvette intermediate
cuvette
intermediate
10 mm 20 mm
30 mm
Fig. 4-3 比色管系統 (a) 側面示意圖,(b) 正面示意圖
2. 清洗 ITO 玻璃
(1) 使用鑽石刀切割適當大小面積之 ITO 導電玻璃。
(2) 利用三用電錶確定 ITO 玻璃之導電面。
(3) 以去離子水清洗玻璃,在清水中以超音波震盪 15 分鐘。
(4) 置換掉去離子水溶液,再於 Isopropanol 震盪清洗 15 分鐘。
(5) 倒掉 Isopropanol 溶液,最後以去離子水溶液清洗 15 分鐘 2 次。
(6) 於室溫無塵環境中靜置待乾。
3. 蝕刻 ITO 玻璃
(1) 將已洗淨且切刻成適當大小的 ITO 玻璃,用抗酸鹼膠帶將欲留 ITO 的部 分貼住,而且要非常緊密的貼住,以防預保留 ITO 的部分也被蝕刻掉,如 Fig. 4-4。
(2) 將 ITO 蝕刻液加熱到 55 ℃,然後將已貼抗酸鹼膠帶的 ITO 玻璃放入溶