政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
台灣產莎草科植物光合作用型多樣性之研究
計畫類別:
■
個別型計畫
□ 整合型計畫
計畫編號:NSC96-2621-B-006-002-
執行期間:
96 年 08 月 01 日至 97 年 07 月 31 日
計畫主持人:郭長生
共同主持人:
計畫參與人員:吳宜蓁
成果報告類型(依經費核定清單規定繳交):
■
精簡報告
□完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年
■
二年後可公開查詢
執行單位:成功大學生命科學系
中
華
民
國
97 年
10
月 31 日
背景與目的
C4 光合作用於被子植物中多次演化。伴隨葉構造的改變、光合作用位置空間上的重新 排列,能讓CO2濃度集中於Rubisco周圍,因此降低經由光呼吸作用損失的能量。C4植物藉 此維持較高的淨光合作用速率而能特別分布於熱帶及亞熱帶地區。C4 植物多次起源於18 個科中,而單子葉植物中,唯一具C4 植物的禾本科及莎草科,一直是研究C4 植物的主要 對象。在台灣這兩科也是維管束植物中的大科。物種多、分布廣、棲地多樣,是探討C4 植 物生態與環境的好題目。國外已有研究,且各自有其地域特性,且不同科之間可能會有不 同的生態意函。台灣的禾本科目前已有較多的C4 植物資料、莎草科則尚待建立。希望藉由 此計畫,研究台灣的莎草科C4 植物多樣性,包括:確認物種光合作用類型、調查記錄其棲 地並配合氣象資料,以探討台灣產莎草科C4 植物的分布與生態之關係。材料與方法
材料 以莎草科莎草族及飄拂草屬材料為主,野外採集健康之正常葉作為解剖特徵鏡檢之用 外。另將採集品製作臘葉標本。無法採集到之材料,可藉由申請取得各標本館所保存之標 本小量樣品。 方法一、光學顯微鏡檢解剖特徵:材料固定於 FAA 樣品。在 PELCO BioWave® 微波爐內,用 經酒精系列脫水、以石蠟包埋後,用切片機製作 8-10 um 厚度之切片,經 xylol 脫蠟 和 Hematoxylin 染色,供顯微鏡檢各種和光合型有關之解剖特徵。
二、穩定碳同位素測定(Leaf carbon isotope discrimination)
三、物種棲地分析:參照 Ueno and Takeda(1992)的棲地分類與 Flora of Taiwan v.2 對物種棲 地描述分為(1)旱地;(2)中濕;(3)濕地/水生;(4)鹽鹼地;(5)遮蔭地。
結果
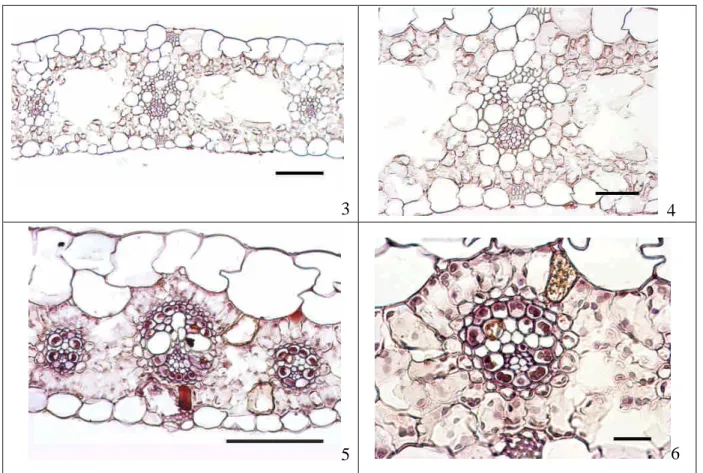
利用葉部解剖光學顯微鏡檢,若為 C4 物種,維管束排列緊密,脈間距離短,通常不超 過 3 個葉肉細胞數。葉肉細胞緊密環繞維管束(圖 1, 2 5, 6)。束鞘在莎草族分為兩層,外層 為纖維構成,內層為薄壁細胞組成,內有大體積的葉綠體,常有後生物質澱粉粒的產生(圖 2)。飄拂草屬的葉部解剖則有三層束鞘,在纖維的外層尚有一層薄壁細胞,其葉綠體與葉 肉細胞形態相同(圖 5, 6)。觀察 C3 物種的葉肉組織,有海綿和柵狀兩種形態,包圍著氣室, 脈間距離較 C4 物種長,沒有緊密圍繞維管束的現象(圖 3, 4)。束鞘亦分為內外兩層,外層 為膨大的薄壁細胞,細胞質少,內層為纖維細胞,並無葉綠體的發現(圖 4)。O’Leary(1988)年指出 C3 植物固定 CO2 的關鍵酵素 Rubisco 分餾值為 30%0,C4 植物則
大約為-28%0,而 C4 植物大約-14%0。本研究共檢驗 48 個物種之穩定碳同位素比值,結果
列於表一,圖 7 可清楚顯示 C3 與 C4 物種間比值有一明顯間隔(gap)。
在台灣莎草族的材料分為五屬:莎草屬(24 種,2 亞種)、水蜈蚣屬(2 種,1 亞種)、磚 子苗屬(5 種)、扁莎屬(6 種),斷節莎屬(1 種),總共 38 種,3 亞種。除了莎草屬的 Cyperus
alternifoliu subsp. flabelliformis、C. difformis、C. diffusus、C. haspan、C. platystylis、C. tenuispica
六種材料為 C3 類型外,其餘皆為 C4 植物。C. imbricatus subsp. elongatus 與 Pycreus sulcinux 未找到材料檢視。飄拂草屬的材料共 19 種,9 變種,皆為 C4 植物。Fimbristylis acuminata、
F. littoralis var. koidzumiana、F. microcarya var. microcarya、F. subinclinata、F. umbellaris 未找到材料檢視。表一為本研究的整理,列表碳同位素比值與葉部解剖結果,並與前人研 究成果比較。
討論
由本研究結果可發現,台灣的莎草科莎草族與飄拂草屬以 C4 物種為大多數,由葉部解 剖、穩定碳同位素的結果與前人研究可相互支持。僅有莎草屬六個物種為 C3 植物,推測原 因應為台灣位居副熱帶與熱帶氣候交界,故多為熱帶與副熱帶親合的 C4 物種。調查這些物 種的棲地發現,大多數物種生長於濕地水生環境,占總物種 65.52%,若加上中溼物種,則 比例高達 82.76%,與一般認為 C4 物種大都分布在高溫乾燥的環境有極大差異(表二)。在乾 燥的旱地,本研究材料僅有 3 種,占總物種的 5.17% (表二)。與其它 C4 植物相同的是,除 了 Fimbristylis shimadana 此一稀有種外,皆生長於開闊的環境,擁有充足的光能供應,符 合 C4 植物適應於高光照的環境(Ueno and Takeda, 1992)。此外,亦有 6 種 C4 物種生長於鹽 鹼地,土壤鹽分的高低,與氣孔調控相關,進而影響水分使用效率,C4 植物可能較有優勢 (Collins and Jones, 1985)。但在此次研究的各屬中,僅有莎草屬 6 個物種為 C3 光合作用類型,缺乏其他分類群物 種的光合作用類型與棲地環境資料的對照。C4 植物由 C3 植物演化出來,為了防止光呼吸 造成的能量損失,因此水分使用效率較一般 C3 植物高,但莎草科 C4 物種的競爭優勢卻與 水分得利用似無關聯,根據 Jones(1987)指出,非洲沼澤區的 C4 優勢物種紙莎草的適應與 氮使用效率高有關,推測在濕地水生環境中,莎草科 C4 物種的競爭優勢應與此有關聯,但 自 Jones 後關於莎草科生理方面的研究報告很少,目前尚不清楚確切原因。 因此,除了繼續台灣其他分類群的光合作用類型研究與棲地探討外,生理與基因調控的研 究也是必需進行的重點,尤其近幾年發現的荸薺屬(Eleocharis spp.)物種具有高度C3/C4形態 與功能上可塑性(Murphy et al., 2007)
自評
本研究已經完成了大部份的實驗,証實了穩定碳同位素在 C3 與 C4 的大量篩選是很有 效率和準確性的,而且和切片的結果也是吻合的。除了少數物種在台灣可能稀少甚至滅絕, 因此無法採集到新鮮的材料而至各大標本館借其物種標本,亦發現標本也可以進行 C3 與C4 的探討。因時間與物種分布不確定的關係,無法完成與氣象因子的比對,之後如果資料 完整而進行比對後可以更了解莎草科的 C3 與 C4 植物在生態上的意義。因此本研究基本上 已達到之前所提出的進度與目的。
參考文獻
Carolin RC, Jacobs SWL, Vesk M. 1977. The ultrastructure of Kranz cells in the family Cyperaceae. Bot Gaz 138:413-419
Collins, R.P., Jones, M.B. 1985. The influence of climatic factors on the distribution of C4 species in Europe. Vegetatio 64: 121-129
Connie Lynn Soros. 1999. Comparative structure, development and evolution of photosynthetic tissue in C3 and C4 Cyperaceae. A PhD thesis of Philosophy Graduate Department of Botany University of Toronto
Editorial Committee of the Flora of Taiwan. 2000. Flora of Taiwan second edition V.5. Department of National Taiwan University
Jones MB. 1987. The photosynthetic characteristics of papyrus in a tropical swamp. Oecologia 71:355-359
Lesley R. Murphy et al. 2007. Diversity and plasticity of C4 photosynthesis in Eleocharis (Cyperaceae). Functional Plant Biology 34: 571-580
Marion H. O'Leary. 1988. Carbon Isotopes in Photosynthesis. BioScience 38: 328-336 O. Ueno and T. Takeda. 1992. Photosynthetic pathways, ecological characteristics, and the
geographical distribution of the Cyperaceae in Japan. Oecologia 89:195-20
Stock, WD, DK Chuba and GA Verboom. 2004. Distribution of South African C3and C4 species of Cyperaceae in relation to climate and phylogeny Austral Ecology 29: 313–319
郭長生, 江蔡淑華. 1984. 飄拂草屬植物葉之比較解剖. Yushania 1(3):29-43
C3/C4 葉部解剖舉例
3 4
5 6
圖 1, 2 Cyperus iria;圖 3, 4 .C. tenuispica;圖 5, 6 Fimbristylis dichotoma。 圖 1, 3, 4, 5 scale bar: 50um. 圖 2, 6 scale bar: 10um
圖 7 48 種材料穩定碳同位素散佈圖 -35 -30 -25 -20 -15 -10 -5 0 0 5 10 15 20 25 30 35 40 45 50 species numbers is o to p e ra ti o (% o ) 表一 研究材料之棲地、碳同位素與葉部解剖總整理 學名 棲地 * 穩定碳同 位素比值 解剖 類型** 本研究 結果 前人研究 結果** 莎草族
Cyperus alternifolius L. subsp.
flabelliformis (Rottb.) Kük.
W -28.295 K-C C3 C3(St)
Cyperus amuricus Maxim. M -11.74 C4 C4(U)
Cyperus compressus L. W K-C C4 C4(U)
Cyperus cuspidatus Humb. W -11.284 C4 C4(S)
Cyperus difformis L. W N C3 C3(U)
Cyperus diffusus Vahl W -32.00 N C3
Cyperus digitatus Roxb. W -10.69 K-C C4 C4(U)
Cyperus distans L. f. W -12.73 K-C C4 C4(U)
Cyperus exaltatus Retz. W -14.447 K-C C4 C4(U)
-13.11
Cyperus haspan L. W N C3 C3(U)
Cyperus imbricatus Retz. W -10.36 K-C C4 C4(U)
-10.96
Cyperus imbricatus Retz. subsp.
elongatus (Boeck.) T. Koyama
W
Cyperus iria L. W K-C C4 C4(C)
Cyperus malaccensis Lam. W -11.28 C4
Cyperus malaccensis Lam. subsp.
monophyllus (Vahl) T. Koyama
W -12.68 K-C C4
Cyperus nutans Vahl subsp.
subprolixus (Kük.) T. Koyama
W -12.52 K-C C4 C4(U)
Cyperus pilosus Vahl W K-C C4 C4(U)
Cyperus platystylis R. Br. W -28.13 N C3 C3(S)
Cyperus procerus Rottb. W -11.989 K-C C4
Cyperus pygmaeus Rottb. W -15.088 K-C C4 C4(B)
Cyperus rotundus L M K-C C4 C4(U)
Cyperus serotinus Rottb. W -13.580 K-C C4 C4(U)
Cyperus stoloniferus Retz. S -9.78 K-C C4 C4(U)
Cyperus tenuiculmis Bock. W -12.209 K-C C4
Cyperus tenuispica Steud. W N C3 C3(U)
Cyperus tuberosus Rottb. D -13.523 K-C C4 C4(S)
Kyllinga brevifolia Rottb. W K-C C4 C4(U)
Kyllinga nemoralis (J. R. & G.
Forst.) Dandy ex Hutch. & Dalzell
W -11.64 K-C C4 C4(U) -8.39
Kyllinga sesquiflora Torr. subsp.
cylindrica (Nees) T. Koyama
M -12.18 K-C C4 C4(U)
Mariscus compactus (Retz.)
Druce
W -12.095 K-C C4 C4(U)
Mariscus cyperinus Vahl M -11.731 K-C C4 C4(U)
-11.63
Mariscus javanicus (Houtt.) Merr.
& Metcalfe
S -12.185 K-C C4 C4(U)
Mariscus radians (Nees &
Meyen) T. Tang & F. T. Wang
S -10.28 K-C C4 -10.07
Mariscus sumatrensis (Retz.) J.
Raynal
M K-C C4 C4(U)
Pycreus flavidus (Retz.) T.
Koyama
W -15.616 K-C C4 C4(U)
Pycreus polystachyos (Rottb.) P.
Beauv.
W K-C C4 C4(U)
Pycreus pumilus (L.) Domin W -8.35 K-C C4
Pycreus sanguinolentus (Vahl)
Nees
W -10.30 K-C C4 C4(U)
Pycreus sulcinux (C. B. Clarke) C.
B. Clarke
D
Pycreus unioloides (R. Br.) Urb. W -12.288 C4 C4(U)
Torulinium odoratum (L.) S.
Hooper
W -14.511 K-C C4 C4(U)
飄拂草屬
Fimbristylis acuminata Vahl W
Fimbristylis aestivalis (Retz.)
Vahl var. aestivalis
W -13.916 K-F C4 C4(S)
Fimbristylis aestivalis (Retz.)
Vahl var. esquarrosa (Makino) T. Koyama
W -13.037 K-F C4 C4(U)
Fimbristylis bisumbellata (Forsk.)
Bubani
W -10.55 K-F C4 C4(U) -8.18
Fimbristylis complanata (Retz.)
Link
W K-F C4 C4(U)
Fimbristylis cymosa R. Br. S -10.39 K-F C4 C4(U)
Fimbristylis eragrostis (Nees &
Meyen ex Nees) Hance
M -10.888 K-F C4
Fimbristylis ferruginea (L.) Vahl
var. ferruginea
S -11.05 K-F C4 C4(S)
Fimbristylis ferruginea (L.) Vahl
var. anpinensis (Hayata) H.Y. Liu
S -13.487 C4 C4(S)
Fimbristylis littoralis Gaudich.
Var. littoralis
W K-F C4 C4(U)
Fimbristylis littoralis Gaudich.
var. koidzumiana (Ohwi) T. Koyama
W
Fimbristylis microcarya F. Muell.
Var. microcarya
W -11.28 C4(S)
Fimbristylis microcarya F. Muell.
var. tainanensis (Ohwi) H. Y. Liu
W -12.12 K-F C4 C4(S) -11.04
Fimbristylis miliacea (L.) Vahl W -11.814 K-F C4
Fimbristylis nutans (Retz.) Vahl W -10.66 K-F C4 C4(U)
-11.72
Fimbristylis ovata (Burm. f.) J.
Kern
M -14.439 K-F C4 C4(C)
Fimbristylis polytrichoides (Retz.)
Vahl
W -15.357 K-F C4 C4(S)
Fimbristylis schoenoides (Retz.)
Vahl
W -12.039 K-F C4 C4(S)
Fimbristylis sericea (Poir.) R. Br. S -12.161 C4 C4(U)
Fimbristylis shimadana Ohwi Sha -16.753 K-F C4 C4(K)
Fimbristylis squarrosa Vahl W -10.89 K-F C4
-8.68
Fimbristylis subinclinata T.
Koyama
D
Fimbristylis tetragona R. Br. W -15.386 C4 C4(C)
Fimbristylis thomsonii Bock. M -13.738 K-F C4
Fimbristylis tomentosa Vahl M K-F C4
Fimbristylis tristachya R.Br. var.
subbispicata (Nees & Meyen) T. Koyama
W -11.938 K-F C4 C4(U)
Fimbristylis umbellaris (Lam.)
Vahl
W C4(K)
***U:Ueno and Taketa (1992);S:Connie Lynn Soros(1999);C:Carolin et al., 1977;St:Stock et al.(2004); K: Kaoh and Tsai (1984)
**K-C:Chlorocyperoid type;K-F:Fimbristyloid type;N:non-Kranz anatomy.
表二 本研究之屬分類階棲地整理 分類階 C3/C4 旱地 中溼 濕地/ 水生 鹽鹼 地 遮蔭 地 Fimbristylis C4 4.1(1) 16.7(4) 62.5(15) 12.5(3) 4.1(1) Cypereae Cyperus C3 0 0 100(6) 0 0 Cyperus C4 5.2(1) (2) 89.5(14) (1) 0 Kyllinga C4 0 66.7(2) 33.3(1) 0 0 Mariscus C4 0 40(2) 20(1) 40(2) 0 Pycreus C4 14.3(1) 0 85.7(6) 0 0 Torulinium C4 0 0 100(1) 0 0 subtotal 5.17(3) 17.24(10) 65.52(38) 10.34(6) 1.72(1)