國立成功大學
優秀新進教師暨研究人員學術研究計畫補助案
計畫結案報告
利用基因轉殖小鼠模式探討小分子干擾核醣核酸技術於 亨丁頓氏手舞足蹈症基因治療的可行性
計畫類別:■個別型計畫 計畫編號:NSC 98-2320-B-006 -041
執行期間: 98 年 11 月 1 日至 99 年 7 月 31 日 執行機構及系所:國立成功大學生理學研究所
計畫主持人:楊尚訓 計畫參與人員:楊尚訓
成果報告類型(依經費核定清單規定繳交):■精簡報告
本計畫除繳交成果報告外,另須繳交以下出國心得報告:
□赴國外出差或研習心得報告
□赴大陸地區出差或研習心得報告
□出席國際學術會議心得報告
□國際合作研究計畫國外研究報告
處理方式:除列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年■二年後可公開查詢 中 華 民 國 九 十 九 年 九 月 十 五 日
(一) 中文摘要
亨丁頓氏手舞足蹈症(Huntington’s disease)是一種罕見人類顯性遺傳疾病,主要影響 中樞神經系統,並造成腦部神經的退化,進而破壞病人的認知及運動功能。目前為止,
對於這疾病並沒有有效的治療方式,而這病人也因此承受相當大的疾病痛苦。近年來,
基因治療的開發為這疾病揭開了治療的楔子,尤其是小分子干擾核醣核酸(short
interfering RNA)的發現,更為治療遺傳性疾病開啟曙光;而目前為止尚無利用基因轉殖小 鼠的模式去探討小分子干擾核醣核酸對於亨丁頓氏手舞足蹈症的影響,因此本計畫預期
利用基因轉殖小鼠的模式進行探討基因治療的可行性;此一新人計畫為期9 個月,目前
已完成的成果包括: (1)篩選出適當的小分子干擾核醣核酸序列,確定能有效抑制突變 HTT 基因的表現;(2)建構含有適當小分子干擾核醣核酸序列的病毒載體,並產製高活性 的病毒;(3) 建立此病毒的基因轉殖模式,確定能有效的轉染亨丁頓氏手舞足蹈症之細 胞,並達到抑制突變 HTT 基因的病理上表現。本階段計畫的執行藉由體外細胞的模式證 明小分子干擾核醣核酸能夠抑制突變 HTT 基因,預期此結果將有助於提供寶貴的小分子 干擾核醣核酸治療實驗動物模式之建立。
關鍵字:亨丁頓氏手舞足蹈症;小分子干擾核醣核酸;基因轉殖小鼠;基因治療;動物 模式。
(二) 英文摘要
Huntington’s disease (HD) is an inherited autosomal-dominant disease which causes neurodegeneration in humans, and affects the central nervous system, resulting in distinct clinical symptoms related to neuron and brain dysfunction. To date, there is no cure of HD, suggesting the importance of developing novel therapy for HD. Several attempts on treating HD have been investigated in order to delay or alleviate the severity of HD, especially gene therapy. After the discovery of RNA interference (RNAi), it has emerged as a hallmarked technique to cure this genetic disease. However, there is no study using the RNAi transgenic mice model to investigate the potentially therapeutic method of RNAi for HD till now.
Therefore, the aim of this study is to use transgenic mice model to demonstrate the effect of RNAi on HD. This study had been conducted for 9 months, and reached the goals, including those (1) identifying the three RNAi which can suppress the expression of mutant HTT, (2) constructing relative RNAi sequences into lentiviral vector, and generating high titer lentiviruses and (3) establishing lentiviral transgenesis in the cell model in vitro, showing the suppression ability of RNAi on mutant HTT. With the above results we finished, we anticipate to facilitate the generation of transgenic mice model for understanding the RNAi therapy approach for HD, and hope to open a way to study the RNAi for the future application in biomedicine field.
Key words: Huntington’s disease (HD); RNA interference (RNAi); transgenic mice; gene therapy; animal model.
(三) 前言、研究目的、文獻探討、研究方法、結果與討論
HD is a terrible neurodegenerative disease which has no cure till now; therefore, developing therapeutic methods is an important investigation for HD. Taking advantage of transgenesis techniques, it would provide a clue for applying gene therapy targeting on these genetic diseases. Not only to alleviate symptoms, but also to prevent the onset of diseases.
Consequently, establishing this first mice model will confer the information in the aspects of molecular, pathological and behavioral alterations of HD, and provide the support for further development of short hairpin RNA (shRNA) therapy on genetic diseases.
Introduction:
Huntington’s disease (HD) is an inherited autosomal-dominant disease which causes neurodegeneration in humans. HD mainly affects the central nervous system, resulting in distinct clinical symptoms related to neuron and brain dysfunction [1]. HD is the phenotype from specific mutant Huntingtin protein (HTT), which is transcribed from mutant Huntingtin (HTT) gene, carrying an abnormal expansion of polyglutamine (polyQ; >37 residues) in the N-terminus [1, 2]. To date, no medicine can be efficient to cure HD, suggesting the importance of novel therapy for HD, especially gene therapy.
Several therapies have been investigated in order to delay or alleviate the severity of HD [3-9]. However, these approaches all target on the co-factors or indirect factors of HD. Since HD is well-characterized to be caused by the mutation of HTT, it would be logical to speculate whether reducing mutant HTT expression directly could release the phenotypes of HD.
Recently, RNA interference (RNAi) has emerged as a hallmarked breakthrough for knocking down target genes in mammalian cells in vitro and in vivo, especially short interfering RNAs (siRNAs) and shRNA. Several reports have been suggested that siRNAs have functioned both in vitro and in vivo to decrease the expression of HTT in post-transcription level [10-14], suggesting RNAi as a highly potential therapeutic strategy for HD. Most of in vivo studies have used stereotaxic injection to investigate the effects of siRNA/shRNA targeting on mutant HTT, which has been very informative; however, it has its limitations, such as heterogenous transduction of cell populations. Therefore, germline transgenesis, in comparison, provides uniform integration of foreign DNA across cells and across generations. This reproducibility facilitates investigation of the physiological mechanisms underlying phenotypic changes due to transgene presence. To date, there is no study using the shRNA transgenic mice model to investigate the potentially therapeutic method of shRNA for HD. Therefore, in this study we expect to use this alternative method, germline transgenic mice model which carries mutant HTT and siRNA, to determine the effect of shRNA function in vivo. In this 9-month study, we have identified three RNAi which can suppress the expression of mutant HTT, constructed relative RNAi sequences into lentiviral vector, generated high titer lentiviruses and established lentiviral transgenesis in the cell model in vitro, showing the suppression ability of three RNAi on mutant HTT.
Materials and methods:
shRNA Lentiviral vector construction
Three short-hairpin RNA (shRNA) sequences targeting to different regions of human HTT were screened for this study (Figure 1A), and then inserted into a lentiviral vector [15], which was regulated by human H1 promoter (pLV-H1-shHD-1, 2 and 3; Figure 1B).
To trace the expression of shRNAs, red fluorescence protein gene (RFP) controlled by human ubiquitin promoter was inserted into same lentiviral vector in the 3’ downstream of shRNA. (pLV-H1-shHD-1-U-RFP, pLV-H1-shHD-2-U-RFP and pLV-H1-shHD-3-U-RFP)..
Generation of high titer lentiviruses and infection of HD-293 FT cells
Lentiviruses were generated by co-transfection of lentiviral vectors of shRNAs, pΔ8.9, and vesicular stomatitis virus glycoprotein vector into 293 FT human embryonic kidney packaging cells (Invitrogen). To generate high titer lentiviruses, the lentiviruses were then concentrated by ultracentrifugation using the method described previously [16, 17]. In order to test the biofunction of shRNAs in vitro, HD-293 FT cells which carry mutant HTT fused with green fluorescence protein gene (GFP) under the control of tetracycline inducible promoter (GFP-HTT130Q) were used for this study.
RNA extraction, reverse transcription (RT), RT-PCR and real-time quantitative PCR (Q-PCR) analysis
RNAs were extracted from different cells using RNeasy Mini Kit (Qiagen), and subjected for RT (Applied Biosystems) to generate cDNAs. cDNAs were then used for Q-PCR via SYBR® Green system (Applied Biosystems), and expression levels of specific genes, including mutant HTT, endogenous HTT and 18S, were quantified using the iQ5 real-time PCR detection system (Bio-Rad).
Western blotting analysis
Total protein was extracted from different cells, and detected by different primary antibodies, mouse monoclonal mEM48 (1:50 dilution) and γ-tubulin (Sigma; 1:2,000 dilution).
Results:
Three sequences targeting on different regions of HTT were used for this study after performing computational analysis of human HTT exon 1 (Figure 1A). shHD-1 and shHD-2 target on the 5’ upstream of HTT exon1 before the polyQ repeats. shHD-3 targetes on the sequence between exon 1 and exon 2 of HTT gene. To construct these sequences into lentiviral vector, sense, stem loop, antisense and terminal sequences were inserted into vector which will synthesize short hairpin RNA interference controlled by human H1 promoter (Figure 1B). In order to deliver transgene into cells or mice, lentiviral transgenesis was used due to the characteristics of high efficiency. Therefore, we contransfected three constructs into 293 FT cells, and generated high titer lentiviruses which reached the titer of 108-109 CFU/mL. To test the infection ability of these lentiviruses and biofunction of shRNAs in vitro, HD-293 FT cells carrying mutant HTT fused with GFP under the control of tetracycline inducible promoter (GFP-HTT130Q) were infected by these lentiviruses in a small scale. Confirming the knock-down functions, Q-PCR analysis was performed to determine the expression level of
HTT. Here, shHD-1 suppressed approximately 75% of mRNA expression, and the other two constructs decreased approximately 50 % mRNA expression of mutant and endogenous HTT, showing the down regulation of both endogenous and mutant HTT in three constructs (Figure 2A), In the protein level, Western blotting result revealed the lower expression level in three shRNA groups compared to the induced HD-293 FT cells, especially shHD-1 (Figure 2B).
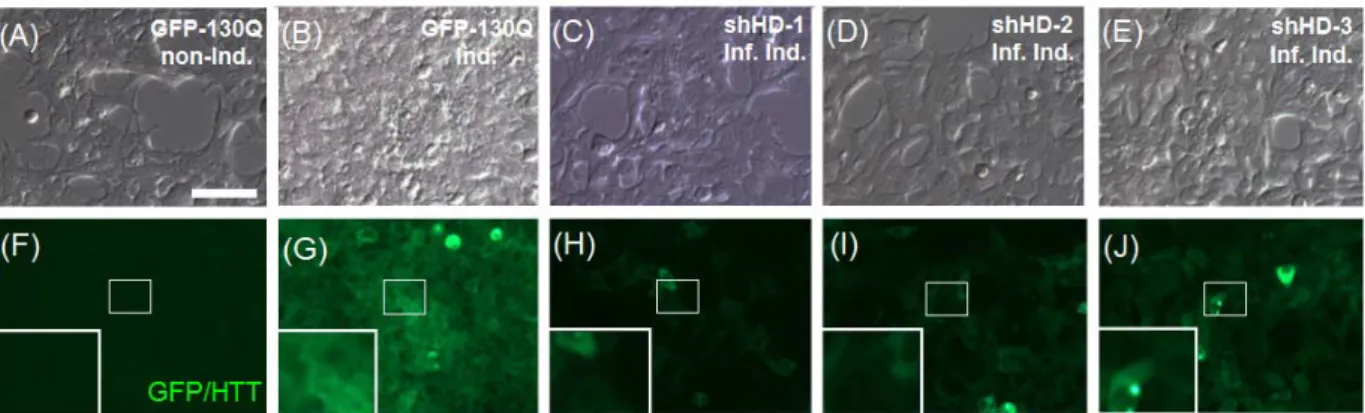
Since the HD-293 FT cells carried the HTT-GFP fusion protein, the GFP expression profiles also suggested the expression level of mutant HTT. With the observation of fluorescent signal under the same exposure time, three shRNA infected groups appeared lower overall intensity of GFP when compared to the induced HD-293 FT cells (Figure 3). Additionally, when we used RFP to trace the expression of shRNAs, we also observed the suppression ability of our constructs, where shRNAs carrying RFP signal revealed lower GFP signal which represented as the expression level of mutant HTT (Figure 4). Based on these results, it suggested the functional ability of infection for three lentiviral vectors with shRNAs in vitro, and these shRNAs could suppress the expression of mutant HTT.
Figure 1. The lentiviral constructs used for this study. Ttargeting sites of three shRNAs were illustrated in (A).
shHD-1 and shHD-2 target on the 5’
upstream of HTT Exon1 before the polyQ repeats. shHD-3 targets on the sequence between Exon1 and Exon2 of HTT gene. Lentiviral vector of shRNA was diagramed in (B). These shRNA constructs, which contain sense, stem loop, antisense and terminal sequences, were inserted into a lentiviral vector.
Figure 2. The suppression of mutant HTT by three shRNA lentiviruses. (A) mRNA expression level was confirmed by using Q-RT-PCR. HD Exon1 (red) represented the mRNA expression of mutant HTT and endogenous HTT, and HD Exon3/5 represented the mRNA expression of endogenous HTT.
“GFP-130Q non-ind.”, which is HD-293 FT cells without induction, and “GFP-130Q ind.”, which is induced HD-293 FT cells without shRNAs, were served as controls.
shRNA treated cells showed the
suppression of mutant HTT and endogenous HTT . (B) Protein expression of the mutant HTT in HD-293 FT cells was detected by Western Blotting analysis using the mEM48 antibody which recognized mutant HTT (top panel) and γ-tubulin antibody was used as an internal control (bottom panel). Non-Induced HD-293 FT cells (GFP-130Q non-Ind), induced HD-293 FT cells (GFP-130 Ind.), and wild-type 293FT cells were used as controls.
Figure 3. Suppression of mutant HTT in HD-293 FT cells infected with shRNA lentiviruses. (A-E) showed transmission light images and (F-J) showed fluorescent images.
GFP-HTT expression levels were revealed via the fluorescent intensity with the same exposure time. Non-Induced HD-293 FT cells (GFP-130Q non-Ind) and induced HD-293 FT cells (GFP-130Q Ind) were used as controls. Images inside small rectangles showed higher magnification. Scale bar: 40μM.
Figure 4. Suppression of mutant HTT in HD-293 FT cells infected with shRNA lentiviruses. (A-C) showed transmission light images and (D-F) showed merged pictures of fluorescent images. The expression of mutant HTT were showed in green due to expression of GFP fusion protein, and the expression of shRNA were indicated in red. Cells expressing red fluorescent signals showed weaker GFP proteins, suggesting the suppression of mutant HTT
via shRNA lentiviruses. Scale bar: 160μM.
Discussion:
The overall objective of this study is to investigate the therapeutic potential and systemic impacts of shRNA-HD on HD in vivo. Since in vivo systems, such as transgenic mice models, are labor-intensive and time-consuming, proper DNA constructs and stringent in vitro tests are required before applying to further in vivo systems. In this 9-month study, three shRNA-HD constructs were tested and exhibited various degree of suppression in overall HTT expression, including at mRNA and protein levels, which were comparable with reports on the down regulation of mutant HTT [10, 11, 13, 18, 19]. Interestingly, latest success in developing siRNAs that specifically target single nucleotide polymorphism in mutant HTT or the structure of HTT has further demonstrated the potential of siRNA therapy in HD [14, 20-22]. Therefore, establishment of one in vivo animal model is an important issue to further investigate the possibility of applying this technique.
To generate transgenic mice via lentiviral transgenesis, titer of viruses is a critical factor determining the transgenic efficiency [23, 24]. High titer lentiviruses reaching the titer of 108-109 CFU/mL were generated in this study, suggesting the feasibility of using these lentiviruses to produce transgenic mice via lentiviral transgenesis. Furthermore, the suppression ability of lentiviruses carrying shRNA-HD had been confirmed based on the epifluorescent images as well. Therefore, we have provided the evidence regarding to lentiviral functions in vitro, including suppression ability and infection ability, in this study, and these results support the convincing evidence to apply these lentiviruses for generating transgenic mice in vivo. For the future study, we would like to conduct lentiviral transgenesis to produce transgenic mice, and then evaluate the in vivo effects of shRNA-HD on this animal model.
With this achievement, we hope to provide a way to study the RNAi therapy for human disease.
專題研究計畫成果報告自評表
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)、是否適 合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
▓達成目標
□ 未達成目標(請說明,以 100 字為限)
□ 實驗失敗
□ 因故實驗中斷
□ 其他原因 說明:
2. 研究成果在學術期刊發表或申請專利等情形:
論文:□已發表 □未發表之文稿 □撰寫中 ■無 專利:□已獲得 □申請中 ■無
技轉:□已技轉 □洽談中 ■無 其他:(以 100 字為限)
3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以 500 字為限)
This 9-month study has finished the in vitro evaluation for shRNA-HD, showing the suppression ability on mutant HTT at RNA and protein level. These comprehensive in vitro results should provide enough confidence for us to generate transgenic mice which display the biofunction of shRNA-HD. shRNA transgenic mice model may be an alternative system to study the molecular, pathological and behavioral mechanisms of HD. Results from this future study should be able to provide more information for applying this potential therapeutic method in human.
Furthermore, this study also raises another issue of “zygotic therapy”, which may be considered as vaccines for specific human diseases, especially diseases caused by the abnormal gene expression. Over-expression or down-regulation of target genes in specific tissues may provide long-term therapy for these diseases, and this purpose is one of most important advantages by using gene therapy.
Another important contribution of this study is that the establishment of an animal model via shRNA down-regulation may provide an evaluation for knock-out study. Since knock-out mice model is a crucial approach to investigate gene functions in biomedical science, how to evaluate the feasibility of these candidate genes has emerged as an important challenge due to time-consuming and labor-intensive for this kind of studies. Generating knock-down mice via shRNA technique is a good preliminary data for knock-out experiments because these two approaches are all trying to decrease the expression of candidate genes. Therefore, if promising result can be seen in the knock-down mice in vivo, it will strongly suggest the feasibility of knock-out mice model. In addition, knock-down models are easier to conduct compared to knock-out models, implying the significant importance of this study for further applications.
References:
1. Li, S. and X.J. Li, Multiple pathways contribute to the pathogenesis of Huntington disease. Mol Neurodegener, 2006. 1: p. 19.
2. Walker, F.O., Huntington's disease. Lancet, 2007. 369(9557): p. 218-28.
3. Ferrante, R.J., et al., Histone deacetylase inhibition by sodium butyrate chemotherapy ameliorates the neurodegenerative phenotype in Huntington's disease mice. J Neurosci, 2003. 23(28): p. 9418-27.
4. Lecerf, J.M., et al., Human single-chain Fv intrabodies counteract in situ huntingtin aggregation in cellular models of Huntington's disease. Proc Natl Acad Sci U S A, 2001. 98(8): p. 4764-9.
5. Ona, V.O., et al., Inhibition of caspase-1 slows disease progression in a mouse model of Huntington's disease. Nature, 1999. 399(6733): p. 263-7.
6. Ferrante, R.J., et al., Therapeutic effects of coenzyme Q10 and remacemide in transgenic mouse models of Huntington's disease. J Neurosci, 2002. 22(5): p. 1592-9.
7. Schilling, G., et al., Coenzyme Q10 and remacemide hydrochloride ameliorate motor deficits in a Huntington's disease transgenic mouse model. Neurosci Lett, 2001. 315(3):
p. 149-53.
8. Schilling, G., et al., Environmental, pharmacological, and genetic modulation of the HD phenotype in transgenic mice. Exp Neurol, 2004. 187(1): p. 137-49.
9. Andreassen, O.A., et al., Creatine increase survival and delays motor symptoms in a transgenic animal model of Huntington's disease. Neurobiol Dis, 2001. 8(3): p. 479-91.
10. Harper, S.Q., et al., RNA interference improves motor and neuropathological abnormalities in a Huntington's disease mouse model. Proc Natl Acad Sci U S A, 2005.
102(16): p. 5820-5.
11. Rodriguez-Lebron, E., et al., Intrastriatal rAAV-mediated delivery of anti-huntingtin shRNAs induces partial reversal of disease progression in R6/1 Huntington's disease transgenic mice. Mol Ther, 2005. 12(4): p. 618-33.
12. Wang, Y.L., et al., Clinico-pathological rescue of a model mouse of Huntington's disease by siRNA. Neurosci Res, 2005. 53(3): p. 241-9.
13. Machida, Y., et al., rAAV-mediated shRNA ameliorated neuropathology in Huntington disease model mouse. Biochem Biophys Res Commun, 2006. 343(1): p. 190-7.
14. Drouet, V., et al., Sustained effects of nonallele-specific Huntingtin silencing. Ann Neurol, 2009. 65(3): p. 276-85.
15. Lois, C., et al., Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science, 2002. 295(5556): p. 868-72.
16. Chan, A.W., et al., Transgenic monkeys produced by retroviral gene transfer into mature oocytes. Science, 2001. 291(5502): p. 309-12.
17. Pinkert, C.A., Transgenic animal technology : a laboratory handbook. 2nd ed. 2002, Amsterdam London: Academic. 359-394.
18. Franich, N.R., et al., AAV vector-mediated RNAi of mutant huntingtin expression is neuroprotective in a novel genetic rat model of Huntington's disease. Mol Ther, 2008.
16(5): p. 947-56.
19. Huang, B., et al., High-capacity adenoviral vector-mediated reduction of huntingtin aggregate load in vitro and in vivo. Hum Gene Ther, 2007. 18(4): p. 303-11.
20. Lombardi, M.S., et al., A majority of Huntington's disease patients may be treatable by individualized allele-specific RNA interference. Exp Neurol, 2009. 217(2): p. 312-9.
21. Pfister, E.L., et al., Five siRNAs targeting three SNPs may provide therapy for three-quarters of Huntington's disease patients. Curr Biol, 2009. 19(9): p. 774-8.
22. Zhang, Y., J. Engelman, and R.M. Friedlander, Allele-specific silencing of mutant Huntington's disease gene. J Neurochem, 2009. 108(1): p. 82-90.
23. Yang, S.H., et al., Enhanced transgenesis by intracytoplasmic injection of envelope-free lentivirus. Genesis, 2007. 45(4): p. 177-83.
24. Yang, S.H., et al., Towards a transgenic model of Huntington's disease in a non-human primate. Nature, 2008. 453(7197): p. 921-4.