國立臺灣大學生命科學院生化科技學系 博士論文
Department of Biochemical Science and Technologhy College of Life Science
National Taiwan University Doctoral Dissertation
台灣紫芝多醣體免疫調節機轉與功能評估 Immunomodulating mechanisms and functional evaluation of the polysaccharides produced from the
submerged culture of Ganoderma formosanum
王正利 Cheng-Li Wang
指導教授:陳俊任 博士 劉文雄 博士 Advisor: Chun-Jen Chen, Ph.D.
Wen-Hsiung Liu, Ph.D.
中華民國 100 年 7 月
July, 2011
中文摘要
靈芝具有多種的藥理功能,在亞洲已有相當久遠的藥用歷史,台灣紫芝是台 灣原生種靈芝,其藥用相關研究卻相當少,因此本研究以液態深層醱酵培養法生 產台灣紫芝胞外多醣體,探討台灣紫芝多醣體之免疫調節功能、作用機制與功能 性評估。
台灣紫芝醱酵液經酒精沉澱和膠體過濾層析純化可分離得 PS-F1、PS-F2 和 PS-F3 三個主要分劃,其各分劃佔比分別為 16.24 ± 2.19 %、55.81 ± 2.97 %和 27.95
± 2.11 %。PS-F2 多醣體分子量約 14 KDa,主要單糖組成為 mannose、galactose、
glucose 及 fucose,分別佔 44.91 %、38.64 %、8.26 %及 8.02 %,而醣基鍵主要以 6-Gal 和 t-Man 鍵結為主。
PS-F2 能夠刺激小鼠巨噬細胞 RAW 264.7 產生 TNF-α 和 NO,並增強巨噬細
胞的吞噬能力,並能刺激巨噬細胞增生。以PS-F2 刺激小鼠腹腔能誘發急性發炎
反應,吸引嗜中性球和單核球來到腹腔。另外PS-F2 亦可刺激小鼠樹突細胞表面
成熟標誌CD40、CD80、CD86 及 MHC II 的表現增加,顯示 PS-F2 具有免疫調 節之活性,能活化先天性免疫反應。
PS-F2 作 用 的 受 體 研 究 方 面 , PS-F2 刺 激 巨 噬 細 胞 產 生 TNF-α 受 到 anti-Dectin-1 抗體、anti-CR3 抗體和 laminarin 顯著性抑制,TLR4 受體缺陷小鼠 BMDM 也降低了 PS-F2 刺激 BMDM 產生 TNF-α 的活性。以上結果顯示 PS-F2 能經由Dectin-1、CR3 及 TLR4 受體刺激巨噬細胞產生 TNF-α。
在 PS-F2 活化巨噬細胞作用機制部分, PS-F2 刺激能使 ERK、JNK 和 p38
絲裂原活化蛋白激酶磷酸化、I-κB 降解和 NF-κB 核移位反應。添加 ERK、JNK、
p38 和 NF-κB 訊息傳遞途徑專一性抑制劑皆可顯著抑制 PS-F2 刺激 RAW 264.7 細胞產生TNF-α,確認 PS-F2 透過活化絲裂原活化蛋白激酶和 NF-κB 訊息傳遞 途徑產生TNF-α。此外,以 Syk 抑制劑 piceatannol 處理後也會抑制 PS-F2 刺激 RAW 264.7 產生 TNF-α、I-κB 降解和 ERK 磷酸化,顯示 PS-F2 刺激 RAW 264.7
細胞會經由Dectin-1 受體透過 Syk 活化下游 NF-κB 和 ERK 訊息傳遞途徑。
在功能性評估方面,C57BL/6 小鼠以 PS-F2 每兩天管餵一次持續四週後,發 現小鼠脾臟細胞之T 淋巴球族群比例提高,血清中之 IgM 及 IgG 抗體濃度均有 顯著增加,顯示PS-F2 會活化 C57BL/6 小鼠體內非特異性免疫反應。
在抗腫瘤功能評估上,發現腹腔注射 PS-F2 可有效抑制 Sarcoma 180、B16 黑色素瘤細胞和C26 腫瘤細胞之生長,抑制率分別達 67.5 % 、23.7 %及 43.3 %;
以管餵方式給予小鼠PS-F2 亦有抗腫瘤之效果。腹腔注射 PS-F2 使接種腫瘤小鼠 脾臟細胞之T 淋巴球族群比例顯著性提高,其中 CD4 與 CD8 之細胞比例均有增 加,特別是CD4 細胞;而表現 CD49b 的 NK 細胞比例及其胞殺活性有些微增加;
而在血清中也發現I gM 抗體濃度有顯著增加。在 C26 腫瘤細胞模式中,腹腔注 射PS-F2 小鼠 CD4 與 CD8 T 細胞活化標誌表現 CD44high CD62Llow,顯示T 細胞 有顯著性活化;PS-F2 在 SCID 小鼠模式不具有抗腫瘤活性,以及 adoptive transfer 試驗也都近一步確認CD4 T 細胞和 B 細胞為主要影響抗腫瘤的免疫細胞。綜合
以上結果顯示PS-F2 能活化先天性免疫相關細胞及後天性免疫反應,透過活化細
胞介導免疫反應和體液免疫反應使腫瘤生長受到抑制,證實PS-F2 具有抗腫瘤活
性。
本研究確認以液態深層醱酵培養台灣紫芝生產之胞外多醣體 PS-F2 具有免 疫調節活性及抗腫瘤功能,確立台灣紫芝多醣體於機能性食品之用途與應用。
Abstract
The fungus of Ganoderma, called ‘Reishi’, has been used as traditional medicine in Asia and has been shown to exhibit various pharmacological functions. In this study, we investigated the immunomodulating activities and antitumor functions of polysaccharides produced from the submerged culture of Ganoderma formosanum, a native species isolated in Taiwan.
The crude polysaccharides could be separated into three main fractions according to their sizes after alcohol precipitation and gel filtration. These three fractions were designated as PS-F1, PS-F2 and PS-F3, and the yields were 21.9% ± 2.19 %, 55.81 ± 2.97 %, and 27.95 ± 2.11 %, respectively. The molecular weight of PS-F2 was about 14 KDa. The monosaccharide composition analyse by HPLC-PDA method demonstrated that PS-F2 was composed of mannose, galactose, glucose and fucose with the mole percentage of 44.91:38.64:8.26:8.02. The mainly glycosyl linkages were 6-Gal and t-Man.
We found that PS-F2 could stimulate RAW 264.7 murine macrophage cells to produce TNF-α and nitric oxide, and to enhance the phagocytic activity and proliferation of macrophages. In vivo, PS-F2 challenge in mice triggered an inflammatory response and led to the recruitment of neutrophils and monocytes. In addition, treatment of bone marrow-derived dendritic cells with PS-F2 resulted in the
enhanced cell-surface expression of CD40, CD80, CD86 and MHC II, suggesting that PS-F2 was able to induce the maturation of immature dendritic cells and activation of innate immune response.
Studies on the immune receptors for PS-F2 stimulated, PS-F2-stimulated TNF-α production in macrophages was significantly reduced in the presence of anti-Dectin-1 Ab and anti-CR3 Ab blocking antibody or laminarin. Moreover, PS-F2 induced TNF-α production by BMDM from C3H/HeN mice, but not C3H/HeJ mice that have a mutated TLR4 molecule, suggesting that TLR4 was also involved in PS-F2-mediated macrophage activation. In conclusion, we demonstrated that PS-F2 induced TNF-α production through the Dectin-1, CR3, and TLR4 membrane receptors in murine macrophages.
About signaling mechanisms of cytokine production upon PS-F2 stimulation, we found that PS-F2 stimulated the phosphorylation of MAP kinases (ERK, JNK and p38), degradation of I-κB and the nuclear translocation of NF-κB. TNF-α production was decreased in the presence of specific inhibitors of MAP kinases and NF-κB in RAW 264.7 cells, suggesting that MAP kinases and NF-κB pathways play a crucial role in activating TNF-α expression upon PS-F2 stimulation. In addition, treatment of macrophages with the piceatannol also inhibited TNF-α production, I-κB degradation and ERK phosphorylation, indicating that Syk functioned upstream of other signaling
events. These results may have important implications for our understanding on the molecular mechanisms in immunomodulating activities of PS-F2.
We further investigated the immunomodulating functions and antitumor activities of PS-F2 in vivo. We examined the effect of oral administration on the immune fnctions in C57BL/6 mice with PS-F2, 50 mg/kg PS-F2 once every two days for 4 weeks. PS-F2 administration resulted in increased in the splenic CD3 T lymphocyte populations and the IgM and IgG levels in serum. It demonstrated that oral administration of PS-F2 activated non-specific immune response in C57BL/6 mice.
Furthermore, we examined the effect of PS-F2 administration on tumor growth in an allogeneic and two syngeneic tumor models. Intraperitoneal administration of PS-F2 markedly inhibited the growth of sarcoma 180, C26, and B16 melanoma in mice and the inhibition rates were 67.5 %, 23.7 %, and 43.3 %, respectively. PS-F2 administration resulted in increases in the splenic CD3, CD4, and CD8 T lymphocyte populations, as well as the serum IgM level. Although PS-F2 administration did not affect the frequency of NK population in the spleen, we observed a slight increase in cytotoxic activity in PS-F2-treated mice. In C26-bearing mice, PS-F2 induced a CD44high CD62Llow phenotype on CD4 and CD8 T cells isolated from spleen of treated mice, suggesting the splenic T cells activated in PS-F2-trated mice. We further
used B16 tumor xenografted SCID mice model and adpotive transfer to find out whether transfer of immune cell from PS-F2-treated mice might be sufficient to prevent tumor development. We found that adoptive fransfer CD4 T cells and serum antibodies into B16 tumor bearing mice significantly inhibited B16 progress in recipient mice. Overall, our results using different tumor cell lines and different mouse strains indicated that PS-F2 administration could protect mice from tumor challenge. PS-F2 administration may exert the anti-tumor effect via modulating the functions of both T and B cells in mice and enhancing cell-mediated immunity and humoral inmunity.
The studies demonstrated that PS-F2 exhibited immunomodulating activities and antitumor response form the submerged culture of G. formosanum. Therefore, the polysaccharides PS-F2 from G. formosanum have the potential to be used as an immunomodulating agent in functional food for tumor therapy.
目錄
中文摘要... I Abstract...IV 目錄...VIII 圖目錄...XI 表目錄...XIII 縮寫表...XIV
第一章 序論...1
一、靈芝的藥用功能...1
1. 認識靈芝...1
2. 靈芝的生理活性...2
3. 靈芝的活性成分...2
二、靈芝的栽培與量產...7
1. 子實體...9
2. 菌絲體...9
三、多醣體的生物活性...10
1. 生物活性多醣體...10
2. 抗發炎活性...12
3. 抗氧化活性...12
4. 護肝效果...14
5. 修復傷口...14
6. 心血管疾病預防...14
7. 減緩自體免疫疾病...15
8. 改善糖尿病症狀...15
9. 抗皰疹活性...16
10. 保護神經功效...16
11. 免疫調節功能...16
12. 抗腫瘤活性...18
四、免疫與腫瘤...20
1. 先天性免疫...20
2. 後天性免疫...22
4. 訊息傳導路徑...28
5. 細胞激素...30
6. 腫瘤的治療...30
7. 生物性抗癌療法...32
1. 低分子量化合物...34
2. 高分子量化合物...34
六、研究架構...41
1. 動機與目的...41
2. 研究架構...42
第二章 台灣紫芝多醣體刺激小鼠巨噬細胞活化及樹突細胞成熟...45
一、前言...45
二、材料與方法...47
1. 實驗菌株、細胞株及動物...47
2. 培養基組成...47
3. 培養方法...47
4. 胞外多醣體之回收及純化...48
5. 測定總醣及蛋白質含量...49
6. 單糖組成分析...49
7. 醣鍵結分析...50
8. 分子量測定...50
9. β-1,3-glucan 含量分析 ...50
10. 多醣體刺激小鼠巨噬細胞產生腫瘤壞死因子...51
11. 多醣體刺激小鼠巨噬細胞產生一氧化氮...51
12. 多醣體刺激小鼠巨噬細胞增生之影響...52
13. 製備小鼠骨髓細胞分化之樹突細胞...52
14. BMDCs 成熟標記之分析 ...52
15. 巨噬細胞吞噬作用分析...53
16. 西方轉漬法及抑制劑試驗...54
17. 製備西方轉漬法細胞質與細胞核萃取液...55
18.參與細胞膜受體之分析...55
19. 製備小鼠骨髓分化巨噬細胞...55
20. 統計分析...56
三、結果...57
1. 以膠體過濾層析分離純化台灣紫芝多醣體...57
2. 台灣紫芝多醣體之單糖組成及分子量...57
3. PS-F2 醣基鍵結分析...61
4. β-1,3-glucan 的含量 ...61
5. PS-F2 刺激巨噬細胞產生 TNF-α...65
6. PS-F2 刺激 RAW 264.7 細胞產生一氧化氮...69
7. PS-F2 可增強 RAW 264.7 細胞的吞噬作用...71
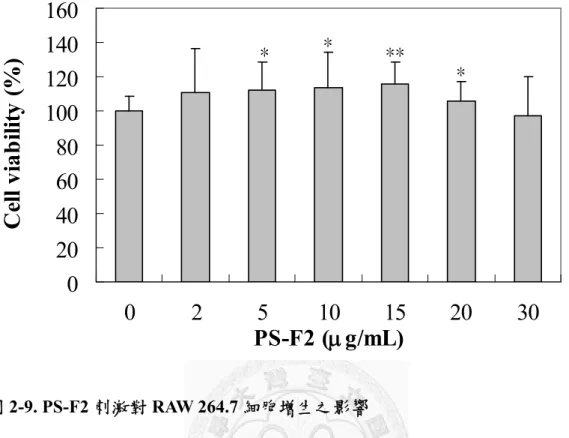
8. PS-F2 會刺激 RAW 264.7 細胞的增生...71
9. PS-F2 刺激未成熟樹突細胞成熟...73
10. PS-F2 會誘發小鼠體內發炎反應...73
11. PS-F2 誘導絲裂原活化蛋白激酶磷酸化...76
12. PS-F2 刺激 NF-κB 的活化 ...80
13. 甘露糖受體在 PS-F2 誘導產生 TNF-α 上扮演的角色...83
14. Dectin-1 受體在 PS-F2 誘導產生 TNF-α 上扮演的角色...83
15. PS-F2 透過 Dectin-1 受體下游的 NF-κB 和 ERK 活化 ...85
16. CR3 受體在 PS-F2 誘導產生 TNF-α 上扮演的角色 ...88
17. TLR4 受體在 PS-F2 誘導產生 TNF-α 上扮演的角色 ...88
四、討論...93
第三章 評估台灣紫芝多醣體之抗腫瘤活性...106
一、前言...106
二、材料與方法...108
1. 菌株、細胞株及動物...108
2. 非特異性免疫反應評估...108
3.抗腫瘤活性之評估...108
4. 血清中抗體之測定...109
5. 脾臟細胞製備...109
6. 脾臟細胞族群比例分析...110
7. NK 細胞毒殺活性之測定...110
8. MTT 法分析細胞存活 ...110
9. 細胞介導毒殺活性分析... 111
10. 免疫細胞純化... 111
11. adoptive transfer...112
12. SCID mice 之抗腫瘤模式...112
13. 統計分析...112
三、結果...113
1. 台灣紫芝多醣體於動物體內引起之非特異性免疫反應...113
2. 台灣紫芝多醣體抗 S180 腫瘤之活性 ...120
3. 台灣紫芝多醣體抗 B16 melanoma 腫瘤之活性 ...124
4. 台灣紫芝多醣體抗 C26 腫瘤之活性...129
5. PS-F2 促進 T 細胞活化 ...142
6. SCID mice 之抗腫瘤模式...149
第四章 結論...163
第五章 參考文獻...168
圖目錄
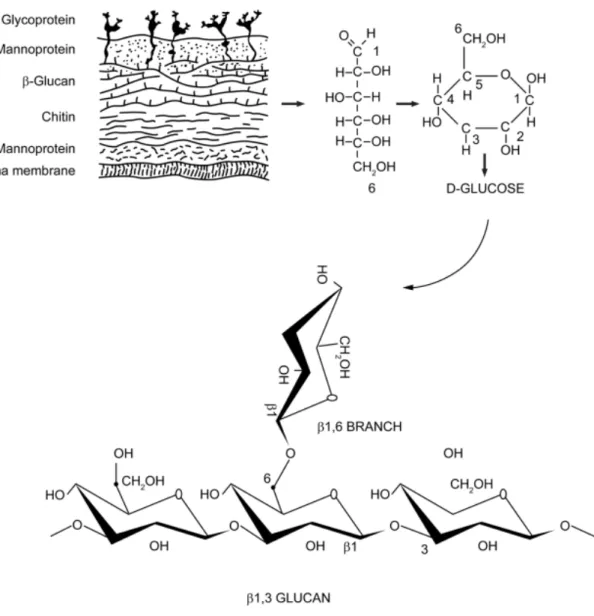
圖1-1. 真菌細胞壁成分及 β-glucan 結構 ...6
圖1-2. 生產赤芝子實體與菌絲體的主要栽培方式 ...8
圖1-3. 先天性免疫反應與後天性免疫反應相關之免疫細胞 ...21
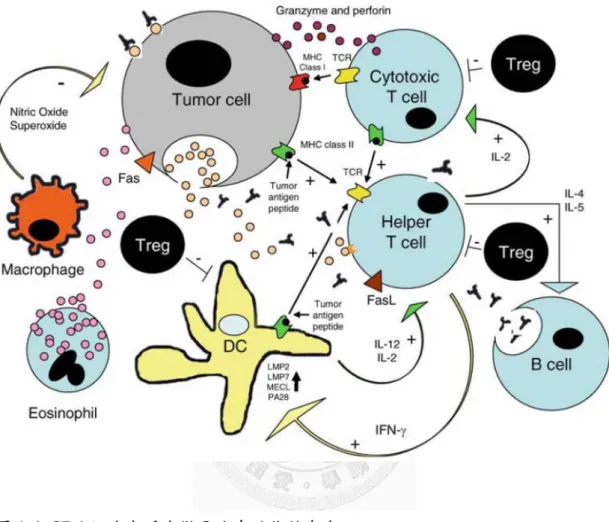
圖1-4. CD4 細胞在腫瘤微環境中的複雜角色...24
圖1-5. Candida albicans 辨識之免疫細胞及其模式辨識受體...27
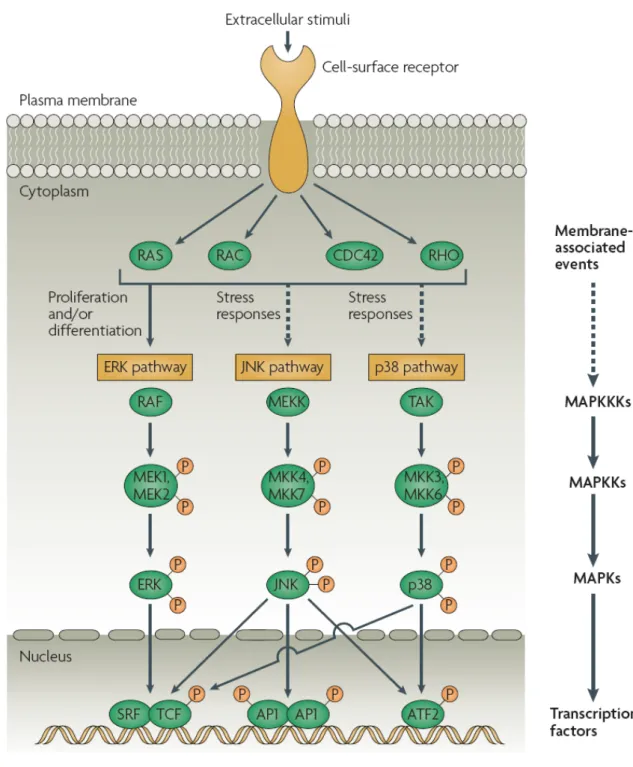
圖1-6. 哺乳動物三種絲裂原活化蛋白激酶訊息傳導途徑簡圖 ...29
圖1-7. NF-κB 的活化路徑 ...31
圖1-8. 食用菇菌中具有抗腫瘤功能之有效成分 ...37
圖1-9 . 靈芝多醣體 GLPS 對免疫細胞之免疫調節機制 ...40
圖1-10. 本研究之基本架構 ...44
圖2-1. 台灣紫芝粗多醣體之 Sepharose CL-6B 層析圖...58
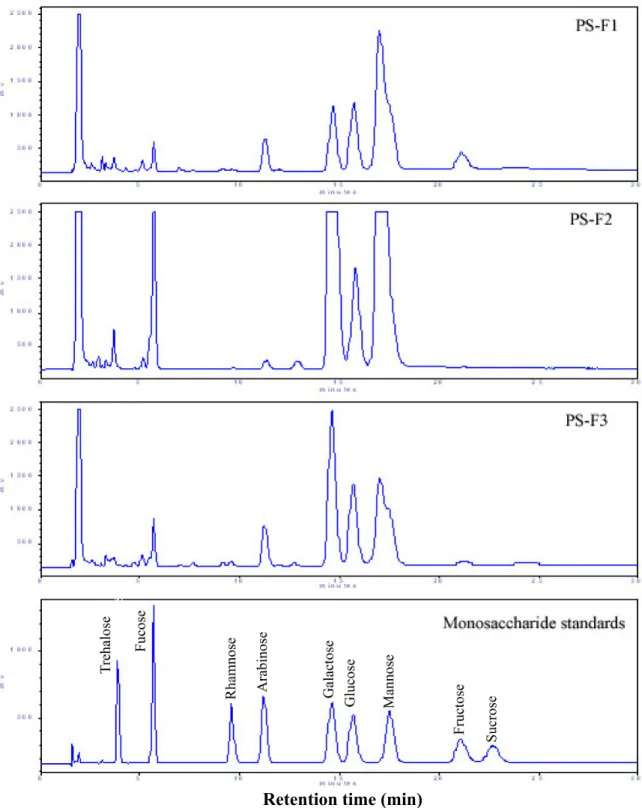
圖2-2. PS-F1、PS-F2 與 PS-F3 之 HPAEC-PAD 層析圖譜 ...59
圖2-3. 以 Sepharose CL-6B 層析管柱測定 PS-F2 分子量...62
圖2-4. PS-F2 之 GC-MS 圖譜...63
圖2-5. 以 aniline blue 方法測定 β-1,3-glucan 含量 ...66
圖2-6. 台灣紫芝多醣體會誘導 RAW 264.7 細胞產生 TNF-α 和 NO ...68
圖2-7. PS-F2 不具 LPS tolerance 效應...70
圖2-8. PS-F2 刺激對 RAW 264.7 細胞吞噬作用之影響 ...72
圖2-9. PS-F2 刺激對 RAW 264.7 細胞增生之影響 ...74
圖2-10. PS-F2 會誘導 C57BL/6 小鼠骨髓分化樹突細胞表面成熟標誌之表現 ...75
圖2-11. PS-F2 會刺激小鼠產生急性發炎反應 ...77
圖2-12. PS-F2 誘導絲裂原活化蛋白激酶磷酸化 ...79
圖2-13. PS-F2 誘導 I-κB 的降解及 NF-κB 之核移位...82
圖2-14. 甘露聚醣對 PS-F2 誘導 RAW 264.7 細胞產生 TNF-α 之影響 ...84
圖2-15. Dectin-1 受體抑制劑對 PS-F2 誘導 RAW 264.7 細胞產生 TNF-α 之影響 ...87
圖2-16. piceatannol 對 PS-F2 誘導 MAPKs 磷酸化及 I-κB 降解之影響 ...89
圖2-17. anti-CR3 中和抗體對 PS-F2 誘導 RAW 264.7 細胞產生 TNF-α 之影響..90
圖2-18. TLR 4 和 Dectin-1 參與 PS-F2 活化骨髓分化巨噬細胞產生 TNF-α...92
圖2-19. 赤芝多醣體的骨架 ...98
圖3-1. 以 PS-F2 管餵小鼠四週期間之體重變化 ...114
圖3-2. 以 PS-F2 管餵 C57BL/6 小鼠對脾臟中細胞比例之影響 ...115
圖3-3. 以 PS-F2 管餵 C57BL/6 小鼠對脾臟中 NK 細胞之影響...116
圖3-4. 以 PS-F2 管餵 BALB/c 小鼠對脾臟中細胞比例之影響...117
圖3-5. 以 PS-F2 管餵 BALB/c 小鼠對脾臟中 NK 細胞之影響...118
圖3-6. 以 PS-F2 管餵小鼠對血清中抗體濃度之影響 ...119
圖3-7. PS-F2 對 S180 之抗腫瘤效果...122
圖3-8. PS-F2 對 S180 腫瘤生長之影響...123
圖3-9. PS-F2 對接種 S180 小鼠脾臟重量之影響...125
圖3-10. PS-F2 對接種 S180 小鼠脾臟細胞族群比例之影響...126
圖3-11. PS-F2 對接種 S180 小鼠脾臟中 NK 細胞之影響 ...127
圖3-12. PS-F2 對接種 S180 小鼠血清抗體濃度之影響...128
圖3-13. PS-F2 對 B16 抗腫瘤之效果 ...130
圖3-14. PS-F2 對 B16 腫瘤生長之影響 ...131
圖3-15. PS-F2 對接種 B16 小鼠脾臟重量之影響...132
圖3-16. PS-F2 對接種 B16 小鼠脾臟細胞族群比例之影響...133
圖3-17. PS-F2 對接種 B16 小鼠脾臟 NK 細胞之影響...134
圖3-18. PS-F2 對接種 B16 小鼠血清抗體濃度之影響...135
圖3-19. PS-F2 對 C26 抗腫瘤之效果 ...137
圖3-20. PS-F2 對 C26 腫瘤生長之影響 ...138
圖3-21. PS-F2 對接種 C26 小鼠脾臟重量之影響...139
圖3-22. PS-F2 對接種 C26 小鼠脾臟細胞族群比例之影響...140
圖3-23. PS-F2 對接種 C26 小鼠脾臟 NK 細胞毒殺活性之影響...141
圖3-24. PS-F2 對接種 C26 小鼠血清 IgM 濃度之影響...143
圖3-25. 接種 C26 小鼠 T 細胞 CD44 的表現情形...144
圖3-26. 接種 C26 小鼠 T 細胞 CD62L 的表現情形 ...145
圖3-27. PS-F2 對接種 C26 小鼠 Treg 細胞比例之影響 ...147
圖3-28. Adoptive transfer 免疫細胞對 B16 腫瘤生長之影響...148
圖3-29. Adoptive transfer 免疫細胞對小鼠存活率之影響 ...150
圖3-30. PS-F2 在 SCID 小鼠抗腫瘤之效果 ...151
圖3-31. 腫瘤碎片刺激 CD4 和 CD8 細胞之反應 ...158
圖4-1. PS-F2 刺激巨噬細胞產生 TNF-α 之受體及訊息傳導途徑簡圖 ...164
圖4-2. PS-F2 抗腫瘤之免疫調節功能 ...166
表目錄
表1-1. 台灣地區常見的野生靈芝種類及特性 ...1
表1-2. 赤芝的生物活性成分及其藥用功能 ...5
表1-3. 藥用真菌類的 β-glucans 活性成分...11
表1-4. 高等擔子菌綱其抗腫瘤化學結構及免疫調節多醣體 ...13
表1-5. 真菌辨識有關之模式辨識受體 ...13
表1-6 食用菇菌中小分子之抗腫瘤有效成分 ...38
表1-7 菇類中具抗腫瘤活性之高分子化合物 ...39
表2-1. PS-F1、PS-F2 與 PS-F3 之單糖組成 ...60
表2-2. 以 GC-MS 分析 PS-F2 之醣基鍵結及滯留時間...64
表2-3. 不同靈芝來源多醣體之單糖組成 ...95
表3-1. PS-F2 對小鼠 CD4/CD8 比率之影響 ...160
縮寫表
7/4 neutrophil- specific rat mAb
AC-PS polysaccharides derived from Antrodia camphorata ALT alanine transaminase
AP-1 activator protein 1
APBP acidic protein bound polysaccharide APCs antigen-presenting cells
B16 B16 melanoma
Bax Bcl-2–associated X protein
BCG Bacillus Calmette-Guerin bcl-2 B-cell lymphoma 2
Blimp-1 B lymphocyte-induced maturation protein BMDCs bone marrow-derived macrophages BRM biological response modifier BSA bovine serum albumin
C26 colon cancer cells
CARD9 caspase recruitment domain protein 9 CD cluster of differentiation
cEPS crude extracellular polysaccharide ConA Concanavalin A COX-2 cyclooxygenase-2 CR3 Complement receptor type 3
DC dendritic cells
Dectin-1 dendritic-cell-associated c-type lectin-1 DMEM Dulbecco's modified Eagle's medium DMSO dimethylsulfoxide
DNA deoxyribonucleic acid DTT dithiothreitol
EAC Ehrlich ascites carcinoma ECL enhanced chemluminescence EDTA ethylenediaminetetraacetic acid EGTA ethylene glycol tetraacetic acid
ELISA enzyme-linked immunosorbent assay ERK extracellular signal-regulated kinase FBS fetal bovine serum FcγR Fcγ receptor
G-CSF granulocyte colony-stimulating factor GLP polysaccharides from Ganoderma lucidum GLP Ganoderma lucidum polysaccharide GLPG Ganoderma lucidum proteoglycan
GLPL low-molecular-weight polysaccharides GLPP Ganoderma lucidum polysaccharide preparation GLPS Ganoderma lucidum polysaccharides
Gl-PS polysaccharide extract isolated from Ganoderma lucidum GM-CSF granulocyte macrophage colony-stimulating factor GOT glutamate oxaloacetate transaminase
GRN grifolan HDAC-1 histone deacetylase 1
HEPES 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid HPAEC-PAD high-performance anion-exchange chromatography-pulse
amperometric detector
HPLC high performance liquid chromatography HRP horseradish peroxidase HSV-1, 2 herpes simplex virus 1 and 2 IFN-γ interferon-gamma IL-1β interleukin-1β
iNOS inducible nitirc oxide synthase ip intraperitoneal ITAM tyrosine-based activation-like motif iv intravenous
I-κB inhibitor protein κB JNK c-Jun N-terminal kinase
LAL Limulus amebocyte lysates LDH lactate dehydrogenase LPS lipopolysaccharide Ly-6G myeloid differentiation antigen Gr-1 LZ Ganoderma extract
MAPK mitogen-activated protein kinase M-CSF macrophage colony-stimulating factor MFI mean fluorescence intensities MG132 N-(benzyloxycarbonyl)leucinylleucinylleucinal MHC II major histocompatibility complex class II MMP matrix metalloproteinase MnSOD manganese superoxide dismutase
MR mannose receptor
MTT 3-(4,5-dimethtithiazol-2-yl)-2,5-diphenyltetrazolium bromide NF-κB nuclear factor κB
NK natural killer cells
NO nitric oxide
OX40 a member of the TNFR-superfamily of receptors PAMPs pathogen-associated molecular patterns
PBS phosphate buffered saline PDA potato dextrose agar
PI3K phosphoinositide 3-kinases PKC protein kinase C
PKC-γ protein kinase C-γ
PMSF phenylmethanesulfonylfluoride Poly I:C polyinosinic:polycytidylic acid
PRRs pattern recognition receptors PSG-1 Ganoderma atrum polysaccharide PSK krestin
PSK protein-bound polysaccharide K ROS reactive oxygen species
RPMI1640 Roswell Park Memorial Institute 1640 RR Rokkaku-reishi
S-180 sarcoma 180
SB202190 4-[4-(4-Fluorophenyl)-5-(4-pyridinyl)-1H-imidazol-2-yl]phenol sc subcutaneous
SCID severe combined immune deficiency
SD standard deviation
SDS-PAGE sodium dodecyl sulfate polyacrylamide gel electrophoresis
SE standard error
SP600125 1,9-pyrazoloanthrone SPF specific pathogen free
Syk spleen tyrosine kinase TBS tris buffered saline
TFA trifluoroacetic acid
THP-1 human acute monocytic leukemia cell line TLR Toll-like receptors
TMB 3,3’,5,5’-tetramethylbenzidine TNF-α tumor necrosis factor-alpha
TRIF TIR-domain-containing adapter-inducing interferon-β
U0126 1,4-diamino-2,3-dicyano-1,4-bis[2-aminophenylthio] butadiene YAC-1 T cell lymphoma
第一章 序論
一、靈芝的藥用功能
1. 認識靈芝
靈芝在中國已有久遠的使用歷史,一直被視為稀有珍貴之藥材。神農本草經 記載,靈芝為草藥中之上藥,「上藥」又稱為神藥,也就是養生之藥,具有調整
體質及增強體魄之功效;而「中藥」是指能預防疾病並補充體力的藥材;「下藥」
則只能用於對症治療,不可經常服用的治療藥。
目前至少有270 種食用菇類被發現具有治療用途之特性。靈芝分類上屬於真
菌界(Myceteae)、無鞭毛菌門(Amastigomycota)、擔子菌綱(Basidiomycetes)、
無蕈褶目(Aphyllophorales)、多孔菌科(Polyporaceae),靈芝屬(Ganoderma)。
子實體為一年生,生長於闊葉樹之腐木,分布廣及熱帶至溫帶地區 (Wasser, 2002)。
靈芝子實體具有光亮的表皮,菌蓋有漆狀分泌物,可利用木質素及纖維素,
擔孢子呈卵形,具雙層壁,會分泌漆化酵素(laccase)和過氧化酵素(peroxidase)。 依據菌蓋外觀顏色的不同可將靈芝分為青、赤、黃、白、黑和紫色六種,由於菇 類的子實體是長期適應環境演化而完成特定繁殖功能的組合,因此在靈芝異種間 的子實體中,無論就外部形態和內部解剖構造上皆有很大的差異,每一種均有不 同種類之功效 (Wasser, 2002; Sanodiya et al., 2009)。古籍中記載靈芝的分佈可分 成兩大區域,赤芝、紫芝和黑芝產於衡山、高夏山及常山等,屬於長江流域;而 黃芝、白芝和青芝生於嵩山、華山及泰山等,屬於黃河流域,顯示靈芝的生態習 性與分佈區域有密切的關係。
大部分的靈芝子實體具有菌蓋和菌柄兩個部份。菌蓋呈半圓形、腎形、貝殼 形等,表面有同心環紋或放射狀縱紋,具有漆樣光澤,有時因生長成熟之程度不 同而同時存在數種不同顏色之區域,一般而言其生長區域位於菌蓋的最外緣,菌 蓋外緣顏色最淡而向內則顏色漸深。菌柄亦具有漆樣光澤,顏色與菌蓋相同或較 深,另外有些靈芝子實體則直接著生於寄主之上,只有短基柄或是無菌柄(表 1-1 )。 台 灣 常 見 的 野 生 靈 芝 有 樹 舌 靈 芝 ( G. applanatum )、 台 灣 紫 芝 ( G.
formosanum)、拱狀靈芝 (G. fornicatum)、赤芝(G. lucidum)、新日本靈芝(G.
neo-japonicum)、熱帶靈芝(G. tropicum)、松杉靈芝(G. tsugae)和小孢子靈芝
(G. microsporum)八種,其外觀及特性整理於表 1-1 (許, 1993)。
2. 靈芝的生理活性
近年來,台灣、日本、中國、韓國及美國等地學者發表相當多的研究報告證 實靈芝的藥用效果,以現代的生物技術、藥學及臨床研究來看靈芝的活性成分及 功能,靈芝對抗腫瘤、提升免疫調節功能、降血壓、降血脂、保肝、抗動脈硬化、
降低自由基、抑制血小板凝集、抗 HIV 病毒、抗細菌、抗寄生蟲、抗過敏、治
療氣喘等 (Lin, 2005; Yuen and Gohel, 2005; Paterson, 2006; Boh et al., 2007;
Sanodiya et al., 2009)都有不錯的效果。其中又以其抗腫瘤及免疫調節功能研究的 最多,被視為最主要的生理功能。
3. 靈芝的活性成分
而靈芝的藥用活性成分已被證實的有三萜類(triterpenoids)、多醣體
表1-1. 台灣地區常見的野生靈芝種類及特性
Table 1-1. The kinds and charatertistics of wild Ganoderma in Taiwan (許,1993).
子 實 體 孢 子
學 名
菌 柄 顏色 菌蓋形狀 漆光 大小(µm) 特性 寄 生 俗 名
樹舌靈芝G. applanatum 無或短柄 灰白至灰褐色 半圓形,不規
則形 無 7-10×4.3-6.2 闊葉樹、針葉
樹、棕櫚
白芝、平蓋靈芝、
古梅靈芝、扁芝 赤芝 G. lucidum 有或短柄 黃褐色、紅褐色 半圓形,腎形 有 8.5-11.2× 5.2-7 闊葉樹 赤芝、相思赤芝、
淺山赤芝
松杉靈芝 G. tsugae 有 紅褐色、紫褐色 腎形,近扇形 有 9-11 × 6-8 針葉樹、闊葉樹 高山赤芝、赤芝
台灣靈芝 G. formosanum 有 暗紫色、黑紫色 近圓形,腎形 有 11.5-13×7-8.5 頂端透
明凸起 楓樹 紫芝、黑芝、玄芝
新日本靈芝
G.neo-japonicum 有 褐色、紫褐色 近圓形,腎形 有 10-12.5 × 7-8.5 頂端透
明凸起 竹類 竹生赤芝、竹芝、
紫芝
拱狀靈芝G. fornicatum 無或短柄 紅褐色至褐色 近圓形,腎形 較弱 8.7-10.4× 5.2-7 闊葉樹 無柄赤芝、紅樹舌
熱帶靈芝 G. tropicum 有 黃褐色、紫褐色 半圓形,漏斗
形 較弱 8.5-11.5× 5.2-6.9 豆科植物 赤芝、淺山赤芝、
相思赤芝 小孢子靈芝
G. microsporum 無或短柄 古銅形、紫黑色 貝殼形,不規
則形 有 6-8.5 × 4.5-5 頂端透
明凸起 柳樹 青芝
(polysaccharides)、核苷酸、蛋白質、胺基酸、生物鹼及多種微量元素等 (Yuen and Gohel, 2005; Paterson, 2006),三萜類與多醣體的功效更是備受矚目。目前從 靈芝分離純化的有效藥用成分已高達 400 種以上 (Shiao, 2003; Sanodiya et al., 2009),表 1-2 為已發表之靈芝活性成分及生理活性,主要成分陳述如下:
3.1 多醣體
真菌來源的多醣體具有刺激免疫系統,保護身體免受病原性微生物或是外來 毒素的侵襲 (Brown and Gordon, 2005)。赤芝多醣體中的 β-1,3-glucan 主鏈和 β-1,6- glucan 支鏈(圖 1-1)也已證實具有抗腫瘤及免疫調節功能 (Miyazaki and Nishijima, 1981)。近來的研究更顯示萃取自子實體、菌絲體或孢子的靈芝多醣體 對巨噬細胞 (Hua et al., 2007; Ji et al., 2007)、B 細胞 (Zhang et al., 2002b; Lin et al., 2006a; Zhuang, 2009)、樹突細胞 (Cao and Lin, 2002; Lin et al., 2006b)及嗜中性白 血球 (Hsu et al., 2003)均具有免疫調節的功能。
3.2 三萜類
三萜類為靈芝的二級代謝產物,也是靈芝苦味的主要來源,超過 150 種以上 的三萜類已從靈芝中被分離鑑定,大多屬於氧化型羊毛固醇類衍生物,像是 ganoderic、lucidenic、ganodermic、ganoderenic、ganolucidic、applanoxidic acids、
lucidones、ganoderals 和 ganoderols 等結構 (Boh et al., 2007)。靈芝中的三萜類 主要功能有抗腫瘤、保肝、抗血管新生、抗高血壓 (Kabir et al., 1988; Lee and Rhee, 1990)、降膽固醇、抑制組織胺釋放、促進血小板凝集及抗 HIV 病毒 (Paterson, 2006; Boh et al., 2007)。
表1-2. 赤芝的生物活性成分及其藥用功能
Table 1-2. Biological activities and pharmacological functions reported for the fungus Ganoderma lucidum (Sanodiya et al., 2009).
Bioactive Compound Therapeutic Effects Protein LZ-8, β-D-glucan, Immunomodulation
Polysaccharides, glycoproteins,
lanostanoid, Triterpenoids , and steroid
Anti-cancer, anti-tumor, chemo and radio prevention
Polysaccharides Anti-herpetic Anti-ulcerogenic Polysaccharides, ganodermin Anti-microbial
Triterpenoids Anti-HIV-1and Anti-HIV-1-protease
Glycans Anti-diabetic Ganoderic acids R and S and
ganosporeric acid A Hepatoprotective
Ganoderic acid Immunomodulation, Anti-inflammatory, Anti-allergic
Ganoderic acid T Anti-proliferative activity ganoderol B Anti-androgenic
Ethanol extract Anti-angiogenic activity Estrogenic
Chloroform extract Anti-oxidant Methanol extract Anti-mutagenic Powdered mycelium and water extract of
mycelium Cardiovascular and circulatory functions
圖1-1. 真菌細胞壁成分及 β-glucan 結構
Figure 1-1. β-glucan is one of the key components of the fungal cell wall (Chan et al., 2009). The basic subunit of the fungal β-glucan is β-D-glucose linked to one another by 1-3 glycosidic chain with 1-6 glycosidic branches. The length and branches of the β-glucan from various fungi are widely different.
3.3 蛋白質及胺基酸
1989 年 Tanaka 等人從赤芝純化出具有免疫調節功能之 LZ-8 蛋白質,研究 顯示LZ-8 具有抗腫瘤及治療自體免疫疾病之效果 (Tanaka et al., 1989),是這幾
年很熱門研究材料。而分離自赤芝的 GLL-M 屬於一種親醣蛋白(lectin)
(Kawagishi et al., 1997);另外也在靈芝發現一些具生物活性之胜肽及胺基酸 (Shiao, 2003; Paterson, 2006; Boh et al., 2007; Sanodiya et al., 2009)。
3.4 核苷酸
從 G. capense 的菌絲體中分離出多種的核苷酸,有 adenine、adenosine、uracil 和 uridine (Boh et al., 2007)。水溶性的核苷酸發現具有抑制血小板凝集的功能 (Shimizu et al., 1985)。
3.5 其他微量元素
微量元素有鉀、鈣、磷、鎂和有機鍺 (水野和川合, 1997; Chiu et al., 2000)。
二、靈芝的栽培與量產
由於靈芝的藥用保健功能,早在中國古代已被視為珍貴藥材,野生的靈芝非 常稀少,在整個保健食品市場的需求日益擴增,野生靈芝早已不符所需,1971 年日本人直井幸雄氏成功建立量產靈芝子實體的方法,人工栽培的靈芝已成為供 應市場的主要來源。
人工栽培靈芝可分成子實體與菌絲體的生產兩種(圖 1-2)。子實體傳統的栽 培方式是以包裝木屑、瓶裝木屑或直接栽種在段木上;這種在段木或以木屑、麥
圖1-2. 生產赤芝子實體與菌絲體的主要栽培方式
Figure 1-2. Main cultivation methods for the production of G. lucidum fruit bodies and mycelia (Boh et al., 2007).
桿及稻桿填充的太空包的農場化栽培方式已發展三十幾年。另一種是在生物反應 器培養的生產技術,以固態基質或是液態基質,在無菌的反應器中進行培養,作 為小型或試驗型的生產工場 (Boh et al., 2007)。
1. 子實體
在過去,靈芝栽培於沒有滅菌的天然原木,可能需要 2-3 年才能獲得成熟的 子實體,這樣的生產方式培養時間長,也需要投入大量勞力。1980 年代後期,
開始把使用的原木裁剪成較短的段木來栽培,菌絲培養只要4-5 個月,而子實體
也可在同年收穫,增加資金的週轉;而使用鋸木屑栽植子實體從 1990 年代開始
發展。赤芝的栽培可利用充填鋸木屑的耐熱性聚丙烯瓶子或袋子、玻璃瓶、玻璃 罐、托盤或苗床中成長,置於可調控溫度與溼度的菇舍栽培,環境及品質更加能 控制 (Boh et al., 2007)。
2. 菌絲體
靈芝子實體的栽培、生長到可採收,一般需要幾個月到一年,這樣的時間及 成本已讓靈芝產品無法充分供應市場所需,使得價格居高不下,為突破這限制,
漸漸有人利用固態基質或液態培養基生產靈芝菌絲體。利用深層液態醱酵培養生
產可以在1-2 週的時間即可回收靈芝菌絲體及醱酵培養液,這樣的生產技術一樣
可以得到具生理功能的活性成分,惟其質與量受到靈芝菌種、培養條件、生長階 段、加工的程序及製備等因素影響 (Boh et al., 2007)。以深層液態醱酵培養技術 生產的培養液、菌絲體或是孢子已確認含有豐富的生物活性物質(Bao et al., 2002a; Bao et al., 2002b)。
三、多醣體的生物活性
多醣體是由多個以上的單糖以糖苷鍵結所組成,連結成直鏈或分支長鏈的大
分子(圖1-1),如澱粉、纖維素、菇菌多醣體等等,均是屬於多醣體,纖維素
和幾丁質為構成植物骨架及蝦蟹、昆蟲甲殼之多醣類,而澱粉和肝醣則為生物體 儲存能量的多醣物質。菇類及真菌中所含的多醣體,因本身分子量較大,多為十 萬以上,甚至上百萬。因此,所含的多醣類一般通稱為「多醣體」。
1. 生物活性多醣體
近年來,很多研究指出從植物、真菌、酵母、菇類及海藻等天然來源的多醣 體具有免疫調節的生理活性,其中的菇類早被人們用於食用及醫藥的應用,表 1-3 列出了藥用菇類的 β-glucan 活性成分。多醣體特殊的結構,如多醣體中心結 構的重複單位、環狀結構、糖苷鍵結位置、變旋異位構形(anomeric-center configurations)等會與病原或微生物表面產生作用,多醣體結構的多變性不同程 度影響生體的反應,具有調控高等生物細胞間互相作用的彈性 (Wasser, 2002;
Avci and Kasper, 2010)。主要具有生物活性的多醣體主要是水溶性的 β-1,3 和 β-1,6-glucan,可以經由添加酒精將多醣體沉澱分離。
一般多醣體的純化可結合多種的純化技術,如酒精沉澱、分劃沉澱、醋酸沉 澱 、 離 子 交 換 層 析 、 膠 體 管 柱 層 析 和 親 和 性 層 析 等 。 基 本 上 , 可 經 由 DEAE-cellulose 離子交換層析管柱分離中性多醣體。多醣體大多是以不同種類的 糖苷鍵結合成的多聚醣,如 β-1,3-glucan、β-1,6-glucan 及 α-1,3-glucan 和一些異 型葡聚醣(heteroglycans)。一般而言,除了glucan 是由 glucose 為骨幹的多醣體,
還有依骨架的單糖不同有galactans、fucans、xylans 和 mannans;異型多醣體(
表1-3. 藥用真菌類的 β-glucans 活性成分
Table 1-3. Selected medicinal mushroom with β-glucans as active components (Chan et al., 2009).
heteropolysaccharides)的側鏈可能接 glucuronic acid、xylose、galactose、mannose、
arabinose 或 ribose 等單糖或不同的組合 (Wasser, 2002)。多醣體骨架除了連接單 糖或寡醣外,也會與胜肽或蛋白質結合成醣蛋白 (Wang et al., 2002; Zhang et al., 2002b)。多醣體已被認為主要透過增強宿主免疫功能而具有抗腫瘤的效果,而不
是直接毒殺腫瘤細胞,表1-4 是在高等擔子菌發現具有抗腫瘤及免疫調節活性的
多醣體化學結構。研究人員發現來自靈芝子實體、孢子、菌絲體及培養菌液的多
醣體已超過200 種以上,不同的分子大小多醣體、單糖組成、單糖鍵結及側鏈修
飾等結構的差異均對生物活性有直接影響 (Huie and Di, 2004; Paterson, 2006;
Avci and Kasper, 2010)。
2. 抗發炎活性
研究發現赤芝的水萃取物可以有效抑制及減緩鹿角菜膠(carrageenan)引起 的水腫,具有抗發炎的活性 (Lin et al., 1993)。從赤芝子實體萃取的 GLPS 可減 緩 indomethacin 和醋酸所誘發小鼠胃粘膜發炎,其修復活性可能透過抑制產生 TNF-α 及誘發 c-myc 和 ornithine decarboxylase 基因的表現 (Gao et al., 2002)。
3. 抗氧化活性
在 UV 誘發光分解產生過氧化氫破壞 DNA 超螺旋 phiX174 的鍵結模式,赤 芝純化的胺基多醣可顯著性抑制脂質過氧化,並且能保護細胞,修復活性氧 (reactive oxygen species, ROS) 造 成 的 傷 害 (Lee et al., 2001) 。 另 以 tert-butylhydroperoxide(tBOOH)誘發 ROS 在體內或體外對巨噬細胞造成的損
害,而赤芝多醣體GLPP 可以有效增加巨噬細胞的存活率,對於細胞內胞器(粒
表1-4. 高等擔子菌綱其抗腫瘤化學結構及免疫調節多醣體
Table 1-4. Chemical structure of antitumor and immunostimulating polysaccharides of higher Basidiomycetes (Wasser, 2002; Ferreira et al., 2010).
Polysaccharide Species Homoglucans
α-(1→3)-glucan
Linear α-(1→3)-glucan α-(1→4)-; β-(1→6)-glucan α-(1→6)-; α-(1→4)- glucan β-(1→6)-glucan
β-(1→6)-; β-(1→3)-glucan β-(1→6)-; α-(1→ 3)-glucan β-(1→3)-glucuronoglucan
Heteroglucans Mannoxyloglucan Galactoxyloglucan Xyloglucan
Xylogalactoglucan Mannogalactoglucan Galactomannoglucan Arabinoglucan Riboglucan glucuronoglucan
Glycans Arabinogalactan Glucogalactan Fucogalactan
α-(1→6)-mannofucogalactan Fucomannogalactan
Mannogalactan Mannogalactofucan Xylan
Glucoxylan Mannoglucoxylan α-(1→3)-mannan Glucomannan
β-(1→2)-; β-(1→3)-glucomannan Galactoglucomannan
Galactomannan
Glucuronoxylomannans
Armillariella tabescens, Auricularia auricula, Agaricus blazei, Amanita muscaria, Agrocybe aegerita
Agaricus blazei Agaricus blazei
Lyophyllum decastes, Armillariella tabescens Agaricus blazei, Grifola frondosa
Agaricus blazei Ganoderma lucidum Grifola frondosa Hericium erinaceus
Grifola frondosa , Polyporus confluens Pleurotus pulmonarius, Agaricus blazei Inonotus obliquus
Pleurotus pulmonarius, Pleurotus cornucopiae Ganoderma lucidum, Agaricus blazei
Flammulina velutipes, Hohenbuehelia serotina, Leucopaxillus giganteus
Ganoderma tsugae
Agaricus blazei, Flammulina velutipes Ganoderma lucidum
Pleurotus citrinopileatus Ganoderma tsugae Sarcodon aspratus Fomitella fraxinea
Dictyophora indusiata, Grifola frondosa Pleurotus pulmonarius
Grifola frondosa Hericium erinaceus
Hericium erinaceus, Pleurotus pulmonarius Hericium erinaceus
Dictyophora indusiata Agaricus blazei Agaricus blazei Lentinus edodes Morchella esculenta Tremella fuciformis
白蛋白誘發人類腎近曲小管上皮細胞的氧化損害及細胞凋亡 (Lai et al., 2006)。
最近的研究更發現赤芝多醣體 GLPP 分離的低分子量多醣體 GLPL1 和 GLPL2 具有抗氧化活性,其中的 GLPL1 對游離自由基的清除與亞鐵離子螯合更 為有效 (Liu et al., 2010)。對於罹患糖尿病小鼠,赤芝 GLP 會影響胰腺抗氧化酵 素活性及脂質過氧化程度,透過調控bcl-2 表現及 bax/bcl-2 比率,與增加抗氧化 酵素活性,GLP 能有效減少氧化的損傷並抑制細胞凋亡 (Yang et al., 2010)。
4. 護肝效果
以赤芝、台灣紫芝和新日本靈芝的水萃物處理 CCl4引起肝損傷的小鼠,發 現能降低血清GOT 及 LDH 酵素活性,其中又以台灣紫芝的效果最好 (Lin et al., 1995)。另以 BCG 或 BCG 加細胞激素處理 BALB/c 小鼠誘發免疫肝損傷,GLP 可以有效減緩肝腫脹,降低血清中 ALT 酵素的釋出與 NO 的產生,抑制 iNOS 蛋白的表現 (Zhang et al., 2002a)。
5. 修復傷口
SACCHACHITIN®膜是由松杉靈芝子實體的加工剩餘物製作的皮膚替代 物,它是一種由60 % β-1,3-glucan 和 40 % N-acetylglucosamine 組成的絲狀結構 聚合物 (Su et al., 1997)。SACCHACHITIN 膜修復傷口的效果是對發炎細胞具有 趨藥性(chemotactic)的效果,而且能影響活化增生細胞細胞核抗原表現、type I collagen 表現、創傷組織的重塑和降低傷口治療組織 transglutaminase 及 MMP 的 表現 (Hung et al., 2001; Hung et al., 2004; Su et al., 2005)。
心血管疾病已是現在人的文明病,Lee 和 Rhee 以麻醉的兔子和大鼠評估赤 芝菌絲體水萃物對心血管作用的相關活性,發現水萃物在不影響心跳下可抑制腎 輸出交感神經活性,而降低血管的收縮壓與舒張壓 (Lee and Rhee, 1990)。
Li 等人也探討黑靈芝(Ganoderma atrum)多醣體 PSG-1 對心肌細胞及缺氧 /複氧(anoxia/reoxygenation, A/R)損傷引起的氧化壓力之影響,發現 PSG-1 能 保護心肌細胞受到缺氧/複氧損傷而死亡,減少缺氧/複氧損傷產生的活性氧、粒 腺體膜電位降低以及 cytochrome c 從粒腺體釋出到細胞質,抑制 caspase-9 和 caspase-3 的活化,改變 Bcl-2 家族蛋白質及增加心肌細胞 MnSOD 的蛋白表現量 (Li et al., 2010c)。
7. 減緩自體免疫疾病
自體免疫疾病(autoimmune diseases)是指生體對自身抗原產生過度免疫反 應而導致自身組織損害的疾病,常見之紅斑性狼瘡即是一種慢性自體免疫引起的 風濕疾病。在實驗動物模式,松杉靈芝萃取物能增加紅斑性狼瘡小鼠(lupus mice)
的存活率,降低尿蛋白及血清中抗雙股DNA 抗體之濃度,顯示靈芝多醣體能減
緩自體免疫病的症狀 (Lai et al., 2001)。而赤芝對於類風濕性關節炎(Rheumatoid arthritis)的症狀也有改善的效果 (Bao et al., 2006)。
8. 改善糖尿病症狀
以赤芝多醣體產品 Ganopoly 針對 71 名罹患第二型糖尿病患者進行臨床的功 效及安全性評估 。服用 Ganopoly 12 週患者的紅血球糖化血色素平均值從 8.4 % 降到 7.6 %,同時也可以改善空腹血糖及餐後血糖值;空腹胰島素、餐後 2 小時
服用Ganopoly 12 週患者結果顯示可有效降低血液中葡萄糖濃度,改善糖尿病症 狀(Gao et al., 2004)。
9. 抗皰疹活性
單純疱疹(Herpes simplex)是一種病毒傳染的疾病。其病原為單純疱疹病 毒第一型(HSV-1)及第二型(HSV-2),感染病毒後會有局部的原發性病灶,痊癒後 仍有復發的可能性。以 HSV-1 和 HSV-2 病毒感染 Vero 細胞,發現赤芝 APBP 多醣體具有抗病毒活性,當 APBP 結合抗皰疹用藥 acyclovir 使用更具有加乘的 治療功效 (Oh et al., 2000);此外赤芝菌絲體純化之 GLPG 也能干擾病毒複製,
進入感染細胞,抑制HSV 病毒感染細胞 (Liu et al., 2004)。
10. 保護神經功效
Zhao 等人探討小鼠皮質神經細胞在缺氧/複氧損傷過程,以 Gl-PS 處理的影 響情形 (Zhao et al., 2004)。Gl-PS (1、10、100 µg/ml)能增加神經細胞在缺氧/複 氧損傷的存活率,同時有效降低 malondialdehyde 含量、ROS 的產生及增強 MnSOD 的活性。
11. 免疫調節功能
健康食品的免疫調節功能評估,是針對包括非特異性及特異性免疫功能之評 估。所謂非特異性免疫力主要包括如嗜中性白血球(neutrophils)及單核球
(monocytes)的吞噬能力或是 NK 細胞的活性,很多研究均顯示多醣體在免疫 調節上的功效,對於各種免疫細胞之影響分述如下:
很多研究發現不同來源的多醣體會影響單核球與巨噬細胞(macrophages)
的活性。以PG-S 和 GLP 刺激人類血液單核球,其刺激培養液能夠誘導單核球增 生並分化為成熟的單核球及巨噬細胞,具有吞噬功能與產生超氧化物;刺激巨噬 細胞產生IL-1β、IL-2、IL-10、TNF-α 和 IL-6,也會使 T 細胞產生 IFN-γ (Lieu et al., 1992; Wang et al., 1997; Zhao et al., 2010)。
赤芝多醣體發現在體外能活化 BALB/c 小鼠 B 細胞和巨噬細胞,但是不能 活性T 細胞 (Shao et al., 2004)。餵食 BALB/c 小鼠 RR(500 mg/kg)14 天,LPS 和ConA 刺激小鼠脾臟貼附細胞產生 IFN-γ 和 IL-12,顯示餵食 RR 能活化脾臟 巨噬細胞及T 細胞,走向 Th1 的免疫反應 (Kohguchi et al., 2004)。
靈芝多醣體也發現能增加小鼠巨噬細胞表面的 CD14 和 TLR4 標誌,促進骨 髓分化巨噬細胞增生,產生NO,誘導細胞內呼吸爆發活性,增加 IL-1β、L-12p35 及IL-12p40 表現 (Wang et al., 2005; Hua et al., 2007; Ji et al., 2007)。顯示靈芝多 醣體,尤其是β-glucan,無論是在體內或體外,對於人類或小鼠巨噬細胞都有刺 激活化的功能。
11.2 增強吞噬作用
以靈芝多醣體處理巨噬細胞能夠增強巨噬細胞 (Lieu et al., 1992; Wang et al., 2005; Pang et al., 2007; Yin et al., 2007; Zhu et al., 2007; Chang et al., 2009)和嗜中 性球 (Hsu et al., 2003)的吞噬能力,靈芝多醣體透過 PI3K、p38 MAPK 和 PKC 訊息傳導路徑增強嗜中性球的吞噬能力。
11.3 活化自然殺手細胞
自然殺手細胞 (NK cells)為毒殺型淋巴球,屬於先天性免疫反應的免疫細 胞,對於抗腫瘤及細胞受病毒感染扮演關鍵角色。很多研究指出實驗小鼠腹腔注
射或餵食靈芝多醣體能夠活化NK 細胞 (Hsu et al., 2002; Hsu et al., 2003; Yin et al., 2007; Zhu et al., 2007; Jeong et al., 2008);刺激小鼠脾臟細胞活化產生 IL-1、
IL-6、IL-12、IFN-γ、TNF-α、GM-CSF、G-CSF 和 M-CSF,而大量誘導產生 IFN-γ 被認為是NK 細胞活化的主因 (Wang et al., 2002; Chen et al., 2004)。
11.4 活化 T 細胞
T 細胞或是稱 T 淋巴球在細胞介導免疫反應(cell-mediated immunity)扮演
重要的功能,在體內或體外的模式中可以發現靈芝多醣體能活化 T 細胞。以 γ
射線照射小鼠使脾臟 T 細胞受損,發現靈芝多醣體對受損 T 細胞的修復效果比
雲芝多醣體PSK 的效果好 (Chen et al., 1995)。此外,在 PS-G(25–100 mg/ml)
處理T 細胞能促進 IFN-γ 的產生 (Wang et al., 1997)。靈芝多醣體也能增強 T 細 胞增生和活化,使樹突細胞成熟,促使走向Th1 反應 (Bao et al., 2002b; Hsiao et al., 2004; Lin et al., 2005; Lin et al., 2006b; Zhu et al., 2007)。對於服用靈芝多醣體 罹癌患者,12 週後發現 CD56 細胞有顯著性增加,而 CD3、CD4 和 CD8 細胞只 些微增加 (Gao et al., 2003)。
11.5 活化 B 細胞
赤芝多醣體能刺激小鼠脾臟 B 細胞增生,增加 3-4 倍的 B 細胞,B 細胞本身 會變大,細胞表面的 CD71 和 CD25,增加免疫球蛋白的分泌和細胞內 PKC 與 PKC-γ 的表現 (Bao et al., 2002a; Zhang et al., 2002b; Shao et al., 2004);另外也發 現F3 多醣體能誘導調控漿細胞分化的轉錄因子 Blimp-1 表現,刺激小鼠 B 細胞 活化,分化成能分泌IgM 的漿細胞 (Lin et al., 2006a)。
12. 抗腫瘤活性
早在1968 年已有報告指出多孔菌科菇類及食用菇類子實體之熱水萃取物,
在接種 Sarcoma 180 的小鼠都有明顯的抗腫瘤效果,而對於靈芝抗腫瘤之科學 研究則在1971 年開始 (Ikekawa et al., 1968; Sasaki et al., 1971)。後續的研究證實 多醣體為其主要活性成分之ㄧ,其中 β-glucan 為有效之結構 (Borchers et al., 1999)。多醣體的抗癌機制並非直接對癌細胞有細胞毒性 (cytotoxicity) 的作用,
而是通過強化宿主的免疫調節功能,抑制腫瘤細胞的增殖或殺滅。抗腫瘤多醣體 可歸類為一種生物反應修飾劑 (biological response modifier, BRM ) (Wasser and Weis, 1999)。目前已有多種菇類多醣體進行抗癌之臨床試驗,如 lentinan (Lentinula edodes) 、 schizophyllan (Schizophyllum commune) 、 PSK (polysaccharide-K, Krestin)、PSP (polysaccharopeptide)(Trametes versicolor) 及 Grifron-D (Grifola frondosa) (Wasser, 2002; Ferreira et al., 2010)。以下分別以細胞模式的體外試驗與 小鼠的體內模式敘述靈芝多醣體的過去研究:
12.1 細胞體外模式
PS-G 處理人類血液單核球細胞的培養液能抑制人類血癌 U937 細胞生長,
此外也可直接抑制Sarcoma 180 細胞的增生 (Lieu et al., 1992)。赤芝孢子多醣體 對小鼠肝癌細胞、Sarcoma 180 細胞和 reticulocyte sarcoma L-II 細胞也有抑制生 長的效果 (Liu et al., 2002; Wang et al., 2009b);對人類結腸癌細胞株(SW480、
HCT116 和 HT29)、血癌細胞、子宮頸癌細胞株(CaSki 和 SiHa)、乳癌細胞、肝 癌細胞、淋巴瘤及多種骨隨瘤細胞都有抑制活性 (Gao et al., 2005a; Muller et al., 2006; Xie et al., 2006)。
比較赤芝、中國紫芝和松杉靈芝水萃物對人類乳癌 MCF-7 和 MDA-MB-231 細胞株的抑制效果,發現松杉靈芝菌柄和野生中國紫芝子實體之多醣體對兩株乳
癌細胞的抑制效果是最好的 (Yue et al., 2006)。
12.2 動物模式
小鼠皮下接種 Sarcoma 180 細胞模式是最常使用的動物模式,以腹腔注射 (Miyazaki and Nishijima, 1981; Sone et al., 1985; Maruyama et al., 1989; Wang et al., 1993; Zhang et al., 1994; Gao et al., 2005a; Bukhman et al., 2007; Pang et al., 2007;
Jeong et al., 2008; Yue et al., 2008; Wang et al., 2009b)或餵食 (Maruyama et al., 1989)靈芝多醣體探討其抗腫瘤活性,其他使用的腫瘤模式還有艾氏腹水瘤 (Ehrlich ascites carcinoma, EAC) (Zhang et al., 2000; Pang et al., 2007)、肺癌細胞 (Cao and Lin, 2004; Nonaka et al., 2008)、肝癌細胞 (Pang et al., 2007)、乳腺腫瘤 細胞 (Nonaka et al., 2008)、血癌細胞 (Chang et al., 2009)和前列腺癌細胞 (Evans et al., 2009),而臨床上也用於肺癌患者的治療 (Gao et al., 2005b)。各種動物腫瘤 模式均顯示靈芝多醣體具有抗腫瘤活性。
四、免疫與腫瘤
免 疫 系 統 是 生 體 防 衛 病 原 菌 入 侵 最 有 效 的 武 器 , 可分成先天性免疫
(innate immunity)與後天性免疫(adaptive immunity),圖 1-3 是參與免疫系統 相關的免疫細胞:
1. 先天性免疫
先 天 性 免 疫 屬 於 非 特 異 性 免 疫 , 為 生 體 第 一 道 防 線 , 透 過 吞 噬 細 胞
(phagocytes)、樹突細胞和NK 細胞等抵禦外來病源體的入侵。先天性免疫之細 胞需經由物種特異性的受體以辨識同源抗原,為達到辨識所有的可能抗原,發展
圖1-3. 先天性免疫反應與後天性免疫反應相關之免疫細胞
Figure 1-3. The innate and adaptive immune response (Dranoff, 2004).
出一套特殊的免疫辨識系統 (Medzhitov and Janeway, 1997)。吞噬細胞包括了嗜 中性白血球、單核球和巨噬細胞。當外來病原菌或微生物侵入淋巴、骨髓或是血 液時,吞噬細胞會受侵入者引發的趨化作用,大量集中到感染發炎部位的附近,
透過吞噬作用殺滅微生物;或是活化產生細胞毒殺物質,像是過氧化氫、一氧化 氮、防禦素(defensin)或 TNF-α 等。NK 細胞則透過與標靶細胞緊密結合,分 泌有毒物質毒殺細胞。
而發炎反應(inflammatory response)會使受傷或微生物入侵的部位發生小 動脈和微血管擴張,增加微血管管壁的通透性,使體液和細胞由充血的血管進入 組織,開始細胞內的消化反應。屬於一種保護反應,讓生體自然啟動修復機制 (Ferrero-Miliani et al., 1997)。嗜中性球和單核球細胞通常出現在急性發炎反應,
嗜中性球受刺激後會產生 prostaglandins 和 thromboxanes;單核球細胞則產生 arachidonic acid 氧化產物,這些免疫細胞在發炎反應均扮演重要的角色(Davies and Allison, 1976; Goldstein et al., 1977)。
2. 後天性免疫
後天性免疫屬於特異性免疫反應,除了非特異性的免疫作用外,免疫系統對 於入侵外來病源視為抗原,對不同的抗原經由隨機基因重組產生專一性的抗體,
專一性的分辨抗原的不同,具有免疫記憶的能力,當再次遇到相同的抗原時,即 可引發更強烈的免疫反應。
特 異 性 免 疫 細 胞 主 要 有 淋 巴 細 胞 ( lymphocytes ) 和 抗 原 呈 現 細 胞
(antigen-present cells, APC)。淋巴細胞又可分成 B 細胞和 T 細胞,B 細胞與抗
原結合活化後分化成記憶 B 細胞和漿細胞。漿細胞可分泌大量的抗體,為體液
免疫(humoral immunity)的主要作用物質。而 T 細胞可分成輔助型 CD4 細胞和 毒殺型CD8 細胞。CD4 細胞主要分泌細胞激素和參與體液免疫與細胞介導免疫 反應;CD8 細胞主要負責清除有病原感染的細胞。
T 細胞中的 CD4 細胞能分泌 IL-4 和 IL-5 細胞激素刺激 B 細胞,使 B 細胞 成熟為漿細胞產生抗體;產生IFN-γ 刺激樹突細胞、巨噬細胞和 B 細胞等抗原呈 現細胞上的MHC II 抗原,CD4 也會直接辨識表現 MHC II 的腫瘤細胞;傳遞訊 息給CD8 清除病原,分泌 IL-2 活化 CD8 細胞的增生及其記憶性(Bennett et al., 1997; Shedlock and Shen, 2003; Gutcher and Becher, 2007; Kennedy and Celis, 2008),而這些反應會被 Treg 抑制(圖 1-4)。
CD8 細胞體內很重要的淋巴細胞之一,主要負責清除細胞內有病原的感染 細胞,或者是清除一些變異的腫瘤細胞。CD8 細胞的活化需要 CD4 細胞提供訊 息給抗原呈現細胞,再傳遞訊息給CD8 細胞;或是 CD4 細胞直接將訊息傳遞給 CD8 細胞(Shedlock and Shen, 2003)。
樹突細胞(dendritic cells)活化過程,MHC II-peptide complexs 會累積在樹 突細胞的表面,將抗原穩定的呈現給CD4 細胞,活化 CD4 細胞,持續產生抗腫 瘤的細胞毒殺T 淋巴球的記憶反應 (Robson et al., 2010)。
抗原呈現細胞包含了樹突細胞、巨噬細胞和 B 淋巴細胞。藉由與抗原結合
進行成熟過程,並將訊息呈現給T 細胞辨識。樹突細胞和巨噬細胞能夠察覺環境
中的入侵微生物,將病原吞入並切成碎片,再將碎片的資訊呈現給T 細胞,組成
免疫示警系統,可說是啟動後天性免疫反應的關鍵,連接了先天性免和後天性免 疫反應(Banchereau and Steinman, 1998; Zanoni et al., 2005)。
圖1-4. CD4 細胞在腫瘤微環境中的複雜角色
Figure 1-4. The complex role of the T helper cell in the tumor microenvironment (Knutson and Disis, 2005).
3. 免疫辨識系統
免疫系統為了有效辨識入侵的微生物或其他抗原,免疫細胞會將大多數微生 物具有高度保守性之結構視為辨識目標,此保守性結構稱為病原相關分子模式
(pathogen-associated molecular patterns, PAMPs),而免疫細胞辨識這些 PAMPs 則是透過模式辨識受體(pattern recognition receptors, PRRs)。
PRRs 可分成細胞膜上的和細胞質內的。細胞膜上的 PRRs 如 Toll-like receptors(TLRs)、甘露糖受體和Dectin-1 等;細胞質內的 PRRs 如 NOD-like 受 體。真菌能辨識之 PRRs 整理如表 1-5;而不同免疫細胞表面所表現的 PRRs 也 有所不同(圖1-5)。
3.1 TLRs
TLRs 因其與果蠅之 Toll 家族蛋白有高度同源性而得名。在人類與小鼠中,
已有11 種 TLRs 被分類鑑定。基本上,TLR1、6 和 9 廣泛存在於各類免疫細胞,
TLR2、4 和 5 分布於除了 T 細胞、B 細胞和 NK 細胞以外的免疫細胞,TLR3 只 在樹突細胞上發現,TLR7 和 TLR8 分布於周邊血液淋巴球細胞 (Nishiya and DeFranco, 2004)。
3.2 TLR4
LPS 是格蘭氏陰性細胞壁的主要成分,也是 TLR4 受體專一性的配體,以 LPS 刺激巨噬細胞時,能同時活化 MyD88 依賴(MyD88-dependent pathway)以 及TRIF 依賴(TRIF-dependent pathway)兩種訊息傳遞途徑,活化 NF-κB 和 AP-1 轉錄因子,產生TNF-α、IL-1、IL-6…等發炎細胞激素 (Takeuchi and Akira, 2001)。
3.3 Dectin-1 (Dendritic-cell-associated C-type lectin-1)
Dectin-1 為一個表現在骨髓細胞上的穿膜受體,具有胞內單一、非典型
表1-5. 真菌辨識有關之模式辨識受體
Table 1-5. Pattern-recognition receptors involved in fungal recognition (Brown, 2006).
*Role as PRR not confirmed. ‡Human studies. §P. Taylor, M. Botto, K. Haynes, S.
Gordon and G.D.B., unpublished observations.
C3, complement component 3; DC-SIGN, dendritic-cell-specific ICAM3-grabbing non-integrin; ND, not determined; SP, surfactant protein; TLR, Toll-like receptor.
圖1-5. Candida albicans 辨識之免疫細胞及其模式辨識受體
Figure 1-5. Cell populations and pattern-recognition receptors involved in Candida albicans recognition (Netea et al., 2008).
The differential expression of PRRs by these cell types is shown. CR3, complement receptor 3; FcγR, Fcγ receptor; MR, mannose receptor; TLR, Toll‑like receptors.
(Brown and Gordon, 2001)。Dectin-1 位於胞內尾端具有一個 tyrosine-based activation-like motif (ITAM)免疫受體,經由依賴 spleen tyrosine kinase (Syk)和 caspase recruitment domain protein 9 (CARD9)途徑能夠傳達各種保護反應,產生 多種細胞激素,甚至會影響CD4 細胞和 B 細胞的作用。在配體的作用下,Dectin-1 能引起氧化爆發、產生發炎細胞激素和吞噬作用等反應 (Brown, 2006; Gringhuis et al., 2009)。
3.4 CR3 (Complement receptor type 3; CD11b/CD18)
CR3 是細胞表面醣蛋白的複合體,屬於 β2 integrin 家族。CR3 的次單位包含 兩個結合部位,一個為結合β-glucan 之凝集素功能區;另一個區域為結合 C3 斷 片iC3b 之功能區。有研究提出酵母菌之 β-1,3-glucan、β-1,6-glucan 以及分離自大 麥之水溶性多醣體β-1,3-glucan 和 β-1,4-glucan 能與 CR3 結合後,刺激 CR3 受體 使白血球細胞毒殺附著 iC3b 之腫瘤細胞 (Vetvicka et al., 1996; Hong et al., 2004)。
4. 訊息傳導路徑(signaling transduction pathway)
4.1 絲裂原活化蛋白激酶(mitogen-activated protein kinase, MAPKs)
MAPKs 在細胞訊息傳導路徑扮演極重要角色,MAPKs 訊息傳導已知有三條 路徑,包括extracellular signal-regulated kinase (ERK1/2)、c-Jun N-terminal kinase (JNK)及 p38-MAPK(p38)(圖1-6)。活化的 MAPK 是透過磷酸化作用而進行訊 息傳導,進而調控細胞增生、細胞分化、壓力反應、細胞凋亡及發炎反應等(Hsu et al., 2004; Liu et al., 2007)。
4.2 NF-κB(nuclear factor κB)
圖1-6. 哺乳動物三種絲裂原活化蛋白激酶訊息傳導途徑簡圖
Figure 1-6. Schematic diagram of the three mammalian mitogen-activated protein kinase signalling pathways (Liu et al., 2007).
胞質的NF-κB 與其抑制分子 I-κB 結合而呈未活化狀態;當細胞受到各種刺激後,
I-κB 受到磷酸化及 ubiquitination,並緊接著被蛋白酶體(proteasome)降解,此 時NF-κB 被活化並移位至細胞核內,接著誘導相關的基因表現(圖 1-7)(Karin and Lin, 2002; Beinke and Ley, 2004)。當細胞受到細胞激素、細菌或病毒感染或是過
氧化物的刺激會藉由 NF-κB 活化而影響細胞的生長和分化,誘發發炎反應。
NF-κB 的調控於先天性免疫反應(如發炎反應)及後天性免疫反應上,具有重要 的功能。
5. 細胞激素
細胞激素(cytokine)是一群負責傳遞各細胞間訊息的分子,由特異性的免 疫細胞所分泌,主要成分為蛋白質、胜肽或醣蛋白,如interferons、interleukins、
TNF-α…等(圖 1-4)。TNF-α 是很重要的發炎因子,在感染或是受傷早期的反應
扮演重要的角色,少量 TNF-α 的釋放在免疫反應中具有正向的功能,但是過多
及持續的釋放則非正常生理現象 (Beutler, 1995)。
6. 腫瘤的治療
目前的腫瘤治療方法主要有四類:(一)手術療法,直接將腫瘤及周圍可能 擴散的地方切除,對擴散性腫瘤效果不佳;(二)放射線療法,以放射線的能量 直接照射腫瘤細胞,但是腫瘤附近的正常細胞也會被殺滅;(三)化學療法,全 身性療法,同時也會殺滅正常細胞,常搭配手術或放射線治療之輔助性療法,副
作用極大;(四)其他療法:生物性療法是指增強自體免疫系統專一性清除腫瘤
細胞以達到抗腫瘤的目的,主要是阻斷腫瘤細胞的分裂複製;或是使用激素類藥
圖1-7. NF-κB的活化路徑
Figure 1-7. Model of the generic NF-κB activation pathway (Beinke and Ley, 2004).
aromatase inhibitors、receptor down- regulators…),可以降低一般治療的副作用,
例如使用干擾素、細胞激素、colony-stimulating factors、單株抗體、疫苗、基因 治療或非專一性的免疫調節劑;血管增生抑制劑治療是防止新血管的形成使腫瘤 細胞無法得到充分的養分增殖;骨髓移植或週邊血液幹細胞移殖,將癌症患者的 病變骨髓替換成健康的骨髓,因為這些免疫細胞,特別是淋巴細胞對淋巴癌或血 癌病患的治療成效良好(Maury et al., 2010);過高熱治療(hyperthermia),讓身體 組織暴露在113 °F高溫以上,以殺死腫瘤細胞;雷射光的應用,以高能量細小的 光 束 雷 射 精 準 的 去 除 腫 瘤 或 未 成 癌 (precancerous ) 生 長 ; 光 力 學 治 療
(photodynamic therapy, PDT),光力學治療是結合光敏感藥物及特殊的光,在細 胞附近產生氧以殺死細胞;標靶癌症治療(targeted cancer therapy)則利用藥物 或 其 他 可 以 阻 礙 癌 細 胞 生 長 或 擴 散 的 物 質 , 防 止 腫 瘤 細 胞 的 生 長 增 生
(http://www.cancer.gov/cancertopics/treatment/types-of-treatment)。
7. 生物性抗癌療法
生物性療法是屬於比較新的治療方法,如特異性抑制劑(targeted therapy)
治療的概念,將原本作用於生體所有的細胞的藥物改成只專一性攻擊腫瘤細胞,
大 大 降 低 過 去 抗 癌 藥 物 的 副 作 用 , 這 些 藥 物 如selective estrogen receptor modulators、kinase inhinbitors、angiogenesis inhibitors、proteasome inhibitors…等,
FDA已核准上市的產品如Fareston®、Arimidex®、Gleevec®、Zolinza®、Avastin®… 等。
抗體治療是將腫瘤細胞視為抗體的標靶細胞,利用抗體封鎖細胞接收生長所
需的信號,或切斷腫瘤細胞需接收信號之相關受體,甚至直接與腫瘤細胞表面的 蛋白質作用,進行標識功能,再由免疫系統加以專一性毒殺,導致腫瘤細胞死亡 以達到治療的效果 (Wolchok and Saenger, 2008; Nelson et al., 2010)。其中又以使 用人類單株抗體為發展趨勢,人類單株抗體不像噬菌體或小鼠生產的抗體,在人 體中不會快速被血液代謝、產生全身的發炎反應或是產生人類抗小鼠抗體等缺 點。目前為止有7種人類單株抗體通過FDA審核,另有3種正在審查中,更有7種 正進行phase III的試驗,81種在phase I或phase II (Nelson et al., 2010)。將自體腫瘤 細胞修飾後製成疫苗、或是轉殖表現細胞激素之基因等,刺激相關細胞分泌細胞 激素,進而抑制腫瘤之生長(Abe et al., 1995; Dranoff, 2004)。
腫瘤治療的策略上大致都是直接切除腫瘤組織或細胞、切斷腫瘤細胞生長所 需的訊息或養份,或是活化生體自身的免疫機制,最終導致腫瘤細胞死亡為基 礎。傳統上,對於腫瘤的處理策略採用化學藥物治療或放射線治療,但是這類的 治療方式不具特異性,無法避免對正常的細胞及組織造成傷害,對身體的副作用 也較廣泛。近年來對於癌細胞分子生物醫學的瞭解,研究學者已積極轉向以細胞 表面抗原為標地的免疫療法,或者利用各種細胞訊息傳遞途徑之作用為目標的治 療方式,另一方面也從天然藥材或傳統使用藥草類去篩選藥效佳、副作用低的有 效抗腫瘤成分。其中,多醣體是植物、動物及微生物來源的天然成分,以往對多 醣體的認識比較偏向能源儲存物質和結構材料上。隨著研究技術的進步,多醣體 的純化、組成分析、分子構形和結構解析都有長足的進展,已知多醣體能參與了 細胞間的辨識、訊息傳導、活化免疫系統和細胞的分化等生物活性,近年來受到 研究人員焦點關注(Schepetkin and Quinn, 2006; Chan et al., 2009; Goodridge et al.,
2009; Wang et al., 2009a; Avci and Kasper, 2010),多醣體在生物醫藥領域新穎功能 的開發更是目前熱門的研究題目。
五、菇類之抗腫瘤成分
自古以來,菇類一直被人作為食用及藥用的材料,而菇類的藥用活性成分及 運用也陸續被研究,活性之成分更能從子實體、液態培養菌絲體或是液態培養液 中純化(Wasser, 2002)。以分子量大小來區分,這些活性成分又可分成小分子和大 分子化合物兩大類(圖1-8)。其中多醣體和三萜類是其中較具代表性的藥用活性 成分,不同來源多醣體的抗腫瘤和免疫調節功能已是具開發潛力的現代藥物。
1. 低分子量化合物
低分子量化合物主要是菇類的二級代謝產物,像是酚類化合物、聚酮類
(polyketides)、三萜類和固醇類等,其中具有抗腫瘤效果的化合物整理如表 1-6(Ferreira et al., 2010)。本研究室從台灣紫芝醱酵液之乙酸乙酯萃取出的小分子 便發現具有抑制腫瘤細胞株之生長(陳,2006),其有效成分之實際化合物則需進 一步鑑定;靈芝類純化的cerebrosides可抑制真核細胞DNA聚合酶(Zaidman et al., 2005; Paterson, 2006);洋菇和香菇中的caffeic acid phenyl ester能抑制專一性抑制 NF-κB結合在DNA上。這些小分子容易穿過細胞膜,進入腫瘤細胞中,對特定的 分子目標作用,具有殺滅腫瘤細胞的活性(Zaidman et al., 2005; Petrova et al., 2008;
Wasser, 2011)。
2. 高分子量化合物