國立臺灣大學生物資源暨農學院森林環境暨資源學系 碩士論文
School of Forestry and Resource Conservation College of BioResources and Agriculture
National Taiwan University Master Thesis
東亞與大洋洲構樹親緣地理學研究
Phylogeography of East Asian and Oceanian Paper Mulberry
劉筱蕾 Hsiao-Lei Liu
指導教授:鍾國芳 博士 Advisor: Kuo-Fang Chung, Ph.D.
中華民國 101 年 8 月
August, 2012
I
II
誌謝
首先誠摯的感謝指導教授鍾國芳博士,以及我的口是委員陳韋仁老師、江友中 老師、以及胡正恆老師,老師們悉心的教導使我得以稍稍理解生物地理學領域的 深奧與有趣之處,不時的討論並指點我正確的方向,使我在這些年中獲益匪淺 (也恭喜老師終於丟掉一個麻煩)。
本論文的完成另外亦得感謝教育部世界南島研究計畫辦公室的大力協助讓 我們得已前往越南、印尼、夏威夷進行採樣,及法國國家植物園Mascarin 常駐 中心Jean Hivert 先生、中研院生多中心張家豪學長與中村剛博士、河野淑子博士 在採樣上的支援。因為有你們的幫忙,使得本論文能夠更完整而嚴謹。
三年裡的日子,實驗室裡共同的生活點滴,學術上的討論、言不及義的閒扯、
每次出隊就可以減肥的生多減肥班、趕作業的革命情感...,這三年我好像多懂 了一點東西,又好像更知道自己得不足之處。感謝眾位學長姐、同學、學弟妹的 共同砥礪,你們的陪伴讓三年的研究生活變得絢麗多彩。
感謝小古、小舟、旨价、忠義、永信學長、姿麟、巽安、群智學姐們不厭其 煩的指出我研究中的缺失,且總能在我迷惘時為我解惑,也感謝家瑜、珮儀、雅 青、阿德的幫忙,恭喜我們順利走過(或即將走過)這些日子。實驗室的儀器們當 然也不能忘記 (不要壞掉喔!!),小二、小三跟 Opal、Triple 的搞笑及老陳的幫忙 我銘感在心,在背後的默默支持更是我前進的動力。
最後,謹以此文獻給我摯愛的雙親。
III
摘要
構樹 (Broussonetia papyrifera) 是南島語族重要的物質文化─樹皮布的原料。
由於構樹在島嶼東南亞與大洋洲無法結果,許多早期研究顯示構樹可能是自東亞 由南島語族人傳播到大洋洲地區。因此,我們希望藉由研究構樹的親緣地理學來 檢測其與史前南島語族遷徙的的假說,並探討造成構樹親緣地理結構的可能原因。
本研究使用核醣體基因 (rDNA) 的內轉錄區間 (internal transcribe spacer, ITS) 與葉綠體 ndhF-rpl32 內含子等兩個片段來重建與推論構樹親緣地理及族群遺傳 結構,研究材料包括採集自台灣、日本、中國南方各省、越南、印尼、菲律賓、
非洲留尼旺島以及大洋洲諸島等共516 株樣本。研究結果發現,越南與印尼的族 群內缺乏遺傳變異,而台灣與中國則涵蓋大多數的遺傳變異。親緣關係重建的結 果兩個片段都呈現星狀結構,顯示東亞以及大洋洲族群大體上呈現族群擴張模式,
與Tajima’s D test 與 nested clade phylogenetic analyses (NCPA) 的結果一致。此外,
而族群親緣網狀圖亦顯示目前分布於夏威夷的構樹族群是經過至少兩次的傳播 事件而來,一次來自台灣南部另一次來自中國廣東、廣西;大洋洲其他島嶼的基 因單型更顯示一個起源於台灣南部的基因單型的分布涵蓋印尼與整個大洋洲,符 合南島語族傳播的途徑。這些結果無法排除構樹是人為自東亞傳播到大洋洲的推 論,更進一步指出台灣較可能是大洋洲構樹的起源,部分符合“Express Train Hypothesis”中對南島文化起源地的描述。
關鍵字:人為傳播、樹皮布、共生者、南島語族、大洋洲
IV
Abstract
Bark cloth (tapa) is one of the most important material cultures of Austronesian peoples. In the Pacific, tapa is made primarily of paper mulberry (Broussonetia papyrifera). Because paper mulberry cannot generate seeds in insular Southeast Asia and Oceania, earlier studies suggested that paper mulberries could have been brought to the Oceania from East Asia along with the expansion of the Austronesian peoples to Oceania. Therefore, we aim to test Austronesian colonization hypothesis through phylogeographic study of paper mulberry and to elucidate how human mediated dispersals have shaped population structure of the species. A total of 516 specimens of paper mulberries were collected from Taiwan, southern China, Vietnam, Indonesia, Philippines, Reunion and the Oceania. Analyses of DNA sequences of both ITS and cpDNA ndhF-rpL32 indicate that populations in Indonesia, Near Oceania and Remote Oceania are devoid of genetic variation, while Taiwan and China harbor the highest genetic diversity. Haplotype networks reconstructed using both DNA markers all show star-like network structure, suggesting a scenario of population range expansion.
This inference is further supported by nested clade phylogenetic analyses (NCPA) that conclude a continuous rang e expansion or long distance colonization for the
Oceanian samples. In the networks, all haplotypes carried by Hawaiian samples are identical or linked directly to haplotypes from southern Taiwan and China separately, suggesting the Hawaiian haplotypes are derived from those two regions and at least 2 colonization events had occurred to shape the population structure there. In the ndhF-rpl32 network, about half of the samples from Remote Oceania carry a haplotype found in southern Taiwan and Sulawesi, Indonesia. Because paper mulberries are planted in Sulawesi, suggesting strongly that the southern Taiwan paper mulberries should have been the direct ancestor of the Remote Oceanic
V
populations. These results are consistent with the idea that paper mulberry in Oceania have been transported from East Asia, especially Taiwan, partially supporting the
“Express Train Hypothesis” that hypothesized Taiwan as an important stepping stone for the Austronesian peoples expansion.
Key words: human-mediated dispersal, tapa, bark-cloth, commensal species, Austronesian, Oceania
VI
目錄
摘要 ... 1
Abstract ... IV 表目錄 ... VIII 圖目錄 ... IX 1 前言 ... 1
1.1 構樹與樹皮布文化 ... 1
1.2 南島語族遷徙理論 ... 3
1.3 利用共生者回溯南島語族的遷徙歷史 ... 6
1.4 利用構樹來推測南島語族文化的起源與傳播 ... 9
2 材料與方法 ... 11
2.1 分類群取樣 ... 11
2.2 DNA 萃取、PCR 擴增及定序 ... 12
2.3 資料分析 ... 14
3 結果 ... 18
3.1 序列變異分析 ... 18
3.2 族群遺傳變異分析 ... 18
3.3 親緣地理關係分析 ... 20
3.4 遺傳與空間結構分析 ... 22
VII
4 討論 ... 24
4.1 東亞不同地理區構樹族群結構的解釋與推論 ... 24
4.2 島嶼東南亞與大洋洲構樹族群遺傳多樣性的分布 ... 29
4.3 人為傳播的探討 ... 33
4.4 與快車說的比較 ... 34
5 結論 ... 37
6 參考文獻 ... 38
VIII
表目錄
表 1、PCR 引子表 ... 49 表 2、PCR 溫度循環表 ... 49 表 3、構樹各地區族群個體數 (N) 、ITS 片段核苷酸歧異度 (nucleotide diversity:
π)、基因歧異度 (haplotype diversity: h )以及族群個體中性檢測 (Tajima’s D test) ... 50 表 4、構樹各地區族群個體數 (N) 、葉綠體 ndhF-rpl32 片段核苷酸歧異度 (nucleotide diversity: π)、基因歧異度 (haplotype diversity: h )以及族群個體中性 檢測 (Tajima’s D test) ... 51 表 5 東亞與大洋洲構樹 ITS 片段兩兩族群間遺傳分化與基因交流指... 52 表 6 東亞與大洋洲構樹葉綠體 ndhF-rpl32 片段兩兩族群間遺傳分化與基因交流 指數... 53 表 7 構樹 ITS 片段的 Nested Clade Analysis (NCA)的推論順序及結果表 ... 54 表 8 構樹葉綠體 ndhF-rpl32 片段的 Nested Clade Analysis (NCA)的推論順序及結 果表... 55 表 9 構樹 ITS 片段基因型在東亞及大洋洲各地區間的分布表... 58 表 10 構樹 ndhF-rpl32 片段基因型在東亞及大洋洲各地區間的分布表 ... 59
IX
圖目錄
圖 1 大洋洲地圖... 61
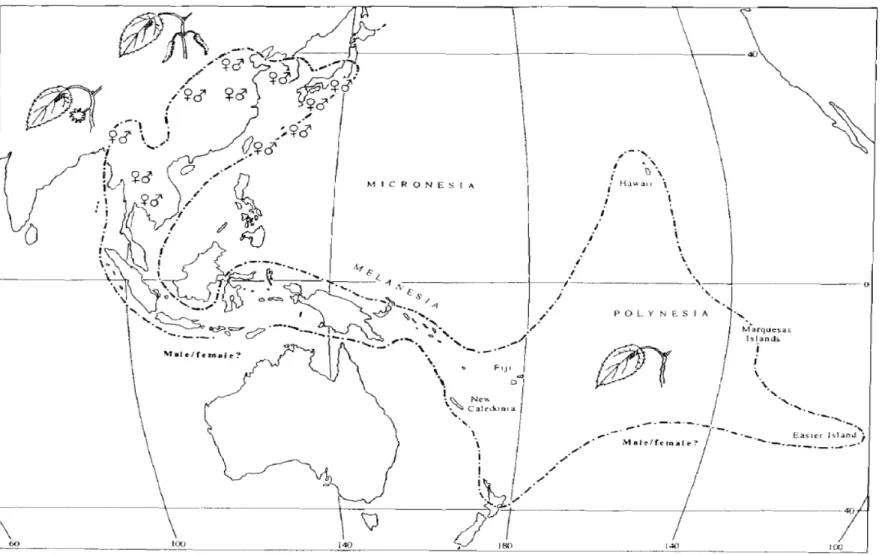
圖 2 Matthew (1996) 所提出的構樹分布圖 ... 62
圖 3 構樹 (Broussonetia papyrifera) 樣本的採樣地點與樣本數圖 ... 63
圖 4 構樹 ITS 片段之親緣關係網狀圖 (haplotype network)與 Nested Clade Analysis 網狀關係圖... 64
圖 5 構樹 ndhF-rpl32 片段之親緣關係網狀圖 (haplotype network) ... 65
圖 6 構樹 ndhF-rpl32 片段之 Nested Clade Analysis 網狀關係圖 ... 66
圖7 構樹 ITS 片段遺傳障礙顯示圖... 67
圖 8 構樹 ndhF-rpl32 片段遺傳障礙顯示圖 ... 68
1
1 前言
1.1 構樹與樹皮布文化
樹皮布 (tapa, bark cloth) 是一種以植物的樹皮為原料,經過拍打技術加工製成 的無紡織布(Matthews, 1996)。製做樹皮布首先要自樹幹剝下樹皮,再利用工具敲 打使纖維軟化,多塊處理過的樹皮經由敲打加以接合,形成我們所看到的樹皮布 (Howard, 2006)。專門敲打樹皮纖維使其軟化的工具─稱為樹皮布打棒或石拍(stone bitter) ,最早的樹皮布石拍出土於西元前 7000 年的華南珠江三角洲,提供了有關 樹皮布文化起源明確的考古證據 (Cameron, 2006, 2008)。石拍也於較晚的年代,在 台灣與中南半島新石器時代的考古遺跡中發現 (Cameron, 2006)。根據新石器時代 考古學的發現,多數研究者認為樹皮布製作的技術是遠在漢族文明入侵中國南部 之前,由南島語族人在發展的結果 (Cameron, 2006; Howard, 2006; Cameron, 2008)。
除此之外,包括東南亞、大洋洲、美洲以及非洲部分地區都有樹皮布製作的 證據 (鄧聰, 1997; Kirch, 2002; Kirch, 2010)。其中東南亞及大洋洲地區由南島語族 發展的樹皮布工藝最為精緻。所謂的南島語族指的是分布在大洋洲、東南亞、台 灣及馬達加斯加,使用南島語系語言的族群,南島語族包含了約1,200 種語言,是 世界現今唯一只分布在島嶼上的語系 (Bellwood, Fox, and Tryon, 1995)。而樹皮布 被認為是南島語族最具代表性的物質文明之一 (Howard, 2006; Cameron, 2008)。雖 然目前在東南亞及大洋洲已經沒有實質的生活需求,然而樹皮布依舊深植在南島 語族人的文化傳統中。樹皮布所製作的樹皮衣在波里尼西亞的高階社會有極大的 象徵意義 (Cameron, 2008),而且直到現在在北蘇拉威西、東加、薩摩亞、斐濟及 夏威夷都為了儀式而持續製作樹皮衣 (Neich and Pendergrast, 1997; Maxwell, 2003)。
值得一提的是,由於民族意識的復興,許多台灣的部落也重新開始注重樹皮布製
2
作這個傳統 (張至善, 2012)。
在東南亞及大洋洲地區,樹皮布的材料主要由桑科的麵包樹屬 (Artocarpus)、
榕屬 (Ficus) 等材料製作,但其中最主要者還是構樹 (Broussonetia papyrifera (L.) L'Hér. ex Vent.) 製成 (Matthews, 1996; Howard, 2006)。構樹為桑科(Moraceae)植 物,俗名鹿仔樹,英文名稱為paper mulberry,普遍自生於台灣各地海平面以上至 中海拔廢耕地、道路兩旁、果園和森林邊緣。雌雄異株,雄花為圓柱狀的葇荑花 序,長4-8 cm;雌花則是呈球形的頭狀花序,花絲為絲狀,子房有肉質柄,多花 聚合果,球形,直徑長約2 cm,未成熟時為綠色,成熟後迸開而呈橘紅色 (Liao, 1996;
Zhou, 2003)。在世界其他地區已經廣為馴化甚至成為入侵植物 (Florece and Colladilla, 2008)。在台灣,構樹是全島海拔 2000 公尺以下最廣泛分布的樹種之一,
能適應廣泛的環境條件 (陳忠義 and 廖天賜, 2007)。構樹也常見於中國大陸黃河 以南,是造紙及醫藥用途的重要經濟樹種 (Zheng et al., 2002)。構樹也是東南亞最 早為人栽種的植物之一 (Matthews, 1996)。由於構樹是作為製作樹皮布的主要材料,
所以也經由南島語族人傳播廣泛分布在太平洋各島 (Matthews, 1996; Prebble, 2008;
Seelenfreund et al., 2010)。許多證據顯示這個樹種經由早期的太平洋島嶼殖民者由 原生地帶到東南亞,再漸次傳到大洋洲 (Matthews, 1996; Seelenfreund et al., 2010)。
除了構樹的植物體及種子並無利於海漂的構造外,也有學者指出,在島嶼東南亞 (Island Southeast Asia) 及大洋洲的構樹不曾發現有性繁殖(Matthews, 1996)。在太平 洋地區,幾乎沒有關於構樹的開花或種子生產紀錄,而且雌雄異株的構樹在該地 區的繁殖完全依賴無性繁殖,像是扦插繁殖,在沒有人為照顧下,部分遠大洋洲 (Remote Oceania) 島嶼的構樹甚至瀕臨絕種,這也顯示大洋洲地區的構樹的生長與 傳播可能須經由人類協助才能進行 (Matthews, 1996; Seelenfreund et al., 2010)。植 物學家也指出在美洲、太平洋島嶼的構樹並非原生植物,很可能是從中國南部至 中南半島經東南亞傳到大洋洲島嶼 (Matthews, 1996; Seelenfreund et al., 2010)。
3
1.2 南島語族遷徙理論
南島族群往遠大洋洲諸島的遷移是人類擴張的最後一個階段 (Gibbons, 2001;
Matisoo-Smith, 2007; Kirch, 2010)。了解南島語族祖先何時、何地與如何克服廣大 的海水阻隔一直是歷史人類學最引人入勝且持續爭議不休的議題 (Diamond, 1988;
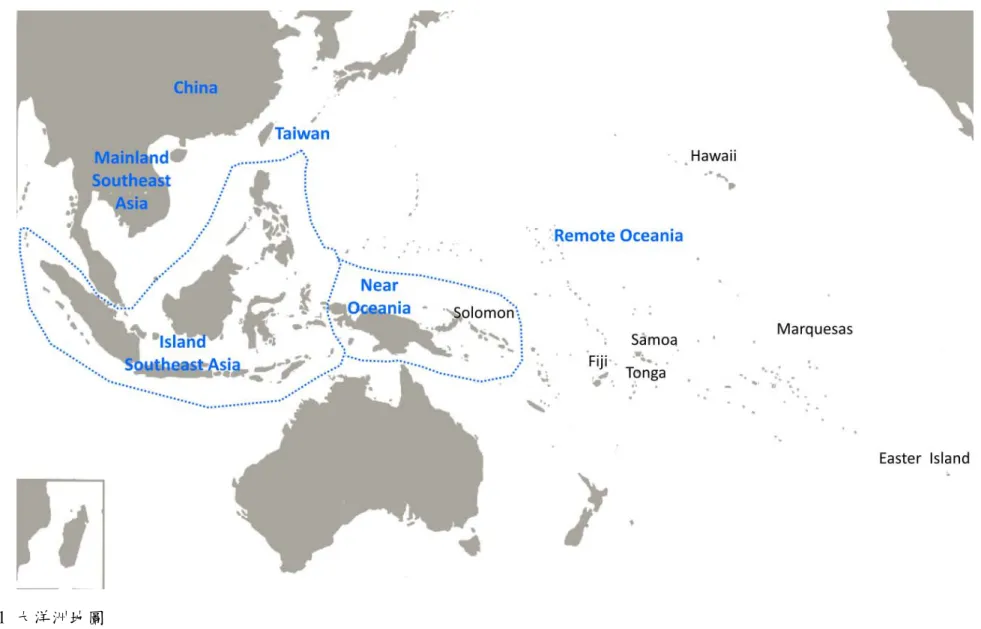
Gibbons, 2001; Matisoo-Smith, 2007; Kirch, 2010)。大洋洲主要由兩個部分組成:太 平洋各島嶼以及澳大利亞大陸。其中太平洋各島嶼根據地理學上的劃分又能分為 密克羅尼西亞 (Micronesia)、美拉尼西亞 (Melanesia) 與波里尼西亞 (Polynesia) 三大島群。然而,這樣的區分並不能代表人類學或是考古學上的分歧,Green (1991) 因而提出近大洋洲 (Near Oceania) 與遠大洋洲 (Remote Oceania) 兩大新地理區 (圖 1)。近大洋洲包含部份美拉尼西亞諸島像是俾斯麥群島與索羅門群島,族群語 言包含南島語和非南島語,而且也是大洋洲中最具生物地理多樣性的地區。而遠 大洋洲中包括所有索羅門群島以東的美拉尼西亞島嶼以及波里尼西亞島群,當地 居民的母語均為南島語 (Kayser, 2010)。根據考古學發現,近大洋洲自 4 萬年前更 新世晚期起就有人類居住,而且也是大洋洲中最具生物地理多樣性的地區。這樣 的區分還有另一層意義─近大洋洲各島間的距離較短,遠大洋洲距離較長,距離 的不同造成人類拓殖的難度差異 (Kayser, 2010)。考古學資料證實遠大洋洲各島直 到距今1500 年之後才開始有人類進入(Kayser et al., 2008)。因此,遠、近大洋洲涵 蓋了太平洋文化史上重大移民擴散事件,適合做為人類學及考古學的分類單位。
這樣獨特的地理環境以及相對較近的人類遷徙歷史使太平洋地區成為了解人 類拓殖及對不同環境適應能力的最佳地點(Gibbons, 2001; Matisoo-Smith, 2007)。目 前關於南島語族傳播有數種不同的假說,其中本研究中較重要的分別是「快車說」
(Express Train To Polynesia Model, ETP)、「慢船說」(Slow-Boat model)。首先 「快 車說」又稱 「出台灣說」 (Diamond, 1988, 1998, 2000),提出台灣可能是現存南島 語族文化的源頭 (Pawley and Ross, 1993)。快車說推測南島語族的祖先由中國華南
4
地區進入台灣,再快速的自台灣移進菲律賓,之後繼續往東至馬來西亞、印尼等 島嶼東南亞 (Island Southeast Asia)地區,向西到達馬達加斯加島,再由近大洋洲 (Near Oceania) 拓殖到遠大洋洲,中途沒有受到當地文化的影響 (Blust, 1988;
Diamond, 1988, 2000; Matisoo-Smith, 2007)。這個假說最初是由考古學證據所支持。
之後的語言學證據也顯示出約1,200 種的南島語經過分析可被分為 10 群,其中 9 群 (共包含 26 種南島語) 只存於台灣 (Pawley and Ross, 1993; Blust, 1996, 1999;
Diamond, 2000)。而第 10 群,也就是馬來-波里尼西亞亞群 (Malayo-Polynesian language group),包含約 1,174 種南島語廣泛分布在除了台灣之外其他所有的南島 語族分布區域 (Blust, 1999; Diamond, 2000)。
根據如此特殊的語言分布,許多研究者提出台灣可能是南島語族原鄉這樣的 想法(Pawley and Ross, 1993; Diamond, 2000)。此外,藉由比較兩種語言之間的變異 程度,可發現台灣島內任兩種南島語之間的差異遠大於台灣之外的南島語所呈現 的變異 (Diamond, 2000)。這種現象暗示南島語族的祖先自原居地(極可能是中國南 部)遷移到台灣之後,曾在台灣停留了一段時間,之後再自台灣向外拓殖,而這樣 的停留造成了台灣南島語的複雜分化,這種推論也被部分考古學證據所支持 (Diamond, 2000)。此外,根據語言分析,在馬來-波里尼西亞亞群內的南島語亦可 發現與非波里尼西亞南島語 (non-Polynesian Austronesian languages) 有明顯的分 別,顯示南島語族再拓殖歷史的第二次「停留」(Diamond, 2000)。根據考古學證據,
第二次停留大約是在西元前1200-200 年之間停留了 1000 年後才往波里尼西亞方向 遷移,完成人類族群的最後一次長距離遷徙 (Diamond, 2000)。除了語言學與考古 學,「快車說」的概念也得到多個人類粒線體DNA 分析結果支持 (Lum and Cann, 2000; Trejaut et al., 2005; Kirch, 2010)。然而,在關於大洋洲緬甸小鼠的研究結果顯 示這些緬甸小鼠的至少三次由島嶼東南亞進入大洋洲,且與台灣的小鼠樣本無關 (Matisoo-Smith and Robins, 2004)。由於緬甸小鼠不會游泳,人類是牠們散布於各 地的唯一媒介。此結果不但突出島嶼東南亞的人類對南島語傳播所扮演的角色,
5
也顯示南島語族傳播過程與快車說的內容可能有所差異。
此外,利用人類 Y 染色體片段所作的遺傳分析數據顯示美拉尼西亞地區的原 住民對南島語族的基因庫帶來極為深遠的影響 (Su et al., 2000; Hunley et al., 2008;
Kayser et al., 2008),支持南島語族遷徙的另一個假說「慢船說」(Oppenheimer and Richards, 2001; Oppenheimer and Richards, 2002)。「慢船說」的內容闡述,波里尼西 亞人的祖先起源於東亞,但曾漸次與美拉尼西亞人的基因混合,然後才向東太平 洋殖民(Kayser et al., 2008)。「慢船說」強調波里尼西亞人的祖先與美拉尼西亞族群 之間的基因交流,這個假說也許可以解釋為何波里尼西亞人發生地中海型貧血的 頻率偏高,由於遠大洋洲幾乎沒有瘧疾,這個基因可能是在波里尼西亞族群祖先 傳播到東太平洋之前,由近大洋洲美拉尼西亞族群傳到波里尼西亞族群祖先身上 (Flint et al., 1986; Kirch, 2010),這個證據指出美拉尼西亞族群與波里尼西亞族群確 實存在基因交流,但卻無法直接證明南島文化的傳播。關於人類Y 染色體多型性 的資料雖支持慢船說,亦將相對極端的假說排除─「俾斯麥群島原居民假說」
(Bismarck Archipelago Indigenous Inhabitants model),主張波里尼西亞人的祖先及文 化是由俾斯麥群島的原住民獨立發展而來 (Matisoo-Smith, 2007)。
最後一個假說─航海廊道說 (Voyaging Corridor Triple I)(Green, 1991) 奠基於 慢船說,但更強調南島語族的前期文化─拉皮塔文化。指出南島文化的起源是由 於「外來文化要素與近大洋洲原有的物質文明整合,而產生全新、獨特的文化要 素」(Matisoo-Smith, 2007)。
6
1.3 親緣地理學的基本概念與應用於了解物種遷徙歷史
不同的遷徙假說及不同學科相互衝突的證據顯示了南島語族文化成型的複雜 歷史。因此,發展一個能結合多個學科所有可用證據的研究方式變得尤其重要 (Kirch, 2002; Hurles et al., 2003; Matisoo-Smith, 2007)。其中一種研究方式是經由研 究太平洋地區共生者物種的遺傳結構 (genetic structure) 與親緣地理學模式
(phylogeographic pattern),藉此作為研究南島語族人遷徙的指標 (Matisoo-Smith, 1994, 2009; Matisoo-Smith and Robins, 2009)。
生物族群的演化受到基因交流及地質事件的影響,在族群內呈現不同程度的 遺傳歧異度以及在族群間呈現不同的遺傳分化。這些演化的過程藉由個體的形態 特徵或遺傳資訊而保留下來。以上資料經過分析,可以做為重建物種親緣地理的 分子標記,以推估造成現今多型性空間分布的可能事件。這就是親緣地理學 (phylogeography) 的基本概念。親緣地理學的重心放在推估過往物種傳播過程與發 生時間,以及了解生物體的遺傳變異在大範圍空間下分布的研究。透過研究自然 族群間的關係與現存的地理分佈,親緣地理學提供了一個整合性概念架構來探討 物種形成和分化的過程。這類的研究經由調查基因流 (gene flow)、遺傳瓶頸 (genetic bottleneck)、群體擴張 (population expansion) 等塑造遺傳變異地理分佈方 式,可以提供我們多方面的資訊,有助於進行生物演化、地球環境變遷、生物育 種、入侵生物及生物傳播方面問題的研究 (Arbogast and Kenagy, 2001)。
而如果因為生物有經濟價值而有人類協助傳播,由於人類的使用偏好影響,
往往會產生族群結構或是遺傳性狀上的特徵,也可做為研究時的參考。像是因強 力的篩選,使得作物快速地達到人類需求的性狀的同時,也強化了瓶頸效應的效 力 (Eyre-Walker et al., 1998)。因此,栽培作物與其祖先種的群體遺傳結構相比,
通常栽培作物的遺傳多樣性更低。以玉米為例,相對於野生玉米,栽培種玉米僅 保存了對偶基因總數的 36% ,同樣地,栽培水稻的遺傳多樣性也較野生稻來得低
7
(Londo et al., 2006)。此外,部分性狀會受到強力人擇的影響。性狀之所以在馴化 時受到選擇,是因為它們符合早期人類的需要,但這些性狀不一定對該物種在天 然環境中的繁衍和擴散有利,所以在沒有人類干預的野生環境中被馴化的作物反 而很難生存 (Clement, 1999)。
目前已知在南島語族人在遠大洋洲四處遷徙殖民時會攜帶一整套的生物包裹 (biological package) 同行,這套包含食用、生活、文化甚至包括疾病的包裹,對航 向物資缺乏的未知島嶼進行殖民的南島語族人至關重要 (Bellwood, 1978;
Diamond, 1998; Kirch, 2002; Matisoo-Smith, 2009)。而被攜帶的生物因為長期與人維 持親近的關係,其生長與傳播往往也要透過人力協助,也因此族群結構也受到影 響,所以被科學家稱為共生者 (commensals, commensal species)。「共生者」的概 念大約是在1994 年,由研究波里尼西亞地區緬甸小鼠的生物人類學家
Matisoo-Smith 首次提出(Matisoo-Smith, 1994; Matisoo-Smith et al., 1998;
Matisoo-Smith and Robins, 2004),當時她正藉由緬甸小鼠來探索南島語族的傳播。
緬甸小鼠原生於中南半島,分布範圍廣布於中南半島及太平洋地區遠及復活節島,
是分布世界第三廣的鼠類。由於緬甸小鼠不會游泳,緬甸小鼠骨骸在太平洋各島 考古遺跡的出現與波里尼西亞人遷徙有莫大的關係 (Matisoo-Smith and Robins, 2004)。
藉由調查緬甸小鼠 (Rattus exulans,波里尼西亞鼠) 的粒腺體 (mitochondria) DNA,Matisoo-Smith 與 Robins 發現 3 群主要的基因單型 (haplotype),而這些基 因單型的地理分布較為符合航海廊道說的推論 (Matisoo-Smith and Robins, 2004)。
此外,島間緬甸小鼠族群間的高度遺傳變化,暗示了太平洋各島的緬甸小鼠族群 可能是自不同的來源多次傳播而來,符合毛利人到紐西蘭的播遷歷史
(Matisoo-Smith, 2007)。
隨著研究的發展,"共生者"的概念被應用在其他南島語族人攜帶的生物上 (Matisoo-Smith and Robins, 2009)。像是親緣地理學研究也顯示一種石龍子 Lipinia
8
noctua 也同時從美拉尼西亞被帶到波里尼西亞,符合快車說的推論 (Austin, 1999)。
關於野豬 (Sus scrofa) 的親緣關係研究,則指出太平洋地區的野豬祖先應該來自中 南半島,而不是台灣或菲律賓 (Larson et al., 2007)。而大溪地的原生蝸牛 (Partula hyaline) 粒線體 DNA 研究發現,這種擁有美麗外殼的蝸牛曾多次被交易或引進澳 洲及庫克群島南部 (Lee et al., 2007)。
共生者研究也被用來調查太平洋地區人類寄生菌的傳播。藉由觀察寄生在人 類消化系統的幽門螺旋桿菌 (Helicobacter pylori) 的基因單型變化,可以發現幽門 螺旋桿菌的DNA,根據地理分布可被明確區分成 hpSahul 與 hspMaori 兩個基因單 型 (Moodley et al., 2009)。hpSahul 的族群分布於新幾內亞與澳洲原住民身上,而 帶有hspMaori 的人主要是台灣原住民及其他使用南島語的族群。在樹形圖中相對 短的分枝顯示帶有hspMaori 的族群經歷了快速的族群擴張,符合快車說的推論 (Moodley et al., 2009)。
9
1.4 利用構樹來推測南島語族文化的起源與傳播
有關南島語族的起源及發展,來自不同的學術領域,闡述多樣且互相衝突的 理論及數據,均顯示了南島語族遷徙歷史的複雜。所以,這方面的研究需要加入 更多新的證據以增加我們對南島語族傳播的了解(Matisoo-Smith, 2007; Kirch, 2010)。為進一步探討南島語族的遷徙歷史,本研究利用「構樹」作為共生生物研 究的題材(Seelenfreund et al., 2010)。
樹皮布是南島語族文化的重要特徵,而身為樹皮布製作重要原料的構樹,也 成為探討南島語族傳播的絕佳素材。作為共生者研究的對象,構樹有許多其他物 種無法達到的優點。首先,構樹為樹皮布原料,且廣布於整個南島語族的分布區,
尤其是快車說所推測的原鄉─台灣,所以構樹相當適合用來檢測快車說對南島語 族傳播的論述。另外,根據野外調查及標本紀錄,構樹在太平洋地區無法自然傳 播,代表構樹在當地的遷徙須藉由人類進行,符合commensal model 的論述。而且 不像其他共生者物種多為食用生物,構樹的調查可以從「衣著」的觀點來研究南 島語族的活動,與南島語族的文化有更緊密的聯結。
智利的研究者 Seelenfreund 在 2010 年也曾提出相似的論述 (Seelenfreund et al., 2010a),也依此論述初步在薩摩亞、東加、斐濟、法屬波里尼西與復活節島等不同 島嶼採集了18 株構樹樣本,並與來自台灣不同水系的 14 株構樹相比較進行親緣 地理學研究 (Seelenfreund et al., 2011),實驗結果發現,所有分布於大洋洲的構樹 均屬於同一基因單型。然而可能是東亞地區的取樣少,也沒有涵蓋構樹所有的天 然分布範圍,研究結果並未釐清大洋洲構樹與東亞、東南亞構樹間的關係。另外,
如果要檢測不同人類傳播的假說,也有必要對島嶼東南亞跟近大洋洲島嶼如印尼、
索羅門等地的構樹進行取樣。而且構樹在大洋洲的拓殖多依靠人類協助無性繁殖,
所以在大洋洲取樣數少不會影響結果,但是構樹在東亞地區多為天然傳播,基因 交流頻繁,若沒有足夠的取樣可能無法正確評估構樹的遺傳多樣性,與重建東亞
10
與大洋洲構樹間之親緣關係。所以,為重建構樹在東亞與大洋洲之間的傳播過程,
對東亞、東南亞以及大洋洲的構樹族群進行廣泛的採樣是必要的。
本研究將利用東亞與大洋洲地區構樹的分子資料比對,探討南島語族分布區 構樹的親緣地理關係。同時討論台灣與其他地區族群可能的擴散模式與遷徙途經。
本研究預期達成下列幾項目標:
1. 東亞與大洋洲不同地區構樹的遺傳結構差異與基因交流 2. 了解東亞與大洋洲構樹的族群變動歷史
3. 分析東亞與大洋洲不同構樹族群的遺傳分化程度與散布歷程
4. 結合不同的分析資料,分析構樹的族群遺傳結構變異是否能看出人為傳播的 跡象
5. 比較構樹的傳播是否能與快車說推定的南島語族拓殖路線相互對照
11
2 材料與方法
2.1 分類群取樣與樣區劃分
本研究針對東亞及大洋洲的構樹進行採樣。研究之採樣涵蓋大部分的構樹天 然分布地以及南島語族文化分布範圍,包含台灣、日本、菲律賓、中國東南、越 南、印尼的蘇拉威西以及南太平洋諸島包括夏威夷、復活節島、馬克薩斯群島 (Marquesas Islands)、薩摩亞 (Samoa)、斐濟 (Fiji)、索羅門群島 (Solomon Islands) 和 印度洋的法屬留尼旺群島 (Réunion)。其中,復活節島、馬克薩斯群島、薩摩亞、
斐濟的樣本為NCBI 下載的 DNA 序列,其他地區的樣本均是由採集者採集再攜回 台灣進行實驗。
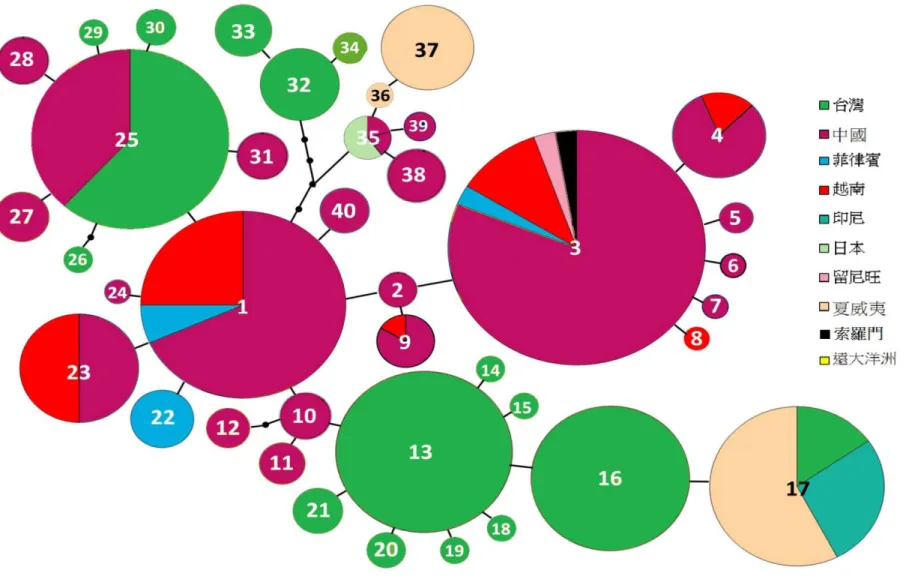
關於採集的分工,筆者主要負責台灣與越南兩地的樣本 (越南的採集由教育部 世界南島研究計畫辦公室所資助);夏威夷與印尼的構樹採樣由史前博物館張至善 先生完成 (世界南島研究計畫辦公室資助);夏威夷之外的遠大洋洲採樣則是智利 大學Deniela Seelenfreund 博士團隊所完成,並提供序列資訊進行本研究;中國的 樣本主要為台大森林系鍾國芳老師、游旨价與中央研究院生物多樣性研究中心張 家豪先生所蒐集;日本的樣本由游旨价提供;索羅門群島的樣本來自科博館楊宗 愈博士研究室;而非洲留尼旺群島的構樹樣本是由法國國家植物園Mascarin 常駐 中心的研究者Jean Hivert 提供 (圖 3)。
在東亞與東南亞地區,由於採集區域大多為海島,所以依據不同島嶼來區分 族群,分為中國、台灣、日本、越南、菲律賓、印尼(蘇拉威西)、留尼旺與大洋洲 等族群。其中由於中國與大洋洲的範圍過於廣大,而且也為了便於追查大洋洲個 體的起源,所以再將中國的樣本依照採集點分為福建、廣東、廣西、雲南、四川、
上海、北京及海南島。此外,由於遠大洋洲與近大洋洲的環境與文化差異,構樹 傳播至遠大洋洲與近大洋洲的途徑不一定相同,所以也將兩地區分開觀察。
隨著樣區中構樹的族群量有所不同,採集策略也有所調整。台灣及中國大陸
12
等構樹密集分布的地區,大約是每20 公里採集 1 到 5 株不同個體的嫩葉作為樣本 及引證標本 (儲存於中央研究院生物多樣性研究中心植物標本館, HAST),同一地 點的個體位置盡量分開以避免來自同源母樹。而構樹族群並不豐富的地區則是發 現個體就立即取樣,外群 (outgroup)則選用與構樹同屬的物種小構樹(B. kazinoki
Siebold et Zucc.)。所有的實體樣本均保存於矽膠乾燥劑中,做為後續DNA 萃取之
用。
2.2 DNA 萃取、PCR 擴增及定序
2.2.1 DNA 萃取
DNA 萃取使用改良的 CTAB 法(Doyle and Doyle, 1987),並根據 Porebski 等人 (1997)對處理富含二次代謝物物種的建議,在實驗過程中加入適量的 PVP (Polyphenolpyrollidine) 以移除構樹組織中的多醣類。DNA 萃取步驟如下:
(1) 秤取 0.1 克的乾燥植物嫩葉 (約小指指甲大小),加入海沙,以研缽磨成細粉放 入2 ml 離心管。
(2) 加入β-mercaptoethanal 4 μl、RNase A 10 μl (分解 RNA)、少量 PVP 粉末及 65。 C 的 CTAB 800 μl。
(3) 以機器震盪離心管,讓組織粉末與緩衝液均勻混合。
(4) 置於水浴槽以 65。C 水浴 1 小時,水浴時不定時搖晃離心管以促進其反應。
(5) 移出水浴槽並等待樣品稍微冷卻,每隻離心管加入 800 μl 的 CIAA (Chloroform : Isoamyl alcohol 24:1)。
(6) 劇烈搖晃離心管三分鐘,使管內溶液呈乳糜狀。
(7) 置於離心機以 13,500g 離心 10 分鐘。
(8) 將離心後之上清液加入新的 1.5 ml 離心管,加入 700 μl CIAA。
(9) 再次劇烈搖晃離心管 1 分鐘,再以 17,000g 的速度離心 10-15 分鐘。
13
(10) 將上清液移至新離心管之後加入 1/10 倍上清液體積的 Sodium Acetate 及 0.54 倍總體積的-20。C 異丙醇 (isopropenol) 幫助 DNA 沉澱並溶出其他雜質。
(11) 輕輕搖勻之後置於-20℃冰箱冰存 2-10 小時。
(12) 以 17,000 g 的速度離心 15 分鐘,移除上清液,並以 75%酒精清洗沉澱物。
(13) 以 17,000 g 的速度離心 1 分鐘,移除上清液,並以 95%酒精清洗沉澱物。
(14) 重複 (13),並至於抽風櫃中氣乾 1-2 小時。
(15) 加入 10-30 μl 的 TE buffer 回溶。
溶於 TE buffer 之 DNA,取 2 μl 進行以 1 % 瓊脂膠 (agarose) 電泳分析,以 DNA 標準品 (marker) 做為比對,估計樣品 DNA 之濃度,並以之為依據,取量進行 PCR 反應。
2.2.2 聚合酶連鎖反應 (Polymerase chain reaction, PCR)
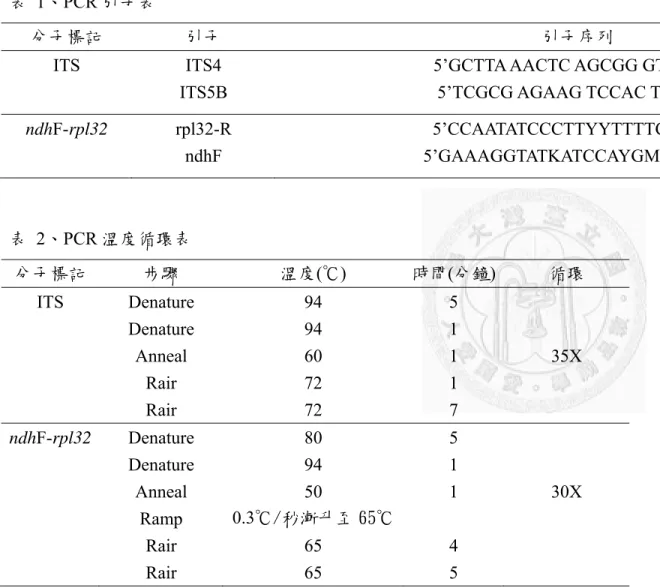
本研究共使用兩段 intergenic spacer 作為分析之用,分別是核醣體核酸基因 (rDNA) 的內轉錄區間 (~600kb) 以及位於葉綠體的 ndhF-rpL32 (~1100kb),使用的 引子 (primer) 如表 1 所列。聚合酶連鎖反應於 200 μl 離心管進行,反應總體積為 20 μl,組成為 2 μl 樣品模板 DNA (template DNA),核酸引子 ITS4、ITS5 Primer 各2 μl 及二次水 (ddH2O) 4μl 進行反應,其他 PCR 所需成分如 MgCl2、dNTPs、
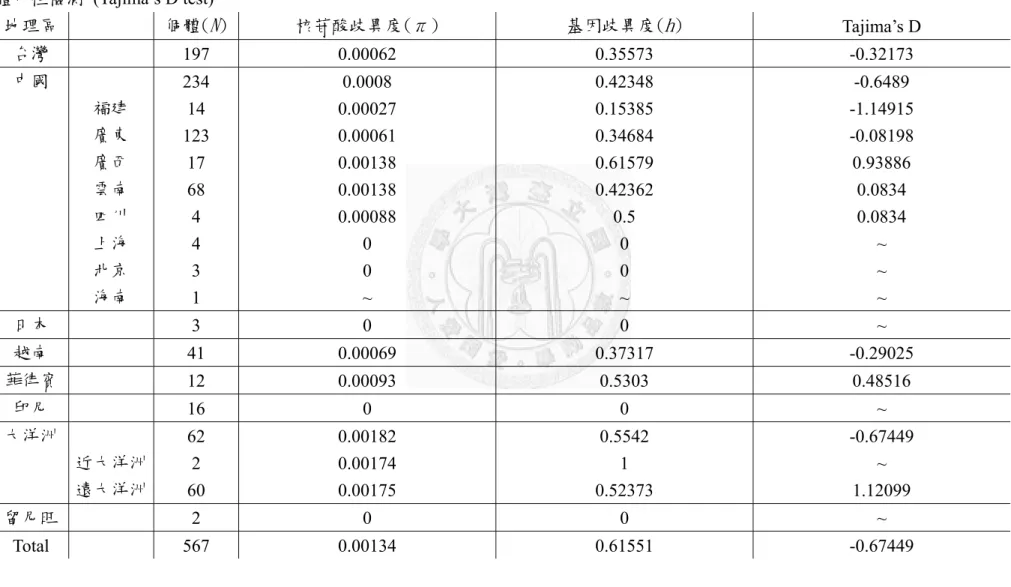
Taq polymerase 等,則使用 2x Taq DNA Polymerase Master Mix RED (Ampliqon) 代 替。此外,ITS 與 ndhF-rpL32 的溫度循環流程分別依照不同的程式進行詳見表 2
2.2.3 洋菜膠電泳分析
將 1.5 g 的洋菜膠與 100 ml 1X TBE buffer 混合並置入微波爐加熱溶解,待稍 涼之後再加入5 μl 的 Healthy Dye。置入至交容器之後,約 1 小時即為 1.5%電泳膠 片。
在電泳槽中放入足量 1X TBE buffer ,再將已用微量吸管加入樣本的膠片置入 電泳槽,開啟電源以100 伏特進行約 20 分鐘的的電泳分析。完成之後自電泳槽取
14
出膠片放入紫外光照相系統拍攝影像與存取。
2.2.4 定序
取 4-7 μl 純化後的 PCR 產物,加入 1.5μl 的 10μM 引子在離心管中,再加二次 水使樣品體積達到10μl,即可送至中央研究院植物暨微生物研究所核酸分析核心 實驗室 (ABI 3730, USA) 或基隆米克斯生物科技有限公司(ABI 3730XL, USA)定 序。每條序列皆訂兩端並以肉眼確認序列正確性,完全相同之序列即判別為同一 基因單型 (haplotype),若有單點位不同鹼基的情形則會再次定序確認。
2.2.5 構樹之 ITS 引子開發
由於構樹葉片在野外極易發生病害感染,一般通用的引子極易同時擴增出構 樹及真菌的ITS 序列,造成直接定序的困難,故本研究根據已知構樹 ITS 序列,
開發構樹ITS 專用引子。研究過程如下:
將構樹的PCR 產物加入 5%洋菜膠的膠片孔中,以 50V 的電壓進行約 1 小時的電 泳分析,電泳結束後再用紫光照相系統,確認目標條帶的位置並將該片段自洋菜 膠切下,以QIA quick Gel Extraction Kit (QIAGEN, Germany) 依照其指示說明進行 純化以進行定序作業。完成的序列資料,利用Primer3 (Rozen and Skaletsky, 2000) 得到合適的引子組合再重新進行PCR 確認該組合的效能,以選取合適的引子組。
2.3 資料分析
2.3.1 序列整理與資料矩陣建立
定序之後所得的資料使用 SeqManTM Pro (DNASTAR, USA) 將同一樣本兩端 組合,手動修正錯誤後,留下引子間的序列存為FASTA 格式以供後續資料處理,
同時也另存ab1 格式留下備份便於後續修改。以 MEGA5 (Tamura et al., 2011) 開啟
15
全部的FASTA 序列,再用 MUSCLE 程式 (Edgar, 2004) 進行比對製作資料矩陣 (data matrix),經過目視調整之後存成 FASTA、NEXUS 等檔案格式進行分析。無 法確認同源性的位置 (ambiguous site) 如大片段的插入或刪除 (insertion and deletion, indel)、微衛星 (microsatellite) 去除不予分析、gap 則處理為 missing data。
2.3.2 族群遺傳變異分析
基因單型多樣性 (haplotype diversity, h)、核苷酸多樣性 (nucleotide diversity, π) 均利用軟體 DnaSP (Librado and Rozas, 2009) 進行計算,來量化顯示族群的遺 傳歧異度。基因單型多樣性 h 的算式為
h=2n(1-Σxi2)/(2n-1) xi是第 i 個基因型的頻率,n 是取樣個體數(Nei, 1987)。而核苷酸多樣性π值的算
式是
π=nΣxixjπij/(n-1)
其中n為取樣的 DNA 數量,πij是第 i 與第 j 基因單型間的序列差異 (Nei, 1987)。
比較兩數值的相對大小可協助推論族群的演化歷史以及得出其他的族群遺傳相關 參數,例如族群間的遺傳分化指數(Fst) 和族群間的基因交流指數 (Nm)。
Fst是族群間遺傳分化指數,主要用來判讀族群間遺傳分化 (genetic differentiation) 的情形。程式 DNAsp 所使用的 Fst為
(Fst)=1-Hw/Hb
Hw是同一小族群內不同序列間的平均差異數量,Hb則是不同小族群間的平均差異 數量 (Hudson, Slatkin, and Maddison, 1992)。而 Nm 值則表示推估的每世代族群遷 徙個體數,該值可間接評估基因交流的大小 (Slatkin, 1985)。Nm 是由該公式估算:
Fst=1/(1+2Nm)
若 Nm 值為 1,表示地區性族群間每世代平均有 1 個個體交流,足以防止個別族群 基因漂變 (genetic drift) 產生的遺傳分化。所以當 Nm 值小於 1,即表示族群間的
16
基因交流較弱,無法抑制漂變所產生的遺傳分化。
此外,以 DnaSP 計算 Tajima’s D test,以檢測族群間的變異是否只受到隨機基 因漂變而不是天擇的影響,並進行1000 次 permutation。Tajima’s D test 主要利用 核苷酸變異出現的頻率 (π) 和核苷酸變異位置的數量 (θ) 相關性作為計算依據,
利用有變異的核苷酸位置數與兩兩序列的核苷酸差異平均數兩者估算的θ值差異。
由於此檢測的虛無假設 (null hypothesis) 為檢測的序列均來自有效族群量穩定 (constant effective population size) 的族群,該段序列的突變也都是中性突變 (selective neutrality)。如果檢測的族群 (序列) 違反任一項假設,檢測的結果就會偏 離 0 拒絕虛無假設。在偏離0 的情況下,如果 Tajima’s D 值為負,表示曾經歷族群 擴張或是平衡選汰 (balancing selection),產生大量的低頻度基因單型。而如果 Tajima’s D 值為正,顯示過去的遺傳變異大多已經消失,表示族群曾經歷瓶頸效應 (bottleneck effect) 或是定向選汰 (directional selection)。所以 Tajima’s D test 也可以 用來推論族群發生的歷史事件。
2.3.3 建構親緣地理網狀圖及親緣地理關係分析
利用統計簡約 (statistical parsimony) 的概念,TCS ver.1.21 (Clement, Posada, and Crandall, 2000) 軟體建構不同基因單型間的最小關聯網狀圖。親緣地理網狀圖 的結構,可以根據以下條件決定各個序列中可能的祖先基因單型。整體來說,祖 先型的判定通常依據以下幾項準則:頻率最高、分布範圍最廣、位於結構中心的 基因單型多,以及與外群關係最近的基因單型,而TCS 程式推定的祖先型則是選 擇位於網狀結構中心的基因單型 (Castelloe and Templeton, 1994; Clement, Posada, and Crandall, 2000; Posada and Crandall, 2001)。
親緣地理網狀支系分析 (Nested Clade Phylogeographic Analysis, NCPA) 用以 推測演化途徑的原則有以下3 項:1. 新的基因單型是由古老的基因單型所衍生。
2. 若缺乏基因交流,則新發生的突變傾向留在原有的地理區。3. 如果所使用的特
17
徵不受演化影響,nested clade 中內部的 clades (internal clades) 會比末端 (tip clades) 古老。再把地理資訊與分子遺傳資料放進GEODIS ver.2.5 (Posada, Crandall, and Templeton, 2000),根據遺傳變異與族群地理位置提供的資訊,我們可以了解不同 族群間的遺傳變異程度與它們地理位置間的關係。經計算clade distance (Dc) 與 nested clade distance (Dn) 來了解每一個 clade 內地理分布的情形,與每個 clade 與 上一階層地理分布距離的值,與這些值統計上是否顯著來評估不同基因單型的傳 播能力,再配合Temploton (2004) 提出的檢索表 (inference key) 來了解遺傳變異 的空間分布以地理關係的了解,並進一步推測過去族群可能發生的歷史事件。
藉由比較相鄰小族群間的遺傳距離,軟體 BARRIER ver. 2.2 可以顯示不同族 群的空間關係,進而了解我們對族群的區分是否符合遺傳上差異。BARRIER 利用 Delaunay triangulation (Manni, Guerard, and Heyer, 2004) 演算法將不同個體間的遺 傳距離矩陣,以及採集時紀錄的地理座標加以分析,並偵測出可能的遺傳屏障出 現在何處。
18
3 結果
3.1 序列變異分析
本研究在東亞、東南亞及大洋洲地區共取得 123 個族群共 516 份 DNA 樣本,
並成功定序571 條 ITS 片段與 516 條葉綠體 ndhF-rpl32 區間,其中由於部分個體 的ITS 片段是由國家生物科技資訊中心 (National Center of Biotechnology
Information, NCBI) 取得、部分個體 PCR 失敗以及部分個體 concerted evolution 不 完全,因此較另一片段為多 (圖 3)。
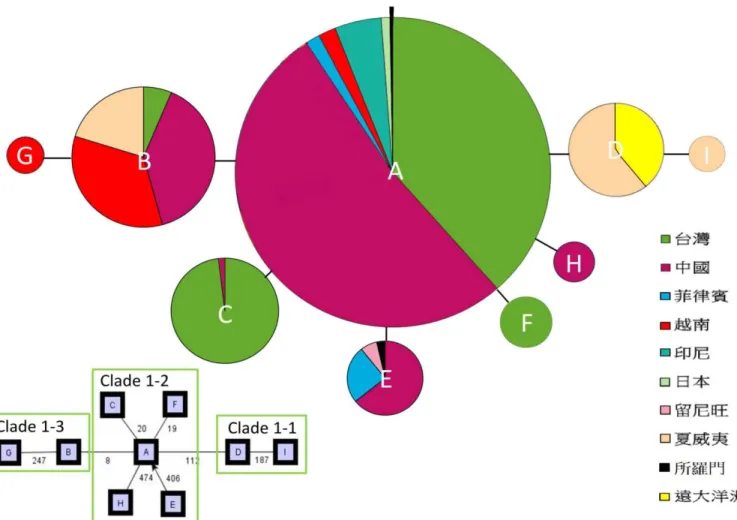
ITS 片段總長為 596 bp(base pair, bp),總共有 9 個多態位點 (polymorphic sites),
可整理出9 個基因單型(圖 4),其中 4 個地區有其獨有的基因單型。1 個分布於中 國廣西,1 個是台灣的獨有型,分布在台灣的中北部。另外,大洋洲也有 2 個獨有 的基因單型,1 個廣泛分布於遠、近大洋洲,1 個則只分布在夏威夷 (表 9)。
而 ndhF-rpl32 片段總長為 1196 bp,有 41 個多態位點,可建出 40 個基因單型 (圖 5)。有 32 個是與地理分區明確相關的獨有基因單型,包括中國各省與台灣分別 獨有14 個基因單型,越南 1 個,菲律賓 1 個及遠大洋洲 2 個基因單型(表 10)。
3.2 族群遺傳變異分析
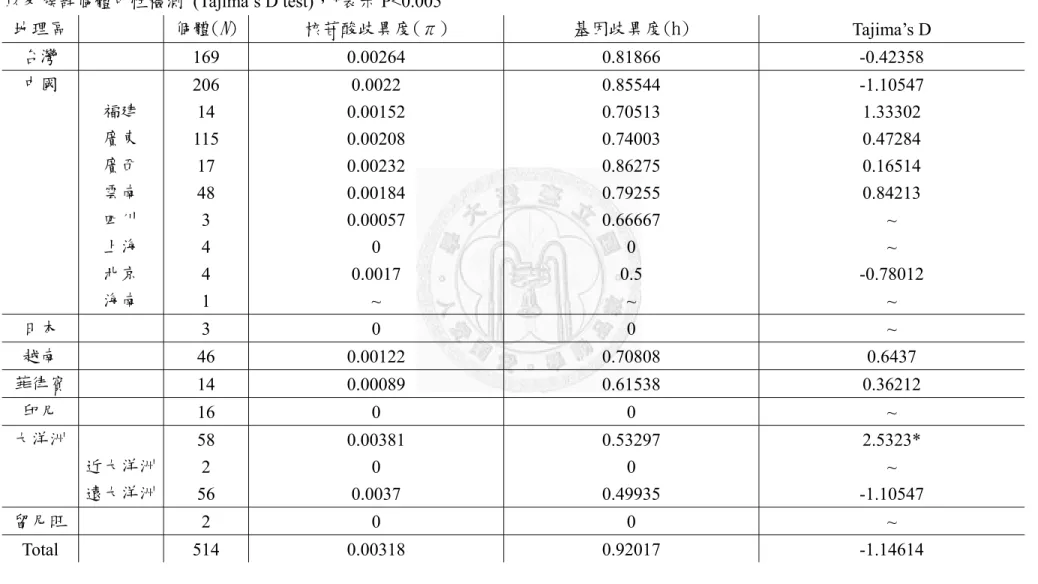
3.2.1 ITS 片段遺傳歧異度分析
將所有取得的構樹族群依照地理分區歸為台灣、日本、中國福建、廣東、海 南島、廣西、雲南、四川、上海、北京,東南亞菲律賓、越南、印尼、位於印度 洋上的留尼旺群島、大洋洲的遠大洋洲以及近大洋洲等16 個族群,利用程式 DnaSP 計算總體與各族群之核苷酸歧異度 (nucleotide diversity, π) 及基因歧異度
(hsplotype diversity, h)(表 3),構樹的核苷酸歧異度π值為 0.00133,基因歧異度 h 值為0.613。所有族群的基因歧異度介於 0 到 0.58788 間,而核苷酸歧異度則是介
19
於0 到 0.00204 之間。東亞及大洋洲各族群中,基因單型多樣性最高的是夏威夷 h= 0.58788,其次是菲律賓 (0.53030) 與台灣 (0.50772)。
另計算以上分群間的遺傳分化指數 (Fst) 與基因交流指數 (Nm) (表 5)。根據分 群間的 Fst 與 Nm,結果顯示南島語族分布區 (印尼、大洋洲、夏威夷與留尼旺群 島)的族群與東亞其他地區的族群間 Fst 值均大於0.15,最小值為台灣與印尼的 0.18,
其次是印尼與廣東 (0.20) 與雲南 (0.22),而跟遠大洋洲的 Fst 最小值是 0.26 (廣西),
顯示東亞與南島語族分布區至少都出現了中度的遺傳分化 (Wright, 1978)。然而南 島語族分布區與其他地理距離相近的族群比較 Fst 反而更大,像是印尼與越南(0.79) 或是菲律賓(0.55),顯示構樹分布似乎不符合距離隔離模式 (isolation by distance)。
以軟體DnaSP 以 1000 次 permutation 計算所有構樹族群與各地理區的 Tajima’s D 值(表 3)。結果顯示整體的 Tajima’s D=-0.67,而各地區不管是整體的或是不同地 理區域,Tajima’s D 值均偏離 0,但是統計上並不顯著(P> 0.1),這樣的結果顯示在 ITS 片段沒有偵測到族群變動,序列也符合中性假說的預測。
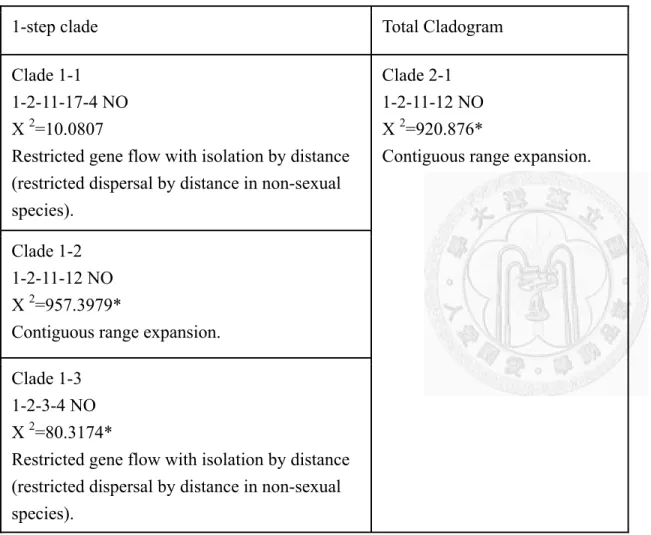
3.2.2 ndhF-rpl32 片段遺傳歧異度分析
利用程式DnaSP 計算 ndhF-rpl32 片段中總體與各族群之核苷酸歧異度及基因 歧異度 (表 4),總體的核苷酸歧異度π=0.00317,基因歧異度值為 0.920。不同族 群的基因歧異度介於0 到 0.86275 間,而核苷酸歧異度則是介於 0 到 0.00370 之間。
東亞及大洋洲各族群中,基因單型多樣性最高的是廣西h= 0. 86275,其次是台灣 (0.81866)與雲南(0.79255)。
而分群間的遺傳分化指數與基因交流指數 (表 6)。可以發現四川、北京、上海、
日本、相關的數值常為0 或負值,這樣的數值可能是由於以上 4 族群的數量較少 的結果。根據分群間的 Fst 與 Nm,結果顯示南島語族分布區的族群與東亞其他地 區的族群間 Fst 值均大於 0.15,最小值為台灣與遠大洋洲的 0.22,其次是近大洋洲 與廣東(0.36),夏威夷與廣西(0.38),顯示東亞與南島語族分布區間至少出現了中度
20
的遺傳分化。東亞與南島語族分布區族群間基因交流指數唯一大於1 的存在於台 灣與遠大洋洲間,顯示遠大洋洲跟台灣間雖然有海洋隔閡但依舊有一定程度的基 因交流。
運用軟體DnaSP 以計算所有構樹族群,與各地理區的 Tajima’s D 值(表 4),結 果顯示不管是整體的Tajima’s D=-1.15,除了某些地區只有一個基因單行或是個體 數少於4,其他地區不管是整體的或是不同地理區域,Tajima’s D 值均偏離 0,但 是統計上並不顯著 (P> 0.1),顯示這樣的偏離結果可能不是單純受天擇選汰所造成,
而是受到族群歷史,像是族群擴張或是瓶頸效應的影響。值得注意的是,整個大 洋洲地區的Tajima’s D 值顯著大於 0 (P< 0.05),顯示構樹的母本可能在大洋洲經 歷過瓶頸效應。
3.3 親緣地理關係分析
3.3.1 從 ITS 片段看親緣地理關係
雖然東亞及大洋洲構樹族群的親緣關係樹,顯示構樹族群大體的樹型結構與 地理區的關係並不明顯。但是各clade 還是能反映出部分與地理位置的相關性。在 ITS 片段的分析結果呈現標準的星狀結構,572 個樣本除了外群之外分為 9 個基因 單型(ITS-A、ITS-B、ITS-C、ITS-D、ITS-E、ITS-F、ITS-G、ITS-H 及 ITS-I) (圖 4)。在全部族群中以 ITS-A、ITS-B 與 ITS-E 的頻率較高分布範圍也較廣,分別有 335、94 和 28 個個體廣布於中國、島嶼東南亞以及大洋洲等地。其中 ITS-A 是程 式TCS 所推論的祖先型,也是分布範圍與頻率最為廣泛的基因單型,除了印度洋 留尼旺島,在中國各省、日本、台灣、菲律賓、越南、印尼與近大洋洲 (索羅門群 島)均有分布,其中印尼(蘇拉威西)的所有個體都是 ITS-A。ITS-B 與 ITS-E 也是廣 布的基因單型,ITS-B 主要分布在中國西南、台灣、中南半島與夏威夷,而 ITS-E 族群分布在廣東、雲南、菲律賓與數個來自近大洋洲(大洋洲)與留尼旺群島(印度
21
洋)的個體。此外,D 與 I 型的所有個體均來自於遠大洋洲,沒有任何東亞個體在 內。
從構樹的親緣網狀圖中,總共包含3 個 1-step clade 與 1 個 2-step clade (表 7)。
(Nested Clade Analysis) 可以用來檢視支系與地理位置之間的關係,在 ITS 片段中,
所有支系均與地理分布相關。根據 inference key 檢索所得到的結果,現今東亞及 大洋洲構樹的族群分布與clade1-2,包含中國、台灣與大洋洲索羅門群島的族群可 能是族群連續擴張 (contiguous range expansion) 的結果。而在 clade1-1 也就是波 里尼西亞諸島的族群間則推論出有限的基因交流並有距離分化效應 (Restricted gene flow with isolation by distance),而在無性繁殖的族群中則可能是傳播受限 (restricted dispersal by distance in non-sexual species) 的結果。clade1-3 的台灣、中 國和夏威夷的族群之間也得到類似的結果。
3.3.2 從 ndhF-rpl32 片段看親緣地理關係
TCS 所建立的親緣地理網狀圖以數個高頻度的基因單型為基礎向外擴張,517 個樣本除了外群之外分為40 個基因單型(1-40) (圖 5)。在全部族群中以 cp-1、cp-3 、 cp-17 與 cp-25 的頻率較高,各自有 60、74、59、47 個體屬於該基因單型。其中 cp-3、cp-17 是所屬個體同時分布在東亞與大洋洲的基因單型,分布範圍涵蓋印度 洋、東亞及大洋洲,也顯示其傳播過程的特殊。cp-3 的樣本主要來自於廣東,還 包含中國其他省份與東南亞 (越南、菲律賓)、近大洋洲 (索羅門群島) 的樣本。
cp-17 則包含了來自台灣、印尼與遠大洋洲的族群。另外 cp-36 跟 cp-37 為遠大洋 洲獨有的基因單型,所有的個體都來自夏威夷諸島,不過與其關係最近的基因單 型 (cp-35) 則是來自日本與中國。值得注意的是在 ndhF-rpl32 片段,所有近大洋 洲與留尼旺群島的樣本跟來自中國(福建、廣東、雲南)、越南的樣本屬於一個 clade,
且不包含任何遠大洋洲的個體。遠近大洋洲的族群來源並不相同,分布在索羅門 群島的個體與夏威夷的個體基因單型不同,夏威夷的族群來源也相當複雜。
22
在 Nested Clade Analysis 的結果中,共有 16 個 1-step clade、7 個 2-step clade 與3 個 3-step clade (圖 6)。經過檢測,除了 clade1-2、1-6、1-8、1-12、1-15、1-16、
2-1 無法檢測,clade1-3、1-4、1-11 與地理相關性不顯著外,共有 16 個支系與地理 相關,以Templeton's inference key 檢索所得的結果如表 8。在 ndhF-rpl32 片段中與 ITS 片段的結果類似,整體的結構顯示構樹的傳播呈現連續的族群擴張(contiguous range expansion),在 clade1-14 中推測台灣東部、夏威夷和印尼的部分族群因為長 距離拓殖後的隔離事件 (long-distance colonisation possibly coupled with subsequent fragmentation) 或是隔離事件後族群擴張 (past fragmentation followed by range expansion.) 而呈現目前的分布狀況。另外,clade1-5 內則推測東亞跟近大洋洲族群 間可能發生過有限的基因交流並有距離分化效應 (restricted gene flow with
isolation by distance)或是無性繁殖的族群中,則可能是傳播受限 (restricted dispersal by distance in non-sexual species) 的結果。
根據以上的分析,雖然所在的地理位置相近,但是近大洋洲與遠大洋洲個體 的來源並不相同,這也符合DnaSP 所呈現的遺傳分化指數與基因交流指數的數據 遠大洋洲與近大洋洲地理距離近但遺傳距離遠的現象。值得注意的是,ITS 親緣關 係網狀圖中的ITS-D 與 ITS-I,與 ndhF-rpl32 網狀圖中的 cp-17 所包含的遠大洋洲 個體幾乎完全一樣,ITS-B 與 cp-36、cp-37 也有一樣的情形。顯示雖然父、母本的 傳播過程理應不相關,但是在大洋洲族群間確有相當類似的分布模式。
3.4 遺傳與空間結構分析
由結果中可以發現,程式 BARRIER 根據兩組 DNA 資料矩陣各建立 7 條屏障 (barrier) (圖 7、圖 8),其中 3 個屏障在兩組資料中都出現,而且有些屏障雖然位置 不盡相同但卻有類似的分布模式 (pattern)。首先,兩片段共同支持的 barrier 全部 位於夏威夷群島,在地形平坦沒有明顯阻隔的夏威夷卻暗示如此多的隔離事件存
23
在,顯示構樹進入夏威夷的機制可能遠比我們想像的來得複雜,所以才形成夏威 夷的族群地理距離近,但是遺傳距離遠的現象。ITS 的某些 barrier 將大陸東南亞與 島嶼東南亞區分開來,而 ndhF-rpl32 片段則是顯示出 barrier 將夏威夷的族群與大 洋洲其他地方以及東南亞族群的遺傳差異,這可能是因為兩片段的傳播機制不同 的結果,也可能是因為葉綠體 ndhF-rpl32 片段在大洋洲的取樣差異很大,所以造 成結果不一致。此外,ITS 與 ndhF-rpl32 片段也分別顯示台灣西南部與台灣北部的 族群可能存在某種屏障。
24
4 討論
構樹的地理分布,廣泛分布於日本、台灣、中國南部、中南半島、泰國、馬 來西亞、印度和太平洋諸島等地 (Liao, 1996)。然而,多項證據顯示構樹的分布受 到人為傳播的影響 (Matthews, 1996; Florece and Colladilla, 2008; Kirch, 2010;
Seelenfreund et al., 2010a; Seelenfreund et al., 2011),所以現在的分布並非全是天然 傳播的結果。Matthews (1996) 認為構樹可自然繁殖的範圍北界在中國大陸的黃河,
南界在泰國至越南,西界位於大陸的喜馬拉雅山區海拔 900 m 至 1,200 m 處至緬 甸北部一帶,東界則在台灣 (圖 2)。而島嶼東南亞地區及大洋洲的構樹族群可能都 是人為傳播的結果。如果將中國、台灣等東亞較可能天然傳播的族群與大洋洲跟 島嶼東南亞這些非天然傳播地區分開討論,應能對構樹的傳播提供較精確的解 釋。
4.1 東亞不同地理區構樹族群結構的解釋與推論
4.1.1 東亞及大洋洲構樹種源中心的推測
台灣與包含華南、中南半島在內的大東亞地區是南島文化產生與向外傳播的 起源地,也是構樹向外傳播的種源中心。如果要利用構樹探討南島文化的傳播,
必須先理解這些地區構樹族群多樣性分布與成因,以推測構樹傳播的方向,也便 於釐清解釋不同南島語族傳播假說的能力與限制。
根據 TCS 所建立的親緣地理網狀圖只能顯示東亞構樹族群的多樣性遠比大 洋洲高,而且台灣、越南、中國各省不同地理區間的關係不明顯 (圖 4、圖 5),不 易解釋這些地區間所有族群的遺傳結構與來源。但經由ITS 與葉綠體所產生的 NCPA 的分析結果 (表 7、表 8)、兩個不同片段 Tajima’s D 在台灣、中國、越南的 結果 (表 3、表 4)與 ITS 與葉綠體的親緣關係網狀圖中程式推定的祖先型 (cp-1 與 ITS-A),顯示大洋洲構樹的起源地位於東亞,包含台灣、越南、中國,再漸次拓殖
25
到島嶼東南亞、大洋洲等地。再者,兩個分子標記顯示核酸歧異度與基因歧異度 最高的地方主要分布在台灣與中國西南,包含福建、廣東、廣西、雲南。所以,
如果進一步比較三個不同地區的遺傳多樣性與族群結構,可以發現相較於越南,
中國與台灣的遺傳多樣性較高,顯示中國與台灣較可能是東亞與大洋洲構樹的種 源中心。前人研究指出,台灣東南部與中國南方在更新世冰河期曾是生物的避難 所 (refugia),像是中國大巴山、雲貴高原東部、南嶺及武夷山以及台灣東南部的 台東、屏東一帶,都被推測為最後一次冰河期時重要的生物避難所(Hwang et al., 2003; Cheng, Hwang, and Lin, 2005; Wang et al., 2009; Kuo et al., 2010; Chou et al., 2011)。當時與構樹棲位相似的亞熱帶植物如牛樟 (Cinnamomum kanehirai)、賽巒 華 (Eurycorymbus cavaleriei) 都曾限縮於台灣與中國南部的不同避難所(Wang et al., 2009; Kuo et al., 2010),所以構樹現今的分布也很可能受到過去冰河期的影響,
在這兩地呈現較高的歧異度 。而構樹遺傳多樣性由分布中心 (中國、台灣、越南) 向外下降的現象,也符合大部分研究所述大洋洲構樹起源於東亞的推論(Matthews, 1996; Zerega et al., 2005; Kirch, 2010; Seelenfreund et al., 2010a)。根據以上結果,也 可以得知,構樹並不適合作為探討南島語族近大洋洲起源的「俾斯麥群島原居民 假說」與專注於島嶼東南亞與大洋洲關係的「西波里尼西亞航海廊道說」的研究 材料。
4.1.2 台灣構樹族群遺傳結構分析
台灣在南島文化傳播中就算不是南島文化的起源,也是重要的傳播中繼站 (陳 堯峰, 陳叔倬, and 許木柱, 2011),語言學研究證明台灣的重要地位(Blust, 1999;
Diamond, 2000),本研究結果也顯示台灣是遠大洋洲構樹族群最重要的起源地之一。
所以對台灣構樹遺傳結構的了解相當重要。然而,本次研究中兩個分子標記卻對 台灣是否屬於種源中心有不同的推論,因此有必要針對台灣構樹的遺傳結構加以 討論。
26
在ITS 片段建立的親緣地理網狀圖中,祖先型包含部分來自台灣的個體 (38.51%),且該 ITS 拷貝廣布於全台。然而葉綠體親緣地理網狀圖的結果卻與 ITS 片段的結果相左,沒有任何台灣的成分。ITS 與葉綠體分子標記常發生不一致,造 成因素可能有:1.雜交事件, 2. 譜系排序不完全 (incomplete lineage sorting), 3.
ITS 協同演化 (concerted evolution)的機制, 4. 由於花粉和種子傳播能力不同造成 不同分子標記所顯現之族群動態不一致。如果因為構樹與其他物種雜交,使分子 標記間親緣地理結構不一致,雜交個體序列會呈現另一親本的ITS 拷貝,但在實 驗過程中並未發現類似的現象。另一方面,雖然無法排除譜系排序不完全與ITS 協同演化對構樹遺傳結構造成的影響,但是如果發生這兩種情況應會觀察到所有 的族群均受影響,而不是侷限於特定地理區。所以花粉和種子傳播能力的差異是 較可能的原因。
由於台灣地形相對複雜,許多平地分布的物種因為山脈 (Yang et al., 1994;
Huang, Hwang, and Lin, 2002)、水系(Liao and Hsiao, 1998; 何坤益 and 張景富, 2006)的阻隔被切分成小族群,因為基因漂變 (genetic drift) 而形成不同的基因單型。
由葉綠體片段的親緣地理網狀圖來自台灣侷限分布的基因單型偏高以及NCPA 分 析來自台灣基因單型,顯示構樹母本在台灣島內基因流受限(表 8,clade 1-13) , 都可以佐證以上的推測。因此,可以合理推斷構樹母系的傳播,因顯現種子傳播 歷史之葉綠體DNA 受到傳播距離影響,而產生地區性分化,但顯現花粉傳播歷史 之細胞核DNA 則可藉由長距離傳播、不受地理隔離而無顯著分化。
4.1.3 中國的構樹族群遺傳結構分析
在親緣地理學研究中,傳播方式不同與族群劃分錯誤,會影響對實驗結果的 分析與推論 (Dupanloup, Schneider, and Excoffier, 2002; Manni, Guerard, and Heyer, 2004; Templeton, 2010)。尤其是非天然分布的族群,因為人為介入,天然的地理屏 障不再是劃分族群的參考。如果天然傳播與人為傳播同時發生,將更難判斷物種
27
的傳播歷史。以中國大陸為例,從表5、表 6 可以發現,中國不同地區間,構樹族 群的關係與地理距離相關性不高,不具明確的地理結構。造成這些地區間族群結 構混亂的原因,除了不同地區取樣的數量不平均外,中國境內四處交互引種種植 構樹也是可能的因素 (李黨法 and 李月風, 2006; Liu, Shu, and Zhang, 2007)。由於 構樹有耐陰、耐汙染等特性,在中國部分地區被當成行道樹或是防沙林引進栽培(李 黨法 and 李月風, 2006; 張秋玉, 李遠發, and 梁芳, 2009),造成族群遺傳結構的改 變。
此外,相較於印尼、台灣等島嶼,易於定義族群,中國占地廣大,不易定義 族群的生長範圍,所以族群的定義不正確,也可能是造成分析結果族群結構模糊 的因素。由於像軟體BARRIER 可利用 Monmonier's algorithm 偵測出相鄰族群間突 然增加的遺傳距離,並將發生的位置顯示於圖上,如果要分析中國境內的族群關 係,利用像BARRIER 這樣的軟體來分群應是較為可行的方式。根據分析的結果 (圖 7、圖 8) ,在中國境內僅有雲南省顯示地理屏障,由於雲南地勢高聳,江河切割 嚴重,造成地形破碎,過去的研究即發現金沙江可能造成兩岸構樹的遺傳分化 (Chen et al., 2008)。另外,因為沒有明顯南北向地理阻隔,兩個分子標記的 Fst分 別顯示,廣西與雲南等南方省分族群與越南採集樣本的關係較密切。至於中國內 部不同地區間構樹族群結構成因與傳播過程,可能還需要後續的採樣或是分析才 能了解。
再者,觀察族群間 Fst值、Nm值的比較可以發現,四川、北京、上海、海南等 地相關的數值常為0 或負值,而中國其他地區間的 Fst值卻偏高。以上結果可能由 兩個因素造成:第一個是取樣不平均,由於四川、北京等族群樣本數均少於5 個,
而其他族群樣本量均多於10 個 ,根據 Hudson (1992)等的 Fst演算法不採用重複取 樣,造成計算上的偏差,也連帶影響 Nm的結果,而造成部分地區間的 Fst值偏高。
而第二個原因可能是族群區劃錯誤。由於高估構樹族群的傳播能力,結果同一族 群內個體變異大於不同族群個體間的變異 (Arnason and Palsson, 1996; Yang et al.,
28
2007)。相對於其他木本植物,如同為桑科的薜荔 (Ficus pumila L.)(Chen et al., 2008),
樺木科的西樺 (Betula alnoides Buch.-Ham. ex D. Don) (Zeng et al., 2003),所顯示的 遺傳分化指數都遠低於構樹 (表 5、表 6),這表示分析中當初設定的族群範圍太大,
也顯示構樹的傳播能力可能極為有限。
4.1.4 越南構樹族群遺傳結構分析與生態觀察
大約在西元前 3500-4000 年,越南北部就出現樹皮布文化,被認為是樹皮布文 化可能的發源地之一 (鄧聰, 1997),所以越南是驗證南島語族傳播假說相當重要的 取樣地區。根據ITS 片段的親緣網狀圖,雖然解析度不足,仍可發現與台灣相比,
中國與越南的個體常同屬一個拷貝,而且該拷貝常涵蓋夏威夷及大洋洲的樣本或 是與之直接相連,無法完全排除中南半島與大洋洲構樹族群間的關係 (圖 5)。然而 解析度較高的葉綠體片段親緣網狀圖中,所有包含越南樣本的基因單型都位於離 遠大洋洲與夏威夷較遠的位置,顯示雖無法完全排除中南半島與大洋洲構樹族群 間的關係,但相對於台灣或是中國大陸,越南與大洋洲族群的交流相對較低。
此外,在前往越南採集的過程中,發現大約在通過北緯16 度線後,構樹的棲 地就逐漸被榕屬的植物取代。越南由於地形狹長,南北兩端的氣候差異相當大,
北部屬於亞熱帶季風氣候帶近似台灣,有春夏秋冬四季,而南部則屬於海洋性熱 帶氣候,一年只有乾濕兩季。而構樹雖然能適應極大範圍的光度條件,然而對乾 旱的忍耐力卻有一定限度 (陳忠義, 2007)。根據以上觀察,發現氣候的因素可能影 響了構樹的生長,也造成構樹在越南分布範圍的限制。這也暗示水分或氣候可能 是限制構樹生長的重要因子。由於不易適應長期缺水的環境,在長期乾旱地區構 樹可能需要人為照料才能生長。這項發現,佐證當初Matthews (1996) 對構樹天然 分布範圍的部分推測,也解釋了為何構樹在印尼無法天然繁殖,以及部分喪失樹 皮布文化的遠大洋洲島嶼構樹族群瀕臨絕種的原因(Seelenfreund et al., 2010a;
Seelenfreund et al., 2011)。
29
4.2 島嶼東南亞與大洋洲構樹族群遺傳多樣性的分布
4.2.1 近大洋洲構樹族群的起源
根據快車說的論述,南島語族自起源地出發後,曾在近大洋洲留了約 1,000 年,
之後才往遠大洋洲擴散 (Diamond, 2000)。如果能得知近大洋洲族群的起源,應能 推測大洋洲構樹的原生地。親緣關係網狀圖顯示近大洋洲的個體與來自福建、廣 東、雲南 (ITS) 與廣東、雲南、菲律賓、越南 (ndhF-rpl32)個體同一個拷貝 (基因 單型),顯示近大洋洲構樹可能來自中國或中南半島,不包含台灣,與廣布於遠大 洋洲的拷貝來源不符。根據文獻指出,部分近大洋洲的構樹族群是近代引進的 (Whistler and Elevitch, 2006),而且本研究在近大洋洲的採集點不是採自製作樹皮布 的村落而是高速公路旁,很可能是作為行道樹之用,而不是與南島文化相關的族 群。所以近大洋洲近代輸入的構樹可能是來自中國、中南半島一帶,如果合併對 菲律賓地區構樹的觀察,更可以確定部分近大洋洲的構樹可能是近代人為傳播的 結果。而與樹皮布相關近大洋洲構樹族群與遠大洋洲構樹的關係可能需要仰賴更 全面的取樣以進一步的研究。
4.2.2 近大洋洲與留尼旺群島構樹族群的關係
此外,圖 4 與圖 5 也可以觀察到印度洋留尼旺島的構樹族群都與近大洋洲同 一拷貝 (基因單型)。由於留尼旺島與馬達加斯加都屬於南島語族的分布範圍,因 此留尼旺群島族群的來源也許能指向南島文化的起源。近大洋洲跟留尼旺島相關 的族群來自福建、廣東、雲南、菲律賓、越南等地,這暗示了兩個結果:1. 留尼 旺群島與部分近大洋洲的構樹種群來源相同 2. 留尼旺群島構樹種源來自中國、越 南或菲律賓。由於菲律賓的構樹很可能不是天然族群,所以中國或越南應是較可 能的來源。許多文獻指出構樹在非洲是外來種 (Sheil, 1994; Newton‐Fisher, 2003;
30
Bosu and Apetorgbor, 2010),因此無法排除近代傳入非洲的可能性。根據以上資料 顯示,很可能部分近大洋洲與留尼旺群島的構樹同樣都是近代傳播,甚至可能是 來自同一種源的結果。
4.2.3 遠近大洋洲構樹族群的關係
要檢測快車說是否符合構樹的傳播,近大洋洲的樣本是否與遠大洋洲一樣的 基因單型 (拷貝) 以及分布模式相當重要,因為快車說強調南島語族位於近大洋洲 的長期停留,所以理論上如果構樹的遷徙符合快車說的論述,近大洋洲出現的基 因單型 (拷貝) 會直接連接到遠大洋洲的基因單型 (拷貝),而且近大洋洲的基因單 型 (拷貝) 會呈現星狀結構。比較不同標記中遠大洋洲的與近大洋洲個體在親緣關 係網狀圖中的位置,可發現遠大洋洲與近大洋洲個體來源不完全一致。ITS 的親緣 關係網狀圖顯示近大洋洲的個體分別來自ITS-A 與 ITS-E,而葉綠體片段中所有的 個體都出自cp-3。其中只有屬於 ITS-A 的個體可以直接連到遠大洋洲的基因單型 (ITS-B、ITS-D),其他近大洋洲的個體都與遠大洋洲沒有直接的關係。因此,雖然 無法排除構樹曾自近大洋洲傳播至遠大洋洲,但支持度不高。而且近大洋洲的樣 本數少,族群在兩個分子標記的多樣性也很低,故無法呈現星狀結構。
回顧過去研究發現的東亞與遠大洋洲的基因單型不一致,Seelenfreund (2011)
提出兩點推測:1. 這些變異可能來自她無法取樣的東亞其他地點,如日本或是大 陸東南亞。2. 這些變異亦可能源自介於台灣與遠大洋洲間的其他地區,即島嶼東 南亞和近大洋洲。由於這次的採樣已涵蓋日本及大陸東南亞,但是依舊沒有找到 相關變異,所以遠大洋洲的基因單型來自日本或是大陸東南亞的可能性較低。本 研究雖然在島嶼東南亞與近大洋洲均有樣本,但是數量與分布較為稀疏。尤其是 近大洋洲只有兩個位於高速公路旁的樣點,不但樣本稀少,而且樣本是作為經濟 作物引入近大洋洲的可能性非常高。根據以上結果,我認為無法追溯祖先族群的 遠大洋洲個體其變異較可能發生在島嶼東南亞與近大洋洲,但是若要確認還需要