行政院國家科學委員會專題研究計畫 成果報告
豬泡疹病毒一型基因體重組機制之研究(2/2)
計畫類別: 個別型計畫 計畫編號: NSC91-2320-B-039-009- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 中國醫藥大學微生物學科 計畫主持人: 項千芸 計畫參與人員: 侯庭鏞, 吳世祿 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 92 年 10 月 29 日
中 文 摘 要
在筆者以往的研究中,證實豬 疹病毒一型(suid herpesvirus 1, SHV-1)去 氧核糖核酸分解 (deoxyribonuclease;DNase)具有類似 RecBCD 的切割活性, 同時定義出 SHV-1 DNase 負責酵素活性及 DNA 結合能力的功能區和關鍵胺基 酸,其次也發現 SHV-1 DNA 結合蛋白質(DNA-binding protein, DBP)具有類似 RecA 促進同源 DNA 配對及兩股交換的能力,這些結果顯示 疹病毒基因體的 重組機制可能是以類似於大腸桿菌 RecBCD pathway 的重組機制進行。因此,本 計畫藉由試管內及細胞內重組模式的建立,探究 DBP 及 DNase 在 疹病毒基因 體重組及複製現象上所扮演的角色。本計畫已完成下列三部分實驗:(一)分析 SHV-1 DBP 及 DNase 在重組過程中參與的角色。我們已建立 helix unwinding 及 homologous pairing 的試管內模式,並發現 DBP 確實具有 helix unwinding 及 homologous paring 這兩項功能,此外,這兩項能力會隨濃度增加而上升。在最佳 反應條件的測試下,發現 SHV-1 DBP 進行 homolo gous pairing 的最佳反應條件為 80 mM NaCl、20 mM MgCl2、pH 7.0~7.8,且過程中不需 ATP 的存在。至於 helix
unwinding 方面,SHV-1 DBP 進行 helix unwinding 時需要鎂離子的存在,但不需 NaCl 與 ATP。進一步探討 DBP 對 DNase 功能的影響,發現 DBP 會促進 DNase 核酸外切 的活性,而且 DBP 是否具有結合 DNA 的能力,與是否會影響 DNase 的活性有關。(二)SHV-1 基因體細胞內重組模式的建立及分析。我們已建立細 胞內重組模式,並證實在單純 疹病毒一型及 SHV-1 感染後,可以成功地啟動 重組的進行,顯示 SHV-1 可辨認來自於單純 疹病毒一型的a 序列,誘發重組
的發生。(三)SHV-1 基因體重組複合物的分析。我們已利用 Biotinylated-ELISA 與 yeast two-hybrid system 求證 DNase 與 DBP 之間是否有直接作用。這兩項實驗 都得到正面的結果,顯示 DBP 確實會與 DNase 產生 physical interaction。因此, SHV-1 DBP 與 DNase會互相作用,以影響彼此的作用。在未來,由對SHV-1 DNase 及 DBP 在病毒重組的角色及功能上的了解,可使研究者對 疹病毒的重組現象 有更合理的推論及解釋,進而提供研究者從中尋求控制的方法。
Abstract
The suid herpesvirus 1 (SHV-1) DNase is an alkaline exonuclease and endonuclease, which exhibits an Escherichia coli RecBCD-like catalytic function. The SHV-1 DNA-binding protein (DBP) promotes the renaturation of complementary single strands of DNA, which is an essential function for recombinase. To investigate the functional and physical interactions between SHV-1 DBP and DNase, these proteins were purified to homogeneity. SHV-1 DBP stimulated the DNase activity, especially the exonuclease activity, in a dose-dependent fashion. Acetylation of DBP by acetic anhydride resulted in a loss of DNA-binding ability and a 60% inhibition on the DNase activity, suggested that DNA-binding ability of SHV-1 DBP was required for stimulating the DNase activity. SHV-1 DNase behaved in a processive mode; however, it was converted into a distributive mode in the presence of DBP, implied that SHV-1 DBP stimulated the dissociation of DNase from DNA substrates. The physical interaction between DBP and DNase was further analyzed by enzyme-linked immunosorbent assay, and a significant interaction was observed. Thus, these results suggested that SHV-1 DBP interacted with SHV-1 DNase and regulated the DNase activity in vitro.
目 錄 中文摘要---I 英文摘要---II 前言---1 研究目的---2 文獻探討---3 研究方法---7 結果與討論---10 參考文獻---13 計畫成果自評---19 圖表---20
前 言
豬 疹 病 毒 一 型 ( suid herpesvirus 1, SHV-1 ), 隸 屬 於 疹 病 毒 科 (Herpesviridae)、阿爾伐 疹 病 毒 亞 科 (Alphaherpesvirinae )、水痘病毒屬
(Varicellavirus),為假性狂犬病(pseudorabies)的致病原(Roizman, 1996)。與
SHV-1同亞科的病毒,包括單純 疹病毒一型、二型( herpes simplex virus 1、2)、 帶狀 疹病毒(varicella- zoster virus)、馬 疹病毒一型(equine herpesvirus 1)、 牛 疹病毒一型(bovine herpesvirus 1)等(Roizman, 1996)。這些阿爾伐 疹病 毒之間,在基因的種類、基因在基因體上的排列位置、基因的轉錄調節,以及病 毒DNA的複製機制都極為相似(Ben-Porat et al., 1983),因此筆者以SHV-1為模 式,研究阿爾伐 疹病毒核酸重組與複製現象的機轉及參與的蛋白質。
研 究 目 的
在筆者以往的研究中,證實SHV-1去氧核糖核酸分解 (deoxyribonuclease, DNase)具有類似RecBCD的切割活性( Hsiang et al., 1998),隨後也定義出SHV-1 DNase負責酵素活性及DNA結合能力的功能區和關鍵胺基酸(Ho et al., 2000), 同時也證實SHV-1 DNA結合蛋白質(DNA-binding protein, DBP)具有類似RecA 促進同源DNA配對及兩股交換的能力(Wu et al., 1998)。這些初步的結果顯示發 生在 疹病毒基因體的重組機制,可能是以類似於大腸桿菌RecBCD pathway的 重組機制進行。因此,本計畫擬藉由試管內及細胞內重組模式的建立,探究DBP 及DNase在 疹病毒基因體重組及複製現象上所扮演的角色。
文 獻 探 討
豬 疹 病 毒 一 型 ( suid herpesvirus 1, SHV-1 ), 隸 屬 於 疹 病 毒 科 (Herpesviridae)、阿爾伐 疹 病 毒 亞 科 (Alphaherpesvirinae )、水痘病毒屬
(Varicellavirus),為假性狂犬病(pseudorabies)的致病原(Roizman, 1996)。與
SHV-1同亞科的病毒,包括單純 疹病毒一型、二型( herpes simplex virus 1、2)、 帶狀 疹病毒(varicella- zoster virus)、馬 疹病毒一型(equine herpesvirus 1)、 牛 疹病毒一型(bovine herpesvirus 1)等(Roizman, 1996)。這些阿爾伐 疹病 毒之間,在基因的種類、基因在基因體上的排列位置、基因的轉錄調節,以及病 毒DNA的複製機制都極為相似(Ben-Porat et al., 1983),因此筆者以SHV-1為模 式,研究阿爾伐 疹病毒核酸重組與複製現象的機轉及參與的蛋白質。在筆者以 往的研究中,證實SHV-1去氧核糖核酸分解 (deoxyribonuclease, DNase)具有 類似RecBCD的切割活性(Hsiang et al., 1998),隨後也定義出SHV-1 DNase負責 酵素活性及DNA結合能力的功能區和關鍵胺基酸(Ho et al., 2000)。同時也證實 SHV-1 DNA結合蛋白質(DNA-binding protein, DBP)具有類似RecA促進同源 DNA配對及兩股交換的能力(Wu et al., 1998;Ho et al., unpublished),這些初步
的結果顯示發生在 疹病毒基因體的重組機制,可能是以類似於大腸桿菌 RecBCD pathway的重組機制進行。因此,本計畫擬藉由試管內及細胞內重組模 式的建立,探究DBP及DNase在 疹病毒基因體重組及複製現象上所扮演的角 色。 基因重組是一個生物基本且必要的過程,所有生物體皆需依賴基因重組來 產生遺傳物質的多樣性、修復由外生性或內因性所造成損傷之 DNA 及確保在減 數分裂時同源染色體能做適當分離並維持染色體的完整性。而重組所造成遺傳物 質的交換,不僅發生在動、植物身上,在原核生物、病毒、質體甚至粒線體 DNA 均會發生。基本上,重組可分為非同源性重組及同源性重組,非同源性重組是指 不 同 源 的 DNA 產 生 的 重 組 行 為 , 如 : bacteriophage λ 的 site-specific recombination,為在 lysogenic cycle 時,使噬菌體 DNA 嵌入宿主 DNA 的一個主 要行為(Landy, 1989),或是在真核生物中,當末端 DNA 發生斷裂時,所利用 的 non-homologous end-joining 來修復 DNA(Paques and Haber, 1999)。而同源性 重組,目前在大腸桿菌的 RecBCD pathway 已被明確的證實,因此在探討其他物 種之重組機制時均以此作為模版。例如在真核生物方面,酵母菌與人類的重組蛋 白質已被證明與細菌的重組蛋白質具有同源性,如:酵母菌的 Rad51、人類的 hRad51 與大腸桿菌的 RecA 在 DNA 序列上及功能上具同源性(Aboussekhra et al., 1992;Basile et al., 1992;Shinohara et al., 1992;Ogawa et al., 1995;Park, 1995)。 目前認為,真核細胞之重組的起始,是在 DNA 上造成了一個雙股 DNA 斷 裂(double strand break, DSB)(Orr-Weaver et al., 1981;Resnisk and Martin, 1976) ,而使重組蛋白可以在 DSB 處開始進行重組。DSB 是時常發生的現象, 例如當細胞暴露在 DNA-damaging agents(e.g. ionizing radiation)、複製叉的突然
停止(Paques and Haber, 1999)、或是細胞遭遇到減數分裂欲發生重組時,會有 DSB 的發生。倘若 DSB 不修復,將會造成突變、基因移位及癌症,而同源染色 體的重組即是 DSB 的一個重要的修復方法。DSB 這重要的重組起點在原核生物 的大腸桿菌也是相同的,在 RecBCD enzyme 存在下,發生重組的機率在 DSB 處 高達 99%(Kowalczykowski and Eggleston., 1994;Arnold and Kowalczykowski, 1999),所以當 DSB 形成後,便正式展開重組。重組的機制可簡單的分為三個步 驟:在 presynaptic phase 時,是由 DSB 的產生而開始的。在 DSB 的末端會有核 酸分解 結合在上面,造成 5'→3'的水解而裸露出 3'端單股的 DNA(asymmetric nuclease activity)(Kowalczykowski, 2000)。之後進入 synaptic phase,由一個單 股 DNA 結合蛋白質結合在 3'端單股的 DNA 上,協助將單股 DNA 插入到同源 DNA 處(Kowalczykowski, 2000),形成 D-loop 的結構,即 Holliday junction structure。最後為 postsynaptic phase,Holliday junction structure 是一個四股 DNA 的結構,為重組過程的一個中間產物,可由 resolvasome complex(RuvA、RuvB and RuvC) 進行 branch migration 及 resolution(Hoess et al., 1987;Kitt and Nash, 1987;Nunes-Duby et al., 1987;Dressler and Potter, 1982;Kowalczykowski, 2000), 以完成重組過程。
本計畫主要藉助試管內及細胞內重組模式的建立,尤其是 presynaptic phase 到 synaptic phase 的階段,探討 DNase 及 DBP 參與的角色。以大腸桿菌為例,參 與大腸桿菌重組的因子有χ site、single-stranded DNA-binding protein (SSB)、 RecA、RecBCD、RecE、RecF、RecG、RecJ、RecN、RecO、RecQ 等十餘種, 但對於重組最重要的因子為 RecA、RecBCD、SSB 和χsite。RecBCD 為一個 heterotrimer,由recB、recC、recD 基因的產物組成(Amundsen et al., 2000),是 一個多功能的酵素,具 3'→5'及 5'→3'外核酸分解 活性,可專一的辨認 DSB, 並緊密的結合在 dsDNA 的末端(Roman and Kowalczykowski, 1989a)。RecBCD 為負責重組起始的蛋白質,當辨認到χsite 時,會產生不對稱的核酸分解 活性, 且具單向雙股 DNA 解旋功能( Telander-Muskavitch and Linn, 1981;Smith, 1988; Taylor, 1988)。RecA 為單股 DNA 結合蛋白,具同源 DNA 配對的功能且會促進 兩股的交換並形成 Holliday junction structure(Kowalczykowski and Eggleston, 1994;Roca and Cox, 1997;Bianco and Kowalczykowski, 1999)。在 ATP 存在下, RecA 會結合在單股 DNA 處,使被結合的那股插入到另一雙股 DNA 的同源區, 而產生 D-loop(Flory et al., 1984)。 當 疹病毒基因體經rolling circle模式複製、產生頭尾相連的concatemer的同 時,會發生高頻率的重組現象(Dutch et al., 1992)。基因體重組現象的發生在 疹病毒是相當常見的。在Kintner等(1995)的研究中,他們將兩株不具毒力的單 純 疹病毒一型同時進行角膜接種,之後再自三叉神經節中進行病毒分離,結果 發現74﹪的分離株都是重組株,而Nishiyama等(1991)發現單純 疹病毒一型 的重組株可能會有毒力增強的現象發生。另外在Glazenburg等(1994)的研究中 也發現,SHV-1突變株在接種到羊及豬之後,都有重組現象的發生。當 疹病毒
DNA複製時,其重組機率可增加高達十倍以上(Dutch et al., 1995;Weber et al., 1988),而且若病毒感染的細胞發生DSB,則會刺激重組的產生(Sarisky and Weber,
1994)。 疹病毒基因體發生高頻率重組現象的特性,對於臨床上,尤其是基因 工程減毒疫苗及基因治療病毒載體的使用上,會造成相當大的困擾及困難。因為 經過不同病毒株分子間或是分子內的重組作用,都有可能會發生基因置換的現 象,而使得不具毒力的疫苗株回復其毒力、增加抗原的變異性、或是 疹病毒載 體回復其致病性,而對患者造成危險性。因此,本計畫擬探討 疹病毒基因體重 組現象的機制、發生的時序性及參與的蛋白質,以便由對重組作用的了解,從中 尋求控制的方法。 在筆者以往的研究中證實,SHV-1 DNase 具有內核酸分解 、5'外核酸分解 及 3'外核酸分解 的活性,單股及雙股 DNA 都可以作為受質(Hsiang et al., 1998),此外利用一連串的系列缺損突變株及定點突變株,也定義出 SHV-1 DNase 負責酵素活性及 DNA 結合能力的功能區和關鍵胺基酸( Ho et al., 2000)。若是將 SHV-1 DNase 與其它核酸分解 的受質及切割模式作一比較,可以發現 SHV-1 DNase 的切割活性與大腸桿菌的 RecBCD 及Neurospora crassa endo-exonuclease 相似,這兩種酵素都已被證實與重組作用有關( Fraser et al., 1989;Smith, 1988)。 大腸桿菌 RecBCD 所主導的重組機制已被研究得很詳細( Taylor et al., 1985),首 先 RecBCD 在接近χsite 時會對雙股 DNA 的一股進行切割及解旋而產生單股
DNA 的尾端,之後 RecA 會與單股 DNA 的尾端結合,形成 D- loop,而進行同源 性 DNA 的交換作用。在筆者的研究中證實 SHV-1 DNase 具有類似 RecBCD 的切 割活性( Hsiang et al., 1998),同時也證實 SHV-1 DNA 結合蛋白質( DNA-binding protein, DBP)具有類似 RecA 促進同源 DNA 配對及兩股交換的能力(Wu et al., 1998;Ho et al., unpublished),這些初步的結果顯示發生在 疹病毒基因體的重 組機制,可能是以類似於大腸桿菌 RecBCD pathway 的重組機制進行,其中 DBP 類似於 RecA,而 DNase 類似於 RecBCD 的角色。雖然如此,可能仍有其它蛋白 質也參與 疹病毒的重組作用,因為單純 疹病毒複製及重組的發生在時序及功 能上都有關連(Dutch et al., 1992),因此負責複製的蛋白質可能也參與重組反 應。所以,更進一步對 SHV-1 基因重組複合體的研究,將有助於了解病毒基因 體重組作用與複製發生的時序性及機制。
單純 疹病毒的 DBP,又名 infected-cell polypeptide 8(ICP8)或單股 DNA 結合蛋白質( single-stranded DNA-binding protein)(Honess and Roizman, 1974), 為 UL29 的基因產物(Weller et al., 1983)。DBP 最早是由 Bayliss 等(1975)以 DNA 親合性色層分析法,自病毒感染的細胞中分離出來。Littler 等(1981)及 Yeo 等(1981)利用胰蛋白 分解法及免疫沉澱法證實 疹病毒(包括單純 疹 病毒、馬 疹病毒、牛 疹病毒等)的 DBP 具有相似的抗原性及胺基酸序列。 由缺損 DBP 基因的單純 疹病毒突變株無法進行病毒 DNA 的複製顯示,DBP 為病毒複製所必須(Littler et al., 1983),此外 DBP 也是單純 疹病毒在 DNA 複 製時,主要的 DNA 結合蛋白,因此可以用它為標的,偵測細胞核內複製區間的
構造(Quinlan et al., 1984)。當單純 疹病毒的 DNA 開始進行複製前,DBP 是 分布在核內被稱為前複製區(prereplication sites)的區域,吸引其他參與 DNA 複製的蛋白,尤其是 DNA 聚合 ,進入到前複製區,並加以組織,為 DNA 複 製作準備( Bush et al., 1991)。此外, DBP 可以藉由刺激 DNA 聚 合 、 helicase-primase complex 及起點結合蛋白質(origin-binding protein)的活性,而 促進病毒 DNA 的複製(Boehmer, 1998;LeGac et al., 1998;Lee and Lehman, 1999)。另外,DBP 也被發現可以降低寡核 酸的黏合溫度及促進兩個互補的單 股 DNA 重新黏合回來,顯示 DBP 可能與單純 疹病毒基因體的重組有關(Dutch and Lehman, 1993)。綜合這些研究指出,單純 疹病毒 DBP 應該類似大腸桿菌 的單股 DNA 結合蛋白質及噬菌體 T4 gene 32 蛋白,為一種典型的單股 DNA 結 合蛋白質(Chase and Williams, 1986)。在生物功能方面,單純 疹病毒 DBP 除 了為病毒複製所必須外,也參與病毒基因表現的調節,使晚期基因的表現量下降 (Chen and Knipe, 1996)。因此 Costa 等(1997)利用這些特性,進行單純 疹 病毒 DBP 缺損株的構築,同時將這種複製缺損株應用在疫苗的使用上。他們發 現,這種缺損株無法進行 DNA 的合成,所以在送到宿主體內後無法產生子代病 毒,但是卻可以表現與野外株等量的晚期蛋白質。進一步經由動物試驗,也證明 這種 DBP 缺損株可以成功地保護小白鼠免於受到單純 疹病毒的攻擊。由這些 結果,可以提供研究者對於單純 疹病毒 DBP 在應用方面的另一種思考方向。 除了單純 疹病毒外,帶狀 疹病毒、馬 疹病毒一型、牛 疹病毒一型
及人類巨細胞病毒等的 DBP 也陸續被證實( Anders et al., 1986; Bell and Whalley, 1993; Davison and Scott, 1986; Hammerschmidt et al., 1988)。筆者在以往的研究 中,藉由 subtractive cDNA 基因庫的建立,定位出 SHV-1 DBP 的位置( Ho et al., 1996a),進一步完成 SHV-1 DBP 的定序、表現及功能分析( Wu et al., 1998)。由 這些 疹病毒及其它物種(包括大腸桿菌、噬菌體 T4、腺病毒)DBP 的胺基酸 序列比較中,可以發現這些 DBP 都含有一段具保留性的鹼性及環狀胺基酸(Wang and Hall, 1990),其中噬菌體 T4 及大腸桿菌的 tryptophan 及 tyrosine、腺病毒的 phenylalanine 及 lysine 為參與 DNA 結合的關鍵胺基酸(Doan et al., 1984; Khamis et al., 1987; Neale and Kitchingman, 1990)。至於 SHV-1 DBP 的功能區及關鍵胺基
酸種類是否也如推測的結果呢?除了參與 疹病毒複製上的角色外,DBP 是否
研 究 方 法
一、分析 SHV-1 DBP 及 DNase 在重組過程中參與的角色 (一)Homologous-pairing assay
將 pUC18 以EcoR Ⅰ切成線形當作受質,取 0.5 μg 的線形 DNA 於 100
℃變性成單股後立刻置於冰上,加入反應緩衝液(20 mM Tris-HCl [pH 7.5], 5 mM DTT,10% Glycerol,0.05 μg/μl BSA)以及 10 pmol 的 DBP,其總 反應體積為 20 μl,混合均勻後,在 37℃下作用 5 分鐘,接著加入 2 μl 的 0.1 μg/μl protease K 於 37℃下作用 10 分鐘,以 stop solution(1.2% SDS, 2% Glycerol,0.05% Bromophenol blue)終止反應,以 1.5%瓊脂糖膠體進行 電泳,經定量分析,其同源性配對能力的計算方式為 ssDNA/dsDNA + ssDNA)。
(二)Helix- unwinding assay
將 M13 forward primer 之 5'端標記32
P 當探針,取 1 pmol 的探針與 0.25 pmol 的 M13 ssDNA 在 TES buffer(10 mM Tris-HCl [pH 8.0],1 mM EDTA, 0.1 M NaCl)下總體積為 20μl,於 65℃反應 10 分鐘,待冷卻至 35℃以下, 使探針與 M13 ssDNA 雜合,接著用 centriflexT M
gel filtration 750 xg 2 分鐘移 除未黏合的探針,儲存於-20℃。將 0.0125 pmol 的雜合 DNA 加入 10 pmo l DBP 與緩衝液(20 mM Tris-HCl [pH 7.5] ,5 mM DTT,10% Glycerol,0.1 μg/ μl BSA),總反應體積為 20 μl,在 37℃下反應 30 分鐘後,再加入 2 μl 的 0.1 μg/μl protease K 於 37℃下作用 10 分鐘,以 stop solution(18 mM EDTA [pH 8.0],1.2 % SDS,6% Glycerol,0.05% Bromophenol blue)終止反應,以 1.0%瓊脂糖膠體進行電泳,經烘乾壓片後分析。
(三)離胺酸(Lysine)化學修飾法
將 0.5 ml 純化的 DBP 與等體積的飽和醋酸鈉(saturated sodium acetate) (15 M)混和,加入 10 μl 的無水醋酸(acetic anhydride)在 4℃反應 30 分 鐘,均分為五管,每管分別再加入第二次不同莫爾比(molar ratio)的無水醋 酸反應 30 分鐘(mol acetic anhydride/mol DBP = 0/1、5/1、10/1、20/1、50/1), 以硝基纖維濾紙結合試驗分析。
(四)硝基纖維濾紙結合試驗(nitrocellulose filter binding assay)
取 10 pmol SHV-1 DBP 野生株或缺損株與 0.05 μg 以 nick translation 標記35
S 的 pUC18 單股線狀 DNA(Koch 1986;Rigby 1977)於 DNA 結合緩 衝液(20 mM Tris-HCl [pH 7.6],1 mM EDTA,125 mM NaCl,1 mM DTT,5 ﹪Glycerol)中,總反應體積為 20 μl,在 25℃作用 20 分鐘,將反應混合物 滴至以 0.5 M KOH 處理過的硝基纖維濾紙上,以 DNA-binding buffer A(10 mM Tris HCl [pH 7.5],1 mM EDTA,150 mM NaCl)沖洗 4 次,待乾後,利 用液態閃爍計數儀測定其 CPM 值。
將 0.5 μg 環形 pUC18 以EcoRⅠ切割成線型 pUC18 作為內切 與外切
的受質,加入純化的 DNase 與 DBP,在 DNase 緩衝液(50 mM Tris-HCl [pH9.0],2 mM MgCl2,10 mM 2-Mercaptoethanol)下 37℃反應 5 分鐘,其總
反應體積為 10 μl,最後以 40﹪sucrose 與 Bromophenol blue 來終止反應,以 1.2 ﹪瓊脂糖膠體電泳分析。
(六)DNase processive mode 測試 利用標記35
S 的 DNA(hot DNA)與未標記的 DNA(cold DNA)當作 受質參與反應。首先將1 pmol DNase 與 hot DNA反應於 37℃ 2 分鐘,使 DNase 充分站在 DNA 上後,再加入 50 倍的 cold DNA,其總反應體積為 100 μl, 反應數分鐘後,利用 100 μl 的 5﹪TCA(trichloroacetic acid)與 10 μl 的 5 mg/ml BSA 在冰上反應 10 分鐘後,以 12000 xg 10 分鐘將大片段的 DNA 沉 澱下來,而被水解的 DNA 會懸浮於上清液中,此時將上清液取出以液態閃 爍計數儀分析結果。 二、SHV-1 基因體細胞內重組模式的建立及分析 (一)受質的構築 細胞內重組模式所使用之受質為具有a 序列之質體 DNA。其中 a 序列的 選殖方法為在病毒感染下,產生頭尾相連的基因體形式後,再利用聚合 連 鎖反應,增殖a 序列。之後在 a 序列兩端連結 BamHI linker,架接進入 pRD107 中,以形成重組用質體 pRD105。pRD105 的結構見圖一。 (二)細胞內重組模式的建立 細胞內重組模式參考自 Dutch 等(1995)的方法。將上述 pRD105 送入 細胞中,以病毒感染後,抽取細胞內的質體 DNA,再送入E. coli 中,最後將 E. coli 塗抹於 X-gal agar 上,藉由計算藍白菌落的數量,以評估重組的效率。
pRD105 上含有兩個同方向的 a 序列,兩個 a 序列中含有一個完整的 lacZ 基 因作為 reporter,當重組發生時,會產生兩個新的質體形式,其中一個具有lacZ
基因但無複製起點ori,另一個不具 lacZ 基因但具有複製起點 ori(見圖一),
因此在送入E. coli 中,只有後者可以成功的複製,且在 X-gal agar 上出現白
色菌落。所以,當重組發生時,白色菌落數會顯著的增加,而我們可藉由計 算藍白菌落的數量,以評估重組的效率。 (三)重組過程與病毒感染的關連 為了探究病毒感染時是否會增加重組的發生,我們將 pRD105 送入細胞 後,再將細胞感染病毒,最後收集細胞內質體 DNA,利用上述方法計算重組 的發生率。 三、SHV-1 基因體重組複合物的分析 ( 一 ) 生 物 素 標 記 結 合 免 疫 吸 附 試 驗 ( Biotinylated enzyme-linked immunosorbent assay;Biotinylated-ELISA)
將 DNase 透析保存在 PBS 溶液中,取 20 倍 DNase 莫爾數比的生物素 (Sulfo-NHS-biotin)(1 μg/μl)加入 DNase 中,放置在冰上兩小時,以 centricon-10 在 2000 xg 離心 20 分鐘兩次,去除未結合的生物素,保存於 4℃, 以此標記生物素之 DNase 進行 結合免疫吸附試驗。首先將 0.5 μg DBP 附 著在孔上,接著以 100 μl 0.5 ﹪脫脂奶粉滴入孔中在 37℃下反應 30 分鐘, 洗淨,甩乾,保存於-20℃,接著再加入標記生物素之 DNase 與附著在孔上的 DBP 反應,在 37℃下 60 分鐘後,洗淨、甩乾,加入標記過氧化 的抗生物 素(avidin)與生物素結合,以受質 ABTS 予以呈色,15 分鐘後測其波長 405 nm 之吸光值。
(二)Yeast two-hybrid system
將筆者以往所架構之 DBP基因,殖入酵母菌表現載體中,利用 yeast two-hybrid system,探究與DBP結合的蛋白質。這一套系統是利用酵母菌為表 現宿主,使用兩種融合蛋白質,分別是與GAL4 DNA-binding domain融合的X 蛋白質及與GAL4 activation domain融合的Y蛋白質,以報導基因的活化與 否,判定X及Y蛋白質是否具有關連性。因此在這個實驗中,我們擬以DBP 基因為X基因,以SHV-1感染細胞後所建立的基因庫為Y基因,進行基因的搜 尋,以便了解DBP可以與那一些蛋白質作用。
結 果 與 討 論
一、分析 SHV-1 DBP 及 DNase 在重組過程中參與的角色
在E. coli 重組過程中,RecA 會造成雙股 DNA 的不穩定,使局部雙股分開
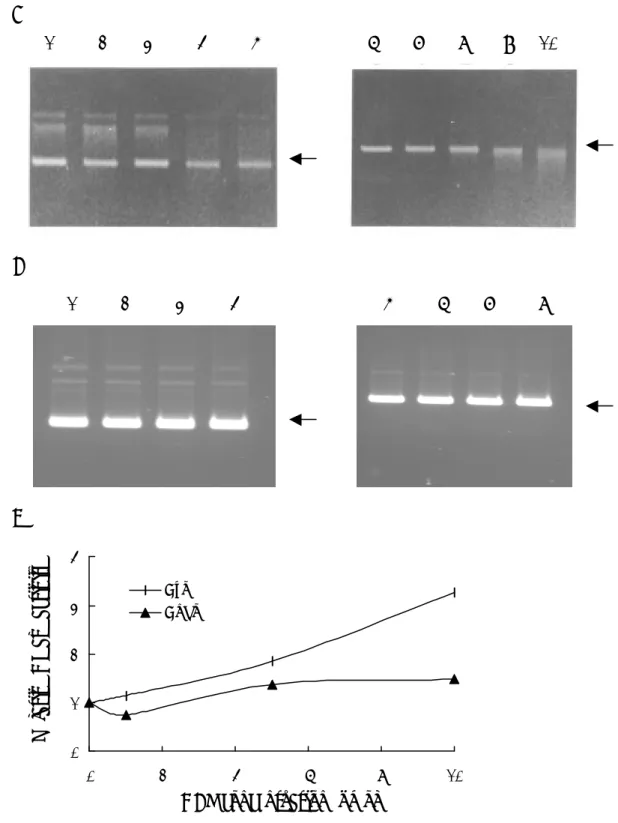
(helix unwinding),而使其他同源單股 DNA 插入進行同源配對(homologous pairing),因此我們建立這兩項重組過程的試管內模式,探討 DBP 是否具有這兩 項功能。圖二顯示 DBP 確實具有 helix unwinding 及 homologous paring 這兩項功 能,此外,這兩項能力會隨濃度增加而上升。在最佳條件的測試下,發現 SHV-1 DBP 進行 homologous pairing 的最佳反應條件為 80 mM NaCl、20 mM MgCl2、pH
7.0~7.8,且過程中不需 ATP 的存在。至於 helix unwinding 方面,SHV-1 DBP 進 行 helix unwinding 時需要鎂離子的存在,但不需 NaCl 與 ATP。
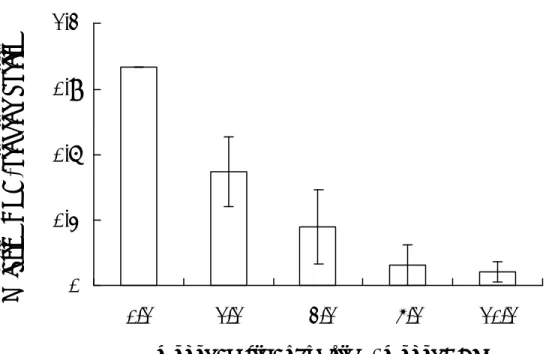
由於 SHV-1 的 DBP 與 DNase 可能參與 SHV-1 的重組過程,本實驗利用不 同濃度之 DBP 探討對 DNase 活性之影響。SHV-1 DNase 具有核酸外切 及核酸 內切 的功能,首先探討 SHV-1 DBP 是否對 DNase 的不同功能上有專一性的影 響,在 DNA 受質方面以 pUC18 超螺旋 DNA 作為核酸內切 功能測試之受質,
EcoRⅠ切成線型的 pUC18 作為核酸外切 功能測試之受質,將 0.5 pmol 的
DNase、0.5 μg 超螺旋或線型 DNA 加入不同濃度的 DBP,在 37℃下反應 5 分 鐘,分別進行核酸內切 與外切 之功能分析。圖三 A 為核酸內切 ,三 B 為 核酸外切 的功能測試,DBP 的量依序為 0、1、5、10 pmol,以瓊脂糖膠體電 泳分析,經定量分析後,以 0.5 pmol DNase水解 DNA的量為 1 倍,結果顯示SHV-1 DBP 對 DNase 的核酸內切 活性並沒有顯著之影響,但對於 DNase 的核酸外切
表現出有促進之影響,活性增加至 3.27 倍。此實驗可明顯證明 SHV-1 DBP 可 專一性的促進 DNase 核酸外切 的活性比內切 高出 2.2 倍,故證實 DBP 會刺 激 DNase 核酸外切 的活性。
為了能顯著證實 DBP 對 DNase 核酸外切 的影響,在預行試驗時,已將 DNase 的量調整為 0.5 pmol 為進行 DNA 水解的閥值以下,表此濃度無法進行水 解。圖四 A 以單獨 0.5 pmol 的 DNase,圖四 B 0.5 pmol DNase 加上 10 pmol DBP, 以 0.5 μg 線型 pUC18 為受質,在 37℃反應 0、1、3、5、10、15 與 20 分鐘後 以電泳膠分析,經定量分析後,結果顯示當只有 DNase 時,水解 DNA 的量未有 所改變,但加入 DBP 後會顯著刺激 DNase 核酸外切 的活性,並證實 DBP 對 DNase 核酸外切 活性的動態影響會隨著反應時間增加而活性不斷上升,且未有 趨於緩和之現象。 SHV-1 DBP 為單股 DNA 結合蛋白質,而 DBP 會促進 DNase 之核酸外切 作用,所以我們進一步研究 DBP 之 DNA 結合功能與 DNase 之核酸外切 活性 有無關連。本項實驗是利用已喪失 DNA 結合功能之 DBP 與 DNase 做核酸外切 活性測試。將 SHV-1 DBP 以 acetic anhydrate 針對離胺酸進行化學修飾,結果 顯示當 DBP 在 5 倍莫爾比的無水醋酸修飾下損失約 90 ﹪的單股 DNA 結合功 能,且已達離胺酸被修飾的極限,故認為離胺酸對 DBP 之 DNA 結合能力有絕
對性的影響(圖五)。將 0.5 pmol DNase、0.05 μg 35
S 標記的線型 DNA、與 10 pmol 野生株 DBP 或已失去 DNA 結合能力之 DBP,於 37℃下作用 15 分鐘,切割下來 的 DNA 片段利用 TCA 沈澱後,經液態閃爍計數儀分析,以單獨 DNase 水解 DNA 反應的量為 1 倍,結果顯示加入野生株 DBP 的 DNase 水解 DNA 反應高達 5 倍, 但以 Lys- modified DBP 與 DNase 反應時其水解 DNA 的量降為 0.5 倍(圖六)。 因此此實驗證明當 DBP 失去 DNA 結合功能時,無法促進 DNase 核酸外切 的 功能,所以 DBP 的 DNA 結合能力對刺激 DNase 的核酸外切 的活性很重要。
進一步研究 DNase 在 DNA 上的移動模式,利用標記 35
S 與未標記的線型 pUC18 DNA 當作受質參與反應。首先將 DNase 與放射線標記的 hot DNA 反應於 37℃ 2 分鐘,使 DNase 充分站在 DNA 上,再加入 50 倍未標記的 cold DNA 做 競爭者,於 37℃反應數分鐘,被切割下來的 DNA 片段經 TCA 沈澱後,取上清 液利用液態閃爍計數儀分析被水解的 hot DNA,當 DNase 為 processive mode 時, 加入 50 倍 cold DNA 並不會改變其產生 CPM 值的速率,若為 nonprocessive mode,加入 50 倍 cold DNA 後 CPM 值速率會下降。在圖七 A 為測試 DNase 的 前進模式,2 分鐘後加入 cold DNA,反應時間為 0、2、5、10、20 分鐘,結果 顯示 cold DNA 加入後並沒有改變其速率,故證實 DNase 為 processive mode,圖 七 B 為 DNase、DBP 與 hot DNA 同時加入,2 分鐘後再加入 cold DNA,反應時 間為 0、2、10、20、40、60 分鐘,結果顯示在加入 50 倍 cold DNA 後 DNase 不 再水解 hot DNA,其水解速率下降,所以證實 DBP 會改變 DNase 的移動模式, 由 processive mode 變為 nonprocessive mode,而此行為會造成 DNase 對受質無法 一次水解完成,產生 DNA 斷端,因此推論此行為會造成高重組頻率的發生。 二、SHV-1 基因體細胞內重組模式的建立及分析
細胞內重組模式參考自 Dutch 等(1995)的方法。將 pRD105 送入 Vero 細 胞中,以 10 m.o.i.的病毒量感染細胞,於 8 小時後抽取細胞內的質體 DNA,再 送入E. coli 中,最後將 E. coli 塗抹於 X-gal agar 上,藉由計算藍白菌落的數量,
以評估重組的效率。pRD105 上含有兩個同方向的a 序列,兩個 a 序列中含有一
個完整的lacZ 基因作為 reporter,當重組發生時,會產生兩個新的質體形式,其
中一個具有 lacZ 基因但無複製起點 ori,另一個不具 lacZ 基因但具有複製起點 ori,因此在送入 E. coli 中,只有後者可以成功的複製,且在 X-gal agar 上出現白
色菌落。所以,當重組發生時,白色菌落數會顯著的增加,而我們可藉由計算藍 白菌落的數量,以評估重組的效率。由表一發現,單純 疹病毒一型如同 Dutch 等(1995)的結果一般,可以顯著地驅動重組的發生,而 SHV-1 也可以成功的 增加重組的機率,顯示 SHV-1 可辨認來自於單純 疹病毒一型的a 序列,誘發 重組的發生。 三、SHV-1 基因體重組複合物的分析 在 證 實 DBP 會 促 進 DNase 核 酸 外 切 的 功 能 後 , 我 們 利 用
Biotinylated-ELISA 與 yeast two- hybrid system 求證 DNase 與 DBP 之間是否有直 接 作 用 。 在 biotinylated-ELISA 方 面 , 首 先 將 Biotin 標 記 在 DNase 上 (biotinylated-DNase),充作為一次抗體,以過氧化 標記的 avidin 作為二次抗 體,來彌補先前 ELISA 過多蛋白質層的缺陷,圖八顯示,DNase 與 DBP 之間會 有直接的 physical interaction,且呈劑量反應作用。在 yeast two- hybrid system 方 面,也證實 DBP 確實會與 DNase 產生 physical interaction。因此,SHV-1 DBP 與 DNase 會互相作用,以影響彼此的作用。
參 考 文 獻
Amundsen, S.K., Taylor, A.F., and Smith, G.R. 2000. The RecD subunit of the
Escherichia coli RecBCD enzyme inhibits RecA loading, homologous
recombination, and DNA repair. Proc. Natl. Acad. Sci. 97: 7399-7404.
Anders, D.G., Irmiere, A., and Gibson, W. 1986. Identification and characterization of a major early cytomegalovirus DNA-binding protein. J. Virol. 58: 253-262.
Arnold, D.A., and Kowalczykowski, S.C. 1999. RecBCD helicase/nuclease. In “Encyclopedia of Life Sciences”, Nature publishing group, London.
Bayliss, G.J., Marsden, H.S., and Hay, J. 1975. Herpes simplex virus proteins: DNA-binding proteins in infected cells and in the virus structure. Virology 68: 124-134.
Bell, C.W., and Whalley, J.M. 1993. Herpesvirus ICP18.5 and DNA-binding protein genes are conserved in equine herpesvirus-1. Virus genes 7: 219-228.
Ben-Porat, T., Veach, R.A., and Ihara, S. 1983. Localization of the regions of homology between the genomes of herpes simplex virus type 1, and pseudorabies virus. Virology 127: 194-204.
Bianco, P.R., and Kowalczykowski, S.C. 1999. RecA protein. In “Encyclopedia of Life Sciences”, Nature publishing group, London.
Boehmer, P.E. 1998. The herpes simplex virus type-1 single-strand DNA-binding protein, ICP8, increases the processivity of the UL9 protein DNA helicase. J. Biol. Chem. 273: 2676-2683.
Boehmer, P.E., and Lehman, I.R. 1993. Herpes simplex virus type 1 ICP8: helix-destabilizing properties. J. Virol. 67: 711-715.
Bush, M., Yager, D.R., Gao, M., Weisshart, K., Marcy, A., Coen, D.M., and Knipe, D.M. 1991. Correct intranuclear localization of herpes simplex virus DNA polymerase requires the viral ICP8 DNA-binding protein. J. Virol. 65: 1082-1089. Chase, J.W., and Williams, K.R. 1986. Single-stranded DNA binding proteins
required for DNA replication. Ann. Rev. Biochem. 55: 103-136.
Chen, Y.M., and Knipe, D.M. 1996. A dominant mutant form of the herpes simplex virus ICP8 protein decreases viral late gene transcription. Virology 221: 281-290. Cooper, J. P., and Hagerman, P. J. 1987. Gel electrophoretic of the geometry of a
DNA four-way junction. J. Mol. Biol. 198: 711-719.
Costa, X.J., Bourne, N, Stanberry, L. R., and Knipe, D.M. 1997. Construction and characterization of a replication-defective herpes simplex virus 2 ICP8 mutant strain and its use in immunization studies in a guinea pig model of genital disease. Virology 232: 1-12.
Davison, A.J., and Scott, J.E. 1986. The complete DNA sequence of varicella- zoster virus. J. Gen. Virol. 67: 1759-1816.
Doan, L.T., Toulme, J.-J., and Helene, C. 1984. Involvement of tryptophyl residues in the binding of model peptides and gene 32 protein from phage T4 to single-stranded DNA. A spectroscopic method for detection of tryptophan in the vicinity of nucleic acid bases. Biochemistry 23: 1202-1210.
Dressler, D., and Potter, H. 1982. Molecular mechanisms in genetic recombination. Ann. Rev. Biochem. 51: 727-761.
Duckett, D.R., Alastair, I., Murchie, H., Diekmann, S., Kitzing, E.V., Kemper, B., and Lilley, D.M.J. 1988. The structure of the Holliday junction, and its resolution. Cell. 55: 79-89.
Dutch R.E., Bianchi V., and Lehman R. 1995. Herpes simplex virus type 1 DNA replication is specifically required for high- frequency homologous recombination between repeated sequences. J. Virol. 69: 3084-3089.
Dutch, R.E., and Lehman, I.R. 1993. Renaturation of complementary DNA strands by herpes simplex virus type 1 ICP8. J. Virol. 67: 6945-6949.
Dutch, R.E., Bruckner, R.C., Mocarski, E.S., and Lehman, I.R. 1992. Herpes simplex virus type 1 recombination: role of DNA replication and viral a sequences. J. Virol. 66: 277-285.
Flory, S.S., Tsang, J., Muniyappa, K., Bianchi, M., Gonda, D., Kahn, R., Azhderian, E., Egner, C., Shaner, S., and Radding, C.M. 1984. Intermediates in homologous pairing promoted by RecA protein and correlations of recombination in vitro and
in vivo. Cold Spring Harbor Symp. Quant. Biol. 49: 513-23.
Fraser, M.J., Hatahet, Z. and Huang, X. 1989. The actions of Neurospora endo-exonuclease on double strand DNAs. J. Biol. Chem. 264: 13093-13101. Glazenburg, K.L., Moormann, R.J.M., Kimman, T.G., Gielkens, A.L.J., and Peeters,
B.P.H. 1994. In vivo recombination of pseudorabies virus strains in mice. Virus Res. 34: 115-126.
Hammerschmidt, W., Conraths, F., Mankertz, J., Pauli, G., Ludwig, H., and Buhk, H.-J. 1988. Conservation of a gene cluster including glycoprotein B in bovine herpesvirus type 2 (BHV-2) and herpes simplex virus type 1 (HSV-1). Virology 165: 388-405.
Ho, T.Y., Wu, S.L., Hsiang, C.H., Chang, T.J., and Hsiang, C.Y. 2000 Identification of a DNA-binding domain and an active-site residue of pseudorabies virus DNase. Biochem J. 346: 441-445.
Ho, T.-Y., Hsiang, C.-Y., and Chang, T. -J., 1996a. Analysis of pseudorabies virus genes by cDNA sequencing. Gene 175: 247-251.
Ho, T.-Y., Hsiang, C.-Y., Wu, K., and Chang, T. -J. 1996b. Rapid screening of pseudorabies virus-specific cDNAs from a cDNA library. J. Virol. Methods 58: 187-192.
Ho, T.-Y., Wu, S.-L., Hsiang, C.-H., and Hsiang, C.-Y. Characterization of pseudorabies virus DNA-binding protein in viral replication and recombination. In preparation.
Hoess, R., Wierzbicki, A., and Abremski, K. 1987. Characterization of intermediates in site-specific recombination. Proc. Natl. Acad. Sci. 84: 6840-6844.
Honess, R.W., and Roizman, B. 1974. Regulation of herpesvirus macromolecular synthesis. I. Cascade regulation of the synthesis of three groups of viral proteins. J. Virol. 14: 8-19.
Hsiang, C.-Y., Ho, T.-Y., Hsiang, C.-H., and Chang, T. -J., 1998. Recombinant pseudorabies virus DNase exhibits a RecBCD- like catalytic function. Biochem. J. 330: 55-59.
Khamis, M.I., Casas-Finet, J.R., Maki, A.H., Murphy, J.B., and Chase, J.W., 1987. Investigation of the role of individual tryptophan residues in the binding of E. coli single-stranded binding protein to single-stranded polynucleotides. J. Biol. Chem. 262: 10938-10945.
Kintner, R.L., Allan, R.W., and Brandt, C.R. 1995. Recombinants are isolated at high frequency following in vivo mixed ocular infection with two avirulent herpes simplex virus type 1 strains. Arch. Virol. 140: 231-244.
Kitt, P.A., and Nash, H.A. 1987. Homology dependent interactions in phage lambda site-specific recombination. Nature 329: 346-348.
Kowalczykowski, S.C. 1991. Biochemical and biological function of Escherichia coli RecA protein: behavior of mutant RecA proteins. Biochemistry 73: 289-304. Kowalczykowski, S.C. 2000. Initiation of generic recombination and
recombination-dependent replication. TIBS 25: 156-164.
Kowalczykowski, S.C., and Eggleston, A.K. 1994. Homologous pairing and DNA strand-exchange proteins. Ann. Rev. Biochem. 63: 991-1043.
Landy, A. 1989. Dynamic, structural, and regulatory aspects of lambda site-specific recombinations. Ann. Rev. Biochem. 58: 913-949
Lee, S.S.K., and Lehman, I.R. 1999. The interaction of herpes simplex type 1 virus origin-binding protein (UL9 protein) with Box I, the high affinity element of the viral origin of DNA replication. J. Biol. Chem. 274: 18613-18617.
LeGac, N.T., Villani, G., and Boehmer, P.E. 1998. Herpes simplex virus type-1 single-stranded DNA-binding protein (ICP8) enhances the ability of the viral DNA helicase-primase to unwind cisplatin- modified DNA. J. Biol. Chem. 273: 13801-13807.
Li, X., Wang, H., and Seeman, N.C. 1997 Direct evidence for Holliday junction crossover isomerization. Biochemistry 36: 4240-4247.
Lilley, D.M.J. 1997. All change at Holliday junction. Proc. Natl. Acad. Sci. 94: 9513-9515
Lin, S.F., Hsu, T.Y., Liu, M.Y., Lin, L.S., Yang, H,L., Chen, J.Y., and Yang, C.S. 1995. Characterization of Epstein- Barr Virus DNase and its interaction with the major DNA binding protein. Virology 208: 712-722.
Littler, E., Purifoy, D., Minson, A., and Powell, K.L., 1983. Herpes simplex virus non-structural proteins. III. Function of the major DNA-binding protein. J. Gen. Virol. 64: 983-995.
Littler, E., Yeo, J., Killington, R.A., Purifoy, D.J.M., and Powell, K.L. 1981. Antigenic and structural conservation of herpesvirus DNA-binding proteins. J. Gen. Virol. 56: 409-419.
Mcllwraith, M.J., Dyck, E.V., Masson, J.Y., Stasiak, A.Z., Stasiak, A., and West, S.C. 2000. Reconstitution of the strand invasion step of double-strand break repair using human Rad51, Rad52, and RPA protein. J. Mol. Biol. 304: 151-164.
Mocarski, E.S., and Roizman, B. 1981. Site-specific inversion sequence of the herpes simplex virus genome: domain and structural features. Proc. Natl. Acad. Sci. 78: 7047-7051.
Namsaraev, E.A., and Berg P. 1998. Branch migration during Rad51-promoted strand exchange proceeds in either direction. Proc. Natl. Acad. Sci. 95: 10477-10481. Neale, G.A.M., and Kitchingman, G.R., 1990. Conserved region 3 of the adenovirus
type 5 DNA-binding protein is important for interaction with single stranded DNA. J. Virol. 64: 630-638.
Nishiyama, Y., Kimura, H., Daikoku, T. 1991. Complementary lethal invasion of the central nervous system by nonneuroinvasive herpes simplex virus types 1 and 2. J. Virol. 65: 4520-4524.
Nunes-Duby, S.E., Matsumoto, L., and Landy, A. 1987. Site-specific recombination intermediates trapped with suicide substrates. Cell 50: 779-788.
Ogawa, T., Shinohara, A., and Ikeya, T. 1995. A species-specific interaction of rad51 and rad52 proteins in eukaryotes. Adv. Biophy. 31: 93-100.
Orr-Weaver, T.L., Szostak, J.W., and Rothstein, R.J. 1981. Yeast transformation: a model system for the study of recombination. Proc. Natl. Acad. Sci. 78: 6354-6358.
Paques, F., and Haber, J.E. 1999. Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 63: 349-404.
radiation in mammalian cells. J. Biol. Chem. 270: 15467-15470.
Quinlan, M.P., Chen, L.B., and Knipe, D.M. 1984. The intranuclear location of a herpes simplex virus DNA-binding protein is determined by the status of viral DNA replication. Cell 36: 857-868.
Resnisk, M.A., and Martin, P. 1976. The repair of double-stranded breaks in the nuclear DNA of Saccharomyces cerevisiae and its genetic control. Mol. Gen. Genet. 143: 119-145.
Roca, A.I., and Cox, M.M. 1997. RecA protein: structure, function, and role in recombinational DNA repair. Prog. Nucleic Acid Res. 56: 129-223.
Roizman, B. 1996. Herpesviridae, p. 2221-2230. In B. N. Fields, D. M. Knipe, P. M. Howley (eds.), Virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia.
Roman, L.J., and Kowalczykowski, S.C. 1989a. Characterization of the helicase activity of the Escherichia coli RecBCD enzyme using a novel helicase assay. Biochemistry 28: 2863-2873.
Smith, G.R. 1988. Homologous recombination in procaryotes. Microbiol. Rev. 52: 1-28.
Taylor, A.F. 1988. In Genetic Recombination. Eds. Kucherlapaati, R. & Smith, G. R., (Am. Soc. Microbiol., Washington), pp. 231-263.
Taylor, A.F., Schultz, D.W., Ponticelli, A.S., and Smith, G.R. 1985. RecBC enzyme nicking at Chi sites during DNA unwinding: location and orientation-dependence of the cutting. Cell 41: 153-163.
Telander-Muskavitch, K., and Linn, S. 1981. In The Enzymes, ed. Boyer, P. C. (Academic, New York), Vol. 14. pp. 233-250.
Thomas, M.S., Gao, M., Knipe, D.M., and Powell, K.L. 1992. Association between the herpes simplex virus major DNA-binding protein and alkaline nuclease. J. Virol. 66: 1152-61.
Wang, Y., and Hall, J., 1990. Characterization of a major DNA-binding domain in the herpes simplex virus type 1 DNA-binding protein (ICP8). J. Virol. 64: 2082-2089. Weber, P.C., Challberg, M.D., Nelson, N.J., Levine, M., and Glorioso, J.C. 1988.
Inversion events in the HSV-1 genome are directly mediated by the viral DNA replication machinery and lack sequence specificity. Cell 54: 369-381.
Weller, S.K., Lee, K.J., Sabourin, D.J., and Schaffer, P.A. 1983. Genetic analysis of temperature-sensitive mutants which define the gene for the major herpes simplex virus type 1 DNA-binding protein. J. Virol. 45: 354.
Wu, S.-L., Hsiang, C.-Y., Ho, T.-Y., and Chang, T-J. 1998. Identification, functional expression, and characterization of the pseudorabies virus DNA-binding protein gene and gene product. Virus Res. 56: 1-9
cross-reactive antigens in the herpesviruses. Virology 108: 256-266.
Zeng, Y. Middeldorp, J. Madjar, J.J., and Ooka, T. 1997. A major DNA binding protein encoded by BALF2 open reading frame of Epstein- Barr virus (EBV) forms a complex with other EBV DNA-binding proteins: DNase, EA-D, and DNA polymerase. Virology. 239: 285-95.
Zheng, S., and Seeman, N.C. 1994. Symmetric Holliday junction crossover isomers. J. Mol. Biol. 238: 658-668.
計 畫 成 果 自 評
本年度計畫進度符合預期,並已完成以下三部份實驗: 一、分析 SHV-1 DBP 及 DNase 在重組過程中參與的角色
我們已建立 helix unwinding 及 homologous pairing 的試管內模式,並發現 DBP 確實具有 helix unwinding 及 homologous paring 這兩項功能,此外,這兩項 能力會隨濃度增加而上升。在最佳反應條件的測試下,發現 SHV-1 DBP 進行 homologous pairing 的最佳反應條件為 80 mM NaCl、20 mM MgCl2、pH 7.0~7.8,
且過程中不需 ATP 的存在。至於 helix unwinding 方面,SHV-1 DBP 進行 helix unwinding 時需要鎂離子的存在,但不需 NaCl 與 ATP。進一步探討 DBP 對 DNase 功能的影響,發現 DBP 會促進 DNase 核酸外切 的活性,而且 DBP 是否具有 結合 DNA 的能力,與是否會影響 DNase 的活性有關。 二、SHV-1 基因體細胞內重組模式的建立及分析 我們已建立細胞內重組模式,並證實在單純 疹病毒一型及 SHV-1 感染 後,可以成功地啟動重組的進行,顯示 SHV-1 可辨認來自於單純 疹病毒一型 的a 序列,誘發重組的發生。 三、SHV-1 基因體重組複合物的分析
我們已利用 Biotinylated-ELISA與 yeast two-hybrid system求證 DNase 與 DBP 之間是否有直接作用。這兩項實驗都得到正面的結果,顯示 DBP 確實會與 DNase 產生 physical interaction。因此,SHV-1 DBP 與 DNase 會互相作用,以影響彼此 的作用。
表 一 細胞內重組模式。 Number of white colony Number of blue colony Number of total colony Recombination efficiency (%)c Controla 0 2 0 0 pRD105 783 17250 18033 4.34 pRD105/HSV-1b 1214 10590 11804 10.30 pRD105/SHV-1b 474 1858 2332 20.33 a Control 為細胞不經任何處理的陰性對照組。 b pRD105/HSV-1、pRD105/SHV-1 表示先將 pRD105 送到細胞後,再以 10 m.o.i. 的病毒量感染。 c Recombination efficiency 為白色菌落數除以總菌落數的百分比。
(A)
(B)
圖 一 細胞內重組的模式圖。(A)pRD105 質體的架構。(B)細胞內重組模式的實 驗流程。
Recombination
No recombination Blue colonies
(A)
(B)
100 13.9 28.6 32.4 40.5 48.2
圖 二 SHV-1 DBP 具有 helix unwinding 及 homologous pairing 的能力。(A) Helix unwinding。將不同濃度的 DBP 與 hybrid DNA 作用 30 分鐘後,以電泳分 析。Lane 1 為 M13 primer,lane 2 為 hybrid,lane 3 為 100℃變性的 hybrid,lanes 4~7 分別為 0、1、5、10 pmol 的 DBP。箭頭部份為 hybrid 及 primer 的位置。(B ) Homologous pairing。將 0.5 μg 的 ssDNA 分別與 0、1、5、10 pmol 的 DBP 混 合,於 37℃作用 5 分鐘後,以電泳分析。Lane 1 代表 dsDNA,lane 2 代表 ssDNA, 箭頭部份為 dsDNA 及 ssDNA 的位置。圖下方的數字代表同源性 DNA 配對的百 分比,計算方式以 ssDNA 除以 dsDNA 與 ssDNA 總量的百分比。
1 2 3 4 5 6 7
← Hybrid
← P r i m e r
dsDNA ssDNA 0 1 5 10 (pmol)
← d s D N A
← s s D N A
100℃
(A)
(B)
(C)
圖 三 SHV-1 DBP對DNase活性的影響。(A)SHV-1 DBP對DNase內切 及外切 活性的影響。Lanes 1~5為利用supercoiled pUC18 dsDNA為受質,lanes 6~10為利 用linear pUC18 dsDNA為受質。SHV-1 DNase (0.5 pmol)與0.5 µg dsDNA和0 (lanes 2, 7)、1 (lanes 3, 8)、5 (lanes 4, 9) 或10 (lanes 5, 10) pmol SHV-1 DBP混合後,於 370C作用5分鐘。(B)SHV-1 DBP內切 及外切 的活性。組別同(A),但反應中 不加入DNase。(C)定量分析。 0 1 2 3 4 0 2 4 6 8 10 DBP concentration (pmol)
Relative DNase activity
Exo Endo
1 2 3 4 5 6 7 8 9 10
(A) (B)
(C)
圖 四 SHV-1 DBP 對 DNase 外切 活性的影響。將 SHV-1 DNase (0.5 pmol)與 0.5 µg linear pUC18 dsDNA 在不含(A)或含有(B) 10 pmol SHV-1 DBP 的情況下, 於 370
C 作用 0 (lane 1)、1 (lane 2)、3 (lane 3)、5 (lane 4)、10 (lane 5)、15 (lane 6) 或 20 (lane 7)分鐘,再以 1.2% agarose gels 分析。(C)定量分析。
0 20 40 60 80 0 5 10 15 20 Time (min)
Relative DNase activity
DNase only DNase + DBP
圖 五 Acetic anhydrate 對 SHV-1 DBP 結合 DNA 能力的影響。將 SHV-1 DBP 與不同莫爾比例的 acetic anhydrate 混合後,於 40 C 作用 30 分鐘,再以硝基纖維 濾紙結合試驗分析。
0
0.3
0.6
0.9
1.2
0/1
1/1
2/1
5/1
10/1
mol of acetic anhydride / mol of DBP
圖 六 SHV-1 DBP DNA 結合能力對 DNase 活性的影響。將 DNase (0.5 pmol)與 50 ng 35S-labeled pUC18 dsDNA 及 10 pmol DBP 、 10 pmol acetic anhydride- modified DBP 或 10 pmol of acetic anhydride-treated E. coli SSB 混合 後,於 370 C 作用 15 分鐘,未切割的 DNA 以 TCA 沉澱後,再測定放射線強度。 0 1 2 3 4 5 6 7
DNase only DBP+DNase Acetic anhydride-treated DBP+DNase Acetic anhydride-treated E. coli SSB
(A)
(B)
圖 七 SHV-1 DBP 對 DNase processivity 的影響。將 SHV-1 DNase (0.75 pmol or 0.5 pmol)與 0.5 µg of 35S-labeled pUC18 dsDNA 在不含(A)或含有(B)10 pmol DBP 的條件下,作用 2 分鐘,再加入過量的 unlabeled pUC18 dsDNA,最後於 370
C 下作用 0、2、10、20、40 或 60 分鐘 (filled squares)。未切割的 DNA 以 TCA 沉 澱後,再測定放射線強度。Open squares 為不加入 unlabeled pUC18 dsDNA 的組 別。 0 100 200 300 400 0 5 10 15 20 25 30 Time (min) cpm (x1000) 0 100 200 300 400 0 5 10 15 20 25 30 Time (min) cpm (x1000)
First layer DNase D B P + + + + + + + + + + + + + + Second layer DNase (ìg) D B P (ìg ) 0 0 0 0 0 0.1 0 0.5 0 1 0 5 0 10 0 0 0 0 0.1 0 0.5 0 1 0 5 0 10 0 Third layer Anti-DNase Ab Anti-DBP Ab + + + + + + + + + + + + + +
圖 八 利用 ELISA 測定 SHV-1 DBP 與 DNase 之間的交互作用。SHV-1 DNase (open bar)或 DBP (close bar)先吸附於 ELISA 平盤上,再加入不同濃度的 DBP 或 DNase。陽性對照組(column 1)為吸附 DNase 或 DBP 的 well 加入相對應的抗體。 陰性對照組為吸附 DNase 或 DBP 的 well 加入對抗 DBP 或 DNase 的抗體。數值 為三次實驗的平均。 0 0.1 0.2 0.3 0.4 1 2 3 4 5 6 7 Absorbance (405 nm)