以 Petri net 技術探討生理時鐘對哺乳類動物機制的影響
陳彥如
a林雅琳
b楊珈芬
b劉郁彧
b陳信志
c*Yan-Ru Chen Ya-Lin Lin Jia-Fen Yang Yu-Yu Liu Austin Chen
a, b ,c
慈濟大學醫學資訊系
a93316145@stmail.tcu.edu.tw
cachen@mail.tcu.edu.tw
*通訊作者:achen@mail.tcu.edu.tw
摘要 近年來發現生理時鐘(Circadian rhythm)是生物中 重要的機制,在一系列細胞中都有表達。它的影響是 全方位(Wide-separated)、多層次的,從組織器官的分 泌運動、基因表達,到對外界刺激的反應等等,均與 生理時鐘存在密切聯繫。 研究指出生理時鐘與糖尿病、肥胖症、癌症有相關 性,故本文目的主要探討哺乳類動物生理時鐘基因 mPer 與癌細胞之間的關係,癌細胞會抑制 mPer 基因 的表現,延長自身的生命週期。就生物實驗中,不易 實作活體實驗,且活體研究中很難控制mPer 的表現量維持穩定,故以Petri net 技術模擬(Simulation),使實
驗可以觀察長時間的週期性表現,呈現穩定狀態。
在本文中模型的確立,是以Petri Net 技術搭配數學微
分方程式,架構出生理時鐘的模型。並模擬各個基因 與蛋白質的交互作用及行為模式,藉由調控正負回饋
機制(positive and passive feedback mechanism),分析模
擬結果,探討對哺乳動物(Mammals)的生理變化有何影 響。本文實驗以調控抑制mPer 的表現量,可發現 PER 的週期明顯拉長。 關鍵字: 生理時鐘、Petri net、模擬、調控、正負回 饋機制、哺乳動物 1. 前言 所有生物都有生理時鐘,各種生理時鐘的週期長短 不同,如心跳、呼吸、昆蟲的翅膀振動,動物的週期 性發情,都有週期性的變化,但最普遍的一種是週期 約廿四小時的「約日」(大概一日)生理時鐘[9]。視 神經交叉上核SCN(suprachiasmatic nuclei)是哺乳動 物的核心時間中樞,是生理時鐘的調整中心[16]。 首先研究的非哺乳類動物為果蠅,因為果蠅的染色 體已都被徹底研究、可大量繁殖、生活史短易研究[8], 第一個哺乳動物的生理時鐘基因由 Takahashi 複製出 來,其名稱為mClock(字首大寫且斜體表示基因,標示 m 代表哺乳類),Takahashi 等科學家所尋找的是能「打 開」生理週期的開關,尤其要找小鼠體內與mClock 基 因所產生的CLOCK(全大寫代表蛋白質)蛋白質結合的 物質。已經發現另外一個產物叫做BMAL 蛋白,兩個 產物可結合成一複合體,這個結合物可開啟小鼠的 mPer、mCry 基因[4,7]。 利用生理時鐘的特性,本文以 Petri net 架構哺乳動 物生理時鐘的模型,可看出生理時鐘呈現約二十四小 時週期性的變化,藉由調控正負回饋的機制,顯示出 異常的生理時鐘產生不同的執行結果,探討對哺乳動 物生理行為上的影響。
Petri net 亦稱斐氏網或派翠網路,起源於 1962 年 C.A. Petri 的博士論文中[13],其概念早期應用於自動裝置 的領域,Petri net 是一種動態模擬的圖形工具,並且 具有分析的特性,利用數學工具模擬與模組系統,可 以藉此驗證系統是否正確合理。在處理同步、非同步 系統、並行系統更加容易,此外,隨著需求不同,延 伸出具有不同特性的Petri net,以其特性加強系統能
力,例如time Petri net [14]、colored Petri net[10]。 Masao Nagasaki 與其研究團隊提出了新概念的 Petri net,稱為「混和功能性派翠網路」(hybrid functional Petri net;HFPN)[11]和擴充版本的「延展型混和功能性派 翠網路」(hybrid functional Petri net with extension;
HFPNe)。此兩種方法擴充與結合了過去發展地許多不 同型態的派翠網路,使此新型的派翠網路更適合用來 處理生物相關的問題。目前已有許多應用例子,如線性 吸收算子的基因規則、果蠅的生理週期、Fas 體引發的 細胞程式化死亡[11]等的模型建製。由於Petri net 對 推論過程有適當的表示方法,使的越來越受到注意。 2. 研究方法 2.1 流程圖 Figure 1.為本文研究流程圖。首先蒐集與生理時鐘 機制相關文獻,了解目前已知生物時鐘背景知識,相
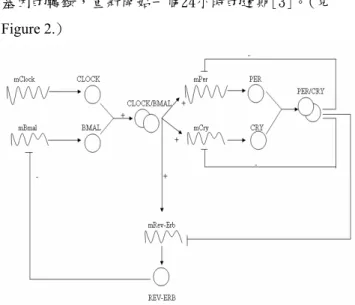
關進展及其背後深遠的醫療意義。就像其他生物學的 問題一樣,我們需要模型系統來提示其運作機制。因 此,我們使用HFPN 用來處理生物相關的問題。從文 獻中找尋可驗證的動力式、數據以便架構出欲實作的 生理時鐘模型。藉由模型系統的模擬結果,我們可以 清楚的看出生理時鐘呈現約 24 小時左右的生理性周期 變化。從最新的研究得知,基因mPer 與癌症發生率有 正相關[15,17]。細胞是透過分裂來增生的,當新細胞 增生的數目足夠之後,會用來取代舊細胞或是癒合傷 口,此時控制細胞週期的蛋白質就會被關掉,細胞週 期也跟著停止。但癌細胞的這種機制出了問題,因此 細胞會繼續分裂下去[5]。我們藉由調控抑制mPer 基 因的閾值以探討生理週期的異常變化。 Figure 1. 研究方向流程圖 2.2 哺乳動物生理時鐘調控機制 蛋白質CLOCK與BMAL結合會趨動時鐘基因mPer 和mCry,於是轉譯出來的PER和CRY蛋白質的量在白 天就會持續增加。通常在黃昏之前,PER和CRY的量 到達某一個程度,兩個蛋白便會結合在一起進入細胞 核,於是抑制了CLOCK/BMAL的作用(負回饋作用)。 此抑制作用便會對日間所增加的PER與CRY進行減量 的工作,而當PER與CRY這兩個蛋白濃度開始下降, CLOCK/BMAL受到的抑制也漸漸減少。當早晨來臨 時,CLOCK/BMAL又再度恢復活性,促進mPer和mCry 基因的轉錄,重新開始一個24小時的週期[3]。(見 Figure 2.) Figure 2. 哺乳動物的生理時鐘機制 3.模型建構 3.1 工具介紹
Cell Illustrator3.0 是建構在 Petri net 的理論上,是一 套細胞圖解器軟體。這款軟體可以將組成大型複雜網 路生物化學過程以視覺化的方式展現給生物化學家。 這套軟體提供了理解這些複雜網路和具有結構上複雜 動 態 聯 繫 的 化 學 組 件 與 過 程 的 關 係 [12] 。Cell Illustrator3.0 可以協助研究人員解決新陳代謝路徑模 型建製,信號轉換分析,基因調整路徑,各種各樣生 物實體例如普通 DNA、mRNA 和蛋白質的動態聯繫等 問題。
在Petri net 中,主要有四個元素,Place 與 Transition 可分為連續式(continuous)、離散式(discrete)與一 般式(generic),Arc 也分為三種分別為正常弧(normal arc)、抑制弧(inhibitory arc)與測試弧(test arc)(見 table 1)。
Place(站區):是一種狀態。可用來表示一個蛋白質、 基因、代謝物或其他單一元素。Transition(渡閘)是 由一個狀態轉成另一個狀態的過程。用來表示反應、 結合、分離、轉錄、轉譯或其他生物化學反應。Arc (弧)用來連結所有這些元件使其形成一個網路架 構,附有加權(weight)且具有方向性。Arc 主要是連接 Place 及 Transition 的一個有方向的線段,可以從 Place 指向Transition 或從 Transition 指向 Place,但 Place 與 Place 或 Transition 與 Transition 之間不能有 Arc 相連, 因為它的狀態不能直接變成另一個狀態,中間一定要 經過一個過程,而過程與過程之間也會存在著一種狀 態。Token(標記)指的就是一個物件,可以代表任何 東西,每個Place 或 Transition 中可以有零個或多個 Token,當 Token 足夠時才能夠由一個狀態變成另一個 狀態[6]。
由Figure 3.及 Figure 4.可清楚知道 Petri net 觸發過 程。在Figure 3.的 e1 初值設為 10,e2 初值為 0,p1 的意思為每2 秒跑 2 給 e2,c1 的 threshold 代表當 e1
的值小於2 時,觸發結束。
Figure 3. Petri net 觸發過程(觸發前)
經過8 秒後,在 Figure 4.中,可看到 e1 值變為 2, e2 的值為 8。
Figure 4. Petri net 觸發過程(觸發後)
3.2 架構模型
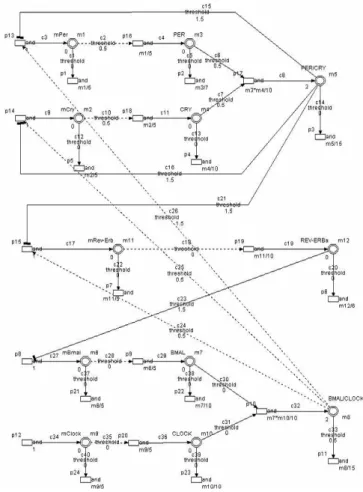
在本文建構的模型中,mPer 和 mCry 基因和其蛋白 質 構 成 主 要 第 一 單 日 負 回 饋 迴 路 , 第 二 迴 路 是 由 mClock 和 mBmal 基因及其蛋白質所組成。連接這兩個
迴路的主要因素,包含 mRev-Erb 基因及其蛋白質,
PER/CRY 複合物抑制 mPer、mCry 和 mRev-Erb 基因 的產量,REV-REBα 蛋白質則抑制 mBmal 基因的表 現,而BMAL/CLOCK 對 mPer、mCry 和 mRev-Erb 有 正回饋作用 [1]。(見 Figure 5.) Figure 5. 使用 HFPN 架構哺乳動物的約日基因調控機 制的模型 透過數學模型,可在電腦中建立各種虛擬實驗,本 文以下列十二個常微分方程建構出哺乳動物生理時鐘 的模型: mPer k mPer k CLOCK BMAL k dt dmPer d t d 1 1 12 / − − = (1)
PER k PER k mPer k dt dPER t d t 2 2 1 − − = (2) mCry k mCry k CLOCK BMAL k dt dmCry d t d 3 3 12 / − − = (3)
CRY k CRY k mCry k dt dCRY t d t 4 4 3 − − = (4) CRY PER k CRY k PER k dt CRY dPER d t t / / 5 4 2 + − = (5)
mRev-Erb k CLOCK BMAL k dt dmRev-Erb t d 6 12 / − =
mRev-Erb k6d −
(6) REV-ERB k mRev-Erb k dt dREV-ERB t t 7 6 − = (7)
mBmal k mBmal k dt dmBmal d t 8 8 1− − = (8)
4. 結果與討論 4.1 正常型哺乳動物的生理時鐘機制 BMAL k BMAL k mBmal k dt dBMAL t d t 9 9 8 − − = (9)
使用 Petri net 技 mClock k mClock k dt dmClock t d 10 10 1− − =
(10)
CLOCK k CLOCK k mClock k dt dCLOCK t d t 11 11 10 − − = (11)
術模擬結果看來,哺乳動物的生理 時鐘機制其中包含的mPer 和 mCry 基因的濃度表現量 約 24 小時的變化,但 mPer、 mCry 被送入細胞質的速率不一樣,故轉譯成的各蛋白 質表現量也因此不同。在mPer 和 mCry 的表現量達到 最高峰之後,形成的蛋白質 PER 與 CRY 的表現量也 達到高峰,週期為約24 小時,可得知基因比蛋白質提
早達到高峰。PER 和 CRY 形成的複合蛋白 PER/CRY,
也有約24 小時週期性的變化,在 PER 和 CRY 到達高 峰準備要下降時,PER/CRY 表現量也達到高峰,由此 可知PER/CRY 對 mPer、mCry 造成負回饋作用,所以 當PER/CRY 表現量最低的下一刻,mPer、mCry 表現 量最高,換句話說,mPer,mCry 的表現量剛開始下降 時,PER/CRY 的表現量逐漸上升。(見 Figure 7.) 和週期完全相同,呈現 CLOCK k BMAL k dt CLOCK dBMAL t t 11 9 / = + CLOCK BMAL k12d / − (12) 每個微分方程式代表此物質的單位時間內濃度的變化 量,方程式的變數與參數皆與此物質有關聯性,負號 代表此物質濃度輸出至另一個物質;正號表示外來物 質濃度輸入至此物質內。1 代表濃度固定量輸入此物 質。 Table 2.參數值 Figure 7. 正常型哺乳動物的生理時鐘機制,第一單日 負回饋迴路的模擬結果。mPer 與 mCry 基因及其蛋白 質之間的關係。橫軸為時間(Hr),縱軸為濃度(nM) 日迴路中的PER/CRY 會抑制 mRev-Erb 的表 第一單 現量,mRev-Erb 與 REV-ERBα 顯示出約 24 小時的週 期變化,REV-ERBα 會抑制第二迴路中的 mBmal 的表 現量,轉譯成的BMAL 在 mRev-Erb 形成的第一個高 峰之後,即呈現穩定狀態,且無週期變化。CLOCK 自 行成穩定狀態,表現量與BMAL 完全相同,且無週期,
BMAL 和 CLOCK 合成的複合蛋白質 BMAL/CLOCK,
在達到一定量時也逐漸形成平穩狀態。(見Figure 8.)
藉由其他文獻參考其動力式及參數值[1,2],並修正某 些數據,建構出本文的哺乳動物生理時鐘模型。
我們將第一單日負回饋迴路中抑制 mPer 基因的閾 值調控為0,抑制 mPer 的產量,使 mPer 及其蛋白無 法表現,結果顯示PER,PER/CRY 表現量皆為 0,但 大約在 369(Hr),產生些微表現量,第二次變化在 738(Hr)發生,由此可知即使將第一單日負回饋迴路中 抑制mPer 基因的閾值調控為 0,因為 BMAL/CLOCK 對mPer 有正回饋作用,所以經過一段長時間交互反應 下,mPer、PER、PER/CRY 仍然有些許表現量產生, 隨後下降為 0,並有週期性。BMAL 呈現週期性的變 化,因PER/CRY 量為 0,則無法抑制 REV-ERBα 的產 量,而REV-ERBα 可抑表現制 BMAL 的形成,所以當 REV-ERBα 表現量最低時,BMAL 表現量達到高峰, 隨著REV-ERBα 表現量上升時,BMAL 產量逐漸下降 為0。CRY、CLOCK、BMAL/CLOCK、REV-ERBα 呈 現週期性變化。(見Figure 10.) Figure 8. 正常型哺乳動物的生理時鐘機制,第二約日 負回饋迴路與mRev-Erb 及其蛋白質的模擬結果 REV-ERBα 是連接第一約日迴路與第二約日迴路 之間的橋樑。被第一約日迴路中的PER/CRY 所調控, 同時也調控著第二迴路中的 BMAL。第一約日迴路中 的 PER/CRY 會 抑 制 REV-ERBα 的 表 現 量 , 而 REV-ERBα 進而會抑制第二約日迴路中的 BMAL 的形 成 。BMAL 和 CLOCK 合 成 的 複 合 蛋 白 質
BMAL/CLOCK,對第一迴路中的 mPer 和 mCry 產生
正回饋作用,同時也促進 mRev-Erb 的生成。PER、 CRY、PER/CRY、REV-ERBα 顯示約 24 小時循環的週 期性,BMAL 和 CLOCK 表現量一致,穩定且無週期, BMAL/CLOCK 達到一定量時逐漸穩定。(見 Figure 9.) Figure 9. PER、CRY、PER/CRY、BMAL、 BMAL/CLOCK、REV-ERBα 蛋白質的模擬結果 4.2 異常型哺乳動物的生理時鐘機制 Figure 10. 異常型哺乳動物的生理時鐘機制,PER、 CRY、PER/CRY、BMAL、BMAL/CLOCK、REV-ERBα 蛋白質的模擬結果 4.3 正常型與異常型的比較 異常型與正常型哺乳動物的生理時鐘機制的差異行 為:將第一約日負回饋迴路中抑制mPer 基因的閾值調 控為 0 (正常閾值為 1.5) ,可抑制mPer 的產量,結 果顯示,PER、PER/CRY 週期拉長,約 369 小時為一 個單位循環,正常值約為24 小時循環,受 mPer 異常 的影響,所有蛋白皆呈現非二十四小時週期性的變 化,而BMAL、CLOCK 和 BMAL/CLOCK 不再有穩 定狀態。
因此,由上述結果可驗證,癌細胞可藉由抑制mPer 的表現,將PER,PER/CRY 的週期拉長,進而延長自 身的細胞週期,延遲細胞凋亡時間,使癌細胞不易死 亡,易於擴散、感染至其他正常細胞。 5. 結論 本文主要針對哺乳類動物進行探討,哺乳類與鳥 類、昆蟲、真菌以及微小細菌的約日系統的歧異性很 大,但其根本機制在生命世界卻很相似,約日系統可 說是無所不在,在醫療行為上的應用不僅有助於相關 藥物的開發,還有助於臨床疾病的預防、診斷和治療。 透過約日節律的瞭解,可讓我們抓準時機採取預防措 施、進行研究治療,將來有可能會利用生理時鐘對疾 病提供有意義的參考。 致謝 本論文承蒙國科會NSC 95-2221-E-320-001 計畫的支 持、贊助,特此致謝。 參考文獻
[1] Becker-Weimann1, S., Wolf, J., Kramer, A., erzel1, H., “A Model of The Mammalian Circadian
Oscillator including REV-ERBα Module”,Genome Informatics, 15(1): 3-12, 2004.
[2] Becker-Weimann1, S., et al., ”Modeling Feedback Loops of the Mammalian Circadian Oscillator”,
Biophysical Journal ,87:3023-3034, 2004.
[3] Bell-Pedersen, D., et al., “Circadian Rhythms from Multiple Oscillators: Lessons from diverse organisms”, Nature Reviews Drug Discovery,doi: 10.1038/nrd1633, 2005.
[4] Darlington, T.K., et al., ”Closing the Circadian Loop: CLOCK-Induced Transcription of Its Own Inhibitors per and tim”,Science, 280: 1599-1603
,
1998.
[5] Foster, R.G., Kreitaman, L., “Rhythms of Life: The Biological Clocks that Control the Daily Lives of Every Living Thing”, ISBN: 1-86197-235-0, 2003. [6] Kao, H.P., Hsieh, B., Yeh, Y.C., “A Petri-net based
approach for scheduling and rescheduling
resource-constrained multiple projects”,Journal of the Chinese Institute of Industrial Engineers, 23:
34-47, 2006.
[7] Ko, C. H., Takahashi1, J.S., “Molecular
components of the mammalian circadian clock”,
Human Molecular Genetics, 15: R271-277, 2006.
[8] Konopka, R., Benzer, S., “Clock Mutants of Drosophila melanogaster”, Proc. Nat. Acad. Sci.
USA, 68: 2112-2116, 1971.
[9] Kramer, A., et al., “Regulation of Daily Locomotor Activity and Sleep by Hypothalamic EGF Receptor Signaling”, Science, 294: 2511–2515, 2001.
[10] Lee, T.E., Wu, J.S., Member, Lin, C.H., “A Colored Petri-Net Model for Load-Transfer in MRT Power Systems”, Volume: 4, on page(s): 6 pp. Vol.4-,
2003.
[11] Matsuno, H., et al, “Biopathways Representation and Simulation on Hybrid Functional Petri Net, In
Silico Biology, 3(3):389-404,2003
[12] Matsuno, H., Chen, L., Miyano, S., “Petri Net Based Descriptions for Systematic Understanding of Biological Pathways”, IEICE, E89-A: 3166-3174, 2006.
[13] Petri, C.A., “Kommunikation mit Automaten”, Bonn: Institut fur Instrumentelle. Mathematik, Schriften des IIM Nr.2, 1962.
[14] Review, B., “A Time Petri Net-based Approach for Hard Real-Time Systems Scheduling Considering Dynamic Voltage Scaling, Overheads, Precedence and Exclusion Relations”.
[15] Sriram, K., Bernot, G., Képès, F., “Discrete Delay Model for the Mammalian Circadian Clock”,
Complexus,3: 185-199
,
2006[16] Stephan, F.K., Zucker, I., “Circadian Rhythms in Drinking Behavior and Locomotor Activity of Rats Are Eliminated by Hypothalamic Lesions”, Proc. Nat. Acad. Sci. USA, 69: 1583-1586, 1972. [17] Walisser, J.A., Bradfield, C.A.,“A Time to Divide:
Does the Circadian Clock Control Cell Cycle”, Cell, 10: 539-540, 2006.