大黃酸誘導人類子宮頸癌細胞Ca Ski細胞週期停滯及細胞凋亡作用之探討; Rhein Induced Cell Cycle Arrest and Apoptosis in Human Cervical Epidermoid Carcinoma Ca Ski Cell Line
138
0
0
全文
(2) 細 胞 膜 內 層 的 phosphatidylserine ( PS ) 的 外 翻 以 及 一 些 cycsteine protease (caspase)的活化。 近年來,有越來越多研究指出大黃酸 rhein 對於腫瘤抑制上的功效,但對於子 宮頸癌的研究上尚未有更進一步的探討。本實驗探討大黃酸對人類子宮頸癌 Ca Ski 細胞株生長之影響,一方面探討其對細胞週期的影響,及對細胞週期相關作用;另 一方面,亦探討大黃酸 rhein 對人類子宮頸 Ca Ski 癌細胞凋亡作用之探討及其凋亡 路徑為何。. 第一節. 實驗架構. 本論文將以細胞模式來探討,觀察大黃酸 rhein 對人類子宮頸癌上皮細胞株(Ca Ski)生長之影響,並探討大黃酸是否具有誘導 Ca Ski 細胞凋亡作用及細胞週期的 調控作用。 一、探討大黃酸 rhein 對 Ca Ski 細胞生長之影響 本實驗為將不同濃度之大黃酸 rhein,直接加入 Ca Ski 細胞之培養液中,於 37°C、5% CO2 及充分溼度下,於 6、12、24、48、72 小時後,收集細胞。評估大 黃酸對 Ca Ski 細胞生長或是存活之影響。 二、探討大黃酸 rhein 對 Ca Ski 細胞週期影響及誘導細胞凋亡之機制 經由存活率之實驗可得之大黃酸 rhein 會抑制 Ca Ski 細胞株之生長,並會誘導 細胞進行細胞凋亡作用,故本實驗將以流式細胞儀檢測 Ca Ski 細胞之細胞週期,並 探討大黃酸 rhein 誘導細胞凋亡之機制。. 2.
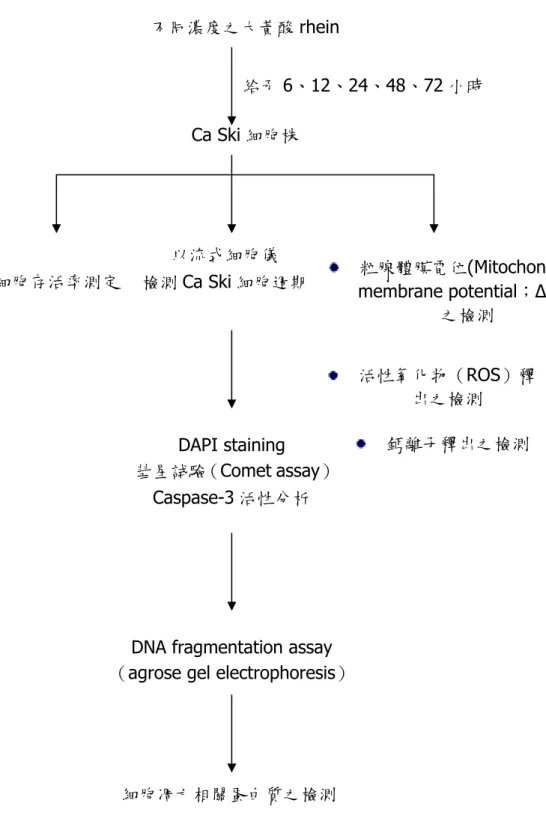
(3) 不同濃度之大黃酸 rhein 給予 6、12、24、48、72 小時 Ca Ski 細胞株. 以流式細胞儀 細胞存活率測定. 檢測 Ca Ski 細胞週期. 粒腺體膜電位(Mitochondria membrane potential;∆Ψm) 之檢測 活性氧化物(ROS)釋 出之檢測. DAPI staining 彗星試驗(Comet assay) Caspase-3 活性分析. 鈣離子釋出之檢測. DNA fragmentation assay (agrose gel electrophoresis). 細胞凋亡相關蛋白質之檢測. 圖 1-1. 大黃酸 rhein 對人類子宮頸癌 Ca Ski 細胞存活率及誘導細凋亡和細胞週期 停滯實驗架構 Figure 1-1. Schematic procedures of rhein on Ca Ski cell viability, cell cycle distribution and apoptosis-induction.. 3.
(4) 第二章 文獻探討 第一節. 大黃酸. 一、大黃酸rhein 的理化特性 (一)化學名 4, 5-dihydroxyanthraquinone-2-carboxylic acid (二)結構式. 圖 2-1. 大黃酸的化學結構式 Figure 2-1. Chemical structure of rhein (三)分子式 C15H8O6 Molecular Weight = 284.23 (四)物理、化學性質 昇華呈黃色粉末,熔點321~322°C。吸收波長為229、258 及435 nm。幾乎 不溶於水,可溶於鹼、吡啶;微溶於乙醇、苯、氯仿、乙醚及石油醚。外型 為紅色的鉀鹽或粉紅色的鈉鹽,和Ca(OH)2及Ba(OH)2也可形成紅色的鹽類 (The Merck Index 13th)。 (五)藥理作用 導瀉利尿作用(Krumbiegel et al., 1993)、抗菌、抑菌作用(Agarwal et al., 2000) 、抗癌作用(Miccadei et al., 1993; Delpino et al., 1992; Lin et al., 2003;. 4.
(5) Wang et al., 2002)。. 二、大黃酸之抗癌作用 大黃酸rhein(4,5-dihydroxyanthraquinone-2-carboxylic acid)是大黃根(Rheum. palmatum L.)中一種蒽醌類(anthraquinone)化合物,大黃根在中國已經廣泛的 使用在當做瀉劑和健胃劑使用,並且在何首烏、决明子等都有大黃酸rhein的存在。 大黃當中含有emodin、emodin-7-methylether(physcion)、aloe-emodin、rhein、 chrysophanol以及一些醣苷類(glucosides)物質。在這些化合物中emodin已經顯 示說具有顯著的抗腫瘤及抗發炎活性;並且於許多研究中已經證實,大黃酸rhein具 有瀉下(Krumbiegel et al., 1993)、抗黴菌(Agarwal et al., 2000)、抗病毒(Barnard. et al., 1992)以及抗癌等作用。近來,研究報導,大黃酸rhein可以藉由降低大鼠肝 臟ATP利用率來抑制某些癌症生長,或者藉由抑制葡萄糖的攝取來抑制Ehrlich腹水 瘤(Ehrlich ascites tumor) 在體內的生長(Castiglione et al., 1993)。1993年, Miccadei等學者研究顯示rhein可以抑制老鼠肝臟腫瘤細胞的生長(Miccadei et al., 1993)以及人類神經膠質瘤細胞的生長(Delpino et al., 1992)。 近來,Lin等人研究報導,藉由小鼠的上皮細胞JB6細胞株體外試驗模式,來探 討rhein癌症化學預防試劑的分子機制。於大黃酸劑量依賴性方法中,顯示出具有抑 制12-O-tetradecanoylphorbol-13-acetate(TPA)誘發的細胞轉型(transformation) 及AP-1(activator protein-1)的活化作用。並且顯示出大黃酸具有效抑制JNK(c-Jun NH2-terminal kinase ) 的 活 性 以 及 c-Jun 蛋 白 的 磷 酸 化 , 但 不 能 抑 制 ERK (signal-regulated protein kinase)和p38 kinase的磷酸化作用,因此這結果說明了 大黃酸抑制AP-1和細胞transformation與抑制JNK和ERK有關,並且是p38非依賴性 的分子機制(Lin et al., 2003)。. 5.
(6) 2002年,Wang等人研究顯示,rhein於囓齒類巨噬細胞株(murine macrophage cell line)RAW 264.7細胞中,具有抑制LPS所誘導的NO(nitric oxide)產生。以LPS (lipopolysaccharide)刺激巨噬細胞iNOS(inducible nitric oxide synthase)活性, 顯示不能有效抑制NO的產生,這結果說明了rhein的抑制作用並非直接抑制iNOS酵 素活性,然而rhein具有降低以LPS誘導的iNOS蛋白表現,但不能抑制COX-2和PGE-2 的產生。由此可知,rhein是有效的iNOS抑制劑,以及可做為有效的抗發炎試劑(Wang. et al., 2002)。 2003年,Lin等人報導rhein會誘導人類原骨髓血癌細胞(HL-60)的凋亡,觀察 到rhein具有誘導caspase的活化、PARP(poly(ADP-ribose)polymerase)的裂解 及DNA fragmentation等典型的凋亡特性產生。而在100 µM rhein、6小時能有效誘 導HL-60細胞凋亡。並且證明了rhein會誘導粒線體膜電位(∆Ψm)的降低、cytochrome. c由粒線體釋出到細胞質以及Bid蛋白的裂解。並且發現rhein也會誘導ROS的產生以 及JNK(c-Jun N-terminal kinase)和p38 kinase的磷酸化。然而,這似乎與rhein誘 導的apoptosis沒有關係,因為透過抗氧化劑NAC(N-acetyl cysteine)、Tiron以及 catalase來阻斷ROS的產生以及JNK和p38 kinase的磷酸化,並不能有效抑制rhein所 誘導之細胞凋亡。因此證明rhein誘導之HL-60細胞凋亡並非是經由ROS依賴性的粒 線體凋亡路徑所致(Lin et al., 2003)。 2004年,Kuo等人,以rhein處理人類肝癌細胞株(Hep G2),結果顯示出rhein 不只抑制Hep G2細胞生長,並且具有誘導細胞凋亡及細胞週期停滯在G1期。由ELISA 分析得到,rhein顯著增加p53和p21的蛋白表現,因此造成細胞週期的停滯。另外, 發現Fas/CD95的表現增加包括mCD95L(mFasL)(membrane-bound CD95 ligand) 以及sCD95L(sFasL)(soluble CD95 ligand),這可能是rhein誘導細胞凋亡的原 因。綜合以上,rhein會藉由增加p53和CD95/CD95 ligand的表現來促使Hep G2細胞 進行apoptosis(Kuo et al., 2004)。由以上可發現,大黃酸rhein對腫瘤的抑制效果,. 6.
(7) 在此,我們以人類子宮頸癌Ca Ski細胞之細胞模式,探討大黃酸rhein對子宮頸癌Ca Ski細胞之誘導細胞凋亡及細胞週期停滯作用機制。. 第二節. 人類子宮頸癌. 根據世界衛生組織的資料統計顯示,子宮頸癌是全球婦女第三常見的癌症,在 台灣根據行政院衛生署的統計,子宮頸癌是台灣婦女最常見的癌症之一,民國八十 九年有近六千名婦女罹患子宮頸癌,導致971名婦女死亡。發生率的排名為女性癌 症的第一位,死亡率的排名卻為第四位。到了九十一年,雖然死亡率降至第五位, 但一千人中還有有高達七人罹患此病,仍是女性癌症罹患的排行榜榜首(衛生署全 國衛生統計資訊網)。全球的排名約在二十幾名,也算得是高發生率的國家之一。 一、子宮頸癌的致病因子 醫學界目前知道,子宮頸癌的病因與人類乳突病毒(Human Papilloma Virus, 簡稱 HPV )有密切關係。而此病毒能干擾正常子宮頸細胞的代謝,增加癌症發生 的機會。人類乳突病毒可因應其基因組合有 100 種以上。不同種的 HPV 會引發不同 的病症。區分為低危險度的 HPV,能導致良性疣的產生。疣是一種皮膚增生所形成 的肉粒,體積大的肉粒呈椰菜花狀,能生長在身體的任何地方,包括生殖器官上。 區分為高危險度的 HPV,可經性接觸而傳播至子宮頸,引致子宮頸細胞變異,造成 癌症的發生。 HPV 感染的主要途徑,與性行為有關。研究顯示,第一次性行為時的年齡愈輕、 性伴侶的數目愈多、或自己的性伴侶有愈多其他性伴侶的婦女,愈容易患上子宮頸 癌。雖然台灣女性的性觀念不如歐美開放,但因為保守的觀念,導致缺乏預防檢查 的實際行動。致使目前罹患子宮頸癌婦女的發生率及死亡率都高居不下。其他可能 發生子宮頸癌的病因包括:. 7.
(8) 1. 年齡:35-60 歲是子宮頸癌的好發年齡。但只要是女性,不論任何年齡皆有可 能發生。 2. 性行為:發生性行為的年齡越早,性行為的次數越多,性行為的對象越複雜,則 發生子宮頸癌的機會就越大。據統計,結婚婦女得子宮頸癌的機會,是單身女子 的二到四倍。 3. 子宮頸炎:子宮頸有破皮、糜爛、發炎的女性較易得子宮頸癌。有人甚至認為子 宮頸炎及子宮頸糜爛,根本就是子宮頸癌的前驅徵候。 4. 家族史:有癌症家族史,甚至有子宮頸癌家族史的女性朋友,得到子宮頸癌的機 會比一般人大。 5. 女性荷爾蒙:有些學者認為黃體素會改變子宮頸上皮細胞的穩定性,而容易發生 不正常的變化,有可能導致子宮頸癌細胞的發展。 二、子宮頸癌的症狀 1. 白帶分泌過多 ,且帶有異味、臭味或混合少許血絲,則應謹慎,並找醫師檢查。 2. 不規則出血、兩次經期間出血、 性交後出血、停經後出血及劇烈運動後出血, 應注意可能已有癌症發生的警訊。 3. 下腹痛及局部疼痛,但也可能是由於單純子宮頸發炎所引起。 4. 子宮頸局部有腫塊,潰瘍或糜爛及組織破壞的現象 。 5. 長久的搔癢,有時亦可能是癌症的早期症狀。 6. 病人體重減輕、胃口不佳或逐漸消瘦。 7. 血尿、小便失禁或便血,這大都是子宮頸癌晚期的症狀。 子宮頸癌的病理癌組織,臨床醫學上依其進行的程度分為: 零期:也稱之為原位癌,癌細胞侵犯在上皮內,尚未到基底膜侵犯真皮組織。 從零期癌到一期,在台灣平均為四年。在此一過程,只要與醫師密切合 作,手術治癒率高達百分之百。. 8.
(9) 一期(I 期):癌細胞已穿過基底膜,但仍侷限在子宮頸內。 二期(II 期):癌細胞已穿過子宮頸,但尚未侵入陰道下三分之一處及骨盆壁。 三期(III 期):癌細胞已侵入陰道下三分之一處及骨盆壁。 四期(IV 期):癌細胞已蔓延到膀胱及直腸的黏膜,甚至發生其他遠隔器官的 轉移。 轉移:子宮頸癌轉移途徑主要是淋巴轉移,及鄰近的淋巴結。此癌亦可直接蔓 延侵犯陰 道,子宮體及子宮頸旁組織。血行轉移多見於晚期,主要轉移 達肺、 肝、骨等處。 表 2-1. 子宮頸癌的臨床分期 (國家衛生研究院,TCOG 子宮頸癌臨床指引,2004) 分期. 子宮頸癌臨床分期. 0. 原位癌. I. 子宮頸癌侷限在子宮. IA1. 微侵襲癌,水平徑不超過7 毫米,子宮頸基質侵襲不超過基底膜下3 毫 米. IA2. 微侵襲癌,水平徑不超過7 毫米,子宮頸基質侵襲為基底膜下3-5 毫 米之間. IB. 肉眼可見腫瘤侷限在子宮頸或顯微病灶範圍超出IA2. IB1. 子宮頸腫瘤直徑不超過4 公分. IB2. 子宮頸最大腫瘤直徑超過4 公分. II. 腫瘤侵襲已達子宮頸外組織,但未達骨盆壁及陰道下端1/3. IIA. 無子宮頸旁組織侵襲. IIB. 已有子宮頸旁組織侵襲. III. 腫瘤侵襲達骨盆壁或達陰道下端1/3 或造成腎臟水腫或無功能腎臟. IIIA. 腫瘤侵襲達陰道下端1/3,未達骨盆壁. IIIB. 腫瘤侵襲達骨盆壁或造成腎臟水腫或無功能腎臟. IVA. 腫瘤侵襲膀胱或直腸之黏膜層,或延展超過真骨盆腔. IVB. 遠處轉移. 9.
(10) 三、醫療與護理 (國家衛生研究院,TCOG 子宮頸癌臨床指引,2004) 子宮頸癌的治療方式有很多種,但在決定最佳的治療方式前,必須考慮很多因 素,包括腫瘤大小,病患年齡及整體健康狀況等。目前醫界最常使用來治療子宮頸 癌的方式,包括外科手術切除、放射線治療及化學治療三種方法。 1. 手術:適用於較早期的癌症,病人的身體狀況良好且無其他合併症時,可開刀將 患部摘除。不過在手術切除時,醫師會考量病患的病情及實際需求,來決定是否 要進行子宮全切除手術,移除整個子宮包括子宮頸,有些也會一併切除卵巢及輸 卵管,還是顧及病患日後生育能力而保留子宮及卵巢。 2. 化學療法:使用藥物來殺死癌細胞,以進行治療的一種方法,通常是癌細胞已轉 移或無法做手術或放射線療法的病人所選擇的治療方法。化學療法可以是靜脈注 射或採取口服的方式,讓藥物可經由血流到達全身,是一種全身性的治療。化學 治療的過程採取週期性的給藥,也就是說,給藥治療後會讓病患有一定的時間進 行恢復,之後才接續下一個療程,至於療程的長短得視病患的病情及治療成果而 定。 3. 放射線治療:利用高能量放射線對癌細胞進行破壞與抑制生長轉移的治療,此方 法適用於 Ib 期及以後各期子宮頸癌病人,並配合內外放療結合治療。放射線治 療屬於局部性的治療,只破壞治療區域的癌細胞。通常醫生所安排治療的時期達 長一個月左右。選擇放射線治療對於早期的子宮頸癌病患來說,其療效與手術治 療不相上下,但若是病患年齡太大或屬晚期的癌症病人,一般醫生會建議以放射 線治療較為可行的方法。 4. 生物製劑療法:生物製劑療法是利用一些物質改變身體免疫系統對抗疾病的一種 療法。通常使用在癌細胞從子宮頸擴散到身體其它部分的情況,最常使用的生物 製劑是干擾素,也常會合併使用化學治療。. 10.
(11) 四、子宮頸癌的預防保健 1. 鼓勵晚婚、少育,婚後婦女,尤其是絕經前後婦女有性交出血或月經異常者,應 及時就醫排除生殖道腫瘤。 2. 凡 30 歲以上的婦女均應定期作子宮頸抹片檢查,早發現、早診斷和早治療。目 前公認子宮頸抹片檢查是最有效預防子宮頸癌發生的最大利器,就台灣的醫療體 系來說,已經由全民健保提供年滿三十歲以上婦女,可享有每年免費接受子宮頸 抹片檢查的福利,因此建議年滿三十歲以上婦女朋友不要放棄接受子宮頸抹片檢 查的機會,要定期接受檢查以保護自己。. 11.
(12) 第三節. 細胞週期(Cell cycle)(Senderowicz, 2004). 細胞開始進行細胞分裂(cell division)到由原來的親代細胞(mother cell)分 裂成兩個子代細胞(daughter cell)時期,稱之為『細胞週期』也稱『細胞分裂週 期』 ,是指一個細胞經生長、分裂而增殖成兩個所經歷的全過程,通常可分為若干階 段,整個週期可分兩各大時期— interphase(即 G1 期、S 期、G2 期)及 Mitosis (M) phase。細胞在 G1 期完成必要的生長和物質準備,在 S 期完成其遺傳物質—染色體 DNA 的複製,在 G2 期進行必要的檢查及修復以保證 DNA 複製的準確性,然後在 M 期完成遺傳物質到子細胞中的均等分配,並使細胞一分為二。 G0 phase︰細胞處於靜止期,此時細胞會停止細胞分裂的進行而去執行一定的生物 功能,這些細胞稱之為靜止期細胞(quiescent cells),經由某些分子的 刺激可進入分化或進入 G1 phase。 G1 phase︰此期細胞開始生長,同時產生 RNA 及合成蛋白質,複製胞器,細胞體積 增加,目的是為 DNA 複製做好準備,此時期染色體數目為 2N。在 G1 phase 進入 S phase 有一個 check point,若細胞走向分裂,則可以進入 S phase 進行 DNA synthesis。 S(synthesis)phase︰此期細胞進行 DNA 合成(DNA synthesis)及複製工作,染 色體數目介於 2N~4N 之間。 G2 phase︰DNA 複製到有絲分裂的期間,此時在細胞核內的的 DNA 含量由 G1 phase 的 2N 變成 4N。而由 G2 phase 進入 S phase 時有第二個 check point, 檢查 DNA 是否完成複製,細胞大小是否生長至合適大小。 Mitosis (M) phase︰細胞行有絲分裂(Mitosis),為細胞週期最短的一期,這階 段的細胞停止生長及蛋白質合成,在 M phase 又可分為五個時期 Prometaphase、Metaphase、Anaphase、Telophase 和 Cytokinesis。而. 12.
(13) 其中 metaphase 為染色體排列於赤道板上,準備進行分裂此時為細胞週 期的第三個 checkpoint(Bruce et al., 2002)。. 圖 2-2. 細胞週期的進行(Secko, D. M., Graphics: Jane Wang) Figure 2-2. Overview of the cell cycle. 當 DNA damage 時,cell cycle 無法通過 check point 而停滯(arrest),此時細 胞會進行 DNA 修復(DNA repaired),一但修補完成時才進入下一期,若是發現無 法彌補的錯誤時,細胞就會選擇走向細胞凋亡(Apoptosis)路徑,以避免錯誤遺傳 至下一代(Smith and Fornace, 1996)。 一、細胞週期調控因子 真核細胞內有一個調控機構,使細胞週期能有條不紊地依次進行。這個調控機 構的關鍵蛋白組成,即細胞週期依賴性蛋白激酶(Cdk)和週期蛋白(cyclin)。 (一)Cyclins and Cyclin-dependent kinases(Cdks) 調 控 細 胞 週 期 進 行 的 蛋 白 稱 之 為 cyclins , 而 調 控 cyclins 活 化 則 是. 13.
(14) Cyclins-dependent kinase(Cdks) ,單獨的 Cdks 並無磷酸化下游分子的能力,這些 Cyclins 與 Cdks 需形成複合物,Cdks 才具有蛋白質激酶的活性,而去磷酸化 (phosphorylation)下游分子,進而調控著 cell cycle 的進行。細胞週期的四個時期 都有特定促使每一時期進行的 Cyclin-Cdks complex;G1 phase 有 Cyclin D-Cdk4/6 complex 及 Cyclin E-Cdk2 complex,S phase 有 Cyclin A-Cdk2 complex,G2/M phase 有 Cyclin B-Cdk1 complex(Sherr, 1996;Sherr, 2000;Ho and Dowdy, 2002)。 (Table2-1) 表 2-2. 調控細胞週期之週期蛋白及其激酶 Table 2-2. Cell cycle regulate of Cyclin and CDKs. Cell cycle stage. Cyclins. CDKs. G0 /G1 phase. Cyclin D. Cdk4, Cdk6. Late G1 phase. Cyclin E. Cdk2. S (synthesis) phase. Cyclin A. Cdk2. G2/M phase. Cyclin B. Cdk1. (二)Cyclins-dependent kinases inhibitors(CdkIs) and p53 Cdk inhibitor 能與 Cdks 結合形成穩定結構而抑制 Cdks 之活性,以抑制 Cdk 之 作用負向調控細胞週期的進行。CdkI 主要可分為兩大家族—CIP/KIP 家族與 INK4 (Inhibitor of Cdk4)家族。CIP/KIP 家族,包括了 p21CIP/WAF1/sdi1、p27KIP1 以及 p57KIP2;INK4 家族則有 p15INK4a、p16INK4b、p18INK4c、p19INK4d。 (Sherr and Robert, 1999)。 CIP/KIP 家 族 中 p21CIP/WAF1/sdi1 、 p27KIP1 此 類 蛋 白 質 可 以 同 時 抑 制 cyclin E/Cdk2、cyclin D/Cdk4 和 cyclin A/Cdk2 的作用,他們會形成 cyclin-Cdk-CdkI 穩定 型式,以遮蔽調 Cdks 的 catalytic site,而不能與下游分子反應,進而抑制細胞週期. 14.
(15) 的進行(Jeffrey et al., 1995)。p57KIP2 則是和細胞的分化較有相關。 INK4 家族,則對 Cdk4 及 Cdk6 具有選擇性的抑制作用,會與 cyclin D 競爭結 合,使 cyclin D 無法活化,而使得細胞週期停滯於 G1 早至中期(McConnell et al., 1999)。 p53 為腫瘤抑制因子,本身為一轉錄因子(transcription factor) ,平時會與 MDM2 protein 結合成為不活化型態,當 DNA 受到傷害時,就會誘使 p53 活化,p53 参與 了細胞修復、細胞週期的停止與細胞凋亡等生物反應。CdkI 當中的 p21 是 p53 主 要下游調控分子,p53 的活化會轉錄 p21,使 p21 蛋白表現量增加,導致細胞週期 的停滯,直到細胞修復完成才使細胞週期正常運行,若 DNA 受損太嚴重時,p53 就 會使細胞進行凋亡(Lundberg et al., 1999)。. p15 p16 p18 p19. p27 p21 p56. 圖 2-3. 細胞週期的調節(John, 1999) Figure 2-3. Regulation of cell cycle. 15.
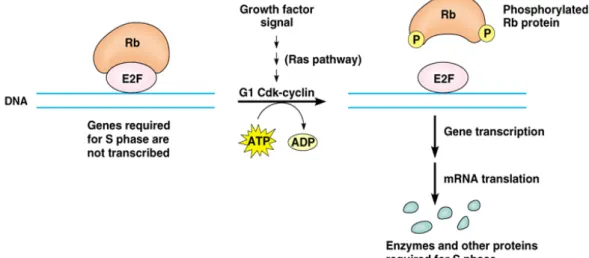
(16) (三)pRb and E2F families pRb(retinoblastoma protein 或 pocket protein)families 有 pRb1/p110、 pRb2/p130 與 p170 三者,E2F(early gene 2 factor)家族則有 E2F-1 至 E2F-6(Dyson, 1998) 。E2F 為轉錄因子(transcription factor) ,平時與 pRb 結合在一起並不具有活 性,當 Rb protein 被上游的 cyclin/Cdk compelex 磷酸化時,就與使 E2F 脫離,使 E2F 具有活性。pRB1 與 E2F-1,-2,-3 結合在一起,pRB2/p130 和 p170 是與 E2F-4,-5 結合在一起,而 E2F-6 則是和 polycomb group proteins(PcG)結合在一起。一般 認為,E2F-4,-5 與 pRB2/p130 以及 E2F-6 與 PcG 的結合是在 G0 與 early G1 期時, 是扮演 transcription repressors 的角色,E2F-1,-2,-3 則作用於 late G1 跟 S 期,為 transcription activators(Stevaux and Dyson, 2002)。. 圖 2-4. Rb protein 與 E2F(Addison, 1999) Figure 2-4. Rb protein and E2F. 16.
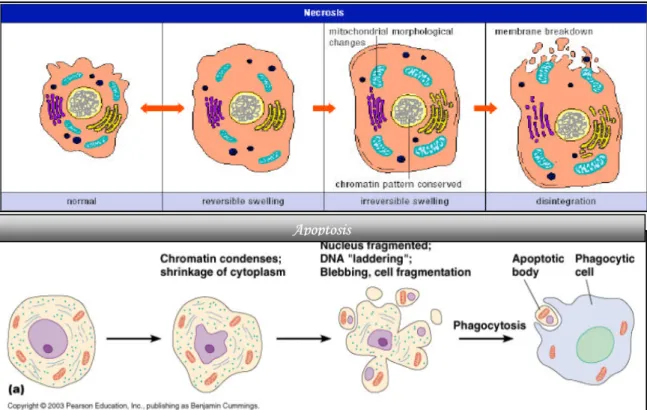
(17) 第四節. 細胞凋亡作用(Apoptosis). 「細胞凋亡」(Apoptosis)又稱為「細胞程式死亡」(programmed cell death) 的正常生理性細胞死亡,一種自然的生命現象;過程中由特定的蛋白質及酵素調控 循序漸進使細胞死亡。正常細胞都擁有感應系統來感應細胞內或外來的訊息,當偵 測到不利其正常生長的訊息時,就引發程式死亡。從生理的觀點上,個體的發生 (development)、生長(growth)、分化(differentation)過程中,細胞凋亡有助 於維持生物體內恆定的功用(Raff et al., 1993) ,若生物體不能正常進行細胞凋亡, 則會引起不正常的增生或導致癌細胞的發生。另一方面,細胞凋亡是細胞對外來病 毒的攻擊或破壞的一種死亡模式,當細胞凋亡失調時,包括不恰當的活化或抑制皆 會導致疾病的發生,像是阿茲海默症(Alzheimer disease)(Behl et al., 2000)、 AIDS、自體免疫疾病等。 一、細胞凋亡的特徵 細胞凋亡的運作,包含著特定的生物化學變化,也與細胞的壞死截然不同。因 此,一些型態學或生化學的特徵,可用來作為探討藥物是否造成細胞凋亡的指標, 例如凋亡過程中會有,細胞型態改變(cellular morphology change)、細胞膜內層 的 phosphatidylserine ( PS ) 的 外 翻 、 細 胞 萎 縮 ( Shrinkage ) 、 DNA 斷 裂 (oligonucleosomal DNA cleavage)、細胞核皺縮(nuclear shrinkage)及染色質 凝結(chromatin condensation),進而細胞膜內陷將細胞分割為多個包膜包覆之 凋亡小體(apoptoic body),再被鄰近的吞噬細胞所吞噬,並無炎症反應產生。故 一般認為誘發細胞進行 apoptosis 是一種較好的抗癌機制,藉此減少藥物可能引起 的副作用。於體外培養因無巨噬細胞,凋亡細胞最終仍會走向壞死(necrosis) (Fadok. et al., 1992)。 活化凋亡相關的蛋白質及酵素,它們有的參與改變粒腺體膜的完整性使粒腺體. 17.
(18) 膜電位(∆Ψm)下降,有的執行分解蛋白質或經特殊調控活化核酸酵素以水解 DNA 使染色體斷裂,可以進行 TUNEL(Terminal deoxynucleotidyltransferase UTP Nick End Labeling)、DAPI staining assay 及 DNA 電泳分析檢測,這些型態學或生化學 的變化上的檢測都是研究 apoptosis 的基礎。此外,細胞凋亡也會啟動一系列的訊 息傳遞串聯(signal transduction cascade)影響各階段的進行,精密地調控程序性 死亡的效果。 二、細胞凋亡與細胞壞死之區別 細胞壞死會同時發生在一群細胞,也可分為早期與晚期兩個時期。在細胞壞死 早期,細胞內之胞器會脹大,細胞膜也會破裂;到了晚期細胞膜也發生破裂(Searle. et al., 1982; Matsuda et al., 1996; Nicotera et al., 1999)。由於細胞發生破裂,因 此會釋放出一些發炎物質而引起局部發炎反應,傷害到鄰近的細胞或組織。. Apoptosis. 圖 2-5. Cell necrosis 與 apoptosis 之比較(Wyllie et al., 1998) Figure 2-5. Compare cell necrosis with apoptosis.. 18.
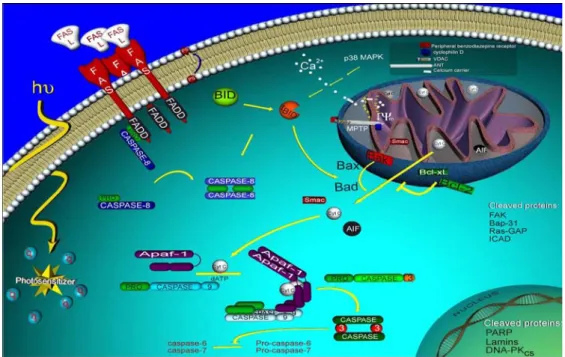
(19) 三、細胞凋亡的調控機轉 (一)內在途徑—粒腺體路徑(Mitochondrial pathway) 引發細胞凋亡最具代表性的粒線體途徑(Parone et al., 2002)。細胞受到細胞 內或細胞外的氧化壓力(stress)如缺氧、放射性傷害及 DNA 損傷時,粒線體膜電 位下降、孔道會打開使 cytochrome c 由粒線體的 intermembrane space 釋放到細胞 質,再與 Apaf-1(Apoptoic protease activating factor 1)、pro-caspase-9 結合成複 合 體 apoptosome , ATP 或 dATP 存 在 下 進 而 活 化 cacsapse-9 , 導 致 下 游 之 caspase-3、6、7 等之活化,最後誘導細胞凋亡發生(Hengartner, 2000)。 1. Bcl-2 families 影響粒線體膜電位最常見的就是 Bcl-2 家族,Bcl-2 家族蛋白在細胞凋亡過程中 扮演一個相當重要的角色,這一群蛋白質可分為促進凋亡分子(pro-apoptoic protein)以及抑制凋亡分子(anti-apoptotic protein)兩大類。 Pro-apoptoic protein 中含有:Bax、Bad、Bid、Bcl-xS、Bak、Bok、Diva、Bik、 Hrk、Nip3、Nix 等。 Anti-apoptoic protein 中含有:Bcl-2、Bcl-xL、Bcl-w、Mcl-1、Nrl3、A1/Bf1-1。 這類蛋白質都含有BH(Bcl-2 homology) domains如BH1、BH2、BH3及BH4, 而促進凋亡蛋白與抑制凋亡蛋白彼此以heterodimers或homodimers的型式結合,調 控細胞凋亡(Jurgensmeier et al., 1998)。Bcl-2存在於粒線體外層,一般抑制凋亡 分子(Bcl-2)平時都存在於粒線體膜上、內質網膜上或核膜上;而促進細胞凋亡分 子(Bax)平時都存在於細胞質當中當收到凋亡訊息後就會轉位(translocation)至 粒線體膜上和促進凋亡分子結合進而影響膜電位的平衡,造成粒線體膜形成孔洞 (pore),粒線體通透性的改變而致使cytocrome c 或AIP( apoptoiis incduced factors)外流至細胞質(Kroemer et al., 1995)。此外,粒線體膜電位下降,粒線. 19.
(20) 體孔洞打開造成大量的ROS聚積,進而造成細胞傷害(Kowaltowski et al., 1996), 另 外 , cytochrome c 釋 放 到 細 胞 質 時 會 與 細 胞 質 中 dATP 、 Apaf-1 結 合 後 活 化 caspase-9,進而引發細胞凋亡。而抗細胞凋亡的Bcl-2家族蛋白,則是直接和Apaf-1 結合,抑制Apaf-1的活性;或經由穩定粒線體膜電位以及其體積之恆定性,以防止 cytochrome c、apoptogenic cofactor之外流,進而保護細胞使其免於步上凋亡之途 (Cosulich et al., 1999)。 2. Caspase enzyme family Apoptosis過程中,會活化一群蛋白水解酶(protease)來執行凋亡之動作。即 caspase(cysteine aspartyl-specific protease)是具有專一性的cysteine protease, 專門切割aspartic acid residues(Alnemri et al., 1996)。Caspase 蛋白系列均具有 相似的氨基酸序列、結構和專一性,都是經非活化態的pro-caspase(約30-50 kDa) 之裂解所致。這些pro-caspase 包含三個主要的部位(domain) :NH2 terminal、large subunit、small subunit。Pro-caspase活化時必須先經由其他caspases 進行一連串 蛋白分解,裂解這些domain,移除pro-domain 和linker region,使得大、小subunit 結合形成複合體,成為活化態的caspase(Nicholson et al., 1997)。Caspase又可 分為initiator caspase和effector caspase,initiator caspase受到外來或細胞內凋亡訊 息傳遞活化後,在進一步活化effector caspase。initiator caspase包含有caspase-2、 8、9、10;而effector casapse包含有caspase-3、6、7。於粒線體路徑中,cyrochrome. c釋出與Apaf-1結合,活化caspase-9,進而活化caspase-3、6、7。結果造成細胞核 膜蛋白(lamin)的分解、PARP(poly(ADP-ribose)polymerase)裂解,PARP具 有修復受損DNA的作用,當caspase-3活化會裂解PARP,則會由116 kDa裂解成85 kDa,而失去修復功能。以及活化的casapse-3會將ICAD(Inhibitor of CAD)切除放 出CAD(caspase-activated Dnase),而造成DNA fragmentation,進而導致細胞凋 亡。. 20.
(21) (二)Apoptosis induction by Ca2+ 一般鈣離子會儲存在內質網(endoplasmic reticulum;ER)中,當死亡訊息或 藥物刺激下,會促使鈣離子從內質網釋放出來,導致細胞內鈣離子恆定(intracellular calcium homostasis)改變(Beyersmann et al., 1997; Shen et al., 2001),近來研 究指出,鈣離子會影響粒線體膜電位的消散,致使 cytochrome c 釋出以及大量 ROS 產生,而導致細胞凋亡(Lemairié et al., 2004;bae et al., 2003)。 (三)Apoptosis induction by ROS 近來研究顯示,活性氧化物(reactive oxide species;ROS)在誘導細胞凋亡過 程中扮演一個重要的角色(Jung et al., 2001)。並且近來許多獻指出,ROS 會由粒 線體所產生,於呼吸鏈反應中產生大量的超氧陰離子(Fleury et al., 2002)。ROS 會造成細胞 DNA 的損傷進而誘導凋亡訊息傳遞。許多刺激像是 TNF-α或抗癌藥物, 皆會刺激細胞產生 ROS 來誘導細胞凋亡(Larrick et al., 1990 and Simizu et al., 1998) 。ROS 的產生會使粒線體的通透性改變,以及導致粒線體的膜電位下降(Xia. et al., 1999),促使粒線體凋亡路徑的發生。. 21.
(22) 圖 2-6. 粒線體凋亡路徑 Figure 2-6. Mitochondrial pathway of apoptosis (2002 Cell Signaling Technology, Inc. www.cellsignal.com) (四)外在途徑—死亡受體路徑(Death receptor pathway) 細胞膜上死亡受體路徑則是經由活化細胞膜上Fas、TNFR-1或TRAIL receptor 等死亡受體與細胞膜上的surface receptor結合,釋放細胞凋亡訊號後開啟。細胞凋. 22.
(23) 亡啟動後會直接活化caspase cascade,而使細胞很快得走向凋亡。 1. Fas/FasL(CD95/CD95L) Fas屬於TNF(tumor necrisis factor) receptor superfamily中type I膜蛋白,而 FasL為TNF receptor superfamily中typeⅡ膜蛋白,當Fas與FasL結合,會活化Fas receptor,並進行trimerization,並吸引FADD(Fas-associated death domain)結合 至receptor 位於細胞質中的domain;另一方面,FADD會進一步吸引多分子的 procaspase-8 形成的DISC(death-inducing signaling complex),此時procaspase-8 會因構形的改變和濃度的提高而活化成caspase-8,具有完整活性的caspase-8 可活 化caspase-3 和caspase-7,而caspase-3會再活化caspase-6,這些effector caspase (包括caspase-3、6、7)會進一步去影響一連串的caspase substrate(Bratton et al., 2000);或者caspase-8的活化促使Bcl-2家族成員Bid的裂解,tBid位移至粒線體, 促 使 粒 線 體 釋 放 出 cytochrome c 接 著 活 化 caspase-9 和 caspase-3 最 後 使 細 胞 apoptotic death(Budihardjo et al., 1999)。. 圖2-7. 經由Fas傳遞的凋亡路徑(Almeida et al., 2004) Figure 2-7. Fas-mediated apoptosis. 23.
(24) 2. TNFR1(tumor necrosis factor receptor 1) TNF是由活化的巨噬細胞或感染反應生成的T細胞所產生的,TNF和細胞膜表面 的TNFR1結合後,會吸引TRADD(TNFR-associated death domain)結合至receptor 膜內的doman,進而活化caspase-8,致使細胞凋亡;另一方面,會吸引RIP(receptor interacting protein)及TRAF2(TNF-associated factor 2)與之結合,活化NF-κB促 使細胞存活(Derradji and Baatout, 2003)。 3. TRAIL(TNF-related apoptosis inducing ligand)(Marsters et al., 1997) TRAIL 所造成的 apoptosis 路徑和 Fas 很相似,唯一跟 Fas 機制不太一樣的是 FasL 的表現要 T cell 與 NK cell 的活化,而 TRAIL 的活化可以藉由許多組織刺激來 表現。 TRAIL 的 receptor 是 DR4 或 DR5,當 TRAIL 與 DR4 或 DR5 結合後會吸引 DR4/DR5 Death domain 結合,進而活化 caspase-8、10,而進行細胞凋亡反應。 而細胞膜上有兩種 receptor 會與 DR4/DR5 競爭 TRAIL 的結合位置,就是 DcR1 與 DcR2,當 TRAIL 與 DcR1 或 DcR2 結合後,則不會啟動細胞凋亡反應。. 24.
(25) 圖 2-8.細胞膜表面受體凋亡路徑 Figure 2-8. Death receptor-mediated apoptosis (2002 Cell Signaling Technology, Inc. www.cellsignal.com). 25.
(26) (五)NF-κB(Nuclear factor kappa B)與細胞凋亡的關係 NF-κB 為 細 胞 核 中 重 要 轉 錄 因 子 , 一 般 位 於 細 胞 質 中 會 與 其 抑 制 劑 IκB (Inhibitor of κB)結合,並不具有活性的,當細胞受 proinflammatory cytokines、 growth factor、oxidative stress 及 ultraviolet light 刺激時,會活化 IκB kinase(IKK) complex,繼而將 IκB 磷酸化,接著會被 E3 ubiquitin ligase 進行 ubiquitination,並 且 IκB 會被 proteasome 降解,而釋出 NF-κB,游離的 NF-κB 即可 translocate 至細 胞核中,影響相關基因轉錄作用。細胞中最主要的 NF-κB 是由 Rel-A(p65) 、NF-κB (p50)形成之 heterodimer(Suh and Rabson, 2004)。受 NF-κB 所調控的相關基 因有,與抑制細胞凋亡有關之 Bcl-xL、Bcl-2(Chiao et al., 2002;Lemarie et al., 2004;Li et al., 1999) ,與細胞生長有關之 IL-6(Zerbini et al., 2004)及 c-myc(Suh and Rabson, 2004)以及 cyclin D1(Albanese et al., 1995)等基因都是受 NF-κB 所 調控。. 26.
(27) 第三章 實驗材料及方法 第一節. 實驗材料. 一、細胞株來源 本實驗所使用之細胞株 Ca Ski 為人類子宮頸癌上皮細胞株(human cervical epidermoid carcinoma) ,由新竹食品工業研究所(Food Industry Research and Development Institute)購得 二、藥品試劑 1. 4,5-dihydroxyanthraquinone-2-carboxylic acid ( Rhein ): 購 自 Sigma Chemical Co. 2. Dimethyl sulfoxide(DMSO):購自 Sigma Chemical Co. 3. RPMI1640 medium:購自 Gibco 4. Fetal bovine serum(胎牛血清, FBS):購自 Gibco 5. L-Glutamin(麩胺酸 , LG):購自 Gibco 6. Penicilllum Streptomycin(PS):購自 Gibco 7. 3,3’-Dihexyloxacarbocyanine iodide (DioC6):購自 Molecular Probes 8. propidium iodide(PI) :購自 Sigma Chemical Co. 9. Trypsin-EDTA:購自 Amersco 10. Trypan blue:購自 Sigma Chemical Co. 11. Disodium hydrogen phosphate(Na2HPO4):購自 Merck 12. Sodium chloride(NaCl):購自 Merck 13. Potassium dihydrogen phosphate(KH2PO4) :購自 Merck. 27.
(28) 14. Potassium chloride(KCl):購自 Merck 15. Propidium iodide(PI) :購自 Sigma Chemical Co. 16. PhiPhiLux®-G1D1 kit:購自 OncoImmunin(Gaithersburg, MD, USA) 17. RNase A(Ribonuclease A):購自 Ameresco 18. Triton X-100:purchase from Sigma chemical Co. 19. Ethanol:購自 TEDIA 20. Ammonium persulfate(APS):購自 Amersco 21. Acrylamide/Bis 40% solution(ACRYL/BISTM29:1):購自 Amresco 22. Bovine serum albumin(BSA):購自 Merck 23. Glycine:購自 Amresco 24. Methanol:購自 TEDIA 25. formaldehyde:購自 Merck 26. ECL kit(Enhanced chemiluminescent kit):購自 Amersham 27. Protein assay-Dye reagent concentrate:購自 Bio-Rad 28. Protein marker:購自 Femantas 29. 10X SDS buffer(Sodium dodecyl sulfate):購自 Amresco 30. TEMED(N,N,N’,N’-Tetramethyl-ethylenediamine):購自 Amresco 31. Tris(Tris(hydroxymethyl)-aminomethane) :購自 Amresco 32. Tween 20:購自 Amresco 33. 顯影劑:購自 Kodak 34. 定影劑:購自 Kodak 35. BioMax Flim:購自 Kodak 36. Agarose Ⅰ:購自 Amresco 37. 核酸純化試劑組(DNA purification kit):購自 Gene Mark. 28.
(29) 38. 蛋白質萃取試劑(protein extraction solution)(PRO-PREP):購自 iNtRON Biotechnology, INC. 39. 5× TBE buffer:購自 Amresco 40. Primary antibody (1°抗體): (a). anti-actin:購自 Oncogen;cat # cp47 (b). anti-p53:購自 Sata cruz Biotechology;Lot# k1102 (c). anti-p21:購自 Calbiochem;cat# op68 (d). anti-p27:購自 Sata Cruz Biotechnology ;Lot: G222 (e). anti-casoase-3:購自 upstate;cat# 05-654 (f). anti-caspase-8:購自 Calbiochem;cat# AM46 (g). anti-caspase-7:購自 upstate (h). anti-caspase-9:購自 upstate (i). anti-cdc25A:購自 Oncogen;Lot# D19466-1 (j). anti-cyclin E:購自 upstate (k). anti-cdk2:購自 upstate cat#5-596 (l). anti-cyclin D3:購自 upstate (m). anti-cdk4:購自 upstate;cat# 06139 (n). anti-cdk6:購自 upstate (o). anti-Fas:購自 upstate (p). anti-NF-κb p50:購自 Zymed;Lot: 30979746 (q). anti-NF-κb p65:購自 Zymed;Lot: 30577746 (r). anti-bcl-2:購自 upstate;cat# 05729 (s). anti-bax:購自 upstate (t). anti-cytochrome c:Calbiochem;cat#PC323. 29.
(30) 41. Secondary antibody(2°抗體): (a). goat anti-mouse IgG ( HRP ) horseradish peroxidase conjugated antibody:購自 Chemicon;AP124P (b). gout anti-rabbit IgG(HRP)horseradish peroxidase conjugated antibody: 購自 Chemicon 三、儀器設備、器材 1. 細胞培養皿:購自 FALCON 2. 細胞培養盤:購自 FALCON 3. 細胞培養箱:購自 Nuaire 4. 細胞計數器(Haemocytometer):購自 Boeco 5. 倒立式位像差顯微鏡(phase-contrast microscope):購自 Olympus 6. 微量天平 (TE-200; MILLTER) 7. 去離子水製造機:購自 Minipore 8. 電源供應器:購自 Amersham 9. 酸鹼值測定計(C831):購自 Consort 10. PVDF membrane:購自 Minipore 11. Mini-3D Shaker:購自 Boeco 12. SDS-PAGE 電泳槽套組:購自 Bio-Rad 13. Transfer Cell Blot 套組:購自 Bio-Rad 14. 加熱板:購自 Lab-Line 15. 流式細胞計數儀(Flow cytometry):購自 Becton Dickinson 16. 高速離心機:購自 HERMLE 17. 分光光度計:from Beckman. 30.
(31) 18. 光學顯微鏡(Olympus CH2) 19. 光學顯微鏡(Nikon LABOPHOT-2) 20. 酵素免疫分析儀(anthos 2020):購自 Anthos Labtec, Australia 21. DNA 電泳槽:購自 Mupid-2. 第二節. 實驗方法. 一、藥品配製 將購自 sigma chemical Co.之 4,5-dihydroxyanthraquinone-2-carboxylic acid 大 黃酸(rhein)純化物,秤取 5.6846 mg 大黃酸,溶於 1ml DMSO,配製成 20 mM stock solution,從 stock solution 分別取不同濃度配製所需濃度,配製濃度如下: 分子式:C15H8O6 Molecular Weight = 284.23 莫耳濃度(M)=分子量/體積(L) 284.23×103 mg/1000 ml=1000 mM 0.28423 mg/ml=1 mM 20 mM=5.6846 mg/ml 表 3-1 大黃酸之配製 20 mM of rhein. DMSO. 0.5 mM. 25 µl. 975 µl. 1 mM. 50 µl. 950 µl. 2.5 mM. 125 µl. 875 µl. 5 mM. 250 µl. 750 µl. 10 mM. 500 µl. 500 µl. 將配製好不同濃度之大黃酸用來進行細胞增生及細胞週期試驗,以及細胞凋亡. 31.
(32) 測試(DNA fragmentation electrophoresis analysis)以及利用西方點墨法(western blot)觀察蛋白質表現等分析。 二、細胞培養 (一)培養條件 人類子宮頸癌細胞株(Ca Ski)以 RPMI1640 培養基添加 10%胎牛血清、1% L-glutamine 及 1% Penicillin Streptomycin 之培養液,置於 5% CO2 及充分溼度下的 37 ℃的細胞培養箱(incubator)中培養,約二天更換一次培養液至細胞長滿。 (二)細胞冷凍保存 Ca Ski 細胞株經大量培養後,可利用液態氮冷凍保存,欲冷凍保存之細胞應在 生長旺盛且存活率高之狀態下進行。細胞冷凍保存方法為,冷凍前應注意細胞生長 情形,並在前一日更換培養基。凍細胞前需先配製冷凍保存溶液:將 DMSO (dimethyl sulfoxide)加入新鮮培養基中,使其最終濃度為 7%,混合均勻,置於 室溫下待用。取少量細胞懸浮液(約 100 µl),計數細胞濃度及凍前存活率。離心 後倒掉上清液,加入事前配製好的冷凍保存液,使其細胞數在 2×106×~5×106 cells/ml,混合均勻後置於以標示好名稱、數目及冷凍日期之冷凍管中,1 ml/管。 冷凍保存方法:冷凍管置於 4 °C,10 min→ -20 °C,30 min → -80 °C,16~18 小時→液態氮槽長期保存。 (三)繼代培養 待細胞長至八分滿時,將培養液取出丟棄,先以 PBS(phosphate buffer saline) 清洗細胞 1~2 次,再加入 3 ml trypsin ,置於 37℃培養箱中處理 2 分鐘後,加入 乾淨培養液以中和 trypsin 之作用,再將瓶中所有液體裝到離心管中以 1500 rpm 離 心 5 分鐘,離心後倒掉上清液,加入 20 ml 新鮮培養液,再使用電動吸量管反覆抽. 32.
(33) 吸使均勻混和,取 20 µl 之細胞懸浮液加入 80 µl trypan blue 溶液,以血球計數盤 (Counting chamber)計算細胞數目(死細胞會被 trypan blue 染成藍色,而活細 胞則不被染色)。 細胞數計算: χ(為所計數的細胞數) ×20 (20 ml 培養液)×5 (trypan blue 5 倍稀釋)×104 依實驗目的不同,計算所需細胞液置於不同培養皿中。 (四)藥物處理 重新繼代的細胞經 24 小時培養後,待貼壁細胞貼覆瓶底後,依實驗所需不同 的時間長短選擇是否加藥前置換培養液,若 24 小時內可不需置換培養液,若超過 48 小時則先置換培養液再加入藥物處理。microplate 每 well 加入培養液體積 2 ml, 則加 20 µl 藥物;3 ml,則加 30 µl,則藥物濃度被稀釋 100 倍為最終濃度。 三、流式細胞儀分析測定 流式細胞儀廣泛的應用於癌細胞臨床樣品檢測當中,是腫瘤生物學研究的重要 工具之一,為癌細胞的早期的藥效評估提供重要資訊。流式細胞分析技術的發展提 供一快速檢測且可靠的方法定量細胞懸浮液,若配合適當的染劑,則可以對藥物處 理細胞後做不同的效果評估,具有準確及快速的優點。 (一)細胞存活率測定 1. 原理 Propidium iodine(PI)是一種核酸染劑,當細胞死亡時會進行壞死或細胞 凋亡的路徑,死亡的細胞膜會失去完整性,使得 PI 可進入細胞內與核酸結合, PI 會與 DNA 雙股螺旋中之 A=T、C≣G 鍵結的氫鍵(hydro bond)接在一起; 存活的細胞因其細胞膜完整 PI 無法和細胞內的核酸結合,經 PI 染色完成的細. 33.
(34) 胞可由流式細胞儀於 488 nm 的雷射光激發後死亡的細胞會呈現較強紅色螢 光,存活細胞會成較弱的紅色螢光以 CellQuest 軟體分析細胞存活率。 2. Survival PI 染劑 PI stock(2 mg/100 ml PBS):10 ml 1×PBS:40 ml 表 3-2. 1 倍磷酸鹽緩衝鹽溶液(1x phosphate buffer saline, PBS;pH. 7.4)之組成. 組成. 重量(g). NaCl. 8.0. KCl. 0.2. Na2HPO4. 1.44. KH2PO4. 0.24. 加 DDW 至總體積 1000 ml. 表 3-3. PI(Propidium iodide)stain 染劑之組成 組成. 濃度. 體積(ml). Propidium iodide. 2 mg/dl. 10. 1× PBS. -. 40 50 ml. 總體積. 3. 實驗步驟 將細胞種植於 12 well plate 中,細胞依照 2×105/well 種植於培養皿中,經 過 24 小時靜置培養後,待細胞貼附後加入不同濃度的 rhein(0.5 mM、1 mM、. 34.
(35) 2.5 mM、5 mM、10 mM) ,分別持續培養 12、24、48 及 72 小時。加藥時間 後,收細胞,將上層液移至離心管中,加入 PBS 清洗細胞一次後,再將細胞以 trypsin 處理,置於 37℃培養箱中處理 2 分鐘後,將細胞打下來,加入 1 ml PBS 以中和 trypsin 之作用,再將所有液體裝到離心管中,1500 rpm 離心 5 分鐘, 去除上清液,再加入 1 ml PBS 清洗細胞,1500 rpm 離心 5 分鐘,去除上清液, 加入 PI 染劑 350 µl(350~550 µl,可依照細胞數做調整 ) ,均勻混合後,transfer 至 FACS 管中,以流式細胞儀進行樣品分析,固定秒數及流數,紀錄細胞增殖 率 gate%存活百分比。 (二)細胞週期分析(Darzynkiewicz et al., 1992) 1. 原理 Proopidium iodide(PI)是一種螢光染劑,可專一的鍵結的核酸而廣泛的應用 於流式細胞儀技術中,正常情況下細胞膜維持完整 PI 無法穿透細胞之細胞膜,故可 以併入其他實驗(如:與 FITC 標記之 annexinV 共同偵測,以區別存活壞死或凋亡細 胞)或者與其他染劑共同使用,評估細胞存活狀態若以酒精將細胞膜打洞固定後, 進入細胞內之 PI 可與核酸進行鍵結,利用流式細胞儀(Flow cytometry; FACS)偵 測 PI 所貢獻之螢光,則可以反應細胞內之 DNA 狀態或進行細胞週期分析。 2. Cell cycle Proopidium iodide(PI)染劑: 表 3-4. Cell cycle Proopidium iodide(PI)染劑之組成. 組成. 濃度. 體積(ml). Propidium iodide. 2 ml/dl 5%. 5. Trion RNase A 1× PBS 總體積. 5. 2 mg/ml -. 1.25 13.75 25 ml. 35.
(36) 3. 實驗步驟 將細胞種植於 12 well plate 中,細胞依照 2×105/well 種植於培養皿中,經 過 24 小時靜置培養後,待細胞貼附後加入不同濃度的 rhein(0.5 mM, 1 mM, 2.5 mM, 5 mM, 10 mM) ,分別培養 12、24、48 及 72 小時。加藥時間後,收細胞, 將上層液移至離心管中,加入 PBS 清洗細胞一次後,再將細胞以 trypsin 處理, 置於 37 ºC 培養箱中處理 2 分鐘後,將細胞打下來,加入 1 ml PBS 以中和 trypsin 之作用,再將所有液體裝到離心管中, 1500 rpm 5 分鐘的離心(重複此步驟 一次) ,去除上清液,置於震盪器輕輕震盪再加入 70% EtOH/PBS,一滴一滴滴 入離心管中以固定細胞,使細胞完全均勻分散於 70% EtOH/PBS 中,靜置-20 ºC 至少一小時,取出細胞後離心(1500 rpm 5 分鐘),倒掉上清液加入 PBS 3ml 清洗細胞,離心去除上清液,加入 cycle PI 染劑 350 µl,將細胞團打散,避光 30 分鐘,以流式細胞儀行樣品分析,所得之結果以 Modfit LT 軟體分析。 (三)粒腺體膜電位(Mitochondria membrane potential; ∆Ψm)之檢測 1. 原理(Petit et al., 1990) 細胞膜電位探針,DioC6(3,3’-Dihexyloxacarbocyanine iodide)是一種可 穿透細胞膜,可專一性的結合並累積在細胞粒腺體中,DioC6在細胞內外的分佈 可反應出細胞膜內外的電位差,其螢光強度的改變就可顯示細胞膜電位改變的 情形,可發射出綠色螢光(green-fluorescent)之陽離子(cationic dye)親脂 性染劑;在不需要固定細胞或其他處理的情況下可以即時、快速的偵測活細胞 內粒腺體膜電位。粒腺體膜功能不良(mitochondrial dysfuction)通常伴隨早 期細胞凋亡發生,而細胞粒腺體膜電位的改變也因此當作早期凋亡偵測上的指 標。. 36.
(37) 2. MMP(∆Ψm)試劑:DioC6(3,3’-Dihexyloxacarbocyanine iodide) 10 µl DioC6/500 µl PBS 3. 實驗步驟 將細胞種植於12 well plate中,細胞依照2×105/well種植於培養皿中,經過24 小時靜置培養後,待細胞貼附後加入5 mM rhein,20 µl/well到12 well plate當中(最 終濃度為50 µM) ,經不同時間培養(1、3、6、12及24 h) ,加藥時間後,收細胞, 將上層液移至離心管中,加入PBS清洗細胞一次後,再將細胞以trypsin處理,置於 37℃培養箱中處理2分鐘後,將細胞打下來,加入1 ml PBS以中和trypsin之作用, 再將所有液體裝到離心管中,1500 rpm離心5分鐘,去除上清液,再加入1 ml PBS 清洗細胞,1500 rpm離心5分鐘,去除上清液,取MMP(∆Ψm)染劑(10 µl DioC6/500 µl PBS)DioC6(3,3’-Dihexyloxacarbocyanine iodide)染劑每管加入500 µl,需有 一管blank不加藥也不加染劑,於37 ºC培養箱避光培養30 min後,transfer至FACS 管中,以流式細胞儀進行樣品分析,每樣品收集10000顆細胞以CellQuest軟體分 析。將blank(格線的peak)把peak調在100~101之間control(斜線的peak)調在 101~102之間,M1 gated約75﹪以上sample上機後,分析MMP(peak往右不產生 apoptosis,往左產生apoptosis) 。. M1. 100 101 102 103 104 圖 3-1 MMP 軟體的分析圖. 37.
(38) (四)活性氧化物產生之檢測 1. 原理 (Royall et al., 1993) 免疫細胞進行需氧性滅菌過程中,會在細胞內啟動一連串的氧化還原反 應,因而產生一些 Oxidative Metabolites,如 H2O2 ,O2- free radical。如欲以 流 式 細 胞 儀 測 量 這 些 代 謝 物 的 產 量 , 可 用 Dihydrorodamine 123 、 2’7’-dichlorofluorescein(for H2O2)或 hydroethidine(for O2- free radical)等 螢 光 染 劑 , 對 細 胞 進 行 染 色 , 藉 由 2’,7’-dichlorofluorescein diacetate (H2DCF-DA)產生螢光來測量 ROS 的產生,H2DCF-DA 是一種具有螢光性質, 可滲透細胞膜特異性的追蹤評估 ROS 的產生。H2DCF-DA 會被細胞內的乙醯酯 酶(esterases)去乙醯化(deacetylated)成非螢光性的 DCFH,DCFH 會在細 胞內被 H2O2 氧化成螢光性質的 DCF,並聚集在粒腺體中,所發散螢光則可反 映出細胞內 H2O2 的濃度。 2. ROS 試劑:2’,7’-dichlorofluorescein diacetate(H2DCF-DA) 1 µl H2DCF-DA /500 µl PBS 3. 實驗步驟 將細胞種植於12 well plate中,細胞依照2×105 /well種植於培養皿中,經過24 小時靜置培養後,待細胞貼附後加入5 mM rhein,20 µl/well到12 well plate當中(最 終濃度為50 µM) ,經不同時間培養(0.5、1、2、4及6 h) ,加藥時間後,收細胞, 將上層液移至離心管中,加入PBS清洗細胞一次後,再將細胞以trypsin處理,置於 37℃培養箱中處理2分鐘後,將細胞打下來,加入1 ml PBS以中和trypsin之作用, 再將所有液體裝到離心管中,1500 rpm離心5分鐘,去除上清液,再加入1 ml PBS 清洗細胞,1500 rpm離心5分鐘,去除上清液,取ROS染劑H2DCF-DA染劑(1 µl. 38.
(39) H2DCF-DA /500 µl PBS)每管加入500 µl,需有一管blank不加藥也不加染劑,只加 入500 µl PBS,在置於37 ºC培養箱避光培養30 min後,transfer至FACS管中,以流 式細胞儀進行樣品分析,每樣品收集10000顆細胞以CellQuest軟體分析。將blank (格線的peak)peak調在100~101之間,control(斜線的peak)調在101~102之間, M1 gated約75﹪以上,sample上機後,分析ROS(peak往右是產生自由基,往左 是抗氧化). M1. 100. 101. 圖 3-2. 102. 103. 104. ROS 軟體分析圖. (五)鈣離子釋出之檢測 1. 原理(Beatrice et al., 1991) 胞內鈣離子作為細胞信號傳遞的信差,是細胞激活過程中重要的功能參 數。螢光染劑(如Fura-Red、Fluo-3和Indo-1等)通過乙醯甲酯(Acetatoxymethyl Ester;AE)導入細胞後,Indo-1會與鈣離子特異性結合。這些螢光染劑的結構 式,一般與EDTA相似,可螯合鈣離子,螯合鈣離子之螢光染劑會有光學特性上 的改變,在(紫外光)UV 的激發下,Indo-1 放出光(emission)的強度會隨 著細胞內鈣離子濃度的改變,而發散出不同強度的螢光,故可用比例法測得或 直接測得的螢光強度得到鈣離子濃度的相對值,實際濃度需經校對後獲得。 2. 鈣離子染劑 [1-[2-amino-5(6-carboxylindol-2-yl)-phenoxy]-2-(2’amino-5-methylphenoxy). 39.
(40) ethane-N,N,N’,N’-tetraacetic acid(Indo-1-AM) 3. 實驗步驟 將細胞種植於12 well plate中,細胞依照2×105 /well種植於培養皿中,經過 24小時靜置培養後,待細胞貼附後加入5 mM rhein,20 µl/well到12 well plate 當中(最終濃度為50 µM),經不同時間培養(0.5、1、2、4及6 h) ,加藥時間 後,收細胞,將上層液移至離心管中,加入PBS清洗細胞一次後,再將細胞以 trypsin處理,置於37℃培養箱中處理2分鐘後,將細胞打下來,加入1 ml PBS 以中和trypsin之作用,再將所有液體裝到離心管中,1500 rpm離心5分鐘,去 除上清液,再加入1 ml PBS清洗細胞,1500 rpm離心5分鐘,去除上清液,取 Indo-1-AM染劑每管加入1000 µl,需有一管blank不加藥也不加染劑,只加入 1000 µl PBS,在置於37ºC培養箱避光培養1h,每10 min上下混合一次,1h後 加入PBS洗2次,1500 rpm離心5 min,倒掉上清液,每管加入400 µl PBS,再 transfer至FACS管中,以流式細胞儀進行樣品分析,每樣品收集10000顆細胞以 CellQuest軟體分析。將blank(格線的peak)把peak調在100~101之間control(斜 線的peak)調在100~101之間,M1 gated約0%,以上sample上機後,分析calcium release(peak往右為鈣離子釋出) M1. 100. 101. 102. 103. 104. 圖3-3 鈣離子軟體分析圖. 40.
(41) (六)Caspase-3 活性分析 1. 原理 利用 PhiPhiLux-G1D1 kit 來檢測凋亡細胞 caspase3 之產生,PhiPhiLux-G1D1 kit 基質是種含有螢光物質之胺基酸序列(amino acid sequence),而活化之 caspase-3 可以裂解胺基酸序列之特定位置,而是螢光物質釋放出來,再經由 流 式 細胞儀分析,可得知 若螢光產量越多則產生活性之 caspase-3 越 多 (Komoriya et al., 2000)。 2. 實驗步驟 將細胞種植於 12 well plate 中,細胞依照 2×105 /well 種植於培養皿中,經 過 24 小時靜置培養後,待細胞貼附後加入 5 mM Rhein 20 µl/well 到 12 well plate 當中(最終濃度為 50 µM) ,經不同時間培養(6、12、24、48 及 72 h); 加藥時間後,收細胞,將上層液移至離心管中,加入 PBS 清洗細胞一次後,再 將細胞以 trypsin 處理,置於 37℃培養箱中處理 2 分鐘後,將細胞打下來,加 入 1 ml PBS 以中和 trypsin 之作用,再將所有液體裝到離心管中,1500 rpm 離 心 5 分鐘,去除上清液,再加入 1 ml PBS 清洗細胞,1500 rpm 離心 5 分鐘, 去除上清液,取 10 µM substrate(Phiphilux green for caspase-3) (Phiphilux red for mitochondria)每管加入 25 µl,在置於 37 ºC 培養箱避光培養 1 h,1 h 後 加入 1 ml PBS 洗 1 次,1500 rpm 離心 5min,倒掉上清液,每管加入 500 µl PBS, 再 transfer 至 FACS 管中,以流式細胞儀進行樣品分析,每樣品收集 10000 顆 細胞以 CellQuest 軟體分析。將 blank(格線的 peak)把 peak 調在 100~101 之間 control(斜線的 peak)調在 101~102 之間,M1 gated 約 75﹪以上 sample 上機後,分析 caspase-3 活性(peak 往右為 caspase-3 產生). 41.
(42) M1. 100. 101. 圖 3-4. 102. 103. 104. caspase-3 軟體分析圖. 四、細胞內分子螢光染色(Intracelluar flouorescence staining) (一)DAPI(4 -6-diamidine-2-phenyl indole)染色 1. 原理 DAPI(4 -6-diamidine-2-phenyl indole)是種核酸螢光染劑,其會專一性 的 binding 在 DNA 雙股螺旋之小溝(minor groove)上,當細胞凋亡時會出現 染色質凝結(chromosomes condensation) 、DNA 斷裂(DNA fragmentation) 情形發生,若細胞凋亡越嚴重,則 DNA 斷裂越多而 DAPI 染劑就會染上越多, 在顯微鏡,下可觀察到白色螢光強度就越亮。. DAPI 染劑. X-ray crystal structure of DAPI bound to DNA. Derived from data submitted to the Protein Data Bank (number PDB 1D30, http://www.rcsb.org/pdb), the structure shows that DAPI binds to DNA in the minor groove.. 圖 3-5. DAPI 染劑鍵結在 DNA 雙股螺旋小溝(minor groove)上. 42.
(43) 2. 實驗步驟 Ca Ski 細胞分盤,種於 6 well plate 當中,每 well 種 1×104/ml(通常會種 3 ml) ,隔日,待細胞貼附後加入不同濃度的 rhein(0.5 mM、1 mM、2.5 mM、 5 mM、10 mM),分別培養 12 和 24 小時。 加 藥 時 間 後 , 收 細 胞 , 吸 取 上 層 液 丟 棄 , 加 入 PBS 洗 3 次,加入 3% Formaldehyde/PBS 固定 10~15 分鐘,再加入 PBS 洗 2 次後,再加入 0.1% Triton X-100/PBS(1 ml)15 分鐘,再用 PBS 洗 2 次,《接下來步驟要避光處理》加 入 DAPI 染液(1 µl/ml)300 µl,置於 37℃水浴槽避光 30 分鐘後,取出,用 PBS 洗 3 次後,於螢光顯微鏡 F200 照相。 (二)彗星試驗(Comet assay) 1. 原理 單細胞電泳分析(single cell gel electrophoresis assay)就是所謂的彗星 試驗(Comet assay) ,可用來分析及定量 DNA 損傷(DNA damage)程度,是 一個簡單、快速以及敏感度高的技術。利用 DNA damage 後發生斷裂,藉由電 泳將斷裂的 DNA 拖出膜外,形成彗星狀而命名之,由此可藉由拖尾的長短,觀 察 DNA 的損傷情形。 2. 實驗步驟 Ca Ski 細胞分盤,種於 6 well plate 當中,每 well 種 1×106/3ml,隔日, 待細胞貼附後加入不同濃度的 rhein(0.5 mM、1 mM、2.5 mM、5 mM、10 mM) , 分別培養 12 和 24 小時,加藥前須先置換新鮮 medium,陰性對照組加入 DMSO;陽性對照組加入 3 µM H2O2 (加入體積為 1%),將 gel 以微波溶解 後放於 55 ℃ 水浴鍋中備用,以 0.1% trypsin 200 µl digestion 2 min,加入. 43.
(44) 1 ml PBS 中和 trypsin 作用後將 cell 吸至 1.5 ml 離心管,以 1500 rpm 離 心 5 min 去上清液,彈散 pellet 加入 1×PBS 200 µl(體積依細胞量調整), 在磨砂載玻片上以鉛筆標上號碼,置 70 µl. 0.5% Normal-melting-point. argarose(NMA)+ 0.5% low-melting-point argarose(LMA)於載玻片上, 將蓋玻片以 45° 角蓋上防止氣泡產生,凝固後拿下蓋玻片,取 10 µl 細胞液 加上 60 µl (80-10) 0.5% LMA 加在第一層膠上,將蓋玻片以 45° 角蓋上 防止氣泡產生,凝固後拿下蓋玻片,配置 lysis buffer。將做好的膠置於 lysis buffer 1 hr,置於冰上,將膠移至 alkalin buffer 中 20 min,將電泳槽置於冰 上,以 alkalin buffer 為電泳液跑 30 min (25 V;300 mA) 。將膠移至 0.4 M Tris buffer 使 pH 回到中性,再將膠置於 methanol 脫水,加 PI( 2.5 µg/ml ) 以螢光顯微鏡觀察,數 100 顆細胞打價數。 表 3-5. Comet assay 之 lysis buffer 配製 Lysis buffer(需新鮮配置). pH=8~10. 5M NaCl. 100 ml. 1M Tris-HCl. 2 ml. 0.5M EDTA. 40 ml. Triton. 2 ml. DDW. 56 ml. Total. 200 ml. 目的:Lysis buffer 將細胞膜上雙層磷脂(phospholipids)結構破壞,以利斷裂的 DNA 被電泳拖出。. 44.
(45) 表 3-6. Alkalin buffer 之配製 Alkalin buffer(pH=13) NaOH. 12 g. EDTA. 0.3724 g. DDW. 1000 ml. 目的:Alkalin buffer 將 DNA 雙股螺旋解開,若斷裂的 DNA 則會被拖出膜外。 表 3-7. Tris buffer 之配製 0.4M Tris buffer(以 HCl 調整 pH=7.5) Tris. 48.456 g. DDW. 1000 ml. 五、DNA 斷裂分析(DNA fragmentation assay) 1. 原理 當細胞 apoptosis 時會有 DNA 斷裂現象發生,其細胞核內 DNA 會裂解成 180~200 bp,可藉由抽取 DNA 後,再經由電泳跑膠,看出是否有 DNA 斷裂 現象,要是有斷裂則電泳跑膠照相後會出現階梯狀(ladder) ,可藉由 marker 來得知其斷裂鹼基對(base pair) 。 2. 材料 Tissue and Cell Genomic DNA Purification Kit(GeneMark) 3. 實驗步驟: 將細胞種植於 10cm plate 中,細胞依照 2×106 /well 種植於培養皿中,每 well medium 總量為 10 ml,經過 24 小時靜置培養後,待細胞貼附後加入 5 mM. 45.
(46) rhein ,100 µl/well 到 6 well plate 當中,經不同時間培養(24、48 及 72 h) ; 加藥時間後,收細胞,將上層液移至離心管中,加入 PBS 清洗細胞一次後, 再將細胞以 trypsin 處理,置於 37℃培養箱中處理 2 分鐘後,將細胞打下來, 加入 1 ml PBS 以中和 trypsin 之作用,再將所有液體裝到離心管中,1500 rpm 離心 5 分鐘,去除上清液,再加入 1 ml PBS 清洗細胞,1500 rpm 離心 5 分鐘, 倒掉上清液 ,加入 200 µl PBS 混合均勻將細胞移至 eppendorf,14000 rpm 離心 1 分鐘,小心移去上清液,加入 200 µl PBS or TE buffer,將細胞懸浮均 勻,再加入 4 µl RNase A(10 mg/ml) ,室溫靜置 5 min,將細胞中 RNA 溶解, 再加入 Proteinase K 20 µl,及 Binding solution 200 µl,置於 70℃水浴,10 min, 再加入 100% EtOH 200 µl,votex 均勻(若此時液體呈現濃稠白色黏液狀,則 細胞數過多,會降低 DNA 抽取的效果),transfer 至 Spin column,14000 rpm 離心 1 分鐘(離心後 DNA 會 binding 到 spin column 中間白色那層) ,再加入 300 µl Binding solution,14000 rpm 離心 1 分鐘,再以 700 µl Washing solution 清洗兩次(14000 rpm 離心 1 分鐘) ,最後一次將 column 中所有液體都離下來, 不加任何 solution 直接 14000rpm 離心 5min,再置於烘箱 3~5 min,將 column 由烘箱中取出後,加入 100 µl Elution solution(Elution solution 需先置於 70ºC 水浴加熱,可增加 DNA 溶解性,直到欲使用前再取出),室溫靜置 2 分鐘後, 14000rpm 離心 5min,離下液體即是含有 DNA 之 solution。 跑膠: 將 DNA 與 6× DNA loading dye 混合(10 µl DNA solution+2 µl 6× DNA loading dye)loading 至 2% agarose (1g agarose、50ml 0.5× TBE buffer、10 µl Eithidium bromide)中。進行電泳,最後放入 UV light 下檢查並照相儲存。. 46.
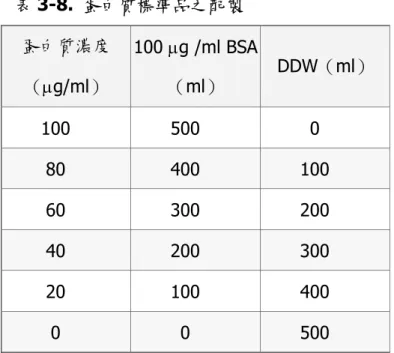
(47) 六、細胞蛋白質之萃取(Protein extraction) (一)細胞總蛋白之製備 1. Protein Extraction Solution(lysine buffer) PRO-PREP for Cell/Tissue 2. 實驗步驟 將細胞種植於 6 well plate 中,細胞依照 5×105 /well 種植於培養皿中,每 well medium 總量為 3 ml,經過 24 小時靜置培養後,待細胞貼附後加入 5 mM rhein ,30 µl/well 到 6 well plate 當中,經不同時間培養(6、12、24 及 48h), 加藥時間後,收細胞,將上層液移至離心管中,加入 PBS 清洗細胞一次後,再 將細胞以 trypsin 處理,置於 37℃培養箱中處理 2 分鐘後,將細胞打下來,加 入 1 ml PBS 以中和 trypsin 之作用,再將所有液體裝到離心管中,1500 rpm 離 心 5 分鐘,去除上清液,再加入 1 ml PBS 清洗細胞,1500 rpm 離心 5 分鐘, 倒掉上清液 ,小心將 PBS 吸乾,再加入 100 µl lysine buffer,將細胞懸浮均勻, 置於-20℃ overnight,之後離心(14000 rpm,20min)取上清液,為 lysine buffer 與細胞蛋白混合之液體,再去進行蛋白質含量測量。 (二)蛋白質濃度測定 1. 蛋白質標準品檢量線製作 以 Bradford 定量法(Bradford, 1976) ,使用胎牛血清白蛋白(Bovine serum albumin; BSA)當作蛋白質標準品,利用酵素免疫分析儀(ELISA reader)在 O.D. 595 nm 來測量蛋白質標準品吸光值來做檢量線(standard curve),以蛋 白質標準品吸光值畫出標準品檢量線,並求出趨勢線方程式及 r2 值。. 47.
(48) 表 3-8. 蛋白質標準品之配製 蛋白質濃度. 100 µg /ml BSA. (µg/ml). (ml). DDW(ml). 100. 500. 0. 80. 400. 100. 60. 300. 200. 40. 200. 300. 20. 100. 400. 0. 0. 500. 先取Bradford染劑2 ml加8 ml二次水(5×稀釋)混合均勻備用,取15 µl配 製好蛋白質標準品(BSA)加735 µl Bradford染劑混合均勻,置於96 well plate 中,每well加入200 µl(三重複),靜置5 min後以O.D.595 nm測量吸光值,測 得標準品之吸光平均值,以O.D. value(Y)對蛋白濃度 µg/mL(X),求出趨 勢線方程式---y=a x + b,r2值要趨近於0.99。 2. 樣品蛋白質定量 取 15 µl sample protein 與 735 µl 的 Bradford 染劑(5 倍稀釋)混合,反 應 5 分鐘後,同蛋白質標準品一起測定吸光值,所得之吸光值平均,帶入 y 值 (y=a x + b),求出該 sample 的蛋白質濃度(µg/ml) 。. 48.
(49) 七、SDS-PAGE 電泳分析 1. 電泳膠片製作 表 3-9. SDS-PAGE 下層膠(Separation gel)之配製及組成 組成. 10% Separation gel (四片量). 12% Separation gel (四片量). H2O 40% Acryamide/Bis(29:1). 9.6 ml 5 ml 5 ml 0.2 ml 0.2 ml 20 µl. 8.6 ml 6 ml 5 ml 0.2 ml 0.2 ml 12 µl. Running buffer 10% SDS 10% APS TEMED. 表 3-10. SDS-PAGE 上層膠(Stacking gel)之配製及組成 組成. Stacking gel (四片量). H2 O 40% Acrylamide/Bis(29:1). 4.06 ml 1.02 ml 1.66 ml. Stacking buffer 10% SDS 10% APS. 66 µl 33.4 µl 12 µl. TEMED. 表 3-11. Running buffer(1.5M Tris-HCl, pH8.8)之組成 組成. 重量. Tris. 36.3 g. DDW. 150 ml 調整至 pH8.8. HCl. 加 DDW 到總體積 200 ml. 49.
(50) 表 3-12. Stacking buffer(0.5M Tris-HCl, pH6.8)之組成 組成. 重量. Tris. 3g. DDW. 40 ml. HCl. 調整至 pH6.8 加 DDW 到總體積 50 ml. 2. 實驗步驟 將蛋白質依分子量分離利用 SDS-PAGE,將配製好的下層膠 Separation gel 注入鑄膠台中,再以 isopropanol 去除氣泡並壓平下膠,靜置一段時間,可觀 察管子當中的剩餘下層膠是否凝固,待下層膠凝固後,將 isopropanol 倒掉, 注入上層膠並插上 comb,並避免氣泡產生,待上層膠凝固後,將鑄好膠體放 置於電泳槽中,加入電泳緩衝液(running buffer;表 3-10),接著將將萃取出 的蛋白質與 5X protein loading dye 混合並以 100℃加熱 10 min 後,冰上冷卻 後離心,依序將標示標準分子量的 Multimaker 5 µl 及各 sample 18 µl 注入膠體 的孔槽中,通以電壓 80 Volts,待樣品通過 stacking gel 後,電壓調為 110 Volt, 進行以電泳,當 SDS-PAGE 染劑跑出 SDS-PAGE 後或可視其需要,即可關掉電 源。 表 3-13. 電泳緩衝液(running buffer)之組成 組成. 重量. 10X SDS buffer(25mM Tris、192 mM. 200 ml. glycine、0.1% SDS) 加 DDW 到總體積 2000ml. 50.
(51) 八、西方點墨法(Western blotting) (一)轉漬步驟: 先將PVDF membrane裁剪好,再以methanol短暫濕潤後,再浸入轉印緩衝液 (transfer buffer;表3-11)中,接著將裁好的濾紙先浸泡在transfer buffer中備用, 將轉漬夾打開後,黑色面朝下,將海綿墊片先以transfer buffer潤濕並鋪在黑夾上, 再將3M濾紙鋪上,接下來裁剪下層膠(separation gel)中所要轉漬的區域後,將 SDS-PAGE gel小心的鋪於3M濾紙上,可在濾紙上加入多量的transfer buffer,再鋪 上SDS-PAGE gel時勿陷入任何氣泡,再依序放上PVDF membrane,同樣避免氣泡產 生,及3M濾紙,最後再放上一片海綿墊片,即可把整個轉漬夾裝好,形成似三明治 夾層狀之構造(圖3-5) 。置入已裝有transfer buffer的電泳槽中將黑夾朝負極,紅夾 朝正極,電泳槽外圍放置足夠冰塊,使整各系統維持低溫狀態。以400 mA、2小時 的條件下進行蛋白質轉漬步驟。 轉印完成後取出轉印膜裁去多餘部分,轉印膜後以0.05% tween 20/1X PBS 清 洗10分鐘共3次。緊接將轉印膜以2% FBS(溶於0.05 % tween 20/1X PBS 中)進 行blocking 步驟,以室溫1小時為條件進行。取出轉印膜後於小盒中以0.05 % tween 20/1X PBS 清洗10分鐘共3次。倒掉清洗液,加入8 ml的一級抗體(溶於新鮮配製 之blocking solution中,稀釋倍數依不同抗體有所不同),4 ℃隔夜進行搖盪。隔天 取出,回收一級抗體,以0.05% tween 20/1X PBS 清洗轉印膜10分鐘共3次。加入8 ml 稀 釋 10000 倍 的 goat anti IgG ( HRP ) horseradish peroxidaseconjugated antibody 二級抗體(溶於含2% FBS的0.05% tween 20/1X PBS中),於室溫下搖 盪進行1小時,最後取出轉印膜後以0.05% tween 20/1X PBS 洗清洗10分鐘共3次。. 51.
(52) (二)壓片步驟:(暗房中進行) 將轉印膜浸泡於ECL 試劑之混合液(每瓶各取1.5 ml等比例混合)中1分鐘反 應。以兩張投影片黏貼固定於cassette 內,轉印膜並正面朝上放置於壓片卡匣 (cassette)兩張投影片中間,以Hyperfilm 軟片置於上層投影片上,對準轉印膜進 行壓片,感光時間依轉印膜上螢光亮度決定時間長短,約5 秒至1 小時不等。感光 完成後放入顯影劑進行顯影步驟(時間依實際觀察決定),再以清水沖洗30秒後放 入定影劑中,過30 秒後再以清水沖洗30 秒。. 表 3-14. 轉印緩衝液(Transfer buffer)之組成 組成. 重量. Tris. 4.5 g. Glycin. 21.6 ml. methanol. 300 ml 加 DDW 到總體積 1500 ml. 紅、正極(+). 海綿墊片 3M paper PVDF membrane SDS-PAGE gel 3M paper 海綿墊片. 黑、負極(-). 圖 3-6 轉漬夾內部組成. 52.
(53) 表 3-15. PBS-tween 20 之組成 組成. 重量. Tween 20. 500 µl. PBS. 1000 ml. 九、統計分析(Statistics analysis) 實驗結果以平均值標準差(mean ± SD)表示,使用Student’s t-test 來決定實 驗組與對照組之差異。*表示p<0.05,表示統計上具顯著差異。. 53.
(54) 第四章 結果 本實驗以探討大黃酸(rhein)對人類子宮頸癌細胞 Ca Ski 細胞之影響,首先 測試大黃酸對 Ca Ski 細胞之生長抑制率及存活率之影響,及藉由型態(morphology) 上觀察細胞之變化,再更進一步利用 DAPI 染色及單細胞凝膠電泳(single cell gel electrophoresis; Comet assay)觀察細胞是否有細胞損傷(DNA damage)情形發 生及細胞凋亡(apoptosis)現象。接下來再探討大黃酸對 Ca Ski 細胞細胞週期影響, 並利用 DNA 斷裂分析(DNA fragmentation) 、粒線體膜電位(∆Ψm)、活性氧化物 產生(ROS products)、鈣離子釋放(calcium release)、以及凋亡蛋白 caspase-3 活性之檢測,來做凋亡路徑探討,最後再以 Western blot 來檢測凋亡蛋白質表現, 以探討大黃酸對 Ca Ski 細胞凋亡之影響。. 第一節 大黃酸(rhein)對人類子宮頸癌細胞株(Ca Ski)抑制增殖作用 將細胞培養於 12 well plate 中,細胞依照 2×105/well 培養於培養皿中,於 37℃、5% CO2 的培養箱中經過 24 小時靜置培養後,待細胞貼附後加入不同濃度的 rhein(0.5 mM、1 mM、2.5 mM、5 mM、10 mM),分別持續培養 12、24、48 及 72 小時。於加藥後第 12、24、48 及 72 小時,分別取出平盤,收細胞後加入 Propidium iodine(PI) ,以流式細胞儀分析大黃酸 rhein 對人類子宮頸癌細胞存活的影響。 經由流式細胞儀分析細胞後可得知細胞存活率及固定時間及流速下的存活細 胞數來得知細胞生長抑制率,由圖 4-1、4-2、4-3、4-4、4-5 得知大黃酸對人類子 宮頸癌細胞株(Ca Ski)具有顯著的存活率降低的現象,隨著劑量的增加及作用時 間的加長,大黃酸(rhein)對人類子宮頸癌細胞株(Ca Ski)的抑制增生作用越強, 呈現劑量時間依賴效應(dose-and time-dependent response)。細胞在投予 5 µM rhein 12 小時後,即有存活率降低的現象,表示 Ca Ski 細胞的生長有被抑制的情形 發生,在 48 小時之後的培養發現細胞數目明顯減少,且在 50 µM rhein 經過 48 小. 54.
(55) 時培養後,細胞存活率顯著降低到 50%以下,在高濃度的 rhein(100 µM )投予 後 72 小時,存活細胞已降到 30%以下,且有許多的漂浮死亡細胞出現。. 第二節 大黃酸(rhein)對人類子宮頸癌細胞株(Ca Ski)型態上的影響 給予不同濃度(5、10、25、50 及 100 µM)之大黃酸對人類子宮頸癌細胞株 Ca Ski 處理 12、24、48、72 小時後,以倒立式位像差顯微鏡觀察型態上之變化(圖 4-1A、4-2A、4-3A、4-4A)。結果可發現 10 µM rhein 處理 48 小時後細胞已有明顯 皺縮情形發生,且隨著濃度的增加,細胞膜變得模糊不清、細胞質空泡化、漂浮許 多的死亡細胞;以及在 50 µM 大黃酸處理 6 小時後之 Ca Ski 細胞,其細胞型態開 始不完整,觀察 12、24、48、72 小時大黃酸處理之細胞,細胞數目明顯少於未處 理之控制組(圖 4-5A),觀察細胞型態(morphology)也變的較皺縮,細胞膜模糊 不清,甚至漂浮著許多的死亡細胞。. 55.
(56) Control. 5 µM. 10 µM. 25 µM. 50 µM. 100 µM. 圖 4-1A. 不同濃度之大黃酸 rhein 處理 Ca Ski 細胞 12 小時之型態變化 Figure 4-1A. Morphology change of Ca Ski cells after treatment with different concentrations of rhein (0, 5, 10, 25, 50, 100 µM)for 12 hours. Photomicrographs are from phase-contrast microscopy magnification, 200X. 12h viability 80 *. *. * *. * *. Viability ( % ). 60. * *. 40. 20. 0 c. 5. 10. 25. 50. 100. Concentrations of Rhein ( µΜ ). 圖 4-1B. 不同濃度之大黃酸 rhein 處理 Ca Ski 細胞 12 小時存活率結果. Figure 4-1B. Viability of Ca Ski cells after treatment with different concentrations of rhein (0, 5, 10, 25, 50, 100 µM) for 12 hours. Result are means ± S.D, n=3. The significant differences between control groups were analyzed by Student’s. t-test. *P < 0.05, **P < 0.01.. 56.
數據

+7




相關文件
• 由於細胞代謝、紫外線的電離輻射、不良的生 活習慣 ( 煙、酒、肥胖 ) 、各種感染物所引起 的中性粒細胞、巨噬細胞的激活, 會催化分子 氧發生單價還原產生的 "
SF12144A 張基晟 比較 Nanoplatin 併用 Pemetrexed 和 Cisplatin 併用 Pemetrexed 作為第一線治 療用於非鱗狀細胞性之非小細胞肺癌
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
Malins, “Squamous cell carcinoma arising in the lining of an epidermoid cyst within the sublingual gland—a case report,” British Journal of Oral and Maxillofacial Surgery, vol..
Persons with a phenotype mediated by one of these MC1R genetic variants are at greater risk of UV-induced skin cancers, because pheomelanin not only provides less effective
Clinical findings and risk factors to oral squamous cell carcinoma in young patients: A 12-year retrospective analysis.. Hellen-Bandeira-de-Pontes Santos 1 , Thayana-Karla-Guerra
While Korean kids are learning how to ski and snowboard in the snow, Australian kids are learning how to surf and water-ski at the beach3. Some children never play in the snow
2.滴加蒸餾水或亞甲藍液製成水埋標本 3.觀察 石細胞 (厚壁細胞)及
