特殊脂肪酸及天然抗氧化物對3T3-L1脂肪細胞生合成影響; The adipogenic effect of uncommon fatty acids and natural antioxidants on 3T3-L1 cell
115
0
0
全文
(2) 目. 錄. 縮寫對照表 ...............................................................................................................Ⅰ 摘要 ...........................................................................................................................Ⅲ Abstract......................................................................................................................Ⅳ 第一章. 前言與研究目的 .......................................................................................1. 第二章. 文獻回顧 ...................................................................................................2. 一、 脂肪細胞生合成 (adipogenesis) ...............................................................2 1、 脂肪細胞分化之轉錄調控 (transcription cascade) .........................3 2、 PPARγ與脂肪細胞分化..................................................................4 二、 3T3-L1 介紹 ...............................................................................................6 三、 食物成份與脂肪細胞生合成相關研究 ....................................................7 (一) 脂肪酸類 ...........................................................................................7 1. 一般脂肪酸(Common fatty acids) ....................................................7 2. 特殊脂肪酸(Uncommon fatty acids) ................................................8 (1) Conjugated linolenic acid ......................................................8 (2) Phytanic acid..........................................................................9 (二) 天然抗氧化物 ...................................................................................10 1、 Epigallocatechin Gallate (EGCG) ..................................................10 2、 Procyanidins ...................................................................................12 3、 Curcumin ........................................................................................12 4、 Naringenin ......................................................................................13 5、 Ascorbic acid ..................................................................................14 6、 Resveratrol......................................................................................14 7、 Apigenin .........................................................................................14.
(3) 8、 Quercetin.........................................................................................15 9、 Genistein .........................................................................................15 第三章. 材料與方法 ...............................................................................................21. 一、 實驗假說 ....................................................................................................21 二、 實驗設計 ....................................................................................................21 三、 細胞培養 ....................................................................................................24 1、 細胞株 ............................................................................................24 2、 培養基組成與配製 ........................................................................24 3、 細胞繼代培養 ................................................................................24 四、 誘導 3T3-L1 脂肪細胞分化 ......................................................................25 五、 各種測試物之處理 ....................................................................................27 六、 MTT 染色法細胞存活率測定...................................................................30 七、 LDH 細胞毒性之測定 ...............................................................................31 八、 細胞內三酸甘油酯測定 ............................................................................32 九、 細胞內 G3PDH 活性測定 .........................................................................33 十、 Lowry’s method 蛋白質濃度測定 .............................................................35 十一、 脂解速率測定 ............................................................................................37 十二、 油紅染色 Oil-Red O stain 測定 .................................................................38 十三、 細胞凋亡測定 ............................................................................................39 十四、 統計分析 ....................................................................................................40 第四章. 結果 ...........................................................................................................41. 一、 實驗一:脂肪酸對 3T3-L1 脂肪細胞脂質生合成影響 ..........................41 1、測試各種溶劑及正對照 TZD 對 3T3-L1 脂肪細胞分化的影響 ...41 2、一般脂肪酸對 3T3-L1 脂肪細胞分化的影響 .................................41.
(4) 3、特殊脂肪酸對 3T3-L1 脂肪細胞分化的影響 .................................41 (1) Uncommon fatty acids 對 3T3-L1 differentiation 之影響.......42 (2) Uncommon fatty acids 對 3T3-L1 前脂肪細胞 proliferation 之影響 ......................................................................................42 (3) Uncommon fatty acids 對 3T3-L1 已分化成熟脂肪細胞脂 質代謝之影響 ..........................................................................43 (4) c9,t11,t13CLN 對 3T3-L1 三階段 (preadipocyte proliferation、 differentiation、terminal differentiation) 細胞凋亡的影響 ...43 二、 實驗二:天然抗氧化物對 3T3-L1 脂肪細胞脂質生合成影響 ..............44 1、 Natural antioxidants 對 3T3-L1 脂肪細胞分化之影響 ................44 2、 多酚類對 3T3-L1 前脂肪細胞 proliferation 之影響 ....................45 3、 多酚類對 3T3-L1 已分化成熟脂肪細胞脂質堆積之影響 .........45 4、 curcumin 對 3T3-L1 三階段 (preadipocyte proliferation、 differentiation、terminal differentiation) 細胞凋亡的影響 .........45 第五章. 討論 ...........................................................................................................67. 第六章. 結論 ...........................................................................................................74. 第七章. 參考文獻 ...................................................................................................76.
(5) 表 目. 錄. 表 2-1. 脂肪酸化學結構 .........................................................................................17. 表 2-2. 天然抗氧化物化學結構 .............................................................................19.
(6) 圖 目. 錄. 圖 2-1. 脂肪細胞生合成 .........................................................................................2. 圖 2-2. 脂肪細胞分化過程 .....................................................................................3. 圖 2-3. PPARγ之活化與作用 .............................................................................4. 圖 3-1. preadipocyte proliferation 實驗流程 ..........................................................28. 圖 3-2. differentiation 實驗流程 .............................................................................29. 圖 3-3. terminal differentiation 實驗流程...............................................................29. 圖 4-1. 各種溶劑及正對照 TZD 對 3T3-L1 脂肪細胞分化 ( 以細胞內 TG 堆積為指標 ) 的影響 ...............................................................................47. 圖 4-2. 一般脂肪酸對 3T3-L1 分化 ( 以細胞內 TG 堆積為指標 ) 的影響.....48. 圖 4-3. 特殊脂肪酸對 3T3-L1 分化 ( 以細胞內 TG 堆積為指標 ) 的影響.....49. 圖 4-4. 特殊脂肪酸對 3T3-L1 分化 ( 以 G3PDH 活性為指標 ) 的影響 .........50. 圖 4-5. 顯微鏡下觀察特殊脂肪酸對 3T3-L1 分化的影響...................................51. 圖 4-6. 油紅染色觀察特殊脂肪酸對 3T3-L1 分化的影響...................................52. 圖 4-7. 特殊脂肪酸對 3T3-L1 前脂肪細胞存活的影響.......................................53. 圖 4-8. 特殊脂肪酸對 3T3-L1 前脂肪細胞之細胞毒性的影響...........................54. 圖 4-9. 特殊脂肪酸對 3T3-L1 成熟脂肪細胞 TG 堆積的影響 ...........................55. 圖 4-10 特殊脂肪酸對 3T3-L1 成熟脂肪細胞 G3PDH 活性的影響....................56 圖 4-11 特殊脂肪酸對 3T3-L1 成熟脂肪細胞脂解的影響...................................57 圖 4-12. c9,t11,t13-CLN 對 3T3-L1 各階段細胞凋亡的影響...............................58. 圖 4-13 天然抗氧化物對 3T3-L1 分化 ( 以細胞內 TG 堆積為指標 ) 的 影響 ...........................................................................................................59 圖 4-14 多酚類對 3T3-L1 分化 ( 以 G3PDH 活性為指標 ) 的影響 .................60.
(7) 圖 4-15 顯微鏡下觀察多酚類對 3T3-L1 分化的影響...........................................61 圖 4-16 油紅染色觀察多酚類對 3T3-L1 分化的影響...........................................62 圖 4-17 多酚類對 3T3-L1 前脂肪細胞存活的影響...............................................63 圖 4-18 多酚類對 3T3-L1 前脂肪細胞之細胞毒性的影響...................................64 圖 4-19 多酚類對 3T3-L1 成熟脂肪細胞 TG 堆積的影響 ...................................65 圖 4-20. curcumin 對 3T3-L1 各階段細胞凋亡的影響.........................................66.
(8) 縮. 縮. 寫. 對. 寫. 照. 表. 全. 名. Abs EtOH. Absolute ethanol. ACO. Acyl-CoA oxidase. ADD1 / SREBP. Adipocyte determination and differentiation factor 1 / Sterol regulatory element binding proteins. AMPK. AMP-activated protein kinase. AOM. Azoxymethane. aP2. Adipocyte specific fatty acid binding protein. BAT. Brown adipose tissue. Cdk2. Cyclin-dependent kinase 2. C/EBP. CCAAT / enhancer binding protein. CLA. Conjugated linoleic acid. CLN. Conjugated linolenic acid. CPT. Carnitine palmitoyltransferase. DEX. Dexamethasone. DMSO. Dimethyl sulfoxide. EGCG. Epigallocatechin gallate. ERK. Extracellular signal-regulated kinase. FAS. Fatty acid synthase. G3PDH. Glycerol-3-phosphate dehydrogenase. GLUT4. Glucose transpoter 4. HSL. Hormone sensitive lipase. I.
(9) 縮. 寫. 對. 縮 寫. 照. 表. 全. 名. IRS. Insulin receptor substrates. JNK. c-Jun N-terminal kinase. LPL. Lipoprotein lipase. MAPK. Mitogen-activated protein kinase. MIX. 1-methyl-3-isobutylxanthine. MUFA. Monounsaturated fatty acid. PA. Phytanic acid. PI3K. Phosphoinositide-3 kinase. PPAR. Peroxisome proliferators activated receptor. PUFA. Polyunsaturated fatty acid. SFA. Saturated fatty acid. STZ. streptozotocin. TZD. Thiazolidinedione. TG. triglyceride. UCP. Uncoupling protrein. II.
(10) 特殊脂肪酸及天然抗氧化物對 3T3-L1 脂肪細胞生合成影響. 摘要. 肥胖是全球普遍之健康問題,從天然食物中尋找抗肥胖成分,不論在疾病預防 及健康食品市場上均有其重要性。本研究旨在利用 3T3-L1 前脂肪細胞測試 2 種特 殊脂肪酸及 5 種天然抗氧化物,對脂肪細胞生合成(adipogenesis)的影響,選擇的二 種特殊脂肪酸為 α-eleostearic acid (c9,t11,t13 conjugated linolenic acid;c9,t11,t13-CLN) 及 phytanic acid (PA),前者存在於苦瓜中,後者在反芻動物肉和乳汁中可發現,5 種多酚為 EGCG、curcumin、resveratrol、apigenin、genistein 存在天然蔬果。分別在 3T3-L1 adipogenesis 的三個階段﹕前脂肪細胞增生、分化及分化末期階段處理細 胞,結果發現脂肪酸都具有促進 3T3-L1 分化的能力,但由於飽和程度不同,所以 促分化能力也會有所不同,分別是 SFA<MUFA<PUFA;因此在特殊脂肪酸部分特 別選用類似的脂肪酸作為對照。當 c9,t11,t13-CLN 與其對照(linolenic acid;C18: 3) 相 較,會顯著抑制前脂肪細胞增生;藉由細胞內 TG (triglyceride)、細胞內 G3PDH (glycerol-3-phosphate dehydrogenase) 活 性 及 油 紅 染 色 觀 察 , 發 現 於 分 化 階 段 c9,t11,t13-CLN 會顯著抑制脂肪細胞分化;在分化末期階段會抑制脂質生合成;除了 成熟脂肪細胞外 c9,t11,t13-CLN 於 100μM 會造成前脂肪細胞及分化階段細胞凋亡, 顯示 c9,t11,t13-CLN 的抑制 3T3-L1 增生及分化作用與誘導細胞凋亡有關;PA 與其 對照(palmitic acid;C16:0) 相較,在前脂肪細胞階段,不會造成細胞數目減少;在 分化階段則會抑制脂肪細胞分化;在分化末期階段 PA 會抑制成熟脂肪細胞 lipogenesis。多酚類的部分與 vehicle control 相比較,發現測試的 5 種多酚類均會抑 制增生且呈現 dose-response,其中 curcumin 於 50μM 會造成細胞毒性;分化階段 除了 EGCG 以外另外 4 種多酚類都會明顯抑制 3T3-L1 分化;在成熟脂肪細胞階段, 3 種多酚類(EGCG、Resveratrol、Genistein)會抑制細胞內 TG 堆積;除了前脂肪細胞 外,curcumin 會造成分化階段及分化末期階段細胞凋亡,顯示 curcumin 抑制 3T3-L1 分化和抑制成熟脂肪細胞 TG 堆積與誘導細胞凋亡有關。由以上可知,特殊脂肪酸 中 c9,t11,t13-CLN 與對照組(C18:3)相比,PA 與對照組(C16:0)相比,均會抑制脂肪細 胞增生、分化、成熟脂肪細胞脂質生合成,而以 c9,t11,t13-CLN 效果最強;在多酚 類當中則是以 Curcumin 之 anti-adipogenesis 效果最強,而這些成分具有抗肥胖潛力, 值得日後進一步研究。 關鍵字:α-eleostearic acid、phytanic acid、adipogenesis. III.
(11) The adipogenic effect of uncommon fatty acids and natural antioxidants on 3T3-L1 cell Tzu-Wen Lai Abstract Obesity is a worldwide health problem. It is important to find the compounds from food with anti-obesity effecct for preventing obesity-related disease and developing functional food. In this study, we investigated the anti-adipogenic effect of two uncommon fatty acids and five natural antioxidants on preadipocyte cell line 3T3-L1. The uncommon fatty acids we chose are α-eleostearic acid (c9,t11,t13 conjugated linolenic acid;c9,t11,t13-CLN) and phytanic acid (PA). c9,t11,t13-CLN is enriched in bitter gourd. Phytanic acid is abundant in ruminant meat and milk. The natural antioxidants we chose are including EGCG, curcumin, resveratrol, apigenin and genistein which are present in fresh vegetables and fruits. These compounds were incubated with 3T3-L1 at three different stages: preadipocyte proliferation, differentiation and terminal differentiation. Results show all common fatty acids could induce the differentiation of 3T3-L1. The potency of inducing adipocyte differentiation was as following, SFA<MUFA<PUFA. During the preadipocyte proliferation stage, c9,t11,t13-CLN reduced the cell number as compared with its control (linolenic acid;C18:3). Compared with C18:3, c9,t11,t13-CLN inhibited 3T3-L1 differentiation as indicated by cellular triglyceride 、 glycerol-3-phosphate dehydrogenase (G3PDH) activity and oil-red O staining. In terminal differentiation stage, c9,t11,t13-CLN reduced lipogenesis in mature adipocytes. c9,t11,t13-CLN induced apoptosis in the preadipocytes and differentiating adipocytes. Palmitic acid was used as a control group of PA. At preadipocyte proliferation stage, PA didn’t decrease cell number as compared with its control. At differentiation stages, PA suppressed TG accumulation and G3PDH activity in differentiating adipocytes as compared with palmitic acid. The same result were observed by oil-red O staining. At terminal differentiation stages, PA reduced lipogenesis in mature adipocytes. When compared with their vehicle control,five polyphenols reduced the preadipocyte cell number during the proliferation stage, especially for curcumin. At differentiation stage, curcumin, resveratrol, apigenin and genistein resulted in an inhibition of adipocyte differentiation. At terminal differentiation stage, EGCG, resveratrol and genistein suppressed TG accumulation in mature adipocytes. Curcumin induced apoptosis in the differentiating and mature adipocytes. It is concluded that uncommon fatty acids c9,t11,t13-CLN and PA may exert its anti-obesity effect by inhibiting preadipocyte proliferation, differentiation and terminal differentiation. c9,t11,t13-CLN was more. IV.
(12) potent than PA. Among the polyphenols we tested, curcumin was the most potent in anti-adipogenesis. Thus, these compounds merit further study for their action mechanism and their contribution in developing the health food for anti-obesity. Key words:α-eleostearic acid、phytanic acid、adipogenesis. V.
(13) 第一章 前言與研究目的. 第一章. 前言與研究目的. 肥胖已經成為全球性普遍之健康問題,近來也發現肥胖年齡層下降及兒童青少 年階層肥胖比例有逐漸增加的趨勢(1-3);可能與目前飲食逐漸西化、生活習慣的改 變 (如高能量飲食攝取增加、體能活動減少) 有關。脂肪組織 (adipose tissue) 的形 成和體內能量恆定調節有關,當能量過多時會導致脂肪細胞內三酸甘油酯貯存增 加,事實上肥胖與脂肪細胞數目增加 (hyperplasia) 和脂肪細胞肥大 (hypertrophy) 均有關,肥胖也成為各種疾病,如高血壓、血脂異常、胰島素抗性、第 2 型糖尿病、 癌症、心血管疾病、動脈粥狀硬化等危險因子(4)。 故此從天然食物中尋找抗肥胖成分,在全民健康疾病預防有其迫切性,在健康 食品市場上也有廣大商機,目前市面上的抗肥胖相關保健食品,如藤黃果酸 (Hydroxycitric acid;HCA)、共軛亞麻油酸 (conjugated linoleic acid;CLA) 以及中鏈 脂肪酸 (medium chain fatty acid),均有許多研究證明其功能及作用機制。 本研究乃利用 3T3-L1 前脂肪細胞嘗試探討食物中存在的特殊脂肪酸及天然抗 氧化物對脂肪細胞生合成 (adipogenesis) 的影響,以找出可能具有抗肥胖潛力之功 能成分。. 1.
(14) 第二章 文獻回顧. 第二章. 文獻回顧. 一、 脂肪細胞生合成 (Adipogenesis) 前脂肪細胞形成成熟脂肪細胞的過程稱之為脂肪細胞生合成 (adipogenesis), adipogenesis 可 分 為 三 個 階 段 , 包 括 為 前 脂 肪 細 胞 增 生 階 段 (preadipocyte proliferation)、分化階段 (differentiation) 及分化末期階段 (terminal differentiation), 分別說明如下:1) preadipocyte proliferation:前脂肪細胞 (preadipocyte) 會進行細胞 的生長使細胞數目增加,長滿後就進入 mitotic clonal expansion 再進行 1-2 次的有絲 分裂後,生長就會停止進入分化階段。2) differentiaiton:此時細胞型態有很大的改 變,由紡綞型改變成圓球形,並且有三酸甘油酯 (triglyceride;TG) 開始產生,除 了型態上的變化,基因表現也與前脂肪細胞大不相同,LPL (lipoprotein lipase) 可作 為分化早期指標(5),之後 C/EBPs (CCAAT / enhancer binding protein)和 PPARγ (peroxisome proliferators activated receptor γ ) 表 現 漸 增 加 (6) 。 3) terminal differentiation:分化末期不斷的 lipogenesis,促使 TG 累積使細胞內油滴變大,形成 成熟脂肪細胞,而成熟脂肪細胞為內分泌器官,會分泌許多脂肪細胞特定荷爾蒙, 如 leptin、adiponectin 等(7)。其過程如下圖 2-1 所示。 proliferation differentiation terminal differentition. preadipocyte. 圖 2-1 脂肪細胞生合成. adipocyte. Fig 2-1 Adipogenesis. 2.
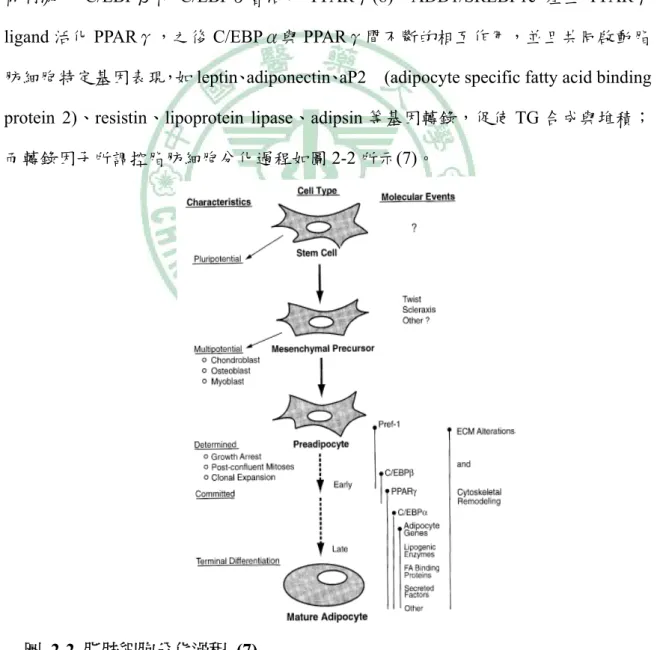
(15) 第二章 文獻回顧. 1、 脂肪細胞分化之轉錄調控 (Transcription cascade) 前脂肪細胞分化為成熟脂肪細胞,是由數種轉錄因子參與調控,而這些轉錄因 子也會相互作用形成 transcriptional factor cascade,參與因子包括 C/EBPs (CCAAT / enhancer binding protein)、ADD1/SREBP1 (adipocyte determination and differentiation factor 1 / sterol regulatory element binding proteins)、PPAR (peroxisome proliferators activated receptor) 。分化起始是由 C/EBPβ和 C/EBPδ啟動,於初期分化二天在荷 爾刺激下 C/EBPβ和 C/EBPδ會活化 PPARγ(8),ADD1/SREBP1c 產生 PPARγ ligand 活化 PPARγ,之後 C/EBPα與 PPARγ間不斷的相互作用,並且共同啟動脂 肪細胞特定基因表現,如 leptin、adiponectin、aP2. (adipocyte specific fatty acid binding. protein 2)、resistin、lipoprotein lipase、adipsin 等基因轉錄,促使 TG 合成與堆積; 而轉錄因子所調控脂肪細胞分化過程如圖 2-2 所示(7)。. 圖 2-2 脂肪細胞分化過程 (7) Fig 2-2 Stages of adipocyte differentiation.. 3.
(16) 第二章 文獻回顧. 2、PPARγ與脂肪細胞分化 Peroxisome proliferator-activated receptors (PPARs) 屬固醇類荷爾蒙核受器家族 (steroid hormone nuclear receptor family) 為一種轉錄因子,當與專一性 ligand 結合可 誘發下游基因的轉錄反應,而早期 PPAR 被選殖出來時,發現可被各種過氧化體增 生劑 (peroxisome proliferators;PP) 活化,而 PP 主要以各種 fibrates 類降血脂藥為主, 如 clofibrate、ciprofibrate 等,Issemann 等學者於 1990 年發現 arachidonic acid 可活 化 mPPAR,並誘發 PPAR 調節之基因轉錄,推測 eicosanoids 在 PPAR 傳訊中扮演重 要角色(9),Gottlicher 等學者也發現 C18:0、C18:1、C18:2、C18:3 等脂肪酸可活化 chimeric receptor 上的 rPPAR(10),直到 Lehamann 證實治療糠尿病藥物 TZDs 可與 mPPARγ結合,PPAR 才首次被認為是 ligand-activated transcription factor(11),而 PPAR 具有三種 isoform 分別是 PPARα、PPARβ和 PPARγ,其中 PPARγ專一表 現在脂肪組織,可受脂肪酸或藥物調控而影響脂質與葡萄糖代謝、脂肪細胞分化相 關基因表現,說明如圖 2-3 (12)。. 圖 2-3 PPARγ之活化與作用 (11) Fig 2-3 The activation of PPARγ and its action effect. 4.
(17) 第二章 文獻回顧. PPARγ最早是在哺乳動物中被發現(13);Spiegelman 學者與其研究團隊也在脂 肪細胞中發現脂肪細胞專一性之 PPARγ,並且發現 PPARγ會大量表現在脂肪組織 中且會於脂肪細胞生合成期間被誘導(14, 15);Tontonoz 等學者將纖維母細胞利用 Retrovirus 異位表現 PPARγ的 gain-of-function 方式,首次指出 PPARγ在脂肪細胞 生合成中扮演重要角色(16)。1999 年 Barak 及 Kubota 等學者利用基因剔除胚胎 PPAR γ的小鼠的模式,發現小鼠於胚胎 10-10.5 天期間由於胎盤不正常而導致胎死腹中 (17, 18)。為了解決 PPARγ-/- embryo 無法存活的問題,Rosen 等學者利用 wild-type ES cells 與完全剔除 PPARγ之 ES cells 製造出 chimeric mice (19);Barak 等學者利用正 常的 tetraploid cells 校正胎盤之缺失製造 chimeric embryo,解剖後發現沒有棕色或白 色脂肪組織(17),二個學者成功製造缺乏 PPARγ小鼠,並證明 PPARγ在脂肪組織 形成之重要性。 之後 He 等學者利用 cre-loxp 技術產生 PPARγ的 knockout mice,其皮下和副睪 脂肪塊、血漿 leptin 和 adiponectin 會減少、血漿游離脂肪酸與三酸甘油酯會增加 (20);Zhang 等人選擇性破壞 PPARγ2 發現會使小鼠腹膜後、腹股溝以及生殖器周 邊脂肪減少(21)。 PPARγ被認為是決定脂肪細胞分化重要因子,證據還包括 Lehmann 等學者發 現高度專一性 PPARγ ligand-抗糖尿病藥物 thiazolidinediones (TZDs),可以促進脂 肪細胞分化(11),另一 PPARγ之 ligand-BRL49653 也證實可促進鼠胚纖維母細胞 C3H10T1/2 分化為脂肪細胞(22)。. 5.
(18) 第二章 文獻回顧. 二、 3T3-L1 介紹 近來前脂肪細胞株如 3T3-L1、3T3F442A(23)、Ob1771(24)以及初代培養前脂肪 細胞 (stromal-vascular cell;SV cell) 為研究脂肪細胞分化常用之體外模式,3T3-L1 及 3T3F442A 都來自於 Swiss 3T3 cell,而 Swiss 3T3 cell 是由 Green 等學者由小鼠胚 胎纖維母細胞 (fibroblasts) cloning 出來具有高度分化為成熟脂肪細胞潛力的前脂肪 細胞株(25)。 在 in vitro 模式為促使前脂肪細胞分化為脂肪細胞,常在細胞長滿後更換為分化 培養基 (differentiation medium),其中添加數種化學藥品或荷爾蒙,以促進分化發 生,以下就這些成分說明: 1、dexamethasone (DEX) dexamethasone (DEX) 屬於 glucocorticoids,Rubin 等學者利用 DEX 及 MIX (methylisobutylxanthine)作為 3T3-L1 cells 分化的 differentiation cocktail 成分(26), DEX 可透過促使 C/EBPβ和 C/EBPδ形成 heterodimers,來活化 PPARγ表現(8, 27);另外在 PPARγ和 C/EBPα transfection 研究也發現 DEX 可誘導 preadipocyte 分化為 adipocyte(28)。 2、methylisobutylxanthine (MIX) MIX 和 DEX 一樣都會促進脂肪細胞的分化作用,它是透過增加 C/EBPβ表現, 抑制 phosphodiesterase 及競爭抑制 A1 adenosine receptor,也透過阻斷 inhibitory regulatory protein G1 來刺激 adenyl cyclase 活性,表示 MIX 可透過增加 cAMP 來促 進分化(7)。 3、insulin insulin 以促進 deoxylglucose uptake,並將 glucose 轉換成 CO2 及 lipid,另外由 於 p38 mitogen activated protein kinase (MAPK) 可以促進 preadipocyte 自發性分化作. 6.
(19) 第二章 文獻回顧. 用(29),加上 insulin 可以刺激 p38 MAPK 活性(30),所以可以促進分化;而在分化 後的第三或第四天起 insulin binding 活性會增加到 6-10 倍(26)。 4、biotin 為脂肪酸合成過程中之關鍵酵素 ACC (acetyl-CoA carboxylase) 的 cofactor,故 添加 biotin 可促進脂肪酸合成,促進分化作用。 三、 食物成分與脂肪細胞生合成相關研究 (一) 脂肪酸類 1、 一般脂肪酸 (Common fatty acid) Medium-chain triglycerides (MCT) 餵食動物會降低脂肪細胞數目及大 小,因為 MCFA 可增加粒線體β-oxidation。另外給予大鼠 MCTs 會降低 大鼠脂肪塊、向下調節 PPARγ、C/EBPα及其下游基因,改善 insulin 敏 感性及葡萄糖耐受性,類似的結果在以 MCFA 處理的 SV cell 中也發現, 與 control 組 long-chain fatty acid (LCFA) 比較,MCFA 會降低 PPARγ和 C/EBPα, 在 3T3-L1 脂肪細胞上也可發現, C10:0 和 C18:1 比較,其促 分化效果較差,且會抑制 3T3-L1 分化及成熟脂肪細胞的 lipogenesis(31)。 同樣是 SFA 的 C8:0 處理 3T3-L1 也會抑制 adipogenic gene,如 PPAR γ、C/EBPα、SREBP-1c,抑制分化(32);visfatin 為內臟脂肪分泌為類 insulin 蛋白質,C16:0 及 C18:1 會向下調節 3T3-L1 前脂肪及成熟脂肪細胞 visfatin mRNA 表現,降低 insulin 促進 glucose uptake 造成 insulin 阻抗(33)。 Amri 等學者以各種脂肪酸處理 Ob1771 脂肪細胞,發現脂肪酸皆會促 進 aP2 gene 表現,且 LCFA (≧C12) 如 C16:0、C18:0 比 SCFA 佳,也發現 PUFA (如 C18:3) 比 MUFA (如 C18:1) 好,SFA 最差(34)。 在 PUFA 的動物實驗證據,Xu 等學者則是給予大鼠魚油,發現 FAS mRNA 表現降低,ACO mRNA 表現增加(35),降低脂肪組織及抑制 rodents. 7.
(20) 第二章 文獻回顧. 肥胖(36);細胞實驗證據有 Hsu 等學者利用豬隻皮下脂肪作初代脂肪細胞 培養,投予 PUFA 包括有 arachidonic acid (AA)、c9,t11 conjugated linoleic acid (CLA)、Docosahexaenoic acid (DHA;C22:6),發現都會增加 FAS、ADD1 mRNA 降解,降低 FAS、ADD1 mRNA 以及促進 ACO mRNA 表現,其中 又以 DHA 100μM 效果最強(37)。另外以 DHA 50-200μM 處理 3T3-L1, 發現會抑制 3T3-L1 分化,增加脂解作用(38),同樣的若給予 t10,c12 CLA 也會抑制 3T3-L1 分化(39)(40)。 2、 特殊脂肪酸 (Uncommon fatty acid) (1) Conjugated linolenic acid conjugated linolenic acid (CLN) 為 18 個碳含有 3 個共軛雙鍵的不飽和 脂肪酸(結構請見表 2-1),存在於特殊種籽油中,如苦瓜 (karela oil) 及桐 油 (tung oil)中含有 60-70% c9,t11,t13 CLN (α-eleostearic acid) ,石榴油 (pomegranate seed oil) 中含 72% c9,t11,c13 CLN ( punicic acid ),樟樹籽油 (Catalpa seed oil) 含 31% t9,t11,c13 CLN ( catalpic acid )、金盞花籽油 ( pot marigold seed oil ) 含 33% t8,t10,c12 CLN ( calendic acid )(41)。 c9,t11,c13 CLN 為苦瓜中主要且特殊的脂肪酸,在生理方面已證實具 有抗癌、降血糖、降體脂等功能。在抗癌方面證據包括,Kohno 等學者給 予 AOM (azoxymethane) 誘導大腸癌之大鼠富含 CLN 苦瓜籽油,發現 0.01-0.1%苦瓜籽油可以增加大腸粘膜上的 PPARγ protein,並且抑制 colon carcinogenesis(42, 43);Yasui 等學者也證實苦瓜中 CLN 可以誘導人類 大腸癌 Caco-2 細胞凋亡(41)。不只苦瓜中 CLN,石榴中的 CLN 也顯示抗 癌功能,如 Kohno 等學者以 AOM 誘發大鼠大腸癌再給予富含 CLN 的石 榴油萃製物 (Pomegranate seed oil ; PGO),發現 PGO 0.01%-0.1%可抑制腫 瘤數量及抑制 adenocarcinoma 發生(44)。. 8.
(21) 第二章 文獻回顧. c9,t11,c13-CLN 降體脂之間接證據包括將苦瓜汁添加於飼料中,發現 可透過增加肝及肌肉 CPT-1 (carnitine palmitoyltransferase 1) 和 acyl-CoA dehydrogenase 活性來增加脂肪酸氧化來降低體脂(45),另外苦瓜具有降低 高糖血症(46, 47),Koba 等學者利用動物模式將 CLN 複合物 (conjugated diene 及 triene) 添加於飲食中,發現 CLN 可降低白色脂肪組織含量(48)。 此外 c9,t11,c13-CLN 在細胞實驗研究包括,Chao 等學者利用苦瓜萃取 物處理肝細胞 H4IIEC3,發現增加肝細胞中 ACO 活性及 mRNA 表現(49), Chuang 等學者同樣利用肝細胞模式,發現苦瓜可透過活化肝臟 PPARα, 促 進 肝 細 胞 中 ACO 活 性 及 其 gene 表 現 的 功 能 成 分 是 苦 瓜 中 的 c9,t11,c13-CLN(50);Nishimura 等學者使用苦瓜萃取出的 c9,t11,c13-CLN 處理 3T3-L1 脂肪細胞,發現 200μM 會誘導 3T3-L1 preadipocyte 細胞凋亡 (51)。 (2) Phytanic acid Phytanic acid (3,7,11,15-tetramethyl hexadocanoic acid;C20H40O2) 為具 有 4 個甲基的支鏈飽和脂肪酸 (結構請見表 2-1),綠色植物中富含大量的 chlorophyll,含有 phytol ( 3,7,11,15-tetramethylhexadec-2-en-1-ol ) 側鏈,經 由反芻類動物瘤胃中之微生物可將葉綠素中一半的 phytol 釋出,並且代謝 成 phytanic acid。因為人體無法將葉綠素中 phytol 釋出,經由放射性標定 chlorophyll 在人類及大鼠的消化道中消化程度實驗,發現食入的 chlorophyll 有 95%以上會排出體外(52, 53),所以透過外源性之飲食來源,如反芻類動 物脂肪、魚類及乳製品,人類可能攝取到 phytanic acid,或者富含葉綠素 食物經加工 (如鹼處理) 可能會使葉綠素之酯鍵斷裂釋出 phytol,人吃入此 類食品後經過體內氧化還原酶之作用轉成 phytanic acid ( PA ) (54)。 PA 經飲食進入體內必須經由過氧化小體進行 α-oxidation 生成 pristanic. 9.
(22) 第二章 文獻回顧. acid (2,6,10,14-tetramethlpentadecanoic acid;C19H38O2),再行 β-oxidation 生 成 propionyl-CoA;phytanoyl-CoA hydroxylase (PAHX 或 PhyH) 是 phytanic acid 進行 α-oxidation 第一個酵素,患有 Refsum、過氧化小體合成缺陷、 rhizomelic chondrodysplasia punctata type I 及 Zellweger syndrome (ZS) 的病 人都缺乏此一酵素,造成 PA 累積(55)(56, 57)。 已知 PA 為 PPAR α 活化劑,會增加大鼠初代肝細胞中 GLUT1、GLUT2 mRNA 表現,增加 glucose uptake(58)。另外在脂肪細胞方面,pristanic acid 及 PA 二者,只有 PA 可促進 UCP-1 (uncoupling protein-1) gene 轉錄,促進 棕色脂肪細胞分化(59),並且共同作用誘導棕色脂肪細胞分化特定 gene 表 現,如 aP2 (adipocyte lipid-binding protein)、LPL、GLUT4、cytochrome c oxidase subunit II 等(60),活化 BAT (brown adipose tissue) 生熱作用。 Schluter 等學者使用 3T3-L1 前脂肪實驗發現,與 vehicle control 相比,PA 可促細胞分化作用(61)。 (二) 天然抗氧化物 1、 Epigallocatechin Gallate (EGCG): EGCG 茶中含量豐富 (結構請見表 2-2),茶葉是來自於 Camellia sinensis 經由不同的製程,可分為未醱酵的綠茶、半醱酵的烏龍茶、包種茶, 以及完全醱酵的紅茶和普耳茶,而東方國家比較常飲用綠茶、烏龍茶和普 耳茶,西方國家則是偏好紅茶(62, 63)。經由醱酵過程會使茶葉中的成分改 變,紅茶及普耳茶中的單體兒茶素減少,catechins 會經由多酚氧化酶形成 quinone , 再 濃 縮 形 成 聚 合 物 theaflavins 如 TF-1 (theaflavin) 、 TF-2a (. theaflavin-3-gallate). ,. TF-2b. (theaflavin-3’-gallate). 、. TF-3. ( theaflavin-3,3’-digallate),和 thearubigins。(64)。兒茶素成分分析其中 (-)-epicatechin (EC)、(-)-epigallocatechin (EGC)、(-)-epigallocatechin gallate. 10.
(23) 第二章 文獻回顧. (EGCG) 三種成分,在綠茶、烏龍茶含量高於紅茶和普耳茶(65)。 EGCG 生 理 功 能 相 關 研 究 包 括 有 , 綠 茶 兒 茶 素 為 polyphenolic flavonoids 過去曾被稱為維生素 P,其特殊結構於 1952 年由 Roberts 等學者 發現,是未醱酵茶中主要的成分,綠茶中兒茶素以 EGCG 為主要活性成分, 已被證實具有抗癌、抗氧化(66)、抗脂質過氧化能力(67, 68),降低體重及 體脂(69)(70)(71),增加肝臟β-oxidation 活性增加脂質代謝(72)。 人體試驗方面 Nakagawa 等學者給予健康受試者綠茶萃取物錠(內含 525mg EGCG、17.5mgEGC)90 分鐘後,發現血漿 EGCG 及 ECG 濃度分別 為 4.3、0.3μM(73),另外口服綠茶萃取物 20mg / kg body weight 溶於 200mL 溫水,約 1 到 2 小時後,血中 EGCG、EGC、EC 濃度分別是 0.17μM、0.5 μM、0.42μM (74)。 降體脂及血脂之動物實驗證據如下,在幾種不同茶中降低肥胖 SD 大 鼠體重及降低血清三酸甘油酯方面,烏龍茶、普耳茶優於綠茶、紅茶,又 以普耳茶最好,但降低膽固醇則是普耳茶、綠茶比烏龍茶、紅茶好(65)(75); 對於飲食誘導肥胖的 C57BL/6J mice,若是給予綠茶兒茶素搭配運動雙管齊 下,對於降低體重、體脂(76)和增加肌肉脂肪酸氧化(77),以及降低高胰島 素血症及 hyperleptinemia 發展是有加成作用(76)。 細胞實驗證據包括有,Chiang 等學者之前證實給予大鼠普耳茶後,發 現不僅降低體重及血脂,也抑制了肝臟 FAS (fatty acid synthase) 表現,所 以作者投予普耳茶萃取物於肝細胞 HepG2 cell ,發現普耳茶是透過 PI3K/Akt 及 JNK (c-Jun N-terminal Kinase) 路徑來抑制 FAS mRNA 及其蛋 白質(75);另外 Waltner 學者同樣使用肝細胞 H4ΠEC3,投予 EGCG 發現 會 抑 制 insulin 誘 導 的 IR (insulin receptor) 及 IRS-1 (insulin receptor substrate-1) 磷酸化作用(78);除此之外也有學者將烏龍茶萃取物投予脂肪. 11.
(24) 第二章 文獻回顧. 細胞,發現茶中 caffeine 會增強脂肪細胞正腎上腺素作用,促進脂肪細胞 脂解作用(79); 此外 EGCG 會抑制 3T3-L1 細胞增生(80),EGCG 100μM 會透過活化 AMPK (AMP-activated protein kinase) (81)和向下調節 resistin mRNA 表現來 抑制 3T3-L1 adipocyte 分化(82) (83),並且誘導成熟 3T3-L1 細胞之細胞凋 亡(84)。 2、 Procyanidins 原花青素 (Procyanidins) 廣泛地存於紅酒、葡萄、可可、蘋果中,是 flavonoids 的一種,由許多的 catechin 聚合形成 oligomeric form(結構請見表 2-2) ,具有抗心血管疾病功能,近來也有和抗肥胖的相關研究。 其生理功能之動物實驗包括﹕給予大鼠口服 procyanidins 200 mg / kg BW,約 1 小時後在血中及肝臟中就可以偵測到體內 procyanidins 達最大濃 度 14 mg / L(85),而葡萄籽中的 procyanidins 可以改善糖尿病的氧化狀態 (86-89),由於 procyanidins 具有 insulinomimetic 活性,可經由 PI3K 及 p38MAPK 路徑並且增加 GLUT4 (glucose transporter-4) 轉位到膜上,促進 葡萄糖汲取並傾向 glycerol 合成(90),降低以 STZ 誘發糖尿病大鼠血糖, 具有抗高血糖症功能(90)。 在細胞實驗證據包括有,Procyanidins 具有類似 TNF-α的功能,會向 下調控 adipocyte marker 如 glycerol-3-phsphate dehydrogenase (G3PDH)、 hormone sensitive lipase (HSL) 活性及 PPSRγmRNA 表現(91),抑制脂肪 細胞分化(92),另外也會增加 3T3-L1 脂肪細胞脂解速率(93)。 3、 Curcumin 微黃色,常見薑科植物之薑黃 (Curcuma longa L.) 中 (結構請見表 2-2) ,在亞洲地區屬於傳統醫療,用於治療發炎及其他疾病,市面薑黃粉. 12.
(25) 第二章 文獻回顧. 末每 100g 含有 2.71-5.18g curcuminoids,市售咖哩粉末每 100g 約含有 0.34-0.47g 薑黃粉末(94)。 在生理方面 curcumin 具有抗氧化(95, 96)、抗癌(97, 98)等功能,Asai 等學者於飼料中添加 1% curcumin 複合物給予 SD 大鼠,可降低體脂、肝 脂、增加肝臟 ACO 活性,(99),Nishiyama 等學者,給多 type2 DM KK-Ay mice 0.1-0.5 g / 100 g diet 薑黃萃製物 4 週後可以降低血糖,薑黃酒精萃製 物可促進人類脂肪細胞分化,另外在 GAL4-PPARγ chimera assay 中發現 薑黃萃製物 5 μg / mL (約為 13.5μM ) 為 PPARγligand(100)。 4、 Naringenin 常見於柑橘類水果及葡萄柚中 (結構請見表 2-2) ,在動物實驗方面 Lee 等學者以 naringenin (0.1%,wt/wt) 合併高脂飲食餵食大鼠,發現可降 低餵食高脂飲食之大鼠血液及肝臟膽固醇(101)。 根據 1997 年美國農業部調查,個人每日柑橘類攝取量約為 68 g,而其 中 59g 來自於果汁,每 100 g 葡萄柚約含有 78 mg naringenin;若食用一個 中 型 葡 萄 柚 相 當 於 攝 取 200 mg naringenin , 血 中 濃 度 達 0.7-14.8 μ M(102);在芬蘭 naringenin 攝取量約為 8.3 mg / day; Erlund 等學者以 HPLC 分析柑橘類及葡萄柚水果中 naringenin 含量,發現柑橘類水果中 naringenin 含量為 151μM (41 mg / L),葡萄柚中 naringenin 含量為 1283μM (349 mg / L),此外又將柑橘類及葡萄柚果汁給予健康受試者 8 mL / kg 每日給予一 次,柑橘類及葡萄柚中 naringenin 在血液中濃度高峰分別為 0.6 ± 0.4 μM 和 6.0 ± 5.4 μM,若一次給予 20 oz. (約 600. mL ) 葡萄柚汁在人體血. 液中 naringenin 高峰為 6μM(102)。 接著 Erlund 等學者又以 6μM 處理成熟脂肪細胞,發現則可降低 insulin 刺激的 glucose uptake 作用,並且維持 24 小時(103),另外 Harmon 等學者. 13.
(26) 第二章 文獻回顧. 使用 3T3-L1 脂肪細胞,投予 naringenin 100μM,發現可以抑制 3T3-L1 preconfluent preadipocyte 增生(104)。 5、 Ascorbic acid 為水溶性維生素 (結構請見表 2-2) ,在細胞實驗相關研究包括有, Kawada 等學者投予 Ascorbic acid 10 - 200μM 於 3T3-L1,則細胞內 TG 含 量增加,促進 3T3-L1 分化(105),同樣 3T3-L1 為模式,給予 0.2 mM ascorbic acid phosphate 發現也會促進 3T3-L1 分化(106)。 由於治療 HIV 藥物 Nelfinavir 會造成 AIDS 病患產生 lipodystrophic syndrome,若合併給予 100mM ascorbate 及 Nelfinavir 於 3T3-F442A 脂肪細 胞,則可改善 70%細胞壞死的情形(107)。 6、 Resveratrol 3,5,4’-trihydroxystibene 存在於葡萄皮、紅酒 (結構請見表 2-2) ,在動 物實驗相關研究包括有,給於高齡 (約 48 週齡大) C57BL/6NIA mice 餵食 22.4 mg / kg / day resveratrol,則可增加 insulin 敏感性、延長壽命改善健康 (108)。 在細胞實驗相關研究有,Picard 等學者發現 50μM resveratrol 可抑制 3T3-L1 成熟脂肪細胞 lipogenesis,增加脂解作用(109);Backesjo 等學者發 現 50μM resveratrol 可抑制 C3H10T1/2 間葉細胞形成的 adipocyte 中 PPAR γ,活化 sirt1 阻斷 adipocyte 的發展(110)。 7、 Apigenin 芹菜配醣體,常見於荷蘭芹、甘菊、車前草等 (結構請見表 2-2) ,生 理實驗相關研究發現,若人體攝取富含 apigenin 的食物-川燙荷蘭芹 2 g ( 濃 度約為 65.8μM ) 72 小時後,血中 apigenin 濃度為 127 nM,而 24 小時後 尿中濃度為 144 nM(111)。. 14.
(27) 第二章 文獻回顧. 動物實驗相關證據包括,Yano 等學者給予 C57BL/6N mice apigenin 則 會抑制 Ig E 及發炎相關 cytokines,如 TNFα 等(112);給予 mice 高脂飲食 誘導肥胖後,再給予 5%柳樹葉 ( Szlix matsudana Koidz ) 萃取出的多酚類 化合物,其中包含有 apigenin-7-O-beta-D-glucopyranoside 可降低肝中總膽 固醇,抑制小腸吸收脂質(113, 114),增強脂肪細胞的脂解作用(113, 115)。 另外 apigenin 也會造成 2 種細胞之細胞凋亡證據如下,Chiang 等學者 發現 apigenin 會經由 p53 路徑誘導 p21 表現,使細胞週期停滯於 G2/M 期, 來使 Hep G2 cell 細胞凋亡(116);在小鼠成骨細胞株 MC3T3-E1 細胞也是 如此(117)。 8、 Quercetin 5,7,3’,4’-hydroxyflavanol 常見於洋蔥、茶 (如蕎麥茶)、蘋果等食物中(結 構請見表 2-2) , 在初代脂肪細胞實驗相關研究包括,Shisheva 等學者使 用大鼠初代脂肪細胞投予 47μM quercetin,發現可以抑制大鼠初代脂肪細 胞約 50% lipogenesis 反應,110μM quercetin 則可抑制 100%(118);10-250 μ M quercetin 可增強 epinephrine 誘 發 的 脂 解 作 用 (119) , 10-100 μ M quercetin 會抑制 glucose uptake(120) ;另外 Kim 等學者則是使用了人類脂 肪 細 織 來 源 之 初 代 細 胞 (human adipose tissue-derived stromal cells ; hADSC),投予 2-10μM quercetin 後發現也會抑制增生,但促進分化作用 (121);此外在 3T3-L1 脂肪細胞方面,Hsu 等學者使用 50-200μM quercetin 發現 quercetin 會透過向下調節 PARP (Poly-ADP-ribose polymerase) 及 Bcl-2 蛋白質,增加 caspase-3、Bax 等蛋白質,來促進 preadipocyte apoptosis(122)。 9、 Genistein 常見於大豆 (結構請見表 2-2) ,genistein 0.1-1 mM 可抑制大鼠初代脂 肪細胞 lipogenesis (123)及抑制 leptin 分泌(124),在 human primary bone. 15.
(28) 第二章 文獻回顧. marrow stromal cells 方面,genistein 也會抑制 adipogenic differentiation(125)。 在 3T3-L1 脂肪細胞方面,100μM genistein 會抑制 preconfluent 及 mitotic clonal expansion 階段增生,抑制分化(126)並促進脂解作用(104),抑 制 glucose uptake(127)。 而若攝取含有高量 genistein (90 mg),體內 genistein 濃度也只會有 0.6 μM(128)。. 16.
(29) 第二章 文獻回顧. 表 2-1 脂 化. 合. 肪. 酸. 物. 化. 學 結. Oleic acid. Linoleic acid. α-Linolenic acid. Stearic acid. Palmitic acid. 17. 結. 構 構.
(30) 第二章 文獻回顧. 表 2-1 脂 化. 合. 肪. 酸. 物. 化. 學 結. c9,t11,t13-conjugated linolenic acid (α-eleostearic acid; 9Z,11E,13E-octadecatrienoic acid; c9,t11,t13-CLN). Phytanic acid (PA). 18. 結. 構 構.
(31) 第二章 文獻回顧. 表 2-2 天 化. 合. 然. 抗. 氧. 化. 物. 物. 化 結. Epigallocatechin gallate (EGCG). Curcumin. Procyanidin. catechin. naringenin. Ascorbic acid. 19. 學. 結 構. 構.
(32) 第二章 文獻回顧. 表 2-2 天 化. 合. 然. 抗. 氧. 化. 物. 物. 化 結. Resveratrol. Apigenin. Quercetin. Genistein. 20. 學. 結 構. 構.
(33) 第三章 材料與方法. 第三章. 材料與方法. 一、實驗假說 食物中特殊脂肪酸包括 c9,t11,t13 conjugated linolenic acid (α-eleostearic acid; 9Z,11E,13E-octadecatrienoic acid;CLN) 和 phytanic acid (PA),以及天然抗氧化物中 或許具有 anti-adipogenesis 功能之成份,可以抑制 3T3-L1 前脂肪細胞增生、分化及 成熟脂肪細胞脂質堆積,或促進(前)脂肪細胞凋亡。. 二、實驗設計 實驗可為二部分,第一部分為脂肪酸對 3T3-L1 脂肪細胞脂質生合成影響,第 二部分為天然抗氧化物對 3T3-L1 脂肪細胞脂質生合成影響,說明如下:. - 21 -.
(34) 第三章 材料與方法. 第一部分:脂肪酸對 3T3-L1 脂肪細胞脂質生合成影響. 測試各種溶劑及正對照 TZD 對 3T3-L1 脂肪細胞分化的影響. Common fatty acid 對脂肪細胞 differentiation 的影響. Uncommon fatty acids 對脂肪細胞 adipogenesis 的影響. Preadipocyte 階段. . MTT assay LDH assay Apoptosis assay. Differentiation 階段. . TG assay G3PDH assay Oil-Red O stain Apoptosis assay. Terminal differentiation 階段. . TG assay G3PDH assay Lipolysis assay Apoptosis assay. - 22 -.
(35) 第三章 材料與方法. 第二部分:天然抗氧化物對 3T3-L1 脂肪細胞脂脂質生合成影響. Nature antioxidants 對脂肪細胞 adipogenesis 的影響. Preadipocyte 階段. . MTT assay LDH assay Apoptosis assay. Differentiation 階段. . TG assay G3PDH assay Oil-Red O stain Apoptosis assay. Terminal differentiation 階段. . TG assay Apoptosis assay. - 23 -.
(36) 第三章 材料與方法. 三、細胞培養 1、 細胞株 前脂肪細胞株 3T3-L1 購自食品工業發展研究所菌種中心/國家衛生研究院 細胞庫,菌種中心編號 BCRC 60159,細胞株來源 ATCC CL-173,組織來源﹕ Mouse embryo 2、 培養基組成與配製 先將 Dulbecco’s Modified Eagle Medium (DMEM;Gibco)粉末溶解,加入 1.5 g sodium bicarbonate ( sigma) ,再以 1N HCl 調整 pH 值為 7.1-7.2 之間,定 量 1L 後以過濾膜 (0.22μm) 過濾。medium 為配好的 DMEM 再加入 10% Bovine serum (BS) (Gibco) 、 2 %. L-glutamine (Biological industries) 及 1x. antibiotic-antimycotic (內含 penicillin G sodium、streptomycin sulfate、amphotericin B;Gibco),此為 maintain medium (10%BS-DMEM)。 3、 細胞繼代培養 細胞解凍後以 4×105cells 接種於 75T flasks,並加入 20% BS-DMEM,待隔 日細胞完全趴附後去除舊的 medium,加入新的 10% BS-DMEM,於 37℃、5% CO2 下培養,每 3-4 天進行繼代培養。 繼代培養時為待細胞約長滿 (約 90 % confluence),移去培養基以 1X PBS 清洗細胞二次後,加入 1mL trypsin-EDTA 潤濕所有細胞後,放入 37℃、5% CO2 下作用約 8-10 分鐘後取出,加入 10% BS-DMEM 9mL 中和反應並使細胞脫落 後,經由 trypan blue solution 染色,以細胞計數盤在倒立式顯微鏡下計數細胞數 後,再將細胞種入新的 75T flask。解凍後細胞均進行繼代培養 2 代後才進行實 驗,實驗使用的繼代數均控制於 p=U+8+11 至 p=U+8+13。. - 24 -.
(37) 第三章 材料與方法. 四、誘導 3T3-L1 脂肪細胞分化 根據許多文獻資料,以 3T3-L1 前脂肪細胞株建立脂肪細胞分化流程 1、 藥品配製: (1) 0.25 mM Dexamethasone (DEX) :1000x stock 藥品. 需要量. conc.. Dexamethasone (SIGMA D-4902). 0.098 g. 0.25 M. DMSO 1 mL 先將 0.098 g DEX powder 溶於 1 mL DMSO 中,即得 0.25 M DEX 藥品. 需要量. Stock conc.. Working conc.. 0.25 M DEX. 10μL. 0.25 mM. 0.25μM. DMSO 10 mL 將 0.25 M DEX 以 DMSO 稀釋為 0.25 mM DEX,分裝於滅菌 eppendorf 中,貯 於-20℃ (2) 0.5 M 3-isobutyl-1-methyl-xanthine (MIX):1000x stock 藥品. 需要量. Stock conc.. Working conc.. 3-isobutyl-1-methyl-xanthine (SIGMA I5879-1G). 0.111 g. 0.5 M. 0.5 mM. DMSO 1 mL 將 0.111 g MIX powder 溶於 1 mL DMSO 中,分裝於滅菌 eppendorf 中,貯於-20℃ (3) 100 μg / mL Biotin:1000x stock 藥品. 需要量. Biotin (SIGMA B-4501) 二次水. 0.01 g. Stock conc. 100μg/ mL. Working conc. 0.1μg / mL. 100 mL 將 0.01 g biotin powder 溶於 100 mL 二次水中,以 0.2μm filter 過濾分裝於滅菌 eppendorf 中,貯於-20℃. - 25 -.
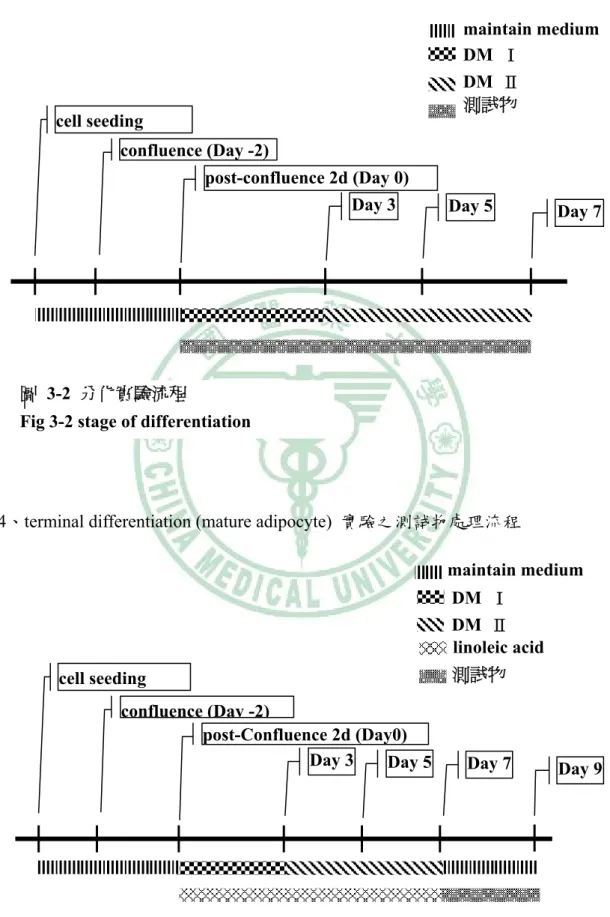
(38) 第三章 材料與方法. (4) 10 mg / mL Insulin:1000x stock 藥品. 需要量. Stock conc.. Working conc.. Insulin (SIGMA I6634). 0.1 g. 10 mg / mL. 10μg / mL. 0.01N HCl 10 mL 將 0.1 g insulin powder 溶於 10 mL 0.01N HCl 中,以 0.2μm filter 過濾分裝於滅 菌 eppendorf 中,貯於-20℃ 2、 培養基: 在分化實驗中所使用的分化 medium 可分為以下二種 Differentiation medium Ⅰ(DM Ⅰ):10 % BS-DMEM 含 0.25μM DEX、0.5mM MIX、10μg/mL Insulin 及 0.1μg/mL Biotin Differentiation medium Ⅱ(DM Ⅱ):10 % BS-DMEM 含 10μg/mL Insulin、0.1 μg/mL Biotin 3、 方法﹕ 誘導分化的實驗,首先將細胞以 2×105 cells/well 接種於 12-well plate 之後,待 細胞長滿後再培養 2 天,就把 maintain medium 換成 DM Ⅰ( 內含 DEX、MIX、 insulin、biotin ),培養 3 天去除舊的 medium 再換成 DM Ⅱ(內含 insulin、biotin)每 2 天更換 medium 至第 7 天。 而 terminal differentiation 實驗,則是將細胞接種於實驗所需的 plate 之後,待細 胞長滿後再培養 2 天,就把 maintain medium 換成 DM Ⅰ(內含 DEX、MIX、insulin、 biotin ) 額外加入 100μM linoleic acid,培養 3 天去除舊的 medium 再換成 DM Ⅱ (內 含 insulin、biotin),此時仍然添加 linoleic acid 至第 7 天誘導細胞形成成熟脂肪細胞。. - 26 -.
(39) 第三章 材料與方法. 五、各種測試物之處理 1、測試物種類 (1) 脂肪酸測試物: Oleic acid. Cayman. Linoleic acid. Cayman. α-linolenic acid. Sigma. Palmitic acid. Cayman. Stearic acid. Sigma. c9,t11,t13 conjuated linolenic acid (CLN). Cayman. Phytanic acid (PA). Cayman. 脂肪酸測試物以絶對酒精配製成 100mM 之 stock solution 進行實驗,實 驗組劑量為 10μM、50μM、100μM、200μM,vehicle control 為絶對 酒精,劑量為 0.01%、0.05%、0.1%、0.2% (2) 天然抗氧化物類測試物: Epigallocatechin gallate. Cayman. Curcumin. Cayman. Catechin. Cayman. Naringenin. Sigma. Ascorbic acid. Sigma. Resveratrol. Cayman. Apigenin. Sigma. Quercetin. Cayman. Genistein. Cayman. - 27 -.
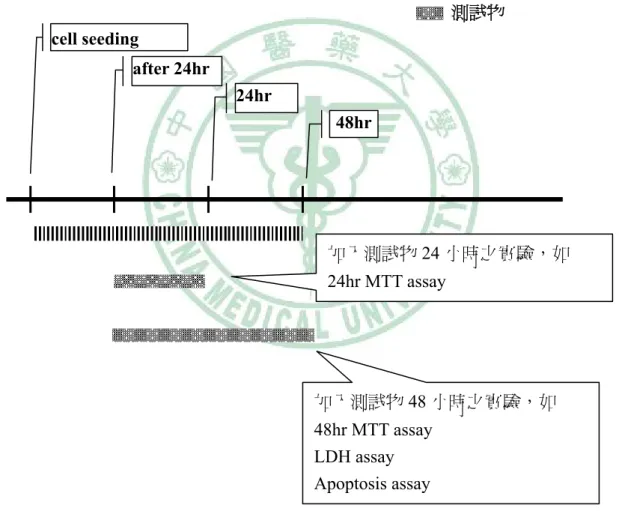
(40) 第三章 材料與方法. 天然抗氧物測試物以 DMSO 配製成 100mM 之 stock solution 進行實驗, 實驗組劑量為 10μM、50μM、100μM,vehicle control 為 DMSO,劑 量為 0.01%、0.05%、0.1%. 2、前脂肪細胞增生實驗之測試物處理流程 maintain medium 測試物 cell seeding after 24hr 24hr 48hr. 加入測試物 24 小時之實驗,如 24hr MTT assay. 加入測試物 48 小時之實驗,如 48hr MTT assay LDH assay Apoptosis assay 圖 3-1 前脂肪細胞增生實驗流程 Fig 3-1. stage of preadipocyte proliferation. - 28 -.
(41) 第三章 材料與方法. 3、differentiation 實驗之測試物處理流程 maintain medium DM Ⅰ DM Ⅱ 測試物. cell seeding confluence (Day -2) post-confluence 2d (Day 0) Day 3. Day 5. Day 7. 圖 3-2 分化實驗流程 Fig 3-2 stage of differentiation. 4、terminal differentiation (mature adipocyte) 實驗之測試物處理流程 maintain medium DM Ⅰ DM Ⅱ linoleic acid 測試物. cell seeding confluence (Day -2) post-Confluence 2d (Day0) Day 3 Day 5. 圖 3-3 分化末期實驗流程 Fig 3-3 stage of terminal differentiation - 29 -. Day 7. Day 9.
(42) 第三章 材料與方法. 六、 MTT 染色法細胞存活率測定 1、. 原理: MTT (3-[4,5-dimethylthiazol-2-y1]-2,5-diphenylterazolium bromide)是種活細. 胞染色法,活細胞粒線體內的 dehydrogenase 會將 MTT 之 tetrazolium ring 打斷 形成 formazan,此時 MTT 由黃色轉變藍色,再用 acid isopropanol 將 formazan 溶出,測定 540nm 吸光值,判定細胞存活數。 2、. 藥品配製: (1) MTT stock solution (5 mg / mL) 藥品. 需要量. MTT (3-(4,5-Dimethyl-2-Thiazolyl)-2,5-Diphenyl Tetrazolium Bromide). 5mg. (USB 19265). 滅菌 1xPBS. 1mL. 將 MTT 粉末溶於滅菌的 1x PBS 中,再以 0.2 μm filter 過濾。分裝於 eppendorf 中,貯存在-20℃. (2) Acidic isopropanol (0.04 N HCl in isopropanol) 藥品. 需要量. Isopropanol 12 N HCl. 1.6mL. 將 1.6 mL 之 12 N HCl 以 Isopropanol 定量至 500 mL,室溫儲存. (3) 滅菌 1x PBS + MTT stock solution (當天配置) 藥品. 需要量. 滅菌 1x PBS. 50 μL / well. - 30 -.
(43) 第三章 材料與方法. MTT stock solution 3、. 5 μL / well. 方法: 在 96 well micro-plate 接種細胞 (1×104 cells / well),待細胞趴附後加入脂肪. 酸標準品或多酚類標準品及其對應的 vehicle control,濃度為 10、50、100、200 μM,培養 24 及 48 小時後進行 MTT 測定。測定當日吸除舊的 medium 後,以 滅菌 1xPBS wash 後吸除,再加入 MTT solution 55μL / well 放入培養箱 37℃反 應 3 小時後,再加入 acidic isopropanol (0.04 N HCl in isopropanol) 100μL / well, 震盪 20 分鐘使染劑溶出後,以 ELISA 測定 540nm 吸光值。 4、. 計算: Cell viability (%) = (test sample / vehicle control) ×100%. 七、 LDH 細胞毒性之測定 1、 原理: Lactate dehydragenase(LDH)是一種穩定存在於各種細胞的酵素,所以若細 胞受到傷害時,細胞膜破裂之後則 LDH 會釋放到 medium 中,測定 medium 中 的 LDH 以表示 cytotxicity 程度。 2、 藥品配製: 使用市售套組 BioVision LDH- Cytotoxicity assay kit Ⅱ Cat.K313-500 3、 方法: 將細胞以 1×104cell/well 種於 96well-microplate24 小時待細胞全部趴附後, 換成各種不同實驗濃度 (10μM、50μM、100μM、200μM) 的各種測試物 100 μL/well 培養 48 小時,溫和 shake 後離心 600xg 10min,將 medium 取到. - 31 -.
(44) 第三章 材料與方法. eppendorf,再從 eppendorf 取出 sample 10μL,並加入 100μL LDH reaction mix 後,室溫反應 30min 測 450nm 吸光。 4、 計算 ( Test sample - Low control). Cytotoxicity(%) =. ×100 ( High control – Low control) Hignt conrol:不以測試物處理之細胞,再以 lysis solution 處理之吸光值 Low control:不以測試物處理之細胞之吸光值. 八、 細胞內三酸甘油酯測定 1、 原理: Triglyceride + H2O Glycerol + ATP. lipase. GK. glycerol + fatty acids. glycerol-3-phosphate + ADP. Glycerol-3- phosphate + O2. GPO. dihydroxyacetone phosphate + H2O2. H2O2 + 4-aminophenazone + 4-chlorophenol. POD. quinoneimine +HCl + 4 H2O. 測定 quinoneimine (紫紅色) 在 500nm 測定吸光值 2、 藥品配製: 採用市售試劑組 (RANDOX, Amtrim, UK),將 Buffer 1 (含 pipes buffer pH7.6,. 4-chlorophenol,. magnesium-ions). 與. Enzyme. regent. 2. ( 含. 4-aminophenazone, ATP, lipase, glycrol-kinase, glycerol-3phosphate oxidase, peroxiase)混合,配成反應試劑備用。 3、 方法: 吸除舊的 medium 後,緩緩加入 1mL 之 1x PBS 潤濕整個培養皿後吸除, 重複二次,再以細胞刮勺將細胞刮下,加入適量 1xPBS 將細胞沖下,細胞液收 集至 1mL eppendorf 中,以 100xg 離心 5 分鐘,小心去除上清液,加入 0.5mL. - 32 -.
(45) 第三章 材料與方法. 二次水打散 pellet 後,以 sonicater 打破細胞,儲存於-20℃。 取 4μL Sample 或 triglycerides standard 於 96 well micro-plate 中,加入 200 μL 上述反應試劑,於室溫反應 10 分鐘,測 500nm 之吸光值。另做 Blank 及 triglycerides standard 作為對照,根據下列公式計算細胞內三酸甘油酯含量。 4、 計算 Triglycerides (mg/mL) = (Asample - Ablank / Astandard - Ablank) × 2 (mg/mL). 九、. 細胞內 G3PDH 活性測定. 1、 原理: 參考 Kozak & Jensen(1974)之方法。細胞質中的 G3PDH 為三酸甘油酯合成之指 標酵素,催化反應如下: DHAP + NADH. G3PDH. glycerol-3-phasphate + NAD+. 2、 藥品配製: (1) Extraction buffer 藥品. 需要量. 最終濃度. Tris-HCl (Merck). 7.88g. 50mM. EDTA (BDH). 0.292g. 1mM. β-mercaptoethanol (Merck). 70μL. 1mM. Triton X-100 (Merck). 5mL. 0.5%. 將 Tris–HCl 和 EDTA 溶於 800 mL 二次水中,再加入 β-mercaptoethanol 和 Triton X-100。調整 pH 值至 7.5,再以二次水定量至 1L,儲存於 4℃. - 33 -.
(46) 第三章 材料與方法. (2) 前 Reaction buffer 藥品. 需要量. 最終濃度. Triethanolamine/HCl (Merck). 18.56g. 100mM. EDTA (BDH). 0.7305g. 2.5mM. β-mercaptoethanol (Merck). 7μL. 0.1mM. 將上述藥品溶於 800 mL 的二次水中,調整 pH 值至 7.5,再以二次水定 量至 1L,儲存於 4℃. (3) 反應受質 NADH stock:1000x stock 藥品. 需要量. 最終濃度. β-nicotinamide adenine dinucleotide reduced (MP Biomedicals). 0.08513g. 120mM. 將 NADH powder 溶於 1 mL 的二次水中(微黃色) ,分裝(100 μL / tube) , 儲存於-20℃. DHAP stock:1000x stock 藥品 dihydroxyacetone phosphate. (Sigma D-7137). 需要量. 最終濃度. 0.017g. 200mM. 將 DHAP powder 溶於 0.5 mL 的二次水中(無色透明),分裝(100 μL / tube),儲存於-20℃. (4) Reaction buffer 藥品. 需要量. - 34 -.
(47) 第三章 材料與方法. 前 Reaction buffer. 1mL. NADH stock. 1μL. DHAP stock. 1μL. 每次依照使用量新鮮配置. 3、 方法: 吸除舊的 medium 後,緩緩加入 1mL 之 1x PBS 潤濕整個培養皿後吸除, 重複二次,再以細胞刮勺將細胞刮下,加入適量 1xPBS 將細胞沖下,細胞液收 集至 1mL eppendorf 中,以 100xg 離心 3 分鐘,小心去除上清液,加入適量冰的 extraction buffer 打散 pellet 後,冰浴 30 分鐘,每隔 5 分鐘 vortex 混勻反應後, 再以 25000xg、4℃離心 1 小時(Hitach CR21, rotor R21 A-39),取出上清液至另 一個 eppendorf,混勻後分裝凍於-80℃。 取適量 sample 加入 1mL Reaction buffer 於 cuvette 中,上下搖晃均勻後放入 分光光度計(U-2000 spectrophotometer, Hitachi),1mL reaction buffer 為空白組, 測 340nm 吸光值,每個 sample 進行 5 分鐘,以下列公式計算下降速率,並以 Lowry’s method 測量蛋白質濃度。 4、 酵素活性計算: NADH : ε340 nm = 6.22 mM-1cm-1 比爾定律 A = εbc (A:吸光;ε:消光係數; b:濾鏡 1cm; c:濃度) c = A / εb G3PDH 酵素活性(nmole / mg prot. min) = (ΔA / min ÷ 6.22 mM-1cm-1) ÷ μg protein × 106 ×(reaction buffer vol. + sample vol.). 十、. Lowry’s method 蛋白質濃度測定. - 35 -.
(48) 第三章 材料與方法. 1、 原理 biuret 法的延伸,當銅離子與胜鏈形成複合物後,可再與 Folin-Ciocalteau 試劑的 phosphomolybdic-phosphotungstate 作用產生藍色物質。 2、 藥品配製 (1) Reagent A (0.5% CuSO4‧5H2O in 1% Na3-citrate‧2H2O) 藥品. 需要量. 最終濃度. CuSO4‧5H2O (聯工). 0.5g. 0.02M. Na3-citrate‧2H2O (USB). 1.14g. 0.04M. 將 0.5 g CuSO4‧5H2O 溶於 80 mL 二次水,再加入 1.14 g 的 Na3-citrate‧2H2O(Na3C6H5O7‧2H2O),最後以二次水定量至 100 mL. (2) Reagent B (2% Na2CO3 in 0.1N NaOH) 藥品. 需要量. 最終濃度. Na2CO3 (聯工). 20g. 0.2M. NaOH (Katayama chemical). 4g. 0.1N. 將 20 g Na2CO3 溶於 800 mL 二次水中,再加入 4 g NaOH,最後以二次水 定量至 1L. (3) Reagent C (Reagent A: Reagent B =1:50) 取 1 mL Reagent A 加 50 mL Reagent B. (4) Reagent D (需避光) (Folin & Ciocalteu’s phenol reagent : 二次水 = 1:1 藥品. 需要量. Folin & Ciocalteu’s phenol reagent. 10mL. - 36 -.
(49) 第三章 材料與方法. 二次水. 10mL. 取 10 mL Folin & Ciocalteu’s phenol reagent 加 10 mL 二次水 Reagent C 和 Reagent D 每次依照使用量當日新鮮配置 (2)Standard 將 1mg BSA(Sigma)溶於 1mL 二次水中製成 1mg / mL stock,實驗時 再利用 stock 做不同濃度之系列稀釋 3、 方法 sample 先以二次水稀釋 50 倍之後,取 30μL sample 或 standard 或二次水加 入 96 well-microplate,加入 150μL reagent C 後於 microplate mixture vortex 上混 勻反應 15 分鐘,再加入 20μL Reagent D 於 microplate mixture vortex 上混勻反 應 30 分鐘。以 ELISA reader (μQuant)測 660nm 吸光值。再以標準曲線公式換 算蛋白質濃度。. 十一、脂解速率測定 1、 原理: Glycerol + ATP. GK. Glycerol-3- phosphate + O2. glycerol-3-phosphate + ADP GPO. dihydroxyacetone phosphate + H2O2. 2H2O2 + 4-aminophenazone + 3,5-dichloro-2-hydroxybenzene sulphonic acid POD. n-(4-antipyryl)-3-chloro-5sulphonate-p-benzoquinoneimine. 以測定甘油的釋出作為指標,根據原理測定 quinoneimine 在 520nm 吸光值 2、 藥品配製: 採用市售試劑組 (RANDOX, Amtrim, UK),將 Buffer 1 (含 pipes buffer pH7.6, 3,5-dichloro-2-hydroxybenzene sulphonic acid, magnesium-ions) 15mL 與 Enzyme regent 2 (含 4-aminophenazone, ATP, glycrol-kinase, glycerol-3-phosphate. - 37 -.
(50) 第三章 材料與方法. oxidase, peroxiase, ascorbic acid oxidase)混合,配成反應試劑備用。 3、 方法: 吸起舊的 medium 收集至 15mL 離心管中,以 1000xg 離心 5 分鐘,吸起上 清液到另一個離心管,混勻後取出 1mL medium 至 eppendorf,再從 eppendorf 取出 100μL medium 至另一個 eppendorf 中,以 70℃加熱 10 分鐘後冰浴,100xg 離心 30 秒,離心後取上清液,儲存於-20℃或進行實驗。 取 40μL sample 或 glycerol standard 於 96 well micro-plate 中,加入 200μL 上述反應試劑,於室溫反應 10 分鐘,測 500nm 之吸光值。另做 blank 及 glycerol standard 作為對照,根據下列公式計算細胞甘油含量。 4、 計算: Glycerol concentration (μmol / L) = (Asample - Ablank / Astandard - Ablank) × 100 (μmol / L). 十二、油紅染色 Oil-Red O stain 測定 1、 原理: 染脂肪細胞內的油滴 2、 藥品配製: 油紅染劑是取 212 mg Oil-Red O 加入 60 mL isopropanol,蓋上蓋子,stir at 4℃ overnight,以 Whatman 3MM paper 過濾。過濾後 Oil-Red O solution : dd water = 3 : 2 混勻後, 放於 4℃ overnight ,最後再以 Whatman 3MM paper 過 濾,貯存於 4℃。 10% formalin/PBS 是取 27.03 mL 37% formaldehyde solution + 72.97 mL PBS 3、 方法:. - 38 -.
(51) 第三章 材料與方法. 吸除 medium,用 PBS(-)5mL 洗細胞兩次後加入 2 mL 10% formalin/PBS(-) 固定細胞 1 hr,再以 5mL dd-water 洗 3 次,加入 1.5 mL 油紅染劑(蓋過細胞 層即可)染 10 min,吸除染劑,以 5mL dd-water 洗數次,室溫倒扣晾乾即可照 相。. 十三、細胞凋亡測定 1、 原理: Formamide 是種溫和的 denaturing reagent,它只會對 apoptotic cells 的 DNA 變性,對 DNA 斷裂的細胞的 necrotic cell 或非 apoptosis cell 都不會作用。所以 本實驗是利用 formamide 將 apoptotic cell 的 DNA 變性之後,再以單株抗體(標 有過氧化酶的二抗)結合上已變性的 DNA(ssDNA)再偵測其吸光。 2、 藥品配製: 使用市售套組 BIOMOL, AK-120;ApoStrandTM ELISA Apoptosis Detection Kit 3、 方法: 由於實驗分成 proliferation、differentiation 及 mature adipocyte 三個階段, 所以將細胞種於 96 well microplate 並依照三階段的流程,以測試物處理後,離 心 200xg 5min 後去除 medium,加入 200μL Fixative 反應 30min 固定細胞,離 心 200xg 5min 去除固定液後於 56℃ oven 20min,再加入 50μL formamide 反應 10min,放入 56℃ oven 30min 後,冷藏 5min,去除 formamide 加入 200μL blocking solution 反應 1 小時,去除 solution 加入抗體反應 30min,以 wash buffer 清洗後加入 peroxidase substate 後反應 60min 測 405nm 吸光。. - 39 -.
(52) 第三章 材料與方法. 十四、統計分析 實驗結果均以平均值 ± 標準差 (Mean ± SD) 表示。數據之統計均須確定 為常態分布,否則轉型為對數值 (Log)。數據以單向變方分析 ( One way ANOVA ) 檢定組間差異之顯著性,再以 Duncan’s multiple range test 及 Dunnet’s test 檢定,統計比較以 p<0.05 為具有顯著之差異。統計分析係以 SAS 軟體(SAS 8.2, Cary, NC, USA)分析。. - 40 -.
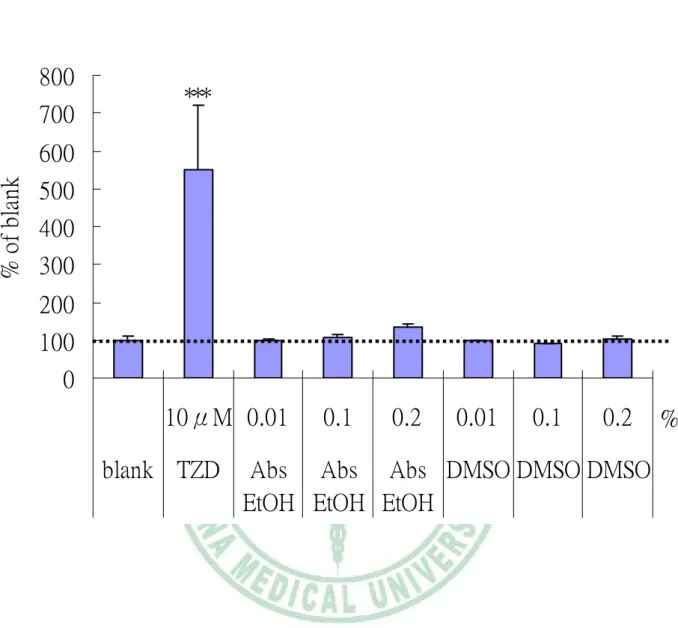
(53) 第四章 結果. 第四章. 結果. 一、 實驗一 脂肪酸對 3T3-L1 脂肪細胞脂質生合成影響. 1、 測試各種溶劑及正對照 TZD 對 3T3-L1 脂肪細胞分化的影響 圖 4-1 各種溶劑及正對照 TZD 對 3T3-L1 前脂肪細胞分化的影響。為確認 3T3-L1 細胞分化流程是否成功,使用已知會促進脂肪細胞分化之 PPARγ促進劑抗 糖尿病藥物 TZD (BRL49653)當成正對照,以細胞內 TG 堆積為指標,發現與未加任 何促進劑的 blank 組相比較,TZD 促分化效果為 blank 組的 5.5 倍;另外又測試二種 溶劑,分別為後續實驗中用來當作脂肪酸及天然抗氧化物之溶劑之 Abs EtOH (absolute ethanol)與 DMSO 對 3T3-L1 分化的影響,同樣以細胞內 TG 堆積作為指標, 發現二種溶劑添加量至 0.2%均與 blank 組沒有差異。 2、 一般脂肪酸對 3T3-L1 脂肪細胞分化的影響 圖 4-2 為以細胞內 TG 堆積為指標測定 common fatty acid 對 3T3-L1 脂肪細胞分 化的影響。使用 common fatty acid 包括有 SFA (saturated fatty acid)- palmitic acid、 stearic acid,MUFA (monounsaturated fatty acid)-oleic acid,PUFA (polyunsaturated fatty acid) - linoleic acid、linolenic acid,以 vehicle control 分化程度作為 100%,發現 5 種 脂肪酸都會促進脂肪細胞分化,並且從 10-200μM 呈現良好 dose-response,而促進 脂肪酸分化程度分別是 SFA 最低其次是 MUFA,促分化程度最好的是 PUFA (p< 0.05)。 3、 特殊脂肪酸對 3T3-L1 脂肪細胞分化的影響 由於前述實驗發現脂肪酸均會促進脂肪細胞分化,因此在測試特殊脂肪酸 c9,t11,t13 conjugated linolenic fatty acid (α-eleostearic acid ; c9,t11,t13-CLN) 和 phytanic acid (PA) 對 3T3-L1 adipogenesis 影響時,決定選用同為 18 碳 3 個雙鍵的一 般脂肪酸 α-linolenic acid 及 16 碳飽和的一般脂肪酸 palmitic acid,分別作為. - 41 -.
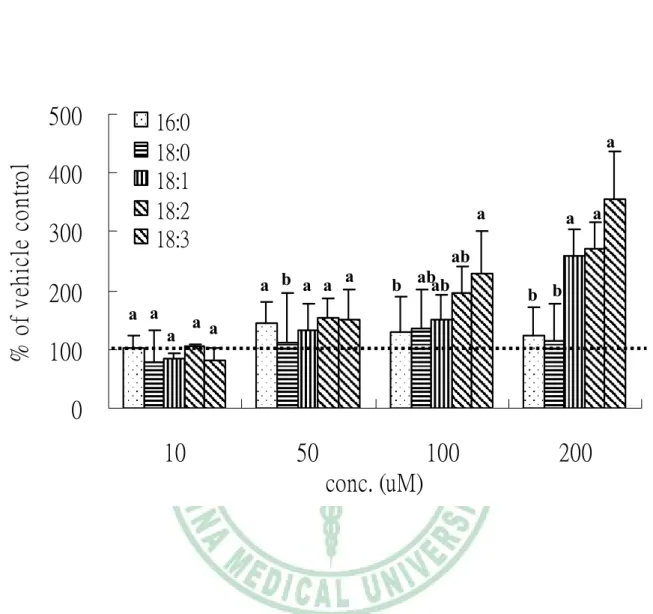
(54) 第四章 結果. c9,t11,t13-CLN 及 PA 之對照,探討二種特殊脂肪酸對 3T3-L1 前脂肪細胞增生、分 化及成熟脂肪細胞脂質代謝之影響。 (1) Uncommon fatty acids 對 3T3-L1 differentiation 之影響 圖 4-3 為以 TG 堆積為指標,測試 uncommon fatty acids 對 3T3-L1 分化 的影響。vehicle control 作為 100%,SFA-C16:0 與 PUFA- C18:3 比較,發現 C16:0 促進分化效果比 C18:3 差 (p<0.05);c9,t11,t13-CLN 及其對照組 C18:3 比較,c9,t11,t13-CLN 顯著抑制細胞內 TG 堆積 (p<0.05);PA 與其對照組 C16:0 比較,PA 亦顯著抑制細胞內 TG 堆積(p<0.05)。 圖 4-4 為以 G3PDH 活性作為指標,測試 uncommon fatty acids 對 3T3-L1 分化的影響。vehicle control 作為 100%與細胞內 TG 堆積結果一致發現 18:3 >16:0,且 c9,t11,t13-CLN 及其對照組 C18:3 比較,c9,t11,t13-CLN 顯著抑制 細胞內 G3PDH 活性 (p<0.05); PA 與其對照組 C16:0 比較,PA 顯著抑制細 胞內 G3PDH 活性 (p<0.05)。 以顯微鏡觀察細胞型態可觀察到, 3T3-L1 細胞進行分化實驗 7 天後細 胞型態明顯改變,轉為圓型內蓄油滴,當 c9,t11,t13-CLN 及其對照組 C18:3 比較,c9,t11,t13-CLN 組油滴數目比較少;PA 與其對照組 C16:0 比較,PA 組 油滴數目比較少,而且又以 C18:3 的油滴數目最多油滴最大,相對的,vehicle 細胞內油滴最少且最小 (圖 4-5)。 油紅染色可染細胞內 TG,更方便觀察細胞內油滴蓄積情形,亦可觀察到 c9,t11,t13-CLN 及其對照組 C18:3 比較,c9,t11,t13-CLN 抑制 TG 累積、油滴 顯著比較少;PA 與其對照組 C16:0 比較,PA 抑制 TG 累積、油滴也是比較 少 (圖 4-6)。 (2) Uncommon fatty acids 對 3T3-L1 前脂肪細胞 proliferation 之影響 圖 4-7 為以 MTT 活細胞存活率試驗測試各種 uncommon fatty acid 對長. - 42 -.
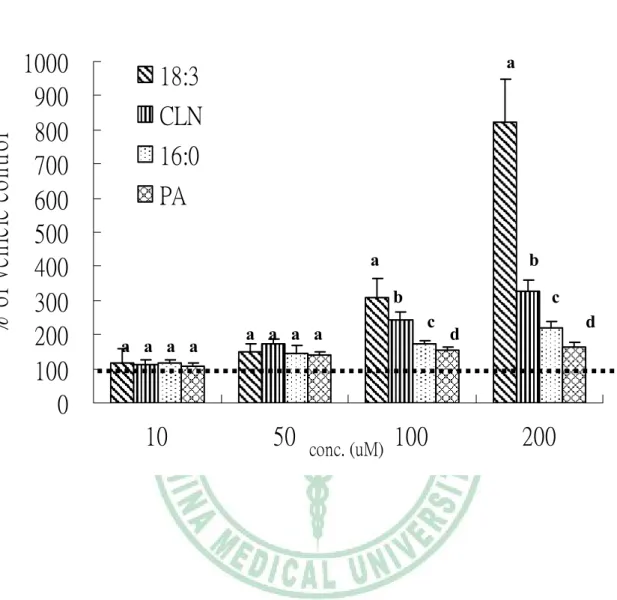
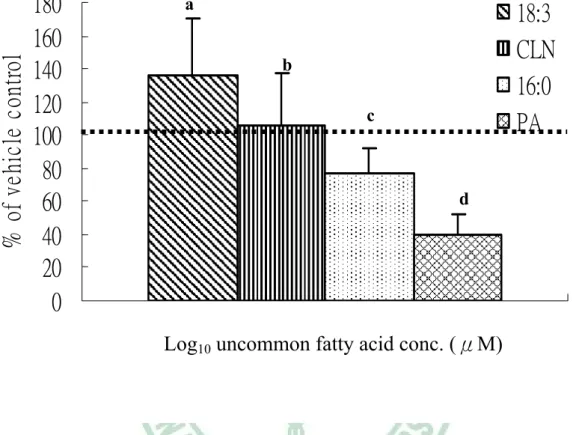
(55) 第四章 結果. 滿前的前脂肪細胞階段 3T3-L1 細胞存活的影響。vehicle control 作為 100%, 圖 4-7(A)可以得知在 24 小時給予測試物處理後,發現 c9,t11,t13-CLN 和 C16:0 於 200μM 時,才會明顯造成細胞數減少 (p<0.05);而在處理 48 小時後 (圖 4-7(B) ) 抑制情形更明顯,c9,t11,t13-CLN 於 10μM 就開始抑制細胞增生 (p <0.05),而且有 dose-response;而 C16:0 於 200μM 才會顯著造成細胞數目 減少 (p<0.05),相對的 C18:3 及 PA 到 200μM 均不會造成細胞數目減少。 由於觀察到 c9,t11,t13-CLN 及 C16:0 會造成細胞數減少,因此利用 LDH release assay 測試 uncommon fatty acid 處理 48 小時,對 3T3-L1 細胞毒性的影 響 ( 圖 4-8 )。發現 c9,t11,t13-CLN 於 10μM 就開始對細胞產生毒性 (p< 0.05),C16:0 則是要在 200μM 時才會顯著造成細胞毒性 (p<0.05),而 C18:3 及 PA 即使在 200μM 也不會對細胞造成毒性。 (3) Uncommon fatty acids 對 3T3-L1 已分化成熟脂肪細胞脂質代謝之影響 圖 4-9 為以 TG 堆積為指標,測試 uncommon fatty acids 對已分化 3T3-L1 成熟脂肪細胞 TG 蓄積的影響。先以 C18:2 及促分化劑誘導脂肪細胞完全分 化,在顯微鏡下可見>95%分化率,此時移去 C18:2 改以不同測試物處理 48 小時。以 vehicle control 作為 100%,可發現 4 種脂肪酸均促進 TG 蓄積, c9,t11,t13-CLN 及其對照組 C18:3 比較,c9,t11,t13-CLN 抑制 TG 累積(p< 0.05);而 PA 與其對照組 C16:0 比較,PA 抑制 TG 累積(p<0.05);另外又以 細胞內 G3PDH 活性為指標,同樣發現 c9,t11,t13-CLN 抑制成熟脂肪細胞內 G3PDH 活性(p<0.05),PA 則有抑制細胞內 G3PDH 活性的趨勢,但無統計上 之差異 (圖 4-10)。在成熟脂肪細胞脂解方面和 C18:3 比較,c9,t11,t13-CLN 有抑制脂解作用 (圖 4-11)。 (4) c9,t11,t13-CLN 對 3T3-L1 三 階 段. ( preadipocyte proliferation 、. differentiation、terminal differentiation ) 細胞凋亡的影響. - 43 -.
(56) 第四章 結果. 由於前述實驗發現 c9,t11,t13-CLN 抑制 3T3-L1 增生、分化效果最明顯, 因此針對 CLN 探討是否造成細胞凋亡,此外 c9,t11,t13-CLN 在前脂肪細胞階 段於 10μM 即造成細胞數減少,在分化階段於 100μM 才會造成細胞數減少 ( 細胞內蛋白質定量結果;結果未示 ) ,而在已分化成熟脂肪細胞處理 100 μM 並不影響細胞數,因此在前脂肪細胞階段 c9,t11,t13-CLN 以 10μM、50 μM、100μM 的劑量,分化階段和成熟脂肪細胞以 100μM 的劑量,分別測 試 c9,t11,t13-CLN 對 3T3-L1 脂肪細胞細胞凋亡的影響,結果發現 100μM c9,t11,t13-CLN 於 preadipocyte 階段及分化階段,都會造成 3T3-L1 細胞凋亡 (p<0.05 ),但 c9,t11,t13-CLN 不會造成成熟脂肪細胞細胞凋亡(圖 4-12)。. 二、 實驗二 天然抗氧化物對 3T3-L1 脂肪細胞脂質生合成影響. 1、 Natural antioxidants 對 3T3-L1 脂肪細胞分化之影響 圖 4-13 以細胞內 TG 堆積為指標,測試 9 種天然抗氧化物對 3T3-L1 脂肪細胞 分化的影響。9 種天然抗氧化包括有 EGCG、curcumin、catechin、naringenin、ascorbic acid、resveratrol、apigenin、quercetin、genistein,vehicle control 為 DMSO, 結果 發現以 4 種天然抗氧化物 (curcumin、resveratrol、apigenin、genistein ) 明顯抑制分 化,因此之後選擇此 4 種加上文獻常被報導可抑制 3T3-L1 分化之 EGCG 做為後續 實驗。 另外以細胞內 G3PDH 活性作為分化指標,測試 4 種多酚類 (curcumin、 resveratrol、apigenin、genistein ) 及 EGCG 在 100μM實驗劑量下比較,vehicle control 分化程度當成 100%,結果發現除了 EGCG 之外其餘 4 種多酚類都會顯著抑制細胞 內 G3PDH 活性,而且以 curcumin 最強 (圖 4-14)。 同樣地以顯微鏡觀察 4 種多酚類及 EGCG 進行 7 天分化實驗之細胞型態,與. - 44 -.
(57) 第四章 結果. vehicle control 比較,發現除了 EGCG 之外,其餘 4 組多酚類幾乎看不到細胞有油滴 產生,curcumin 組細胞型態明顯的改變,與正常的 3T3-L1 型態不同. (圖 4-15)。. 再將各組細胞進行油紅染色,可觀察到 EGCG 油滴的數量和 vehicle 差不多,而其 餘 4 組多酚類油滴數很少,且以 curcumin 為最少 (圖 4-16)。 2、 多酚類對 3T3-L1 前脂肪細胞 proliferation 之影響 之後特別對分化實驗篩選出的 4 種多酚類及 EGCG 進行分析,測試對長滿前 preadipocyte 增生影響。以 5 種多酚類處理 24 小時後,可觀察到 5 種多酚類均會抑 制細胞存活率且有 dose-response,其中以 curcumin 最為明顯,在 50μM 顯著低於 它組 ( p<0.05);而在以 48 小時處理後,抑制效果更為顯著,同樣又以 curcumin 為最強,在 50 μM 處理下顯著低於它組( p<0.05) (圖 4-17)。 另外以 LDH release assay 測試 5 種多酚類及 vehicle 對 3T3-L1 前脂肪細胞細胞 毒性的影響,其中只有 curcumin 對 3T3-L1 前脂肪細胞產生細胞毒性,且於 50μM 即會產生,並且呈現 dose-response(圖 4-18)。 3、 多酚類對 3T3-L1 已分化成熟脂肪細胞脂質堆積之影響 使用 linoleic acid 促分化劑誘導 3T3-L1 形成成熟脂肪細胞後,再給予 100μM 5 種多酚類及 vehicle 處理 48 小時,vehicle control 處理之 TG 程度當成 100%,發現 抑制細胞內 TG 堆積以 EGCG、resveratrol、genistein 較明顯 ( p<0.05)(圖 4-19)。 4、 curcumin 對 3T3-L1 三階段 ( preadipocyte proliferation、differentiation、terminal differentiation ) 細胞凋亡的影響 由於前述實發現 curcumin 抑制 3T3-L1 增生、分化效果最明顯,因此針對 curcumin 探討是否造成細胞凋亡,此外 curcumin 在前脂肪細胞階段於 50μM 即造 成細胞數減少且具有細胞毒性,在分化階段於 100μM 會造成細胞數目減少 ( 細胞 內蛋白質定量結果;結果未示),而在已分化成熟脂肪細胞處理 100μM 並不影響細 胞數,因此在前脂肪細胞階段 curcumin 以 10μM、50μM、100μM 的劑量,分化. - 45 -.
(58) 第四章 結果. 階段和成熟脂肪細胞以 100μM 的劑量,分別測試 curcumin 對 3T3-L1 細胞凋亡的 影響。結果發現 100μM curcumin 會於分化及成熟脂肪細胞階段對 3T3-L1 脂肪細胞 造成細胞凋亡 ( p<0.05 ) (圖 4-20)。. - 46 -.
(59) 第四章 結果---實驗一. 800. ***. 700 % of blank. 600 500 400 300 200 100 0 10μM 0.01 blank TZD. 圖 4-1. 0.1. 0.2. 0.01. 0.1. 0.2. %. Abs Abs Abs DMSO DMSO DMSO EtOH EtOH EtOH. 各種溶劑及正對照TZD對3T3-L1前脂肪細胞分化 ( 以細胞內TG堆積為 指標 ) 的影響. Figure 4-1 Effect of solvent and positive control TZD (BRL49653) on differentiation of 3T3-L1. Cellular triglyceride (TG) was measured after the initiation of differentiation for 7d. Values are means±S.D. (n=3). The significance of differences between the treatment (TZD, Abs EtOH, DMSO) and blank group was analyzed statistically by Dunnet’s test. *p<0.05, **p<0.005, **p<0.0005.. - 47 -.
數據



+7
Outline
相關文件
SPO has high fatty acids, therefor it has to carry out the acid catalyzed esterification to convert of the fatty acids in SPO to biodiesel first, and then used the
L1:add eax,C_minutesInDay ; totalMinutes+=minutesInDay call WriteString ; display str1 (offset in EDX) call WriteInt ; display totalMinutes (EAX) call Crlf. inc days
The predicted expression profiles of 24 chemokines and immunosuppressive biomarkers for SCC4, SCC15, and SCC25 were used in a decision tree format to sort cell lines into those
而其他性質又不假設?假設的依據為何?然而這些假設內
使用限制 海水、硬水 皂化反應.
[r]
Parry,“ Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures, ” Aquaculture, Vol.211, No.1-4,
add ax,[edi] ; add an integer add edi,TYPE intarray ; point to next loop L1 ; repeat until ECX = 0 The following code calculates the sum of an array of 16-bit integers... Copying
