碳源與生長速率對大腸桿菌DNA 超螺旋結構、ATP生合成基因與細胞分裂基因調控之研究
95
0
0
全文
(2) 碳源與生長速率對大腸桿菌 DNA 超螺旋結構、ATP 生合成基因與細胞分 裂基因調控之研究 學生:鄭惠文. 指導教授:曾慶平 博士 國立交通大學. 生物科技研究所 碩士班 摘. 要. 微生物可利用不同受質獲取能量,大腸桿菌代謝碳源產生能量的途徑與生長 速率亦隨受質不同而改變。本論文以醋酸、葡萄糖、甘油與琥珀酸四種不同碳源 作為大腸桿菌之生長基質,結果發現不同碳源會造成細胞生長速率改變,相對地 代謝途徑上與 ATP 生合成相關基因的表現量也發生變化(pgk、 pyk、ackA、 sucCD、atpI)。由於 DNA 超螺旋結構受 ATP/ADP 比例所調控,研究發現細 胞內 ATP 與 ADP 含量以及 ATP/ADP 比例會隨碳源不同而上升或下降,當生 長速率加快時,因 ATP/ADP 比例與 gyrase 表現量受到調控而上升,因此促使 DNA 超螺旋結構較生長速率緩慢時來得緊密。 此外 ppGpp 為大腸桿菌在貧瘠環境下的重要調控因子,實驗結果顯示 ppGpp 並不影響大腸桿菌體內 ATP/ADP 比例與 DNA 超螺旋結構的改變,但 卻觀察到 ppGpp 會使細胞分裂基因 ftsZ 表現上升,且對 minC 與 minD 基因 進行負向調控。本研究也發現細胞大小會隨生長速率加快而變大,以不同碳源為 生長基質時,造成細胞分裂基因(ftsZ, minC, minD)的表現有明顯差異。當大 腸桿菌生長在碳源貧乏的環境時,菌體型態會趨向長絲狀並呈現鍊狀結構,因此 推測 ppGpp 有可能透過影響細胞分裂基因的表現而改變細胞大小以及菌體型 態。本實驗藉由研究碳源如何改變大腸桿菌生長速率, ATP 與 ADP 含量及細 胞內 ATP/ADP 比例、 DNA 超螺旋結構、細胞分裂等結果,可作為基礎分子生 物學研究提供一個重要的模式發現。. i.
(3) ABSTRACT. To understand the expression of ATP generation and cell division genes in different carbon sources, we individually used acetate, glucose, glycerol or succinate as a sole carbon for energy source. The results of this study showed that the expression of ATP generating genes in metabolic pathway varied with carbon sources and ATP concentration increased with cell growth rate. Comparision with the wild-type strain and relA spoT double mutant, the growth rate and ATP yields were changed, but ATP/ADP ratio remained at the same level. DNA supercoiling was dependent on ATP/ADP ratio. Whereas ppGpp did not change DNA supercoiling. Under various growth conditions, fast-growing E. coli cells were larger than slowly growing ones. Starvation of the cells resulted in filamentous morphology. In addition, the results clearly showed that relA spoT double mutant had more filamentous than wild-type cells regardless acetate or glucose as carbon substrates. It was also notable that the filamentous features provided the phenotypic clues for ppGpp function. However, the morphology raised the possibility of indirect, rather than direct, effects of ppGpp. It indicated that a link between the levels of ppGpp and cell division, which ppGpp could act as a positive regulator of the expression of ftsZ gene. Deficiency of ppGpp (relA spoT double mutant) drastically reduced the expression of minC and minD. These results also suggested that ppGpp was important factor involved in the regulation of cell cycle of E. coli under starvation condition.. ii.
(4) 誌謝. 在此衷心感謝指導教授曾慶平老師在實驗與生活上悉心指導與啟發導,讓我 受益良多,並感謝彭慧玲老師與楊昀良老師的細心指導與寶貴意見,使我得到更 多科學上的觀點與想法,本論文獻上最誠摯的謝意。 實驗室裡每一位深具個人特色的伙伴,感謝你們陪我走過漫漫的研究歲月, 使我感受到研究生活中動人又有趣的另一面,不管實驗或生活上的點點滴滴都深 深烙印在心裡,還記得說話與實驗都超快的鐘達、深具智慧的育甄,慈祥又美麗 的傅老師、認真又滿懷搞笑細胞的校賢、寬容又博學多聞的煜沛、堆肥王國裡的 羽球王子--國領、熱心負責的宗益、聰明又具耐心的慧貞、鬼靈精怪的鈞毅、能 言善道的承宏、活潑善良的慧珊、高人一等的上進青年--岳賢,以及可愛又美麗 的同學們--靜雪、慧君、郁婷、巧璇,還有一群嗷嗷待哺又充滿活力的學弟妹們 --繼元、愛雁、翊維、冀冬和智彬,另外,要感謝大學的學長姐及同窗好友們, 因為你們的存在,我才能順利完成論文,你們的幫助與鼓勵真是令我感動不已。 最後要感謝父母與家人精神上的支持,默默陪我走過最低潮的時候,過去的 衝突與挫折,因為你們的包容,讓我得以在人生的路途上不斷前進,以本論獻上 最深的愛意與敬意,滿心感激每一位伴我成長的伙伴們!. iii.
(5) 目錄 頁次 中文摘要…………………………………………………………………………………. i. 英文摘要…………………………………………………………………………………. ii. 致謝…………………………………………………………………………………........ iii. 目錄…………………………………………………………………………………........ iv. 圖表目錄……………………………………………………………………………….... vii. 壹 序言………………………………………………………………………………….. 1. 1.1 研究緣起…………………………………………………………………………. 1. 1.2 研究目的…………………………………………………………………………. 2. 貳、文獻回顧……………………………………………………………………………. 4 2.1 碳源與生長速率對大腸桿菌能量狀態與 DNA 超螺旋結構之影響………….. 4. 2.1.1 碳源對大腸桿菌細胞生長速率之影響……………………………………... 4. 2.1.2 生長速率對大腸桿菌細胞組成及其生理狀況的影響……………………... 4. 2.1.3 細胞生長速率對基因表現的影響…………………………………………... 5. 2.1.4 生長速率調控機制的探討…………………………………………………... 6. 2.1.5 細胞內主要來源 ATP……………………………………………………….. 7. 2.1.6 碳源對代謝產生能量途徑之影響…………………………………………... 8. 2.1.7 ATP/ADP 比例………………………………………………………………. 9. 2.1.8 DNA 超螺旋結構(DNA supercoiling)與 topoisomerase ………………. 10 2.1.9 細胞內能量因子調控 DNA 超螺旋結構…………………………………… 12 2.2 貧乏培養(starvation)與 ppGpp 對大腸桿菌生理之影響………………… 13 2.2.1 迫切反應(stringent response)與 ppGpp 對大腸桿菌之生理調控…….. 13 2.2.2 ppGpp 作用機制與對細菌生理影響………………………………………. 14. iv.
(6) 2.3 大腸桿菌之細胞分裂基因……………………………………………………….. 15. 2.3.1 細胞分裂 ( cell division )………………………………………………….... 15. 2.3.2 細胞隔板的形成(septum formation)…………………………………… 16 2.3.3 細胞隔板形成之抑制者……………………………………………………... 17. 參 基本理論……………………………………………………….. …………………... 18. 3.1 批次培養(batch culture)之細胞生長模式. …………………………………. 18. 3.1.1 批次培養下細胞之生長曲線………………………………………………... 18. 3.1.2 菌體倍增時間………………………………………………………………... 18. 3.2 連續式培養之動力學模式………………………………………………………. 20. 3.3 聚合酶連鎖反應(Polymerase chain reaction,PCR)之原理……………….. 22 肆 材料、設備與方法……………………………………………………….. ………… 24 4.1 實驗材料…………………………………………………………………………. 24. 4.1.1 菌株質體與噬菌體…………………………………………………………... 24. 4.1.2 藥品與試劑…………………………………………………………………... 24. 4.1.3 緩衝液………………………………………………………………………... 26. 4.1.4 培養基與培養液……………………………………………………………... 27. 4.1.5 電泳膠體……………………………………………………………………... 28. 4.2 實驗方法………………………………………………………………………….. 28. 4.2.1 大腸桿菌生長條件與培養方法……………………………………………... 28. 4.2.2 細胞倍增時間的測定………………………………………………………... 31. 4.2.3 細胞轉型(transformation)…………………………………………………. 32 4.2.4 大量抽取細菌質體…………………………………………………………... 33. 4.2.5 DNA 超螺旋結構電泳分析………………………………………………….. 34. 4.3.6 細胞內能量狀態測定……………………………………………………….. 35. 4.3.7 突變株的構築與篩選……………………………………………………….. 36. v.
(7) 4.3.8 大腸桿菌總 RNA 製備方法……………………………………………….. 38. 4.3.9 北方墨點轉漬分析法(Northern blotting analysis)………………………. 39 4.3.10 革蘭氏染色法……………………………………………………………….. 44. 伍 結果……………………………………………………….. ………………………... 45. 5.1 大腸桿菌生長速率與能量變化之關係…………………………………………. 45. 5.2 貧乏培養(starvation)與 ppGpp 對大腸桿菌能量變化之影響…………… 45 5.3 DNA 超螺旋結構之電泳分析………………………………………………….. 46. 5.3.1 不同碳源對大腸桿菌 DNA 超螺旋結構之影響…………………………… 47 5.3.2 貧乏培養(starvation)與 ppGpp 對大腸桿菌 DNA 超螺旋結構之影響… 48 5.4 生長速率與碳源對不同 topoisomerase 基因表現之影響…………………... 49. 5.5 不同碳源對 ATP 合成酵素基因表現之影響…………………………………… 49 5.6 碳源與 ppGpp 對細胞分裂基因之影響………………………………………. 50. 5.7 碳源與 ppGpp 對大腸桿菌菌體型態之影響…………………………………. 51. 陸. 討論……………………………………………………….. ………………………. 53. 6.1 碳源與生長速率對大腸桿菌細胞內能量之關係………………………………. 53. 6.1.2 不同碳源與生長速率對大腸桿菌細胞內能量狀態的影響……………….. 53 6.1.3 貧乏培養(starvation)與 ppGpp 對大腸桿菌細胞內能量狀態之影響….. 55 6.1.4 碳源對大腸桿菌 ATP 產生相關酵素基因之影響………………………... 56. 6.2 碳源與生長速率對大腸桿菌 DNA 超螺旋結構之關係……………………….. 59. 6.2.1. 碳源、生長速率與 ATP/ADP 比例對 DNA 超螺旋結構之影響………… 59. 6.2.2. 碳源與生長速率對於 topoisomerases 基因表現之影響………………. 6.3 生長速率與大腸桿菌細胞分裂之關係………………………………………… 6.3.1 碳源對大腸桿菌細胞分裂基因 ( ftsZ, minC, minD ) 的影響……………. 60 61 61. 6.3.2 ppGpp 對大腸桿菌細胞分裂基因(ftsZ, minC, minD)與菌體型態之影響 62 柒 參考文獻……………………………………………………….. …………………... vi. 79.
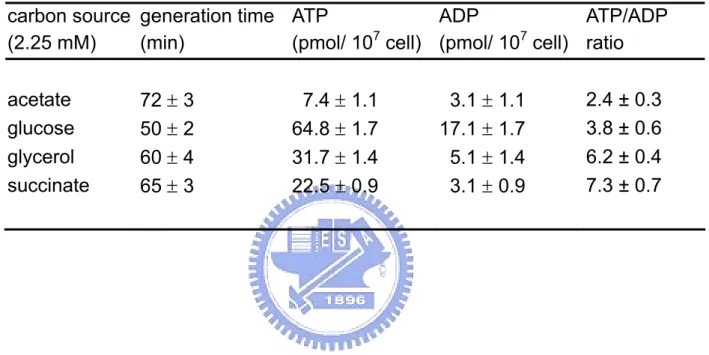
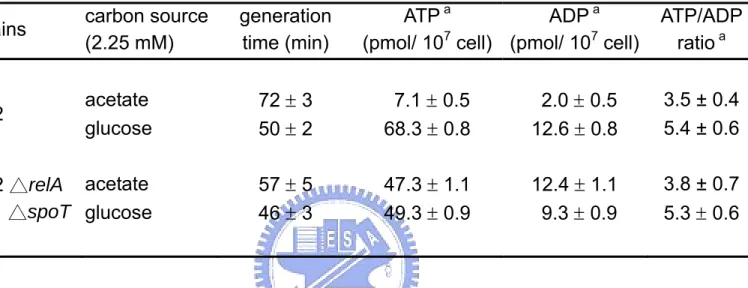
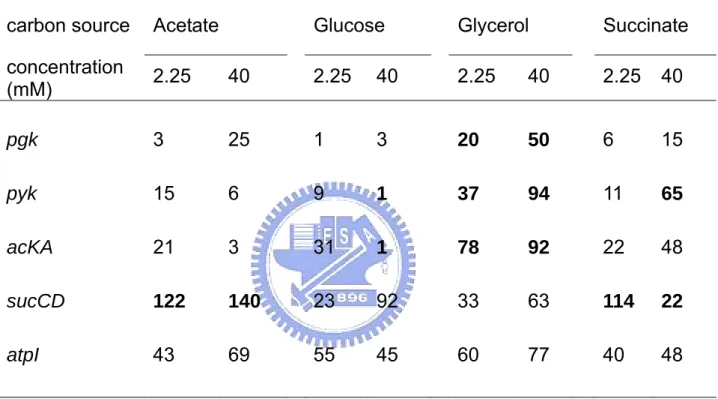
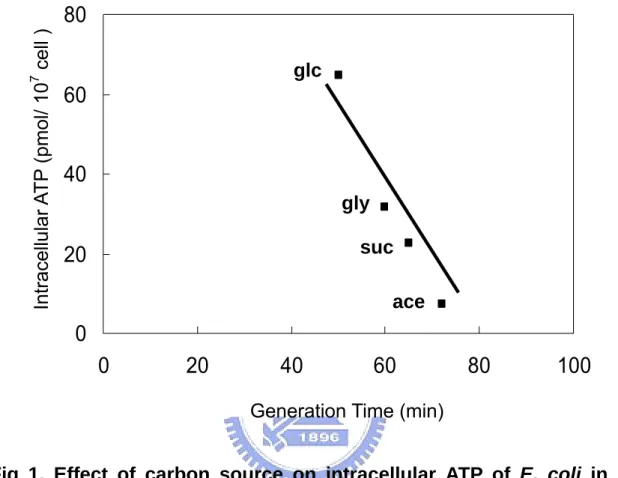
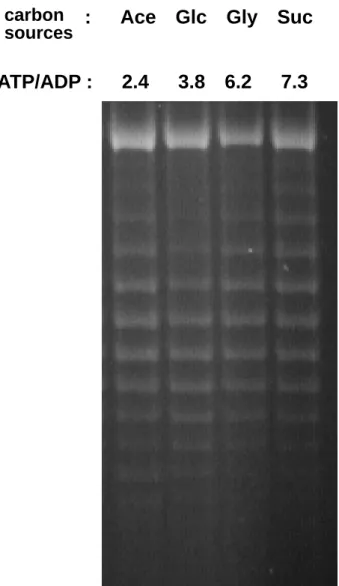
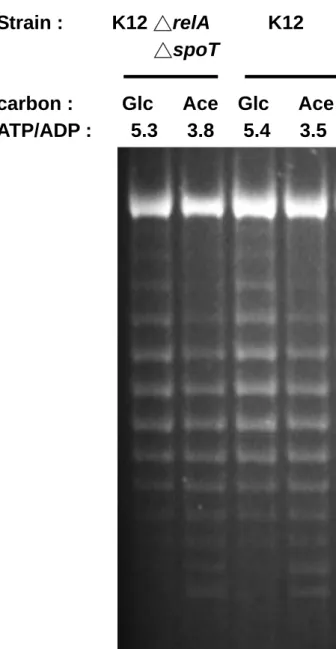
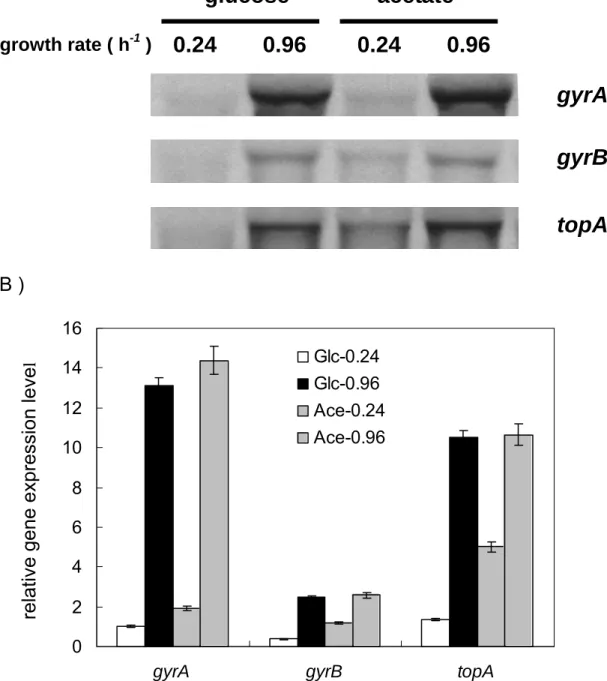
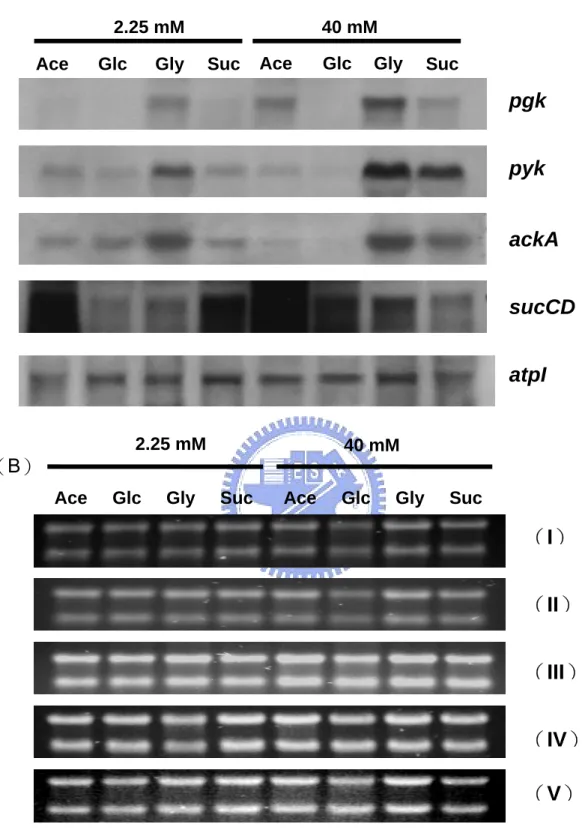
(8) 圖表目錄 Table 1. Effect of carbon source on energy state of E. coli in batch culture.. 65. Table 2. Effect of starvation and ppGpp on energy state of E. coli in batch culture.. 66. Table 3. The changes of ATP generating gene expression in various carbon medium.. 67. Fig 1. Effect of carbon source on intracellular ATP of E. coli in batch culture.. 68. Fig 2. Effect of carbon source on DNA superocoiling of E. coli in batch culture.. 69. Fig 3. Effect of starvation and ppGpp on DNA superocoiling at different carbon sources in E. coli in batch culture.. 70. Fig 4.Effect of carbon source and growth rate on the expressions of topoisomerase genes ( gyrA, gyrB, topA ) in E. coli.. 71. Fig 5. Effect of carbon source on the expressions of ATP generating genes ( pgk, pyk, ackA, sucCD, atpI ) in E. coli.. 72. Fig 6. The quantitative northern blot analysis of pgk, pyk, ackA, sucCD and atpI. 73 Fig 7. The metabolic pathways of glycolysis, tricarboxylic acid cycle and electron transfer chain. Fig 8. Effect of carbon source on the expressions of cell division genes(ftsZ, minC, minD) of E. coli.. 74. Fig 9. Effect of starvation and ppGpp on the expressions of cell division genes (ftsZ, minC, minD)at different carbon sources in E. coli.. 76. Fig 10. Morphology of wild-type K12 and ppGpp mutant under different carbon sources in minimal medium.. 77. Fig 11. Morphology of wild-type K12 and relA spoT mutant in carbon-starved minimal medium.. 78. vii. 75.
(9) 壹、序言. 1.1 研究緣起. 細菌生長過程從生長、分裂以至死亡各階段中,每個細胞明顯地受到環境中 物理以及化學因子所影響(Rothfield, 1996),我們可清楚地觀察到當細胞生長在 養分充足或適宜的環境時,細胞生長與分裂較為快速,反之則變得較緩慢,而在 生長快速與緩慢細胞之間有明顯不同的生理變化。已知環境中的養分、溫度、含 氧量、酸鹼度、滲透壓等因子改變均會造成細胞生長及生理變化(Bloch et al., 1980; Epstein and Schultz, 1965; Herendeen et al.; 1979, Felle et al., 1980) ,而且細胞 生長速率改變時會影響細胞內巨分子(如 DNA 、RNA)的組成(Rothfield, 1996) 。 目前已證實許多基因會受生長速率影響(growth rate-dependent regulation) ,如 生 長 速 率 加 快 時 , glucose-6-phosphate dehydrogenase 、 enolase 、 DNA methyltransferase 等基因的表現量會隨之上升(Rasmussen et al., 1995; Rowley et al., 1991 ), 而. fumarase reductase 與. TCA cycle 中 succinate. dehydrogenase、malate dehadrogenase 及 fumarate dehydrogenase 等基因的 表現卻隨著生長速率加快而下降。 此外細胞生長與分裂需要能量供應,而在不同環境下細胞內的能量狀態亦有 所不同。目前多數研究以 ATP/ADP 比例作為能量對細胞生理狀況影響的指標 (Neidhardt et al., 1990) ,細胞體外實驗顯示經純化後的 DNA gyrase 活性受到 ATP/ADP 比例影響,細胞內 ATP/ADP 比例增加時,質體 DNA 超螺旋結構亦 隨之旋緊(Workum et al., 1996)。細胞內 ATP 和 ADP 含量會有一定比例關 係,細胞接受外界環境的刺激,例如改變溫度,滲透壓及含氧量等因子均會使得 ATP/ADP 比例發生變化,並且同時觀察到 DNA 超螺旋結構相對應改變(Hsieh et al., 1991; Camacho-Carranza et al., 1995),此訊息透露 ATP/ADP 比例對. 1.
(10) DNA 超螺旋結構的調控扮演重要角色。由於 DNA 超螺旋結構與 ppGpp 均為 細菌體內重要的調控因子,目前已知在大腸桿菌細胞內至少有 88 種以上蛋白質 的表現受 DNA 超螺旋結構影響(Notley-McRobb et al., 1997) ,且細胞內 DNA 超螺旋結構也影響了 DNA 複製、轉錄等機制,此外大腸桿菌體內 ppGpp 在營 養素缺乏( starvation )時對菌體生理之調控已被廣為研究( Gallant, 1979; Hernandez and Bremer, 1990)。因此本論文經由探討不同碳源對細菌生理所造 成之變化,藉此了解是否不同碳源可經由改變細胞內能量狀態、代謝途徑與生合 成反應,以及 DNA 超螺旋結構或是 ppGpp 層次變化,造成細胞分裂基因表現 與菌體型態改變,進而影響了細胞生長速率。. 1.2 研究目的. 已知生長速率可調控細菌某些基因的表現,但是基因表現受細胞生長速率影 響的機制尚未明確,由於 DNA 超螺旋結構改變與 ppGpp 濃度改變等已證實會 影響許多基因表現,因此藉由連續式培養操作技術與分子生物學結合,來研究大 腸桿菌不同碳源及生長速率對能量狀態、 DNA 超螺旋結構、細胞分裂與 ppGpp 的影響。大腸桿菌目前為研究基因調控最有效的系統之一,由其所建立的模式可 作為研究其它生物細胞相關機制之參考。藉由探討不同碳源對大腸桿菌生長速率 與基因調控之間的關係,可為基礎分子生物學發現整體相互調控作用,並對微生 物遺傳與生理之間的關係有更深一層的瞭解。. 綜合上述,我們以下列關係圖顯示此論文目的與研究方向之關聯。. 2.
(11) 研究目的 :不同碳源與生長速率對細胞內 ATP/ADP 比例及 DNA 超螺旋結構的影響,並偵測代謝途徑上 ATP 生合成基因之 表現,以及對細胞分裂基因與菌體型態調控之關係。. 研究方向. 1.不同碳源與starvation 時 ppGpp 對 菌 體 ATP/ADP 比 例 與 DNA 超螺旋結構改變。 2.不同碳源與生長速率 下 偵 測 topoisomerase ( gyrA、gyrB、topA) 基因表現。. 不同碳源下偵測代謝 途徑ATP生合成基因 (pgk、pyk、ackA、 sucCD、atp)表現變 化。. 3. 不 同 碳 源 與 starvatio時 ppGpp 對細胞分裂基因 ( ftsZ 、 minC 、 minD)表現與菌體 型態變化。.
(12) 貳、文獻回顧. 2.1. 碳源與生長速率對大腸桿菌能量狀態與 DNA 超螺旋結構之影響. 2.1.1 碳源對大腸桿菌細胞生長速率之影響. 大腸桿菌可利用不同基質(substrate)來生長,不同的營養成份如碳源及氮 源經微生物攝取後可經由細胞內不同的代謝途徑加以利用,進而產生不同細胞生 長速率。例如大腸桿菌 K12 菌株培養在以葡萄糖(glucose)為碳源的培養基時, 其細胞細胞倍增所需的時間(doubling time)約為 0.5 小時。若培養在僅以醋酸 (acetate)為碳源之培養基時,其細胞倍增時間則增長為 2 小時。 Wolf 等人曾 利 用 不 同 養 分 培 養 液 證 明 大 腸 桿 菌 體 內 6- 磷 酸 葡 萄 酸 鹽 去 氫 酶 ( 6-phosphogluconate. dehydrogenase ) 及 葡 萄 糖 -6- 磷 酸 去 氫 酶. (glucose-6-phosphate dehydrogenase)等酵素活性均受到細胞生長速率所調 控,並證實此二酵素活性的改變是直接由生長速率改變所造成,而不是由於碳源 的專一性所影響造成(Rowley et al., 1991)。同樣利用不同養分培養基組成也證 明了 ompA 基因、elongation factor Tu 及 gua 操縱子的表現會受到生長速率的 調控(Furano, 1975; Cozzarelli et al., 1990; Georgellise et al., 1992) ,本實驗即 利用不同碳源進行批次培養以研究生長速率的變化。. 2.1.2 生長速率對大腸桿菌細胞組成及其生理狀況的影響. 細胞生長速率會影響細胞內的生理狀況,特別是細胞內巨分子組成,包括 DNA 、RNA 及蛋白質等均會隨著細胞生長速率不同而改變。 Dorman 等人在研. 4.
(13) 究傷寒沙門氏桿菌(Salmonella typhimurium)中首先證明細胞內巨分子組成隨生 長速率上升而增加(Dorman et al., 1988) 。陸續研究亦發現在大腸桿菌 B/r 菌株 中,當生長速率由 0.6 doubling/hr 增快至 2.5 doubling/hr 時,RNA 含量及核糖 體(ribosome)增加近 10 倍,而 DNA 含量增加近 2 倍(Ingraham, 1987)。 由於生長速率改變對於細胞的巨分子組成造成了重大改變,意謂著生長速率對於 細胞生理而言是複雜且重要的調控因子。 除了巨分子外,許多細胞內生理因子亦被生長速率所調控。如 RNA 聚合酶 活性也隨著生長速率增快而增加。生長速率加快同時也造成了細胞轉譯頻率 ( translational frequency ) 增 加 , 複 製 時 間 與 複 製 起 始 重 量 下 降 等 的 情 形 (Ingraham, 1987; Wold et al., 1994),由於上述各種因子均受生長速率所調控, 因此細胞生長速率與基因表現之關係一直受到重視。. 2.1.3 細胞生長速率對基因表現的影響. 在菌體中有許多基因會受生長速率所影響,這些基因的表現量與酵素活性高 低一般可分為兩種情形: 1. 隨著生長速率加快而上升。 2. 隨著生長速率下降而增加。 例如 zwf (glucose-6-phosphate dehydrogenase) 、ompA (out membrane protein II)等基因屬於前者(Rowley et al., 1991; Georgellis et al., 1992) ,而 rmf (ribosome modulation factor)、frdABCD (fumarate reductase)以及 lacZ 基 因則屬於後者(Yamagish et al., 1993; Tseng et al., 1994a; 2001b)。除此之外, 以下再舉出幾個受生長速率調控的基因為範例加以介紹: 在 TCA cycle 中,將琥珀酸轉換成延胡索酸(fumarate)的琥珀酸去氫酶. 5.
(14) (succinate dehydrogenase, sdhCDAB)與催化蘋果酸(malate)和草醋酸 (oxaloacetate)之間轉換的蘋果酸去氫酶(malate dehydrogenase, mdh)此二 酵素,無論在有氧或無氧環境下基因表現量均受生長速率調控(Park et al., 1995a; 1995b)。本研究室亦發現於有氧環境下催化延胡索酸與蘋果酸轉換的延胡索酸酶 A(fumA)及延胡索酸酶 C(fumC)之表現量也均會隨著生長速率加快而降低。 DNA methyl-transferase(dam)是基因複製時負責甲基化 GATC 序列且為 錯誤配對(mismatch)修復時的必需酵素。目前已證實 dam 基因中的主要啟動 子 P2 在 高 生 長 速 率 下 有 較 高 的 轉 錄 頻 率 ( transcriptional frequency ) (Rasmussen et al., 1995)。而在磷酸五碳糖反應途徑(pentose phosphate pathway)中,將乳糖(lactose)轉換為核酮酸-5-磷酸(ribulose-5-phosphate) 之 6-phosphogluconate dehydrogenase(gnd),在大腸桿菌 K12 菌株中發現以 葡萄糖為碳源時, gnd 基因表現量是以醋酸為碳源時的 3 倍,因此生長速率增加 時, gnd 基因的表現量亦增加(Wolf et al., 1979)。. 2.1.4 生長速率調控機制的探討. 細胞生長速率對細胞生理功能的影響深遠複雜,因此生長速率對基因的調控 一直是許多研究的著眼點,目前對於生長速率改變時對基因調控的機制仍不是十 分清楚,相關的研究亦未臻成熟。我們已了解在不同的生長速率下,影響酵素活 性及其產量高低的機制可能是在轉錄或轉錄後過程受到調控(transcription level 或 post-transcription level)。例如:產生 pentose phosphate pathway 中催化第 一個步驟酵素的 zwf 基因,在生長速率改變時,可藉由轉錄的步驟來調控(Rowley et al., 1991);而產生大腸桿菌外膜蛋白質的 ompA 基因隨著生長速率提高而增 加表現量,則是因為生長速率的不同影響了 ompA 基因 mRNA 的穩定性(Nilsson. 6.
(15) et al., 1984; Georgellis et al., 1992)。 由於生長速率調控基因的方式不一,亦沒有特定的啟動子需求,目前仍有不 明確的調控機制存在,因此本論文由探討不同碳源對細菌生理所造成之變化,藉 此了解是否不同碳源可經由改變細胞內能量狀態、代謝途徑與生合成反應,以及 DNA 超 螺 旋 結 構 改 變 影 響 細 胞 生 長 速 率 , 或 是 鳥 苷 四 磷 酸 ( guanosine tetraphosphate, ppGpp)造成細胞分裂基因表現改變與菌體型態變化,進而影響 了細胞生長速率。. 2.1.5 細胞內主要來源 ATP. 細菌吸收並分解環境或培養基中的養分後,以 ATP 形式存在於細胞內,這些 ATP 隨即被消耗運用於溶質運送、單體(單醣、核苷酸、脂肪酸)與大分子(核 酸、脂質、蛋白質、多醣類)合成等各種需能反應上。這些反應所製造出的物質 不但造成細胞重量增加,也使得細菌能持續分裂與生長,因此微生物的生長與 ATP 有直接關係。 細菌體內合成 ATP 的機制有二(Gottschalk, 1988): 1. 氧化磷酸化反應(oxidative phosphorylation):利用電子從氧化還原電位較低 的供給者轉移到電位較高的接受者時,伴隨 ADP 與無機磷酸根(Pi)結合產 生 ATP。這一部份的反應多發生於呼吸鏈或光合成反應中。 2. 受質水平磷酸化反應(substrate level phosphorylation) :在有機受質被分解成 小分子中間物時,放出的高能磷酸鍵與 ADP 結合而生成 ATP。. 7.
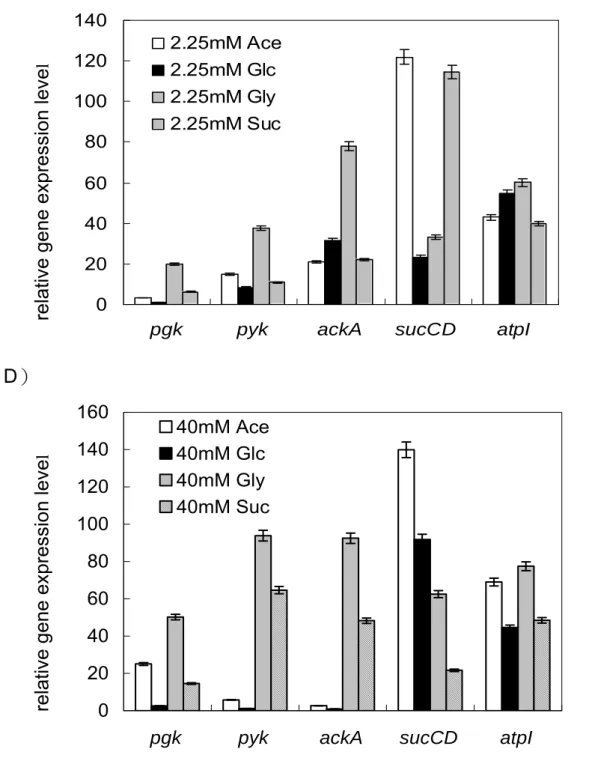
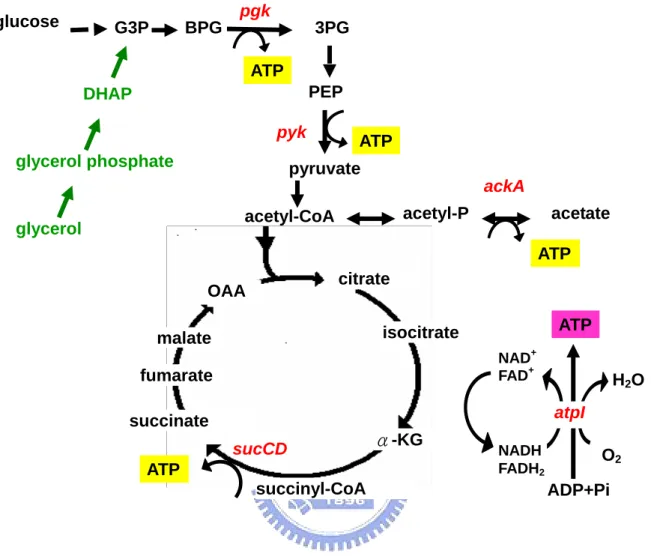
(16) 2.1.6 碳源對代謝產生能量途徑之影響. 不同環境下微生物的代謝產能途徑會隨之改變,大腸桿菌利用不同受質經呼 吸酵素產生 NADH,再經由細胞內 quinone pool 傳給電子接受者,以進行氧化 磷酸化反應來產生 ATP 供給生長所需。在有氧環境下,若以一分子葡萄糖為例, 由 Fig 7 代謝途徑可知葡萄糖經由糖解作用(glycolysis)生成兩分子丙酮酸 (pyruvate)的過程中,先後分別由磷酸甘油激酶(phosphoglycerate kinase, pgk) 與丙酮酸激酶(pyruvate kinase, pyk)催化,可經由受質水平磷酸化反應產生 2 個 ATP 。接著 pyruvate 以乙醯輔酶 A(acetyl-CoA)為主要的分解代謝產物,進 行完整的 TCA cycle 後氧化成二氧化碳,在此過程中,由琥珀醯硫激酶(succinyl thiokinase, sucCD)將琥珀醯硫 A(succinyl-CoA)催化成琥珀酸(succinate) 步驟中,可生成一個高能 ATP 分子,同時經由電子傳遞鏈(electron transfer chain) 伴隨進行氧化磷酸化作用,由 ATP 合成酶(ATP synthase, atp)產生 38 個 ATP 分子。若是處於無氧的環境下,同樣是一分子的葡萄糖僅可藉由發酵作用 (fermentation)或厭氧呼吸產生 ATP,在此過程中大腸桿菌是以醋酸激酶 (acetate kinase, ackA)將帶有高能磷酸根的乙醯磷酸(acetyl-phosphate)轉變 為醋酸,並伴隨一個 ATP 分子產生。 綜合以上所述,在代謝途徑上與 ATP 直接合成的相關酵素共有五種,分別是 磷酸甘油激酶(phosphoglycerate kinase, pgk)、丙酮酸激酶(pyruvate kinase, pyk) 、琥珀醯硫激酶(succinyl thiokinase, sucCD) 、ATP 合成酶(ATP synthase, atp)與醋酸激酶(acetate kinase, ackA) 。本實驗將探討不同碳源為生長基質時, 對於 pgk、pyk、ackA、sucCD 與 atp 等基因的影響與表現量改變情形。. 8.
(17) 2.1.7 ATP/ADP 比例. ATP 是生物系統能量可供利用的形式,作為自由能的直接供應者而非用來長 期儲存自由能。 ATP 轉換速率(turn over rate)在細胞中極為快速,一分子 ATP 在形成後一分鐘內即被消耗,唯有持續將 ADP 與 Pi 結合再生 ATP 才能提供細 胞所需的能量(Stryer, 1995) 。因此產能與耗能的反應不斷交替進行著,使得細胞 內 ATP 與 ADP 二者間的濃度比例與細胞內生理狀態有關。例如:在肌細胞中 ATP/ADP 比例可調控肌動蛋白(actin)的作用(Carlier et al., 1993) ;反之,肌 細胞內的肌酸激酶(MM creatine kinase)可調控 ATP/ADP 比例(Korge et al., 1993) 。目前已知在有氧環境下,大腸桿菌細胞內 ATP/ADP 比例較生長於無氧下 為低(Hsieh et al., 1991) 。許多活體外試驗均證實 ATP/ADP 比例影響 DNA 超 螺旋結構。當培養環境之含氧量降低時 ATP/ADP 比例提高, DNA 超螺旋隨之 旋緊;或將菌體由一般培養基中轉移至高鹽(NaCl)環境下,ATP/ADP 比例會立 刻增加且 DNA 超螺旋結構也變得較為緊密(Hsieh et al., 1991)。推論其作用為 ATP/ADP 比例影響了 gyrase B 的活性,進而影響細胞 DNA 超螺旋結構。再 者,也發現 ATP/ADP 比例會影響 RNA 的穩定性,磷酸酶(phosphorylate kinase, PPK)可藉由磷酸根的轉移來調控 mRNA 穩定性,當 RNA 處在分解狀態下, PPK 會將 ATP 水解且於 RNA 的 5’ 或 3’ 端形成多磷酸鏈(polyphosphorylate chain) ,此作用可增加 RNA 的穩定性;但加入 ADP 時, PPK 則會催化 ATP 合 成而不進行加磷酸鏈反應。由此可知 ATP/ADP 比例高時 RNA 較穩定,反之則 較不穩定(Blum et al., 1997)。 Rohwer 等學者亦發現藉由誘導 atp 操縱子 (H+-ATPase)表現量的不同而改變 ATP/ADP 比例時,會影響大腸桿菌攝取葡 萄 糖 的 PTS ( phosphoenolpyruvate phosphotransferase system ) 活 性 , 當 ATP/ADP 比例低時會降低 PTS 活性,反之則使 PTS 的活性升高(Rohwe et al., 1996) 。由這些例證均可肯定 ATP/ADP 比例的確可反應細胞內的生理狀況,因此. 9.
(18) ATP/ADP 比例已成為相關研究中的反應參考指標。. 2.1.8 DNA 超螺旋結構(DNA supercoiling)與 topoisomerase. 由電子顯微鏡觀察到 DNA 分子具有雙股雙股之間緊密纏繞的現象,此種 DNA 形式稱為超螺旋結構(supercoiling)。實驗證實菌體 DNA 以自由能最小的 negatively supercoiling 狀態存在(Worcel and Burgi, 1972; Sinden and Pettijohn, 1981) ,且在 negative 狀態下,有利於雙股 DNA 在複製、重組及轉錄時的分離。 為了解決 DNA 半保留複製造成的雙股解旋(untwining)與鏈結(catention) 現象,不論在原核或真核細胞中皆有某些酵素可以藉由改變 DNA linking number 來解決這些問題。而這些酵素亦可使 DNA 形成各種拓樸異構物,因此就統稱它 們為拓樸異構酶(topoisomerases)。 目前所發現的 topoisomerases 以它們的作用形式大致上可分為兩類: type I topoisomerase 利用切斷雙股 DNA 的一股使其穿過另一股,再以 ligase 將其 連接;而藉由將兩股 DNA 完全切斷,讓兩股相互繞過再結合的則屬於 type II topoisomerase(Wang, 1985)。目前在大腸桿菌中已發現 topoisomerase I、II、 III、IV 四種形式;其中 topoisomerase I、III 屬於 type I,而 topoisomerase II、 IV 則屬於 type II。已知 DNA topoisomerase III(topB)在細胞中含量非常低, 且 DNA topoisomerase III 突變株對 DNA 超螺旋結構並沒有顯著的影響。至於 DNA topoisomerase IV 目前尚無豐富的相關研究,僅知此酵素只在 topA 突變株 中才會經由 parC 及 parE 基因轉錄(Drlica, 1992)。因此生物體內 DNA 超螺 旋結構最主要是由 DNA topoisomerase I 與 II 來調控,所以多由此二酵素進行 相關研究。以下簡要地介紹這兩個重要的酵素。. 10.
(19) 1. DNA topoisomerase I 在 1971 年所發現的 DNA topoisomerase I 是由 topA 基因所轉譯,早期一 直被稱為 ω 蛋白質,為 110 kDa 大小的單一多肽鏈(Gellert, 1981)。 DNA topoisomerase I 可將雙股 DNA 切斷其中一股,此酵素會與斷裂的 DNA 形成共 價中間物,繞著另一股鬆開後再以 ligase 連接。 DNA topoisomerase I 僅作用 於 negatively supercoiling DNA,對 positively supercoliling DNA 並無作用,且 DNA topoisomerase I 並不能將 DNA 超螺旋結構完全鬆開,研究顯示當 DNA topoisomerase I 存在時,若加入 DNA gyrase 的抑制劑,DNA 超螺旋結構鬆開 的速率並不受影響。因此推論 DNA topoisomerase I 的功能主要是為了協調 DNA gyrase 作用,避免 DNA 超螺旋過度纏繞(Wang, 1985)。. 2. DNA topoisomerase II(DNA gyrase) 1976 年在大腸桿菌中分離出可將鬆散結構 DNA 纏繞成超螺旋形式的 DNA gyrase,因為緊繞的 DNA 具有較高自由能,所以在 DNA gyrase 作用時需要水 解 ATP 來提供反應所需要的能量。 DNA gyrase 含兩個次單元(subunit): gyrase A(gyrA)與 gyrase B (gyrB) ,每一分子 DNA gyrase 由二個 gyrase A 與二個 gyrase B 次單元所組成,在細胞內二者皆以雙聚合體(dimer)的形式存 在,且只有當這兩個次單元結合形成四聚體(tetramer)一起作用時, DNA gyrase 才有完整的活性。具有旋緊 DNA 功能的是 gyrase B,而 gyrase A 則有放鬆 DNA 之活性(Cozzarelli, 1980)。 DNA gyrase 可水解 ATP 以誘發 DNA 雙股 的斷裂,切開後的 DNA 其 5’ 末端會固定在 gyrase A 上,避免其自由旋轉,待 斷裂缺口繞過另一段 DNA 後,再加以接合(Wold et al., 1994)。 在細胞內 gyrase A 及 gyrase B 兩個次單元可相互協調,來保持細胞內 DNA 超螺旋結構的恆定狀態,若太緊時才由 DNA topoisomerase I 進行鬆散的工 作。由於 gyrase B 是細胞內唯一具有將 DNA 超螺旋結構纏繞的酵素活性,因. 11.
(20) 此細胞內不能完全沒有 gyrase B 的活性(Gellert, 1981)。. 2.1.9 細胞內能量因子調控 DNA 超螺旋結構. 目前發現 DNA gyrase 是菌體中唯一具有纏緊 DNA 超螺旋結構功能之酵 素,這個反應必須伴隨著 ATP 水解以獲得能量,目前有愈來愈多的研究證實細胞 內 ATP/ADP 比例是影響 DNA 超螺旋結構的重要因子。當細胞內 ATP 缺乏時, DNA 超螺旋結構會變得鬆散,若 ATP/ADP 比例增加,質體 DNA 超螺旋結構亦 隨之旋緊(Workum et al., 1996) 。此外體外實驗中也已證實純化後的 DNA gyrase 活性受 ATP/ADP 比例影響,所以目前推論 ATP/ADP 比例是藉由改變 DNA gyrase 活性而控制 DNA 超螺旋結構。從此觀點可了解在不同生長環境下改變 DNA 超螺旋結構的機制: 1. 含氧量:將大腸桿菌培養環境從有氧迅速變成無氧後,ATP/ADP 比例逐漸上 升,且在不同的時間點抽取質體觀察也發現,其 DNA 超螺旋結構呈現愈來愈 緊的現象(Hsieh et al., 1991)。 2. 溫度:在 37℃ 培養時,大腸桿菌質體 DNA 扭轉程度較 17℃ 時緊,而當溫 度從 30℃ 增加至 42、45、47 或 50℃,觀察質體 DNA 超螺旋程度也是漸 增的,這之間的變化亦伴隨著 ATP/ADP 比例而上升(Goldstein and Drlica, 1984; Camacho et al., 1995) 。 3. 滲透壓:在 0.5 M NaCl 影響下,大腸桿菌之 ATP/ADP 比例較高,且質體 DNA 超螺旋結構為較緊密的狀態( Hsieh, et al., 1991 )。. 12.
(21) 2.2. 貧乏培養(starvation)與 ppGpp 對大腸桿菌生理之影響. 2.2.1 迫切反應(stringent response)與 ppGpp 對大腸桿菌之生理調控. 細菌生活在自然界中,常會因為環境因素的不穩定而遭受到相當的壓力 (stress),因此細菌會因應環境而產生不同的 global regulator 來調節基因表 現。細胞內常見的 global regulator 為 ppGpp 與 cAMP 等物質,統稱為警報素 (alarmone) ,可透過此類調控子來改變生理狀況來適應不同環境。當大腸桿菌處 於胺基酸貧乏的環境時,體內會產生一系列迫切反應(stringent responses)以應 付不利環境。迫切反應的調控是由 ppGpp 所影響,透過 ppGpp 抑制核糖體和其 他大分子合成等應急反應,活化某些胺基酸操縱子的轉錄表達,抑制與胺基酸運 轉無關的系統,活化蛋白水解酶等,以節省或開發能源度過難關。 當 培 養 基 缺 乏 必 需 胺 基 酸 時 , 大 腸 桿 菌 能 合 成 鳥 苷 五 磷 酸 ( guanosine pentaphosphate, pppGpp)或鳥苷四磷酸(guanosine tetraphosphate, ppGpp) ,。 這是由於菌體在缺乏營養的環境下,會啟動 relA 基因促使 ppGpp 合成。 ppGpp 的 合 成 在 菌 體 的 生 理 與 生 化 代 謝 上 扮 演 重 要 功 能 , 包 括 抗 藥 性 ( antibiotic resistance)與抗殺蟲劑(biocide resistance)的功能也與 ppGpp 有關(Greenway and England, 1999; Xu et al., 2002)。 與 ppGpp 合成的相關基因除了 relA 基因外,另有 spoT 基因。 SpoT 具 有 synthase 與 hydrolase 兩種區域(domain) ,可負責 ppGpp 的合成與分解。 通常於缺乏碳源與磷酸時會活化 spoT 基因,進行 relA-independent 方式來合成 ppGpp ,同時 SpoT 也具有將 ppGpp 分解的功能(Murray and Bremer, 1996) 。. 13.
(22) 2.2.2 ppGpp 作用機制與對細菌生理影響. 在旺盛生長的細胞中約有 65~90% tRNA 是載有胺基酸的。當胺基酸缺乏 時,亦即不負載胺基酸的 tRNA 增多,這種不負載胺基酸的 tRNA 是細胞產生迫 切反應之重要信號,仍能與核糖體 A 位結合。在正常的蛋白質合成過程中,將負 載有胺基酸之 tRNA 運轉到正在延伸的多肽鏈上需要 GTP,由於多肽鏈的停止使 得大量 GTP 便被用作合成 ppGpp 的前趨物。 自 1979 年 Gallant 學者探討細菌迫切反應之機制,已知當胺基酸貧乏時,未 負載胺基酸的 tRNA 進入核糖體 A 位會導致兩種效應:一是空載的 tRNA 與 mRNA 密碼子結合而中止蛋白質生成,二為刺激 relA 基因的啟動而產生迫切控 制因子(stringent factor, 簡寫為 SF) ,SF 為核糖體相關蛋白質,會促使 GTP 或 GDP 與 ATP 反應,進而合成 pppGpp 或 ppGpp (Gallant, 1979) 。而 ppGpp 累積會抑制 rRNA 基因(rrn)P1 啟動子的轉錄,透過 ppGpp 與 σS 因子結合 以降低 RNA 聚合酶與 P1 啟動子的親合力,藉以降低 rRNA 的轉錄。由此可知, ppGpp 會與 RNA 聚合酶結合以抑制其活性,使基因的轉錄下降,或是可能影響 RNA 聚合酶與啟動子結合的專一性,從而改變了基因轉錄的效率,進而調控蛋白 質的合成能力(Gallant, 1979; Hernandez and Bremer, 1990)。 在細菌正常生長期間 ppGpp 含量極低,當進入對數生長期時, ppGpp 含量 會隨生長速率下降而上升;而生長速率對於具高穩定性的 RNA(tRNA 及 rRNA) 基因所存在的正比表現關係是藉由 ppGpp 所調控(Neidhardt et al., 1990)。 除了胺基酸貧乏時會引發 ppGpp 合成,碳源與磷酸的缺乏以及生長速率下降 時都會促使 ppGpp 產生,以降低新的蛋白質合成進而回饋抑制穩定性 RNA 的 轉錄,因此使各種酵素表現量也跟著下降(Joseleau-Petit et al., 1999) 。受 ppGpp 所調控的基因不只是一個或幾個操縱子,而是影響一連串迫切反應,主要包括:(1) 節流:如抑制大部分蛋白質、 rRNA 、 tRNA 、脂肪、磷酸與核苷酸等合成;(2). 14.
(23) 開源:如促進 lac、ara、his、trp 及 arg 等操縱子開放,活化蛋白質水解系統以 提供胺基酸. 2.3. 大腸桿菌之細胞分裂基因. 2.3.1 細胞分裂 ( cell division ). 大腸桿菌以二分裂的方式進行繁殖,其分裂是從染色體複製開始。為了使遺傳 物質能均等地傳给兩個子細胞(daughter cell) ,染色體複製與細胞分裂必須有嚴 謹的調控。正常生長狀態下,若染色體複製未完成則細胞就不能分裂,複製一旦 完成細胞隨即開始分裂。在生長緩慢時,大腸桿菌待染色體複製完成且分離後才 進行細胞分裂,分裂完成之菌體才開始下一次染色體複製。在快速生長時,染色 體複製一次尚未结束時,在尚未分開的染色體上又開始了新的複製,導致染色體 複製的時間大為縮短,也因此細胞分裂的倍增時間大為減少(Donachie, 1993) 。 接種到新鮮培養基中的大腸桿菌細胞,從周圍環境中選擇地吸收營養物質。在 細胞内合成所需的 RNA、DNA、蛋白質及酵素等大分子物質,細胞體積不斷增大, 待染色體複製完成後即進行分裂過程。首先是以複製完成之染色體開始分離,同 時細胞中央的隔板(septum)也開始對稱地向中心凹陷生長,使染色體均等地分 配到凹陷兩側。隨著細胞膜的向内凹陷,母細胞的肽聚糖層也跟著由四周向中心 生長,把細胞膜分為兩層,每層分别成為子細胞的細胞膜。肽聚糖層也分為兩層, 直至中央會合,形成由細胞質膜和肽聚糖組成之隔膜。隔膜完全形成後,兩個子 細胞分開,完成了一次細胞分裂(Romberg and Levin, 2003)。. 15.
(24) 2.3.2 細胞隔板的形成(septum formation). 在大腸桿菌中,當染色體複製與分離完成後會在細胞中間形成隔板 (septum) ,並分裂成兩個子細胞(Donachie, 1993a; 2001b) 。隔板是由 FtsZ 蛋 白質所構成的環狀多聚體環(Z ring) ,Z ring 的形成至少有九種蛋白質參與,分別 為 FtsZ、FtsA、FtsQ、FtsL、FtsI、FtsW、FtsK、FtsN 與 ZipA 等蛋白質,其 中 FtsZ 為周邊膜蛋白,屬於 actin/hsp70/sugar kinase family,可水解 ATP 以促 使隔板形成,而 ZipA 是一種與 FtsZ 交互作用的膜蛋白,也會促進 Z ring 的聚 合,其餘與細胞分裂有關之蛋白質皆具有至少一個 membrane domain ,可與細 胞膜進行交互作用以幫助細胞隔板的形成。 FtsZ(filamenting temperature sensitive)是從大腸桿菌溫度敏感型突變體中 分離到的基因,該基因與大腸桿菌細胞分裂密切相關。自 1980 年首次從大腸桿菌 中選殖(clone)到該基因後(Lutkenhaus et al., 1980),又陸續在多種生物中發 現了 ftsZ 基因的存在。隨後研究發現 FtsZ 蛋白為原核細胞的骨架蛋白,通常在 細胞的分裂位點聚集形成一個 Z ring ,從而引發並控制原核細胞分裂過程。 FtsZ 為細胞隔板形成最重要的蛋白質,其蛋白質結構與真核生物的微管蛋白(tubulin) 同源,同樣具有 GTPase 活性,可水解 GTP 以進行 Z ring 的聚合作用。在正 常的細胞分裂周期中, ftsZ 基因發揮著不可替代的作用,如果胞内 ftsZ 基因的 表現異常,則會導致細胞分裂受阻並形成不分裂的長絲狀(filamentous)細胞(Bi and Lutkenhaus, 1991; Addinall et al., 1996)。 當 FtsZ 過量時會聚合過多的 Z ring ,而在細胞兩端也形成隔板導致 minicell 產生,若 FtsZ 含量再更多時則會完全抑制細胞分裂,這是因為 FtsZ 與 FtsA 之間的比例在正常分裂時是固定的,若之間比例變動過大時則導致細胞隔板 無法形成以致不能進行細胞分裂(Dewar et al., 1992)。. 16.
(25) 2.3.3 細胞隔板形成之抑制者. 隔板是由 FtsZ 蛋白質所構成的 Z ring ,形成的位置目前推論至少有兩種機 制,一為核酸聚集(nucleoid occlusion) ,當染色體複製完成後,將兩個子染色體 由細胞中央分別移動至兩極, 核酸會佔據 Z ring 形成的位置,因此限制了 Z ring 形成的位置:另一為 MinCD 複合物, MinCD 複合物是 Z ring 抑制者,由 MinC 與 MinD 兩種蛋白質所組成,分布於細胞的兩端,會抑制 FtsZ 在此處聚合,使 得 Z ring 只在細胞中央形成隔板。 MinC 的 C 端會與 MinD 結合, N 端則與 FtsZ 結合以抑制 Z ring 合成; MinD 具有 ATPase 活性,可分解 ATP 在細胞 膜內側形成雙聚體分子,並與 MinC 結合以抑制 FtsZ 在膜上聚合 (Rothfield and Garcia-Lara, 1996; Pichoff and Lutkenhaus, 2001; Hu et al., 2003; Lackner et al., 2003)。. 在本論文中,除探討 ATP/ADP 比例對大腸桿菌生長速率與 DNA 超螺旋結 構之間的關係,以及偵測代謝途徑上與 ATP 產生相關酵素之基因的研究,也將一 併探討碳源與其他調控因子對細胞分裂基因 ( ftsZ, minC, minD ) 的影響。. 17.
(26) 參、基本理論. 3.1 批次培養(batch culture)之細胞生長模式. 批次培養法是指在細胞的培養過程中不再添加任何新鮮培養液,僅提供一定 量培養液供細胞生長所需。由於細胞生理狀況、培養液組成養分及細胞代謝產物 等因素,在批次培養過程中均會影響細胞的生長。因此當進行批次培養時,細胞 的生長具有一定模式。. 3.1.1 批次培養下細胞之生長曲線. 在批次培養系統下細菌的生長過程可分為六個時期(Lee, 1992):. 1. 遲滯期(lag phase): 微生物開始適應新的環境,菌體的重量會增加,但細胞不會分裂繁殖。 2. 加速生長期(accelerated growth phase); 菌體開始分裂,細胞數目開始增加,且細菌生長速率逐漸增加,而達到最大 生長速率。 3. 對數生長期(exponential growth phase); 細胞以最大速率進行分裂,且在一定時間間隔內分裂一次,此時期菌體的數 目會隨時間呈對數增加。 4. 減速生長期(decelerated growth phase); 菌體生長及分裂速率均開始降低。 5. 靜止期(stationary phase) 細胞的增殖速率與死亡速率相等,細胞數目維持一定。. 18.
(27) 6. 死亡期(death phase) 因環境中的養分已被耗盡,而代謝產物累積對菌體產生毒害。細胞死亡速率 較增殖速率快,細胞數目開始降低。. 上面所述細胞生長曲線的六個時期中,一般研究細菌生理及生長速率時,均 取對數生長時期的菌體加以測定。. 3.1.2 菌體倍增時間. 菌體自開始培養,到最後總菌群為 N 時,可依下列公式表示:. N = N0 × 2n----------(3-1). 其中 N:最後菌數 No:起始時間菌數 n:菌體培養時的世代數. 對 3-1 式兩邊取對數. log10N= log10N0+n log102 ↓ n = (log10N-log10N0)/ log102----------(3-2). log102=0.301 代入 3-2 式. 19.
(28) n = (log10N-log10N0)/ 0.301 ↓ n =3.3 (log10N-log10N0)----------(3-3). 菌群加倍所需時間為倍增時間,以下列公式表示:. g=t/n----------(3-4). 其中 g:菌體倍增時間 t:發生 n 次世代所需時間. N0 和 N 為已知,則可由 3-3 式得到 n 值,將其代入 3-4 得到 g 值 即菌體的倍增時間。. 3.2 連續式培養之動力學模式. 基於研究或工業上生產之需要,必須使菌體維持在對數生長期(log phase) , 利用連續式培養則可達到此目的。連續式培養法是控制新鮮且無菌之培養液流入 槽體內的速率,並且以相同速率將槽體內多餘的培養液排出,以維持反應槽內培 養液體積。連續式培養可以依對流量控制方式不同而分成兩種形式。一種是利用 可調式幫浦(pump)直接控制流進基質的量,同時從反應槽中移除相同體積的 菌液,以維持反應槽之培養體積。流入基質成份中,除了最基本的營養成份以供 細胞生長外,還含有特定的限量基質(limited nutrient) ,而此限制生長的營養因 子控制了在穩定狀態(steady state)下細胞族群大小,此種方式之培養稱為化學. 20.
(29) 恆定(chemostat)的連續式培養(chemostat 是指在穩定狀態時培養基內的化 學組成維持一定)。 另一種方式稱為濁度恆定(turbidostat)連續式培養,其培 養液之流量是以光電管來控制,這種光電管可以很敏銳的察覺反應槽中細胞濃度 的變化,當培養液中菌體的吸光度超過設定值時,可依據光電管發出的訊號來添 加基質以維持一定的細胞濃度(Wang et al., 1981)。本論文中採用化學恆定的連 續式培養法來控制細胞生長速率。. 連續式培養之動力學模式如下(Wang et al., 1981):. 當連續式培養達穩態時,對反應槽中細胞質量作質量平衡:. cell in - cell out + cell growth - cell death = cell accumlation. FX0/V-FX/V+µX-αX=dX/dt---------(3-5). 其中 X0: 進料時的細胞質量 X: 細胞質量 µ: 比生長速率 (hr -1) α: 比死亡速率 (hr -1) F: 培養液流量 (hr -1) V: 培養液體積 (L) t: 時間 (hr). 由於進料培養液已滅菌,所以為無菌狀態(即 X0=0) ,而且在連續式培養中,比 生長速率遠大於死亡速率(µ>>α),所以 3-5 式可以簡化如下:. 21.
(30) (µ- F/V)X = dX/dt----------(3-6). 當反應槽達穩定狀態時,單位時間內菌體質量變化為零,即. dX/dt = 0. 所以由(3-6)式可得 µ= F/V-------(3-7). 又(3-7)式中 F/V 可定義為 F/V = D ------------(3-8). 且 D = 1/T. 其中 D:稀釋速率 T:培養液流經槽體的平均滯留時間. 也就是在 chemostat 的連續式培養中,操作者只需決定培養液進料時的流 量(F)或稀釋率(D)便可以控制細胞的比生長速率。所以在低稀釋速率時,相 對槽體內所含的限制營養成分濃度降低,細胞生長速率減慢;在稀釋速率高時, 相對槽體內所含的限制營養成分濃度提高,細胞生長速率加快。. 3.3 聚合酶連鎖反應(Polymerase chain reaction,PCR)之原理. 聚合酶連鎖反應是在活體外,以基因組 DNA 當模板(template) ,將某段特 定序列之套數擴增至數百萬倍,利於實驗進行分析。PCR 原理係利用高溫將雙股 基因組 DNA 分成單股(denature) ,並以兩段單股的引子(primer)與單股基因 組 DNA 互補接合(primer annealing) ,經由耐熱性高的 Taq DNA polymerase. 22.
(31) 產生聚合反應,進行引子下游的序列合成(extention),至此稱為一個週期。之 後重新進行 DNA 分股步驟,依序進行多個週期。當進行 n 個週期時,便產生 2n 條特定序列雙股 DNA,藉此達到擴增大量特定序列之目的。. 23.
(32) 肆、材料、設備與方法. 4.1 實驗材料. 4.1.1 菌株質體與噬菌體. 1. 菌株 Escherichia coli K12(W3110) :大腸桿菌野生菌株。 Escherichia coli K12(W3110)△relA △spoT:大腸桿菌 ppGpp 突變株。 2. 質體 pBR322(Ampr Tetr). 4.3 kb. 3. 噬菌體 P1 vis lysate. 4.1.2 藥品與試劑. 1. Sigma Chemical Co. ethylenediaminetetraacetic acid(EDTA), rifampin, deoxyribonucleic acid, pyruvate kinase, adenylate kinase, sodium dodecyl sulfate ( SDS ) , chloroquine, ampicillin, tetracyclin, ethidium bromide ( EtBr ) , diethyl pyrocarbonate(DEPC), dextran sulfate 2. E. Merck. N,N-methylformamide, ethanol, phenol, isoamyl alcohol, chloroform, phosphoenopyruvate, glacial acid, formamide, formaldehyde, ammonium. 24.
(33) acetate, 3-N-Morpholino propanesulfonic acid(MOPS), sodium acetate, potassium citrate, Denhardt's solution 3. Promega Biotec. EcoR I 4. Difco Laboratories yeast extract, tryptone, agar, casamino acid 5. J. T. Baker HCl, NaOH, glacial acid, MgCl2‧H2O, NaCl, H2SO4, (NH4)2SO4, CaCl2, CaCl2‧2H2O,. MgSO4‧7H2O,. FeSO4‧7H2O,. K2HPO4,. KH2PO4,. NaH2PO4‧H2O, Na2HPO4‧7H2O, dextrose(glucose), 2-propanol, Tris (Base), glycerol, dimethyl sulfoxide(DMSO), potassium acetate, sodium acetate, citric acid‧H2O, NaNH4HPO4‧H2O, boric acid 6. Riedel-de Haen C4H4Na2O4‧7H2O 7. Roche anti-DIG-AP, positively charged nylon membrane, CSPD, PCR DIG Probe Synthesis Kit, DIG Wash and Block Buffer set, ATP Bioluminescence Assay Kit HSII 8. MRC TRI reagent 9. Amresco agarose, ammonia persulfate 10. Becton Drive BBLTM gram stain kit ( crystal violet solution 、 iodine solution gram decolorizer solution and safranin solution). 25.
(34) 4.1.3 緩衝液. 1. TAE buffer 40 mM Tris acetate, 1 mM EDTA 2. TBE buffer 45 mM Tris borate, 1 mM EDTA 3. 10X formaldehyde gel running buffer:0.2 M MOPS, 80 mM sodium acetate, 10 mM EDTA(pH 7.0) 4. RNA gel loading dye(Ambion cat#8552) 5. 20X SSC, pH 7.0:3 M NaCl, 300 mM sodium citrate 6. 20X SSPE, pH 7.0:3 M NaCl, 0.2 M NaH2PO4⋅H2O, 0.02 mM EDTA (pH 7.4) 7. Hybridization buffer:5X Denhardt's solution, 33% formamide, 6.6X SSPE, 1% SDS, 50﹪dextran sulfate, sterilized by filter 8. Tris-glycine electrophoresis buffer:25 mM Tris-base, 250 mM glycine(pH 8.3), 0.1% SDS 9. Rifampicin solution:1 g rifampicin in 10 ml DMSO, sterilized by filter 10. DEPC treated water:1 ml DEPC in 1 L ddH2O, reacted 10 hrs, sterilized by autoclave 11. ATP assay reagent:75 mM potassium phosphate buffer, 15 mM MgCl2 12. ATP+ADP assay reagent:75 mM potassium phosphate buffer, 15 mM MgCl2, 0.5 mM phosphoenopyruvate(PEP), 4U pyruvate kinase. 26.
(35) 4.1.4 培養基與培養液. 1. 基礎培養液(minimal medium)(/L):每公升 solution I 加入 1 ml solution II Solution I:KH2PO4 5.44g, K2HPO4 10.49 g, (NH4)2SO4 2 g, casamino acid 0.5 g Solution II:MgSO4‧7H2O 50 g, MnSO4‧H2O 5 g, FeSO4‧7H2O 0.125 g, CaCl2‧2H2O 0.735 g (以 0.2N H2SO4 為溶劑) 2. LB (Luria-Bertani)培養液 1% tryptone, 0.5% yeast extract, 1% NaCl pH 7.0 3. LB – ampicillin 培養液 1% trpytone, 0.5% yeast extract, 1% NaCl pH 7.0, 80 µg/ml ampicillin 4. LB 培養基 1% tryptone, 0.5% yeast extract, 1%NaCl pH 7.0, 1.5% agar 5. LB – ampicillin 培養基 1% tryptone, 0.5% yeast extract, 1% NaCl pH 7.0, 80 µg/ml ampicillin, 1.5% agar 6. LB − ampicillin − tetracyclin 培養液 1% tryptone, 0.5% yeast extract, 1% NaCl pH 7.0, 80 µg/ml ampicillin, tetracyclin 20 mg/ml 7. LB ( ampicillin ( tetracyclin 培養基 1% tryptone, 0.5% yeast extract, 1% NaCl pH 7.0, 80 µg/ml ampicillin, tetracyclin 20 mg/ml, 1.5% agar 以上培養基(液)若須添加碳源、抗生素或指示劑時皆待 121℃ 高溫高壓滅菌後冷卻 至 55℃後再加入。. 27.
(36) 4.1.5 電泳膠體. 1. DNA gel 1.0 % agarose 溶於 1 X TAE buffer 中 2. RNA gel 1.2 % agarose, 1 X formaldehyde gel running buffer, 2.2 M formaldehyde. 4.2 實驗方法. 4.2.1 大腸桿菌生長條件與培養方法. 1. 繼代培養 第一代菌株皆以 LB 為培養液,挑單一菌落培養於培養液,在 37℃ 恆溫震 盪箱以轉速 200 rpm 培養。菌株之繼代保存則以 LB 作為繼代保存培養基, 利 用劃線法將菌株接種於平板培養基,在 37℃ 恆溫箱培養到適當大小的菌落,置 於 4℃ 保存。. 2. 批次培養 為了使菌體適應生長環境,所以需先作二次繼代培養,然後再於第三代時培 養在所需條件。 第一代培養:挑取單一菌落接種至 5 ml LB 培養液,培養 6 ~ 8 小時使菌 體活化。. 28.
(37) 第二代培養:取 50µl 菌液接種到 5 ml 基礎培養基(minimal medium), 並以最終濃度 2.25 mM 或 40 mM 葡萄糖作為碳源,然後於 37℃ 隔夜培養, 此培養目的在使細菌適應新的生長環境。 第三代培養:取適量菌液至含不同碳源的基礎培養基中進行培養,培養時所 加的菌量在 OD600 上升範圍控制於 0.02~0.025 之間。本實驗所使用的碳源為 醋酸(acetate)、葡萄糖(glucose)、甘油(glycerol)與琥珀酸(succinate)。 之後於 37℃ 200 rpm 培養至 OD600 吸光值為 0.4~0.45 時,置於冰上 10 分 鐘後,以 5,000 rpm 離心 10 分鐘收取菌體,目的為取得處於對數生長期的菌 體進行 RNA 萃取與 ATP 與 ADP 含量分析。. 3. 連續式培養 本實驗連續式培養是採用化學恆定的連續式培養( chemostat continuous ,其最大的優點在於只需決定稀釋速率(D)便可以得到比生長速率(µ) culture) (詳見 3.2) ,而稀釋率由可調式幫浦便可決定,因此可控制細菌的比生長速率。. (1) 培養基的製備 化學恆定連續式培養使用之培養基與批次培養之基本培養基成分相同,唯碳 源的添加為每公升 2.25 mM,在此以 carbon limitation 來限制菌體生長。由於 連續式培養須消耗大量培養液,所以我們以 20 公升的儲水桶(Nalgene Cat. No. 2250-0050)為單位,每桶裝 17 公升基礎培養基,配製數桶後於高溫高壓下滅 菌 90 分鐘,於滅菌後待培養液溫度下降至室溫時再添加碳源. (2) 連續式培養系統之裝置 連續式培養系統主要可分為四個部份:槽體、流速測定管、培養液供應桶、 廢液收集桶。. 29.
(38) a. 槽體之組裝: 槽體為連續式發酵之主體,加入 1 公升去離子水,將上蓋裝配鎖緊後,於 冷凝管出口及進氣口接上 0.22 µm 濾菌膜,各出口及接頭均用鐵夾封閉並在管 口處包上鋁箔紙。以打氣幫浦測試槽體是否有漏氣情況,直到調整至完全封閉後 才可加以滅菌。滅菌時須打開冷凝管出口以防止滅菌時壓力的變化。. b. 流速測定管: 將滴定管頂部以棉花塞住並包上鋁箔紙。滴定管之下方接上三插管,再接 上矽膠管而分成兩個接頭,其中一個與槽體相接,另一個則與進料管相接。並在 管口處包上鋁箔紙,二接頭均用鐵夾封閉。. c. 培養液供應桶: 內裝有 17 公升培養液,桶上的蓋子有兩個接頭,一為負責進料之接頭,另 一則與過濾器相接,目的在平衡桶子內外的壓力並過濾空氣,所有出口及接頭均 用鐵夾封閉並在管口處包上鋁箔紙,以防止滅菌後污染。. d. 廢液收集桶: 以 20 公升空桶作為收集廢液用。桶上蓋子也有兩個接頭,一個與發酵槽出 料管相接以收集廢液,另一個接頭與裝有棉花之注射筒的筒身相接,以平衡桶子 內外的壓力並過濾空氣,出口及接頭均用鐵夾封閉並在管口處包上鋁箔紙,以防 止滅菌後污染。 槽體、流速測定管、廢液收集桶三者可同時置於 121℃,1 大氣壓下滅菌 60 分鐘或個別滅菌,而培養液供應桶則需 121℃,1 大氣壓下滅菌 90 分鐘。滅菌 後便可以將此四部份所有接頭相連接,所有操作過程皆需快速完成並過火滅菌以 避免污染。. 30.
(39) (3) 接菌前的準備及接菌 連續式培養系統於每次裝置完成後,由於安裝於蠕動幫浦之矽膠管長度可能 會有些許的不同,所以每次接菌前須先利用流速測定管測定本次蠕動幫浦的流 速,以作為往後調控培養液之流動速率的依據。並且在接菌之前需先啟動連續式 培養系統,將槽體內的去離子水置換成培養液,本實驗的通氣量是利用氣體流量 計來控制,而氣體流量計在使用之前需用排水集氣法校正。另外欲進行連續式培 養的菌株需先培養在 5 ml LB 培養液,於 37℃ 恆溫振盪培養至對數期後注入 發酵槽內,再調整蠕動幫浦的流速以控制生長速率進行連續式培養。. (4) 連續式培養的生長條件 本實驗是利用 BIOFLO 發酵槽及控制器來控制生長條件(New Brunswick 。槽體體積為 1.5 公升,培養體積為 1 公升,培養溫度為 37℃, Scientific Co.) 攪拌速率為 500 rpm,通氣量控制在 2 L/min,pH 值則維持在 6.9 ( 0.1。. (5) 細胞生長速率之控制與菌體收集 利用調節培養液流動的快慢以控制細胞生長速率,在每次變換培養液流動速率時, 至少須相隔 6 個反應槽的滯留時間(reactor residence time, τ),以確定菌體的生長 到達新的穩定期(steady state)。當菌體生長達穩定狀態後才可以收集樣本。. 4.2.2 細胞倍增時間的測定. 本實驗之生長條件及培養方法均與批次培養相同,但需每隔 0.5~1 小時用分光光 度計在波長 600 nm 下測定菌體之光學密度。由於光學密度主要是用來測定細胞濃 度,所以需先用分光光度計作細胞濃度和吸光度相互關係的校正曲線,再由吸光值轉換. 31.
(40) 成細胞濃度,則可求得細菌之生長曲線。由菌體的生長曲線可知當對數生長期中細胞數 目在 t1 與 t2,時間分別為 N1 與 N2 時,由 3-3 式得 n = 3.3(logN2 - N1) ,而細 胞倍增時間則由 3-4 式得 g = t / n 求得,其中 g 為菌體倍增時間, t 為發生 n 次. 世代所需時間。. 4.2.3 細胞轉型(transformation). (1) 原理 將質體導入細胞的過程稱為轉型。目前我們使用 CaCl2 將細胞穿孔,再以 heat-shock 的方式將質體送入。. (2) 實驗流程如下 a. 製備勝任細胞(competent cell) 活化欲轉型之細胞挑取單一菌落培養於 37℃ 14~18 小時後,將菌液稀釋 1/20 至新 LB 培養液中培養,至 OD600 吸光值約 0.4~0.6 時以 6,000 rpm 離 心 10 分鐘,去除上清液後,以 5 ml 4℃ 0.1 M CaCl2 懸浮菌體,置於冰上 1 小 時後同以上條件離心並去除上清液,加入 0.5 ml 4℃ 0.1 M CaCl2 懸浮菌體, 仍置於冰上 1 小時後,加入甘油(終濃度 30%)分裝,迅速以 -80℃ 儲存。. b. 勝任細胞的轉型 取出勝任細胞置於冰上,待其溶解後加入 0.1 ~ 1.0 µg 質體 pBR322 DNA (質體體積不得超過 10µl) ,過程中需有對照組,除不加質體外其餘操做均相同。 置於冰上 40 ~ 60 分鐘後利用 42℃水浴槽加熱處理 2 分鐘,再置於冰上 5 分 鐘後加入 0.2 ml LB 培養液,以 37℃ 恆溫培養。. 32.
(41) c. 篩選轉型細胞 由於大腸桿菌 K12 野生株並不能生長於含有抗生素的培養基,又質體 pBR322 DNA 含 有 抗 ampicillin 及 tetracyclin 之 基 因 , 因 此 以 LB-ampicillin-tetracyclin 培養基篩選 pBR322 轉型細胞,只有轉型成功的細胞 才能生長,對照組的細胞在培養基上則不會生長。. 4.2.4 大量抽取細菌質體(Sambrook et al., 2001). (1) 原理 利用 alkaline lysis 的方式將細胞打破,以 phenol-chlorofrom 將多餘的蛋白 萃取出,再以酒精將 DNA 沈澱並以水將其溶出。. (2) 溶液的製備 a. Solution I: 含 50 mM 葡萄糖、25 mM TriCl (pH 8.0)及 10 mM EDTA (pH 8.0), 高溫高壓滅菌後儲存於 4℃。 b. Solution II: 含 0.2 N NaOH(由 10 N 溶液稀釋)及 1% SDS,使用前配置此溶液。 c. Solution III: 含 5 M potassium acetate 60 ml、glacial acid 11.5 ml 及 dH2O 28.5 ml,高溫高壓滅菌後儲放於 4℃。 d. phenol:chlorofrom:isoamyl alcohol(25:24:1)混合液 將中和後的酚與 chlorofrom、isoamyl alcohol 以 25:24:1 比例混合, 避光以 4℃儲放一個月。. 33.
(42) (3) 抽取步驟 收取生長穩定期之菌體,立即加入 rifampin(終濃度 50 µg/ml)後冰浴 10 分鐘,以 6,000 rpm 離心 15 分鐘收集菌體,接著加入適量的 solution I 將菌塊 均勻打散後移至 50 ml 離心管中。加入 2 倍體積之 solution II 翻轉離心管使 液體充分混合,並置於室溫下 5 分鐘,待菌體溶解澄清後加入 1.5 倍體積之 solution III,翻轉離心管後置於冰上 10 分鐘後以 12,000 rpm 離心 20 分鐘,留 取上清液,加入等倍體積之 phenol:chloroform:isoamyl alcohol(25:24:1) 溶液,輕搖使兩相液體充分混合,以 12,000 rpm 離心 5 分鐘,助兩相完全分離 後留取上層(水相),此步驟重複兩次以去除多餘蛋白質。最後,吸取上清液加 入 2 倍體積之 95% 乙醇並置於冰上 1 小時幫助質體沈澱,以 12,000 rpm 離 心 20 分鐘後棄置上清液,加入 5 ml 70% 乙醇潤洗 pellet 後以 12,000 rpm 離 心 5 分鐘,所得 pellet 以適量含 RNase A 的無菌水溶解沈澱質體,於 37℃ 反 應 30 分鐘後置於 –20℃ 備用。. 4.2.5 DNA 超螺旋結構電泳分析. 質體 DNA 的超螺旋結構可利用含 chloroquine agarose gel 電泳加以分 析。製備 20 × 20 cm2 1% agarose gel(含有 25 µg/ml chloroquine),使用同 樣含有 25 µg/ml chloroquine 之 TBE 緩衝液作為導電液以進行電泳分析。將質 體 DNA 混合 0.2 倍體積之 loading buffer 後注入瓊脂糖凝膠 well 內,通電 50 V 使 DNA 泳動 45 小時。完成後將瓊脂糖凝膠取出置於去離子水內 1 小時 以除去 chloroquine,再以 0.5 µg/ml ethidium bromide 染色 30 分鐘。於紫外光 下觀察並照相記錄。. 34.
(43) 4.3.6 細胞內能量狀態測定(Chapman et al., 1971). (1) 原理 本實驗中以 ATP/ADP 比例作為細胞內能量的指標,可藉由偵測細胞內 ATP 與 ADP 的含量求得。 luciferase 會利用 ATP 將 luciferin 氧化產生激光反應, 當受激發的分子回復到基態時,部分能量將以光 (photon)的方式釋出。因過 程中以化學反應方式激發分子而不以熱激發,最後釋放出冷光。由於 ADP 無法 直接測得,因此以 pyruvate kinase 將 ADP 轉換成 ATP 後,再扣除 ATP 含量 求出。反應式如下:. luciferase ATP + luciferin + O2. oxyluciferin + AMP + Pi + CO2 + light. pyruvate ADP + PEP. ATP + pyruvate kinase. (3) 實驗步驟 此實驗採用 Roche 公司所生產 ATP Bioluminescence Assay Kit HSII 。首 先收取 0.5 ml 到達穩定期之菌液,立即加入等量的溶菌劑,均勻混合置於冰上。 分別取 200 µl 溶菌液加入 ATP assay reagent 及 ATP+ADP assay reagent 中(以 200 µl 溶菌劑作為空白對照組) ,利用 37℃ 水浴 15 分鐘,使酵素充分 反應,置於冰上等待測試。以冷光儀混合 luciferin、luciferase mix reagent 與待 測樣本,反應 10 秒,積分讀值 10 秒,以標準曲線判斷偵測結果。. 35.
(44) 4.3.7 突變株的構築與篩選. 1. P1 lysate 挑選單一菌落(single colony)宿主細胞至 5ml LB 培養液中。宿主細胞為 含有突變基因的菌株當作 donor cell ,即利用 P1 phage 將它所帶有的特定基 因片段攜出,再轉給另一隻 recipient cell 。將宿主細胞培養於 37℃ 恆溫震盪 培養箱 14~18 小時後,取出 50 µl 菌液至含 0.2 ﹪葡萄糖及 5 mM CaCl2 之 LB 培養液中(此時提供含 CaCl2 的新鮮培養液除了可使宿主細胞快速分裂生長 外,還可增加細胞膜的通透性,且鈣離子是 P1 phage 在吸附宿主細胞時所必需 的離子)。置於 37℃ 恆溫震盪培養箱培養 30 分鐘後,以下列比例加入 P1 vir lysate。. 試管編號. 噬菌體體積(µl). 1. 0(對照組). 2. 5. 3. 10. 4. 30. 5. 50. 加入 P1 vir lysate 後以 40 rpm 旋轉培養 2~3 小時,此時噬菌體吸附宿主 細胞之後將本身 DNA 送入宿主細胞內。 P1 phage 在宿主細胞內若處於 lysogenic state 時會以環狀質體獨立存在於宿主細胞內;而當宿主細胞 DNA 複 製完畢進行細胞分裂時,噬菌體的質體可平均分佈到各個子細胞內,而不如一般 噬菌體是將本身的 DNA 嵌入宿主細胞染色體中。 P1 phage 在宿主細胞內進行 lytic cycle 時會將宿主細胞的染色體 DNA 分成小片段,並將之裝配至蛋白質外 鞘中,此時被包入外鞘中的 DNA 可能包括噬菌體原有 DNA ,亦可能包括宿主. 36.
(45) 細胞內的 DNA 。以 P1 phage 而言,其蛋白質外鞘可包裏 91.5 Kb 大小的 DNA ,一隻 P1 phage 可容納 2.4 ﹪大腸桿菌之染色體 DNA 。因此於本實驗 中使用 P1 phage 將 relA 及 spoT 突變 DNA 片段送至大腸桿菌 K12 菌體內。 當 P1 phage 進行 lytic cycle 路徑時,比較加入 P1 phage 與對照組之宿 主細胞,培養 3~4 小時後即可明顯看出宿主細胞被水解(lysis)而呈較透明狀。 於已呈透明狀的試管每管加入 0.1 ml chloroform 震盪混合均勻後,以 5,000 離心 10 分鐘,此時被水解及未被水解的細胞因而沉澱下來。離心後小心地取含 P1 phage lysate 的上清液,將每 1 ml 上清液置於 2 ml 冷凍保存瓶中,再加 入 0.1 ml chloroform 混合均勻後即可放入 4℃ 冰箱保存。. 2. P1 transduction 將宿主細胞作為 recipient cell(K12 菌株)以接受 P1 phage 所攜帶之外來 DNA 。 recipient cell 於 37℃ 恆溫震盪培養 14~18 小時,將菌體離心 10 分 鐘後以 2.5 ml 10 mM MgSO4 (含 5 mM CaCl2)懸浮菌體。並以下列比例之 P1 phage(可能帶有 ∆relA 及 ∆spoT)加入微量離心管中:. 試管編號. 細胞體積(µl). 噬菌體體積(µl) 0(對照組). 1. 100. 2. 100. 10. 3. 100. 30. 4. 100. 50. 5. 100. 100. 6. 0. 10 (對照組). 混合均勻置於 37℃ 培養 30 分鐘後,加入 0.1 ml 1 M sodium citrate ,由 於鈣離子是 phage 吸附宿主細胞所必需,而 citrate 又可與鈣離子緊密結合,. 37.
(46) 故 sodium citrate 的加入可使 P1 phage 不再吸附宿主細胞。接著於 37℃ 以 100 rpm 懸轉培養 1 小時,使 P1 phage 之 DNA 與宿主細胞染色體進行重 組,則 phage 就可將所帶有特定基因片段送到宿主細胞染色體 DNA 上。因 ∆relA 及 ∆spoT 基因分別含有抗 kanamycin 與 chloramphenicol 的片段,故 1 小時後即可將其塗抹在含 kanamycin 與 chloramphenicol 的培養基中以篩選 ∆relA ∆spoT 雙突變株。. 4.3.8 大腸桿菌總 RNA 製備方法. 收集 5 ml 菌液(OD600 = 0.4 ~ 0.45)置於離心管內,迅速插於 -80℃酒精 桶內冰浴 10 秒鐘後迅速插於冰上,並立刻以 4℃ 6,000 rpm 離心 10 分鐘後去 除上清液,加入 0.75 ml TRI reagent 將菌塊震盪溶解,隨即將溶解之菌液置於 微量離心管內,於 65 ℃ 水浴槽內水浴 10 分鐘(每隔 2.5 分鐘以手搖方式溫 和混勻),接著於室溫下加入 200 µl chloroform,以溫和手搖方式充份混勻後靜 置於室溫下 15 分鐘,再以 4℃ 12,000 rpm 離心 15 分鐘,棄置下層液,將上清 液吸取至另一乾淨微量離心管內,加入 600 µl. 2-propanol 及 40 µl. 3 M. sodium acetate(pH 5.2),置於 -20℃ 冰箱內 30 分鐘以幫助 RNA 析出。然 後於 4℃ 12,000 rpm 離心 10 分鐘,棄置上清液後留下 RNA 沉澱物,加入 75% 乙醇 1 ml 清洗一次,再於 4℃ 12,000 rpm 離心 10 分鐘,完全吸乾上清 液後將沉澱之 RNA 進行真空乾燥 10 分鐘,待沉澱物成半溶解狀時加入適量 DEPC‧ddH2O,測定 D. 260 及. OD. 280 吸光比值以確定. 之 RNA 樣本則須立刻置於 –80 ℃ 冰箱內保存。. 38. RNA 濃度及純度。剩餘.
數據
+7
相關文件
IRB 編號 SE19403A-1 計畫主持人 陳周斌 計畫名稱 大腸直腸癌患者的生活品質與創傷後成長的關係之追蹤研究 審查意見
In this report, formats were specified for single, double, and extended precisions, and these standards are generally followed by microcomputer manufactures using
• 是細胞不正常增生,且這些增生的細胞可
基因編輯技術以人工核酸酶辨識特定 DNA 位置,並於此處切斷雙股 DNA。DNA 斷
基於 TWSE 與 OTC 公司之特性,本研究推論前者相對於後者採取更穩定之股利政 策 (Leary and Michaely, 2011; Michaely and
• Since the term structure has been upward sloping about 80% of the time, the theory would imply that investors have expected interest rates to rise 80% of the time.. • Riskless
• A put gives its holder the right to sell a number of the underlying asset for the strike price.. • How to
• A call gives its holder the right to buy a number of the underlying asset by paying a strike price.. • A put gives its holder the right to sell a number of the underlying asset
