生物學報 (2007) 42(2): 105-117
阿里山地區阿里山山椒魚的分布與族群監測
賴俊祥 呂光洋
* 國立臺灣師範大學生命科學系 (收稿日期:2007.3.12,接受日期:2007.8.6) 摘 要 從 2002 年 4 月至 2006 年 3 月於阿里山地區進行阿里山山椒魚的族群分布與族群監測的研究。 調查了阿里山鄰近地區共 36 個林班地中的山葵田,在其中 9 個林班地的山葵田有發現山椒魚,這 些地點共有的特色是坡度平緩及穩定的水源。在族群監測方面,在阿里山姐妹潭的永久樣區內共調 查到 266 隻次山椒魚,其中 94 隻次為再捕獲。利用 Jolly-Seber 法估算每月族群數量在 13-224 之 間,族群數量與前人對在此樣區的估算值相比較尚稱平穩。年齡結構方面,樣區個體的組成以成體 為主,亞成體的出現不規則,而幼體僅在春季出現。個體活動範圍的估算值在 0-509 m2之間,平 均值 64.9 ± 29.6m2 (n = 19)。利用 Cormack-Jolly-Seber 模型估算存活率在成體為 0.996,亞成體為 0.997,皆屬於高存活率。估算的捕獲率每個季節都不同,捕獲率與濕度及溫度呈正相關。再利用 Leslie matrix model 來模型化族群的動態,估計的族群增長率為 1.077,表示此族群為穩定而略呈增 長的族群。進一步分析增長率的敏感度及彈性值,都顯示成體的存活率是影響族群增長率最重要的 因子。關鍵詞:阿里山山椒魚、分布、族群監測、族群數量、Cormack-Jolly-Seber 模型、存活機率、捕獲 機率、Leslie matrix model、族群增長率
緒 言
阿里山山椒魚(Hynobius arisanensis)為兩生 類有尾目(Urodela)、小鯢科(Hynobiidae)的種類。 在台灣廣泛分布於中央山脈中南段、玉山山脈及 阿里山山脈,北起丹大山、南至北大武山;海拔 分布在 2000 m 以上,且分布區的年平均溫度在 20℃以下。一般認為牠是台灣典型的冰河孑遺物 種(glacial relict) (陳及呂,1987)。阿里山山椒魚 在習性上,一般生活在森林底層較潮溼處,又以 小溪流或天然滲水處附近較為常見。習性隱蔽, 白天躲藏在石頭或朽木下,鮮少於地表面活動, 故不易觀察(陳及呂,1986)。 阿里山地區的山椒魚生態研究最早由陳及 呂(1986)進行,他們的研究開始於 1982 年 12 月, 持續至 1985 年 6 月,當時即選定了阿里山森林 遊樂區內的山葵田進行研究。葉等(1994)除接續 研究外,亦將整個研究地點擴展到玉山國家公 園。他們的結果顯示阿里山及自忠地區的單位面 積捕獲率最高,同時阿里山的族群量及族群的年 齡結構最為穩定。由於二個樣區皆為山葵田,因 此是否山葵田有較多的山椒魚分布呢?是來自 於山葵田的農作型態?或者是農作型態外的其 他棲地因子?本研究即是對阿里山地區的山葵 田進行山椒魚分布狀況的調查,瞭解影響山椒魚 分布的可能主要因子。 再者,葉等(1994)對阿里山地區山椒魚族群 的調查距今已在十年以上,阿里山山椒魚族群的 數量經過這些年後是增加還是減少?因此我們 對現有族群量作估算,以作為未來長期監測的基 礎數據,同時也能提供管理單位有關特有物種族 群數量的數據,以利保育及經營管理。此外,藉 由長期監測的標放再捕捉研究,捕捉個體時可以 得到許多有用的個體資訊,如體長及體重的分 布、年齡結構的月變化、活動範圍等等。 事實上,族群應該關注的參數更包括了存活 機 率 (survival probability) 及 族 群 增 長 率 (population growth rate) (Williams et al., 2001)。以 往國內少有研究使用建立模型且經過統計步驟 來獲得的估算值的方法,因此我們利用本研究的 捕獲-再捕獲(capture-recapture)的數據率先嚐試 用此種方法來估算族群的重要參數。族群參數的 模型的建立,在合適的假設下,可以獲得的資訊 相 當 多 , 包 括 存 活 率 、 捕 獲 機 率 (Captureprobability)、年資機率(seniority probabilities)、徵 召 率(recruitment rate) 、 轉 移 機 率 (transition probalility)、陷阱反應(trap response)等等(Williams et al., 2001)。存活率可以利用標記個體的捕獲歷 史資訊,透過模型化及模型的選擇後,可以得知 (Williams et al., 2001)。最常用於存活機率估算的 是 Cormack-Jolly-Seber Model (CJS 模 型 ) (Lebreton et al., 1992),此模型為將標記個體的捕 獲歷史數據變為一連串存活機率及捕獲機率的 多項式乘積,然後找出每個機率參數的最大似真 估計值(maximum likelihood estimators)。此模型是 通用模型(general model),即存活機率及捕獲機率 皆為時間特定(time-specific)也就是每次調查時的 數值皆不同。而存活機率及捕獲機率有可能在時 間規模的變化是恆定的(constant)、每年或每季不 同,這些模型都只使用較少的參數故稱為減參數 模型(reduced-parameter model)。因為有許多模 型,因此就必需選擇最能和數據適配的模型,判 斷的標準是使用 Akaike's Information Criterion (AIC)值,具有最低 AIC 值者視為最能適配數據 的 模 型 。 而 模 型 間 可 更 進 一 步 以 適 合 度 檢 測 (goodness-of-fit tests) 和 似 然 比 檢 測 (likelihood ratio tests)進行統計比較。
至 於 族 群 增 長 率 可 以 使 用 Leslie matrix model(矩陣模型)(Caswell, 2001)。這個模型首先 必需具備研究族群各年齡或各生活史時期(life history stage)的重要比率(Vital rate)--存活率及生 殖力(Fertility)的數據,數據來源可以實際調查或 估 算 得 來 的 。 這 些 數 據 放 入 族 群 投 影 矩 陣 (Population projection matrix, A),則可以用某時期 的族群內各年齡層的數目(n(t)),來推測下一時期 各年齡層的數目:即 n(t+1) = A n(t)。矩陣 A 的 最大特徵值(largest eignevalue)即是固定的族群增 長率(deterministic population growth rate, λ);右特 徵向量(right eigenvector)為穩定年齡分布(Stable age distribution)、左特徵向量(left eigenvector)為 繁殖價值(Reproductive value)。而族群增長率對 重 要 比 率(θ) 或 矩 陣 輸 入 值 (aij) 變 異 的 敏 感 度 (sensitivity)則是對二者的偏微分值分別為(∂λ/∂θ) 及(∂λ/∂aij),以及敏感度的彈性值(elasticities)則是 對數刻度的敏感度值(Caswell, 2001)。因此我們利 用前項估算得到的存活機率及相關文獻資料,建 構族群投影矩陣將族群的動態模型化,以評估族 群未來的趨勢。
材料與方法
阿里山山椒魚在阿里山地區山葵田的分布情況 調查 調查範圍 調 查 範 圍 依 據 農 委 會 林 務 局 的 林 班 地 分 區,計包括了阿里山事業區第1 至 11、16 至 20 林班以及大埔事業區第190、191、195、196、197、 206 至 220 林班等共 36 個林班,海拔範圍由 1714 至2500 m(表一)。 調查方式 由於山椒魚的表層活動在雨季(三至八月)時 較為明顯,因此在雨季時由林務局人員的帶領至 各林班地調查,調查時間為2002 年至 2004 年。 在每個林班地中,隨機選取山葵田,紀錄進行調 查山葵田的面積(概估)、坡向、海拔高度,並依 據農民或現場工作人員的資訊記錄主要水源狀 況(穩定、季節性乾涸、無水源)等。接著尋找山 葵耕作範圍及附近地區中任何適合山椒魚棲息 的棲地,包括山葵田、小溪流、石壁滲水處或潮 溼處等等,在這些地點內徒手翻找山椒魚慣於躲 藏的石塊及枯木。計數各林班地中發現到的山椒 魚數量。 阿里山地區阿里山山椒魚族群動態 研究樣區 永久樣區位置在姐妹潭北方的山谷內(地理 位置圖見葉等(1994)),屬阿里山事業區 2 林班。 整個樣區中間被溪流及樹木分隔為上下兩個不 連續的部分,因此將樣區分為A、B 兩區,兩區 間相距約20 m。 A 區大致與陳及呂(1986)及葉等(1994) A 樣 區位置相當,但田畦形狀及耕作狀態有差異。A 區總面積約為3000 m2,長約100 m,寬約 30 m, 為 柳 杉 (Cryptomeria japonica) 和 紅 檜 (Chamaecyparis formosensis)混合林地,由東南向 西北方傾斜約15 度,樣區內有三條主要小水道, 一在樣區的東南邊,二條在樣區的西北邊,二者 皆由南向西流。地表為當地居民依山勢闢為梯 田,種植山葵(Eutrema japonica),田畦四周以石 塊或木板或枕木堆砌而成,共計 61 畦山葵田, 其中有52 畦在種植,9 畦為休耕地。但 2003 年 起全數休耕,地表的其他植物以草本植物、苔蘚 植物和蕨類等蔭性植物為主。 B 區約在 A 區北方 20 m 處,概為西北向東 南的長方形,長約80 m、寬約為 40 m,總面積阿里山地區阿里山山椒魚的分布與族群監測
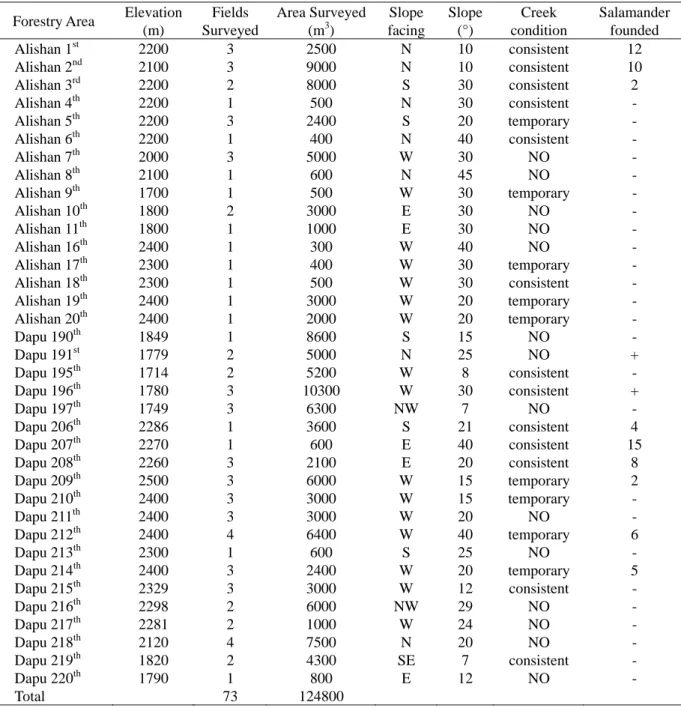
表一、阿里山地區林班地調查的山葵田相關資料與尋獲阿里山山椒魚數量表。
Table 1. Characteristics of wasabia field surveyed at each forestry area in Alishan, and number of salamander found.
Forestry Area Elevation (m) Fields Surveyed Area Surveyed (m3) Slope facing Slope (°) Creek condition Salamander founded Alishan 1st 2200 3 2500 N 10 consistent 12 Alishan 2nd 2100 3 9000 N 10 consistent 10 Alishan 3rd 2200 2 8000 S 30 consistent 2 Alishan 4th 2200 1 500 N 30 consistent - Alishan 5th 2200 3 2400 S 20 temporary - Alishan 6th 2200 1 400 N 40 consistent - Alishan 7th 2000 3 5000 W 30 NO - Alishan 8th 2100 1 600 N 45 NO - Alishan 9th 1700 1 500 W 30 temporary - Alishan 10th 1800 2 3000 E 30 NO - Alishan 11th 1800 1 1000 E 30 NO - Alishan 16th 2400 1 300 W 40 NO - Alishan 17th 2300 1 400 W 30 temporary - Alishan 18th 2300 1 500 W 30 consistent - Alishan 19th 2400 1 3000 W 20 temporary - Alishan 20th 2400 1 2000 W 20 temporary - Dapu 190th 1849 1 8600 S 15 NO - Dapu 191st 1779 2 5000 N 25 NO + Dapu 195th 1714 2 5200 W 8 consistent - Dapu 196th 1780 3 10300 W 30 consistent + Dapu 197th 1749 3 6300 NW 7 NO - Dapu 206th 2286 1 3600 S 21 consistent 4 Dapu 207th 2270 1 600 E 40 consistent 15 Dapu 208th 2260 3 2100 E 20 consistent 8 Dapu 209th 2500 3 6000 W 15 temporary 2 Dapu 210th 2400 3 3000 W 15 temporary - Dapu 211th 2400 3 3000 W 20 NO - Dapu 212th 2400 4 6400 W 40 temporary 6 Dapu 213th 2300 1 600 S 25 NO - Dapu 214th 2400 3 2400 W 20 temporary 5 Dapu 215th 2329 3 3000 W 12 consistent - Dapu 216th 2298 2 6000 NW 29 NO - Dapu 217th 2281 2 1000 W 24 NO - Dapu 218th 2120 4 7500 N 20 NO - Dapu 219th 1820 2 4300 SE 7 consistent - Dapu 220th 1790 1 800 E 12 NO - Total 73 124800 +: Farmer’s information 約 3200 m2,由東西兩側向來自 A 區的溪流凹 陷,兩側坡度約 20 度。B 區的位置約當陳及呂 (1986)的 B 樣區,但面積較大。本區共有 24 畦山 葵田,亦為柳杉及紅檜的造林地,但種植較為稀 疏,樣區中央一部分整日陽光直射,其底層植被 狀況似 A 區,但以樣區四週樹木的下層較多。 調查方式 從 2002 年 4 月至 2006 年 3 月約每月一次進 行 調 查 , 實 驗 設 計 上 使 用 固 定 面 積 調 查 法 (Area-constraint survey method),即以徒手翻找樣 區內所有可能被山椒魚利用為遮蔽物的土表物 體,直到將所有的遮蔽物皆翻找過一次為止。尋 獲的個體紀錄日期、時間及位置後,接著以 0.1% benzocaine 麻醉,再測量個體的吻肛長、尾長、 頭長、體重等,若可分辨出性別亦記錄下來。依 Martof (1953)的去趾標記系統進行編號,由於認 為剪趾會影響到兩棲類的存活率,從 2005 年 11
開 始 使 用 軟 性 螢 光 標 誌 (Visible Implant Fluorescent Elastomers (VIE tags), Northwest Marine Technologies, Inc.),以紅、橙、黃、綠四 種顏色的螢光標誌注射於山椒魚四肢及尾部的 腹面皮下,藉以辨視個體(未再進行剪趾)。操作 完成後的個體,放入清水中,待其甦醒後,放回 原來的遮蔽物下,並儘量回復原有棲地形態。 族群組成與年齡結構 由 於 阿 里 山 山 椒 魚 沒 有 明 顯 的 雌 雄 二 型 性,並且能間接鑑定性別的個體太少,所有後續 分析都將二個性別的資料合併分析。我們分析了 捕獲個體體長及體重的分布、體長及體重的相關 性分析及年齡結構。年齡類別依據葉等(1994)由 吻肛長(SVL)歸類,其成體之 SVL>45 mm、亞成 體為25 mm < SVL < 45 mm、幼體則是 SVL<25 mm。 族群量估計 由於在長期的研究中,族群會有出生、死 亡、遷出及遷入,這樣的族群是個開放族群(Open population),因此使用 Jolly-Seber 法 (Krebs, 1999) 來估算每月的族群數量。由於山椒魚每年僅在春 季進行生殖,在此之後族群不再大量增加,若將 出生之外的數值視為可以忽略,就可以使用封閉 族群(Closed population)的方法,因此使用具多次 標放的Schnabel 法(Krebs, 1999)來估算每年的族 群量,本研究中對季節的界定是春天為 2、3、4 三個月,依此類推,三個月為一季,而年的界定 是每年的二月至翌年一月。 活動範圍 依據個體被紀錄的出現點,以最小凸多邊形 法 (minimum convex polygon method, MCP)(Stickel, 1950)及適應核心法(adaptive kernel, ADK)(Worton, 1989)計算個體的活動範圍(home range),後者能更進一步瞭解個體活動中心的大 小。活動範圍的計算使用 Calhome 軟體(U.S. Forest Service, Pacific Southwest Research Station)。
存活分析
存活機率及捕獲機率模型化依據 Cormack- Jolly-Seber model (Lebreton et al., 1992)。我們將 通用模型與幾個減參數模型進行比較,具有最低 AIC 值者視為最能適配數據的模型(Williams et al., 2001)。依據本調查的特性在存活率(Φ)的備選 模型(alternative models)上會有季節效應、年效應 及恆定,同時再依年齡分為成體及亞成體二組。 發現率(p)則依據山椒魚的特性,我們僅選擇有季 節效應及年齡分組的備選模型。調查中雖然發現 幼體,但由於此估算法是以標記動物為材料,而 考慮到幼體進行剪趾標記可能會產生很大的傷 害,故未進行標記,因此幼體的資料未納入此分 析中。模型的比較與選擇使用軟體MARK (White & Burnham, 1999)。
族 群 動 態 模 型 化 (Modeling the population
dynamics)
我們使用Leslie matrix model 來建構以生活 史時期為結構的族群模型。將阿里山山椒魚的生 活史分為四個時期:(1)卵及水生幼體;(2)陸生幼 體;(3)亞成體;(4)成體。在矩陣中輸入各生活史 時期的重要比率--存活率及生殖力。第 1 期至第 2 期的存活率依據 Lue & Chuang (1992)的估計 值。亞成體及成體的存活率來自於存活分析的估 算值。生殖力的估算值來自 Kakegawa et al. (1989)。圖一中為此模型的生活史圖及族群投影 矩陣。從矩陣分析求得固定族群增長率、穩定年 齡分布、繁殖價值、敏感度(sensitivity)及彈性值 (elasticities)。接著再以每年捕獲的各生活史時期 個體的平均數,投影(projection)10 年中各生活史 時期個體數的變動。
結 果
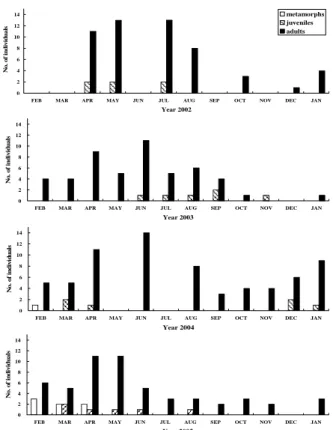
阿里山山椒魚在阿里山地區山葵田的分布情況 調查 研究期間共調查了36 個林班地,共計 73 處 山葵田總面積達 124800 m2。各林班地進行調查 山葵田的數量、概略面積、坡向、坡度、水源狀 況、調查到山椒魚數量見表一。有山椒魚分布的 林班地有9 個,其中 1 個並非在山葵田中發現, 而是在步道的兩側。發現山椒魚的9 個林班地海 拔分布在2100 至 2500 m,各種坡向都有,坡度 的範圍10 至 40 度,6 個具有穩定水源,3 個的 水源會有乾涸的現象。調查到的山椒魚數量從 2 隻到15 隻。 阿里山地區阿里山山椒魚族群動態 研究期間在姐妹潭樣區內共調查到266 隻次 山椒魚。每次捕獲量,因月別而異(表二),2002 年5、7 兩月 15 隻最高,2003 年 12 月、2005 年 12 月 0 隻最低。在個體被捕捉次數方面,133 隻阿里山地區阿里山山椒魚的分布與族群監測 A 0 0 0 F41 P12 0 0 0 nt+1 = 0 P23 0 0 nt 0 0 P33 0 0 0 P34 P44 圖一、上圖:阿里山地區阿里山山椒魚族群的生活史 時期圖及各時期重要比率的數值。下圖:族群投影矩 陣 A 乘上描述族群在 t 時狀態的向量 n(t),得到 t+1 時的狀態。
Figure 1. Upper figure: Life cycle graph and stage-based
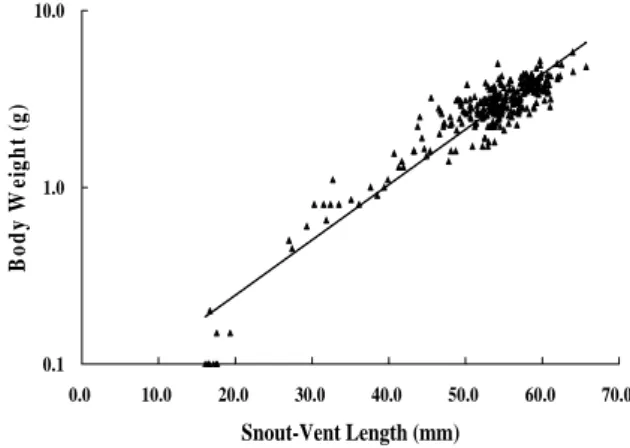
vital rates for the Alishan salamander populations at Alishan. Lower figure: Population projection matrix A is multiplied to n(t), a vector describing the stat of the population at time t, to obtain the state of the population at t+1. 個體僅被捕獲一次,48 隻個體有再捕獲的記錄, 而單一個體被捕獲次數最高為 10 次,平均每隻 捕獲 1.54 次。個體捕獲間隔時間由 13 天(半個月) 至 1193 天(39 個月)不等,平均捕獲間隔日數為 197.5 日。以捕獲間隔日數的分布而言,最多的 個體在 100 天(3 個月)以內再次被捕獲,15 隻個 體再捕獲的間隔達一年以上。個體的持續捕獲時 間(最初及最後捕獲的時間差),最高為 1307 天, 最低為 14 天,在再捕獲的個體記錄中,持續捕 獲以 1 年內者佔大多數,但將近三分之一已存活 天數高於 1 年,最長者已將近 4 年。 族群組成與年齡結構 捕獲個體的各形值之均值及標準差在吻肛 長為 52.1 ± 9.4 ㎜、尾長 36.4 ± 8.0 ㎜、頭長為 12.9 ± 2.1 ㎜、頭寬為 8.0 ± 1.3 ㎜。不同月份及 年度的吻肛長分布如圖二,樣區內捕獲的個體主 要為成體,幾乎每個調查的月份都可捕獲。亞成 體在調查中偶有發現,但是在出現的月份上沒有 規律。而幼體在每年的 2、3、4 月發現。 捕獲個體的體重範圍從 0.1-5.8 g,平均體重 為 2.9 ± 1.1 g。體重的分布略呈常態分布,以 2.5-3.0 g 的個體最多(62 隻),剛變態完成的幼體 體重為 0.1 g,大型的個體體重可達 5.0 g 以上。 山椒魚的體重隨著吻肛長的增加而呈指數增加(y = 0.0581e0.072x, R2 = 0.8847)(圖三)。 0 2 4 6 8 10 12 14
FEB MAR APR MAY JUN JUL AUG SEP OCT NOV DEC JAN
Year 2002 N o . o f i n di vi dua ls metamorphs juveniles adults 0 2 4 6 8 10 12 14
FEB MAR APR MAY JUN JUL AUG SEP OCT NOV DEC JAN
Year 2003 N o . of i ndi v idual s 0 2 4 6 8 10 12 14
FEB MAR APR MAY JUN JUL AUG SEP OCT NOV DEC JAN
Year 2004 N o . of i n d ivi d u al s 0 2 4 6 8 10 12 14
FEB MAR APR MAY JUN JUL AUG SEP OCT NOV DEC JAN
Year 2005 N o . of i n d ivi d u a ls 圖二、各年度各月份捕獲山椒魚的年齡結構圖。年齡 以吻肛長(SVL)來決定:(1)幼體是 SVL 小於 25 mm; (2)亞成體的 SVL 介 25 至 45 mm 之間;(3)成體的 SVL 大於 45 mm (葉等,1994)。
Figure 2. Age structure of salamander captured in each
month. Age group determined by snout-vent length (SVL): (1) metamorphs, SVL smaller than 25 mm; (2) juveniles, SVL between 25 to 45 mm; (3) adults, SVL larger than 45 mm (Yeh et al. 1994).
Aquatic
Larvae
Metamorphs
Juveniles
Adults
P12 = 0.3 P23 = 0.3 P34 = 0.45
F41 = 2.4
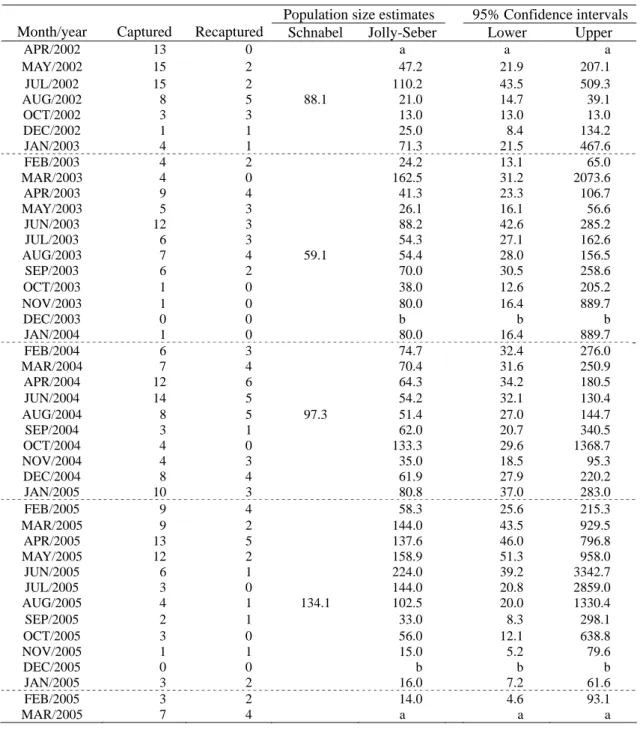
表二、姐妹潭樣區各月份捕捉數及再捕獲數、Schnabel 年族群估算量、Jolly-Seber 月族群估算量與 95%信賴區 間。
Table 2. Number of individuals captured and recaptured of each month at Alishan study site. Population size estimated
including Schanbel estimates (yearly), and Jolly-Seber estimates (monthly) with 95% confidence intervals. Population size estimates 95% Confidence intervals Month/year Captured Recaptured Schnabel Jolly-Seber Lower Upper
APR/2002 13 0 a a a MAY/2002 15 2 47.2 21.9 207.1 JUL/2002 15 2 110.2 43.5 509.3 AUG/2002 8 5 88.1 21.0 14.7 39.1 OCT/2002 3 3 13.0 13.0 13.0 DEC/2002 1 1 25.0 8.4 134.2 JAN/2003 4 1 71.3 21.5 467.6 FEB/2003 4 2 24.2 13.1 65.0 MAR/2003 4 0 162.5 31.2 2073.6 APR/2003 9 4 41.3 23.3 106.7 MAY/2003 5 3 26.1 16.1 56.6 JUN/2003 12 3 88.2 42.6 285.2 JUL/2003 6 3 54.3 27.1 162.6 AUG/2003 7 4 59.1 54.4 28.0 156.5 SEP/2003 6 2 70.0 30.5 258.6 OCT/2003 1 0 38.0 12.6 205.2 NOV/2003 1 0 80.0 16.4 889.7 DEC/2003 0 0 b b b JAN/2004 1 0 80.0 16.4 889.7 FEB/2004 6 3 74.7 32.4 276.0 MAR/2004 7 4 70.4 31.6 250.9 APR/2004 12 6 64.3 34.2 180.5 JUN/2004 14 5 54.2 32.1 130.4 AUG/2004 8 5 97.3 51.4 27.0 144.7 SEP/2004 3 1 62.0 20.7 340.5 OCT/2004 4 0 133.3 29.6 1368.7 NOV/2004 4 3 35.0 18.5 95.3 DEC/2004 8 4 61.9 27.9 220.2 JAN/2005 10 3 80.8 37.0 283.0 FEB/2005 9 4 58.3 25.6 215.3 MAR/2005 9 2 144.0 43.5 929.5 APR/2005 13 5 137.6 46.0 796.8 MAY/2005 12 2 158.9 51.3 958.0 JUN/2005 6 1 224.0 39.2 3342.7 JUL/2005 3 0 144.0 20.8 2859.0 AUG/2005 4 1 134.1 102.5 20.0 1330.4 SEP/2005 2 1 33.0 8.3 298.1 OCT/2005 3 0 56.0 12.1 638.8 NOV/2005 1 1 15.0 5.2 79.6 DEC/2005 0 0 b b b JAN/2005 3 2 16.0 7.2 61.6 FEB/2005 3 2 14.0 4.6 93.1 MAR/2005 7 4 a a a
a: The Jolly Seber method doesn't compute this value. b: Not estimate. 族群數量 利 用 Jolly-Seber 法 估 算 的 月族 群量 於表 二,表中亦包括了族群量估值的 95%信賴區間。 估計的族群量最高為 2005 年 6 月,族群數量為 224 (39.2-3342.7)隻,最低為 2002 年 10 月,數量 為 13 (13-13)隻,族群數量在 2002 年至 2003 上 半年呈劇烈變動形式,然後持續平穩約一年左 右,至 2004 下半年略呈下降的趨勢。2005 年初 開始增加,但是 6 月開始又開始減少。 以 Schnabel 法估算的年族群量(表二),2002 年 88.1 隻、2003 年 59.1 隻、2004 年 97.3 隻、2005 年 134.1 隻,除 2003 年數值較低外,其餘年度數
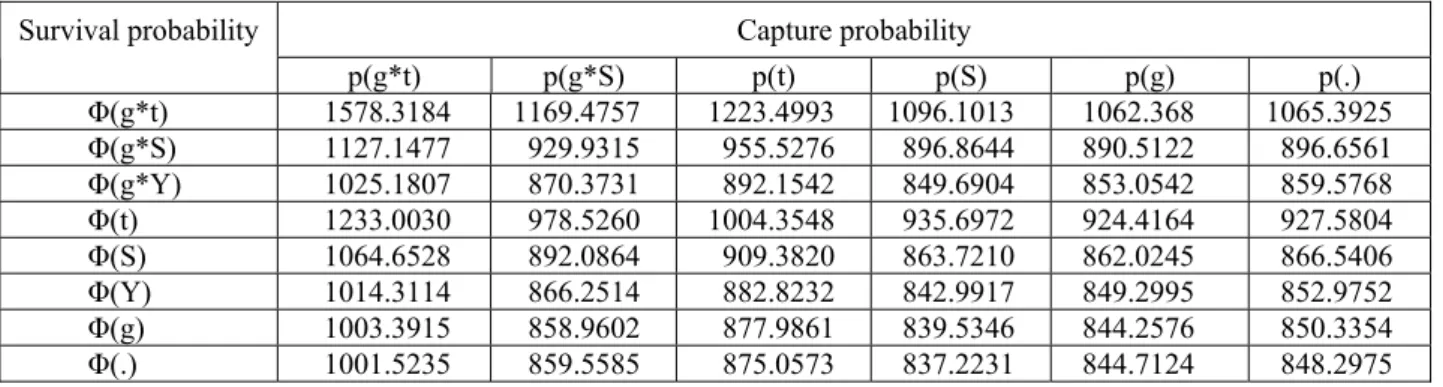
阿里山地區阿里山山椒魚的分布與族群監測 值都在100 隻上下。 水平分布與活動範圍 將A、B 二樣區內捕獲 3 次以上的個體共計 19 隻,依其出現點以最小凸多邊形法(MCP)及適 應核心法(ADK)估算其活動範圍。A 樣區個體的 活動範圍圖如圖四,B 樣區個體如圖五。估算的 個體活動範圍,在MCP 法在 0 至 509.4 m2之間, 平均值64.9 ± 29.6m2 (n = 19)。在 ADK 法中,近 活動中心的50%的出現點面積在 0 至 417.5 m2之 間,平均值58.6 ± 27.8 m2。用以計算的位置點數 最多出現在5 位點(捕獲 9 次),而有 2 隻個體的 捕獲皆在同一點(圖四)。而個體的位點間的平均 距離在0.0 至 52.7 m 之間,個體間平均值為 14.2 m。調查中,曾有三次在同一遮蔽物下同時發現 兩隻個體,而僅有一遮蔽物先後發現不同的個 體。 能計算活動範圍的個體皆為成體,吻肛長 (個體最大測量值)從 45.4 至 64.0 mm,平均值為 56.3 mm,而捕捉次數在三次以下的個體(不含幼 體)的吻肛長平均值為 52.3 mm,二者間呈顯著差 異(F1,166=5.81, p<0.05)。 A 樣區內個體的活動範圍在邊緣的部分有 重疊(圖四),然而由於個體出現的年度不同,將 不同年度以不同顏色標示後發現,個體的活動範 圍基本上是不重疊的,若是重疊也僅在邊緣的部 分。B 樣區的 5 隻個體,也呈現相同的情形(圖五)。 存活率分析 存活率及捕獲率模型比較的結果如表三,具 有最低AIC 值的最適配數據的模型為「存活率恆 定、捕獲率季變化」{Φ(.) p(S)},次佳模型「存 活率組變化、捕獲率季變化」{Φ(g) p(S)}。這表 示存活率在調查期間是恆定的,或者二個不同年 齡組的存活率有差異也可接受。在{Φ(.) p(S)}模 型中存活率的估計值0.9976,在{Φ(g) p(S)}模型 中亞成體存活率估計值為0.9975,而成體的存活 率估計值為0.9976。捕獲率在前三個適配數據的 模型皆呈現明顯的季節效應(表三),因此捕獲率 會隨著季節而變動。將捕獲率分別對每季的氣 溫、雨量及溼度作相關分析,捕獲率和季累積雨 量無相關(r = 0.40, p<0.152),與季平均溫度(r = 0.79, p = 0.002)及溼度(r = 0.61, p = 0.034)呈正相 關。
族 群 動 態 模 型 化 (Modeling the population
dynamics) 0.1 1.0 10.0 0.0 10.0 20.0 30.0 40.0 50.0 60.0 70.0 Snout-Vent Length (mm) Bo d y W ei g h t ( g ) 圖三、姐妹潭樣區調查捕獲山椒魚體重—吻肛長關係 圖,實線為指數迴歸線y = 0.0581e0.072x, R2 = 0.8847。
Figure 3. Relationships between body weight and
snout-vent length of captured salamanders. Exponential regression line indicated by solid line, y = 0.0581e0.072x,
R2 = 0.8847.
使用Leslie matrix 來建構以生活史時期為結 構的族群模型。在矩陣中輸入各生活史時期的重 要比率--存活率及生殖力(圖一)。第 1 期至第 2 期的存活率(P12)依據 Lue & Chuang (1992)的估計
值 0.35-0.48,取其低值 0.3。第 2 期至第 3 期的 存活率(P23)因為沒有野外資料,假設值為 0.4。亞 成 體 存 活 率 在 存 活 分 析 中 得 到 的 估 算 值 為 0.997,在本分析中取為 0.9,由於標放記錄顯示 亞成體長大為成體的時間在一年以上,故假設每 年會有一半仍停留在亞成體,一半則成長為成 體,因此兩個部分的比率值皆為0.45 (P33及P34)。 成體的存活率同樣輸入值為0.9。生殖力(F41)的估 算值來源如下:Kakegawa et al. (1989)估計台灣山 椒魚的平均窩卵數(Clutch size)為 16;存活率為 0.9;假設族群中性別比為 1:1,故繁殖機率為 0.5,而保守推測每年僅有 1/3 雌性個體進行繁 殖,四個數值相乘得生殖力值為2.4(圖一)。
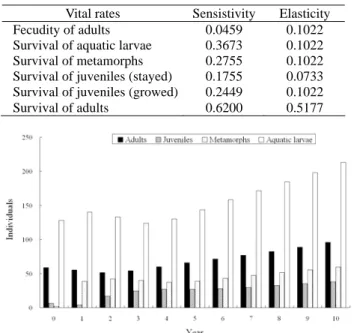
利用Leslie matrix model 估計得到的族群增 長率(λ)為 1.077,表示此族群為穩定而略呈增長 的族群。增長率敏感度值列於表四,對族群增長 率敏感度最高者為成體的存活率(0.6200),其次為 水生幼體存活率(0.3673)。敏感度值的彈性值仍以 成體存活率最高(0.5177),而停留的亞成體存活率 最低為0.0733(表四)。 族群矩陣投影後,得到未來每一年的每個時 期的個體數量(圖六),族群的年齡分布相當穩 定,水生幼體佔最大部分,而在陸地生活時期以 成體佔最主要。每年的個體總量持續緩慢的增 加,總量介於200 至 350 之間。族群達穩定年齡
組成時(矩陣的右特徵向量),水生幼體佔 52.5%、 陸生幼體佔 14.6%、亞成體佔 9.3%及成體佔 23.6%。估計的世代長度(generation time)11.45 圖四、姐妹潭 A 樣區捕獲三次以上個體的活動範圍 圖。每個多邊形分代表不同個體的活動範圍,圓點則 表示此個體一直停留在同一點,顏色代表不同的年度 (紅:2002 年;藍:2003 年;紫:2004 年;棕:2005 年),數字為個體編號。
Figure 4. Home range of individuals captured more than
three times in plot A of Sisters pond study site. Each polygon represents home range of different individuals. Dots are individuals not moved. Color indicate capture year: red-2002; blue-2003; purple-2004; brown-2005. Identification number showed beside each polygon.
年,平均年齡為13.81 年。繁殖價值(矩陣的左特 徵向量)在水生幼體為 3.6%、陸生幼體佔 12.9%、 亞成體佔34.8%及成體佔 48.6%。 圖五、姐妹潭 B 樣區捕獲三次以上個體的活動範圍 圖。每個多邊形分代表不同個體的活動範圍,顏色代 表不同的年度(紅:2002 年;藍:2003 年;紫:2004 年;棕:2005 年),數字為個體編號。
Figure 5. Home range of individuals captured more than
three times in plot B of Sisters pond study site. Each polygon represents home range of different individuals. Dots are individuals not moved. Color indicate capture year: red-2002; blue-2003; purple-2004; brown-2005. Identification number showed beside each polygon.
表三、姐妹潭樣區族群存活率(Φ)及捕獲率(p)的模型選擇及 AIC 值,包括了時間效應(t)、季節效應(S)、年效應 (Y)、組效應(g)及恆定值(.)。
Table 3. Model selection and values of Akaike’s information criterion (AIC) for the survival (Φ) analyses of Sister pond
population, given various capture probalities (p), including time effect (t), season effect (S), year effect (Y), group effect (g) and constant (.).
Survival probability Capture probability
p(g*t) p(g*S) p(t) p(S) p(g) p(.) Φ(g*t) 1578.3184 1169.4757 1223.4993 1096.1013 1062.368 1065.3925 Φ(g*S) 1127.1477 929.9315 955.5276 896.8644 890.5122 896.6561 Φ(g*Y) 1025.1807 870.3731 892.1542 849.6904 853.0542 859.5768 Φ(t) 1233.0030 978.5260 1004.3548 935.6972 924.4164 927.5804 Φ(S) 1064.6528 892.0864 909.3820 863.7210 862.0245 866.5406 Φ(Y) 1014.3114 866.2514 882.8232 842.9917 849.2995 852.9752 Φ(g) 1003.3915 858.9602 877.9861 839.5346 844.2576 850.3354 Φ(.) 1001.5235 859.5585 875.0573 837.2231 844.7124 848.2975
阿里山地區阿里山山椒魚的分布與族群監測
表四、姐妹潭樣區族群的族群增長率的敏感度值與敏 感度的彈性值。
Table 4. Sensitivity and elasticity of population growth
rate of each vital rate of Sister pond population. Vital rates Sensistivity Elasticity Fecudity of adults 0.0459 0.1022 Survival of aquatic larvae 0.3673 0.1022 Survival of metamorphs 0.2755 0.1022 Survival of juveniles (stayed) 0.1755 0.0733 Survival of juveniles (growed) 0.2449 0.1022 Survival of adults 0.6200 0.5177
圖六、族群矩陣投影的結果,顯示未來 10 年各生活史 時期個體數的動態變化。
Figure 6. Result of 10-year population size projection.
討 論
阿里山山椒魚在阿里山地區山葵田的分布情況 調查 經過山葵田地山椒魚的調查,發現的山椒魚 事實上非常的少。僅在阿里山 1, 2, 3 林班及大埔 206、207、208、209、212 林班調查到山椒魚(表 一)。經過分析發現,有山椒魚分布的山葵田共同 特色為坡度平緩且有穩定水源。而在陡峭的林班 地中,在步道兩側平坦處也可以發現。山葵田一 般以梯田的方式種植,事實上在每一塊田坡度都 在 5 度以內,故坡度的影響較為有限,因此我們 認為穩定水源的有無是較為重要的因素。另外, 調查的過程中我們還注意到農民圍田畦方式也 會有影響,許多地點的山葵田,圍田是以多層的 石頭堆疊,有些以大型枯木堆疊,這都會影響到 調查者發現山椒魚。依據 Lai & Lue (2008)有關阿 里山山椒魚棲地利用的特性,他們偏好使用木塊 作為遮蔽物且有苔類及高等植物附著者。山椒魚 在棲息地的基質方面,偏好單純的泥土基質。綜 合以上結果,山椒魚的分布主要是受到坡度及水 源特色的影響,並非是山葵田的存在。但是當坡 度及水源特色的適合,山葵田的耕作方式—堆疊 枯木、石塊及除草等等,會創造山椒魚喜歡利用 的微棲地,這也就是在山葵田的樣區中會有較多 山椒魚棲息的主因。 阿里山地區阿里山山椒魚族群動態 比較各年度捕獲個體的年齡分布,每個月皆 以成體的數量最多(圖二),說明此地是山椒魚成 體主要的棲息地點。亞成體出現的時間沒有規則 性。山椒魚在冬季進行生殖後,從春季開始,就 會有各種體型的個體出現在樣區內,至春末許多 體型較小的個體就可能遷移到其他地點,僅體型 較大者停留在樣區內,一直到下個生殖季才又開 始新的循環。就 2003 年度而言,由於該年雨量 少使得山椒魚生殖現象不明顯,未發現任何的幼 體,直到隔年才又恢復出現。 從持續捕獲的時間差異,可以概略推估山椒 魚的存活時間,在樣區中山椒魚最長的持續捕獲 時間將近 3 年,這表示山椒魚在野外的壽命至少 是 3 年的時間。3 隻存活接近 3 年者,有 2 隻最 初捕獲時吻肛長已達 60 mm,而三年來吻肛長增 加僅 1~2 mm,所以可說是已達最大吻肛長,另 一隻吻肛長則增加了近 5 mm,亦達到 60 mm 左 右。從這些有限的數據推測,山椒魚的壽命應該 遠高於 3 年,若從 50 mm 成長到 60 mm 需 3 年, 至少就有 6 年的壽命。在本研究中,捕獲的剛變 態幼體吻肛長僅 18 mm 左右,而至下一生殖季前 捕獲的幼體吻肛長為 28 mm (僅 1 隻),故僅成長 了 10 mm,依此速度,幼體從 17 mm 成長至 50 mm 的時間估計也需 3 年,山椒魚應該能存活 9 年, 而這個數值受到骨骼鑑齡法(Skeletochronology) 支持(賴及呂, 未發表資料)。 陳及呂(1986)使用 Weight mean 法估計此地 的族群量在 1 至 105 之間(A 樣區及 B 樣區合計), 這與我們依據 Jolly-Seber 法所估算的族群量接 近,表示族群的穩定。然而葉等(1994)使用相對 指標估算阿里山的族群密度值為 3 隻/月.1000m3 左右,以同樣方法求算這四年的族群密度值為 1 隻/月.1000m3,變成原來數值的三分之一。我們 推測是發現率減低的緣故。山葵田在 2003 年開 始休耕後,在缺少人為清除雜草下,草本植物的 生長幾已包住了大部分的遮蔽物,植物的根部伸 入,造成了基質特性明顯地漸不適合山椒魚棲息 (Lai & Lue, 2008)。雖然基質特性不適合山椒魚棲 息,但估計的存活率卻仍保持恆定且達 0.99,山 椒魚應當是遷出這個地區而非死亡。發現率減低 就造成相對指標的差異。再者 Jolly-Seber 法的信賴區間很大,也應歸因於發現率的減低使得估計 的樣本數變得很小,相對的信賴區間就變大了。 然而不論山椒魚是減少或移出,都必需進行提高 山椒魚棲息量的工作,可行的一個方法是除去遮 蔽物旁的雜草,另一個方法可以增加遮蔽物或粗 質殘材(coarse woody debris) (McKenny et al., 2006)來增加山椒魚的族群量,然而方法的合適性 仍有待後續的研究。不同的族群量估算法卻呈現 不同的趨勢,使用Schanbel 法估算的族群量呈現 先下降後上昇的趨勢,這與族群動態模擬所得到 的結果類似。 在本研究中,MCP 估計的 64.9 m2與 ADK 估計的58.6 m2遠高於葉等(1994)用 MCP 法 4.51 ± 3.64 m2。由於本研究中有些個體呈長距離的移 動,造成了平均值的升高。但這種長距離的移動 究竟是生殖遷移或者是日常的活動範圍我們無 法得知,故皆視為日常活動範圍。大部分陸棲有 尾 兩 生 類 日 常 的 移 動 皆 為 短 距 離 , 例 如 攀 螈 (Aneides vagrans)範圍在 2.0 - 20.3 m (Davis, 2002)、奧林螈(Rhyacotriton cascade)的平均值為 2.4 m (Nijhuis & Kaplan, 1998)。阿里山山椒魚的 平均移動距離14.2 m 介於二者之間。 大部分個體的50%活動範圍都小於 10 m2且 活動範圍不重疊的現象都指出山椒魚成體可能 會建立領域,而領域行為在陸棲有尾類是常見 的。已知具有領域行為的有尾類包括無肺螈科 (Plethodontidae) 的 攀 螈 屬 (Aneides) 、 脊 口 螈 屬 (Desmognathus)、 劍 螈屬 (Ensatina)及 無 肺 螈屬 (Plethodon)(Cupp, 1980; Keen and Reed, 1985; Jaeger et al., 1982; Wiltenmuth, 1966),以及鈍口螈 科 (Ambystomatidae) 的 鈍 口 螈 (Ambystoma) (Smyers et al., 2002)。陸生的蠑螈領域的形成可能 導因於個體在以下因子間的競爭,包括食物或覓 食區域、提供涼爽及潮溼微棲地的遮蔽物或地底 棲 所 、 地 下 產 卵 地 、 獲 得 配 偶 等 等(Marvin, 1998)。由於具有固定的活動範圍是體型較大的成 體,在Marvin (1998)對無肺螈的研究中,顯示領 域的保持者通常是體型較大的成體,本研究也發 現能測量活動範圍的個體體型顯著較捕獲個體 均值為高,所以這種現象也可能是山椒魚具有領 域的間接證據。至於亞成體及幼體在樣區內通常 僅發現一次,其狀況就無法得知了。Lai & Lue (2008)進行棲地偏好性分析的結果,成體在微棲 地偏好現象最為明顯,亞成體次之,幼體則幾乎 沒有偏好性。我們推測成體的明顯偏好是在找到 合適的棲地後,建立領域以利攝食,而趨趕體型 較小的個體,使這些個體僅能選擇品質較差的棲 地。 存活分析 在存活分析中,山椒魚的成體及亞成體的存 活率皆在 0.99 以上,這遠較一些動物的估算值 0.8~0.9 高(例如帝王企鵝(Aptenodytes forsteri)為 0.94~0.95、雪海燕(Pagodroma nivea)為 0.79-0.82 (Jenouvrier et al., 2005a,b)、箱龜(Terrapene)為 0.97(Bowen et al., 2004))。高存活率的原因推測是 此地區的環境穩定。這個樣區我們從 1982 年至 今斷斷續續的進行調查,基本上每個月的捕捉數 量及趨勢沒有很大的變化,這些年間的颱風及豪 雨都未曾對樣區造成重大損害,因此在穩定的棲 息地的狀況下,個體的存活是非常穩定的。另一 個原因有可能此地掠食者不多。根據我們的觀察 會 捕 食 山 椒 魚 的 動 物 有 菊 池 氏 龜 殼 花 (Trimeresurus gracilis) 、 高 砂 蛇 (Elaphe
mandarinus) 、 史 丹 吉 氏 斜 鱗 蛇 (Pseudoxendon stejnegeri)和標蛇類(Achalinus spp.)等活動於高山 的蛇類及地表啄食性的雉類如帝雉(Syrmaticus mikado)及藍腹鷴(Lophura swinhoii)。在長久的研 究期間中未曾在樣區內見過雉類也未曾發現或 翻找過高山蛇類,因此有可能因為掠食者稀少而 使存活率升高。 捕 獲 率 很 明 顯 的 受 到 季 節 及 溫 濕 度 的 影 響,在春季及夏季等氣溫及濕度較高的季節山椒 魚比較會到地表來活動。此時恰為牠們生殖活動 結束,需要大量的食物來補充因為生殖活動所損 失的能量,且高濕度時表土較為濕潤,能使牠們 在活動時少有失水的危險。而其餘季節,只要是 濕度較高,出現的也較多。因此未來在長期監測 時,應該考慮儘量在下雨過後的時間進行調查, 以使捕獲的個體數增加,進而減小估算值的信賴 區間。
族群增長率估算(Population growth rate)
利用Leslie matrix model 估計得到的族群增 長率(λ)為 1.077,表示此族群為穩定而略呈增長 的族群,但是能否證明此地的族群是健康而穩定 的呢?這需要其他數值來進一步說明。首先可以 用敏感度分析,此分析探討族群增長率如何受到 重要比率的改變的影響,並且能提供幾個有用的 資訊:(1)某個重要比率對族群增長率的重要性; (2)評估重要比率若未能在野外研究時準確估算 時產生的影響;(3)量化環境變動的影響,且將環
阿里山地區阿里山山椒魚的分布與族群監測 境變動與各時期的存活率及生殖力作連結;(4) 管理者可依據這些資訊來決定保育資源的著力 點(Caswell, 2001)。族群增長率的敏感度值在生殖 前期的存活率佔了 61.49%,而生殖期的數值為 0.6200 佔了總數的 35.85%,生殖力僅佔 2.66%, 因此存活率對族群的生存十分的重要。 由於敏感度值會因為不同重要比率的測量 刻度不同而產生偏差,使用敏感度值的彈性值可 以看重要比率的比例變化對敏感度值的影響。彈 性質的總和為 1,能評估關鍵生活史時期及轉換 期 , 以 及 生 殖 及 存 活 對 動 物 的 相 對 重 要 性 (Caswell, 2001)。表 4 中彈性分析的結果中,成體 存活率具有最高的敏感度彈性值(0.5177),表示此 時期的存活率是需要受到最大關注的項目。生殖 力所佔彈性值的比例為 10.2%,存活率依然佔較 高比例達 89.8%。成體存活率對族群的增長率而 言是最重要的項目。成體存活率經由矩陣分析後 對許多動物而言都是最重要的項目,包括帝王企 鵝、雪海燕、南方管鼻蒦鳥(Fulmarus glacialoides)、 深藍鶯(Dendroica cerulea)、黑熊(Ursus americans) 等 (Hebblewhite et al., 2003; Jenouvrier et al., 2005b; Jones et al., 2004)。然而這種現象在兩棲爬 行動物方面,目前沒有相關的文獻可供比較。阿 里山地區山椒魚族群的成體存活率很高,因此族 群仍呈現穩定的狀態。由於調查樣區是山椒魚成 體在非生殖季時主要的活動地區,保持樣區的穩 定對整個族群的永存是很需要的。山椒魚在阿里 山地區出現的地點,都在小溪流二側的高平坦地 上,因此要確保整個大阿里山地區山椒魚族群的 長存,這類的環境必需保持良好,以提高山椒魚 成體的存活率。 本研究提供了矩陣分析的初步模型,作為日 後的研究基礎。本文僅建構了生活史時期為基礎 的分析,未來每隻捕獲的個體若能以骨骼鑑齡法 確知年齡,再加上幼體現已可以使用螢光劑上 標,就能估算各年齡的存活率,使整個動態分析 能更為清楚。再者山椒魚在野外的生殖狀況,一 直都十分難以研究,這也是對瞭解山椒魚長期趨 勢 需 要 解 決 的 問 題 。 本 分 析 使 用 簡 單 的 deterministic model , 未 來 尚 可 進 行 重 要 比 率 stochastic model 的分析。這個研究法被許多學者 利用來研究族群的長期趨勢,同時能探討氣候變 遷、聖嬰現象、食物量對於族群動態的影響。山 椒魚是台灣高山動物的代表,近年來由於全球暖 化使高山氣候產生變遷,未來應該持續而長期對 山椒魚族群進行監測,瞭解高山氣候變遷對高山 動物各重要比率的影響。
誌 謝
本研究承林務局嘉義林區管理處(林務局保 育研究計畫 92-05-6-01; 93-05-8-1; 94-05-8-1)與 行政院農業委員會(92 農科-4.1.4-林-r1(8)、93 農 科-4.1.1-務-e2(2)、94 農科-9.1.7-務-e1(1))提供經 費。野外工作感謝實驗室同仁梁高賓、張俊文、 黃詠承、張雅淇、王昭均、許浩瑩、張天祐、柳 椒惠、蔡欣倫;嘉義林管處許碧如、蕭明學、劉 洋辰、魏世昌、劉啓同等人的幫助。參考文獻
陳世煌、呂光洋,1986,台灣產山椒魚之研究 ( 二 )— 阿 里 山 地 區 山 椒 魚 之 族 群 生 態 研 究。師大生物學報 21: 46-72。 陳世煌、呂光洋,1987,台灣產山椒魚之研究 (一)—研究歷史、分布和形態學之初步研 究。野生動物保育研討會專集(一)國家公園 和自然保留區之野生動物。頁 79-104。 葉明欽、呂光洋、賴俊祥,1994,阿里山及玉山 國家公園台灣山椒魚族群生態研究。師大 生物學報 29(2): 79-87。Bowen KD, Colbert PL, Janzen FJ. 2004. Survival and recruitment in a human-impacted population of ornate box turtles, Terrapene Ornata, with recommendations for conservation and management. J. Herpetol. 38: 562-568.
Caswell H. 2001. Matrix population models. 2nd ed. Sinauer Associates, Sunderland, Massachusetts, USA.
Cupp PV Jr. 1980. Territoriality in the green salamander, Aneides aeneus. Copeia 1980: 463-468.
Davis TM. 2002. Microhabitat use and movements of the wandering salamander, Aneides
vagrans, on Vancouver Island, British
Columbia, Canada. J. Herpet. 36: 699-703. Hebblewhite M, Percy M, Serrouya R. 2003. Black
bear (Ursus americanus) survival and demography in the Bow Valley of Banff National Park, Alberta. Biol. Conserv. 112: 415-425.
Jaeger RG, Kalvarsky D, Shimizu N. 1982. Territorial behaviour of the red-backed
salamander: expulsion of intruders. Anim. Behav. 30: 490-496.
Jenouvrier S, Barbraud C, Weimerskirch H. 2005a. Long-term contrasted responses to climate of two Antarctic seabird species. Ecology 86: 2889-2903.
Jenouvrier S, Barbraud C, Cazelles B, Weimerskirch H. 2005b. Modelling population dynamics of seabirds: importance of the effects of climate fluctuations on breeding proportions. Oikos 108: 511-522. Jones J, Barg JJ, Sillett TS, Veit ML, Robertson RJ.
2004. Minimum estimates of survival and population growth for Cerulean Warblers (Dendroica cerulea) breeding in Ontario, Canada. Auk 121: 15-22.
Kagekawa M, Iizuka K, Kuzumi S. 1989. Morphology of egg sacs and larvae just after hatching in Hynobius sonani and H.
formosanus from Taiwan, with an analysis of
skeletal muscle protein compositions. In "Current Herpetology in East Asia." ed. by Matsui M, Hikida T, Goris RC. Herpetological Society of Japan, Kyoto. pp.147-155.
Keen WH, Reed RW. 1985. Territorial defence of space and feeding sites by a plethodontid salamander. Anim. Behav. 33: 1119-1123. Krebs CJ. 1999. Ecological Methodology, 2nd ed.
Addison-Welsey Educational Publishers, Inc. 620pp.
Lebreton JD, Burnham KP, Clobert J, Anderson DR. 1992. Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecol. Mono. 62: 67-118.
Lai JS, Lue KY. 2008. Microhabitat Preference of Alishan Salamander (Hynobius arisanensis) in Taiwan. Zool. Stud. (Submitted)
Lue KY, Lin CY, Chuang KS. 1992. Notes on larva of Hynobius formosanus found on Yu-Shan National Park. Acta Zool. Sinica 31: 68-70.
Martof BS. 1953. Territoriality in the green frog,
Rana climitans. Ecology 43: 165-174.
Marvin GA. 1998. Territorial behavior of the plethodontid salamander Plethodon kentucki: Influence of habitat structure and population density. Oecologia 114: 133-144.
McKenny HC, Keeton WS, Donovan TM. 2006. Effects of structural complexity enhancement on eastern red-backed salamander (Plethodon
cinereus) populations in northern hardwood
forests. For. Ecol. Manage. 230: 186-196. Nijhuis MJ, Kaplan RH. 1998. Movement patterns
and life history characteristics in a population of the Cascade Torrent Salamander (Rhyacotriton cascadae) in the Columbia River Gorge, Oregon. J. Herpet. 32: 301-304. Smyers SD, Rubbo MJ, Townsend VR Jr., Swart
CC. 2002. Intra- and interspecific characterization of burrow use and defense by juvenile Ambystomatid salamanders. Herpetologica 58: 422-429.
Stickel LF. 1950. Population and home range relationship of the box turtle, Terrapene c.
carolina (L.). Ecol. Mono. 20: 353-378.
White GC, Burnham KP. 1999. Program MARK: survival estimation from populations of marked animals. Bird Study 46(Supplement): 120-138.
Williams BK, Nichols JD, Conroy MJ. 2001. Analysis and management of animal populations. Academic Press, San Diego, California, USA.
Wiltenmuth EB. 1996. Agonistic and sensory behaviour of the salamander Ensatina
eschscholtzii during asymmetrical contests.
Anim. Behav. 52: 841-850.
Worton BJ. 1989. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70: 164-168.
BioFormosa(2007)42(2): 105-117
Distribution and Population Dynamic of Alishan Salamander
(Hynobius arisanensis) in Alishan Areas
June-Shiang Lai, Kuang-Yang Lue*
Depatment of Life Science, National Taiwan Normal University Taipei, Taiwan
(Received: 12 March 2007, accepted: 6 August 2007)
ABSTRACT
From April 2002 to March 2006, we studied the distribution and population dynamic of Alishan salamander (Hynobius arisanensis) in Alishan Areas. We surveyed 73 sectors of wasabia plantation at 36 forestry areas around the Alishan areas, and found Alishan salamander in nine of them. Common characters of these localities were having a gradual rise slope and with small creeks. We use the area-constrained method to monitoring population dynamic in the Sister Ponds population. A total of 266 salamanders were captured in four years periods, with 94 recaptured salamanders. We used the Jolly-Seber method to estimate the monthly population size, it ranged between 13 and 305 individuals. It seemed that population size does not decreased as comparing with those estimation made by the former researchers. Most captured salamanders were adults, while juvenile were captured irregularly, and metamorphs only appeared in spring. Estimated home ranges of Alishan salamanders in the Alishan areas ranged 0-509 m2(64.9 ± 29.6m2 (n = 19)). We used the Cormack-Jolly-Seber model to modeling the survival and capture probability. The survival probability were constant, adults and juveniles were 0.996 and 0.997, respectively. Capture probabilities were seasonal-specific, the capture probability were significantly positive correlated with seasonal rainfall. Using the Leslie matrix model to investigate the population dynamics and to determine population growth rate of this population, it was 1.077, indicating that the population is stable and slow growth. The sensitivity and elasticity population growth rate both indicate that the adult survival rate was the most important vital rate for the population stability.
Key words: Hynobius arisanensis, distribution, population monitoring, population size, Cormack- Jolly-
Seber model, survival probability, capture probability, Leslie matrix model, population growth rate