桑黃粗抽物誘導血管平滑肌細胞(A10)凋亡及其相關機制; Phellins merrillii ( PM ) induces apoptosis of smooth muscle cells ( A10 ) and it’s related mechanisms.
108
0
0
全文
(2) 總目錄 頁次 中文摘要-------------------------------------------------3 英文摘要-------------------------------------------------5 表目錄---------------------------------------------------7 圖目錄---------------------------------------------------8 致謝----------------------------------------------------10 英文縮寫------------------------------------------------11 研究目的------------------------------------------------13 第一章、前言--------------------------------------------14 第一節、粥狀動脈硬化之介紹-----------------------14 第二節、粥狀動脈硬化的形成機制-------------------16 第三節、過度增生的平滑肌細胞與粥狀動脈硬化之關聯-19 第四節、細胞凋亡之介紹---------------------------20 第五節、桑黃之介紹-------------------------------24 第二章、材料方法----------------------------------------28 第一節、實驗材料---------------------------------28 第二節、實驗方法---------------------------------31 第三節、統計方法---------------------------------39. 1.
(3) 第三章、實驗結果-----------------------------------------40 第一節、PM 對 A10 細胞型態上的影響------------------40 第二節、PM 對 A10 細胞存活率的影響------------------41 第三節、PM 對 A10 細胞是否造成細胞凋亡之現象--------42 第四章、討論---------------------------------------------46 第一節、結果討論----------------------------------46 第二節、其他相關性討論----------------------------54 第三節、研究限制----------------------------------56 第五章、結論---------------------------------------------58 第六章、參考文獻-----------------------------------------59. 2.
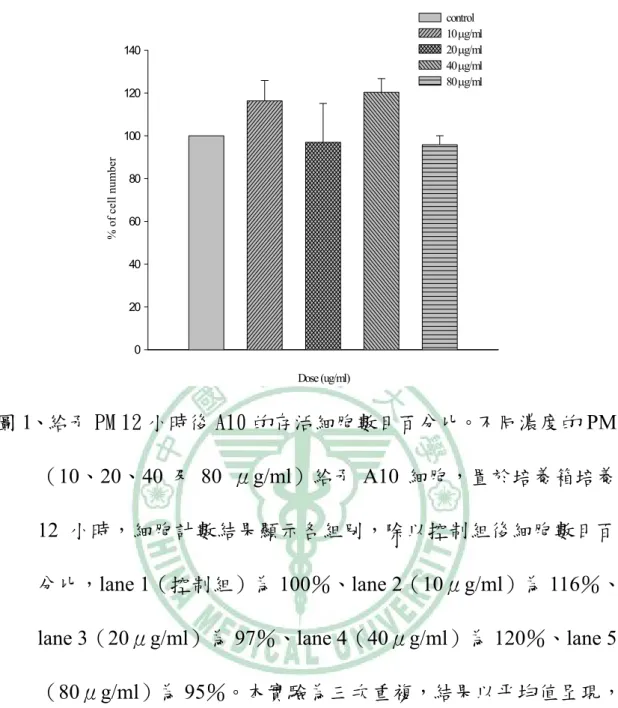
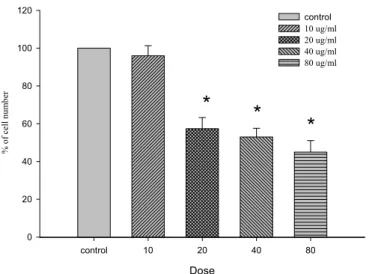
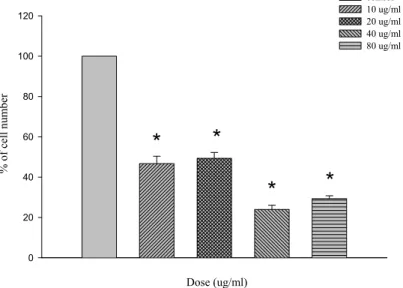
(4) 中文摘要 本 篇 論 文 主 要 的 研 究 目 的 , 在 探 討 桑 黃 Phellinus merrillii ( PM )誘導平滑肌細胞 ( A10 ) 凋亡 ( Apoptosis ) 的機 制。實驗過程中使用體外的(in vitro)細胞培養方式,給予 A10 細胞 不同濃度的 PM ( 10、20、40 及 80μg/ml ),分別處理 12、24、 48 及 72 小時之後,首先藉由細胞計數 ( cell count ) 方式觀察細 胞存活數目,結果顯示,20、40 及 80μg/ml 之 PM 給予 A10 細胞 24、 48 及 72 小時後與控制組 ( control ) 相較有明顯存活率下降 ( p <0.05) 的情形。然而給予 A10 細胞不同濃度的 PM 12 小時,並無 明 顯 之 存 活 率 的 改 變 。 接 下 來 我 們 藉 由 DNA 裂 解 ( DNA fragamentation ) 方式觀察凋亡情形,結果顯示 40、80μg/ml 之 PM 給予 A10 細胞 48 小時後可明顯地誘導 DNA 裂解。而進一步我們也 分別使用 MC540 / Propidium iodide 方式,以確定 PM 給予 A10 細胞 後所造成之凋亡情形,經由流式細胞儀的檢測,發現 A10 細胞經 PM ( 20、40 及 80μg/ml ) 處理 24、48 及 72 小時後,有明顯的凋亡 細胞發生,分別約為 22、31 及 33 %。最後為了探討 PM 誘導 A10 進行細胞凋亡的相關路徑,我們以不同濃度的 PM ( 10、20、40 及 80 μg/ml ) 分別處理 24、48 及 72 小時之後,抽取蛋白質以西方 墨點法進行分析。研究結果發現許多與細胞凋亡相關的蛋白在 A10. 3.
(5) 細胞給予 PM 後會有顯著性的表現增加。凋亡相關路徑中 p53 蛋白表 現,進而使得下游的 p21 也被活化,且與凋亡相關的蛋白如 Fas、Bax 及 p38,也會隨著 PM 濃度增加而有較多的蛋白表現量。這顯示由 PM 所 誘 導 的 細 胞 凋 亡 路 徑 中 , 所 參 與 的 可 能 角 色 包 含 了 Extrinsic pathway. 及 Intrinsic pathway 。 綜 合 以 上 結 果 , 我 們 發 現 桑 黃. Phellinus merrillii ( PM ) 可誘導平滑肌細胞 ( A10 )發生細胞 凋亡 ( Apoptosis ) 進而抑制細胞增生。. 4.
(6) 英文摘要 To investigate how Phellinus merrillii ( PM ) affects the smooth muscle cell ( A10 ) growth and study their mechanisms in inducing apoptosis are the main purposes of the research. In. in vitro study,. various concentrations of PM ( 10、20、40 and 80 μg/ml ) were applied in different time period ( 24、48 and 72 h ).. Firstly, We. observed the viable cells through cell count, and, data showed that 20、 40、80 μg/ml of PM could significantly reduce the viability of A10 cells after 24、48、72 h . Secondly, through DNA fragmentation assay results showed that 40、80 μg/ml of PM obriously induced DNA fragmentation in A10 cells. Then, through MC540 / Propidium iodide assay, analyzed by flow cytometry, results showed that 20、40、80 μg/ml of PM could significantly induce A10 cells apoptosis after 24、48、72 h, (22、31、 33 %, respectively) Finally, total protein extracted from A10 cells treated with PM in different time and concentration periods were subjected to Western blot analysis. Results showed PM could induce p53、p21、Fas、Bax、p38 expression. These suggest that PM could induce its apoptosis through both extrinsic. and intrinsic pathways. In. conclusion, evidences shown above suggest that Phellinus merrillii. 5.
(7) ( PM ) could induce apoptosis of A10 cells, therefore inhibit the proliferation of A10 cells.. 6.
(8) 表目錄 頁次 表 1、細胞凋亡與細胞壞死之比較-----------------------------------------63 表 2、Phosphate buffer saline(PBS)配製法-----------------------------66 表 3、蛋白質量化所使用之配方表------------------------------------------63 表 4、DNA marker 的製作方式----------------------------------------------64. 7.
(9) 圖目錄 頁次 圖 1、給予 PM 12 小時後 A10 的存活細胞數目百分比-----------------71 圖 2、給予 PM 24 小時後 A10 的存活細胞數目百分比-----------------72 圖 3、給予 PM 48 小時後 A10 的存活細胞數目百分比-----------------73 圖 4、給予 PM 72 小時後 A10 的存活細胞數目百分比-----------------74 圖 5、 PM 對 A10 細胞存活率依時間和劑量相關性的影響------------75 圖 6、給予 PM 後 24 小時 A10 細胞型態---------------------------------76 圖 7、給予 PM 後 48 小時 A10 細胞型態---------------------------------77 圖 8、給予 PM 後 72 小時 A10 細胞型態----------------------------------78 圖 9、給予 A10 細胞 PM 後 48 小時 DNA fragmentation 的影響-------79 圖 10、給予 PM 後 24 小時細胞凋亡偵測試驗------------------------------80 圖 11、給予 PM 後 48 小時細胞凋亡偵測試驗-----------------------------82 圖 12、給予 PM 後 72 小時細胞凋亡偵測試驗------------------------------84 圖 13、給予 PM 細胞凋亡偵測試驗,依時間和劑量相關性的影響-86 圖 14、Annexin V 及 propidium iodide 雙染-------------------------------87 圖 15、p53 蛋白 Western blot 的表現量-------------------------------------88 圖 16、p53 蛋白 Western blot 表現量依時間和劑量相關性的影響--90 圖 17、Bax 蛋白 Western blot 的表現量-------------------------------------91. 8.
(10) 圖 18、Bax 蛋白 Western blot 表現量依時間和劑量相關性的影響---93 圖 19、p21 蛋白 Western blot 的表現量-------------------------------------94 圖 20、p21 蛋白 Western blot 表現量依時間和劑量相關性的影響---96 圖 21、Fas 蛋白 Western blot 的表現量--------------------------------------97 圖 22、Fas 蛋白 Western blot 表現量依時間和劑量相關性的影響---99 圖 23、p38 蛋白 Western blot 的表現量-------------------------------------100 圖 24、p38 蛋白 Western blot 表現量依時間和劑量相關性的影響--102 圖 25、caspase 3 Western blot 的表現量-----------------------------------103 圖 26、caspase 3 Western blot 表現量依時間和劑量相關性的影響--105 圖 27、桑黃造成 A10 細胞凋亡可能之路徑-------------------------------106 圖 28、桑黃---------------------------------------------------------------------107. 9.
(11) 誌謝 首先,最感謝的是我的指導教授 許明志老師在實驗過程中非常 有耐心的指導,及論文撰寫時相當細心的教導與討論,使得本論文可 以順利完成,並且讓我在研究所兩年的時間當中,獲益良多。文稿始 成之初,承蒙中興大學廖俊旺老師與本校梁育民老師等口試委員,撥 冗審閱,大力釜正,提供諸多寶貴意見,使內容得以更加完善在此致 上衷心的謝意。此外就讀期間承蒙生理學科蔡主任宗統老師、陳卓昇 老師、林靜茹老師、鄭雅興老師、黃怡萍老師、鞏志力老師、運醫系 王慧如老師、生化科李妙蓉老師、惠華姐、麗如姐在研究上及生活上 的關心。另外感謝張恆源學長提供桑黃 Phellinus merrillii(PM) 。也 感謝學長姐義忠、曉妘、佩玉、馨葶、怡文…等及同學們玉潔、芝奇、 文坽、蓉蓉、祐暄、立仁、菁秀…等在課業及實驗上的協助。最後, 要感謝我的父母、大姐、二姐、小妹、國安於生活上持續的鼓勵與支 持。 謹將此論文獻給所有關心、鼓勵和協助我的師長、同學和家人。. 呂雅亭 謹致於 中國醫藥大學醫學研究所 中華民國九十五年六月. 10.
(12) 英文縮寫 APS. Ammonium persulfate. Caspase. Cysteine dependent aspartate specific protease. DISC. Death-inducing signaling complex. DMEM. Dulbecco’s Modified Eagle’s Medium. DMSO. Dimethyl Sulfoxide. DTT. Dithiothreitol. ECL. Enhanced chemilminescence. FBS. Fetal bovine serum. IL-1. Interlukin-1. IL-4. Interlukin-4. IL-10. Interlukin-10. INF-γ. Interfiron-γ. LPC. Lysophosphatidylcholine. MAP-1. Monocyte chemotactic protein-1. M-CSF. Marophage colony-stimulating factor. 10-ODA. 10-oxo-trans-8- decenoic acid. PCNA. Proliferating cell nuclear antigen. PVDF. Polyvinylidene fluoride. 11.
(13) PBS. Phosphate buffer saline. PG. Phellins givus. PI. Propidium iodide. PL. Phellinus linteus. PM. Phellinus murrillii. PR. Phellinus rimosus. SDS. Sodium dodecyl sulfate. SOD. Superoxide dismutase. TNF. Tumor Necrosis Factor. TNF-α. Tumor necrosis factor-α. VCAM. Vascullar cell adhesion molecule-1. VSMCs. Vascular smooth muscle cells. 12.
(14) 研究目的 由於國人的生活飲食習慣日漸西化,使得心臟及腦血管疾病已經 分別躍居 2004 年國人十大死因之亞軍與季軍,其中粥狀動脈硬化則 是誘發心血管疾病的危險因子之一,故研發心血管疾病的治療藥物也 成為大家所重視的主題。在中草藥典籍上,有許多適用於治療心血管 疾病的中草藥研究,例如:丹參 (salvia miltiorrhiza)、厚朴酚 (magnolol),還有銀杏的粹取物 (ginkgo biloba extract)…等,在 實驗動物模式中證實可以減緩粥狀動脈硬化的發生與進行,但如何藉 由中草藥之特性與發展,提升粥狀動脈硬化的治療與預防,將是值得 我們深入討論的議題。 本研究選擇中藥當中的藥用菇菌-桑黃 ( Phellinus merrilliis; PM)屬多孔菌科,相關研究指出其具有誘導癌細胞死亡、抑制發炎 反應、增強免疫能力…等特性。由於,粥狀動脈硬化疾病形成機制中 包含血管平滑肌細胞異常增生情形,其異常增生情形與癌症細胞相 似,因此本論文利用桑黃具有誘導癌症細胞死亡之特性,實驗中主要 研究目的將著重於探討桑黃對於血管平滑肌細胞(A10)之生長抑制 作用及其誘導 A10 細胞凋亡的機制。. 13.
(15) 第一章、前言 第一節、粥狀動脈硬化之介紹 由於國人的飲食習慣日漸西化,生活變得緊湊而忙碌,使得一些 會導致心血管疾病的因素,包括高血壓、糖尿病、高血脂等發生的比 例逐漸上升,根據衛生署統計處所公佈 2004 年國人十大死亡主因, 心臟及腦血管疾病分別囊括了第二、三名(62),可見其盛行率與發 生率皆相當高。此外在 2004 年全民健保之國民醫療保健支出中,用 於循環系統疾病約有 650 億元,佔 11%,排名第三,為社會成本之 沉重的負荷,所以對於心血管疾病的預防與治療方法的改進和研發, 是為一個極待解決與研究的課題。 心血管疾病是心臟病與血管疾病的總稱,在人體心臟為主要動力 提供者,而血管為體內各組織器官養分輸送和廢物排除的主要溝通管 道,當心臟無法維持正常功能或血管因為某些原因而造成阻塞會使得 血流無法通過,即會造成損傷而發生生命危急狀況。粥狀動脈硬化 ( Atherosclerosis ) 則是誘發心血管疾病的危險因子之一,其中成人最 主要的心血管疾病就是由其所造成的阻塞或狹窄,有時候會造成心肌 缺氧或壞死等嚴重後果。臨床上較易出現的病況有心絞痛、急性心肌 梗塞、心臟衰竭、心律不整,甚至是猝死等症狀。 根據流行病學的研究指出動脈硬化疾病的危險因子,主要是高膽. 14.
(16) 固醇血症、高血壓、糖尿病、年紀大、抽煙及早發性動脈硬化疾病家 族史、而肥胖及男性亦被視為重要的危險因子。具有這些危險因子的 人,大皆被證實會傷害內皮細胞或使其功能失常,因而容易發生動脈 硬化。 對於粥狀動脈硬化的預防應多為日常生活習慣的改善,包含避免 攝取含有高油脂食物、控制體重、養成運動習慣等等。現今對於已經 有動脈硬化現象的人,傳統上的治療方式分為:血管粥狀動脈硬化引 起血管狹窄程度小於 60 % 時,通常只採用藥物治療,意即控制高血 壓、糖尿病、高血脂症,再加上抗血小板聚集劑或抗凝血劑;如果超 過60%的動脈狹窄,則必須考慮積極治療,如施行血管內膜剝離術或 放置頸動脈支架等手術治療。. 15.
(17) 第二節、粥狀動脈硬化的形成機制 基本上而言,粥狀動脈硬化是一種脂質與發炎細胞的聚積並伴隨 著平滑肌細胞 ( smooth muscle cell, SMC ) 增生與細胞外間質液分泌 ( extracellular matrix secretion ) 所引起的細胞內膜纖維變性 ( intimal fibrosis ) 。造成之動脈壁增厚、變硬或失去彈性的的病理變化。動脈 硬化可依據其硬化方式分為:粥狀動脈硬化、動脈中層鈣化及小動脈 硬化;臨床上以粥狀動脈硬化為主。 粥狀動脈硬化形成的機制,主要包括四個階段,分別為內皮細胞 層功能不良 ( endothelial dysfunction ) 、脂肪條的形成 ( fatty-streak fomation ) 、壞死中心與纖維帽 ( necrotic core and Fibrous-cap ) 的 形成及不穩定的纖維斑塊 ( unstable fibrous plaques ) 。這四個階段 都是影響粥動脈硬化疾病嚴重度的重要的因素。 在初期內皮層功能不良的階段中,粥狀動脈硬化形成的原因,主 要是血流所產生的擾流與不規則剪力 ( Shear Stress ) ,使內皮細胞物 理因素改變,發炎細胞(1)的黏著堆積而釋放出趨化因數,導致細 胞通透性增加,是早期動脈硬化症形成的關鍵步驟。另一方面若血液 中過多的脂蛋白 ( 例如:低密度脂蛋白;LDL ),經由血液動力學 的影響,被動的經由通透性增加的內皮細胞間隙,穿過血管內皮細胞 層,而進入到血管內膜中。這些進入血管壁中的 LDL 會與結構性蛋. 16.
(18) 白 相 黏 合 , 且 極 易 受 到 血 管 內 皮 細 胞 所 釋 放 之 活 性 氧 與 12-LO ( 12/15 lipoxygenase ) 攻 擊 , 而 氧 化 為 氧 化 態 的 低 密 度 脂 蛋 白 ( ox-LDL )(2) 。 第二個階段中,ox-LDL 會刺激血管內皮細胞產生許多的黏附趨 化因數,例如 MCP-1 ( monocyte chemotactic protein )、 ICAM-1 ( Intercellular adhesion molecule-1 ),VCAM-1 ( vascullar cell adhesion molecule-1 ) 與 E-selectin 等,使內皮細胞變得容易被血液 內 的 細 胞 吸 附 。 同 時 ox-LDL 亦 會 吸 引 血 液 中 的 單 核 球 細 胞 ( monocyte ) 向 內 皮 的 表 面 黏 附 而 進 入 血 管 內 皮 細 胞 下 ( subendothelial ), 一 方 面 分 泌 細 胞 激 素 ( cytokines ) , 如 Interlukin-1 ( IL-1 ),Interlukin-4 ( IL-4 ),interfiron-γ ( 干擾素-γ; IFN-γ ),Tumor Necrosis Factor ( TNF ) 進行發炎反應(1、3)。 另 一 方 面 則 是 接 著 受 到 血 管 內 皮 細 胞 所 釋 放 的 M-CSF ( marophage colony-stimulating factor ) 分化為巨噬細胞。這些存 在 於 血 管 壁 中 的 巨 噬 細 胞 表 面 具 有 清 道 夫 受 體 ( scavenger receptor ) ,對於 ox-LDL 具有極高的親合力,形成一含巨噬細胞之泡 沫細胞 ( macrophage foam cell ) 也稱為 ”泡沫細胞” 。此時平滑肌細 胞會受到泡沫細胞的刺激,穿過彈性纖維層進入內膜增生,然後泡沫 細胞及平滑細胞,則由內層移位至內膜增生形成脂肪線. 17.
(19) ( Fatty-streak )(3)。 第三階段則由於巨噬細胞不斷的吞噬 ox-LDL,產生大量的泡沫 細胞,極易堆積在血管壁並且死亡(4)。並藉由 Interferon 的作用使 細胞走向凋亡,再與其他脂質碎片形成一個壞死中心 ( necrotic core ) 。血管壁中層膠原蛋白與平滑肌細胞過度增生,遷移至血管內 膜層形成纖維帽 ( Fibrous-cap ) ,其功能在於隔開血管腔以及受損 區域。 動脈硬化最後一個階段中,血管壁會形成動脈硬化斑塊。動脈硬 化斑塊是由平滑肌、泡沫細胞、低密度脂蛋白及其他脂質所組成,也 可以稱為粥狀動脈腫 ( atheroma )。此時血管的管腔會因為硬化斑塊 的產生而狹窄,持續擠入活化的巨噬細胞會釋放出分解蛋白的酵素, 這些酵素會分解基質,使原本覆蓋於壞死中心上的平滑肌細胞亦受到 刺激進行凋亡之機制(4) 。而此機制結果會導致斑塊不穩定,甚至於 破裂,增加粥狀動脈疾病之嚴重度。. 18.
(20) 第三節、過度增生的平滑肌細胞與粥狀動脈硬化之關聯 有相當多的文獻指出,平滑肌細胞在粥狀動脈硬化形成過程中, 扮演了很重要的角色(63、64、65)。其中原因為粥狀動脈硬化形成 過 程 中 , 血 管 內 皮 細 胞 所 釋 放 的 M-CSF (. marophage. colony-stimulating factor ) 分化為巨噬細胞,對 ox-LDL 具有極高的 親合力,形成一含巨噬細胞之泡沫細胞 ( macrophage foam cell ),進 而刺激平滑肌細胞穿過彈性纖維層進入內膜增生(3) ,增生的平滑肌 細胞再與泡沫細胞、低密度脂蛋白及其他脂質形成動脈硬化斑塊,造 成血管的管腔狹窄(4)。由於平滑肌細胞是動脈最主要的構成物之 一,且當異常增生的平滑肌細胞對粥狀動脈硬化疾病的形成有很重要 的影響,因此很多的文獻極力的在探討,如何抑制過度增生的平滑肌 細胞,普遍方法為造成平滑肌細胞的細胞週期停滯使其無法 proliferation ;或者造成細胞的 Apoptosis(65、63、64)。 由上述可知,抑制異常增生的平滑肌細胞已成為探討粥狀動脈硬 化疾病的重要實驗模式,而文獻中,使用 體外( in vitro )實驗 的設計中,促成平滑肌細胞異常增生的方式包含:實驗時增加細胞培 養時的血清比例(63) ,或者給予細胞生長因子等方式(64) 。本論文 實驗過程中,採用將細胞培養時的血清比例,由原本的 10 % 增加 至 15 %,模擬平滑肌細胞異常增生的情形。. 19.
(21) 第四節、細胞凋亡之介紹 細胞凋亡 ( Apoptosis )首先被 Kerr, Wyllie, 及Currie 等病理 學家於1972年用來描述一種過程被嚴密控制的細胞死亡機制且與細 胞壞死 ( necrosis ) 不同的現象【表1】,指出當細胞受環境刺激後, 在基因調控之下所產生的自然死亡現象,故亦稱之為細胞計劃性死亡 ( programmed cell death )(5、6)。 一般而言細胞死亡的形式分兩大類:一類是細胞壞死 ( necrosis ) 指細胞受到外在激烈且有害的刺激,通常為非生理性的損傷如:毒藥 作用、缺氧、發炎等反應時;或細胞內在環境嚴重的錯亂而造成細胞 急劇的死亡,所以正常的生理狀態下細胞死亡是不走此途徑的,過程 中細胞膜通透性增加使水分大量進入細胞中,細胞因而腫脹破裂,典 型的特徵有:細胞膨脹、被分解、細胞核及細胞質的內涵物被釋放到 細胞外等現象。另一類細胞死亡的形式則為細胞凋亡 ( apoptosis ), 是一種藉由基因的調控,維持體內細胞數量動態平衡、清除不需要或 病變細胞的基本措施。 細胞凋亡在多細胞生物中扮演幾個生理角色,包括:挑選最適細 胞 ( the fittest cells )、塑造個體及器官形態的機制、原狀恆定 ( homeostasis )、剔除危險性細胞等功能,可見細胞凋亡在生物體之正 常發育及生理功能是所不可或缺的,過多或過少的細胞凋亡可能導致. 20.
(22) 疾病的發生(6)。 細胞凋亡的形態學特徵包括有:染色質濃縮 ( pyknosis )、細胞 質濃縮、細胞形態變得不規則而且有斷裂 ( fragmentation )、及由凋 亡細胞脫落出細胞凋亡體 ( apoptotic body ) 等情形。而在生化學特徵 包括了:細胞膜變化、凋亡蛋白酵素 ( caspases )的活化、與去氧 核醣核酸的斷裂 ( DNA fragmentation )。細胞膜變化:凋亡中的細胞 會在細胞膜上表現出一些 “請吞噬我” 的信號來告知其鄰近細胞或 巨噬細胞。這些信號已知有 phosphatidylserine ( PS )由細胞膜內 翻轉至膜外的表層, anionic thrombospondin binding sites,與一些醣 分子。這些信號可能在細胞凋亡的早期便會出現(7)。凋亡蛋白酵 素的活化,這些凋亡蛋白酵素的作用造成大部份細胞凋亡的典型表 徵,標的受質(target substrates)可歸納成三大類:凋亡蛋白酵素的 前質、凋亡過程中必須去活化的蛋白質、與凋亡過程中必須活化的蛋 白質。去氧核醣核酸的斷裂:凋亡細胞所產生的斷裂去氧核醣核酸會 在膠片電泳上呈現 “階梯” 圖案,這個階梯圖案被認為是細胞凋亡 的生化學特徵之一(8),因此看到去氧核醣核酸的斷裂,為現今偵 測細胞凋亡有用的方法,例如 terminal deoxynucleotidyl transferase mediated dUTP nick end labeling ( TUNEL ) 可用來測定細胞凋亡。 另外,利用 ELISA 的斷裂去氧核醣核酸測定法更可用來定量細胞凋 21.
(23) 亡的程度。 細胞凋亡在細胞內的分子機制大致可分為二種:包含有 extrinsic pathway 及 intrinsic pathway。extrinsic pathway 是由death ligand 鍵 結至death receptor,進而將死亡的訊息送到細胞內,誘發一連串凋亡 事件的產生。而 intrinsic pathway 則是經由 DNA damage、hypoxia、 和生長因子的缺乏所引發,其主要過程中會活化 apoptosis proteins 且抑制 antiapoptosis proteins 的表現(9)。 細胞凋亡的過程中涉及許多相關的基因和蛋白,其中研究最多的 包含:caspase家族、p53、Fas等。(A)Caspase 為半胱氨酸蛋白酶, 當細胞接受來自藥物或細胞激素( TNF, FasL, IFN )的刺激,將引 發細胞內死亡訊息的傳遞,最後會活化caspase,活化後主要會切斷細 胞內的結構蛋白質,並破壞某些參與代謝的酵素和訊息傳遞的蛋白 質。如:被活化的caspase-8、9能引起Caspase 級聯反應,進而降解結 構蛋白和功能蛋白,引起凋亡,此外細胞中也具有caspase的抑制因 子 , 這 些 因 子 屬 於 一 個 龐 大 的 蛋 白 家 族 稱 為 IAPs ( inhibitors of apoptosis proteins ) ,他們能與caspase 結合,抑制其活性,藉由此抑 制以相互調節。(B)p53為一抑癌基因,存在於細胞核內為一核蛋 白,當細胞的DNA受到損傷、生長因子短缺、或缺氧…等情況,進 而會觸發凋亡的進行,過程中活化的p53會進一步促進下游凋亡相關 22.
(24) 基因的轉錄如:pro-apoptosis Bax protein、Fas/CD95 ligand、PIGs ( p53- induced genes )等,導致 apoptosis的進行;且其亦可活化抑 制存活相關蛋白的表現如:IGF-BP3 protein來促使apoptosis的發生。 (C)Fas屬於TNF家族,具有三個富含半胱氨酸的胞外區和一個死亡 結合區 ( Death domain ),當FasL (Fas ligand )與Fas結合後, 會使胞內的Death domain 改變並與pro-caspase-8(或-10)結合,形成 DISC(death-inducing signaling complex),進而激活caspase( 8、9、 and 10 ),引起Caspase 級聯反應,最後降解結構蛋白和功能蛋白, 引起細胞凋亡(7、10、11)。. 23.
(25) 第五節、桑黃(Phellinus merrillii)之介紹 一、藥用菇類簡介 中要典籍很早即記載部分真菌(菇類)之子實體及其菌絲體均可 被食用或應用於醫療上。而近年來許多文獻指出靈芝、樟芝、松茸、 舞茸或巴西蘑菇等菇類具有抗腫瘤、降血壓、降膽固醇或增強免疫力 等生物活性,因此如何從中得到有效的抗氧化或抗腫瘤成分,以製造 抗心血管疾病或抗腫瘤新藥,是目前及未來的研究趨勢(12)。 菇是一種具有明顯子實體( fruit body )的大型真菌,能形成此 種子實體之菇類,少數為子囊菌綱真菌,大部分屬於擔子菌綱,其中 大多為食藥用菇類,少數為毒菇。因食藥用菇之子實體集菌絲體富含 有蛋白質、多醣、有機酸、核苷酸和酵素等成分及抗腫瘤、降血壓等 生物活性物質,現已成為高經濟價值及學術價值之產品(13、14)。 近來的研究中發現,菇類的子實體萃取液或是菌絲體發酵液中, 含有相當多的生理活性成分,如水溶性多糖體、超氧歧化酵素 ( superoxide dismutase, SOD ) 及 10-ODA ( 10-oxo-trans-8decenoic acid )等,分別具有免疫調節、自由基清除等特性(10、11) 。 此外還發現菇類萃取物還具有抗腫瘤(15、16) 、增加免疫力(20)、 降血壓(17)等生理活性功能。特別在抗腫瘤方面,菇類的子實體萃 取液或是菌絲體發酵液中具有多醣體(β-葡聚醣以及與蛋白質結合. 24.
(26) 的葡萄糖)、triterpene、agglutinin、dietary fiber 等,主要功用為恢復 或增強體內免疫能力,經由活化多種免疫細胞如:刺激巨噬細胞、嗜 中性細胞、T 細胞、B 細胞及自然殺手細胞等(20),以達抗腫瘤之 效果。 二、桑黃之生物特性 桑黃為一種寄生於桑樹上的多年生覃菇類,在分類上桑黃是屬於 真菌界( Fungi )、擔子菌門( Basidiomycotina )、真正擔子菌綱 ( Hymenomycetidae ) 、平滑臥孔菌目( Aphyllophorales ) 刺革 菌科( Hymenochaetaceae )、火木層孔菌屬( Phellinus )、桑黃種 ( Phellinus merrillii ) 。本論文所使用之桑黃,經大陸中國科學研究 院戴玉成博士及台中科學博物館植物組主任吳聲華博士確認為 Phellinus merrillii 。 桑黃特徵為馬蹄型,長約 9 公分,寬 6~15 公分,厚 2~5 公分, 傘的表面具有,呈現淡紅褐色或暗褐色,有明顯的環溝與縱溝形成龜 裂紋。邊緣的新生部分為鮮黃色,底部則由鮮黃色逐漸變成黃褐色, 而底部的管孔是多層黃色或黃褐色的楔形,會生長厚膜的剛毛,藉此 可和其他種類區別。 生長時主要寄生於桑樹的古木上,但也會寄生於其他的闊葉樹 上,是會讓心材腐朽枯死的木層孔菌型的白色腐朽菌,原本為瘤狀,. 25.
(27) 慢慢的生長為扇狀。主要產地為中國、菲律賓、北美、中南美等地。 『本草綱目』將桑黃歸納為木耳的項目中,指其為硬蕈類,記載 療效包含:治療子宮出血、白帶、月經不順等婦女病,支氣管炎、氣 喘、肺結核等呼吸系統疾病,胃痛、下痢等消化系統疾病,以及過度 疲勞、淋巴腫瘤等疾病。 桑黃子實體的主要成分包含有:多醣體β-(1-3)D 葡聚糖、β(1-6)D 葡聚糖、蛋白質結構的酸性雜聚糖α-(1-4)、三帖類 ( truterpenoids ) 、 腺苷、脂肪酸( C22 及 C24 的飽和脂肪酸 ) 及氨基酸。 三、桑黃之生物活性 目前國內外關於桑黃的研究中,主要是針對其子實體的藥理作 用,包含有:具有抗肝纖維化作用,結果顯示桑黃能明顯改善肝功能, 提高 SOD 活性,表現出清除自由基的作用。也具有增強自體免疫能 力之作用,研究顯示桑黃可以誘發γ-干擾素( IFN-γ )的能力 「30」、刺激巨嗜細胞( marcrophage )活化(24)、刺激巨噬細胞 產生 interleukin-10( IL-10 )「26」、誘導殺手細胞(27)活化進而 調節自體免疫能力,抑制腫瘤細胞的增殖作用(30)。 四、桑黃的其他相關研究 綜合文獻發現桑黃除了抑制癌症及抗氧化的作用外,尚包含具有. 26.
(28) 免疫調節、增加傷口癒合、降低血糖等作用。首先,在免疫調節方面, 文獻指出桑黃可以增強免疫能力,藉由促進活化免疫細胞如:T 細 胞、B 細胞、自然殺手細胞、巨噬細胞等以增強免疫能力(24、26、 27) 。此外桑黃亦具有減低過敏反應的作用(28) ,2003 年 kim 等學 者提出桑黃可藉由減低血清中的 IgG 及 IgG2a 且減低 tumor necrosis factor-α( TNF-α)與 interferon-γ( IFN-γ)以達到減低過敏之 作用(29)。而也有學者提出桑黃的減低敏感作用中,interferon-γ ( IFN-γ)的減少可能與 IgE 的減少有關聯(30) 。再來,在增加傷 口癒合方面,文獻指出體內( in. vivo )的實驗方式,給予桑黃的. 組別其傷口癒合會較控制組快(34),且於糖尿病鼠的傷口癒合試驗 中,給予桑黃的組別其傷口癒合會較控制組快(31)。其次在減低血 糖方面,研究顯示使用體內( in. vivo )的實驗方式,在糖尿病鼠. 中,給予桑黃的組別其體重、血糖濃度及飲水量等,皆會較控制組別 良好(33、36)。. 27.
(29) 第二章、材料與方法 第一節、實驗材料 一、儀器 無菌操作臺(造鑫,Taiwan) 細胞培養箱(Nuaire,USA) 光學顯微鏡(Motic,Japan) 離心機(Beckman) 細胞計數器(Haemocytometer;Boeco,Germany) 酸鹼值測定計(C831;Consort,UK) 微量天平(GR–200;A&D,Japan) 乾浴槽(Model 110001;購自 Boekel) 水浴槽(TKS,Taipei,Taiwan) 去離子水製造機(Minipore,USA) 超高速離心機(Himac CS 120GX) 蛋白質轉漬電泳套組(Bio–Rad,USA) 流式細胞儀(FACSCalibur,BD,USA) 影像分析儀器(AlphaImagerTM2200,USA) Power supply(Hoefer,San Francisco,CA,USA) ELISA reader(ANTHOS–2020,Salzbrug,Austria). 28.
(30) Vortex–genie 2(SCIENTIFIC INDUSTRIES,NY,USA) SDS–PAGE 電泳槽套組(Bio–RAD,USA) 二、材料 細胞培養皿 (騰達行,Taiwan) 細胞培養盤 (騰達行,Taiwan) PVDF 轉漬膜 (Minipore,USA) 蓋玻片 (Kimble,USA) 載玻片 (Marriefeld,Germany) 冷凍管 (騰達行,Taiwan) 微量離心管 (季勗,Taiwan) 離心管 (季勗,Taiwan) X–film (Kodak,Boston,MA,USA) 三、試劑 Acrylamide/Bis(Amresco,OH,USA) Annexin V-FITC Apoptosis Detectopn Kit(BioVision,CA,USA) APS (Ammonium persulfate;Amresco,OH,USA) Bradford reagent (Amresco,OH,USA) Caspase 3 substrate reagent Kit(OncoImmunin,MD,USA) DMSO (Dimethyl Sulfoxide,Sigma,MI,USA) DMEM (Dulbeccco’s Modified Eagle’s Medium;GIBCO,USA) 29.
(31) ECL kit (Perkinelmer,Boston,MA,USA) FBS (Fetal Bovine serum;Invitrogen,CA,USA) Glycine (Amresco,OH,USA) Glyerol (Amresco,OH,USA) Hydrochloric acid (RDH,SE,Germany) Methanol (景明化工,Taiwan) Penicillin-Streptomycin (Invitrogen,CA,USA) PI (Propidium iodide;Invitrogen,CA,USA) Protein assay-Dye reagent concentrate (Bio-Rad,MA,USA) Protein maker (Amersham,UK) RNase A (Invitrogen,CA,USA) SDS (Sodium dodecyl sulfate;Amresco,OH,USB) Sodium pyruvate (GIBCO,USA) TBA (2-Thiobarbituric acid;Sigma,MI,USA) TCA (Trichloroacetic acid;Sigma,MI,USA) TEMED(Amresco,OH,USA) Transfer Buffer(10X TG Buffer)(Amresco,OH,USA) Tris (Amresco,OH,USA) 脫脂奶粉(安佳,New Zealand) 顯影劑(Kodak,Boston,MA,USA) 定影劑(Kodak,Boston,MA,USA). 30.
(32) 第二節、研究設計 一、 藥物配製 將桑黃【圖 28】子實體傘部,以 70 %酒精冷泡 48 小時,取出 溶有桑黃之酒精,將酒精減壓濃縮至膏狀,再將膏狀物置烘箱中使其 烘至塊狀,將塊狀之桑黃研磨至粉狀,完成粗粹步驟。秤取桑黃粉末 32 mg,加入細胞培養等級之 DMSO ( Sigma 編號:D2438)溶解, 先配製好濃度 32. mg/100μl 之 PM 溶液,作為平日儲存的 Stock 濃. 度,實驗前新鮮配製成 10、20、40、80 及 160 μg/ml 等濃度備用。 二、 細胞株培養 血管平滑細胞株 (購自財團法人食品工業發展研究所生物資源 保存與研究中心,生資中心編號:60127,細胞株名稱:A10)來自 胎鼠胸主動脈 ( the thoracic aorta of DB1X embryonic rat ),多應用於 血管平滑肌 ( vascular smooth muscle cells; VSMCs ) 實驗模型。 使用之培養基為 Dulbecco’s Modified Eagle’s Medium 內含胎牛血 清 ( Fetal bovine serum ; FBS;一般培養使用 10 % FBS、實驗進 行時使用 15 % FBS、同步化時使用 0 % FBS;血清購回後需先水浴 56℃加熱 30 分鐘後使用 ) 、 HEPES Buffer Solution 使用劑量 1 %、 Strpotomycin/penicilline 使用劑量 1 %。 將存放血管平滑肌細胞的冷凍小管自液態氮中取出後插上浮. 31.
(33) 板,迅速放入 37℃ 水浴槽中使其快速解凍,將解凍之細胞懸浮液從 冷凍管中放入10 公分培養皿,並加入約10 ml 含10 %胎牛血清之培 養基,混合均勻後放入細胞培養箱 (37 ℃、95 % O2、5 %CO2 ) 中 進行培養,每二至三天更換一次培養基。當細胞長約9分滿時,則進 行繼代培養。繼代時將舊的培養廢液抽吸乾淨,加入7 ml 1x PBS【表 2】緩衝液 ( pH:7.4 ) 清洗一次,加入Trypsin- EDTA 1ml incubate. 2. 分鐘,加入 1 ml 10% FBS- DMEM 2 ml 中和Trypsin-EDTA 作用,以 抽吸方式將細胞沖散,將培養皿液體吸換至離心管後加入10 % FBSDMEM共10 ml,離心1200 rpm,5分鐘,計算細胞數目後進行日常分 盤或實驗用分盤。處理後再將細胞置於細胞培養箱 (37 ℃、95 % O2、 5% CO2 ) 中進行培養。 三、 細胞週期同步化(synchronize) 將細胞分到實驗適合之dish或plates後 (例如:10cm dish 細胞數 目約1 x106 cells/ well),以含10 % 胎牛血清(FBS)的培養液培養細 胞,並在放入培養箱前稍微搖晃使細胞均勻分佈於dish上。待12~24 小時細胞適應環境並貼覆上 plates 穩定後吸去培養廢液,再以1x PBS緩衝液 (pH:7.4) 2 ml/well 小心清洗1 次,最後加入10 ml/well 的0 %胎牛血清培養液,經24小時後即完成細胞同步化步驟。 四、 細胞生長抑制試驗(cell count). 32.
(34) 應用 Trypen blue exclusion 實驗計數樣品中的細胞數,因活細胞 會排斥 trypan blue 染劑,僅死細胞會被染成藍色,因此計數亮色的 細胞數可相對得知其細胞存活率。 將細胞分盤至10 cm dish培養,種植細胞數約1×106/dish,以15% FBS DMEM1ml 培養24小時後,以1 x PBS 10ml 沖洗1次,加入 0﹪ FBS-DMEM 10 ml/dish同步化24小時,以含不同濃度藥物之培養基培 養至時間點,再將10 公分培養皿內之培養液吸去,以約10 ml 的1x PBS 緩衝液小心清洗細胞兩次後加入約1 ml 的1x trypsin-EDTA 置 入37 ℃培養箱內,待2 分鐘後取出。以輕拍培養皿底部的方式將細 胞完全脫離培養皿懸浮起來。加入1 ml 的培養液中和 trypsin-EDTA 活性,並用1 ml 的pipette 以緩慢來回抽吸方式將細胞打散。將打散 後之細胞換到15 ml 離心管中,並加入10 ml培養液稀釋混勻。如所含 細胞密度過高,則算出的數目誤差較大,需再加入更多的培養液稀 釋。取出100 μl 細胞懸浮液和10 μl typan-blue,在96-well內混合均 勻,共取2次。分別從混合液中各取出10 μl 的細胞懸浮液放在細胞計 數盤上下兩個凹槽上,利用蓋上蓋玻片時的虹吸作用將細胞均勻平均 分散於細胞計數區域。在顯微鏡下計數細胞數目(細胞顏色較亮者為 存活的細胞)並算出細胞計數盤內9大格中5格 (左上角及右上角區域) 之活細胞總數 ( N 值)。N 值除上10,再乘上1.1 (稀釋倍數)x10. 33.
(35) (細胞懸浮液總量)x 104 即可得細胞總數目 ( cells/10ml )。 五、 DNA 抽取與電泳分析 將 A10 細胞同步化並給予藥物 PI(20、40 及 80 μg/ml)後, 於 48 小時後,使用 trypsin-EDTA 將細胞收起,以 DNA ladder kit (GeneMark,Atlanta,USA)將細胞中的 DNA 抽出,並將 DNA 暫 放置於冰上,再取 Agarose 粉末與 1x TAE 溶液放入血清瓶中,以微 波每次 30 秒加熱致澄清為止。將滾燙的液態膠倒入 50ml 離心管中, 以手觸摸離心管外壁帶膠體溫度約降至 50oC 左右後加入 EtBr ( 量為 1μl/10ml Gel ),混合均勻。將膠體緩慢倒入造膠台中,並以 tip 將膠 上的氣泡趕走。待膠硬化後,加入 300~35 ml 的 1x TAE 溶液當作 Running buffer。Loading 6~10ul 的 DNA Marker 【表 4】 ,再 Loading sample ( DNA 與 6x Loadong buffer 以 5:1 之比例混勻 )。最後設定 100V、30 分鐘為跑膠條件。 六、 細胞凋亡偵測試驗 1、使用 MC540 及 propidium iodide 進行偵測 首先將 MC540 溶在 50% EtOH 中,配製成濃度為 1mg/ml 之溶 液,而 propidium iodide 則溶在 PBS 中,配製成濃度為 5ug/ml 之溶液 備用。將 A10 細胞同步化 ( 24 小時)並給予藥物 PM(20、40 及 80μg/ml)後,於 24、48 及 72 小時後,使用 trypsin-EDTA 將細胞收. 34.
(36) 起,再加入 0.375 ml 之 10﹪FBS-DMEM,接著分別加入 125 ul 的 propidium iodide ( 濃度為 5ug/ml ) ,及 2.5 ul 的 MC540 ( 濃度為 1mg/ml ) ,混合均勻,靜待約 1 分鐘後,上機使用流式細胞儀 ( flow cytometer ) 檢測凋亡細胞的比率。 2、使用 Annexin V-FITC Apoptosis Detection Kit 進行偵測 將 A10 細胞同步化並給予藥物 PM (20、40 及 80 μg/ml )48 小時後,使用 trypsin-EDTA 將細胞收起,加入 500 ul 的 Binding Buffer,再加入 5 ul 的 Annexin V-FITC 及 5 ul 的 propidium iodide, 避光靜置於室溫中 5 分鐘,混合後放於載玻片上,蓋上蓋玻片,使用 螢光顯微鏡進行凋亡細胞之檢測。當被 Annexin V-FITC 染到將呈現 綠色螢光,而被 propidium iodide 染到將呈現紅色螢光。 七、 西方墨點法 ( Western blot ) 偵測相關蛋白變化 1、細胞蛋白質抽取 將培養於 10cm dish 的細胞取出,加入 5 ml 1x PBS 沖洗 2 次, 加入 500 μl 的 lysis buffer 反應 3 分鐘,使用細胞刮杓將細胞刮下並 將細胞液體收集至微量離心管放入乾浴加溫器內加溫 ( 95℃、5 分 鐘 ) ,隨即放置於冰上 30 秒以上,4℃離心 ( 13000 rpm、10 分鐘 ) , 取上清液,進行蛋白質定量或冰存於 -20℃。 2、蛋白質定量. 35.
(37) 使用 Bradford 製作蛋白質檢量線【表 3】,Bradford 為蛋白質染 劑,有毒性,使用時需戴手套。加入染劑混合均勻後需反應 5 分鐘在 測吸光值 。吸光值測定時每個樣本數需 3 重複,使其數值穩定。以 測定出來的標準品吸光質與蛋白質濃度畫出檢量線,並求出趨勢線方 程式即 R2 值。R2 值需 > 0.99 以上的準確度。算出蛋白質檢量線並求 出趨勢線方程式,經帶入測得之吸光值 ( y ),則可求出蛋白質濃度 ( μg/ml )。 樣品蛋白質濃度測定時,取 10 μl 的樣品與 790 μl 的 DDW 混合, 再加入 200 μl 的 Bradford 染劑,均勻混合 5 分鐘後以 O.D 590 測定吸 光值。將樣品吸光值 ( y ) 帶入算出趨勢線方程式,則可算出樣品蛋 白質濃度 ( x )。 3、蛋白質電泳( SDS-PAGE ) 組合鑄膠檯,注入 DDW 確定無滲漏之情形,依照下列配方先配 製 12% 下層膠。下層膠注入造膠檯內約八分滿,剩餘的空間先用 DDW 填滿去除上面的氣泡並壓平膠之上緣,等待凝固約 30 分鐘, 造上層膠,TEMED 則須等下層膠凝固後將 DDW 倒出後加入,下層 膠凝固後,注入上層膠,插入 comb 等待凝固約 30 分鐘將 comb 取 出後先用 DDW 清洗 well,將膠檯放入電泳槽內裝滿 Running buffer, 將 loading sample 先加熱 ( 95℃、5 分鐘 )立即放置於冰上冷卻 30 秒以上,將 Marker ( 5~10μl )和 sample ( 10 μl )loading 到 well 內,利用 100V 跑電泳約 2 個半小時,取出下層膠準備進行 蛋白質轉漬至 PVDF 膜 Western blot 步驟。 36.
(38) 4、西方墨點法(Western blot) PVDF membrane 需先用 Methanol 潤濕 30 秒,將下層膠自水龍頭 下取去下,並將膠體放至轉漬夾上其順序如下: 白底網夾 海綿 Filter paper. 正極(+). PVDF membrane 膠體(gel) Filter paper. 負極(-). 海綿 黑底網夾. 將轉漬夾合上放入 Transfer box 蓋上蓋子放置冰桶中並加滿冰塊,放 入 4℃冰箱內,Transfer 條件設定 60 valtage,轉印時間 2 小時,將轉 印後的 PVDF membrane 放入 5 % fat-free milk 室溫 bloclking 2 小 時,0.1 % PBST 搖盪 5 分鐘,共 3 次,把 membrane 放入含有一級 抗體的小盒內,4℃冰箱搖晃至隔天。接著取出 membrane 放入 0.1 % PBST 搖盪 5 分鐘,共 3 次,將 membrane 放入含有二級抗體,室溫 搖盪 1 小時,取出 membrane 放入 0.1 % PBST 搖盪 5 分鐘,共 3 次 後進行暗房底片顯影。 5、暗房底片顯影(壓片) 37.
(39) 準備用物:dropper、cassete、鑷子、感光底片、剪刀、透明膠片、 ECL kit、Developer、Fixer,先將 membrane 浸泡於 ECL (比例為 1:1 )混合液中約 30 秒,將 membrane (正面朝上)放置於透明膠 固定好,剪適當大小的底片,曝光時間依 membrane 上冷光亮度決定, 數秒至 1 小時,曝光後用顯影劑及定影劑洗底片,後將底片風乾保存 並進行分析。. 38.
(40) 第三節、統計方法 實驗結果以 SPSS 軟體之單因子變異數分析(one way ANOVA) 計算分析。數據結果以 mean ± SE 表示各項值。 (圖中*表示與 15% FBS-DMEM 相比,P<0.05) 。. 39.
(41) 第三章、實驗結果 第一節、PM 對 A10 細胞型態的影響 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間分別為 24、48 及 72 小時。藉由倒立光學顯微鏡,放大倍率 40 倍觀看細胞型態上之變化。 發現給予 PM ( 24 及 48 小時 ) ,濃度 20、40 及 80 μg/ml 時,細 胞型態上呈現較不規則且偏圓形,與正常 A10 細胞之紡垂狀不同【圖 6、圖 7】 。而在 PM ( 72 小時 ) 時,濃度 10、20、40 及 80 μg/ml 時,細胞型態上呈現較不規則【圖 8】 。. 40.
(42) 第二節、PM 對 A10 細胞存活率的影響 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (10、20、40 及 80 μg/ml),放入培養箱培養時間分別為 12、24、 48 與 72 小時。藉由 trypan blue 染劑進行細胞計數 ( cell count ) 而判定細胞存活率之高低。結果顯示,不同濃度之 PM 於 A10 細胞 12 小時並無明顯抑制細胞生長【圖 1】 ;將時間增加至 24、48 小時後, 濃度 20(p<0.05) 、40(p<0.05)及 80(p<0.05) μg/ml 有抑制 細胞生長情形【圖 2、圖 3】 ;於 72 小時之作用時間點,發現 10(p <0.05)、20(p<0.05) 、40(p<0.05)及 80(p<0.05) μg/ml 濃 度之 PM 皆會對 A10 細胞產生抑制的作用【圖 4、圖 5】 。. 41.
(43) 第三節、PM 對 A10 細胞是否造成細胞凋亡之現象 一、以 DNA 裂解方式探討 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml) ,放入培養箱培養 48 小時。將細胞中的 DNA 抽出後,利用 agraros DNA 電泳分析方式,觀察是否有因為細胞凋亡, 所造成的典型 DNA 裂解片段( DNA fragmentation )的現象。結果 顯示,PM( 40 及 80 μg/ml )作用 48 小時,有較控制組明顯之 DNA 裂解片段( DNA fragmentation )的現象發生【圖 9】 。 二、以 MC540 / Propidium iodide 方法探討 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間分別為 24、48 及 72 小時。使用 MC540 及 Propidium iodide 雙染之方式,藉由流式細胞儀 的檢測下,觀察早期及晚期凋亡細胞。結果顯示,PM ( 24、48 及 72 小時 ) 在不同濃度下皆會造成 A10 細胞,早期及晚期細胞凋亡 比例明顯增加,早期加上晚期凋亡細胞比率分別約為 23 %、30 % 及 33 %【圖 10、圖 11、圖 12、圖 13】。 三、以 Annexin V-FITC 方法探討 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM ( 20、40 及 80 μg/ml ) ,放入培養箱培養時間分別為 48 小時。使. 42.
(44) 用 Annexin V-FITC 之方式,藉由螢光顯微鏡的檢測下,觀察早期及 晚期細胞凋亡細胞。結果顯示,PM ( 20、40 及 80 μg/ml )於 48 小時,與控制組相較細胞核會帶有紅色螢光,且細胞膜有表現綠色螢 光,表示細胞有細胞凋亡之現象【圖 14】。 四、以細胞凋亡蛋白探討 (一)p53 蛋白與 A10 細胞凋亡之關係 A10 細胞在同步化後,給予不同濃度的 PM ( 20、40 及 80 μ g/ml ) ,放入培養箱培養時間為 24、48 及 72 小時,以西方墨點法分 析藥物對於細胞中 p53 蛋白的表現變化。結果顯示,PM ( 20、40 及 80 μg/ml ) 於 24 小時會顯著增加 A10 細胞中 p53(p<0.05) 的表現,然而,於 48 及 72 小時,儘管有觀察到 p53 蛋白的表現量增 加,但並沒統計上的意義【圖 15、圖 16】 。 (二)Bax 蛋白與 A10 細胞凋亡之關係 A10 細胞在經過同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml) ,放入培養箱培養時間為 24、48、72 小時,以西方墨點法分 析藥物對於細胞中 Bax 蛋白的表現變化。結果顯示,PM ( 20、40 及 80 μg/ml )於 24 及 48 小時會顯著增加(p<0.05)A10 細胞中 Bax 的表現量,然而,於 72 小時,儘管有觀察到 Bax 蛋白的表現量 增加,但並沒統計上的意義【圖 17、圖 18】 。. 43.
(45) (三)p21 蛋白與 A10 細胞凋亡之關係 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間為 24、48、72 小時, 以西方墨點法分析藥物對於細胞中 p21 蛋白的表現變化。結果顯示, PM ( 40 及 80 μg/ml )於 24 小時會顯著增加 A10 細胞中 p21 的 表現(p<0.05) ,然而,於 48 及 72 小時,儘管有觀察到 p21 蛋白的 表現量增加,但並沒統計上的意義【圖 19、圖 20】 。 (四)Fas 蛋白與 A10 細胞凋亡之關係 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間為 24、48、72 小時, 以西方墨點法分析藥物對於細胞中 Fas 蛋白的表現變化。結果顯示, PM 會增加 A10 細胞中 Fas 的表現,但並沒統計上的意義【圖 21、圖 22】 。 (五)p38 蛋白與 A10 細胞凋亡之關係 A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間為 24、48、72 小時, 以西方墨點法分析藥物對於細胞中 p38 蛋白的表現變化。結果顯示, PM 會增加 A10 細胞中 p38 的表現,但並沒統計上的意義【圖 24】。 (六)Caspase 3 蛋白與 A10 細胞凋亡之關係. 44.
(46) A10 細胞在經過 24 小時細胞週期同步化後,給予不同濃度的 PM (20、40 及 80 μg/ml),放入培養箱培養時間為 24、48、72 小時, 以西方墨點法分析藥物對於細胞中 caspase 3(17 kda)蛋白的表現變 化。結果顯示,PM 會增加 A10 細胞中 caspase 3(17 kda)的表現, 但並沒統計上的意義【圖 25、圖 26】 。. 45.
(47) 第四章、討論 第一節、結果討論 本研究主要以體外(in vitro)方式為主軸,實驗進行中將培養基 的血清濃度由原本的 10 % 提高至 15 %,主要是利用血清本身含有 生長因素可以促進細胞增生,模擬粥狀動脈血管疾病中平滑肌細胞增 生的狀態,進而探討給予桑黃( Phellinus merrilli PM )是否可誘導 增生的平滑肌細胞(A10)產生細胞凋亡的情形,進而減少細胞增殖。 一、桑黃對於平滑肌細胞存活率的影響 粥狀動脈硬化是一種慢性發炎伴隨著自由基傷害、脂質堆積與平 滑肌不正常細胞增生的血管性疾病,不正常的細胞增生就如同癌細胞 一樣。在本論文中,將增生的平滑肌細胞加入不同濃度的 Phellinus merrillii,經由細胞計數後發覺在 20μg/ml 的濃度下,培養 24 小時 開始即開始產生存活率下降、細胞數減少作用,且具有統計上的意義 (p<0.05) 【圖 2】 ,此外在 20μg/ml 的濃度下培養 24 小時後,細胞 形態改變 (包括細胞皺縮、細胞質濃縮、細胞核型態改變),因此認 為異常增生的平滑肌細胞給予桑黃後會造成其細胞凋亡。 文獻中有非常多的報告指出,真菌這種高等植物可以有效的抑制 腫瘤的增生,而它的效果大多是透過宿主的防衛功能增強來達到目 的。這種防衛功能的增強是因為高分子多醣體的影響。實驗結果發. 46.
(48) 現,靈芝可顯著促進抗體產生增強自然殺手細胞(NK Cell)之免疫 功能,同時證實口服、腹腔內注射與靜脈注射給藥都有效,而且呈現 良 好 的 劑 藥 反 應 ( dose-dependent manner ) 與 時 程 反 應 (time-dependent manner)關係(66) 。靈芝也能促進造血功能及抑 制人血癌細胞(HL-60)增殖。由抗腫瘤有效成份分析結果證實,靈 芝這種防衛功能的增強,同樣也是一種叫高分子多醣體的關係。這種 成份可以增加抗體、誘導干擾素、增加自然殺細胞,自然而然的增強 宿主的免疫功能,抑制不正常的腫瘤細胞。最近實驗結果更發現靈芝 能激發人類巨噬細胞產生細胞激素。於誘導人血癌細胞(Leukemic U937)分化效應的實驗中發現靈芝能刺激人血液單核細胞(Blood Monouclear Cell)分沁細胞激素如 TNF-α、INF(interferon) 、IL-1β 等明顯的抑制人血癌細胞(U973)分裂(Proliferation)並誘導其 分化(Differentiation)完全。而於靈芝抗肺癌實驗時發現靈芝能 有效延長肺癌動物存活時間與增加存活率。若與抗癌藥 Adriamycin、 Fluorouracil、Thioguanine、Methotrexate 、Paclitaxel 或人工 合成免疫功能增強劑 Imexon 合併使用更能有效的延長動物存活時間 達三倍於控制組並顯著地降低動物死亡率。因此靈芝有明顯增進抗腫 瘤免疫功能對腫瘤防治具有重大意義。除了我們對於靈芝於抑制癌症 方面的了解以外,其實其它蕈類對於癌症的預防以及治療有下列諸多. 47.
(49) 的研究。 而文獻中桑黃對癌症的影響,以體外 ( in. vitro)方式中,. 而與 Phellinus merrillii (PM)同屬不同種的桑黃,也具有相似 的效果,2005 年有研究指出不同種名之桑黃 Phellinus igniarius(PI) 萃取出的 phelligridimer G 對於人類卵巢癌細胞 ( A2780 )及人類 結腸癌細胞 ( HCT-8 )皆有抑制作用(22)。同年也有研究指出 Phellinus igniarius 萃取出的 phelligridimer A 於 10.2 μM 可造成肝 癌細胞 50% 的致死率(21) 。而 Phellinus linteus (PL)文獻指出其 可抑制多種癌症細胞,並造成細胞凋亡的發生,包含黑色素瘤細胞 ( B16BL6 ) 、結腸癌細胞( SW480 )等,無論是無論體內(in vivo) 或體外的方式(in vitro) (35、37、38、39、40) ,並且細胞內機制可 能包含活化 ERK、p38、NF-κB 所造成(39、43)或是活化 cytochrome c 及抑制 Bcl-2 所造成(38) 。Phellins givus(PG)有關的研究也指出 其可造成人類胃癌細胞(TMK-1)proliferation 的減少及 Apoptosis 的 發生,並可抑制 MCA-102 細胞生長,並在體內(in vivo)的實驗中 發現 PG 可造成腫瘤體積變小(27、37) 。Phellinus rimosus(PR)相 關的研究也指出其可造成淋巴癌細胞及肝癌細胞發生抑制的情形 (41、42)。 二、桑黃誘導平滑肌細胞產生細胞凋亡. 48.
(50) 為了要了解 PM 誘發 A10 細胞生長抑制的可能基轉,我門進一 步研究 PM 是否會誘發細胞凋亡的產生。因此使用流式細胞儀及螢光 顯微鏡偵測細胞凋亡早期變化,卽細胞膜上磷脂質絲氨酸 ( Phosphatidyl Serine;PS )的外露;健康的細胞膜具不對稱性,磷 脂質絲氨酸只存在於細胞內膜層而不外露,並且在晚期時會有細胞膜 不完整與 DNA 斷裂的情形,因此在本論文中使用 Annexin V 及 Propidium iodide 進行雙染,其中 Annexin V 會與翻轉至膜外的磷脂質 絲氨酸特異性結合,而發出綠色螢光,另外晚期 Propidium iodide 會 與斷裂的 DNA 結合。所以早期的凋亡只有 PS 翻轉只表現 Annexin V 的綠色螢光,而晚期的細胞凋亡則除了 PS 翻轉表現的綠色螢光外, 還有細胞膜不完整所以 Propidium iodide 會進到細胞中與 DNA 結合發 出紅色螢光。而本論文還使用 MC540 與 Propidium iodide 進行雙染, 其中 MC540 與 Annexin V 為相同作用。在實驗結果中,藉由流式細 胞儀得知,在平滑肌細胞中給予桑黃可以增加凋亡細胞的百分比(p <3 個不同時間點的百分比將之列出來並且註明其 p value 統計)【圖 10、11、12】,且在螢光顯微鏡的使用下,看到只有在給予桑黃情形 下,在細胞核才會出現紅色螢光,而細胞膜才會產生綠色螢光【圖 13】。因此我們可以推論給予桑黃後,會造成增生的平滑肌細胞走向 細胞凋亡。. 49.
(51) 相關聞文獻提及,醫藥蘑菇Ganoderma lucidum (G. lucidum) 在東 方社會被使用為癌症的預防。其中高分子多醣體乃為一免疫促進劑試 教為人所知的一部份,事實上,其它主要組成部分包括三萜類。在此 篇報告中,作者了解從G. lucidum萃取出三萜類的萃取物WEES-G6對 於癌症的作用。發現WEES-G6可以抑制人類的hepatoma Huh-7 細胞 成長, 而且,其不會抑制正常人肝臟細胞株。WEES-G6藉著活化JNK 和p38 MAP kinases,降低protein kinase C,可以致延長細胞周期的G2 期,在正常肝臟細胞沒有了這些現像。磨菇多聚糖,從Phellinus linteus 之水層萃取物有抗腫瘤和免疫調節的物質,特別是,酸性多聚糖 (aPL)。aPL於動物體內顯著誘導NO之產生。藉由NO之產生,aPL增 加B16細胞的死亡率。這些結果建議, PL刺激NO產生為tumoricidal, 並且過程也使PL導致它的治療作用的機制(67)。由Phellinus linteus 分 離 出 來 的 protein-bound 多 聚 糖 細 胞 毒 素 抑 制 人 類 結 腸 癌 細 胞 SW480的擴散和增殖。流式細胞儀分析表示, PL促進細胞的凋亡,並 且其sub-G1和G2/M兩者階段的細胞數目都有增加的情形產生,細胞 的凋亡和成長抑制可能與Bcl-2的減退, 細胞色素c釋放的增加聯繫在 一起,和減少了cyclin B1表現有關。這些結果建議, PL可以有抗腫瘤作 用。從Phellinus linteus分離出來之proteoglycan (PG)可以因為活化 CD11c+CD8+DC 以 及 type-1. helper. 50. T 細 胞 , 進 而 抑 制.
(52) MCA-102-tumor-bearing 癌症的成長(68)。從Phellinus linteus的果 實或菌絲體水層萃取物有抗腫瘤和免疫調節的活性(69) 。從Phellinus linteus被分離出來之酸性多聚糖(aPL)可以刺激增殖T淋巴細胞和B細 胞之活化,和抑制腫瘤成長和轉移(70)。據早先報導, 從Phellinus linteus多聚糖PL刺激了細胞調節和體液免疫。PL單獨增大延長了 B16F10變種老鼠的生存率,抑制腫瘤生長在NCI- H23-變種的裸體老 鼠,和減少了B16F10 肺黑瘤轉移率。Adriamycin顯著地抑制了腫瘤成 長,但輕微地抑制了轉移。PL和adriamycin的組合式療法更有效在抑制 腫瘤成長,但不是轉移。PL沒有誘導直接毒性在癌細胞裡,這是免疫化 學療法的特性。總而言之, PL也使用在癌症的免疫化學療法由於它的 有效的活力在腫瘤成長和轉移通過患者的免疫效力沒有毒力(71)。 關於Phellinus gilvus (P.G.)在黑色素瘤方面,其抽出物PG抑制B16F10 黑 色 素 瘤 細 胞 株 的 增 殖 並 且 造 成 其 細 胞 凋 亡 ( 72 ) 。 Polypore macrofungus, Phellinus rimosus (Berk) Pilat.之乙酸乙酯、甲醇和含水萃 取物對於Dalton,slymphoma腹水(DLA)及Ehrlich,s腹水癌(EAC)細胞株 具有細胞毒性和抗癌活性。研究結果推論,對這個蘑菇的潛在的用途 作為抗癌症使用,也有學者發現Phellinus linteus的抽出物可以抑制血 管新生,並有抗氧化的作用。在以上諸多對於靈芝以及桑黃的研究 中,我們可以發現蕈菇類在癌症方面的研究非常的豐富,那麼到底桑. 51.
(53) 黃對於異常增生的平滑肌細胞影響為何呢,是我們研究的重點。 三、桑黃誘導平滑肌細胞走向凋亡的路徑 細胞凋亡的程式進行可藉由外在途徑,例如:Fas / Fas ligand、 死 亡 接 受 器 ; 或 內 在 途 徑 , 例 如 , Bax / Bcl-2 、 粒 線 體 釋 放 出 cytochrome C 等調控,最後皆會活化 caspase 3 表現進行細胞凋亡。 血管平滑肌細胞凋亡參與粥狀動脈硬化行程過程即包括此二條途 徑。文獻指出Shear stree 引起血管平滑肌細胞凋亡是經由 Fas / Fas ligand 途徑(60) ;Chae 等人也在2004年研究報告指出, Nitric oxide 也會引起大鼠血管平滑肌細胞進行凋亡,而這個機制是經由短暫的 cytochrome C,活化 caspase-3 及 Bax / Bcl-2的表現所引起的(61)。 在本實驗中,為了探討細胞在凋亡過程中的機制,運用了西方墨點法 的方式,找出參與其中的相關蛋白,發現當中有一些相關的蛋白有表 現增加的情形,綜合文獻可能推論為p38蛋白活化後造成 p53 蛋白的 表現量增加,進一步促進下游凋亡相關基因的轉錄如:pro- apoptosis Bax protein、Fas / CD95 ligand,使胞內的 Death domain 改變並與 pro-caspase-8 ( 或 -10 ) 結 合 , 形 成 DISC ( death-inducing signaling complex),進而激活 caspase-8(-10),引起Caspase 級聯反應,最 後降解結構蛋白和功能蛋白引起細胞凋亡,並活化下游p21使細胞週 期停滯造成細胞無法proliferation。也或者 p38 活化後造成MAPK級. 52.
(54) 聯反應而造成細胞凋亡(45、46、60、61)。但在本實驗中有增加表 現的蛋白有p38、p53、Fas、Bax、caspase 3、p21等蛋白,因此顯示 細胞凋亡所走的路徑可能為, p38 造成 p53 蛋白表現增加,進而使 得下游的 p21 也被活化,且與凋亡相關的蛋白如 Fas、Bax,最後活 化 caspase 3 而使細胞凋亡。這顯示由PI所誘導的細胞凋亡路徑中, 所參與的可能角色包含了 Extrinsic pathway 及 Intrinsic pathway。. 53.
(55) 第二節、其他相關性討論 一、血管平滑肌增生與自由基之關係 近年研究報告對於粥狀動脈硬化的形成原因中,對於血管平滑肌 細胞增生的探討我們可以發現,有許多的因素可以引起血管平滑肌的 增生。如 ox-LDL 和 acetyl-LDL 引起巨噬細胞吞噬,形成泡沫細胞 (44);另有文獻指出 LDL 與 ox-LDL 可以藉由活化訊息傳遞路徑 MAPK patheway,或藉由 ERK1/2 之間的 cross link 引起細胞的增生 及遷移(47、48、49、50) ;另外 ox-LDL 也可以引起細胞內生長因 數,如 FGF-2 之釋放,達到刺激細胞增生。Marjorie 等學者提出, ox-LDL 刺激平滑肌細胞增生,主要是引起細胞週期蛋白質的表現。 細胞週期是由許多不同的蛋白質所共同調控,包括:cyclin D1、 cyclin-dependent kinase 或 PCNA 等(51)。Proliferating cell nuclear antigen (PCNA)是 DNA polymerase δ 輔助蛋白,主要分佈於核質, 其主要扮演的角色是與 cyclin 或 cdks 及 P21 相互作用以調控細胞週 期的進行並與 DNA 合成有關(46);而 ox-LDL 氧化產物包括 4-hydroxynonenal、lysophosph atidylcholine 也會刺激平滑肌細胞的增 生(46、52) 。 2000 年亦有文獻指出,內皮細胞的 ox-LDLreceptor-1(LOX-1) 與 ox-LDL 結合後會產生 ROS,並且隨著 ox-LDL 濃度增加 ROS 的. 54.
(56) 量亦會隨之增加(53) ,血管平滑肌細胞亦被發現有 LOX-1 而 ox-LDL 脂 質 氧 化 產 物 lysophosphatidylcholine 會 增 加 血 管 平 滑 肌 細 胞 LOX-1mRNA 的表現(55) ,推測這個現象的原因可能是因為 ox-LDL 在不同濃度所產生自由基的總量不同引起不同的細胞外訊息傳遞路 徑因而導致增生的結果;另一方面也可能是 ox-LDL 脂質氧化代謝產 物不同或代謝產物總量的差異引起細胞毒性而導致不同的結果。 抗氧化劑具有保護身體,防止自由基形成、去除體內所產生的自 由基 ( freeradicals );這些自由基是由氧氣和體內的各種物質反應 而產生,會造成一連串有害的連鎖反應,而對體內組織造成各種傷 害。抗氧化劑,例如:脂溶性維他命E、水溶性維他命C,雖然抗氧 化機制不同但是對於降低氧化傷害(55、56、57),及抑制血液中膽 固醇含量都有顯著的成效(58),並且可以降低並預防心血管疾病及 粥狀動脈硬化疾病進展(59)。而文獻上,也指出桑黃也具有抗氧化 的作用,顯示桑黃可以活化超氧歧化酵素 ( superoxide dismutase, SOD )進而清除自由基。. 55.
(57) 第三節、研究限制 粥狀動脈硬化血管疾病是高死亡率的現代文明病的一種,許多研 究學者致力於研究其治病機轉,尋找可以預防或降低此疾病的發生 率,而本實驗使用了藥用菇類-桑黃 ( Phellinus merrillii )進行實 驗。結果顯示桑黃在粥狀動脈硬化中平滑肌細胞增生具有誘導凋亡的 作用,這也是本研究的主旨,對於粥狀動脈硬化疾病的進展可以藉由 自然界中的產物而達到預防或降低的效果,更可以減少因服用化學合 成藥物副作用所造成的更進一步的傷害。 在研究的限制上,本論文採體外的實驗方式進行,沒有考慮藥物 是否經過體內代謝後有了不一樣的效果,因此在往後的體外方式中, 或許可以在細胞中添加肝臟的代謝酵素,證實桑黃的效果在經過代謝 後會不會產生變化。 此外也可以計算給予桑黃後細胞之 LC50 的藥物濃度及給藥時間 等,以更一步確認給予桑黃後之抑制增生的平滑肌細胞之效果。 在給藥濃度方面,經由細胞計數後發覺在 20μg/ml 的濃度下, 培養 24 小時開始即開始產生存活率下降、細胞數減少作用,且具有 統計上的意義(p<0.05)【圖 2】 ,但若將藥物濃度提高至 40、80μ g/ml 並無隨著藥物濃度上升而有加劇的情形。而在本實驗中曾將藥物 濃度提高至 160、320μg/ml,但發現此濃度之桑黃溶解度不佳,且給. 56.
(58) 藥後細胞會似壞死般突然大量死亡,無法進行細胞凋亡試驗。故在本 論文中將濃度設定於 20、40 及 80μg/ml。 而未來的實驗上也可朝體內模式的方向前進,方法可以將動物施 行頸動脈的氣球擴張術,利用此術後會造成平滑肌細胞異常增生之特 性,建立平滑肌細胞增生之動物模式,再經口餵食桑黃,觀察餵食桑 黃後是否減少平滑肌細胞增生之情形。. 57.
(59) 第五章、結論 首先在存活率的實驗中發現,A10 細胞經由 PM 處理後,細胞生 長情形明顯受到抑制【圖 2、3、4、5】 ,且細胞型態 ( Morphology ) 在給藥後也明顯改變,細胞膜變的不規則、皺縮 ( shrinkage ) 【圖 6、7、8】 。然後藉由 DNA fragmentation 的電泳分析中,了解 48 小 時作用時間點時,給藥組 (40 及 80 μg/ml )會較控制組有 DNA 的裂解片段【圖 9】 。再來,我們為了更進一步確定細胞凋亡的可能, 運用二種細胞凋亡偵測方式,經由流式細胞儀及螢光顯微鏡檢測下, 結果顯示給予 PM 組別(20、40、80 μg/ml)與控制組相比,明顯 會有較多的凋亡細胞被偵測出【圖 10、11、12、13】 。最後,為了明 白給藥後 A10 凋亡相關蛋白之表現情形,藉由西方墨點法,發現 p53 【圖 14】 、Bax【圖 15】 、p21【圖 16】、Fas【圖 17】 、p38【圖 18】、 Caspase 3(17 kda) 【圖 19】蛋白,於給予 PM 後似乎有較多的表現 量。推測凋亡相關路徑中 p53 蛋白表現增加,進而使得下游的 p21 也 被活化,且與凋亡相關的蛋白如 Fas、Bax 及 p38,也會進而有較多 的蛋白表現量【圖 20】 。 故根據上述的研究結果顯示,桑黃 Phellinus merrillii(PM)於體 外實驗(in vitro)方式中,具有誘發平滑肌細胞(A10)走向細胞凋 亡(Apoptosis)的情形。. 58.
(60) 第六章、參考資料 1. Libby P. Inflammation in atherosclerosis. Nature 2002;420:868-74. 2. Xu W, Takahashi Y, Sakashita T, Iwasaki T, Hattori H, Yoshimoto T. Low density lipoprotein receptor-related protein is required for macrophage-mediated oxidation of low density lipoprotein by 12/15lipoxygenase.The Journal Of Biological Chemistry 2001;276:36454-59. 3. Berliner J A, Navab M, Fogelman A M, Frank J S, Demer L L, Edwards P A, Watson A D, Lusis A J. Atherosclerosis: basic mechanisms. Oxidation, inflammation, and genetics. Circulation 1995;91:2488-96. 4. Hegyi L, Skepper J N, Cary N R, Mitchinson M J. Foam cell. apoptosis and the development of the lipid core of human atherosclerosis. J Pathol 1996;180(4):423-29. 5. Hengartner M O. The biochemistry of apoptosis. Nature 2000;407:770-75. 6. Kerr J F R, Stephenson J R, Heisterkamp N. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer 1972;26:239-57. 7. Zorning M, Hueber A O, Evan G. Apoptosis regulators and their role in tumorigensis. Biochemica et Biophysica Acta. 2001;1551:1-37. 8. Yu X H, Perdue T D, Tones A M, Mitochondrial involvement in tracheary element programmed cell death. Cell Death and Differentiation 2002;9:189-98. 9. Ricky W, Johnstone A, Astrid A, Apoptosis: A link between cancer 59.
(61) genetics and chemotherapy. Cell 2002;108:153-64. 10. Antonsson B, Martinou J C. The Bcl-2 protein family. Experimental cell research 2000;256:50-57. 11. Adams J M, Cory S. The Bcl-2 protein family: arbiters of cell survival. Science 1998;281:1322-26. 12. Chang S T, Miles P G. Recent trend in world production of cultivated mushrooms. Mushroom J 1991;503:15-18. 13. Eyal J. Mushroom mycelium grown in submerged culture-potential food applications. Biotechnology and food ingredients 1991;31-64. 14. Mizuno T. Bioactivity biomolecules of mushrooms: food function and medicinal effect of mushroom fungi. Food Rev 1995;11:7-21. 15. Misaki A M, Nasu M, Sone Y, Kishida E, Kinoshita C. Comparison of structure and antitumor activity of polysacchrides isolated from Fukurotake, the fruiting body of Volvariella volvacea. Agric Biol Chem 1986;50:2171-83. 16. Wasser S P, Weis A L. Therapeutic effects of substances occurring in higher basidiomycetes mushrooms: a modern perspective. Immunol 1999;19:65-96. 17. Jong S C, Birmiminghum J M. Medicinal and therapeutic value of the shiitake mushroom. Appl Microbiol 1993;39:152-84. 18. Kim W G, Lee I K, Kim J P, Ryoo I J, Koshino H, Yoo I D. New indole derivatives with free radical scavenging activity from Agrocybe Cylindracea. J Nat Prod 1997;60:721-23. 19. Liu F V, Ooi E C, Chang S T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci 1997;60(10):763-71. 20. Mizuno T, Saito H, Nishitobai T, Kawagishi H. Antitumor-active 60.
(62) substances from mushrooms. Food Rev Int 1995;11(1):23-61. 21. Ying W, Wang S J, Mo S Y, Shuai L, Yang Y C, Shi J G. Phelligridimer A, a highly oxygenated and unsaturated 26-Metabolite macrocyclic metabolite with antioxidant activity from the fungus Phellinus igniarius. Organic Letters 2005;7(21):4733-36. 22. Ying W, Wang S J, Mo S Y, Shuai L, Yang Y C, Shi J G. A unique highly oxygenated pyrano[4,3-c] [2] benzopyran and cytotoxic activities from the fungus Phellinus igniarius. Organic Letters 2005;7(9):1675-78. 23. Naoki I, Tomonori S, Tomokazu L, Tomohiro S, Hiroyuki T, Tomoyuki N, Yukihito A, hirokazu K, Hiroichi N. Inhibition of IgG-dependent mouse triphasic cutaneous reaction by a boiling water fraction separated from mycelium of Phellinus linteus. eCAM 2005;2(3):369-74. 24. Song K S, Cho S M, Lee J H, Kim H M, Han SB, Ko K S, Yoo I D.B-lymphocyte-stimulating polysaccharide from mushroom Phellinus linteus. Chem Pharm Bull 1995;43:2105–8. 25. Kim H M, Han S B, Oh G T, Kim Y H, Hong N D, Ypp I D. Stimulation of humoral and cell mediated immunity by polysaccharide from mushroom Phellinus linteus. Int J Immunopharmacol 1996;18:295-303. 26. Kim GY, Oh Y H, Park Y M. Acidic polysaccharide isolated from Phellinus linteus induces nitric oxide-mediated tumoricidal activity of macrophages through protein tyrosine kinase and protein kinase C. Biochem Biophys Res Commun 2003;309:399-407. 27. Park S K, Kim G Y, Lim J Y, Kwal J Y, Bae Y S, Lee J D, Oh Y H, 61.
(63) Ahn S C, Park Y M. Acidic polysaccharides isolated from Phellinus linteus induce phenotypic and functional maturation of murine dendritic cells. Biochem Biophys Res Commun 2003;3012:449-58. 28. Kim S H, Song Y S, Kim S K, Kim B C, Lim C J, Park F H. Anti-inflammatory and related pharmacological activities of the n-BuOH subfraction of mushroom Phellinus linteus. J Ethnopharmacol 2004;93:141-6. 29. Kim GY, Kim S H, Hwang S Y, Kim H Y, Park Y M, Park S K, Lee M K, Lee S H, Lee T H, Lee J D. Oral administration of proteoglycan isolated from Phellinus linteus in the prevention and treatment of collagen-induced arthritis in mice. Biol Pharm Bull 2003;26:823-31. 30. Lim B O, Kim S H, Hwang S Y, Kim H Y, Park Y M, Park S K, Lee M K, Lee S H, Lee T H, Lee J D. Comparative study on the modulation of IgE and cytokine production by Phellinus linteus grown on germinated brown rice, Phellinus linteus and germinated brown rice in murine splenocytes. Biosci Biotechnol Biochem 2004;68:2391–4. 31. Jae S B, Kwang H J, Hee K J. Polysaccharides isolated from Phellinus gilvus enhances dermal wound healing in streptozocin-induced diabetic rats. J Vet Sci 2005;6(2):161-64. 32. Jae S B, Kwang H J, Hee K J. Effects of natural polysaccharides on the growth and peritoneal carcinomatosis of human gastric adenocarcinoma in a nude mose model. Cancer Letters 2005;(5):1-9. 33. Hye J H, Kim S W, Lim J M, Joo J H, Kim H O, Kim H M, Yun J W. Hypoglycemic effect of crude exopolysaccharides produced by a medicinal mushroom Phellinus baumii in streptozotocin-induced 62.
(64) diabetic rats. Life Sciences 2005;76:3069-80. 34. Bae J S, Jang K H, Park S C, Jin H K. Promotion of dermal wound healing by polysaccharides isolated from Phellinus gilvus in rats. Laboratory Animal Science 2005;67(1):111-114. 35. Hyo J L, Lee H J, Lim E S, Ahn K S, Shim BS, Kim H M, Gong S J, Kim S H. Cambodian Phellinus liteus inhibits experimental metastasis of melanoma cells in mice via regulation of urokinase type plasminogen activator. Biol Pharm Bull 2005;28(1):27-31. 36. Soo B C, Chun H P, Mi K C, Dong W J, Sunmin P. Improvement of insulin resistance and secretion by water extracts of cordyceps militaris, Phellinus linteus, and paecilomyces tenuipes in 90% pancreatectomized rats. Biosci Biotechnol Biochem 2004;68(11):2257-64. 37. Kim G Y, Oh W K, Shiin B C, Shin Y I, Park Y C, Ahn S C, Lee JD, Bae Y S, Kwark J Y, Park Y M. Proteogycan isolated from Phellinus linteus inhibits tumor growth through mechanisms leading to an activation of CD11c CD8 DC and type I helper T cell-dominant immune state. FEBS Letters 2004;576:391-400. 38. Li G, Kim d H, Kin T D, Park B J, Park H. D, Park J I, Na M K, Kim. H C, Hang N D, Lim K, Hwang B D, Yoon W H. Protein-bound polysaccharide from Phellinus linteus induces G2/M phase arrest and apoptosis in SW480 human colon cancer cells. Cancer Letters 2004;216:175-181. 39. Kim G Y, Han M G, Song Y S, Shin B C, Shin Y I, Lee H J, Moon D O, Lee C M, Kwak J Y, Bae Y S, Lee J O, Park Y M. Proteoglycan isolated from Phellinus linteus toll-like receptors 2- and 4- maturation 63.
(65) of murine dendritic cells via activation of ERK, p38, and NF-κB. Biol Pharm 2004;27(10):1656-62. 40. Kim G Y, Choi G S, Lee S H, Park Y M. Acidic polysaccharide isolated from Phellinus linteus enhances through the up-regulation of nitric oxide and tumor necrosis factor-α from peritoneal macrophages. Journal of Ethnopharmacology 2004;95:69-76. 41. Ajith T A, Janardhanan K K. Cytotoxic and antitumor activities of a polypore macrofungus, Phellinus rimosus (Berk) pilat. Journal of Ethnopharmacology 2003;84:157-62. 42. Ajith T A, Janardhanan K K. Antioxidant and antihepatotoxic activities of Phellinus rimosus (Berk) pilat. Journal of Ethnopharmacology 2002;81:387-91. 43.Cho J H, Cho S D, Hn H, Kim S H, Lee S K, Lee Y S, Kang K S. The roles of ERK1/2 and p38 MAPK kinases in the preventive mechanisms of mushroom Phellinus liteus against the inhibition of gap junctional intercellular communication by hydrogen peroxide. Carcinogenesis 2002;23(7):1163-69. 44.Han S B, Lee C W, Jeon Y J, Hong N D, Yoo I D, Yang K H, Kin H M. The inhibitory effect of polysaccharides isolated from Phellinus linteus on tumor growth and metastasis. Immunopharmacology 1999;41:157-164. 45. Cominacini L, Rigoni A, Pasini A F, Garbin L, Daoli A, Campagnola M, Pastorino A M, Lo Cascio V, Sawamura J. The binding of oxidized low density lipoprotein (ox-LDL) to ox-LDL receptor-1 reuces the intracellar concentration of nitric oxide in endothelial cells through an 64.
數據

相關文件
Keratinizing squamous cell carcinoma composed of recognizable squamous tumor cells with pronounced keratinization (B) (original magnification ×200).6. Dark blue punctate (dotlike,
Aim: This study examined and classified patients who were treated at the Faculty of Dentistry at Ankara University Dentistry to determine the incidence of different types of
Giant cells are also seen in other diseases like giant cell granuloma of the jaws, traumatic bone cyst, aneurysmal bone cyst, and jaw tumor of
stained negative for a-smooth muscle actin because the main body of the tumor consisted of epithelioid or epithe- lial cells that contained few positive smooth muscle cells.. We
Stain (-) because the main body of the tumor consisted of epithelioid or epithelial cells that contained few positive smooth muscle cells. No clear ductal structures or
A9 Median employment earnings of the employed population in July 2016 by statistical district A10 Non-Macao born land-based population by gender, age group and statistical
The majority (4,075 units valued at MOP9.2 billion) of these transactions were residential units that accounted for 55.5% of the total number of building units; besides, there were
The prepared nanostructured titania were applied for the photoanodes of dye-sensitized solar cell.. The photoanodes were prepared by the doctor blade technique and the area
