異戊烯轉移酶(ipt)基因轉殖青花菜與近緣亞種作物
雜交潛力及後代之性狀探討
丁柏瑜
1、陳烈夫
2、陳榮芳
3、杜元凱
4,*
1亞洲大學保健營養生技學系及綠寶生物科技有限公司
2行政院農委會農業試驗所技術服務組
3中央研究院植物暨微生物學研究所
4行政院農委會農業試驗所生物技術組
摘要
基因轉殖作物之基因流佈可能造成轉殖
基因雜交後代植株競爭優勢,為生態安全評
估 主 要 議 題 之 ㄧ 。 本 研 究 異 戊 烯 轉 移 酶
(isopentenyl transferase; ipt)基因轉殖青花
菜
103 轉殖系與 7 種近緣亞種作物作為花粉
接受親進行田間自然雜交潛力評估。田間自
然雜交率範圍在
09%,屬於種間雜交之小白
菜及小油菜皆未發現雜交現象,證實種內基
因流佈存在。為進一步探討轉殖株雜交後代
植株之競爭能力,乃於溫室利用基因轉殖
ipt
青花菜與自交系
104 (CK)為花粉貢獻親,分
別 與 近 緣 亞 種 作 物 進 行 人 工 雜 交 。 整 體 而
言,F
1後代植株性狀以
103 轉殖系雜交 F
1後
代株高較高,多數性狀仍在
104 (CK)雜交 F
1後代、近緣亞種及
104 (CK)範圍內,但每果
莢種子數顯著高於
104 (CK)雜交 F
1後代及雜
交親本,顯示基因轉殖
ipt 青花菜雜交近緣亞
種
F
1後代仍具有相當的雜交優勢。
關鍵詞︰雜交親和力、生長勢、雜種優勢、
花粉活性、生物安全評估。
Study on Hybridization Potential
between Transgenic Isopentenyl
Transferase (ipt) Broccoli (Brassica
oleracea var. italica) and Related
Subspecies and Comparison of
Horticultural Traits for the Hybrid
Offspring
Po-Yu Ting
1, Lit-Fu Chan
2, Long-Fang
O. Chen
3and Yuan-Kai Tu
4,*
1 Department of Health and Nutrition Biotechnology, Asia University and Green Global State Biotech LTD., Wufeng District, Taichung City 41354, Taiwan ROC
2 Technology Service Division, Taiwan Agricultural Research Institute, Wufeng District, Taichung City 41362, Taiwan ROC
3 Institute of Plant and Microbial Biology, Academia Sinica, Nankang, Taipei 11529, Taiwan ROC
4 Biotechnology Division, Taiwan Agricultural Research Institute, Wufeng District, Taichung City 41362, Taiwan ROC
ABSTRACT
Genetically modified crops may cause transgene distribution of hybrid plants. Issues about the ecological security assessment become more and more important. In this study, broccoli (Brassica oleracea var. italica) transformed with the ipt (isopentenyl transferase) gene, designated as line 103 was used to investigate hybridization extent by pollination with seven closely related varieties. The range of fertility rates followed by nature pollination were 09%, while no hybridization was found for ‘Pakchoi’ and ‘Rapa’
* 通信作者, [email protected] 投稿日期:2014 年 4 月 29 日 接受日期:2014 年 6 月 6 日
作物、環境與生物資訊 11:88-104 (2014)
Crop, Environment & Bioinformatics 11:88-104 (2014) 189 Chung-Cheng Rd., Wufeng, Taichung 41362, Taiwan ROC
as pollen recipients. Results showed that transgenic DNA insertion did not alter potential risk for outcross among broccoli varieties. In order to study the competitive advantage of F1 progenies bred from transgenic ipt broccoli crossing with four closely related varieties, transgenic line 103 and 104 (CK) inbred line were used as pollen donor and hybridized with related varieties by artificial pollination. Results of transgenic ipt F1 showed that plant height was higher than that of 104 hybrid F1. Seed numbers per pod of transgenic ipt F1 were higher than those of 104 hybrid F1, parents and 104 (CK). Results indicated that F1 progeny from transgenic broccoli hybrid exhibited the phenomenon of heterosis.
Key words: Cross compatibility, Growth vigor, Heterosis, Pollen viability, Biosafety assessment.
前言
近 年 來 , 基 因 轉 殖 作 物 在 全 球 大 量 種
植,卻也帶來生態風險可能性。主要風險之
一,係抗性基因透過花粉流佈
(pollen flow)
轉移到野生近緣植物,導致這些植物產生抗
性,從而形成「超級雜草」
。因此,在基因轉
殖作物商品化生產之前,有必要對基因轉殖
作物與近緣植物間潛在的基因流佈進行評估
(Wang et al. 2011)。青花菜(Brassica oleracea
var. italica Plenck)屬十字花科(Cruciferae)蕓
薹屬(Brassica)作物,在自然界中構成蕓薹屬
的 三 種 基 本 種 群
(species) , 分 別 為 小 油 菜
Brassica campestris (n = 10, 染色體組 AA)、
黑芥
B. nigra (n = 8, 染色體組 BB)及甘藍 B.
oleracea (n = 9, 染色體組 CC)。其後,又經過
生物種類間自然雜交及長時間演化,衍生出
三類共存的野生種群,包括小油菜與黑芥雜
交經染色體加倍而成新物種芥菜
B. juncea (n
= 18, AABB)、小油菜與甘藍雜交經染色體加
倍而成新物種蕪菁甘藍或大油菜
B. napus (n
= 19, AACC)及黑芥與甘藍雜交經染色體加
倍而成新物種阿比西尼亞芥子
B. carinata (n
= 17, BBCC),且與其相關的近緣種作物在生
物學上具有雜交親和力
(cross compatibility)
(Prakash and Hinata 1980, Roy 1980,
Scheffler and Dale 1994, Li et al. 2012)。
關於十字花科作物基因流佈之研究報告
甚多,主要以甘藍型油菜(B. napus L.)與白菜
型油菜(B. rapa L.)等同屬作物,以及與基因轉
殖 抗 除 草 劑 油 菜 與 野 生 蘿 蔔
(Raphanus
raphanistrum L.)、野芥(Sinapis arvensis L.)、
白芥(S. alba L.)、黑芥(Hirschfeldia incana L.)
等異屬植物間均可進行雜交(FitzJohn et al.
2007, Li et al. 2008, Mitsuko et al. 2011)。其
中 , 基 因 轉 殖 抗 除 草 劑 基 因 甘 藍 型 油 菜
(B.
napus)在自然情況下亦容易與 B. rapa 及 B.
juncea 兩 近 緣 種 雜 交 (Jørgensen et al.
1996)。Metz et al. (1997)亦獲有相同的結果,
且在
F
1代與
B. rapa 回交 4 代後,仍能檢測到
抗除草劑植株,這也顯示出外源基因能夠以
一 定 的 頻 率 從
B. napus 的 基因轉移 至 B.
rapa。Heinrich and Antje (2012)研究指出由
於油菜種子採收後其殘留於田間之種子發芽
演變成實生苗(volunteer)或進入休眠狀態,
透過耕作深埋種子,處在黑暗及乾旱環境中
亦能長時間存活。Beckie et al. (2006)指出加
拿大之抗除草劑基因轉殖油菜之種子可存活
達
10 年,並演變成實生苗,增加其轉殖基因
油菜植株適應性,故實生苗極可能與野生族
群產生基因流佈。據此進而推論,可產生花
球之花椰菜、青花菜的種子特性亦易受基因
轉殖影響而改變,在植株生長過程至花球採
收前,或種子採收後之殘株,其生長情形將
可 能 造 成 植 株 野 化 或 再 次 與 近 緣 種 作 物 雜
交,而造成基因流佈之可能。
然而,基因轉殖植物能否成功地流佈到
近 緣 種 , 取 決 於 兩 個 層 面
(Chapman and
Burke 2006, Mercer et al. 2007):(1)基因轉殖
作 物 和 近 緣 種 的 親 和 性
(sexual compati-
bility),此為決定目標基因能否轉移到野生近
緣種的首要條件;(2)雜交後代與回交代的適
合度(fitness)是否下降,適合度主要受到親本
的 基 因 型
(Campbell and Waser 2001,
Gueritaine et al. 2002) 、 測 試 環 境 條 件
(Campbell and Snow 2007, Vacher et al.
2004)及兩者間相互作用(Johannessen et al.
2006, Whitney et al. 2006)之影響。Mercer et
al. (2007)發現栽培型向日葵與野生向日葵的
雜交種,在逆境下的適合度會增加。
Cao et al.
(2009)發現 2 個抗蟲基因轉殖水稻與 3 個野生
稻種植田間,其雜交之
F
1後代在株高、分蘖、
穗數、千粒重等性狀皆較親本野生稻為高,
說明了基因轉殖雜交後代相較於親本具有較
高競爭能力。當轉入具有更強適應性的基因
時,藉由基因流佈使得親本或近緣種作物具
有 更 強 的 生 存 競 爭 能 力
(Andow and
Claudia 2006, Mikkelsen et al. 1996,
Mohammad 2011)。
目前已有許多研究報告,發現基因轉殖
ipt 植物不但能延遲植株老化且較具抗逆性
(Adhikary et al. 2010, Liu et al. 2011,
Merewitz et al. 2011)。因此,本研究探討基
因轉殖
ipt 青花菜與其受體青花菜(綠王品種)
及其近緣亞種作物在田間自然雜交,以分析
插入
ipt 基因對青花菜異交潛力的影響。又為
了進一步瞭解基因轉殖
ipt 青花菜雜交後代
植株之競爭能力,乃於溫室評估同時帶有
ipt
與
nptII 基因雜交後代之花粉活性、雜交結實
率、種子數及植株性狀之差異性,以此作為
野化能力評估指標。
材料與方法
試驗材料
本研究係以自交五代(T
5)園藝性狀優良
且具延遲花球黃化之中生優良轉殖青花菜自
交系
(103 轉殖系)與對照組「綠王」自交系 104
(CK)、青花菜商業品種「綠王」及其他 5 種
近緣亞種作物,分別為芥藍菜(B. oleracea var.
alboglabra)之白花芥藍「富悅」(Fu Yue; FY)
與黃花芥藍「白格林」
(Bai Ge Lin; BGL)、
花椰菜(B. oleracea var. botrytis)之「麗雪」(Li
Syue; LS)、小白菜(B. rapa var. chinensis)及小
油菜(B. rapa),作為試驗材料,以檢測其自交
系間、品種間及種間之雜交能力。
田間基因轉殖青花菜與近緣亞種作物自
然雜交試驗
試驗於
2007-2008 年在行政院農委會農
業試驗所隔離設施內進行。由於青花菜屬性
喜冷,故採秋作種植,即於每年
11 月初在隔
離溫室育苗,種子先播種於穴盤,經培育
3
週當本葉達
4 片葉時,定植於隔離試驗田。
田間配置採逢機完全區集設計
(randomized
complete block design; RCBD),將基因轉殖
青花菜自交系
(103 品系)種植於田區中央,共
600 株,田間四周設置 4 區集,各區集逢機種
植
2 種非 GM 青花菜及 5 種近緣亞種作物,
每重複小區畦長
10 m 及寬 1.5 m (含畦溝寬
度
),株距 45 cm、行距 60 cm,各重複種植
15 株。試驗期間記錄各參試材料的始花期及
終花期所需天數,並調查各品種(系)間之花期
重疊日數。同時,青花菜在田間進行自然授
粉,於受體材料成熟後,按品種(系)收穫小區
之種子,若每株
1,000 粒以下時取一半種子,
1,000 粒以上時取 1/3 量種子(95%信賴水準
下最大取樣誤差為
±1.9%)再進行播種。播種
培育後,利用
PCR 分析對幼苗葉片進行 DNA
檢測,其檢測
ipt 及 nptII 基因之 PCR 方法可
參考
Chan et al. (2009),再根據 PCR 檢測結
果具有外來
ipt 或 nptII 之轉殖株估算其雜交
頻率,計算公式如下(Funk et al. 2006):
雜交頻率(%) = (含轉殖基因株數/總檢測
株數
) × 100
溫室基因轉殖青花菜及非基因轉殖青花
菜分別與近緣亞種作物雜交
隔離溫室所使用的試驗材料與田間試驗
相同,係於隔離溫室育苗,種子先播種於穴
盤(4.5 cm × 4.5 cm × 5 cm),經培育 3 wk 當
本葉達
4 片葉時,移植至盆栽(26 cm × 29 cm
× 45 cm)。盆內栽培介質混合複合肥料一號
(台灣肥料股份有限公司出品),其施用量為
N:P
2O
5:K
2O = 270:150:200 kg
-1·ha,以
每隔
20 d 分 4 次施用,其間並送至人工光源
室進行為期
1 mo.的春化處理(日/夜溫為 20
℃/10℃),待至抽苔時再分別移入隔離溫室
進行人工授粉試驗,每一品種(系) 4 重複。
自交五代(T5)園藝性狀優良且具延遲花
球黃化之中生優良轉殖青花菜自交系(103 品
系)與對照組「綠王」自交系 104(CK)為花粉
貢獻親,與前述
4 種近緣亞種作物為花粉接
受親(pollen recipient)進行人工雜交,並獲得
F
1後代作為試驗材料。經
PCR 檢測證實 103
轉殖系為花粉貢獻親與近緣亞種雜交之
F
1植
株,均含有
ipt 及 nptII 基因,進行以下雜交
F
1後代之性狀探討。
雜交
F
1後代植株園藝性狀調查
試材於溫室育苗,將小苗移植至盆栽定
植
45 d 後,將檢測帶有 ipt 及 nptII 基因之雜
交 後 代 調 查 其 植 株 性 狀 , 包 括 株 高
(plant
height) 、 株 寬 (plant width) 、 莖 長 (stem
height)、 莖 寬 (stem width)、 柄 長 (petiole
length) 、 柄 寬 (petiole width) 、 葉 長 (leaf
length) 、 葉 寬 (leaf width) 與 葉 數 (leaf
number)等性狀,每供試品系 3 重複,每一
重複調查
5 株。
雜交
F
1後代花球性狀調查
1. 乙烯釋放量之測定
將 採 收 後 花 球 放 入
350 mL 之樣品瓶
中,每一樣品瓶
3 顆花球,在 25℃下密封 2
h,以注射針筒抽取 1 mL 之氣體,利用氣相
層 析 儀
(Gas chromatograph, VARIAN
430-GC, USA)進行乙烯釋放量之測定。其偵
測 器 為 火 焰 離 子 式
(Flame Ionization
Detector; FID),注射孔(injector: 1177)溫度
為
200℃,層析管柱為毛細管柱(層析管柱長
27.5 m、管柱內徑 0.53 mm、管柱外徑 0.7
mm)層析管柱溫度為 40℃,其載流氣體為氮
氣。乙烯釋放量單位以
μL·g
-1·h
-1表示。每一
品系(種) 4 重複。
2. 色差儀之檢測:
利用色差儀測定雜交
F
1及親本花球之
L、a、b 值(Σ80 Color Measuring System,
Nippon Denshoku Ind. Co., LTD., Japan)。
色差儀將顏色分為三個參數,L 值代表明亮
度,範圍
0100,愈近 0 表示樣品愈近黑色,
愈近
100 表示樣品愈接白色;a 值為色相,顏
色為紅-綠色,a 值愈接近正數表示紅色,負
數表示綠色,愈接近+100 愈紅,愈接近-100
愈綠;b 值代表黃-藍色,b
色相顏色為藍色-黃色,愈接近正數表示黃色,負數表示藍色,
愈接近+100 愈黃,愈接近-100 則愈藍。
雜交
F
1後代花粉活性、花粉性狀及果莢
結實率調查
1. 花粉活力檢定
在近緣亞種之雜交後代花粉活性檢測,
利用螢光染色法(fluorescent diacetate; FDA)
(Heslop-Harrison and Heslop-Harrison
1970)及 TTC 法(2,3,5-triphenyltetrazolium
chloride; TTC) (Norton 1966)進行花粉活力
檢定,每一雜交
F
1後代及雜交親本為
4 重
複,每一重複取
4 朵花測定其活性。
2. 近緣亞種雜交 F
1之花粉量與花粉粒大小調查
依據
Snow et al. (2010)之方法將花藥放
入
1.5 mL 離心管內,加入 100 mL ddH
2O,
震盪充分混均後,於顯微鏡底下計數其朵花
之花粉量與花粉粒直徑。每一雜交
F
1後代及
雜交親本為
4 重複,測量花粉量取樣方式為
每一重複取
4 朵花;而花粉粒大小為每一重
複取
4 朵花,每一朵花調查 10 粒花粉。
3. 調查果莢結實率
每品系(種) 4 重複,每重複授粉 15 朵
花,至果莢成熟後採收並計算之。果莢結實
率(%) = (結莢數 / 授粉花朵數) × 100
統計分析
雜交
F
1後代與親本間在植株生育性狀、
花 球 性 狀 及 花 粉 活 性 測 定 的 差 異 顯 著 性 測
驗 , 係 利 用
SAS EG 統 計 分 析 軟 體 (SAS
Institute, Inc. 2006)進行變方分析(analysis
of variance; ANOVA) 及 最 小 顯 著 差 異 性
(least significant difference; LSD)測驗。
結果
田間自然雜交試驗
以轉殖
103 品系為父本,「綠王」、104
(CK)、「富悅」、「麗雪」、「白格林」、小白菜
及小油菜為母本,進行田間自然雜交後,將
收穫種子催芽播種。小苗經
PCR 檢測後,後
代只含有
ipt 或 nptII 單一基因或同時帶有此
兩個基因三種現象,大部分雜交
F
1後代仍同
時含有
ipt 及 nptII 基因(Table 1)。合併 2007
與
2008 年田間自然雜交結果進行綜合變方分
析前,先以
F 檢定進行年度均方同質性測
驗,結果差異顯著(F 值 = 3.008*),表示年度
均方並非同質,不符合綜合變方分析之使用
前提,故乃進行各年度變方分析。結果顯示
2007 年品種間雜交頻率達顯著差異(Table
1),經 LSD 測試結果,以花椰菜-「麗雪」、
青花菜-「綠王」及自交系 104 (CK)之雜交頻
率無差異,最低為黃花芥藍-「白格林」。試
驗兩年期間除了白花芥藍
-「富悅」在 2007
年無雜交現象,2008 年發生 2.42%雜交率之
外,小白菜及小油菜皆未發現種間雜交現象。
雜交
F
1後代植株園藝性狀調查
利用
T
5代之
103 轉殖系及 104 (CK)作為
花粉貢獻親,與近緣亞種芥藍類(「富悅」與
「白格林」、「綠王」商業品種及花椰菜「麗
雪」進行雜交,以評估轉殖株或其
F
1後代植
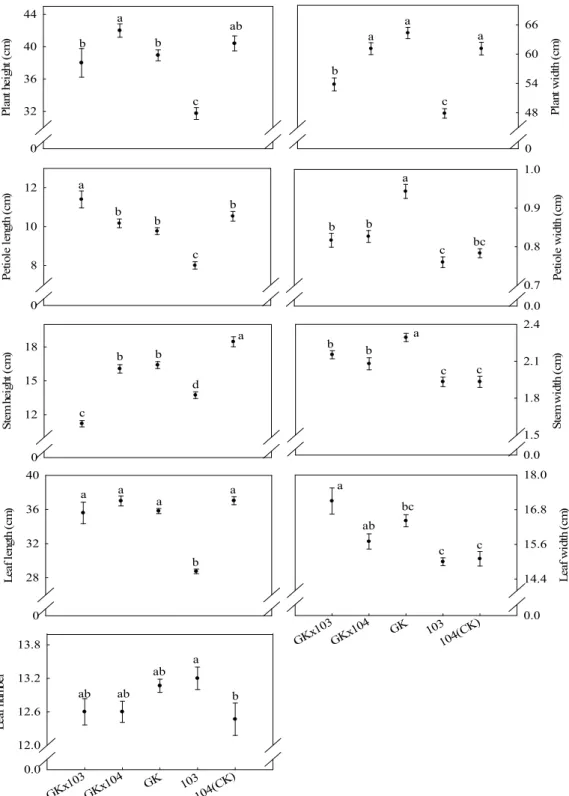
株性狀之變異性。結果顯示,
104 (CK)之植
株性狀變異係數(CV 值)均較 103 轉殖系大,
即
103 轉殖系植株性狀較為穩定(Table 2)。
經
103 或 104 (CK)雜交所雜交近緣亞種 F
1後
代,大部份植株變異係數大於雜交親本。然而,
「白格林」 × 103 雜交 F
1後代之株高、莖長、
葉長和葉寬,以及「綠王」
× 103 雜交 F
1後代
之株高、株寬、葉長和葉寬,其等性狀變異係
數大於雜交親本或
104 雜交後代,「富悅」 ×
103 和「麗雪」 × 103 大部分性狀變異程度小
於
104 雜交後代(Table 2)。
由於
103 轉殖系本身株高較 104 (CK)
矮,與
4 個近緣亞種作物相較,除了「富悅」
株高顯著高於
F
1雜交後代及青花菜品系[103
轉殖系及
104 (CK)]外,以 104 (CK)為花粉貢
獻親所雜交「白格林」、「綠王」及「麗雪」
之
F
1後代株高為最高,顯示
103 轉殖系為花
粉貢獻親所雜交
F
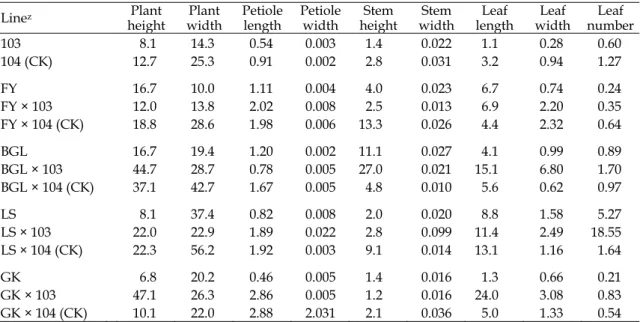
1後代未高於親本(Fig. 1)。
Table 1. Hybrid frequency and distribution ratio between transgenic ipt broccoli and seven close relativessubspecies by natural pollination from 2007 to 2008.
Variety (line)
2007 2008 Distribution ratio Distribution ratio
Hfz nptII ipt n+i Hf nptII ipt n+i
(%) (%) (%) (%) (%) (%) (%) (%)
B. oleracea var. italica–Green King 6.4±1.8 ay 0.9 1.2 4.2 7.5±1.0 a 0.2 1.0 6.2
B. oleracea var.italica– 104(CK) 2.1±1.5 ab 0.2 0.1 1.9 4.9±1.4 a 0 0.4 4.6
B. oleracea var. alboglabra– Fu Yue 0.0±0.0 b 0 0 0 2.4±0.3 a 0 0 2.3
B. oleracea var. botrytis– Li Syue 9.0±4.8 a 1.4 1.6 6.0 6.1±2.9 a 0.2 0.6 5.3
B. oleracea var. alboglabra– Bai Ge Lin 1.1±0.5 b 0.7 0.1 0.3 2.9±0.3 a 0 0.2 2.7
Brassica rapa –Edible rape 0.0±0.0 0 0 0 0.0±0.0 0 0 0
Brassica rapa var. chinensis – Pakchoi 0.0±0.0 0 0 0 0.0±0.0 0 0 0
z Hf: Hybrid frequency; nptII: Neomycin phosphotransferase II (kanamycin resistance); ipt: isopentenyltransferase; n+i: nptII+ipt.
y Mean±standard error (n = 4); the data of edible rape and pakchoi values are zero for both years.
Means within each column followed by the same letters are not significantly different at 5% level by LSD test. All data was square-root transformed prior to analysis.
Table 2. Comparison of the coefficient of variation (%) among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli crossed with four closely related varieties.
Linez Plant
height width Plant Petiolelength Petiolewidth heightStem widthStem length Leaf width Leaf numberLeaf
103 8.1 14.3 0.54 0.003 1.4 0.022 1.1 0.28 0.60 104 (CK) 12.7 25.3 0.91 0.002 2.8 0.031 3.2 0.94 1.27 FY 16.7 10.0 1.11 0.004 4.0 0.023 6.7 0.74 0.24 FY × 103 12.0 13.8 2.02 0.008 2.5 0.013 6.9 2.20 0.35 FY × 104 (CK) 18.8 28.6 1.98 0.006 13.3 0.026 4.4 2.32 0.64 BGL 16.7 19.4 1.20 0.002 11.1 0.027 4.1 0.99 0.89 BGL × 103 44.7 28.7 0.78 0.005 27.0 0.021 15.1 6.80 1.70 BGL × 104 (CK) 37.1 42.7 1.67 0.005 4.8 0.010 5.6 0.62 0.97 LS 8.1 37.4 0.82 0.008 2.0 0.020 8.8 1.58 5.27 LS × 103 22.0 22.9 1.89 0.022 2.8 0.099 11.4 2.49 18.55 LS × 104 (CK) 22.3 56.2 1.92 0.003 9.1 0.014 13.1 1.16 1.64 GK 6.8 20.2 0.46 0.005 1.4 0.016 1.3 0.66 0.21 GK × 103 47.1 26.3 2.86 0.005 1.2 0.016 24.0 3.08 0.83 GK × 104 (CK) 10.1 22.0 2.88 2.031 2.1 0.036 5.0 1.33 0.54
z 103: transgenic ipt inbreed line; 104 (CK): nontransgenic inbreed line; BGL: B. oleracea var. alboglabra– Bai Ge Lin; FY: B. oleracea var. alboglabra– Fu Yue; LS: B. oleracea var. botrytis– Li Syue; GK: B. oleracea var. italica– Green King.
以
103 轉殖系雜交近緣亞種 F
1後代性狀
優於親本、非轉殖雜交
F
1後代或
104 (CK)者
為「富悅」
× 103 之柄長和莖寬性狀,但與
「富悅」、
104 (CK)或「富悅」 × 104 (CK)
無顯著差異。除葉數外,大部份性狀仍以「富
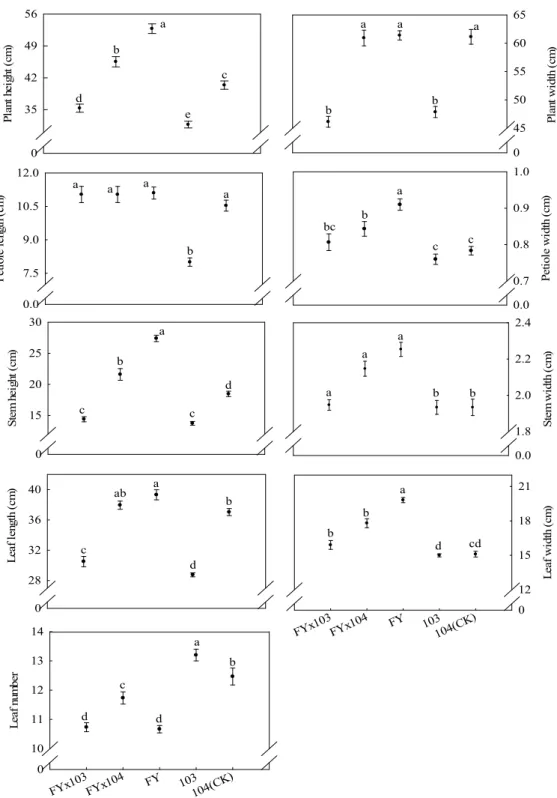
悅」白花芥藍較佳(Fig. 1)。「白格林」 × 103
之
F
1後代葉長、葉寬及葉數性狀雖優於非轉
殖系
F
1後代及
104 (CK),葉長與葉寬卻與「白
格林」親本無顯著差異(Fig. 2)。
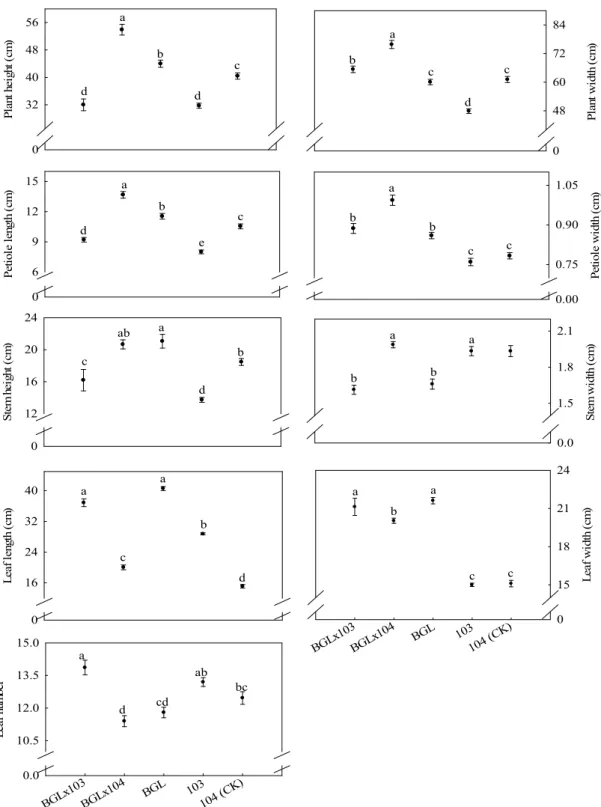
「麗雪」
× 103
性狀僅葉數性狀較
103 轉殖系、
「麗雪」
× 104
或
104 (CK)佳,但無優於「麗雪」(Fig. 3)。
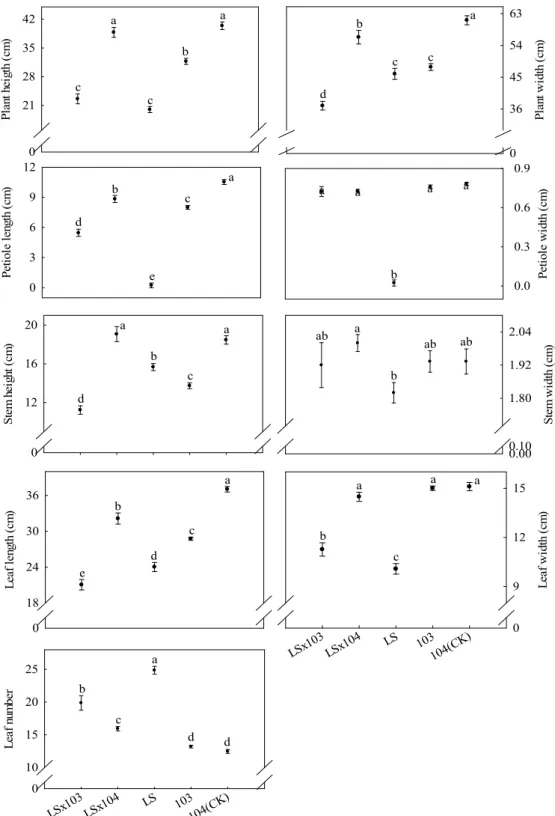
「綠
王」
× 103 之柄長及葉寬顯著高於「綠王」 ×
104、104 (CK)、103 轉殖系及「綠王」(Fig. 4)。
綜合
F
1後代性狀評估,103 轉殖系雜交 F
1後
代相較於親本,多數園藝性狀改變甚微。
雜交
F
1後代花球生理變化
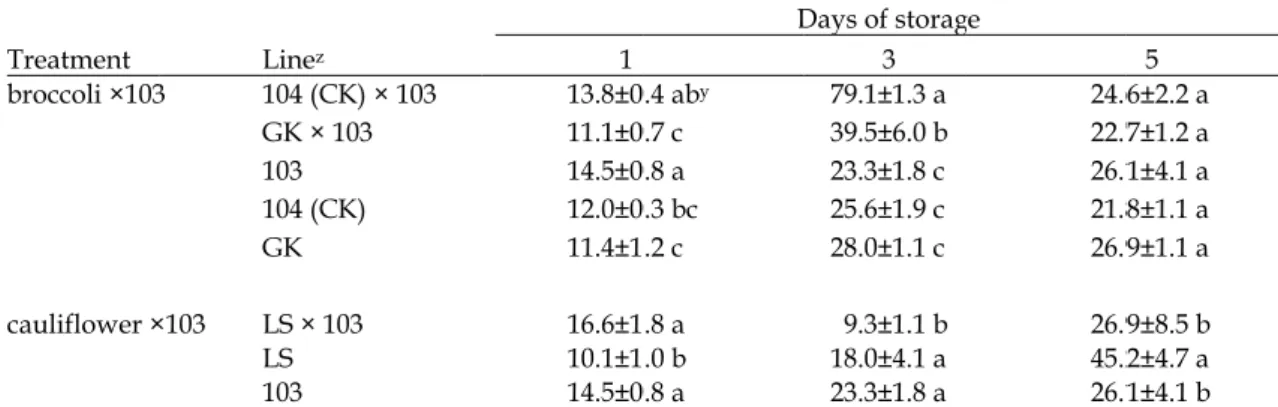
1. 乙烯生成量
採收後貯藏於
25±2℃室溫下 1 d 之青花
菜花球尚未黃化,
103 轉殖系乙烯生成量已顯
著高於
103 雜交之 F
1後代及非基因轉殖青花
菜[「綠王」及 104 (CK)];在第 3 天花球開始
黃化,以
104 (CK) × 103 雜交種之黃化面積
較其他雜交後代多,此時乙烯生成量顯著高
於其他品系及親本,其次為「綠王」 × 103,
而
103 轉殖系及非基因轉殖青花菜[「綠王」
及
104 (CK)]為最低;至第 5 天 103 雜交之 F
1後代及親本花球的黃化面積無顯著差異。又
花椰菜-「麗雪」乙烯生成量顯著高於「麗雪」
× 103 與 103 轉殖系,但「麗雪」 × 103 後代
花球已黃化,說明了
103 轉殖系之延遲老化
效果仍較雜交後代為佳
(Table 3 及 Fig. 5)。
2.色差
運用色差計分析雜交
F
1後代與親本花球
顏色,測定花球的
L、a、b 值, L 值代表明
亮度,愈接近
0 表示樣品愈接近黑色,愈接
近
100 表示樣品愈接近白色。Table 4 結果顯
示,
103 雜交 104 (CK)及「綠王」之 F
1後代、
非轉殖青花菜及
103 轉殖系親本,其等花球
採收當天及貯藏第
5 天時,以 103 轉殖系明
亮度顯著低於雜交
F
1後代及非轉殖青花菜;
而「麗雪」顯著高於
103 轉殖系及「麗雪」 ×
103 之 F
1後代,顏色接近白色(Table 4)。
Pl an t h ei gh t ( cm ) 0 35 42 49 56 a b c e Plan t w id th ( cm ) 0 45 50 55 60 65 b b a a a Pe tio le le ngt h ( cm ) 0.0 7.5 9.0 10.5 12.0 a a a a b Pe tiol e w idth ( cm ) 0.0 0.7 0.8 0.9 1.0 a b bc c c St em h ei gh t ( cm ) 0 15 20 25 30 a b c c d d Leaf le ng th ( cm ) 0 28 32 36 40 ab a b c d FYx10 3 FYx10 4 FY 103 104(CK ) Leaf w id th ( cm ) 0 12 15 18 21 a b b cd d FYx10 3 FYx10 4 FY 103 104(CK ) Leaf n um be r 0 10 11 12 13 14 a b c d d St em w id th ( cm ) 0.0 1.8 2.0 2.2 2.4 a a a b b
Fig. 1. Comparison of the traits among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli
crossed with Chinese kale-Fu Yue (white flower). Error bar is the standard error of mean (n = 4). Means with the same letters are not significantly different at 5% level by LSD test. LSD: least significant difference.
St em h ei ght ( cm ) 0 12 16 20 24 Pl an t he ight ( cm ) 0 32 40 48 56 a b c d d a ab b c a Pe tiol e l en gth ( cm ) 0 6 9 12 15 a b c d e Pe tiol e wi dth ( cm ) 0.00 0.75 0.90 1.05 a b b c c St em w id th ( cm ) 0.0 1.5 1.8 2.1 a b a b Pl an t w id th ( cm ) 0 48 60 72 84 a b c c d d BGLx1 03 BGLx1 04 BGL 103 104 (C K) Leaf w id th ( cm ) 0 15 18 21 24 a a b c c Le af le ng th ( cm ) 0 16 24 32 40 a a b c d BGLx1 03 BGLx1 04 BGL 103 104 (C K) Leaf n um ber 0.0 10.5 12.0 13.5 15.0 a ab bc cd d
Fig. 2. Comparison of the traits among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli
crossed with Chinese kale-Bai Ge Lin (yellow flower). Error bar is the standard error of mean (n = 4). Means with the same letters are not significantly different at 5% level by LSD test.
Pl an t h ei gth ( cm ) 0 21 28 35 42 a a b c c Pl an t w idt h ( cm ) 0 36 45 54 63 a b c c d Pe tio le le ngt h ( cm ) 0 3 6 9 12 a b c d e Petiol e w idt h (c m ) 0.0 0.3 0.6 0.9 a a a a b St em he ight ( cm ) 0 12 16 20 a a b c d St em w id th ( cm ) 0.00 0.10 1.80 1.92 2.04 a ab ab ab b Le af le ngt h (c m ) 0 18 24 30 36 a b c d e LSx10 3 LSx10 4 LS 103 104(CK ) Leaf n um be r 0 10 15 20 25 a b c d d LSx10 3 LSx10 4 LS 103 104(CK ) Leaf w id th ( cm ) 0 9 12 15 a a a b c
Fig. 3. Comparison of the traits among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli
crossed with cauliflower-Li Syue. Error bar is the standard error of mean (n = 4). Means with the same letters are not significantly different at 5% level by LSD test. LSD: least significant difference.
Le af l engt h (c m ) 0 28 32 36 40 Pl an t wi dt h ( cm ) 0 48 54 60 66 Pl an t he ight ( cm ) 0 32 36 40 44 a ab b b c a a a b c P et io le l engt h (c m ) 0 8 10 12 a b b b c Pe tiol e wi dt h ( cm ) 0.0 0.7 0.8 0.9 1.0 a b b bc c St em w id th ( cm ) 0.0 1.5 1.8 2.1 2.4 a b b c c St em h ei gh t ( cm ) 0 12 15 18 b a b c d b a a a a GKx10 3 GKx10 4 GK 103 104(CK ) L eaf w id th ( cm ) 0.0 14.4 15.6 16.8 18.0 a ab bc c c GKx10 3 GKx10 4 GK 103 104(CK ) Leaf n um be r 0.0 12.0 12.6 13.2 13.8 a ab ab ab b
Fig. 4. Comparison of the traits among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli
crossed with broccoli-Green King (GK). Error bar is the standard error of mean (n = 4). Means with the same letters are not significantly different at 5% level by LSD test. LSD: least significant difference.
Table 3. The with Treatment broccoli ×103 cauliflower × z 103: transge King; LS: B. y Mean±stand letters are si
由
Tab
值,表示皆
與親本間無
系顯著低於
之
F
1後代及
-「麗雪」所
及
103 轉殖
b 值顯
轉殖系顯著
(CK)]之 F
1後
其親本。10
e ethylene pro h closely relate Linez 3 104 (C GK × 103 104 (C GK ×103 LS × LS 103 enic ipt inbree . oleracea var. b dard error (n ignificantly dile 4 得知,所
皆為綠色,貯藏
無顯著差異,至
於
104 (CK) ×
及非轉殖青花
所測得值顯著
殖系,顯示顏色
顯示花球偏黃色
著低於雜交青花
後代或雜交
03 轉殖系花球
duction of flo ed varieties of z CK) × 103 × 103 CK) 103 ed line; 104 (C botrytis– Li Sy = 4), within tr ifferent at 5% l Fig. 5. Flo所測得花球
a
藏期間
103
至第
5 天以
103、「綠王
花菜,即較綠
著高於「麗雪
色為最淺綠
色,貯藏第
花菜[「綠王
「麗雪」之
球第
5 天之
oral heads amo f storage at 25 1 13.8±0 11.1±0 14.5±0 12.0±0 11.4±1 16.6±1 10.1±1 14.5±0 CK): nontrans ue. reatment for e level by LSD t
ower heads yel
值皆為負
雜交後代
103 轉殖
王」 × 103
;花椰菜
雪」 × 103
。
5 天 103
王」或
104
F
1後代及
L、a、b
ong F1 progen 5±2℃after harv D 0.4 aby 0.7 c 0.8 a 0.3 bc .2 c .8 a .0 b 0.8 a sgenic inbreed each sampling test. LSD: leas llowing in F1 p三個測值
為較綠色
度最小,
黃化效果
雜交
F
1後
種子數及
分別
與
104 (C
花粉活性
TTC 染色
nies from tran vested from g Days of storag 3 79.1±1.3 a 39.5±6.0 b 23.3±1.8 c 25.6±1.9 c 28.0±1.1 c 9.3±1.1 b 18.0±4.1 a 23.3±1.8 a d line; GK: B. g time, line me st significant d progenies.
值,分別為
4
,且
a 及 b
即葉綠素降
。
後代花粉活性
及花粉性狀
利用基因轉殖
CK)為花粉貢獻
結果如
Tab
色法分析
103
nsgenic ipt bro reen house. ge 24.6 22.7 26.1 21.8 26.9 26.9 45.2 26.1 oleracea var. i eans followed difference.
6.8、-10.1、
值相較於第
解最少,顯示
性測定、果莢
殖
ipt 青花菜
獻親,其雜交
le 5 所示。
轉殖系為花粉
occoli crossed 5 6±2.2 a 7±1.2 a 1±4.1 a 8±1.1 a 9±1.1 a 9±8.5 b 2±4.7 a 1±4.1 b italica– Green d by different19.6,得知
1 天上升幅
示具有延遲
莢結實率、
菜
103 轉殖系
交
F
1後代之
以
FDA 及
粉貢獻親雜
Table 4. The L, a, and b values of floral heads among F1 progenies from transgenic ipt broccoli crossed with closely related varieties of storage at 25±2℃after harvested from green house.
Treatment Linez Days of storage L a b 1 5 1 5 1 5 broccoli ×103 104(CK) ×103 43.8±0.3 a y 54.3±0.3 cy -8.7±0.2 a y -4.3±0.8 a 8.3±0.3 d y 30.0±0.7 bc GK × 103 43.8±0.5 a 55.7±0.7 b -9.8±0.2 bc -5.7±0.9 b 10.5±0.4 bc 32.0±0.9 b 103 43.8±0.4 a 46.8±0.5 e -11.2±0.4 d -10.7±0.3 d 15.0±1.2 a 19.6±1.1 d 104 (CK) 42.2±0.2 b 53.4±0.5 d -10.2±0.3 c -7.7±0.5 c 12.0±0.6 b 29.1±0.6 c GK 42.1±0.2 b 57.9±0.3 a -9.4±0.1 b -5.1±0.4 ab 9.9±0.3 cd 35.3±0.4 a cauliflower × 103 LS × 103 60.3±1.8 b 63.5±1.5 b -15.9±0.3 c -12.8±0.8 c 36.9±1.2 a 37.0±0.7 a LS 86.5±1.5 a 84.1±1.4 a -4.7±0.4 a -3.0±0.4 a 24.8±1.4 b 22.0±1.2 b 103 43.8±0.4 c 46.8±0.5 c -11.2±0.4 b -10.7±0.3 b 15.0±1.2 c 19.6±1.1 c
z 103: transgenic ipt inbreed line; 104 (CK): nontransgenic inbreed line; GK: B. oleracea var. italica– Green King; LS:
B. oleracea var. botrytis– Li Syue.
y Mean±standard error (n = 4), within treatment for each sampling time, line means followed by different letters
are significantly different at 5% level by LSD test. LSD: least significant difference.
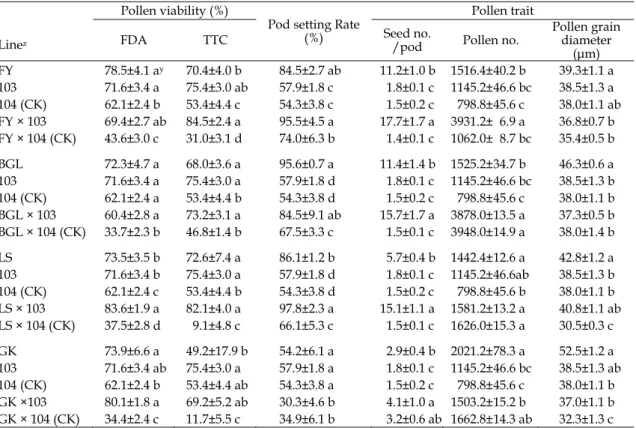
Table 5. Comparison of the pollen viability, pod setting rate, seed number per pod and pollen traits among F1 progenies from transgenic ipt broccoli and non-transgenic broccoli crossed with closely related varieties.
Linez
Pollen viability (%)
Pod setting Rate (%)
Pollen trait
FDA TTC Seed no./pod Pollen no. Pollen grain diameter (μm) FY 78.5±4.1 ay 70.4±4.0 b 84.5±2.7 ab 11.2±1.0 b 1516.4±40.2 b 39.3±1.1 a 103 71.6±3.4 a 75.4±3.0 ab 57.9±1.8 c 1.8±0.1 c 1145.2±46.6 bc 38.5±1.3 a 104 (CK) 62.1±2.4 b 53.4±4.4 c 54.3±3.8 c 1.5±0.2 c 798.8±45.6 c 38.0±1.1 ab FY × 103 69.4±2.7 ab 84.5±2.4 a 95.5±4.5 a 17.7±1.7 a 3931.2± 6.9 a 36.8±0.7 b FY × 104 (CK) 43.6±3.0 c 31.0±3.1 d 74.0±6.3 b 1.4±0.1 c 1062.0± 8.7 bc 35.4±0.5 b BGL 72.3±4.7 a 68.0±3.6 a 95.6±0.7 a 11.4±1.4 b 1525.2±34.7 b 46.3±0.6 a 103 71.6±3.4 a 75.4±3.0 a 57.9±1.8 d 1.8±0.1 c 1145.2±46.6 bc 38.5±1.3 b 104 (CK) 62.1±2.4 a 53.4±4.4 b 54.3±3.8 d 1.5±0.2 c 798.8±45.6 c 38.0±1.1 b BGL × 103 60.4±2.8 a 73.2±3.1 a 84.5±9.1 ab 15.7±1.7 a 3878.0± 13.5 a 37.3±0.5 b BGL × 104 (CK) 33.7±2.3 b 46.8±1.4 b 67.5±3.3 c 1.5±0.1 c 3948.0± 14.9 a 38.0±1.4 b LS 73.5±3.5 b 72.6±7.4 a 86.1±1.2 b 5.7±0.4 b 1442.4±12.6 a 42.8±1.2 a 103 71.6±3.4 b 75.4±3.0 a 57.9±1.8 d 1.8±0.1 c 1145.2±46.6ab 38.5±1.3 b 104 (CK) 62.1±2.4 c 53.4±4.4 b 54.3±3.8 d 1.5±0.2 c 798.8±45.6 b 38.0±1.1 b LS × 103 83.6±1.9 a 82.1±4.0 a 97.8±2.3 a 15.1±1.1 a 1581.2± 13.2 a 40.8±1.1 ab LS × 104 (CK) 37.5±2.8 d 9.1±4.8 c 66.1±5.3 c 1.5±0.1 c 1626.0± 15.3 a 30.5±0.3 c GK 73.9±6.6 a 49.2±17.9 b 54.2±6.1 a 2.9±0.4 b 2021.2±78.3 a 52.5±1.2 a 103 71.6±3.4 ab 75.4±3.0 a 57.9±1.8 a 1.8±0.1 c 1145.2±46.6 bc 38.5±1.3 ab 104 (CK) 62.1±2.4 b 53.4±4.4 ab 54.3±3.8 a 1.5±0.2 c 798.8±45.6 c 38.0±1.1 b GK ×103 80.1±1.8 a 69.2±5.2 ab 30.3±4.6 b 4.1±1.0 a 1503.2± 15.2 b 37.0±1.1 b GK × 104 (CK) 34.4±2.4 c 11.7±5.5 c 34.9±6.1 b 3.2±0.6 ab 1662.8± 14.3 ab 32.3±1.3 c
z 103: transgenic ipt inbreed line; 104 (CK): nontransgenic inbreed line; BGF: B. oleracea var. alboglabra– Bai Ge Lin; FY: B. oleracea var. alboglabra– Fu Yue; LS: B. oleracea var. botrytis– Li Syue; GK: B. oleracea var. italica– Green King.
y Mean±standard error (n = 4), the means given within a column for each closely related variety followed by different letters are significantly different at 5% level by LSD test. LSD: least significant difference.
交近緣種
F
1後代中,分別只有「麗雪」
× 103
和「富悅」 × 103 之之 F
1後代花粉活性顯著
高於親本、
「麗雪」
× 104 (CK)及 104 (CK),
且此兩近緣亞種之
F
1後代果莢結實率亦最
高。而
103 雜交「白格林」及「綠王」之 F
1後代,花粉活性與親本(103 轉殖系及近緣亞
種本身)無差異,但顯著高於 104 (CK)雜交 F
1後代或
104 (CK)。由兩種分析方法結果,均
以
104 (CK)花粉貢獻親分別與 4 種近緣亞種
作物雜交
F
1後代的花粉活性顯著最低,而果
莢結實率只有雜交綠王之
F
1後代(「綠王」 ×
103 及「綠王」 × 104 分別為 30.3%與 34.9%)
顯著低於雜交親本(親本範圍 54.257.9%)。
但每果莢種子數皆以
103 轉殖系雜交近緣亞
種
F
1後代顯著高於
104 (CK)雜交 F
1後代及雜
交親本,顯示基因轉殖
ipt 青花菜雜交近緣亞
種
F
1後代仍具有相當的雜交優勢(Table 5)。
以「富悅」 × 103 雜交 F
1後代檢測估算
每朵花之花粉量平均達
3,931.2 粒,顯著高於
雜交親本[「富悅」、103 轉殖系、104 (CK)]
及「富悅」
× 104 之花粉粒數 798.8-1,516.4
粒範圍間;而白格林與
103 轉殖系或 104 (CK)
雜交之
F
1後代花粉粒數,分別為
3,878.0 粒
及
3,948.0 粒,均顯著高於雜交親本[「白格
林」、103 轉殖系、104 (CK)]之 798.8-1,525.2
粒間。
103 轉殖系和 104 (CK)分別與這 4 個近
緣亞種作物雜交
F
1後代之花粉粒直徑,顯著
小於近緣亞種本身或
103 轉殖系,僅「麗雪」
× 103 介於親本之間。以 103 轉殖系雜交 F
1後代相較於
104 (CK)雜交 F
1後代及雜交親本
中,只有「富悅」 × 103 花粉量增多,整體
而言,其後代對花粉性狀影響甚微(Table 5)。
討論
一般而言,雜草的特質包括花粉易傳播
授粉、種子易萌芽、壽命長及多產等優勢的
繁殖潛力,雜草亦具有幼苗生長快速及植株
耐逆境等強勢的競爭力(Baker 1974)。基因轉
殖作物野化之可能途徑有二:一為由於導入
的基因,使其產生具有雜草特質的適應力及
競爭力;另一為基因轉殖作物與其近緣種雜
交,經由花粉傳播形成基因流佈,而成為具
有 雜 草 特 質 優 勢 的 近 緣 種
(Chapman and
Burke 2006)。
目前有關基因轉殖十字花科蕓苔屬作物
油菜之基因流佈研究居多,皆顯示轉殖抗蟲
基因
B. napus 與同屬不同種 B. juncea 雜交後
具有雜種優勢(Huangfu et al. 2011)。在自然
條件下,蕓苔屬間之甘藍型油菜
B. napus (2n
= 38, AACC)與白菜類 B. rapa (2n = 20,
AA)、芥菜類物種 B. juncea (2n = 36, AABB)
容易發生自然雜交,但與甘藍類
B. oleracea
(2n = 18, CC) 植 物 極 難 發 生 天 然 雜 交
(Hauser et al. 1998, Jørgensen et al. 1998,
Metz et al. 1997, Scheffler and Dale 1994)。
然而,本研究也發現
C 染色體組之 B.oleracea
var. italica 與 A 染色體組的 B. rapa 不同種間
無法雜交,而無探討其雜交後代,此與蕓苔
屬種間雜交的
A、B 及 C 基因組間之同源性
有關,A 與 C 基因組遺傳背景差異大,故不
親 合
(McGrath and Quiros 1991, Prakash
and Chopra 1990)。
進一步探討
103 轉殖系雜交 F
1後代植株
競爭能力,由於
103 轉殖系株高較 104 (CK)
矮,但產量佳(Chan et al. 2009)。本研究發現
103 轉殖系之雜交 F
1較
104 雜交 F
1後代為
矮);其 103 轉殖系為花粉貢獻親所雜交「白
格林」F
1後代之株高、莖長、葉長和葉寬及
「綠王」 × 103 後代之株高、株寬、葉長和
葉 寬 之 性 狀 變 異 係 數 大 於 雜 交 親 本 或
104
(CK)雜交後代,「富悅」 × 103 和「麗雪」 ×
103 大部分性狀變異程度小於 104 (CK)雜交
後代。整體而言,103 轉殖系雜交近緣亞種
F
1後代多數性狀仍在雜交親本、104 (CK)雜
交
F
1後代及
104 (CK)範圍以內,性狀改變甚
微。此可能為外源基因的表達產物並不是植
物生長發育所必需的,其表達過程必然消耗
植株的能量,打破原有的能量供給與消耗平
衡 , 造 成 一 些 性 狀 表 達 所 需 能 量 的 相 對 不
足,影響生理代謝而導致轉基因雜交後代性
狀的變化(Yu et al. 2006)。另一方面,轉殖基
因植株的植株性狀在後代會同時出現有利變
異和不利變異,可能是由於轉入的外源基因
打破了受體原有遺傳物質之間的平衡,使受
體基因的表達受到影響,進而影響性狀的改
變(Guo et al. 2006)。
另一個值得關注的,係基因轉殖
ipt 青花
菜與其近緣亞種雜交
F
1後代之花粉活性、果
莢結實率、種子數,相較於親本均產生改變。
本試驗發現
103 雜交後代活性顯著高於 104
雜交後代,但並沒有超過非基因轉殖青花菜
的變幅範圍,其果莢結實率高於親本或與親
本無差異。說明了花粉活性高並不全代表其
雜交率高,而只是具有相當的雜交潛力。根
據其他相關研究指出,不同作物與其野生種
之間的雜交率不同,同一作物在不同條件下
與其野生種的雜交率也多少存在差異,如各
種植物的花期、生長勢、隔離情形、昆蟲分
布及氣候都會影響其表現(Chèvre et al. 1997,
2000, Mikkelsen et al. 1996)。此是否揭示
出,每一個基因轉殖材料的花粉活性與其相
近緣亞種雜交之稔實率具有不一致的現象,
且可能與整個雜交稔實過程中諸多影響因素
有關,有待進一步的探研。
由本研究果莢結實率顯示,青花菜 × 青
花菜
(花粉活性除外)相較於芥藍 × 青花菜
或花椰菜 × 青花菜雜交率較低,有可能是同
為
B. oleracea L. var. italica Planck,其遺傳背
景較為相似。不同物種間具有雜交優勢,而
使得芥藍 × 青花菜或花椰菜 × 青花菜的雜
交 率 較 高 。 蕓 苔 屬 作 物 之 雜 交 過 程 極 為 複
雜,有時雜交適合度高於親本,可反映出雜
交親本的拮抗作用與基因頻率之間的遺傳差
異程度(Hauser et al. 2003)。又由 103 雜交後
代種子數高於
104 (CK)雜交後代或親本,此
結果說明
103 轉殖系較 104 (CK)具雜交優勢
表。花粉性狀因作物親緣性不同,造成雜交
後代花粉型態的改變,花粉粒較大者花粉存
活可能性越高(「綠王」花粉數量多,但活性
不高可能有效花粉數量少)。目前還無直接的
證據解釋這些性狀改變的原因,是由於所轉
入
ipt 基因所造成的,但過去研究報告指出,
ipt 基因能在花朵部位促進細胞分裂素生物合
成 或 內 源 性 細 胞 分 裂 素 的 累 積 以 延 遲 老 化
(Chang et al. 2003)。ipt 基因轉殖株系之產量
較非基因轉殖植株更高(Chan et al. 2009, Liu
et al. 2011, Ting et al. 2010),以及 ipt 基因轉
殖青花菜花球採收後會累積逆境反應相關蛋
白
(Liu et al., 2011)。此外,由於隨機插入
DNA 序列的性狀改變為不可預知的結果,從
而 改 變 了 受 體 作 物 原 本 的 特 性
(Wei et al.
2005),此結果提供基因轉殖 ipt 青花菜與近
緣種雜後代競爭能力相關訊息。
在加拿大、日本等國家發現,由於油菜
種子採收後的脫落種子殘留於田間演變成實
生苗,使轉基因油菜植株適應性增加,其實
生苗再與野生族群造成基因流佈,演變成雜
草(Beckie et al. 2006, Mitsuko et al. 2011)。目
前利用轉基因甘藍(CC)與非基因轉殖白菜雜
交,從而獲得
C 染色體轉基因甘藍型油菜的
新策略,可降低基因散佈風險。利用農桿菌
轉殖法將抗除草劑
bar 基因轉殖到芥藍(B.
oleracea var. alboglabra, CC)中,以轉殖 bar
基 因 的 芥 藍 為 父 本 , 非 基 因 轉 殖 的 白 菜
(B.
rapa, AA)為母本,透過種間雜交、子房培養
及染色體加倍,
bar 基因已結合(integrate)到
人工合成甘藍型油菜的基因組中,當外來
bar
基因插入甘藍型油菜
C 染色體上時,回交後
代中位於
C 染色體組上的 bar 基因在多代以
後 會 消 失 , 而 降 低 基 因 擴 散 風 險
(Li et al.
2006)。雖然青花菜食用部位為花球,若栽培
者將花球採收後,殘株繼續留在田間,由側
芽再開花結果,或是育種者採收種子過程,
仍可能造成種子脫粒於田間,生成實生苗。
本試驗
ipt 基因於 F
1後代花球延遲黃化效果
仍以轉殖系親本較佳,造成此原因推測,透
過雜交方式使植株基因組重組,在有性階段
經植株萌芽、生長開花、減數分裂、授粉授
精 、 合 子 形 成 及 胚 發 育 等 一 系 列 之 複 雜 過
程。再者,外界環境條件和植物生長發育狀
況,都有可能影響目標基因表現異常、不表
現或表現低(He et al. 2007)
然而,
103 轉殖系雜交 F
1後代相較於
104
(CK)雜交 F
1後代及親本具雜交優勢,若後代
的種子中具有較強的生命力,目標基因可能
從雜交後代再次流佈到近緣種作物。再者,
由於
ipt 基因對逆境耐受性較佳(Adhikary et
al. 2010, Merewitz et al. 2011),演變成野化
之風險可能性也相對提高,本研究結果可做
為將來基因轉殖十字花科作物共存可行性及
野化能力評估方法之參考。由於本試驗是在
溫室條件下所獲得結果,未來可進一步探討
在田間對於轉基因雜交後代或其親本的種子
於土壤中之休眠性及是否演變成實生苗之可
能性。
引用文獻
Adhikary M, S Ganguli, HJ Chakraborty (2010) Computational analysis of the SAG-13 gene encoding an alcohol dehydrogenase. J. Pharm. Biomed. Sci. 2:14.
Andow DA, Z Claudia (2006) Assessing environmental risks of transgenic plants. Ecol. Lett. 9:196214.
Baker HG (1974) The evolution of weeds. Annu. Rev. Ecol. Syst. 5:124.
Beckie HJ, KN Harker, SM Hall, SI Warwick, A Legere, PH Sikkema, GW Clayton, AG Thomas, JY Leeson, G Seguin-Swartz, MJ Simard (2006) A decade of herbicide-resistant crops in Canada. Can. J. Plant Sci. 86:12431264.
Campbell LG, AA Snow (2007) Competition alters life history and increases the relative fecundity of crop-wild radish hybrids (Raphanus spp.). New Phytol. 173:648660.
Campbell DR, NMWaser (2001) Genotype- by-environment interaction and the fitness of plant hybrids in the wild. Evolution 55: 669676.
Cao QJ, H Xia, X Yang, BR Lu (2009) Performance of hybrids between weedy rice and insect-resistant transgenic rice under field experiments: Implication for environmental biosafety assessment. J. Intl. Plant Biol. 51:11381148.
Chan LF, LFO Chen, HY Lu, CH Lin, HC Huang, MY Ting, YM Chang, CY Lin, MT Wu (2009) Growth, yield and shelf-life of isopentenyl transferase (ipt)-gene transformed broccoli. Can. J. Plant Sci. 89:701711.
Chang H, LJ Michelle, MB Gary, GC David (2003) Overproduction of cytokinins in petunia flowers transformed with PSAG12-IPT delays corolla senescence and decreases sensitivity to ethylene. Plant Physiol. 132:21742183.
Chapman MA, JM Burke (2006) Letting the gene out of the bottle: The population genetically modified crops. New Phytol. 170:429443. Chèvre AM, F Eber, A Baranger, M Renard (1997)
Gene flow from transgenic crops. Nature 389:924.
Chèvre AM, F Eber, H Darmency, A Fleury, H Picault, JC Letanneur, M Renard (2000) Assessment of interspecific hybridization between transgenic oilseed rape and wild radish under agronomic conditions. Theor. Appl. Genet. 100:12331239.
FitzJohn RG, TT Armstrong, LE Newstrom-Lloyd, AD Wilton, M Cochrane (2007) Hybrid isolation within Brassica and allied genera: evaluation of potential for transgene escape. Euphytica 158:209230.
Funk T, G Wenzel, G Schwarz (2006) Outcrossing frequencies and distribution of transgenic oilseed rape (Brassica napus L.) in the nearest neighborhood. Europ. J. Agron. 24:2634. Gueritaine G, M Sester, F Eber, AM Chèvre, H
Darmency (2002) Fitness of backcross six of hybrids between transgenic oilseed rape (Brassica napus) and wild radish (Raphanus raphanistrum). Mol. Ecol. 11:141926.
Guo JF, YX Huang, SH Jiang, HX Yuan, XP Xu, JZ Zhang (2006) The effect of transgenic restorer lines on agronomic traits of F1 hybrid rice. Southwest China J. Agric. Sci. 19:772776. Hauser TP, C Damgaard, RB Jørgensen (2003)
Frequency-dependent fitness of hybrids between oilseed rape (Brassica napus) and weedy B. rapa (Brassicaceae). Amer. J. Bot. 90:571578.
Hauser TP, RB Jørgensen, H Østergárd (1998) Fitness of backcross and F2 hybrids between weedy Brassica rapa and oilseed rape (B.napus). Heredity 81:436443.
He ZZ, M Huang, LY Chen (2007) Genetic expression of exotic genes and their effects on
non-target traits in transgenic rice. Hybrid Rice 22:610.
Heinrich T, DP Antje (2012) Marker-assisted identification of oilseed rape volunteers in oilseed rape (Brassica napus L.) fields. 25th German Conference on Weed Biology and Weed Control 434:361367.
Heslop-Harrison J, Y Heslop-Harrison (1970) Evaluation of pollen viability by enzymatically induced fluorescence; intracellular hydrolysis of fluorescein diacetate. Biotech. Histochem. 45:115120.
Huangfu CH, S Qiang, XI Song (2011) Performance of hybrids between transgenic oilseed rape (Brassica napus) and wild Brassica juncea: An evaluation of potential for transgene escape. Crop Prot. 30:5762.
Johannessen MM, BA Andersen, RB Jørgensen (2006) Competition affects gene flow from oilseed rape (♀) to Brassica rapa (♂). Heredity 96:360367.
Jørgensen RB, B Andersen, L Landbo, TR Mikkelsen (1996) Spontaneous hybridization between oilseed rape (Brassica napus) and weedy relatives. Acta Hort. 407:193200. Jørgensen RB, B Andersen, TP Hauser (1998)
Introgression of crop genes from oilseed rape (Brassica napus) to related wild species: An avenue for the escape of engineered genes. Acta Hort. 459:211217.
Li C, BW Zhou, XL Guo, CH Dong, XJ Hu, MS Hou, SY Liu (2008) Pollen-mediated gene flow in Chinese commercial fields of glufosinate- resistant canola (Brassica napus). Chinese Sci. Bull. 53:23332341.
Li J, XP Fang, Z Wang, J Li, LX Luo, Q Hu (2006) Transgene directionally integrated into C-genome of Brassica napus. Chinese Sci. Bull. 51:15781585.
Li YJ, CM Lu, XJ Wang, SR Jia, ZX Wang (2012) Data survey and analysis of the transgene flow frequencies and distances in major crops V. Rapeseed. (in Chinese with English abstract) J. Agric. Sci. Tech. 14:4956.
Liu MS, HC Li, YM Chang, MT Wu, LFO Chen (2011) Proteomic analysis of stress-related proteins in transgenic broccoli harboring a gene for cytokinin production during postharvest senescence. Plant Sci. 181:288299. McGrath JM, CF Quiros (1991) Inheritance of
isozyme and RFLP markers in Brassica
campestris and comparison with B. oleracea. Theor. Appl. Genet. 82:668673.
Mercer KL, DA Andow, DL Wyse, RG Shaw (2007) Stress and domestication traits increase the relative fitness of crops wild hybrids in sunflower. Ecol. Lett. 10:383393.
Merewitz EB, T Gianfagna, BR Huang (2011) Photosynthesis, water use, and root viability under water stress as affected by expression of SAG12-ipt controlling cytokinin synthesis in Agrostisstolonifera. J. Exp. Bot. 62:383395. Metz PLJ, E Jacobsen, JP Nap, A Pereira, WJ
Stiekema (1997) The impact on biosafety of the phosphinothricin-tolerance transgene in inter-specific B. rapa×B. napus hybrids and their successive backcrosses. Theor. Appl. Genet. 95:442450.
Mikkelsen TR, B Andersen, RB Jørgensen (1996) The risk of crop transgene spread. Nature 380:3133.
Mitsuko A, S Wakiyama, M Nagatsu, Y Kaneko, T Nishizawa, N Nakajima, M Tamaoki, A Kubo, H Saji (2011) Seeds of a possible natural hybrid between herbicide-resistant Brassica napus and Brassica rapa detected on a riverbank in Japan. GM Crops 2:201210.
Mohammad SK (2011) Future challenges in environmental risk assessment of transgenic plants with abiotic stress tolerance. Biotechnol. Mol. Biol. Rev. 6:199213.
Norton JD (1966) Testing of plum pollen viability with tetrazolium salts. Proc. Amer. Soc. Hort. Sci. 89:132134.
Prakash S, VL Chopra (1990) Reconstruction of allopolyploid Brassica through non- homologous recombination: introgression of resistance to pod shatter in Brassica napus. Genet. Res. 56:12.
Prakash S, K Hinata (1980) Taxonomy, cytogenetics and origin of crop Brassicas, a review. Opera Bot. 55:157.
Roy NN (1980) Species cross ability and early generation plant fertility in interspecific crosses of Brassica. Sabrao J. 12:4353.
SAS Institute Inc. 2006. SAS/STAT E.G User's Guide. Version 8. SAS Institute Inc. Cary NC 5136pp.
Scheffler JA, PJ Dale (1994) Opportunities for gene transfer from transgenic oilseed rape (Brassica napus) to related species. Transgenic Res. 3:263278.
Snow AA, TM Culley, LG Campbell, PM Sweeney, SG Hegdeand, NC Ellstrand (2010) Long-term persistence of crop alleles in weedy populations of wild radish (Raphanus raphanistrum). New Phytol. 186:537548.
Ting PY, H Chang, YW Kao, HY Lu, LFO Chen, MT Wu, LF Chan (2010) Pollen viability and cross ability of isopentenyl transferase (ipt)-gene transformed broccoli (Brassica oleracea var. italica) and its relatives. (in Chinese with English abstract) J. Taiwan Agric. Res. 59:261274.
Vacher C, AE Weis, D Hermann, T Kossler, C Young, ME Hochberg (2004) Impact of ecological factors on the initial invasion of Bt transgenes into wild populations of birdseed rape (Brassica rapa). Theor. Appl. Genet. 109:806814.
Wang ZX, XJ Wang, SR Jia (2011) Data survey and analysis of the transgene flow frequencies and distances in major crops.Ⅰ.The background, aim and general consideration. (in Chinese with English abstract) J. Agric. Sci. Tech. 13:2629.
Wei XH, XP Yuan, HY Yu, YP Wang, SX Tang, XY Liao (2005) Effects of transgenes insertion on pollen vigor and hybrid seed set of rice. (in Chinese with English abstract) Chinese J. Appl. Ecol. 16:115118.
Whitney KD, RA Randell, LH Rieseberg (2006) Adaptive introgression of herbivore resistance traits in the weedy sunflower Helianthus annuus. Amer. Nat. 167:794807.
Yu, C., Y. Feng, Q. Xue (2006) Comparative study on the variation of agronomy traits of transgenic insect resistance rice. (in Chinese) Chinese Agric. Sci. Bull. 22:579