國 立 交 通 大 學
生物資訊及系統生物研究所
碩 士 論 文
由結構推導蛋白質與蛋白質接觸面的動力學特性
Structure-derived dynamic properties of
protein-protein interfaces
研 究 生:林子琳
指導教授:黃鎮剛 教授
i
由結構推導蛋白質與蛋白質接觸面的動力學特性
研究生: 林子琳 指導教授: 黃鎮剛 教授 國立交通大學 生物資訊及系統生物研究所摘 要
蛋白質的交互作用完成了許多生物功能。序列研究指出蛋白質與蛋白質作用 區富含厭水性胺基酸;然而,目前為止蛋白質的結構資訊尚無法清楚地區分蛋白 質與蛋白質的接觸面與其他蛋白質表面。我們在這個研究中分析了兩項與動力學 相關的結構資訊: 蛋白質與蛋白質接觸面的中心與蛋白質的質心距離比一般蛋 白質表面與蛋白質的質心距離小。蛋白質與蛋白質接觸面的中心的加權接觸數目 比一般蛋白質表面的加權接觸數目大。這表示蛋白質與蛋白質接觸面的中心是靠 近蛋白質的質量中心並且處於擁擠的堆疊狀態。ii
Structure-derived dynamic properties of protein-protein interfaces
Student: Zih-Lin Lin Advisor: Dr. Jenn-Kang HwangNational Chiao Tung University
Institute of Bioinformatics and Systems Biology
Abstract
Many biological functions result from the interactions between proteins. From
sequence information, studies have revealed that the interaction interfaces are
conserved in hydrophobic environments. However, structure information is still not
clear enough to differentiate interaction interfaces from protein surfaces. In this study,
we analyzed two structural properties related to protein dynamics: the core interface
residues are closer to the centroid of protein, and the weighted contact numbers of
core interface residues are larger than that of surface residues. The results suggest that
the core interface residues are nearness to the protein centroid and in a crowded
iii
誌 謝
謝謝大家!!感恩幫助我完成論文,兩年以來給我支持與鼓勵的所有人!!首先 要感謝的是提供我研究機會的黃鎮剛老師,給予我研究上的指教,指點我學術上 的迷津,並且鼓勵我以樂觀的想法與不同的角度看待自己的實驗成果。感謝老師 讓我了解作研究應有的態度與方法也教導我待人接物的道理。接著我要感謝實驗 室研究同仁們:景盛,志豪,志鵬,建華,存操,啟文,彥龍,士中,儼毅,仙 蕾,人維對我在程式與實驗設計方面的指導與幫助以及協助我的口試的進行。還 有實驗室的研究同仁們:少偉,志杰,肇基,瓊文,松桓,曉芬,乃文,儷芬在 我研究生活上的幫忙助益。還要感謝系上朋友們:智先,志偉,慶恩,宗翰,彥 修,敬立,廖芹,佳達,昭昉,致宏,在我研究遇到瓶頸時給我的支持與鼓勵。 另外,我要特別感謝:志鵬學長,存操學長,啟文學長,士中學長,仙蕾學姊協 助我設計實驗也教導我程式邏輯與應用,並且幫助我撰寫論文,教了我實驗低潮 時應抱持正面樂觀的態度。我也要感謝我的家人在我背後的支持,還有之義在我 旁邊幫我搖旗加油,以及我的愛鼠賈修在新竹陪伴我。紙短情長,再次感謝所有 幫助我與陪伴我的人們。iv
Contents
摘 要... i Abstract ... ii 誌 謝... iii Contents ... iv Introduction ... 1Materials and Methods ... 5
ProMate database ... 5
ZW database... 6
Surface and interface definition ... 7
Amino acid components ... 8
Secondary structure definition ... 8
Centroid Model (CM) ... 9
Weighted Contact-Number model (WCN) ... 10
Z-score ... 10
Two-sample t-test... 11
Results and Discussions ... 12
Amino acid components ... 12
Secondary structure constituents ... 12
Accessibilities and delta accessibilities ... 13
Evolutionary conservation ... 13
B-factors ... 14
Centroid Model (CM) ... 16
Weighted Contact Number model (WCN) ... 17
Summary ... 18
References ... 20
Figures... 22
1
Introduction
Classifying residues as surface exposed and buried, based on their solvent
accessibilities, is a simple but important step towards understanding the contributions
of the residues to the structural integrity1-2. Surface exposed residues are often crucial
for interactions with other proteins and play functional roles while the buried residues
contribute more towards stability of the tertiary structures3. Proteins perform their
function by interacting with other molecules, such as small ligands, lipids, nucleic
acids, and other proteins4. The recognition of protein-protein interaction sites can be
used to identify functionally important amino acid residues, facilitate experimental
efforts to catalog protein interactions, enhance computational docking studies and
drug designs, as well as enable functional annotation for the growing number of
structurally resolved proteins of unknown functions4.
Identification of the interface between interacting proteins is an important clue to
the function of a protein. In general, the problem of recognition of protein–protein
interaction sites can be cast as a classification problem, that is, each amino acid
residue is assigned to one of two classes: interacting (interfacial) or non-interacting
(non-interfacial) residues4. The experimental methods such as yeast-two-hybrid
screening, immune-precipitation assays and Föster resonance energy transfer (FRET)
2
but it is still difficult to use above experimental methods to identify which residues
are in the interaction region, called interface. By analyzing the surfaces of proteins,
the interfaces can be differentiated from the surfaces of component subunits in protein
complexes.
The characteristics of interface residues have been systematically studied. Lo
Conte and Chothia et al.8 have analyzed the amino acid compositions of
protein-protein complexes. They discovered the interface residues are more aliphatic
and aromatic than the rest part of protein surface. Neuvirth et al.9 noticed that polar
and hydrophobic residues are more plentiful in the interface than the rest part of
surface. Tyr, Met, Cys, and His are favored on binding interface9. Zhou and Shan et
al.10 observed that the interface residues are apparently more conserved than the
non-interfacial surface residues.
The protein dynamics tell us the information about how the protein moves. It is
well-known that protein dynamics is highly correlated to protein function. The
experimental measurement of the oscillations of an atom around its mean position in a
protein structure is called B-factor. The B-factor (also called crystallographic
temperature factor and Debye-Waller factor) in protein X-ray structures is an
experimental evidence to protein structure dynamics and is closely related to the
3
B-factor of proteins in free form and revealed that the B-factor of interface is slightly
lower than that of the surface in the unbound state9. This result is consistent with the
finding that an interfacial surface region is less flexible than the rest of the protein
surface in unbound state12.
The protein dynamics are usually calculated by mechanical models. Molecular
dynamics (MD) simulation is one of the most famous mechanical models used to
describe protein flexibility. Molecular dynamics computes the movements of proteins
based on bond stretching, bond angle bending, bond twisting, van der Waals and
electrostatic interaction13-15. The main drawback of MD simulation is its high
computational cost16-17. Several prediction methods for protein dynamics elaborated
based on protein structures overcame this limitation. For example, the centroid model
(CM)18 and weighted contact number model (WCN)19 developed by our group. The
CM computes protein dynamics directly from the protein geometrical shape. The CM
method is based on the observation that the deeper an atom is buried inside a protein
structure, the less it will fluctuate around its equilibrium position18. The CM only
computes the coordinates of Catoms and plainly defines the center of mass of a protein. The distance square between the C atoms to the center of mass of the protein is accordant to the thermal fluctuation. The atomic fluctuation is in fact
4
protein18. The weighted contact number model (WCN) calculates the number of
neighbor atoms which is weighted by inverse distance between two atoms of each pair.
The WCN computes protein dynamics from the protein packing. If an atom is more
crowded in a protein structure, the less it will fluctuate around its equilibrium
position19. We use the CM and WCN to analysis the differences between interfaces
and the rest part of the surfaces. We applied the two methods to protein-protein
complexes and found the correlation between protein dynamics and interface residues.
In the work presented here, the distance to the protein centroid and the weighted
contact number were analyzed for both interfaces and surfaces. These two structural
5
Materials and Methods
We use the ProMate database and ZW databases to analyze the components of
protein interface residues and surface residues. And we have further defined the
interface residues as core interface residues and peripheral interface residues. Core
interface develops the center region of interface and peripheral interface contrasts the
rim area of interface. The core interface and peripheral interface together form the
interface. We analysis the components and tendencies of core interface and peripheral
interface.
ProMate database
ProMate database9 contains 57 protein-protein interaction structures. The
database consists of both the unbound and bound states for transient protein-protein
hetero-dimers derived from the PDB20. The unbound and bound states of proteins
were determined by X-ray crystallography or NMR. The ProMate database has 42
X-ray structures protein structures in unbound form.
The ProMate database was extracted from a database of 92 bound monomers
longer than 85 AA. The combinatorial extension method (CE)21 was prosecuted to
find each possible pair of monomers and one of them would be executed from
6
The unbound structures were then derived from the bound structures. 57
monomers in bound form were found to have a highly homologous unbound form in
the PDB20 by using BLAST22, with more than 70% sequence identity.
ZW database
The ZW dataset contains 101 transient protein-protein complexes. The 101
transient protein-protein complexes were retrieved from the Zhiping Weng’s transient
databases23. The Zhiping Weng group had collected 212 non-redundant transient
hetero-dimeric X-ray structures of protein-protein interactions24.
To obtain data for hetero-dimers, the Zhiping Weng group only kept the records
of X-ray structures better than 3.25 Å and all the chains in the database are longer
than 25 amino acids. They eliminated all homomeric records using the
BLASTCLUST algorithm25. A homomeric record was defined which all chains have
85% sequence identity to each other and at least 50% of the sequence was aligned. To
receive a non-redundant database of protein complexes, they used pairwise BLAST22
to check each pair of all chains in the database and deleted one complex of the pair
with 25% sequence identity.
And we have further used global sequence alignment to check each pair of each
7
identity. We remained 101 transient protein-protein complexes in the ZW database.
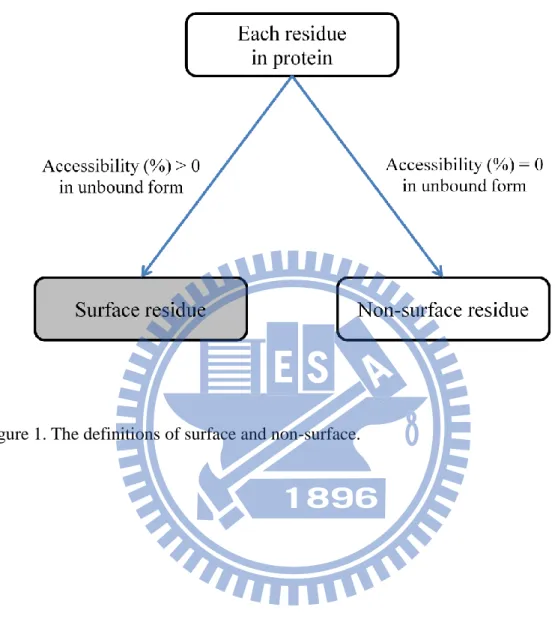
Surface and interface definition
We took the bound state complexes apart and treated the subunits as the unbound
state of complexes. After clarifying the corresponding residues between unbound state
and bound state for protein-protein complexes, we explored the surface residues and
interface residues.
The accessibility(%) plays an important role in definition of surface and interface.
Accessibility(%) is presented as
𝐴𝑐𝑐𝑒𝑠𝑠𝑖𝑏𝑖𝑙𝑖𝑡𝑦𝑖(%) =
𝑆𝐴𝑖
𝑆𝑡𝑎𝑛𝑑𝑎𝑟𝑑 𝑆𝐴𝑖100% (1)
Accessibility(%) of the i-th residue, 𝐴𝑐𝑐𝑒𝑠𝑠𝑖𝑏𝑖𝑙𝑖𝑡𝑦𝑖(%), is defined as the ratio of the
solvent exposed surface area of the i-th residue. SAi is the solvent exposed surface
area of the i-th residue and Standard SAi is the standard value of the solvent exposed
surface area for this kind of amino acid26. We use the DSSP program27 to calculate
exposed surface areas in unbound state and bound state of protein-protein complexes.
We defined the surface residue based on the accessibility(%) in unbound state and the
interface residue based on the delta accessibility(%) upon complex formation28. A
residue was categorized as a surface residue if its accessibility(%) in free form is
8
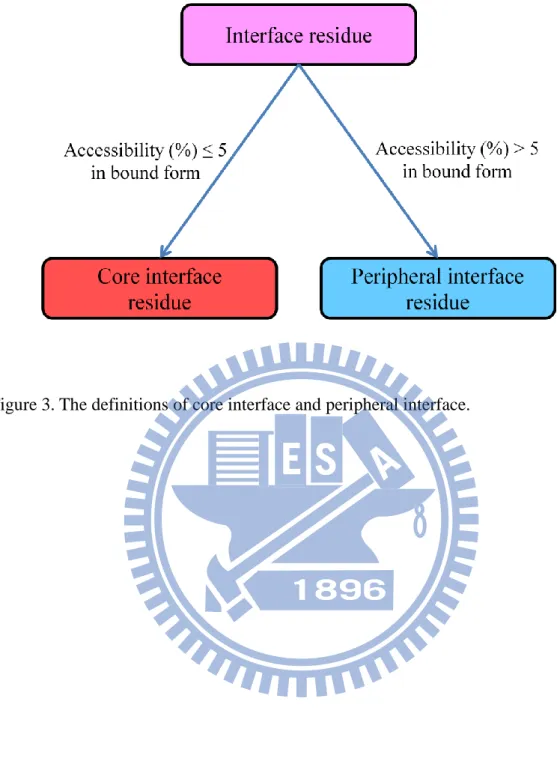
upon complex formation. And we further separated interfaces as core interfaces and
peripheral interfaces based on the accessibilities(%) in bound state. If an interface
residue has the accessibility(%) in the complex smaller than or equal to 5, it is a core
interface residue, else it is a peripheral interface residue. An interface residue is taken
as either a core interface residue or a peripheral interface residue. The definition of
surface, interface, core interface and peripheral interface are showed in Figure
1~Figure 3, and one example is pictured in Figure 4.
Amino acid components
We have calculated the amino acid propensities using the following equations: 𝑃𝑟𝑜𝑝𝑒𝑛𝑠𝑖𝑡𝑦𝑇𝑎𝑟𝑔𝑒𝑡 = 𝑂𝑐𝑐𝑢𝑟𝑒𝑛𝑐𝑒 𝑇𝑎𝑟𝑔𝑒𝑡
𝑂𝑐𝑐𝑢𝑟𝑒𝑛𝑐𝑒𝑆𝑢𝑟𝑓𝑎𝑐𝑒 (2)
Where 𝑃𝑟𝑜𝑝𝑒𝑛𝑠𝑖𝑡𝑦𝑇𝑎𝑟𝑔𝑒𝑡 is the amino acid type propensity of the target residues.
𝑂𝑐𝑐𝑢𝑟𝑒𝑛𝑐𝑒𝑇𝑎𝑟𝑔𝑒𝑡, 𝑂𝑐𝑐𝑢𝑟𝑒𝑛𝑐𝑒𝑆𝑢𝑟𝑓𝑎𝑐𝑒 are the amino acid type occurrences of the
target residues and the surface residues.
Secondary structure definition
The secondary structure is defined by DSSP27 program. DSSP recognizes eight
types of secondary structure, depending on the pattern of hydrogen bonds and 3D
9 H: -helix
B: residue in isolated -bridge
E: extended strand, participates in ladder G: 3/10 helix
I: -helix
T: hydrogen bonded turn
S: bend
U: undefined
These eight types are usually grouped into three larger classes: helix (G,H, and I),
strand (E and B), and loop (all others).
Centroid Model (CM)
Let X0 be the center of mass of the protein, which is express as
𝑋0 = 𝑚𝑘 𝑘𝑋𝑘/ 𝑚𝑘 𝑘 (3)
Where mk and Xk are the mass and the crystallographic position of C atom k, respectively. The distance of the C atom i from the center of mass of the protein is expressed as
𝑟𝑖2 = Xi− X0 Xi− X0 (4)
10
of size N has the square distance of each C atom given by (𝑟12, 𝑟22, … , 𝑟𝑛2). The r2 profile is closely related to the thermal B-factor, which is given as
𝐵𝑖 = ( 8𝜋2
3 )(𝛿𝑋𝑖𝛿𝑋𝑖) (5)
The centroid model suggest the following interesting relation,
𝛿𝑋𝑖𝛿𝑋𝑖 ~ 𝑋𝑖 − 𝑋0 𝑋𝑖− 𝑋0 (6)
And equation (5) and (6) suggests that the fluctuation of a residue is usually
proportional to the distance between center of mass and its position.
Weighted Contact-Number model (WCN)
When the neighboring contact number of an atom is larger, the fluctuation of the
atom will be smaller. We can define WCN model as
𝑉𝑖 = 𝑁𝑗 ≠𝑖( 1/𝑟𝑖𝑗2) (7)
The equation (7) defines 𝑉𝑖 , the number of C atoms which surround the 𝑖𝑡 residue.
The influence of atom j to the atom i is attenuated by the factor 1/𝑟𝑖𝑗2. rij is the
distance between C atoms of residues i and j.
Z-score
On the mission to compare the results, we would normalize the 𝑟𝑖2 of CM and
11
𝑧𝑥𝑖 = (𝑥𝑖 − 𝑥 )/𝜎𝑥 (8)
𝑥 is the mean of x and is the standard deviation of x, where the 𝑥𝑖 represents
the 𝑟𝑖2 of CM and 𝑣
𝑖 of WCN.
Two-sample t-test
To compare the differences between interfaces and the rest part of the surfaces,
we use the two-sample t-test. 𝑡 = 𝑥 −𝑦
𝑠𝑥 2𝑚+𝑠𝑦 2
𝑛
(9)
Where 𝑥 and 𝑦 are the sample means, sx and sy are the sample standard deviations,
and m and n are the sample sizes of the two groups. When a t is determined, a p-value could be decided from the Student’s t-distribution table. If the p-value is lower than 0.05, the two sample have differences.
12
Results and Discussions
Amino acid components
The amino acid distributions of core interfaces, peripheral interfaces and
interfaces could give a general indication of relative importance of different amino
acids. Percentage frequencies and propensities of amino acids were calculated for
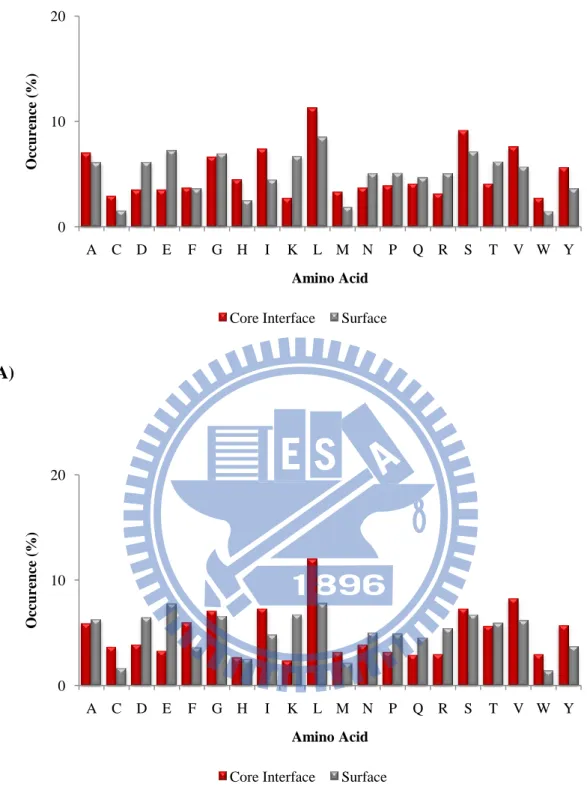
each amino acid type and the results were illustrated in Figure 5~10.
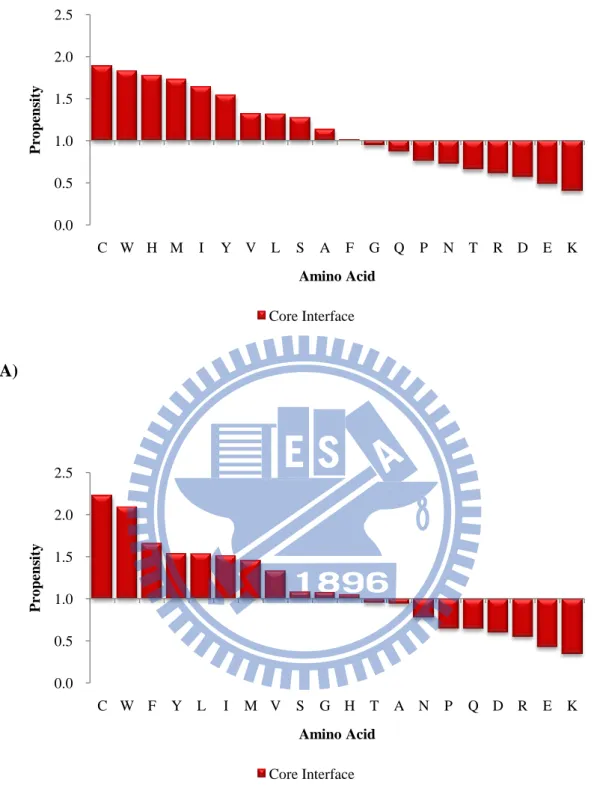
Cys, Trp, His, Met, Ile are the most dominant amino acid types in the core
interface in the ProMate database and Cys, Trp, Phe, Tyr, and Leu are the most
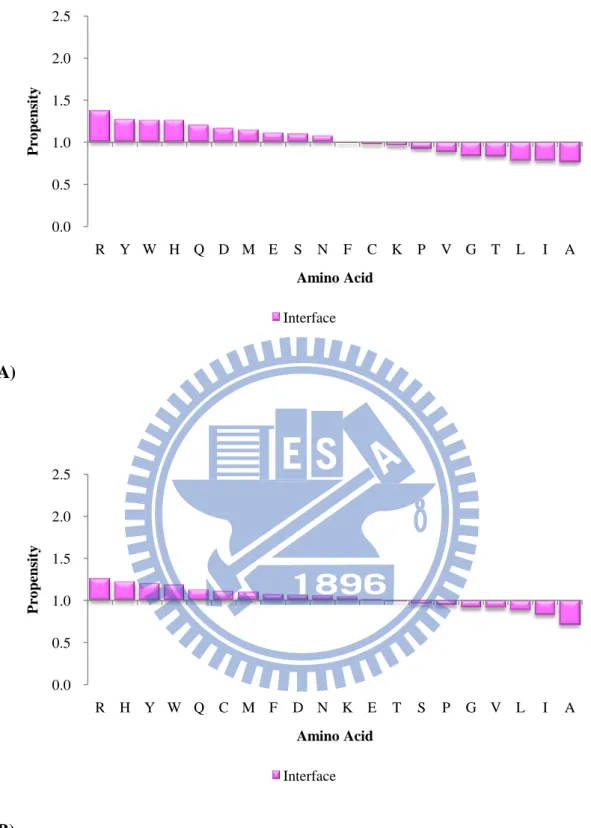
abundant amino acids in the core interface in the ZW dataset. Arg, Asp, Glu, Gln, and
Asn are the most dominant amino acids in the peripheral interfaces in the ProMate
database. Arg, Glu, Lys, His and Glu are the most dominant amino acids in the
peripheral interfaces in the ZW database. We could observed that the core interface
prefers polar, uncharged and aromatic amino acids whereas the peripheral interface
likes charged residues.
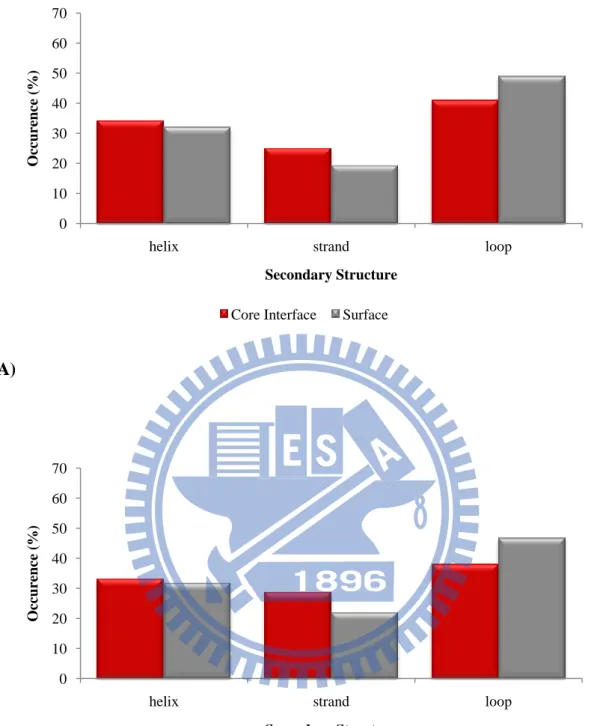
Secondary structure constituents
The occurrences of secondary structures are represented in Figure 11~13. The
rigid secondary structures, helices and strands, are preferred in the core interfaces
13
in the core interfaces contract to surfaces. The flexible secondary structures, loops, are
preferred in the peripheral interfaces compared with surfaces and the rigid secondary
structures, helices and strands, are unfavorable in the peripheral interfaces contract to
surfaces.
Accessibilities and delta accessibilities
The accessibility distributions in unbound state are pictured in Figure 14~16. The
accessibilities of core interfaces are lower than that of surfaces and the accessibilities
of peripheral interfaces are higher than that of surfaces. The analysis of accessibilities
revealed that the peripheral interfaces are significantly more accessible than the rest of
surfaces.
The delta accessibilities in complexation are pictured in Figure 17~18. There is
no difference of delta accessibility distributions between core interfaces, peripheral
interfaces, and interfaces.
Evolutionary conservation
It is interesting to research the conservation degree of proteins if the
protein-protein interactions play important part in function. We measured the
14
corresponds to its evolutionary rate. The residues evolve slowly are directed to be
conserved residues. The lower the conservation score obtained from ConSurf, the
higher the conservation degree the residue has.
We could see that the conservation scores are much lower in the core interfaces
in Figure 19. The p-values between core interfaces and surfaces in the ProMate
database and the ZW database are both 0.00. Figure 20 represents that there is no
difference between the conservation scores of peripheral interface and surface. The
p-values between peripheral interfaces and surfaces in the ProMate database and the
ZW database are 2.39× 10-2, and 5.30 × 10-3 sequentially. Figure 21 shows the
tendencies of conservation scores of interface and surfaces. We could see that the
conservation scores are lower in the interfaces than that in the surfaces. The p-values
of conservation scores of the ProMate database, the ZW database are 1.33 × 10-5,
1.66× 10-11 sequentially. The analysis of conservation scores revealed that the core
interfaces play important roles in protein-protein interactions and the core interfaces
are more conserved than the rest part of surfaces.
B-factors
The X-ray crystallization structures from Protein Data Bank offer the B-factor
15
experiments and related to protein structure dynamics. The higher the B-factor the
residue has, the more flexible the residue is.
We analyzed the B-factor distributions in unbound form of the ProMate database.
We used the EMBOSS Pairwise Alignment Algorithm30 to search for regions of local
similarity and homologous residues between the two sequences of unbound state and
bound state for protein-protein complexes. We found that the B-factors in unbound
form of core interfaces are significantly lower than that of surfaces (Figure 22) and
the t-test comparing core interfaces and surfaces gave a p-value of 1.07 × 10-10. The
B-factors in unbound form of peripheral interfaces are slightly higher than that of
surfaces (Figure 23) and the p-value is 1.31 × 10-10. The B-factors of interfaces have
no difference with that of surfaces (Figure 24) and the p-value is 3.97 × 10-2. We
could notice greatly dissimilarity between core interface and surfaces. We could
observed from the B-factor presences that the core interfaces are more rigid than the
surfaces and the peripheral interfaces are slightly elastic than the surfaces in unbound
state.
Figure 31 depicts the protein structure of pdbid 1tmq. We could exam the
B-factors of core interface and peripheral interfaces of the chain A of 1tmq in the
upper figure. The B-factors of 1tmqA were obtained from the homologous protein of
16
expresses low B-factor values respectively. We could contract the upper and lower
figures and perceive that the B-factors of core interfaces are much lower than that of
the rest of the surfaces.
Centroid Model (CM)
The centroid model (CM) only computes the distance square between each C atom to the center of mass of the protein. The CM method is based on the observation
that the deeper an atom is buried inside a protein structure, the less it will fluctuate
around its equilibrium position.
We measured the CM distributions of protein-protein interactions in unbound
state. The CM of the core interfaces is significantly lower than the CM of the surfaces
in unbound form (Figure 25) and the p-values of ProMate database, ZW database both
are 0.00. The CM of the peripheral interfaces is lightly higher than the CM of the
surfaces in unbound form (Figure 26) and the p-values of ProMate database, ZW
database are 2.77 × 10-6, and 0.00. The CM distributions have no significant
difference between the interfaces and the surfaces (Figure 27). The two sample t-test
gave p-values between the interfaces and surfaces of the two databases with 3.16 ×
10-1, 2.70 × 10-4. The core interfaces are close to the center of mass of protein
17
center of mass of protein structures are slightly longer than that between surfaces and
the center of mass of protein structures.
Visualization of the example of CM model was described in Figure 32. The
average distance between surfaces and the center of mass of 1tmqA is 22.62 Å . The
distance between the core interface residue, W56, and the center of mass is 16.79 Å .
The distance between the peripheral interface residue, E229, and the center of mass is
24.62 Å . The distances of the core interfaces are shorter than that of the surfaces.
Weighted Contact Number model (WCN)
The weighted contact number model (WCN) estimates protein dynamics by
calculating the protein packing denseness. The WCN model computes the number of
neighboring atoms which is weighted by inverse distance between two atoms of each
pair. The WCN is based on that the more crowded an atom is around its environment
in a protein structure, the less it will swing around its equilibrium position.
Examining Figure 28 reveals that core interfaces have much higher WCN than
the whole surfaces. The two sample t-test contrast the core interfaces and the whole
surfaces and gave both p-values of 0.00 in the ProMate database and the ZW database.
Figure 29 shows that peripheral interfaces have much lower WCN than the whole
18
surfaces and gave both p-values of 0.00 in the ProMate database and the ZW database.
Observing Figure 30 reveals that the interfaces have lower WCN than the whole
surfaces. The two sample t-test contrast the interfaces and the whole surfaces and
gave p-values of 7.54 × 10-13
, 0.00 individually in the ProMate database, the ZW
database. The core interfaces have high packing densities and the peripheral interfaces
have low packing densities in unbound state.
We could also observe from Figure 33 that the core interfaces are sunken on the
surface and the peripheral interfaces are protruding on the surface. The example of
Figure 33 agrees with the results of WCN model. It shows that core interface is in a
crowded environment and the peripheral interface is not.
Summary
The results of B-factors, WCN, CM suggest that the core interfaces are rigid and
the peripheral interfaces are plastic on the surface. The analysis of protein secondary
structures also supports the dynamic observations. The evolutionary conservation
measurements exposed that the core interfaces are more conserved in the surface
whereas the peripheral interfaces are not. It means the core interfaces play important
role in protein-protein interactions.
19
protein-protein interaction are rigid. And we could further use these structure
20
References
1. Pollastri G, Baldi P, Fariselli P, Casadio R. Prediction of coordination number and relative solvent accessibility in proteins. Proteins 2002;47(2):142-153. 2. Rost B, Sander C. Conservation and prediction of solvent accessibility in
protein families. Proteins 1994;20(3):216-226.
3. Chan HS, Dill KA. Origins of structure in globular proteins. Proc Natl Acad Sci U S A 1990;87(16):6388-6392.
4. Porollo A, Meller J. Prediction-based fingerprints of protein-protein interactions. Proteins 2007;66(3):630-645.
5. Young KH. Yeast two-hybrid: so many interactions, (in) so little time. Biol Reprod 1998;58(2):302-311.
6. Crowell RE, Du Clos TW, Montoya G, Heaphy E, Mold C. C-reactive protein receptors on the human monocytic cell line U-937. Evidence for additional binding to Fc gamma RI. J Immunol 1991;147(10):3445-3451.
7. Jares-Erijman EA, Jovin TM. FRET imaging. Nat Biotechnol 2003;21(11):1387-1395.
8. Lo Conte L, Chothia C, Janin J. The atomic structure of protein-protein recognition sites. J Mol Biol 1999;285(5):2177-2198.
9. Neuvirth H, Raz R, Schreiber G. ProMate: a structure based prediction program to identify the location of protein-protein binding sites. J Mol Biol 2004;338(1):181-199.
10. Zhou HX, Shan Y. Prediction of protein interaction sites from sequence profile and residue neighbor list. Proteins 2001;44(3):336-343.
11. Halle B. Flexibility and packing in proteins. Proc Natl Acad Sci U S A 2002;99(3):1274-1279.
12. Jones S, Thornton JM. Protein-protein interactions: a review of protein dimer structures. Prog Biophys Mol Biol 1995;63(1):31-65.
13. Levitt M, Warshel A. Computer simulation of protein folding. Nature 1975;253(5494):694-698.
14. Warshel A. Bicycle-pedal model for the first step in the vision process. Nature 1976;260(5553):679-683.
15. McCammon JA, Gelin BR, Karplus M. Dynamics of folded proteins. Nature 1977;267(5612):585-590.
16. Pandey BP, Zhang C, Yuan X, Zi J, Zhou Y. Protein flexibility prediction by an all-atom mean-field statistical theory. Protein Sci 2005;14(7):1772-1777. 17. Rueda M, Ferrer-Costa C, Meyer T, Perez A, Camps J, Hospital A, Gelpi JL,
21 2007;104(3):796-801.
18. Shih CH, Huang SW, Yen SC, Lai YL, Yu SH, Hwang JK. A simple way to compute protein dynamics without a mechanical model. Proteins 2007;68(1):34-38. 19. Lin CP, Huang SW, Lai YL, Yen SC, Shih CH, Lu CH, Huang CC, Hwang JK.
Deriving protein dynamical properties from weighted protein contact number. Proteins 2008;72(3):929-935.
20. Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, Shindyalov IN, Bourne PE. The Protein Data Bank. Nucleic Acids Res 2000;28(1):235-242. 21. Shindyalov IN, Bourne PE. Protein structure alignment by incremental
combinatorial extension (CE) of the optimal path. Protein Eng 1998;11(9):739-747.
22. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 1997;25(17):3389-3402.
23. Mintseris J, Weng Z. Structure, function, and evolution of transient and obligate protein-protein interactions. Proc Natl Acad Sci U S A 2005;102(31):10930-10935.
24. Mintseris J, Weng Z. Atomic contact vectors in protein-protein recognition. Proteins 2003;53(3):629-639.
25. Johnson M, Zaretskaya I, Raytselis Y, Merezhuk Y, McGinnis S, Madden TL. NCBI BLAST: a better web interface. Nucleic Acids Res 2008;36(Web Server issue):W5-9.
26. Rose GD, Geselowitz AR, Lesser GJ, Lee RH, Zehfus MH. Hydrophobicity of amino acid residues in globular proteins. Science 1985;229(4716):834-838. 27. Kabsch W, Sander C. Dictionary of protein secondary structure: pattern
recognition of hydrogen-bonded and geometrical features. Biopolymers 1983;22(12):2577-2637.
28. Jones S, Thornton JM. Analysis of protein-protein interaction sites using surface patches. J Mol Biol 1997;272(1):121-132.
29. Landau M, Mayrose I, Rosenberg Y, Glaser F, Martz E, Pupko T, Ben-Tal N. ConSurf 2005: the projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res 2005;33(Web Server issue):W299-302.
30. Smith TF, Waterman MS. Identification of common molecular subsequences. J Mol Biol 1981;147(1):195-197.
22
Figures
23
24
25
(A)
(B)
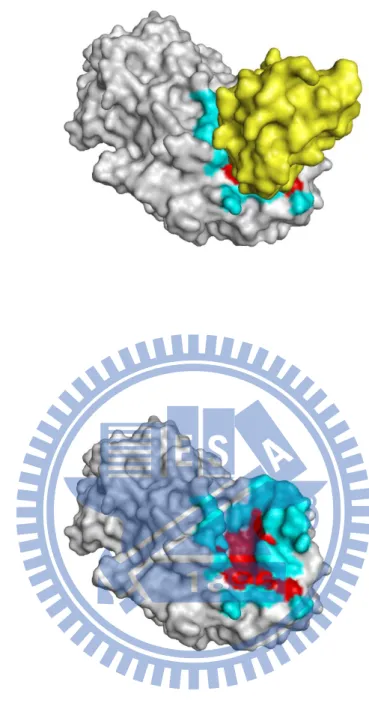
Figure 4. Visualization of the core interface and the peripheral interface (PDB entry 1tmq) (A) Chain A and chain B. (B) Chain A only. Visual graphics tool
Pymol was used to visualize the core interface and peripheral interface of 1tmqA. The
chain A was shown in gray and chain B was shown in yellow. The core interface and
26
(A)
(B)
Figure 5. Comparison of the amino acid distributions between core interfaces and
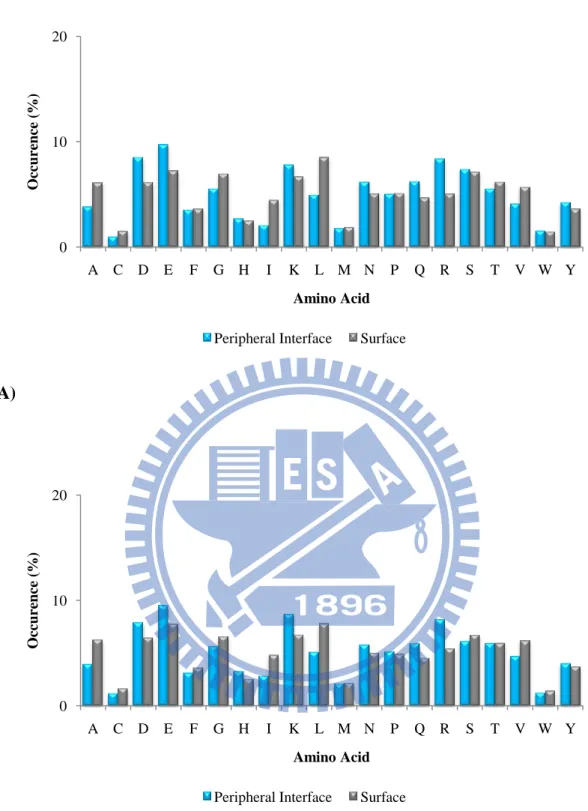
surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Core Interface Surface
0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Core Interface Surface
27
(A)
(B)
Figure 6. The amino acid propensities of core interfaces in unbound state of (A) the
ProMate database (B) the ZW database. 0.0 0.5 1.0 1.5 2.0 2.5 C W H M I Y V L S A F G Q P N T R D E K P ro pens it y Amino Acid Core Interface 0.0 0.5 1.0 1.5 2.0 2.5 C W F Y L I M V S G H T A N P Q D R E K P ro pens it y Amino Acid Core Interface
28
(A)
(B)
Figure 7. Comparison of the amino acid distributions between peripheral interfaces
and surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Peripheral Interface Surface
0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Peripheral Interface Surface
29
(A)
(B)
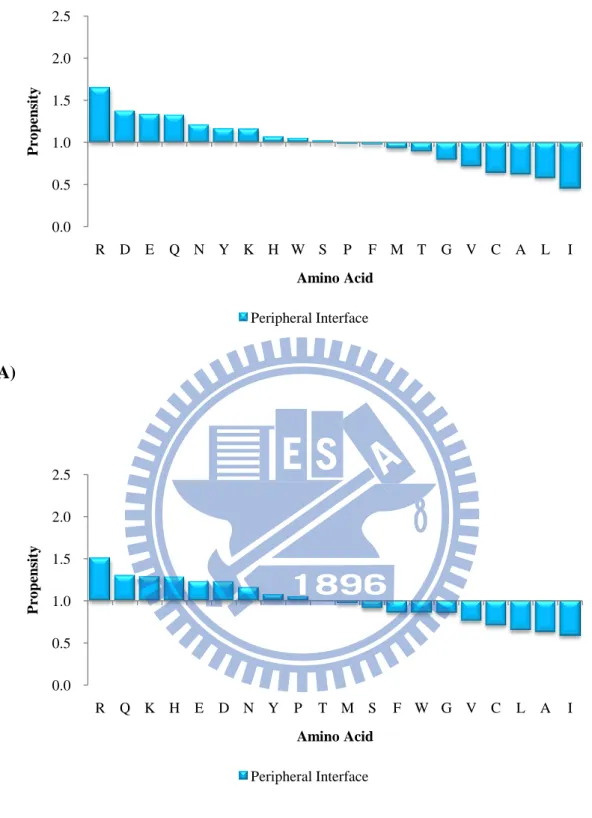
Figure 8. The amino acid propensities of peripheral interfaces in unbound state of (A)
the ProMate database (B) the ZW database. 0.0 0.5 1.0 1.5 2.0 2.5 R D E Q N Y K H W S P F M T G V C A L I P ro pens it y Amino Acid Peripheral Interface 0.0 0.5 1.0 1.5 2.0 2.5 R Q K H E D N Y P T M S F W G V C L A I P ro pens it y Amino Acid Peripheral Interface
30
(A)
(B)
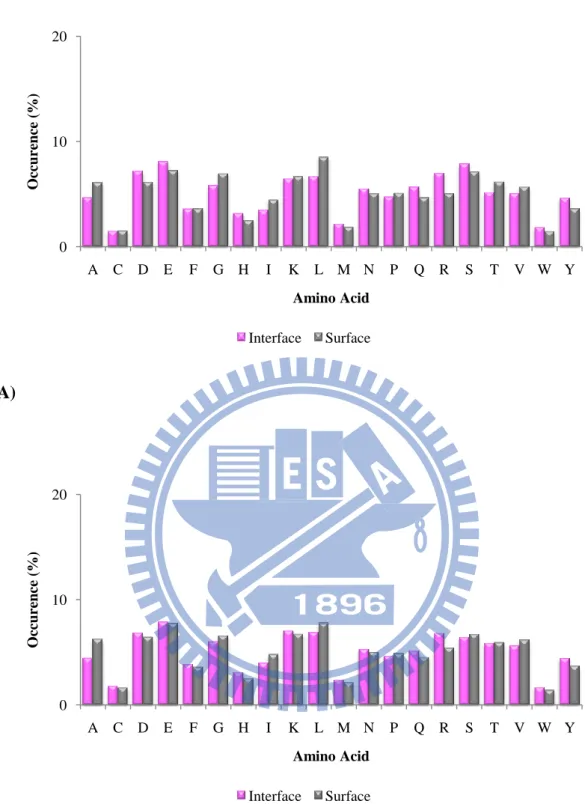
Figure 9. Comparison of the amino acid distributions between interfaces and surfaces
in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Interface Surface 0 10 20 A C D E F G H I K L M N P Q R S T V W Y O cc urence ( %) Amino Acid Interface Surface
31
(A)
(B)
Figure 10. The amino acid propensities of interface in unbound state of (A) the
ProMate database (B) the ZW database. 0.0 0.5 1.0 1.5 2.0 2.5 R Y W H Q D M E S N F C K P V G T L I A P ro pens it y Amino Acid Interface 0.0 0.5 1.0 1.5 2.0 2.5 R H Y W Q C M F D N K E T S P G V L I A P ro pens it y Amino Acid Interface
32
(A)
(B)
Figure 11. Comparison of the secondary structure distributions between core
interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure Core Interface Surface
0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure
33
(A)
(B)
Figure 12. Comparison of the secondary structure distributions between peripheral
interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure Peripheral Interface Surface
0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure Peripheral Interface Surface
34
(A)
(B)
Figure 13. Comparison of the secondary structure distributions between interfaces and
surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure Interface Surface 0 10 20 30 40 50 60 70
helix strand loop
O cc urence ( %) Secondary Structure Interface Surface
35
(A)
(B)
Figure 14. Comparison of the distributions of solvent accessibility between core
interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 5 10 15 20 O cc urence ( %) Accessibility (%) Core Interface Surface
0 5 10 15 20 O cc urence ( %) Accessibility (%) Core Interface Surface
36
(A)
(B)
Figure 15. Comparison of the distributions of solvent accessibility between peripheral
interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 5 10 15 20 O cc urence ( %) Accessibility (%) Peripheral Interface Surface
0 5 10 15 20 O cc urence ( %) Accessibility (%) Peripheral Interface Surface
37
(A)
(B)
Figure 16. Comparison of the distributions of solvent accessibility between interfaces
and surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 5 10 15 20 O cc urence ( %) Accessibility (%) Interface Surface 0 5 10 15 20 O cc urence ( %) Accessibility (%) Interface Surface
38
(A)
(B)
Figure 17. Comparison of the distributions of accessibility change between core
interfaces and interfaces in complexation of (A) the ProMate database (B) the ZW
database. 0 5 10 15 20 25 O cc urence ( %) DAccessibility (%) Core Interface Interface
0 5 10 15 20 25 O cc urence ( %) DAccessibility (%) Core Interface Interface
39
(A)
(B)
Figure 18. Comparison of the distributions of accessibility change between peripheral
interfaces and interfaces in complexation of (A) the ProMate database (B) the ZW
database. 0 5 10 15 20 25 O cc urence ( %) DAccessibility (%) Peripheral Interface Interface
0 5 10 15 20 25 O cc urence ( %) DAccessibility (%) Peripheral Interface Interface
40
(A)
(B)
Figure 19. Comparison of the distributions of amino acid conservation scores between
core interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Core Interface Surface
0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Core Interface Surface
41
(A)
(B)
Figure 20. Comparison of the distributions of amino acid conservation scores between
peripheral interfaces and surfaces in unbound state of (A) the ProMate database (B)
the ZW database. 0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Peripheral Interface Surface
0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Peripheral Interface Surface
42
(A)
(B)
Figure 21. Comparison of the distributions of amino acid conservation scores between
interfaces and surfaces in unbound state of (A) the ProMate database (B) the ZW
database. 0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Interface Surface 0 10 20 30 40 -3 -2 -1 0 1 2 3 4 O cc urence ( %)
Conservation Score (normalized) Interface Surface
43
Figure 22. Comparison of the B-factor distributions between core interfaces and
surfaces in unbound state of the ProMate database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) B-factor (normalized) Core Interface Surface
44
Figure 23. Comparison of the B-factor distributions between peripheral interfaces and
surfaces in unbound state of the ProMate database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) B-factor (normalized) Peripheral Interface Surface
45
Figure 24. Comparison of the B-factor distributions between interfaces and surfaces in
unbound state of the ProMate database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) B-factor (normalized) Interface Surface
46
(A)
(B)
Figure 25. Comparison of the CM distributions between core interfaces and surfaces
in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Core Interface Surface
0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Core Interface Surface
47
(A)
(B)
Figure 26. Comparison of the CM distributions between peripheral interfaces and
surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Peripheral Interface Surface
0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Peripheral Interface Surface
48
(A)
(B)
Figure 27. Comparison of the CM distributions between interfaces and surfaces in
unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Interface Surface 0 10 20 30 -2 -1 0 1 2 3 4 5 6 O cc urence ( %) CM (normalized) Interface Surface
49
(A)
(B)
Figure 28. Comparison of the WCN distributions between core interfaces and surfaces
in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Core Interface Surface
0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Core Interface Surface
50
(A)
(B)
Figure 29. Comparison of the WCN distributions between peripheral interfaces and
surfaces in unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Peripheral Interface Surface
0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Peripheral Interface Surface
51
(A)
(B)
Figure 30. Comparison of the WCN distributions between interfaces and surfaces in
unbound state of (A) the ProMate database (B) the ZW database. 0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Interface Surface 0 10 20 30 -4 -3 -2 -1 0 1 2 3 O cc urence ( %) WCN (normalized) Interface Surface
52
(A)
(B)
Figure 31. Visualization of the B-factor of the core interface and the peripheral interface (PDB entry 1tmq) (A) The chain A of 1tmq was colored by B-factors. (B)
The chain A of 1tmq was colored in gray. The core interface and the peripheral
53
Figure 32. Visualization of the example of CM model (PDB entry: 1tmqA) The
core interface and peripheral interface were shown as sticks and colored in red and
cyan independently. The center of mass of chain A of 1tmq was pictured as sphere and
colored in green. The distance of the core interface residue, W56, and the center of
mass was 16.79 Å . The distance of the peripheral interface residue, E229, and the
54
55
Appendix
Homologous monomer Equivalent bound Homologous monomer Equivalent bound Homologous monomer Equivalent bound1a19A 1brsD 1eza_ 3ezaA 1pne_ 1hluP
1a2pA 1brsA 1eztA 1agrE 1poh_ 1ggrB
1a5e_ 1bi7B 1f00I 1f02I 1ppp_ 1stfE
1acl_ 1fssA 1f5wA 1kacB 1qqrA 1bmlC
1ag6_ 2pcfA 1fkl_ 1b6cA 1rgp_ 1am4A
1aje_ 1am4D 1flzA 1euiA 1selA 1cseE
1ajw_ 1cc0E 1fvhA 1dn1A 1vin_ 1finB
1aueA 1fapB 1g4kA 1ueaA 1wer_ 1wq1G
1avu_ 1avwB 1gc7A 1ef1A 1xpb_ 1jtgA
1aye_ 1dtdA 1gnc_ 1cd9A 2bnh_ 1a4yA
1b1eA 1a4yB 1hh8A 1e96B 2cpl_ 1ak4A
1bip_ 1tmqB 1hplA 1ethA 2f3gA 1ggrA
1ctm_ 2pcfB 1hu8A 1ycsA 2nef_ 1avzB
1cto_ 1cd9B 1iob_ 1itbA 2rgf_ 1lfdA
1cye_ 1eayA 1j6zA 1c0fA 3ssi_ 2sicI
1d0nA 1c0fS 1jae_ 1tmqA 6ccp_ 2pcbA
1d2bA 1ueaB 1lba_ 1aroL 1jtgB
1ekxA 1d09A 1nobA 1kacA
1ex3A 1cgiE 1nos_ 1nocA
1ez3A 1dn1B 1pco_ 1ethB
56
2prg B:C 1ebd AB:C 1g0y I:R
1h59 A:B 1e6e A:B 1ijk A:BC
1c4z A:D 1gaq A:B 1n2c AB:EF
1evt A:C 1f80 A:E 1mah A:F
1dn1 A:B 1stf E:I 1gcq B:C
1xdt R:T 1f02 I:T 1www VW:X
1t7p A:B 2btc E:I 1i2m A:B
1go4 A:G 1gh6 A:B 1kgy A:E
1i85 B:D 1rlb ABCD:E 1c1y A:B
1jma A:B 1aro L:P 1gl4 A:B
1kac A:B 1ak4 A:D 1d2z A:B
1gc1 C:G 1i3o ABCD:E 3ygs C:P
1f51 AB:E 1atn A:D 1cs4 AB:C
1kmi Y:Z 1dkg AB:D 1efu A:B
7cei A:B 1b6c A:B 3sgb E:I
1bvn P:T 1qo0 A:DE 1fqv A:B
1qkz A:HL 1ugh E:I 1k3z AB:D
1dpj A:B 1df9 B:C 1m4u A:L
1f83 A:BC 1jiw I:P 1m2o AC:B
1fak HL:T 1f93 AB:EF 1mbu A:C
1jw9 B:D 1noc A:B 1fc2 C:D
1jtd A:B 1hwg A:BC 1ml0 A:D
1d5x A:C 1fg9 AB:C 1gvn AC:B
1i4e A:B 1ebp A:CD 1o6s A:B
1ib1 AB:E 1du3 A:DEF 1h2k A:S
2pcc A:B 1euv A:B 1m1e A:B
1f3v A:B 1de4 CF:A 1o94 AB:CD
1lpb A:B 1ghq A:B 1nf5 A:B
1ay7 A:B 1flt VW:X 1gzs A:B
1kkl ABC:H 1gxd A:C 1nbf A:D
1dev A:B 1ycs A:B 1mr1 A:D
1l0o AB:C 1gla F:G
1dfj E:I 2sic E:I
1g4y B:R 1jsu AB:C
1jch A:B 1is8 ABEJCIDHGF:KLOMN