白英Solanum lyratum THUNBERG 粗抽物誘導人類大腸腺癌細胞(Colo 205)細胞毒性及細胞凋亡之分子機轉; The molecular mechanism of extracts of Solanum lyratum THUNBERG induced cytotoxicity and apoptosis in human colon adenocarcinoma cell lines (Colo 205)
53
0
0
全文
(2) 細胞毒作用,在經驗上 fluorouracil 合併使用包括 methotrexate、cisplatin、 N-(phosphon acetyl) -l-aspartic acid (PALA)、thymidine、interferon 及 leucovorin。 早期研究中宣稱具有很高的 regression rate。但是最近對於 fluorouracil 合併 cisplatin (6,7)、high-dose PALA(8)、high-dose thymidine (8)或 interferon a (9)治療的幾種 隨機臨床試驗結果,和單獨使用 fluorouracil 結果比較,對於大腸癌的治療並沒 有益處。 最近被提出治療方法中,對於大腸癌的存活率及減低局部的復發率較為肯定 的為 fluorouracil 對 radiation 治療,臨床前研究中指出 a continuous infusion of fluorouracil 比 the short courses of bolus administration 可能更有效率的增加放射治 療的敏感度(radiosensitization)(10),在為期三年的臨床試驗追蹤發現,在放射線治 療中同時給與 fluorouracil 利用 continuous infusion 比 bolus administration 具有 意義的減少復發率及增加存活率,並且局部復發比率也被降到只有 8%(11)。 Fluorouracil 為抗代謝抗癌藥物,對於源自於乳癌、頭頸部位、胃腸道、卵巢 的上皮細胞腫瘤(epithelial tumors)具有廣效的活性,單一藥物反應率(response rates)為10-30 %。細胞內fluorouracil被thymidine phosphorylase先轉換成floxuridine (FUdR),接著又被 thymidine kinase 磷酸化為活性成分5-fluoro-2'-deoxyuridine monophosphate (FdUMP)(12)。對於細胞週期影響fluorouracil可使細胞停於S期,不 影響G0或G1期,短暫誘導增加p53、p21蛋白,細胞週期cyclin A及cyclin A-cdk2 kinase activity增加 (13,14)。 在 chemopreventive agents 作用機轉方面,如 aspirin、other nonsteroidal anti-inflammatory drugs 及 selective cyclooxygenase-2 (COX-2) inhibitors,它們可 以藉由 cyclooxygenase (COX) 家族 COX-1 及 COX-2(15),將 arachidonic acid 轉 換成 prostaglandins。 aspirin 及 sulindac 可同時抑制 COX-1 及 COX-2,然而 celecoxib and rofecoxib 對於 COX-2 具有選擇性抑制作用,這些藥物可經由 COXdependent and COX-independent mechanisms 誘導 apoptosis;COX-2 的抑制可導致 arachidonic acid 增加,因此可刺激細胞膜上的 sphingomyelin 轉換成 ceramide, 2.
(3) ceramide 可誘導 apoptosis 產生 (16)。COX-2 的抑制也可經由改變 prostaglandin production 並且降低 angiogenic factors(17,18),達到誘導 apoptosis 作用。目前 sulindac sulfone(a metabolite of sulindac)可抑制 COX-1 及 COX-2 造成 coloncarcinoma cell lines 的 apoptosis 已經進入動物模式 (19),有一部分的作用可能是透 過抑制 NF-?B(20)或干擾 PPARd(peroxisome-proliferator-activated receptor)結合到 DNA 上進而誘導細胞產生凋亡作用 (21)。 白英( Solanum lyratum THUNB. ) 始載於神農本草經列為上品,功用為清熱利濕 解毒,臨床上用於濕熱黃疸、瀉痢、白帶、水腫、風濕關節疼痛、濕疹瘙癢、癰 疽惡瘡;現今用於多種癌症,包括肺癌、胃腸道癌症、子宮頸癌、膀胱癌… 等。 目前較為接受的作用有抗炎(anti-inflammation )、抗癌(anti-oncosis、anti-cancer), 可治療腸胃炎(enterogastritis)、咽喉炎(laryngitis)、扁桃腺炎(tonsillitis)、肝炎 (hepatitis)及關節炎(arrthritis and rheumatism)(22)。 因此,本實驗利用野生採集的白英,基原鑑定後採收種子,於田間栽培,採 收後用不同溶媒抽取,以不同溶媒的粗抽物作為實驗材料,處理人類大腸腺癌細 胞株 Colo 205,利用流式細胞計數儀體外評估細胞毒殺作用及誘導細胞凋亡的作 用機轉,並且利用 5-FU 為正對照組做比較,在分子機轉方面,使用 RT-PCR 及 Western Blot 方法,初步探討其抗腫瘤活性與初步的分子機轉,作為白英治療癌 症之參考。. 3.
(4) 第二章. 總. 論. 第一節 白英之文獻考察部分 一、白英之本草學考察 (一)白英藥名之考訂 白英之名最早見於神農本草經. (23). (24). 一名? 菜,爾雅稱苻、鬼目。別錄 稱. (36). (37). 白草。本草綱目 釋名中始有排風、白幕。綱目拾遺 稱天燈籠、和尚頭草。 植物名實圖考. (38). 有望冬紅、酸尖菜之名。綱目云:「白英謂其花色,毅菜象其. 葉文,排風言其功用,鬼目象其子形。」說文:「? ,細縛也。」縛,即白色 細絹,用以形容白英被覆細長的柔毛。因此以白英為其植物名較恰當。. (二)白英形態與產地之考訂 白英形態描述首見於爾雅注釋: 「似葛,葉有毛,子赤如耳璫珠,若云子 熟黑,誤矣。」新修本草. (26). 云:「此鬼目草也。蔓生,葉似王瓜,小長而五椏。. 實圓若龍葵子,生青,熟紫黑。」然而開寶重定本草. (29). 今按陳藏器本草云: 「爾. 雅云:苻,鬼目。注:似葛,葉有毛,子赤如耳璫珠,若云子熟黑,誤矣」 。綱 目. (36). 收於草部,集解中引別錄曰:「鬼目一名來甘。實赤如五味,十月採。」. 時珍曰:「此俗名排風子是也。正月生苗,白色,可食。秋開小白花,子如龍 (37). 葵子,熟時紫赤色。」綱目拾遺 云:「莖、葉皆有白毛,八、九月開花藕合 色,結子生青熟紅,鳥雀喜食之。」 綜合諸家本草所述形態,為蔓生植物,葉片心形或有五淺裂覆有白色細 毛,果實成熟為紅色多數,花白色,再參閱綱目及植物名實圖考. (38). 之附圖,與. 今茄科植物白英 Solanum lyratum THUNBERG 特徵相符。本草言產地為益州山 谷,為今四川成都山谷。. (三)性味、藥能之考訂 (23). 白英為本經. 上品藥,本草皆言味甘性寒。唯植物名實圖考 4. (38). 言其味酸。.
(5) (23). 別錄以降正統本草皆言無毒。本經 :「主寒熱八疸,消渴,補中益氣,久服 輕身延年。」 。本草經集注 解勞。」。本草拾遺 寶本草. (29). (27). (25). (26). :「葉作羹飲,甚療勞。」 。新修本草. :「煮汁飲,. :「主煩熱,風疹,丹毒,瘧瘴寒熱,小兒結熱。」 。開. :「別本注云:莖葉煮粥,極解熱毒。」 。綱目拾遺. (37). :「止血淋、瘧、. 疝氣。汁滴耳中,止膿不乾。入藥內,保腫毒不大。治瘰? 串,用煮牛肉精者 (38). 食之。」「清濕熱,治黃疸水腫,小兒蛔結腹痛。」。植物名實圖考 :「治腰 痛。」 總結本草所言白英功能為清熱利濕、解毒消腫、袪風止痛、益氣止渴。 主治寒熱消渴、風疹丹毒、瘧瘴黃疸、頭風頭痛、血淋腹痛、耳膿。. (四)白英修治方用及禁忌之考訂 (24). 白英莖、葉、花、根、果實皆可用,因此名醫別錄 云: 「白英無毒。春 (25). 採葉,夏採莖,秋採花,冬採根。」 。但陶弘景集注神農本草經 云:「諸方藥 不用。…又有白草,葉作羹飲,甚治勞,而不用根、華。」只用葉不用根,嘉 (36). 祐用莖、葉,綱目. 附方引聖濟錄用果實治風熱上攻,因此白英莖、葉、花、. 根、果實皆可用,應該全草入藥。地上部份應該於春夏採收,日曬乾,煮汁飲。 綜合諸家本草所述形態,確定為茄科植物白英 Solanum lyratum THUNBERG。 味甘性寒,無毒。功能清熱利濕、解毒消腫、袪風止痛、益氣止渴。主治寒熱 消渴、風疹丹毒、瘧瘴黃疸、頭風頭痛、血淋腹痛、耳膿。藥用部位為莖、葉、 花、根、果實皆可用,可全草入藥。地上部份應該於春夏採收,日曬乾,煮汁 飲。. 本草綱目. 植物名實圖考長編. 圖 一 白英之本草附圖 5.
(6) 後漢 神農本草經. 白英。? 菜。. 爾雅 苻。 魏晉 名醫別錄. 南北朝 陶弘景集注神農本草經(500AD). 唐 新修本草(659AD). 白英。白草。. 白英。? 菜,白草。. 白英。? 菜、白草、鬼目草。. 本草拾遺 白英。白幕、排風。 重廣英公本草 白英。? 菜、白草、鬼目草。. 宋 開寶重定本草(974AD). 白英。? 菜、白草、鬼目草、苻。. 宋 嘉祐補注神農本草經(1061AD). 白英。? 菜、白草、鬼目草、苻。. 聖濟錄 白英。排風子。. 宋 經史證類備急本草(1098AD). 白英。? 菜、白草、鬼目草、苻。. 宋 經史證類大觀本草(1108AD). 白英。? 菜、白草、鬼目草、苻。. 宋 重修政和經史證類備用本草(1249AD). 白英。? 菜、白草、鬼目草、苻。. 明 本草品彙精要(1505AD). 白英。? 菜、白草、鬼目。. 明 本草綱目(1583AD) 白英。? 菜、白草、白幕、排風 、子名鬼目。 清 本草綱目拾遺(1725AD) 白英。天燈籠、和尚頭草。. 清 植物名實圖考(1848AD). 白英。苻、鬼目、排風子、鬼目菜、望冬紅、酸尖菜。. 清 植物名實圖考長編(1848AD). 圖 二 白英之本草系統圖. 6.
(7) 二、白英之生藥學考察 1. 1970 年東丈夫等報導白英之生藥學研究,建立莖部及根部的藥材組織構造圖 (39)。 2. 1995 祁振聲認為花鏡中藤本蔓延,莖生白毛的雪下紅決非忍各科莢? 屬的高大 灌木或喬木,亦非紫金牛紫金牛屬、產於冬無降雪地區的直立灌木,而是與白 英形態極相近,僅葉形有區別的茄屬細弱草質藤本千年不爛心 Solanum cathayanum。從植物名實圖考的文字描述及插圖可以判斷,千年不爛心是一種 與白英的形態特徵截然不同植株光滑無毛、葉邊緣決不分裂的蔓生灌木,其原 物不可能是莖葉被白毛、與白英僅有葉形不同 S. cathayanum,而只能是現代 分類學上的海桐葉白英 S. pittosporifolium(40)。 3. 1995 祁振聲認為白英的絕大多數性狀均與花鏡中雪下紅的描述吻合,只是白英 的時常裂為琴形,與山茶的葉形差異較大。而現名千年不爛心的同屬植物 S. cathayanu C.Y. W U et S.C.H UANG,植株各部形態均與白英相近;而且葉為心形 或卵狀披針形,多數全緣,稀基部戴形 3 裂,與山茶的倒卵形葉片較接近。故 可確認,該植物即雪下紅的原植物 (40)。 4. 2000 年張樹人認為始載於神農本草經列為上品的白英和列為中品的蜀羊泉,不 該當一種藥物白毛藤使用。現代藥學專著將白英稱蜀羊泉或白毛藤,將千年不 爛心又稱蜀羊泉或白毛藤,如此混稱不妥。古本草記載的白英應為茄科茄屬植 物白英 Solanum lyratum,而蜀羊泉為同屬的千年不爛心 Solanum dulcamara(41)。 5. 1981、1985 年 Murakami K, et al. 分離白英莖中主要含有 steroid glycoside 分為 (42), (1)SL-a 又名 SL-0 含有 4 種成分,分別為﹝(5a,25ß)-furostan-3,22,26-triol﹞、 ﹝(5a,25a)-furostan-3,22,26-triol﹞、﹝(25ß)-furost-en-3,22,26-triol﹞、 ﹝(25a)-furost-5-en-3,22,26-triol﹞的﹝3-O-ß-D-glucopyranosyl (1? 2 )-ß-Dglucopyranosyl ( 1? 4 )-ß-D-galactopyranoside-26-O-ß-D-glucopyranoside﹞。 (2) SL-b 又名 SL-1 含有 4 種成分 tigogenin、neotigogenin、diosgenin、yamogenin 皂? 元的﹝3-O-ß-D-. 7.
(8) glucopyranosyl (1? 2 )-ß-D-glucopyranosyl ( 1? 4 )-ß-D-galacto- pyranoside﹞ 組成。 (3) SL-c 含有 2 種成分 ﹝(25?) – solanidan-3ß,23ß-diol﹞、﹝(25?) – solanid-5-en-3ß,23ß-diol﹞的 ﹝3-O -ß-D- glucopyranosyl (1? 2 )-ß-D-glucopyranosyl ( 1? 4 )-ß-D-galacto pyranoside﹞。 (4) SL-d 含有 2 種成分 ﹝(25?) – solanidan-3ß,23ß-diol﹞、﹝(25?) – solanid-5-en-3ß,23ß-diol﹞的 ﹝3-O-ß-D-glucopyranosyl (1? 2 )-ß-D-glucopyranosyl ( 1? 4 )-ß-D –galactopyranoside﹞。 6. 1985、1986、1989 年 Yanara S, et al. (43)分離出白英含 aspidistrin、methylprotoaspidistrin、﹝(22R)3ß,16ß,22,26-tetrahydroxycholest-5-ene- 3-O-a-L- rhamnopyranosyl (1? 2 )-ß-D-galactopyranoside﹞、26-O-ß-D- lucopyranosyl- (22?,25R)3ß,22,26-trihydroxy-furost-5-ene-3-O-a-L-rhamno pyranosyl (1? 2 )-ß-Dglucopyranosyl ( 1? 3 )-ß-D-glucuronopyranoside﹞、26-O-ß- D-glucopyranosyl(22?,25R)-3ß,26-dihydroxy-furost-5-ene-3-O- a-L-rhamno pyranosyl (1? 2 )-ß-Dglucuronopyranoside﹞、26-O-ß-D-glucopyranosyl- (22?,25S)-3ß,26-dihydroxyfurost-5-ene-3-O-a-L-rhamnopyranosyl (1? 2 )- ß-D-glucuronopyranoside﹞、 3-O-a-L-rhamnopyranosyl (1? 2 )-ß-D- glucurono pyranosyl-3ß-hydroxy-(25R)spirost-5-ene﹞、3-O-a-L-rhamnopyranosyl (1? 2 )- ß-D-glucuronopyranosyl-3ßhydroxy- (25S)-spirost-5-ene﹞。 7. 1997 年 Yung-Yung LEE 等從新鮮的地上部分,分離出兩種 steroid alkaloidal glycosides 命名為 solalyratine A and B(44)。 8. 2002 楊敬之等從乙酸乙酯萃取層中分離出 caffeic acid、vanillic acid、 N(p-hydropheneethyl) p-coumaramide,從水飽合正丁醇萃取層中分離出 rutin(45)。 8.
(9) 三、白英藥理作用文獻考察 (一) 抗腫瘤作用 1965 年 Kupchan S. H.等提出,在體外,SL-c 和 SL-d 對人子宮頸癌 JTC 26 細胞有明顯抑制作用,8 µg/mL 抑制率可達 100%,SL-b 也表現抑制作用,15 µg/mL 抑制率可達 100%,但化學結構與 SL-b 相似的 SL-a 對 JTC 26 卻沒有抑 制作用 (46)。 1988 年曹濟遠及譚湘陵報導,利用融合技術研究白英對 CHO 細胞 G2-PC 染色體損傷的觀察,發現白英濃度 0.5∼1.0 kg/L,37 ℃處理 45 分鐘後,G2-PC 染色體有明顯損傷,可明顯阻斷 G2 期細胞,但各個劑量白英對 M 期細胞無明 顯作用 (47)。 1989 年 Ushio R, et al.提出白英熱水提取物具有抗腫瘤作用 (48)。 1989 張永健報導白英複方注射液對人胃癌細胞 BGC-823 和人乳腺癌 MCF-7 均有較強的殺傷作用,流 式細胞計數儀測定,可使 BCG-823 細胞停留 在 G1 期,導致 S 期細胞顯著減少,用 3H-TdR 試驗發現能抑制 BCG-823 細胞 DNA 合成 (49)。 1996 年 Hsu, Shu-Hui 報導,solamagine 對於 human hepatocyte( Hep 3B) 具 有強的細胞毒殺作用,且具有誘導細胞凋亡的作用,作用機轉可能和 TNF receptorⅠ上調有關 (50)。 1998 年 Chang, Li-Ching 報導,solamargine 分子結構中的 2' rhamnose moiety 對於細胞凋亡機轉,可能扮演重要的作用點,藉由 solamargine 的分子 模式發現不同的 carbohydrate 部分對於 TNFR Ⅰ和Ⅱ 表現亦不相同,且只 有 solamargine 具有誘導細胞凋亡作用 (51)。 2001 年單長民等提出,白英乙酸乙酯提取物對人肝癌 BEL-7404 細胞有誘 導凋亡作用,在培養基終濃度為 2.5 µg/mL、5.0 µg/mL、10 µg/mL 有誘導肝癌 BEL-7404 細胞凋亡作用,光學顯微鏡觀察及凝膠電泳均有細胞凋亡的特徵(52)。 9.
(10) 2002 年施文榮及劉豔報導,白英水提液在 1 mg/mL 濃度及以上時對人白 血病細胞株 HL-60 的生長也表現出顯著的抑制作用。白英水提液具有較強的 體外抑瘤活性,而其抑瘤活性並不限於直接的細胞毒作用 (53)。. (二) 抗過敏作用 (54) 1998 年 Kang, Bookyung 提出,白英水抽物具有強烈 antianaphylactic activity。白英水抽物在 1.0 mg/g body weight (BW)劑量時,可抑制由 compound 48/80 誘導的 anaphylactic shock 反應,於 0.0001-1.0 mg/g BW 劑量下血漿中的 histamine levels 減低且有劑量依賴效應,口服白英水抽物對於 passive cutaneous anaphylaxis 有抑制作用;再者白英水抽物對於藉由 compound 48/80 所誘導的 histamine 釋放具有抑制作用且有劑量依賴效應,機轉可能和增加 cAMP 量有關。. (三) 對於 NO 合成的影響 (55) 1999 年 H.M. Kim 提出,白英本身不會誘導 NO 合成,但熱水粗抽物合併 recombinant interferon-? 處理 mouse peritoneal macrophages 可導致 NO 合成增 加,因此白英對 interferon-? 誘導 NO 合成增加有明顯的協同作用,且有劑量 遞增效應(dose-dependent manner),於合併處理後 6 小時達到最大作用,其作 用機轉為透過 single transduction pathway of PKC activation。. (四) 抗肝損傷作用 (56) 1998 年 Kang SY 提出,Scopoletin (7-hydroxy-6-methoxycoumarin)在體外 有意義的降低由四氯化碳引起的大鼠肝細胞釋放 glutamic pyruvic transaminase and sorbitol dehydrogenase。 scopoletin 有意義的保留細胞內 glutathione 含 量及 superoxide dismutase 活性﹐並且可以抑制 malondialdehyde 產生。. 10.
(11) 第二節 細胞週期(Cell cycle)調控之影響 一、細胞週期 細胞由一個增殖為二的過程稱為細胞週期。細胞開始進行細胞核分裂到細 胞分裂成兩個獨立子細胞時期稱為細胞分裂期Mitosis (M) phase,兩個細胞分 裂期之間稱為間期(Interphase),間期又可分為Gap 0, Gap 1, S , G 2 phase。因此 完整的細胞週期包含兩個時期為間期Interphase (G1, S, G2 phase) 及 細胞分裂 期Mitosis (M) phase。細胞週期進行當中,可有3個檢查點checkpoints來決定細 胞週期的走向是停滯(arrest) 或前進(progression)。 Gap 0 ( G0 phase):細胞處於靜止期 sensecence,可能為暫時性或永久性的停 止生長,若細胞已發展到最後階段便不再生長(如 neuron)。 Gap 1 ( G1 phase ):此期處於 M phase 及 S phase 之中間期,細胞開始生長, 同時產生 RNA 及合成蛋白質,複製胞器,細胞體積增 加,目的是為 DNA 複製做好準備。 G1 進入 S 期為第一個 checkpoint。 Synthesis (S) phase:DNA 合成及複製使 DNA 含量增加一倍。 Gap 2 ( G2 ) phase:DNA 複製到有絲分裂的期間,細胞會持續生長並產生新 的蛋白質。G2 進入 M 期為第二個 checkpoint。 Mitosis (M) phase:細胞週期中最短的時期。細胞核及細胞質分裂。Mitosis phase 又可細分為五個時期 Prometaphase、Metaphase、 Anaphase、Telophase、Cytokinesis 。染色體排列於赤道 板上時期稱為 Metaphase,此期為第三個 checkpoint (59)。 當 DNA 受損時,週期便無法通過 checkpoint 而停滯 ( arrest),此時細胞 內會進行 DNA 的修復,一旦修補完成時才進入下一期,若是發現有無法彌補 的錯誤時,細胞可能選擇走向凋亡 (apoptosis),以避免錯誤遺傳至下一代(60)。 11.
(12) 二、細胞週期調控相關分子 1. Cyclins 與 Cyclin-dependent kinases(Cdks) 調控細胞週期進行的蛋白主要為 cyclins,而調控 cyclins 活化的則是 cyclin-dependent kinases(Cdks)。當 cyclins 與 Cdks 形成複合物時,Cdks 才具有 激? 活性,可活化下游蛋白分子的作用,因此 cyclins-Cdks 複合物的結合與分解 調控整個細胞週期的進行。 在真核細胞中,可經由連續的細胞訊號,經過活化,與一系列特殊的 cyclins-Cdks(cyclins-Cdks complexes)的活化來調控細胞週期進行,使細胞增生。 但是細胞也會因為 DNA 損傷 (DNA damage)、細胞分化 (differentiation)、接觸 性抑制 (contact inhibition)、和衰老 (senecence),產生抑制增生(antipro- liferative) 訊號,促使 Cdks 的抑制者(CdkIs)負向調控細胞週期的進行 (61,62)。 Cyclins 家族,主要在細胞週期中被合成。目前已知至少有 8 種 cyclins 的存 在,分別為 A、B1,2,3、C、D1,2,3、E、F、G 及 H,他們在 N-端與 Cdks 鍵結的地 方都具有約 150 個胺基酸的相同區域,稱之為 cyclin-box。Cyclin C、D 以及 E, 主要存在於 G1 時期,並在 G1-S 的過渡期間被分解。而 cyclins A 與 B 被稱為 mitotic cyclins,穩定的存在於間期,但在 mitosis 時會迅速分解 (63)。 Cyclin-dependent kinases (Cdks)是一群蛋白激? 家族,會和特殊的 cyclins 結合而被活化。目前已知共有 7 種 Cdks 被發現,分別是 cdc 2(又稱為 Cdk1)、 Cdk2、3、4、5、6、7 。細胞在 G0/G1 時期,Cdk4、6 會和 cyclin D 家族結合; 而 Cdk2 也會與 cyclin D 家族結合,但主要還是在 G1 及 G1-S 過渡期與 cyclin A 及 E 結合。cdc 2 主要存在 S、G2 以及 M 時期,並與 cyclin A 及 B 結合。而 cyclin H 則會和 Cdk 7 形成一具有酵素活性的複合物,進而活化 cdc 2 (Cdk1) 及 Cdk2(64)。. 2. Cyclin-dependent kinases inhibitors(CdkIs) 細胞週期調控過程中,除了 cyclins 與 Cdks 的正向調控因子外,另有一家 12.
(13) 族擔任負向調控的角色,稱之為 cyclin-dependent kinases inhibitors(CdkIs) (65)。 CdkIs 主要可分為兩大族群,一為 INK4(means Inhibitors of Cdk4)家族,另 一為-CIP/KIP 家族 (66)。 在 INK4 家族中,主要成員有 p14、p15(INK4b),p16(INK4a),p18(INK4c)以及 p19(INK4d),其功能為選擇性抑制 cyclin D/Cdk4 與 cyclin D/Cdk6,主要機轉為 INK 家族蛋白上具有 ankyrin repeat domains 會和 cyclin D 共同競爭 Cdk4、Cdk6 上 的結合部位,使細胞停滯於 G1 期。 另一個族群為 KIP/CIP 家族,包括了 p21(CIP1/WAF1/SDI1),p27(KIP1)以及 p57(KIP2)。p21(CIP1/WAF1/SDI1) 和 p27(KIP1)對於細胞週期影響的範圍較 INK4 家族為 廣,其所調控的蛋白包括所有的 cyclin-Cdk complexes 包括 cyclin E/Cdk2、 cyclin D/Cdk1、cyclin D/ Cdk6、cyclin A/Cdk2 及 cyclin B/cdk1 等。主要作用機 轉為 p21、p27 和 cyclin-Cdk 形成 cyclin-Cdk-CdkI 三元體形式,遮蔽掉 Cdks 的 催化部位,使 cyclin-Cdk complexes 無法磷酸化下游分子,使細胞停滯。. 3. p53(67,68) p53 是一種腫瘤抑制基因,其所轉錄的蛋白 p53 則是一種能夠抑制腫瘤的蛋 白,此蛋白為一種轉錄因子,平時存在於細胞質中,並且和 MDM2 蛋白結合, 形成不活化狀態。當細胞的 DNA 發生損傷、缺氧、被致癌因子活化時,p53 會 被活化,p53 和 MDM2 分開,p53 單離出來,進入細胞核,座落到相關基因上 影響下游基因的表現,其影響包括 DNA 修復、細胞週期停止與細胞凋亡等。如 p53 活化可增加 WAF gene 轉錄作用,p21WAF1 蛋白質量增加,促使細胞週期停 留在 G1/S 期,直至 DNA 修復後才進入正常的週期。但是當 DNA 受損太嚴重 時,p53 則會促使細胞進入細胞凋亡。. 三、轉錄因子 NF-?B protein family 與 I-?B 調控 NF-?B 屬於Rel family為一種轉錄因子(transcription factors) ,在細胞發炎作. 13.
(14) 用、免疫反應、細胞週期的調控、細胞分化及細胞產生凋亡時受到活化產生 (69,70)。 哺乳類中NF-?B家族成員有 Rel family:Rel-A (p65)、Rel-B、c-Rel、NF-kB1 (p50:p105)及NF-kB2 (p52:p100) (71-76),NF-?B家族成員胺基酸末端均有延伸的 amino-terminal region 稱為 Rel homology domain (RHD),RHD 由leucine zipper dimerization domains、細胞核局部信號nuclear localization sequence(NLS)和 DNA-binding domain 組成 (77),此外Rel-A, c-Rel 及 Rel-B 具有carboxy-terminal transactivating domains (reviewed in refs.) (78)。 NF-?B protein 存在於細胞質中,並且和其抑制劑 I-?B family 形成二聚 體,I-?B family 有 I-?Ba 、I-?Bß 及 I-?Be,I-?B 蛋白可遮蓋掉 NF-?B protein 的核轉錄信號部位( NLS),當細胞受到刺激(TNF-a 或 IL-1ß),可引起一連串 的活化作用,使兩個主要的I-?B kinases〔IKK-1(IKKa)及IKK-2(IKKß)〕活化, IKK-1及IKK-2會活化I-?Bs的特定氨基酸殘基,被磷酸化的 I-?Bs 會藉由 E3 ubiquitin ligase 產生ubiquitination作用,接著I-?Bs 被 proteasome 分解,游離的 NF-?B 便可進入細胞核,影響相關基因轉錄作用,影響範圍包括細胞分化、細 胞存活、發炎反應,在細胞週期方面 NF-?B 會透過 cyclin D1 增加影響 G1期 (79). 。 目前研究中最為複雜的為 NF-?B1(p50:p105) 是一種多二聚體結構 (80)。. Rel-A(p65)可保護細胞免於凋亡。 NF-?B dimers 的活化必須先使 I-?B 降解,因 此 I-?B 的降解為控制 NF?B 活性最重要的機轉。稍後的步驟包括磷酸化反應及 和p300:CBP、p:CAF、members of the p160 coactivator family及proteins of the basal transcriptional apparatus的交叉活化反應(transactivation),如此可使NF-kB 訊號 和 其它信號傳導途徑做整合及關聯 (81-83)。然而,不少文獻亦指出這些化學藥物在引 起癌細胞的凋亡同時,會相對的活化了 transcription factor NF-?B或其他因子,進 而引發與促進細胞存活(cell survival)有關的基因的表現、並阻止癌細胞的凋亡 (84-86). 。. 14.
(15) 第三節 細胞凋亡(Apoptosis;programmed cell death) 一、細胞凋亡的定義 Kerr, Wyllie 及 Currie 於 1992 年提出 apoptosis 這個名詞,apoptosis 是從 希臘字 apo(away from) 及 ptosis(falling) ,意思為葉片從樹上凋落下來 (87)。細 胞凋亡 Apoptosis 又稱為計畫性死亡 programmed cell death,apoptosis 在生物體 內扮演相當重要的角色。從生理上的觀點,在個體的發生( development) 、生長 (growth)、分化(differentation)過程中,細胞凋亡有助於維持生物體內恆定(homeostasis)的功用 (88)。 從病理方面觀點,apoptosis 是細胞對於外來病毒攻擊或破壞的一種死亡模 式;然而,過度的細胞凋亡(excess of apoptosis)通常伴隨著大量細胞數目的 減少及病變(例如 neurodegenerative disease) (89)。反之,若生物體有缺陷而不 能正常的進行細胞凋亡( defects of apoptosis) ,則會引起細胞不正常的增生或導 致癌細胞的發生。 目前,一部分的癌症化學治療藥物之作用機轉是利用抑制癌細胞複製之機 制,誘導癌細胞的細胞週期停滯和細胞凋亡以達到其抗癌的目的(90)。然而,不 少文獻指出這些化學藥物在引起癌細胞凋亡的同時,亦會相對的活化 transcription factor(NF-?B)或其它因子,進而引發與促進細胞存活(cell survival)有 關基因的表現,並阻止了癌細胞的凋亡 (91-94)。另外由於癌細胞本身基因的不穩 定性,造成了 tumor suppressor gene p53 的不活化(inactivation) (95,96),或是持 續活化 oncogene family 如 Bcl-2 families genes(97,98),而增加了癌細胞的抗藥性, 這些現象都會導致藥效降低。因此抗癌藥物的藥效除了多重藥物抗藥性蛋白 multidrug resistance proteins( MRP)的影響外 (99),亦與癌細胞的抗凋亡(resistance to apoptosis)能力有關。. 15.
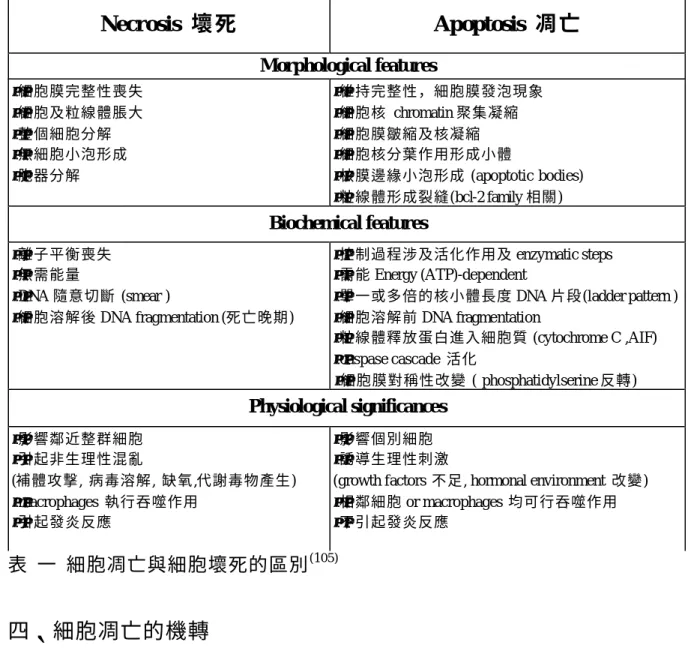
(16) 二、細胞凋亡的形態學變化 細胞凋亡基於形態學的表現來定義,雖然基因及生化標誌不斷的增加取代 了形態學方面的地位,但是目前並無標準的黃金準則(100)。細胞凋亡形態學上變 化,早期時,會在細胞核周圍出現染色質濃縮( chromatin condensation),細胞 膜周圍突起會回縮,細胞變圓,接著細胞核分葉作用產生( fragmentation of nuclear ),當 lamins and actin filaments 被分解後細胞開始皺縮( shrinkage ),細 胞膜形成小泡( membrane blebbing );到了晚期,細胞核會開始發生裂解,然後 形成凋亡小體( apoptotic bodies )。在細胞凋亡過程中,並不會發生胞器或是胞 膜破裂,但是粒線體膜電位下降及通透性增加可能發生(101)。當細胞產生凋亡 時,粒線體膜電位下降失去他原有製造 ATP 的能力時,此時細胞膜無法維持其 內外層的對稱性,此時大量分佈於細胞膜內層的 phosphatidylserine( PS ) 會翻 轉到外層,外層分佈較多的( PS ),此時在體內可為吞噬細胞所吞噬( phagocytosis )。體外培養凋亡細胞最後仍走向壞死( necrosis )方向 (102)。. 三、細胞凋亡與細胞壞死( necrosis )之區別 細胞壞死會同時發生在一群細胞,也可分為早期與晚期兩個時期。在細胞 壞死早期,細胞內之胞器會脹大,細胞膜也會破裂;到了晚期細胞膜也發生破 裂。由於細胞發生破裂,因此會釋放出一些發炎物質而引起局部發炎反應,傷 害到鄰近的細胞或組織 (103,104)。 細胞凋亡與細胞壞死( necrosis )之區別 如下表 (105). 16.
(17) Necrosis 壞 死. Apoptosis 凋 亡 Morphological features. •細胞膜完整性喪失 •細胞及粒線體脹大 •整個細胞分解 •無細胞小泡形成 •胞器分解. •維持完整性,細胞膜發泡現象 •細胞核 chromatin 聚集凝縮 •細胞膜皺縮及核凝縮 •細胞核分葉作用形成小體 •核膜邊緣小泡形成 (apoptotic bodies) •粒線體形成裂縫(bcl-2 family 相關). Biochemical features •離子平衡喪失 •無需能量 •DNA 隨意切斷 (smear ) •細胞溶解後 DNA fragmentation (死亡晚期). •控制過程涉及活化作用及 enzymatic steps •需能 Energy (ATP)-dependent •單一或多倍的核小體長度 DNA 片段(ladder pattern ) •細胞溶解前 DNA fragmentation •粒線體釋放蛋白進入細胞質 (cytochrome C ,AIF) •caspase cascade 活化 •細胞膜對稱性改變 ( phosphatidylserine 反轉). Physiological significances •影響鄰近整群細胞 •引起非生理性混亂 (補體攻擊, 病毒溶解, 缺氧,代謝毒物產生) •macrophages 執行吞噬作用 •引起發炎反應. •影響個別細胞 •誘導生理性刺激 (growth factors 不足, hormonal environment 改變) •相鄰細胞 or macrophages 均可行吞噬作用 •不引起發炎反應. 表 一 細胞凋亡與細胞壞死的區別 (105). 四、細胞凋亡的機轉 目前已知有兩個機轉可以調控細胞凋亡現象,第一個作用機轉是經由 caspase( caspase-dependent )機轉活化而引起細胞凋亡 (106)。第二個作用機轉 是經由 caspase-independent 機轉。 (一) caspase dependent 機轉: 又包含兩種途徑 (1)The extrinsic apoptotic pathway 外在路徑 經由細胞外死亡接受體(death receptor)如 TNFR1(tumor necrosis factor receptor)、CD95(Fas)或 TRAIL( TNF-related apoptosis inducing ligand)等許 多訊息傳遞,外在路徑在細胞凋亡的過程中扮演相當重要的角色,最後活化 17.
(18) caspase cascade 而引起細胞凋亡,引起凋亡的速度較快。 1. CD95 (Fas receptor) 為 tumor necrosis factor receptor superfamily 家族成員,Fas receptor 和 Fas ligand 結合後產生 multimerisation,recruitment of adapter molecules 及 caspaseactivating complex。Fas ligand 為三合體結構,和 Fas receptor 結合促進 Fas receptor 形成三合體化(trimerisation),接著細胞膜內側的 receptor 稱 death domains(DD)開始成簇聚合,death domains 成簇聚合後使得轉接者蛋白 FADD(Fas-associated death domain )和 death domains 利用其上的 homologous death domains 相互作用結合。接下來 FADD 透過另一個 domain 將訊號轉送 給 DED(Death effector domain) ,具有 caspase-8 zymogen 。FADD-DD-DEDpro-caspase-8 complex 會使 pro-caspase-8 切除並且活化,被活化的 caspase -8 接著活化 caspases-3 and -7。經由此途徑產生凋亡的作用稱為 Death Induc- ing Signaling Complex 或 DISC。DISC 於生理性的細胞凋亡扮演重要角色 (107)。 2. TNFR1(tumor necrosis factor receptor 1) TNF 為被活化的 T 細胞及吞嗜細胞所產生,TNF 和位於細胞膜表面的 TNFR1 結合後,導致 TNFR1 產生 trimerisation 並且膜內側的 death domains 成 簇聚集,這個動作會使膜內側的轉接分子 adapter molecule 稱 TRADD (TNFR-associated factor 2)和 Death domains 產生相互作用。TRADD 可使 多數不同的蛋白恢復形成活化的 receptor,例如可使 RIP(receptor interacting protein)及 TRAF 2(TNF-associated factor 2)恢復活性,導致 NF-?B pathway 活化,誘導細胞存活;並且 TRADD 和 FADD 相關,可經由 pro-caspase-8 切 除,誘導細胞凋亡。 3. TRAIL(TNF-related apoptosis inducing ligand) TRAIL 會結合到 DR4 或 DR5 receptor,快速誘導 apoptosis,DR4 或 DR5 蛋 白位於細胞膜內側部分具有 death domains,但 DR4、DR5 無轉接蛋白,因此 DR4、DR5 receptors 直接啟動 apoptosis。 18.
(19) 自從 DR4、DR5 的 mRNA 表現在許多組織中被找到後,顯示 TRAIL 也有 保護細胞免於細胞凋亡的誘導,其中可能的機轉是基於一套 decoy receptors , decoy receptors 被稱為 DcR1 and DcR2,DcR1 and DcR2 可伴隨 DR4、DR5 和 TRAIL 結合,並且可相互競爭 TRAIL,但是 DcR1 and DcR2 和 TRAIL 結合並 不會啟始 apoptosis(108)。 ( 2.) The intrinsic apoptotic pathway 內在路徑 細胞內粒線體釋放出 cytochrome c 和 Apaf-1 結合,進而活化 caspase-9 , 最後啟動 caspase-3 活化而引起細胞凋亡。所以 caspase-3 的活性偵測,通常 是一個細胞凋亡的指標。 (二) caspase-independent 途徑而引起細胞凋亡,如透過 endonuclease G 或 apoptosis -inducing factor (AIF)等來啟動細胞凋亡。. 第四節. Topoisomerase 相關基因表現. DNA topoisomerases (topo)在許多影響DNA功能的步驟當中扮演必要的角 色,它可藉由改變雙股DNA的立體結構來影響到DNA的功能 (109)。在哺乳動物 細胞中基於? 的反應形式又分為 topo I 和 topo II,topo II 又區分為兩個不同 的isoforms為topo IIa and topo IIß (110)。 Topoisomerase II 它可以可以在雙股DNA間自由進出(111),topoisomerase II 對 DNA metabolism 重要的影響包括染色體組織化(chromosome organization) 及分離(segregation)並且為整個真核生物存活所必需 (112,113)。Topoisomerase II 為同功二聚? ( homodimeric enzyme )分子量大小從 160 to 180 kDa,其每個單 體氨基酸序列可分成三個明顯的蛋白功能區塊 (114)。. Topoisomerase II 的 n-terminal domain 接近前端 660 amino acids 區域,和 19.
(20) DNA gyrase B subunit為同源蛋白區塊,含有一致的蛋白序列作為 ATP binding 部位(115,116)。Topoisomerase II 的中央部位將近氨基酸1200序列,和A subunit of DNA gyrase為同源蛋白,含有 tyrosine residue 的活化部位,當 DNA切斷時可 和DNA形成暫時的共價鍵 (117)。C-terminal domain of topoisomerase II 不同種之 間不相同並且外觀呈現和DNA gyrase不同的同源區域( region of homology ) , 這個變異區為高度的帶電性並且含有NLS (nuclear localization sequences) (118-120) ,在體內這個區域被磷酸化作用接上磷酸 (121,122)。 直接作用於 topoisomerase II 藥劑為可以干擾至少一個步驟的催化循環 catalytic cycle,目前當為抗癌藥物使用,可用來處理乳癌、肺癌、攝護腺、肉 瘤(sarcomas)、血癌(hematological malignancies)。另外,可以穩定共價的covalent DNA topoisomerase II complex 藥物傳統上稱為 topoisomerase II poisons,然而 作用在其他 catalytic cycle中的任一步驟稱做 catalytic inhibitors;因此catalytic topoisomerase II inhibitors 定義為多種化和物可以干擾 DNA 和 topoisomerase II 之間的鍵結,或穩定非共價的 DNA topoisomerase II complexes,或是抑 制ATP binding作用。 Topoisomerase IIa and ß 對細胞的角色,可能影響生理的控制是基於? 的 特性(123)。topoisomerase IIa 和整個細胞的生長週期相關; topoisomerase II ß 和 細胞處於增殖狀況下相關,ß isoform 在細胞處於G2/M期時濃度增加2-3倍 (124)。 a isoform 相當於 type II enzyme,它在細胞複製時不會連結到姊妹染色體上, 因此,a isoform被認為為抗癌藥物重要的靶點 (125)。. 20.
(21) 第三章 實驗材料與方法 第一節 白英之採集 91 年 9 月 南投縣信義鄉東埔所採集開花原植物,經形態學鑑定為植物白英 Solanum lyratum THUNBERG,作為藥材性狀和組織鑑定標本。 91 年 11 月 信義鄉東埔所採集成熟白英果實,曬乾後浸泡於水中搓洗,去掉 浮於水面上的種子,過濾收集成熟種子,做為田間栽培材料。. 第二節 白英粗抽物備製 (一) 製備白英酒精粗抽物(SLE) 取地上部份 200 克以 50% e thanol,加熱抽取 60 分鐘,過濾,收集濾液, 重複抽取一次,合併濾液,60 ℃濃縮成流浸膏,40 ℃充分烘乾,得浸膏片 18.980 克,研粉後備用,生藥和粗抽物比值為 9.54 : 1 ,藥物 stock solution 的配製為 秤取 90 mg 粉末,加少量 DMSO 溶解後定量到 1 mL,再用 DMSO 稀釋到濃度 為 40、30、20、10、5 mg/mL,為 stock solution,以藥物終濃度 1%, DMSO 為 控制組 control。. (二) 製備白英熱水粗抽物 取田間採收乾燥後的白英地上部分 20 克加入 200 mL 二次水,加熱 100℃ 抽取 30 分鐘,過濾,收集濾液,重複抽取一次,合併濾液,100℃濃縮,以二 次水定量到總體積為 20 mL,1 mL 藥液中含有生藥 1 g,再用二次水稀釋到濃 度為 600、500、400、300、200、100、50 mg 生藥/mL 為 stock solution,白英 熱水粗抽物組以二次水為控制組。. (三)白英成分初步分離 取田間採收乾燥後的白英地上部分 200 克,95% ethanol 浸泡 7 天後,40℃ 減壓濃縮至呈現浸膏狀,n-hexane 脫脂後,利用酸水提取總生物鹼方法,分離 出總生物鹼及非鹼性脂溶性成分。非鹼性脂溶性成分 95% ethanol 溶解後,減壓 21.
(22) 濃縮,用不同極性溶媒 n-hexanol、ethyl acetate、n-butanol 分層粗抽,得到白英 分層粗抽物;總生物鹼再利用不同酸鹼度初步分離總生物鹼,得到不同酸鹼度 的生物鹼。藥物 stock solution 的配製為秤取不同粗抽物重,加少量 DMSO 溶 解後定量到 1 mL,再用 DMSO 稀釋到濃度為 5 mg/mL,為 stock solution,再使 稀釋成一系列給藥濃度,1% 給藥濃度為藥物最終濃度,1% DMSO 為 control。. 1. 總生物鹼提取及不同極性溶媒分層粗抽 (126,127) ↓1000 g 地上部份用 95% ethanol 浸漬 7天 ↓過濾 8 liters 濾液 ↓45℃減壓濃縮至200 mL ↓600 mL n-hexane 萃取 ↓不溶物以1.0% acetic acid 150 mL 水溶液溶解 ↓過濾. 酸 性 水 溶 液 (總 生 物 鹼). 不 溶 物 (非鹼性脂溶性雜質) 95% ethanol 溶解 濃縮到乾 加水溶解 加 飽合 n-hexanol 萃取. 水層. n-hexanol layer n-H= 0.1579 g. 加 ethyl acetate 萃取. ethyl acetate layer AA= 0.5408 g. 水層 加 n-butanol 萃取. n-butanol layer n-B= 1.5834 g (白英總皂? ) 22. 水層 丟棄.
(23) 2.生物鹼的初步分離 (126,127) 酸性水溶液(總生物鹼) 175 mL +CHCl3 100 mL 萃取. CHCl3 layer (弱鹼性alkaloid). acid layer (強鹼性或中強鹼性alkaloid) NH4 OH 鹼化 pH 9.3. +1.5% NaOH 100 mL. +CHCl3 萃取. CHCl3 layer (Non-phenol 弱鹼性alkaloid) NPWA=0.1225 g. basic H2 O +acetic acid 酸化 pH=4. acid aquous layer. CHCl3 layer +1.5% NaOH 50 mL. basic H2 O layer (水溶性 alkaloid). +CHCl 3 萃取. H2O layer 丟棄. CHCl3 layer (phenol 弱鹼性alkaloid). CHCl3 layer (Non-phenol 中強鹼性alkaloid) NPMA=0.0903 g. basic H2 O layer +冰醋酸 酸化 pH=5 +CHCl3 萃取. PWA=0.0534 g CHCl3 layer (phenol 強鹼性alkaloid) PSA=0.0217 g. 親脂性總生物鹼. 23. H2 O layer 丟棄.
(24) basic H2O layer (水溶性 alkaloid). 加 n-butanol 萃取. 水層? 棄. n-butanol layer 減壓濃縮. ? 留物(水溶性總生物鹼). BH=2.6359 g. 第三節 基原鑑定方法 1.. 外觀性狀之鑒別 運用五官鑒別法及立體顯微鏡檢視實驗之材料,並以照相方式記錄實 驗材料之外觀影像。. 2.. 生藥組織之鑒別 運用徒手切片法將實驗材料製成臨時組織切片並加入顯微鑒定試劑作 顯微化學反應。組織切片先加入透明劑水合氯醛試液溶解澱粉粒,再加入 藤黃酚試液和濃鹽酸作木化反應,木化細胞壁會呈現紅色,最後加入甘油水試液作封鎖劑。本實驗利用光學顯微鏡來觀察實驗材料所製成的組織橫 切及縱切切片,包括:組織的顯微構造、細胞形態以及細胞內含物的特徵, 以照相方式記錄實驗材料之組織影像、並以顯微測微器測量其大小,作為 實驗材料顯微鑒別之依據。. 24.
(25) 第四節 實驗儀器設備及試劑 (一)儀器設備、器材 1.. 細胞培養皿 (購自 FALCON). 2.. 細胞培養盤 (購自 FALCON). 3.. 細胞計數器 (Haemocytometer; 購自 Boeco). 4.. 倒立式位像差顯微鏡 (phase-contrast microscope; 購自 Olympus). 5.. 微量天平 (TE-200; MILLTER). 6.. 去離子水製造機 (購自 Minipore). 7.. 電源供應器 (購自 Amersham). 8.. 酸鹼值測定計 (C831; 購自 Consort). 9.. PVDF membrane (購自 Minipore). 10. Mini-3D Shaker (購自 Boeco) 11. SDS-PAGE 電泳槽套組 (購自 Bio-Rad) 12. Transfer Cel Blot 套組 (購自 Bio-Rad) 13. 流式細胞計數儀 (Flow cytometry; 購自 Becton Dickinson) 14. DNA 電泳槽 (購自 Mupid-2) 15. 高速離心機 (購自 HERMLE) 16. PCR Minicycler T M ( from MJ Research) 17. 分光光度計( from Beckman ) 18. EYELA ROTARY EVAPORATOR N-100 19. EYELA COOLACE CCA-1100 20. Heidolph VV2000 21. PERKIN ELMER Gene Amp PCR System 9600 22. 光學顯微鏡(Olympus CH2) 23. 光學顯微鏡(Nikon LABOPHOT-2) 25.
(26) 24. 立體顯微鏡(Nikon SMZ-2T) 25. 顯微測微器(Micrometer) 26. 電子測微器(Digimatic caliper DC-6). (二)藥品試劑 1.. RPMI medium 1640 (購自 GIBCO). 2.. fetal bovine serum (購自 GIBCO). 3.. penicillin-streptomycin (購自 GIBCO). 4.. L-glutamine. 5.. dimethyl Sulfoxide (購自 Sigma). 6.. trypan blue (購自 Sigma). 7.. disodium hydrogen phosphate (Na2HPO4; 購自 Merck). 8.. sodium chloride (NaCl; 購自 Merck). 9.. potassium dihydrogen phosphate (KH2PO4; 購自 Merck). (購自 GIBCO). 10. potassium chloride (KCl; 購自 Merck) 11. PI (Propidium iodide; 購自 Sigma) 12. 5-FU (購自 Sigma) 13. RNase A (Ribonuclease A; 購自 CLONTECH) 14. triton X-100 (購自 Sigma) 15. ethanol (購自 TEDIA) 16. APS (ammonium persulfate; 購自 Amresco). 17. acrylamide/Bis 40% solution (ACRYL/BIST M29:1; 購自 Amresco) 18. bovine serum albumin (BSA; 購自 Merck) 19. glycine (購自 Amresco) 20. methanol (購自 Merck). 26.
(27) 21. formaldehyde (購自 Merck) 22. ECL kit (Enhanced chemiluminescent kit;購自 Amersham) 23. GelCode® commassie blue (購自 PIERCE) 24. glyerol (購自 Scharlau) 25. hydrochloric acid (購自 Merck) 26. protein assay-dye reagent concentrate (購自 Bio-Rad) 27. 4X protein maker (購自 Femantas) 28. SDS (sodium dodecyl sulfate; 購自 Amresco) 29. TEMED (N,N,N’,N’-Tetramethyl-ethylenediamine; 購自 Amresco) 30. Tris (tris(hydroxymethly)-aminomethane; 購自 Amresco) 31. tween 20 (購自 Amresco) 32. 脫脂奶粉 (安佳) 33. 顯影劑 (購自 Kodak) 34. 定影劑 (購自 Kodak) 35. BioMax Flim (購自 Kodak) 36. 6X gel loading buffer (購自 EMI) 37. agarose I (購自 Amresco) 38. PRO-PREP protein extraction solution (購自 iNtRON Biotechnology) 39. 5X TBE buffer (購自 Amresco) 40. 10X SDS-PAGE running buffer (TG-SDS buffer; 購自 Amresco) 41. 4X protein loading dye (購自 Amresco) 42. Rneasy Mini kit (from QIAGEN) 43. Dneasy Mini kit ( from QIAGEN) 44. 人類大腸癌細胞株 (colo 205 : Human colon adenocarcinoma cell line; 購自 新竹食品工業研究院) 45. 一級抗體: 27.
(28) (a). anti-Cyclin A (#05-374; 購自 Upstate) (b). anti-Cyclin B1 (#05-373; 購自 Upstate) (c). anti-cdk1/cdc2 (#06-923; 購自 Upstate) (d). anti-cdk2 (#05-596; 購自 Upstate) (e). anti-actin (MAB1501; 購自 Chemicon) (f). anti-p27Kip1 (MS-256-P0; 購自 NeoMarkers) (g). anti-p21WAF1 (MS-891-P0; 購自 NeoMarkers) (h). anti-caspase-8(RB-1200-P0; 購自 NeoMarkers) (i). anti-caspase-9(RB-1205-P0; 購自 NeoMarkers) (j). anti-caspase-3(RB-1197-P0; 購自 NeoMarkers) (k). anti-NF-?B(p50) (購自 Upstate) (l). anti- NF-?B(p65) (購自 Upstate) (m). anti- Bcl-2 (購自 Upstate) (n). anti- Bax (購自 Upstate) 47. 二級抗體 (a). goat anti-mouse IgG (HRP) horseradish peroxidase conjugated antibody (AP124P; 購自 Chemicon) (b). goat anti-rabbit IgG (HRP) horseradish peroxidase conjugated antibody (購 自 Chemicon) 48. 水合氯醛試液(chloral hydrate solution) 49. 藤黃酚試液(phloroglucinol-alcohol solution) 50. 濃鹽酸(concerned hydrochloric acid) 51. 甘油-水試液(glycerin-water)(1:1) 54. 甘油-酒精-水試液(glycerin-alcohol-water)(1:1:1). 28.
(29) (三) PCR primers: Primer name. Sequence F: forward primer R: reverse primer 5’. Cdk1. Chk1. Chk2. cdc25c. cdc25b. Wee 1. MDM2. MAPK. Smad 4. Apaf-1. TNF60. FasR. TRAILRⅡ. TRAIL. DcR1. DcR2. 3’. F R F R F R F R F R F R F R F R F R F R. TTTTCAGAGCTTTGGGCACT. F R F R F R F R F R. TTAAGGGCACTGAGGACTCAG. F. CGGAAGAAATTCATTTCTTACCTC. R F. TTCTTCCAGTGTTGCCGAGGC. R. CATCTTCTTCCAGATGGT. 產物大小 (bp) 142. AAATCCAAGCCATTTTCATCC GCATTGTTTTGGAGGCTGGTT 130 AGAACTGGCCACGCAGTC TTTCCTTTCCCGATAAGCTCTG 201 CTACTGGTTCGGGAGGGATAAA GCAGCCTTGAGTTGCATAGAG 131 GACAACGCTCTTGCATAGCC TTGAACCAAGGCCCTAACAG 125 CAAGAGGAAATGTCACAGAAGC ACTGATAGAATCCAGTTTGC 321 ATGCAATGCCTACAAAGTGC ATGGCCTGCTTACATGTGC 209 TGAATTGAGGCATTTTCTCAC GAAGTTGAACAGGCTCTGGC 201 CAGTCCTCTCTGAGCCCTTGTC GAATAGCAGGACCTATCAAAGAGC 129 CAGATTAAGTGAAAACAGCATATGTG AGAGTTTCAAAACATACAAGAGAGA 161 TTCATGCAGTCGGTCA 294 TATAGGTGGAGCTGGAGGTGA CAGAACTTGGAAGGCCTGCATC 680 TCTGTTCTGTGTCTTGGAC GTCGGTGATTGTACACCTTG 579 and 666 CTTTATCAGCATCGTGTACAAG CAGGATCATGGCTATGATGG 863 TCCAGGTCAGTTAGCCAACT GGATCCCCAAGACCCTAAAGT 473 TTCCACAGTGGCATTGGCAC 369 CATGAAGACAGGGGCCCTT. Bax. 517 or 560. 29.
(30) Primer name. 5’ Bcl-2. NF-?B. I-?B. Topoisomerase Ⅰ. TopoisomeraseⅡ. TopoisomeraseⅡ a. TopoisomeraseⅡ ß. TopoisomeraseⅢ a. GAPDH. cyclin D1. cyciln B1. Cylin E. p21. p27. p53. 產物大小. Sequence F: forward primer R: reverse primer 3’. F R F R F R F R. CGACTTCGCCGAGATGTCCAGCCAG. F. GGAGATTTGTACAAACATAC. R F R F R F R F R F R F R F. AAAATGTCTGCCTTACAA. R. GGCCCTCCACAGCTTCAAGC. F R F. AGTGGACAGCGAGCAGCTGA. R. CGCTTCCTTATTCCTGCGCATTG. F. CAGCCAAGTCTGTGACTTGCACGTAC. R. CTATGTCGAAAAGTGTTTCTGTCATC. (bp) 389. ACTTGTGGCTCAGATAGGCACCCAG GATGAATATGTGTGTATCCG 658 TTTGTTGTGCTTGAGAACC CATGTTTCACACACACAATCAGC 240 GGAGTAGGAGACTTTCACAAGTGG TCTTCCCATGAAACTATAAATGGC 224 CAAAGCTGAAAACCGCAGG 186 ACACTCAGCCTCTTATGTGC 272 ATCAAATGTTGTCCCCG GCCCAAAGAGCACAAACATT 204 CAGACAGTAGTGACATTTCAATG CCAGCTGAAGCCCACTGT 160 CCTTGCATTACACCGTCCTT CCATGGAGAAGGCTGGGG CAAGAGTTGTCATGGATGACC GAGACCATCCCCCCTGACGGC TCTTCCTCCTCCTCGGCGGGC AAGGCGAAGATCAACATGGGC AGTCACCAATTTCTGGAGGG GTTGCACCAGTTTGCGTATGTG. TAGAAATCTGTCATGCTGGTCTG AAACGTGCGAGTGTCTAACGGGA. 30.
(31) 第五節 實驗方法 一.人類大腸腺癌細胞 Colo 205 細胞培養 Colo 205 細胞培養於 RPMI-1640 培養基中(含 10% fetal bovine serum、100 units/mL penicillin、100 µg/mL streptomycin、2 mM L-glutamine),置於 37 ℃、 5% CO2 的培養箱中培養,並控制細胞的數目,使其細胞密度維持在 2-5×10 5 cells/mL。. 二. 血球計數器評估存活細胞數 計數 Colo 205 細胞,接種於 12 well 培養盤中,每 well 含培養液總體積 6 小時、12 小時、24 小時、48 小時分別為 2mL,72 小時為 3mL,每 well 細胞總 數為 1.5×10 5 cells,培養 24 小時使貼壁後,分別給不同濃度藥物,加等藥物體 積的 DMSO 為 control 組,分別持續和藥物培養 6 小時、12 小時、24 小時、48 小時、72 小時後,收集全部細胞,trypsin 使細胞游離瓶底,1X PBS 清洗 2 次 後,加入 500 µL 的等量 1X PBS,每個給藥濃度取 50 mL 細胞懸浮液與 50 mL trypan blue ( 0.4% w/v trypan blue)等體積混合均勻於 1.5 mL 小離心管中(每個濃 度取 3 次)。取少許混合液﹙約 10 mL﹚自血球計數盤 chamber 上方凹槽加入, 蓋上蓋玻片,於 100 倍倒立顯微鏡下觀察。計數四個大方格之細胞總數,再除 4,乘以稀釋倍數,最後乘以 104,即為每 mL 中細胞懸浮液之細胞數。. 三. 利用流式細胞計數儀計數存活的細胞數 計數 Colo 205 細胞,接種於 12 well 培養盤中,每 well 含培養液總體積 6 小時、12 小時、24 小時、48 小時為 2 mL,72 小時為 3 mL,每 well 細胞總數 為 1.5×10 5 cells,於 37 ℃、5% CO2 的培養箱(incubator)進行培養 24 小時, 使貼壁後,分別給不同濃度藥物,加等藥物體積的 DMSO 為 control 組,分別 持續和藥物培養 6 小時、12 小時、24 小時、48 小時、72 小時後,收集全部細 31.
(32) 胞,加 2 drops trypsin 使細胞游離瓶底,離心 1500 轉 5 分鐘,1X PBS 清洗 2 次後,再離心 1500 轉 5 分鐘,倒掉上清液,加 400 µL propidium iodine( 0.004 mg/mL PBS )上流式細胞計數儀,計算存活細胞數,流式細胞計數儀條件為固定 低流速(35 µL/min),抓取 20 秒(escape time),計錄 R2 events 當作相對的存活 細胞數,不同濃度之間比較差異。計錄 R2 gated percentage 當作細胞存活率指 標,不同濃度之間比較差異。並且以 0 時間(給藥同時收細胞上流式細胞計數儀 計數存活率)的存活細胞數目為初始給藥點細胞基準線,如果給藥後細胞存活數 目低於初始給藥點細胞基準線,表示藥物對於細胞具有毒殺作用,若給藥後細 胞存活數目介於控制組的細胞數,表示藥物具有抑制細胞增殖作用。. 四. 利用流式細胞計數儀評估細胞週期及 sub-G1 的影響 計數 Colo 205 細胞,接種於 12 well 培養盤中,每 well 含培養液總體積 6 小時、12 小時、24 小時、48 小時為 2 mL,72 小時為 3 mL,每 well 細胞總數 為 1.5×10 5 cells,於 37 ℃、5% CO2 的培養箱(incubator)進行培養 24 小時, 使貼壁後,分別給不同濃度藥物,加等藥物體積的 DMSO 為 control 組,分別 持續和藥物培養 6 小時、12 小時、24 小時、48 小時、72 小時後,收集全部細 胞,加 2 drops trypsin 使細胞游離瓶底,離心 1500 轉 5 分鐘,1X PBS 清洗 2 次後,再離心 1500 轉 5 分鐘,倒掉上清液,將細胞完全打散後,置於震盪器上 以 shake 3 速度震盪,4 ℃的 70% 乙醇一滴一滴緩慢滴入,進行細胞固定,固 定後置於-20 ℃冰箱隔夜存放。隔天,將樣品取出後離心 1500 轉 5 分鐘,以除 去乙醇,將細胞完全打散,加入 2 mL 的 1X PBS ( 1X phosphate buffer saline (PBS):NaCl 8.0 g、KCl 0.2 g、Na2 HPO4 1.44 g、KH2PO4 0.2 g 加二次水到總體 積 1000 mL。pH=7.4 ) 緩衝液清洗 1 次後將細胞完全打散。最後在每個試管中 加入 300 µL 的 PI stain 染劑( 0.002 % propidium iodide 5 mL,5 % triton 5 mL,2 mg/mL RNase A 1.25 mL,1X PBS 13.75 mL )其量可視細胞數目增減,避光反應 30 分鐘。最後以流式細胞計數儀進行樣品分析,一秒細胞數不超過 300 顆細胞, 32.
(33) 每個數據收集 12000 顆細胞,數據以 Modfit LT軟體進行處理分析。記錄 G1, S,G2/M phase,sub-G1 比率。. 五. 利用倒立式位像差顯微鏡評估細胞形態 計數 Colo 205 細胞,接種於 12 well 培養盤中,每 well 含培養液總體積 2 mL,每 well 細胞總數為 1.5×10 5 cells,於 37 ℃、5 % CO2 的培養箱( incubator) 進行培養 24 小時,使貼壁後,分別給不同濃度藥物,加等藥物體積的 DMSO 為 control 組,分別持續和藥物培養 6 小時、12 小時、24 小時後於 200 倍倒立 式位像差顯微鏡下照相。. 六. DNA 的萃取(DNA extraction) 計數 Colo 205 細胞,接種於 6 well 培養盤中,每 well 含培養液總體積 6 小時、12 小時、24 小時、48 小時為 3 mL,每 well 細胞總數為 2×106 cells,於 37℃、5% CO2 的培養箱(incubator)進行培養 24 小時,使貼壁後,分別給不 同濃度藥物,加等藥物體積的 DMSO 為 control 組,分別持續和藥物培養 6 小 時、12 小時、24 小時、48 小時後,無菌吸管吸出培養盤中細胞懸浮液至 15 mL 離心管收集全部細胞,無菌的 1X PBS 清洗 1 次後,加 3 滴 trypsin 使細胞游 離瓶底,離心 1500 轉 5 分鐘,1X PBS 清洗 2 次後,再離心 1500 轉 5 分鐘, 倒掉上清液,將細胞完全打散後,加 1X PBS 使細胞懸浮液為 200 µL,加 20 µL QIGEN Protease (or proteinase K) 到離心管底部,加 200 µL buffer AL 到 sample 內,混合均勻震盪 15 秒,置於 56 ℃ 水浴鍋 10 分鐘,離心 1500 轉 5 分鐘, 加 200 µL absolute ethanol (無水) 震盪 15 秒,然後離心 1500 轉 10 秒,取混 合液 600 µL 進入 the QIAamp spin column 內,於 10000 轉離心 1 分鐘,更 換乾淨的 2 mL collection tube,加 500 µl buffer AW1 solution,於 10000 轉離心 1 分鐘,更換乾淨的 2 mL collection tube,加 500 µl Buffer AW 2,於 13000 轉離 心 3 分鐘(離心到乾),更換 QIAamp spin column 成新的 2 mL collection tube,. 33.
(34) 於 13000 轉離心 1 分鐘,更換 QIAamp spin column 成新的 2 mL 無菌的 collection tube,加 50 µL buffer AE 置於室溫 1 分鐘,10000 轉離心 1 分鐘,收 集濾液即為 DNA。. 七. Total RNA 分離抽取 ( Isolation of total RNA ) 計數 Colo 205 細胞,接種於 6 well 培養盤中,每 well 含培養液總體積 4 小時、8 小時、12 小時、24 小時為 3 mL,每 well 細胞總數為 2×106 cells,於 37 ℃、5% CO2 的培養箱(incubator)進行培養 24 小時,使貼壁後,分別給不 同濃度藥物,加等體積藥物的 DMSO 為 control 組,分別持續和藥物培養 4 小 時、8 小時、12 小時、24 小時後,無菌吸管吸出培養盤中細胞懸浮液至 15 mL 離心管收集全部細胞,滅菌 PBS 清洗 1 次後,加 3 drops trypsin 使細胞游離瓶 底,離心 1500 轉 5 分鐘,滅菌 PBS 清洗 2 次後,再離心 1500 轉 5 分鐘,倒掉 上清液,不打散細胞,用 pipet 吸掉多餘的 PBS,吸乾後打散細胞,加 600 µL buffer RLT 上下混合均勻呈澄清黏稠均質狀態 (每 1 mL buffer RLT 預先加 10 µL 2-mecaptoethanol ),加 600 µL 70% ethanol 固定,使用 pipet 混合均勻,total 700 µL 吸到 RNeazy mini QIAshredder column,然後離心 13000 轉 15 秒,倒掉收集 管濾液,剩餘 600 µL 再吸到 RNeazy mini QIAshredder column,離心 13000 轉 15 秒,倒掉收集管濾液,加 700 µL Buffer RW1,離心 13000 轉 15 秒,倒掉 下層濾液,加 500 µL buffer RPE,離心 13000 轉 15 秒,倒掉下層濾液,加 500 µL buffer RPE,離心 13000 轉 2 分鐘,上層濾膜管柱移到另一新 1.5 mL 離心管, 加 40 µL RNase-free water,靜置 1 分鐘,離心 13000 轉 1 分鐘,得到 total RNA 40 µL。 取已滅菌 1.5 mL eppendorff tube,先取 198 µL DEPC-H2O,再取 2 µL sample,上下輕柔混合均勻,置冰上,於 285 nm 測 OD value。OD value=1 相當 RNA=40 ng/µL,sample RNA 濃度=O.D. value × 100( 稀釋 100 倍 ) × 40 ng/µL,每個 sample 取 3000 ng RNA 量做 Reverse Transcription。. 34.
(35) 八. First-strand cDNA synthesis (Reverse Transcription) 取 0.5 mL sterilized-eppendorf tube 置於冰上,加入適量滅菌過的二次水, 二次水量由 10-(3000/RNA 濃度) µL,sample RNA 量 (µL)= 3000/RNA 濃度,加 dNTP mix 1 µL、oligo dT 1 µL,輕揉的上下混合完全,置於 PCR 機器 well 上, 加熱 65℃ 5mins,取出離心 2000 rpm×4 秒,加入 5× first-strand buffer 4 µL、0.1 M DDT 2 µL、RNAase out recombinant ribonuclease inhibitor(可用二次水代替) 1 µL,輕柔混合,total 19 µL 加熱 42℃ 2mins,加入 superscript Ⅱ transferase 1 µL,加熱 42℃ 50 mins 接著加熱 70℃ 15 mins,得 cDNA first strand 4℃冰箱保 存。. 九. Polymerase Chain Reaction (PCR) 取滅菌過的 0.5 mL eppendorf tube 置於冰上,加已滅菌的二次水 38.1 µL、10× PCR buffer 5 µL、50 mM MgCl 2 1.5 µL、10 mM dNTP mixture 1 µL、10 mM amplification primer 3’end 1 µL、10 mM amplification primer 5’end 1µL,加 2 µL cDNA sample, 輕柔均勻混合,離心 2000 轉 1 分鐘,加 Tag DNA polymerase 0.4 µL(加 Tag polymerase 時直接加到底部,不必混合),總量 50 µL 離心 2000 轉 15 秒,跑 PCR。PCR 條 件為為預熱 95℃ 5 分鐘, denaturation 溫度 95℃ 1 分鐘, annealing 溫度 55℃ 1 分 鐘, extension 溫度 72℃ 1 分鐘 35-40 cycles,最後以 72℃ 10 分鐘退火。 Agarose Gel Electrophoresis 取 25 µL PCR 產物,加入 5 µL 的6X loading buffer 混合均勻,總量30 µL 注入 2 % agarose gel (溶於0.5X TBE buffer PH=7.4) 電壓設定50 voltage跑60分 鐘,取出 agarose 以 5 µg/mL ethidium bromide 染色 30 分鐘。. 十. 蛋白質之定性及定量分析 (一) 細胞總蛋白質抽取 35.
(36) 計數 Colo 205 細胞,接種於 6 well 培養盤中,每 well 含培養液總體積為 3 mL,每 well 細胞總數為 3×10 6 cells,於 37 ℃、5 % CO2 的培養箱( incubator) 進行培養 24 小時,使貼壁後,分別給終濃度 300 µg/mL 藥物,加等藥物體積的 DMSO 為 control 組,每個重複 3 well,分別持續和藥物培養 4 小時、8 小時、 12 小時、24 小時後,吸管吸出培養盤中細胞懸浮液至 15 mL 離心管收集全部細 胞,PBS 清洗 1 次後,加 3 drops trypsin 使細胞游離瓶底,離心 1500 轉 5 分 鐘,1X PBS 清洗 2 次後,再離心 1500 轉 5 分鐘,吸乾上清液,將細胞完全打 散後,加入 200 µL 的 RIPA lysis buffer〔 50 mM Tris-Hcl, pH 7.4,150 mM NaCl, 1 mM ethylenediamine-tetra-ethyl acid (EDTA) , 1 mM ethylene glycol - bis (aminoethylether) - tetra-ethyl acid (EGTA),0.5 mM dithothreitol,1% NP-40,0.3% deoxycholate,10 µg/mL leupeptin,10 µg/mL aprotinin,10 µg/mL soybean trypsin inhibitor,0.5 mM phenylmethylsulfonylfluoride (PMSF)〕。劇烈震盪後,置於冰 上 30 分鐘,轉移至 1.5 mL 的 eppendorf tube,再經離心 13,000 轉 15 分鐘, 取上清液即為總蛋白質。. (二) 蛋白質的定量 蛋白質標準品檢量線製作: 以 Bradford 方法 (128),以胎牛血清白蛋白(bovine serum albumin; BSA) 為蛋白 質標準品,精確稱量 50 mg,用二次水 1 mL 溶解後,稀釋成一系列濃度( 7 µg/µL、6.5 µg/µL、5.5 µg/µL、5 µg/µL、3.5 µg/µL ),取 5 µL 的蛋白質樣品與 250 µL 的 Bradford 染劑(稀釋 5 倍),混勻 5 分鐘後,放入分光光度計 590 nm 測 定其吸光值平均值,依並 OD value (Y) 對蛋白濃度 µg/mL (X),求出趨勢線方 程式 y=0.1246X+1.2263 R2=0.9933。 樣品蛋白質定量: 取 5 µL 的蛋白質樣品與 250 µL 的 Bradford 染劑(稀釋 5 倍),混勻 5 分鐘 後,放入分光光度計 590 nm 測定其吸光值平均值,蛋白樣品 OD value 用 RIPA. 36.
(37) lysis buffer 稀釋到檢量線範圍內,利用檢量線公式則可求得實際蛋白質樣品濃 度。. (三) 變性電泳 ( SDS-PAGE ) 配置 1.5 mm 厚度的 10 % discontinuous acrylamide gel,gel 分上下兩層,依 照下面表格依序配製下層膠和上層膠: 下層膠. 上層膠. (10% Separation gel). (5% Stacking gel). 4 片量. 4 片量. 二次水. 14.4 mL. 4.7 mL. 1.5 M Tris ( pH8.8 ). 7.5 mL. -. 1.0M Tris ( pH6.8 ). -. 2.5 mL. 10% SDS. 300 µL. 60 µL. 40% acrylamide/Bis (29:1). 7.5 mL. 1.22 mL. 10% ammonium persulfate (APS). 300 µL. 60 µL. TEMED. 10 µL. 6 µL. 組. 成. 下層膠注入鑄膠玻璃後,加滿二次水去除氣泡並壓平下層膠之上緣,靜置 30 分 鐘待膠體凝固。隨後,將上層膠注入後小心地插入樣品槽梳子(comb)到未凝固 的膠體中,避免氣泡出現於樣品槽(wells)下緣。等待約 30 分鐘上層膠凝固後拔 出樣品槽梳子,並以二次水小心沖洗樣品槽內避免雜質殘留。將鑄好的膠體放 入電泳槽中,加入 running buffer ( Tris 30 g、glycine 14.4 g、SDS 10 g 加二次水 至 1 liter)。鑄好的膠體先以 100 v、300 mAmp 為條件預跑 15 分鐘。sample 加 入 4X protein loading dye(8 % SDS,0.01 % bromophenol blue,40 % glycerol, 250 mM Tris pH 6.8,5 % 2-mercaptoethanol)於蛋白質樣品中,置於 95 oC 乾浴 中變性(denature)10 分鐘,冰浴冷卻後快速離心,每個 sample 取蛋白總量為 30 µg,將各樣品及 5 µL 標準分子量的 multimarker 分別注入膠體的孔槽( well) 中,以 40 voltage 跑 30 分鐘,再以 80 voltage 跑 100 分鐘。. 37.
(38) (四) 西方墨點法 (Western Blotting) 轉印步驟:將轉印夾打開後黑色面朝下放,取出一片海綿墊片浸泡轉印緩衝液 transfer buffer ( Tris 3.3 g、glycine 14.4 g 先加二次水到 800 mL,待 Tris 及 glycine 完全溶解後再加入 200 mL 的 100% 甲醇即可)。後放於其上面,並依序在海綿 片上放上 3M 濾紙、SDS-PAGE gel、PVDF 轉印膜 (先用 100 %甲醇濕潤 15 秒、 再放於 transfer buffer 中浸泡 1 分鐘以上)、3M 濾紙,最後再放上一片海棉墊片 後夾上轉印夾(夾層中間不可有氣泡),形成類似三明治夾層的構造。 海綿墊片 3M paper PVDF SDS-PAGE Gel 3M paper 海綿墊片. 紅、正極(+). 黑、負極(-). 將轉印夾放入電泳槽中後放入冰盒並加滿轉印緩衝液。於轉印槽外圍放入足夠 的冰塊,使整個系統保持低溫狀態。以 100 voltage、400 m Amp、1.5 小時為條 件,進行蛋白質轉印步驟。轉印完成後取出轉印膜裁去多餘部分,轉印膜後以 0.05 % tween/1X PBS 清洗 10 分鐘共 3 次。緊接將轉印膜以 5 % 脫脂奶粉(溶於 0.05 % tween/1X PBS 中)進行 blocking 步驟,以室溫 1 小時為條件進行。取出轉 印膜後於小盒中以 0.05 % tween/1X PBS 清洗 10 分鐘共 3 次。倒掉清洗液,加 入 8 mL 的一級抗體(溶於含 5 %脫脂奶粉的 0.05 % tween/1X PBS 中),4 ℃隔夜 進行搖盪。隔天取出,回收一級抗體,以 0.05 % tween/1X PBS 清洗轉印膜 10 分鐘共 3 次。加入 8 mL 的 10000 倍 goat anti IgG (HRP) horseradish peroxidase conjugated antibody 二級抗體 (溶於含 5 %脫脂奶粉的 0.05 % tween/1X PBS 中),於室溫下搖盪進行 1 小時,最後取出轉印膜後以 0.05 % tween/1X PBS 清. 38.
(39) 洗清洗 10 分鐘共 3 次。 (五)壓片步驟:(暗室中進行) 將轉印膜浸泡於 ECL 試劑之混合液(每瓶各取 1.5 mL 等比例混合)中 1 分鐘 反應。以兩張投影片黏貼固定於 cassette 內,轉印膜並正面朝上放置於壓片卡匣 (cassette)兩張投影片中間,以 Hyperfilm 軟片置於上層投影片上,對準轉印膜進 行壓片,感光時間依轉印膜上螢光亮度決定時間長短,約 5 秒至 1 小時不等。 感光完成後放入顯影劑進行顯影步驟(時間依實際觀察決定),再以清水沖洗 30 秒後放入定影劑中,過 30 秒後再以清水沖洗 30 秒。. 十一. 統計分析 實驗結果以平均值±標準差(mean ± SD)表示,使用 Student’s t-test 來決定 實驗組與對照組之差異。*表示 p<0.05,**表示 p<0.01,表示統計上具顯 著差異。. 39.
(40) 第四章. 結. 果. 一、白英種子播種於腐植土後 2 週可發苗,小苗於第 4 週可移栽於田間。溫室內 生長情形比田間生長快速,溫室內植株葉片較寬大,數目多於田間栽培植 株,植株生長較為健壯。溫室內栽培施肥與未施肥(台農一號有機氮肥)比 較,植株生長情形無顯著差異,移栽 3 個月後施肥植株近莖基部的葉,葉片 較容易脫落。白英種子播種後 3.5 個月後開花、5 個月後結果,達採收標準。 採收後乾燥秤重新鮮品含水量約為 77.5%,結果如圖 四。. 二、田間採收之白英其組織學鑑定結果如圖 三 所示。 外觀性狀: 莖粗大者直徑約 0.5~1.2 cm 表面呈灰褐色,曲折彎曲,表皮具光澤性 之縱皺紋,偶有皮層剝落而露出木部及韌皮纖維。莖節上具瘤狀突起或莖 痕。細莖呈黃綠色與葉及葉柄均同樣密生軟毛,莖同樣不易折斷。莖破折面 呈纖維性,導管與位於中心部之髓部裂隙;莖細者斷面中空。氣香味稍苦甘。 組織構造: 莖橫切面顯微鏡下鏡檢發現,表皮密生腺毛,最外層為角質層(cu), 下方為一層表皮細胞(ep)所構成,再下方由 3~6 層呈等徑性之橢圓~長方形 之柔細胞(p)而成,呈切線方向排列。順次移向內方為包圍其外篩部(le)之內 鞘纖維束(fb)一層,此內鞘纖維束由 1~5 列之韌皮纖維所構成。外篩部發達 而明顯,形成層(c)不明顯。木部由導管(有緣孔紋導管,螺旋紋導管,偶有 環紋導管),假導管(孔紋),木纖維,髓線所構成,導管圓大,其徑可達 180 µ 內外,多為有緣孔紋導管(vp),在髓冠部出現之導管多為螺旋紋導管(vs)。 髓線(mr)乃由 1~2 列具孔斑之髓線細胞而成,並呈放射狀排列。木部內側 與髓部外側交接處有顯著而發達之內篩(le)存在,由木部延長之螺旋紋導管 (vs)於內篩部出現。內篩與髓之交接處附近有 1-3 列之纖維散在。內篩內側 即為具有孔斑之髓細胞所構成之髓部(m),髓部的中心為廣大之空腔(cav)。 40.
(41) 三、利用流式細胞計數儀評估不同濃度( control、50 µg/mL、100 µg/mL、200 µg/mL、300 µg/mL、400 µg/mL )之白英 50%乙醇粗抽物( SLE )對人類大腸癌 細胞株 Colo 205 處理 6 小時、12 小時、24 小時、48 小時、72 小時後觀察細 胞存活率、細胞週期及 sub-G1 的變化。結果如表 二、圖 五、圖 六 顯示, SLE 有明顯的抑制細胞增生作用且有劑量及時間依賴效應。細胞週期的變化, 結果如圖 七、圖 八 顯示,SLE 處理 12 小時、24 小時後細胞週期停止於 S-phase,處理 48 小時後停止於 G2/M phase。在 sub-G1 的變化方面,結果如表 三、圖 九、圖 十、圖 十一 顯示,SLE 在濃度為 300 µg/mL、400 µg/mL 給 藥 48 小時後開始有 sub-G1 peak 出現,持續給藥 72 小時後誘導 sub-G1 比率呈 現劑量依賴效應。. 四、利用倒立式位像差顯微鏡下觀察不同濃度( control、100 µg/mL、200 µg/mL、 250 µg/mL、300 µg/mL、400 µg/mL )之白英 50%乙醇粗抽物( SLE )對人類大 腸癌細胞株 Colo 205 處理 6 小時、12 小時、24 小時後形態學方面的改變。結 果如圖 十二 顯示,SLE 處理 6 小時後高濃度( 300 µg/mL、400 µg/mL )時細 胞已有明顯變化,細胞膜呈現模糊不清,細胞質空泡化,部分細胞變淡死亡; 處理 24 小時後於低濃度( 200 µg/mL、250 µg/mL )細胞膜呈現模糊不清,細胞 質空泡化,部分細胞變淡死亡。. 五、利用流式細胞計數儀評估不同濃度( control、50 µg/mL、100 µg/mL、200 µg/mL、300 µg/mL、400 µg/mL )之白英 50 %乙醇粗抽物( SLE )對人類大腸癌 細胞株 Colo 205 處理 6 小時、12 小時、24 小時、48 小時、72 小時後觀察存 活細胞數的變化。結果如圖 十三 顯示 100 µg/mL、200 µg/mL 的 SLE 抑制細 胞增生作用,SLE 濃度為 300 µg/mL、400 µg/mL SLE 對人類大腸癌細胞株 Colo 205 具有細胞毒殺作用。. 41.
(42) 六、利用 trypan blue 染色細胞計數器評估不同濃度( control、50 µg/mL、100 µg/mL、200 µg/mL、300 µg/mL、400 µg/mL )之白英 50%乙醇粗抽物( SLE ) 對人類大腸癌細胞株 Colo 205 處理 12 小時、24 小時、48 小時、72 小時、96 小時後觀察存活細胞數的變化。結果如圖 十四 顯示 100 µg/mL、200 µg/mL 的 SLE 抑制細胞增生作用,SLE 濃度為 300 µg/mL、400 µg/mL 時對人類大 腸癌細胞株 Colo 205 具有細胞毒殺作用。正對照組 100 µg/mL 的 5-FU 於 24 小時、48 小時細胞存活數介於 200-300 µg/mL SLE 之間、結果和使用流式細 胞計數儀計數存活細胞結果相符合。. 七、利用流式細胞計數儀評估不同溶媒分層粗抽及總生物鹼不同酸鹼度分層粗抽 物對人類大腸癌細胞株 Colo 205 處理 48 小時後細胞存活率、細胞週期及 sub-G1 的變化。結果如圖 十五 顯示,不同溶媒分層粗抽物及總生物鹼不同酸 鹼度粗抽物於濃度 50 µg/mL 處理 48 小時後存活率沒有明顯減少。細胞週期的 變化,結果如圖 十六 顯示,白英 n-butanol 層粗抽物及 n-hexanol 層粗抽物 給藥 48 小時後細胞週期停止於 S-phase,正對照組 300 µg/mL SLE 處理 48 小 時後停止於 G2/M phase。在 sub-G1 的變化方面,結果如圖 十六、圖 十七 顯 示,n-butanol 層粗抽物在濃度為 50 µg/mL 給藥 48 小時後開始有 sub-G1 peak 出現,和正對照組 300 µg/mL SLE 誘導凋亡比率相當。. 八、利用流式細胞計數儀評估不同濃度( control、20 µg/mL、40 µg/mL、60 µg/mL、 80 µg/mL、100 µg/mL )之白英 n-butanol 粗抽物及 250 µg/mL 乙醇粗抽物( SLE 250 )、80 µg/mL 5-FU 對人類大腸癌細胞株 Colo 205 處理 24 小時、48 小時、 72 小時後觀察細胞存活率的變化。結果如表 四、圖 十八 顯示,白英 n-butanol 粗抽物有明顯的抑制細胞增生作用且有劑量及時間依賴效應。細胞週期的改 變,結果如圖 十九 A 顯示,白英 n-butanol 粗抽物、SLE 250 給藥 24 小時後 對於細胞週期影響不大,但是 80 µg/mL 5-FU G1 phase 細胞增加。如圖 十九 B. 42.
(43) 顯示,48 小時後白英 n-butanol 粗抽物、250 µg/mL 乙醇粗抽物、80 µg/mL 5-FU 細胞週期均停滯於 G1 phase。 在 sub-G1 peak 方面,結果如圖 二十、圖 二十 一 顯示,高濃度時白英 n-butanol 粗抽物於 24 小時出現 sub-G1 peak,比 80 µg/mL 5-FU 更早出現,給藥 72 小時後 80 µg/mL 之白英 n-butanol 粗抽物大部 份細胞均產生凋亡,誘導凋亡的比率大於 80 µg/mL 的 5-FU。. 九、利用流式細胞計數儀評估不同濃度( control、10 µg/mL、20 µg/mL、40 µg/mL、 80 µg/mL )之 5-FU 對人類大腸癌細胞株 Colo 205 處理 6 小時、12 小時、24 小 時、48 小時、72 小時後觀察細胞存活率、細胞週期及 sub-G1 的變化,且利用 白英 300 µg/mL 乙醇粗抽物( SLE 300 )為正對照組做比較。結果如圖 二十二 A 顯示,5-FU 各濃度對 Colo 205 cell 具有抑制增殖作用,但各濃度間無明顯 差異,正對照組 SLE 300 µg/mL 抑制細胞增生作用和 80 µg/mL 的 5-FU 相當。 細胞週期的改變,結果如圖 二十三 顯示,不同濃度之 5-FU 對細胞週期影響 於給藥 12 小時後 G1 phase 上升,24 小時後 S phase 上升,72 小時後 G1 phase、 G2/M 上升、S phase 下降。SLE 300 µg/mL 於 12、48 小時細胞週期停滯於 G2/M 期。 sub-G1 peak 方面,結果如表 六、圖 二十四、圖 二十五 顯示, 高濃度 80 µg/mL 時 5-FU 於 48 小時出現 sub-G1 peak,SLE 300 µg/mL 於 48 小時出現 sub-G1 peak,給藥 72 小時 SLE 300 µg/mL 誘導凋亡的比率大於 80 µg/mL 的 5-FU。. 十、利用流式細胞計數儀評估不同濃度( control、0.25 mg/mL、0.5 mg/mL、1 mg/mL、2 mg/mL、3 mg/mL、4 mg/mL、5 mg/mL )之白英熱水粗抽物對人類 大腸癌細胞株 Colo 205 處理 24 小時、48 小時後觀察細胞存活率的變化。結果 如圖 二十六 顯示,白英熱水粗抽物,藥物處理 24 小時及 48 小時後,對於 Colo 205 細胞有抑制增殖作用,且有劑量依賴性。. 43.
(44) 十一、利用 DNA fragementation electrophoresis assay 評估不同濃度( control、50 µg/mL、100 µg/mL、200 µg/mL、300 µg/mL、400 µg/mL )之白英乙醇粗抽物 SLE 對人類大腸癌細胞株 Colo 205 處理 48 小時後觀察 DNA ladder 的變化。 結果如圖 二十七、二十八 顯示,300 µg/mL、400 µg/mL 的 SLE 處理 48 小 時後,可見到明顯的 DNA ladder partern 並且 genomic DNA 大部份被切除。 300 µg/mL SLE 處理 12 小時、24 小時後,具有明顯的 DNA ladder partern。比 較不同濃度的 300 µg/mL SLE、80 µg/mL n-butanol 粗抽物、80 µg/mL 5-FU 於 藥物處理 24 小時、48 小時後,結果如圖 二十九 顯示,300 µg/mL SLE、80 µg/mL n-butanol 粗抽物處理具有明顯的 DNA ladder partern,80 µg/mL 5-FU 處理的 DNA ladder partern 不明顯。. 十二、利用 RT-PCR 方法觀察不同濃度( control、100 µg/mL、200 µg/mL、300 µg/mL、400 µg/mL )之白英 50%乙醇粗抽物( SLE )對人類大腸癌細胞株 Colo 205 處理 24 小時後檢測基因表現的影響。 細胞週期相關基因表現的影響,結果如圖 三十 顯示,對於基因具有上調表現 ( up-regulation )為 Cdk1、p27;對於基因具有下調表現( down-regulation )為 Chk1、Chk2、MDM2;對 Colo 205 細胞基因有表現但對藥物作用沒有影響 的基因為 P21、p53、Cdc25b、Cdc25c、Wee1、cyclinB1。 細胞凋亡相關基因表現的影響,結果如圖 三十一 顯示,對於基因具有下調表 現為 Bcl-2、Apaf-1;對 Colo 205 細胞基因有表現但對藥物作用沒有影響的 基因為 BAX、Caspase-3、Caspase-6、Caspase-8、Caspase-9、Smad 4 及 MAPK。 對 Death Recepter 相關基因表現的影響,結果如圖 三十二 顯示,基因具有下 調表現為 TRAIL-2、DcR-2,Colo205 細胞基因有表現但對藥物作用沒有影 響的基因為 TNF60、TRAIL。 對 NF-?B 相關基因表現的影響,結果如圖 三十三 顯示,白英 50%乙醇粗抽物 於低濃度時上調表現,高濃度時具有下調表現,I-?B 有表現但對藥物作用沒 44.
(45) 有影響。 Topoisomerase 相關基因表現的影響,結果如圖 三十四 顯示,基因具有下調表 現為 topoisomerase Ⅱa,對 topoisomerase Ⅰ基因上調,基因有表現但對藥物 作用沒有影響的基因為 topoisomerase Ⅱβ。. 十三、利用 Western Blot 方法觀察濃度為 300 µg/mL 之白英乙醇粗抽物( SLE )對 人類大腸癌細胞株 Colo 205 處理 4、8、12、24 小時後檢測相關蛋白表現的影 響。結果如圖 三十五、圖 三十六、圖 三十七顯示,增加蛋白表現的為,p27、 caspase-3、casepase-9,減少蛋白表現的為 Cdk1、cyclin B1、NF-?B(p50)、 NF-?B(p65),無影響的為 p53、cyclin E、caspase-8。. 45.
數據
Outline
相關文件
SF11310 張基晟 比較 Ipilimumab 加上 Etoposide/Platinum 與 Etoposide/Platinum
- 絕對數值為零
眼睛是心靈之窗,心靈是眼神之源。在眼球後方 感光靈敏的角膜含有 1.37 億個細胞,將收到的信 息傳送至腦部。這些感光細胞,在任何時間均可 同時處理
• 是細胞不正常增生,且這些增生的細胞可
基因編輯技術以人工核酸酶辨識特定 DNA 位置,並於此處切斷雙股 DNA。DNA 斷
細胞外液(extracellular fluid;ECF) 約佔
•自內分泌系統分泌的激素利用血液輸送到 目 標器官。類固醇和甲狀腺激素是非極 性,且為脂 溶性,所以能通過細胞膜進入 目標細胞。大部分
辣椒 有利於促進胃黏膜的再生,維持胃腸 細胞功能,防治胃潰瘍。常食可降低 血脂、血糖減少血栓形成,且有效地
