行政院國家科學委員會專題研究計畫 成果報告
德國蜚蠊生物時鐘掩蓋機制探討(3/3)
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-002-058- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學昆蟲學系暨研究所 計畫主持人: 李後晶 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 93 年 11 月 2 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※
※
※ 德國蜚蠊生物時鐘掩蓋機制探討 ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:■個別型計畫 □整合型計畫
計畫編號:NSC90-2313-B-002-331-
NSC91-2313-B-002-274-
NSC92-2313-B-002-058-
執行期間:90 年 8 月 1 日至 93 年 7 月 31 日
計畫主持人:李後晶
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
■出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立台灣大學昆蟲學系
中 華 民 國 九 十 三 年 十 月 二十 日
1
行政院國家科學委員會專題研究計畫成果報告
德國蜚蠊生物時鐘掩蓋機制探討
The masking mechanisms of the biological clock in the
German Cockroach
Blattella germanica
計畫編號:NSC90-2313-B-002-331-
NSC91-2313-B-002-274-
NSC92-2313-B-002-058-
執行期限:90 年 8 月 1 日至 93 年 7 月 31 日
主持人:李後晶 國立臺灣大學昆蟲學系
一、中文摘要 由於德國蜚蠊活動行為的表現上,雌 蟲會因為卵巢的發育,而造成活動行為日 週律動被掩蓋的現象,本計劃嘗試深入探 討此掩蓋機制。首先我們進行觸角電位圖 譜 (Electroantennogram, EAG) 的偵測,當 作第二個生物時鐘外在表徵:利用雌蟲之 觸角電位圖譜的偵測,我們可以了解雌蟲 活動行為之掩蓋現象其作用之機制位於生 物時鐘系統的哪一部位。另外因為德國蜚 蠊活動行為的掩蓋現象是生殖行為的生理 活動運作的結果,我們同時選取交尾活性 (mating activity) 作為另一個生物時鐘的外 在表徵來研究掩蓋現象。 其次,德國蜚蠊與雙紋姬蠊這兩個同 胞種在交尾前期的日週律動的表現有很大 的差異,是探討「日週律動掩蓋機制」很 好的材料。在基因層次的研究方面,針對 時鐘基因 per 在德國蜚蠊與雙紋姬蠊之 DNA 序列上做比較,希望從時鐘基因 per 的分子演化上了解影響生物時鐘表現的因 子為何。 最後,由於掩蓋現象是生物時鐘節律 器 (pacemaker) 的 時 間 訊 息 在 輸 出 (output) 的過程中受到生殖因子的影響而 造成活動行為日週律動消失,為了想要知 道生物時鐘與生殖的關係在其它物種的情 況為何,我們比較不同職務的白蟻(生殖型 與 非 生 殖 型 ) 之 PER 、 PDF 以 及 Corazonin 三種蛋白質在不同時間點的表 現差異(包含日夜變化),分析其作用位置與 活動行為日週律動訊息傳遞路徑的關係, 進而了解此掩蓋作用之機制,同時藉此研 究對昆蟲之生物時鐘與生殖的關係有更進 一步的了解。 關鍵詞: 掩蓋效應、活動行為、日週律動、 時鐘基因、視網膜電位圖譜、德國蜚蠊、 雙紋姬蠊、交尾活性、去氧核醣核酸序列、 白蟻。 AbstractThe locomotor circadian rhythm was masked by the development of ovaries in female German cockroach (Blattella
germanica). This study explores the
masking mechanisms of the biological clock in cockroach. Firstly, we proceed the detection of Electroantennogram (EAG) as the second external indicator of circadian clock. We realize the localization of masking mechanism of biological clock based on EAG results. Since the masking phenomenon is result from physiological activity of reproduction, we choose the mating activity as an other indicator to study the masking effect.
Secondly, Blattella germanica and B.
bisignata are close-related species, but they
show huge differences in the expression of circadian rhythm s before mating. The comparative study of the expression of circadian rhythm between two species, is a good theme in the investigation of ecological significance of circadian rhythm. Comparing the DNA sequences of per gene between B. germanica and B. bisignata, we understand which factor influences the
expression of circadian clock based on the molecular evolution of per gene.
Finally, in order to understand the effect of reproductive factor on the output pathway and masking the circadian rhythm of locomotion, termite are used to investigate the relationship between reproduction and circadian clock in non-cockroach species. We compare the expression of three clock gene (per, pdf and corazonin) in individual of different casts at different time point. This result shows the locations where these clock genes the interaction takes place between the expression and transmission pathway of time signal for locomotion. Based on the above research, the masking mechanism are understood clearly.
Keywords: masking effect, locomotor
behavior, circadian rhythm,
period gene, Electroretinogram , Blattella germanica, Blattella bisignata, mating activity, DNA
sequence,termite, PDF, Corazonin. 二、緣由與目的 基因表現、生理週期性變動與行為表 現上的日週律動,都是由生物個體內的調 律器(pacemaker)所控制,也由於調律器屬 於細胞層次,因此也被稱為時鐘細胞(clock cell)。生物時鐘運作系統主要有三個組成 (Takahashi, 1995) : ( 一 ) 調 律 器 (pacemaker),是產生時間訊息之所在。(二) 環境訊息輸入路徑(input pathway),負責傳 遞外界環境訊息至調律器中樞,以使調律 器能與外界環境時間同步。(三)時間訊息輸 出路徑(output pathway) ,傳遞調律器所生 之時間訊息至下游,在生化上、生理上、 行為上表現出日週律動現象。目前,在生 物時鐘運作機制研究的領域裡,已建立出 調律器中樞時間訊息產生的分子機制。整 個日週律動系統是由自我調節的迴饋循環 (autoregulatory feedback loop) 所 控 制 (Dunlap, 1999)。 第一個生物時鐘基因 per,由 Konopka 及Benzer 於 197l 年所發表,他們以黃果蠅 (Drosophila melanogaster)羽化為日週律動 之觀察指標,找到了 per 基因的突變型。 從此以後,開始生物時鐘分子機制研究的 新紀元。per 基因在多種果蠅和許多昆蟲都 有發現,而且胺基酸序列有相似的保守性 (Colot et al., 1988)。在日夜二十四小時交替 下的LD 環境中 perRNA 及 PER 蛋白質表 現量呈穩定約二十四小時週期,而最大的 表現量都在夜晚。在全暗 DD 的環境下, 也就是沒有外界環境時間訊息的情況下,
per RNA 及 PER 蛋白質表現量呈現一個週
期 接 近 二 十 四 小 時 的 自 由 律 動 (free-running)。這種現象行為觀測的結果相 符。 德國蜚蠊雌性成蟲的卵巢在發育的過 程中,會抑制雌性成蟲活動行為的日週律 動;但是卵巢摘除後的雌成蟲則恢復具有 日週律動的活動行為(Lin and Lee, 1998)。 關於卵巢掩蓋活動行為日週律動現象的機 制並不十分清楚。掩蓋的機制有兩種可能 性:(一)卵巢存在擾亂了調律器的運作,故 活動行為不具日週律動現象。(二)卵巢存在 阻礙訊息輸出路徑,故活動行為不具日週 律動現象。藉由研究日週律動被掩蓋的現 象,有助於我們找出正常的訊息輸出路 徑,進一步建立訊息輸出路徑的可能機制。 要解答以上的問題可以 per 基因表現 情形作為分子指標,常用的方式是用北方 墨點法(northern blotting)偵測 RNA 的表 現,或是用西方墨點法(western blotting)偵 測蛋白質的表現;此外也可藉由找尋其他 指標進行偵測判斷。我們用觸角神經圖譜 (elctroretinogram (ERG))以及交尾活性 (mating activity) 這兩種指標來瞭解掩蓋作 用與訊息輸出路徑的關係。 觸角神經圖譜已經被證實是測量嗅覺 感受性很有效的方法,已有許多昆蟲利用 ERG 來測量視覺感受性的日週律動(Rosén et al., 2004)。因為 EAG 是一種很好的測量 生理反應日週律動的指標,本計畫利用嗅 覺感受性的日週律動系統來瞭解德國蜚蠊 的日週律動系統的調控。尤其雌蟲活動行 為日週律動的掩蓋現象,是否會造成EAG 的不規律表現,可以證實卵巢發育之掩蓋 機制是否位於調律器上或時間訊息的輸出 路徑上。 另一個指標是交尾活性。Sakai and
3 Ishida (2001) 證實果蠅的交尾活性呈現出 日週律動,而且該現象是受時鐘基因 per 控制。同樣地,為了證實卵巢發育之掩蓋 機制是否位於調律器上或時間訊息的輸出 路徑上,我們想知道雌蟲活動行為日週律 動的掩蓋現象,是否會造成交尾活性的非 日週律動表現。 本計畫也將從德國蜚蠊與雙紋姬蠊活 動行為規律性之差異來研究掩蓋效應的成 因。德國蜚蠊在性接受期間,不僅夜晚的 活動量高,白天的活動量亦很高,沒有表 現 出 明 顯 的 晝 夜 律 動 (Lee and Wu, 1994)。雙紋姬蠊雌蟲的活動模式則顯然與 德國蜚蠊不同:雙紋姬蠊在性接受期間活 動量也會增高,但大部份的活動仍侷限於 夜晚,因此仍表現日週律動;由於德國蜚 蠊與雙紋姬蠊是同胞種,但在活動行為日 週律動之掩蓋現象上卻有截然不同的表 現,因此將以雙紋姬蠊為材料探討掩蓋現 象不發生的原因。因為在種化的過程中常 常 伴 隨 而 來 的 是 遺 傳 組 成 上 的 分 化 (Genetic differentiation),為了對雙紋姬蠊的 日週律動的分子調控機制有更深的了解, 必須先對雙紋姬蠊 per 基因有基本的認 識,此部份的研究首先要進行的是雙紋姬 蠊 per 基因的 DNA 序列分析。 許 多 實 驗 證 明 pigment-dispersing factor (pdf)基因與日週律動活動之訊息輸 出有關, 也就是節律器用來影響其下游目 標細胞的作用分子。將PDH 注射入馬德拉 蟑螂腦內的 accessory medulla 附近,結果 會使得馬德拉蟑螂每日活動開始之時間提 前或延後(Petri and Stengl, 1997)。 pdf 基因 突變的果蠅,受到光暗周期的導引作用 時,能表現近似正常的活動日週律動(其 light-off activity 提前 1 小時開始);若將其 置於完全黑暗的環境中,前二天還能表現 近似正常的活動日週律動,但自第三天 起,活動之日週律動就消失了(Renn et al., 1999)。此外 pdf 基因的表現受到許多 clock 基因的調控;研究發現當果蠅的 clock 或 cycle 基因發生突變,均會影響 pdf RNA 的 生成,所以推測 pdf 基因的轉錄應是受到 CLOCK 、CYCLE 這兩種轉錄因子的調控 (Park et al., 2000);而 period 或 timeless 基
因突變果蠅之PDF 含量,則有週期異常或
含量持續過低的現象,由此可推論,period 及 timeless 基因之產物對 pdf 基因的轉錄 並 無 影 響 , 其 作 用 之 層 次 應 是 在 轉 譯 (translation)的階段 (Park et al., 2000)。
另一個與日週律動活動之訊息輸出有 關 的 基 因 為 corazonin (cr) , 其 產 物 (Corazonin) 是 昆 蟲 的 神 經 調 節 物 (neuropeptides) , 有 [Arg7]-corazonin 和 [His7]-corazonin 2 種 homologs,最初是在 美洲蜚蠊的心側腺 (corpora cardiaca, CC) 中被發現;corazonin 普遍存在於各種昆蟲 內,但有多種功用 (Veenstra, 1989; Tanaka
et al., 2002; Roller et al., 2003)。目前猜測
corazonin 可能是日週律動生物時鐘訊息 輸出路徑的調節物,因為 (1) 在 Manduca
sexta 中,[Arg7]-corazonin 和 PER 的表
現 位 置 相 同 ;(2) 在 Bombyx mori 中, corazonin 和 DBT 的 表 現 位 置 相 同 (Qi-miao et al., 2003)。 本 計 畫 打 算 用 西 方 墨 點 法(western blotting)偵測 PDF 與 Corazonin 蛋白質的 表現,進而了解這些Output 基因與掩蓋效 應的關係。 此部份的研究所用的材料為白 蟻 (termite) ,因為白蟻有社會結構,族群 中的個體有生殖型與非生殖型 (工蟻或兵 蟻)兩類;利用這個特性我們可以比較這兩 種個體之時鐘基因的表現,藉此對昆蟲之 生物時鐘與生殖的關係有更進一步的了 解。 三、結果與討論 (一) 觸角電位圖譜 (Electroantennogram, EAG) 日週律動的偵測: 1. 給予德國蜚蠊一秒鐘之醋酸乙酯(ethyl acetate)刺 激 的 觸 角 電 位 圖 譜 反 應 ( 圖 一) ,其反應強度的變異值約為 0.04mV (-0.08~-0.12)(重覆六次)。 -1 0 1 2 3 4 5 6 -0.14 -0.12 -0.10 -0.08 -0.06 -0.04 Ampl it ud e ( m V) Time (S) 圖一、 德國蜚蠊雄蟲對醋酸乙酯的觸角電
位圖譜。橫軸代表時間,縱軸是反應強度 (amplitude). 2. 醋 酸 乙 酯 的 稀 釋 反 應 曲 線 (dilution response curve) 見圖二。結果顯示醋酸乙 酯在 稀釋到十倍以上後就無法引起德國蜚 蠊的觸角電位圖譜反應。 -5 -4 -3 -2 -1 0 0.00 0.01 0.02 0.03 0.04 0.05 A m plitude (mV ) Concentration (log 10) 圖二、德國蜚蠊雄蟲對不同濃度之醋酸乙 酯的觸角電位圖譜。橫軸是醋酸乙酯的濃 度,縱軸是反應強度;每個點是六次重覆 的平均(每次重覆間隔五分鐘) 3. 圖三是德國蜚蠊雌蟲在不同時間的醋酸 乙酯刺激下,觸角電位圖譜反應的日週律 動分析圖。結果顯示刺激時間愈長,觸角 電位反應愈大。 圖三、. 德國蜚蠊雌蟲的觸角電位圖譜。橫 軸是醋酸乙酯的刺激時間,縱軸是反應強 度。 4. 圖四是德國蜚蠊觸角電位圖譜反應的日 週律動分析圖。圖中顯示雌性與雄性的德 國蜚蠊對 100% 的醋酸乙酯在不同的時 間點有不同的觸角電位圖譜反應強度。 圖四、五天大之德國蜚蠊(A)雄蟲與(B)雌蟲 對醋酸乙酯之觸角電位圖譜的日週律動分 析。.實驗條件為 16 小時光照 8 小時黑暗 (圖上方的白色長方形代表光期,黑色代表 暗期,灰色代表自由律動期)。將德國蜚蠊 先在有日夜變化的實驗條件下先進行導引 (entrainment) ,接著進入全暗期讓其自由 律動。每兩小時取樣一次。圖中的平滑線 是 觸角電位反應之變化的最適曲線(best fitted line). (二) 、德國蜚蠊交尾活性之日週律動 圖五的結果顯示德國蜚蠊之交尾活性 (mating activity) 有日週律動,也就是在該 特徵上並沒有掩蓋效應出現。我們發現德 國蜚蠊的交尾高峰出現在暗期快結束與光 期開始。 圖五、德國蜚蠊交尾活性的日夜變化。實 驗條件為 16 小時光照 8 小時黑暗。 (三)、雙紋姬蠊 per 基因之 DNA 序列分 析: 抽取雙紋姬蠊頭部之 mRNA 後,再利 用 degernerate primer 進行反轉錄 PCR 反 應 (Reverse-transcription polymerase chain reaction) ,最後跑膠得到一個大小約 1Kb
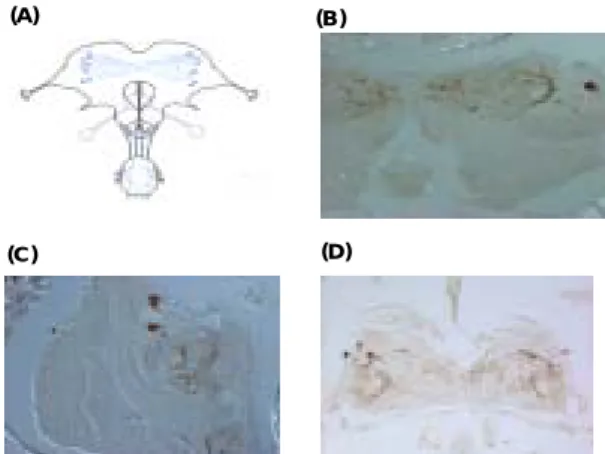
5 的產物。將該產物進行核酸定序後, 與實 驗 室 之 前 找 到 的 德 國 蜚 蠊 per 基 因 之 DNA 序列做比對,結果顯示在 759 bp 長 度的比對序列中,兩者的 DNA 相似度 (identities) 約為 93%;接著再以美洲蜚蠊 (Periplaneta americana)做外群 (outgroup) 做DNA 序列比對,結果顯示在 575 bp 的 比 對 序 列 中 , 兩 者 的 DNA 相 似 度 (identities) 約為 74%。 由此可知雙紋姬蠊 per 基因之 DNA 序列與德國蜚蠊非常相 似,符合預期。 (四)、各種職務之白蟻的頭部神經組織的免 疫染色 (immunostaining) 我們針對與日週律動相關的兩種蛋白 質 (PDF、 Corazonin),對各種職務之白蟻 的頭部神經組織進行免疫染色,希望了解 社會性昆蟲白蟻的生物時鐘是否因職務不 同而不同。取 Pseudogates、Solider (兵蟻) 以及reproductive nymph 之佛羅里達濕木白 蟻 Prorhinotermes simplex,針對不同時間 點之白蟻進行 anti-PDF、anti-corazonin 免 疫染色,以判斷 PDF、corazonin 兩種蛋白 質之表現是否具有日週律動。此外還比較 在 長 日 照 (L:D= 16:8) 與 短 日 照 (L:D =10:14)的不同處理下三種蛋白質之表現。 圖六的結果顯示Pseudogates、Reproductive nymph 和 Solider 染出的結果是一樣的(不 同時間點、不同性別、不同光期處理)。 A B C D
圖 六 、(A). Pseudogate (B) Reproductive nymph 以及 (C) Solider 三種職務之白蟻 anti-PDF 免疫染色結果示意圖;(D) Solider
切片,腦部表現 PDF 之細胞與軸突。
在 Corazonin 的結果方面(圖七),
Pseudogates 和 Reproductive nymph 以及 Solider 三者染出的結果是一樣的出的結果 是一樣的(不同時間點、不同性別、不同光 期處理)。染出的細胞數目各有差異,推測 應是個體差異。 (A) (B) (C) (D)
Fig. 7. (A). Pseudogate 之 anti-Corazonin 免疫染色結果示意圖(B) Pseudogate (C) Reproductive nymph 以及(D) Solider 三種 職務之白蟻的切片,腦部表現 Corazonin 之細胞與軸突。 四、計畫成果自評 本計畫為三年期計畫,研究內容皆依 原計畫期程分年執行,與原計畫大致相 符。在計畫執行期間,我們達成了下列預 期目標: (一)、完成了德國蜚蠊觸角電位圖譜 (Electroantennogram, EAG) 日週律動 的偵測,由此可知造成活動行為日週 律動消失的因子並未作用在觸角的 嗅覺反應上。 (二)、德國蜚蠊之交尾活性表現出日週律 動,顯示雖然雌蟲交尾前期的活動 行為的日週律動因為卵巢的發育而 被掩蓋,但掩蓋並未作用在交尾活 性的表現上。 (三)、比較雙紋姬蠊與德國蜚蠊 per 基因後 發現兩者之 DNA 序列差異不大, 推測兩者在活動行為日週律動表現 上的差異可能是 per 基因表現上 的差異,或是生物時鐘節律器的時 間訊息在輸出的過程中受到其它因 子的影響而造成德國蜚蠊活動行為 日週律動消失。 (四)、比較生殖型與非生殖型白蟻頭
部織的免疫染色,結果顯示兩者在 PDF 與 Corazonin 的表現上在各 個處理組間並無不同,同時在光期 與暗期的表現上也沒有差異。 五、參考文獻
Colot, H. V., J. C. Hall, and M. Rosbash. 1988. Interspecific comparasion of
period gene of Drosophila reveals
large blocks of non-conserved coding DNA. EMBO 7: 3929-3937.
Crispi, S., E. Giordano, P. P. D’Avino, and M. Furia. 1998. Cross-talking among Drosophila nuclear receptors at the promiscuous response element of the ng-1 and ng-2 intermolt genes. J. Mol. Bio. 275: 561-574.
Darlington, T. K., K. Wager-Smith, M. F. Ceriani, D. Staknis, N. Gekakis, T. D. L. Steeves, C. J. Weitz, J. S. Takahashi, and S. A. Kay. 1998. Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim. Scinece 280: 1599-1603.
Dunlap, J. C. 1999. Molecular bases for circadian clocks. Cell 96: 271-290. Glossop, N. R. J., L. C. Lyons and P. E.
Hardin. 1999. Interlocked feedback loops within the Drosophila circadian oscillator. Science 286: 766-768. Helfrich-Forster C., M. Stengl, and U.
Homberg. 1998. Organization of the circadian system in insects. Chronobiol. Int. 15: 567-594.
Hogenesch, J. B., Y. Gu, S. Jain and C. A.
Bradfield. 1998. The Basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 95: 5474-5479.
Honig, M. G., and R. I. Hume. 1989. DiI and DiO: versatile fluorescent dyes for neuronal labeling and pathway tracing. TINS 12: 333-341.
Jindra, M., F. Malone, K. Hiruma, and L. M. Riddiford. 1996. Developmental profiles and ecdysteroid regulation of the mRNAs for two ecdysone
receptor isoforms in the epidermis and wings of the tobacco hornworm,
Manduca sexta. Dev. Bio. 180:
258-272.
King-Jones, K., G. Korge, and M. Lehmann. 1999. The helix-loop-helix proteins dAP-4 and daughterless bind both in
vitro and in vivo to SEBP3 sites
required for transcriptional activation of the Drosophlia Gene Sgd-4. J. Mol. Bio. 291: 71-82.
Koehler W. K., and G. Fleissner. 1978. Internal desynchronization of bilaterally organized circadian oscillators in the visual system of insects. Nature 174: 708-710.
Konopka, R. J. and S. Benzer. 1971. Clock mutants of Drosophila
melanogaster. Proc. Natl. Acad. Sci.
7
Koontz, M., and J. S. Edward. 1980. The projection of neuroendocrine fibers in the brain of three Orthopteroid insect. J. Morphol. 165: 285-299.
Lee, C., K. Bae, and l. Edery. 1999. PER and TIM inhibit the DNA binding activity of a Drosophila CLOCK-CYC/dBMALI heterodimer without disrupting formation of the heterodimer: a basis for circadian transcription. Molecular and Cellular Biology 19: 5316-5325.
Lee, H. J. and Y. L. Wu. 1994. Mating effects on the feeding and locomotion of the German cockroach, Blattella
germanica. Physiol. Entomol. 19:
39-45.
Lezzi, M., T. Bergman, J. -F. Mouillet, and V. C. Henrich. 1999. The ecdysone receptor puzzle. Arch. Insect Bio. Physio. 41: 99-106.
Lin, T. M., and H. J. Lee. 1996. The expression of locomotor circadian rhythm in female German cockroach,
Blattella germanica (L.).
Chronobiol. Int. 13: 81-91.
Lin T. M., and H. J. Lee. 1998. Parallel control mechanisms underlying locomotor activity and sexual receptivity of the female German cockroach, Blattella germanica (L.). J. Insect Physiol. 44: 1039-1051.
Lococo, D. J., and S. S. Tobe. 1984a. Neuroanatomy of the retrocerebral
complex, in particular the pars
intercerebralis and pars lateralis in the cockroach Diploptera punctata. Int. J. Insect Morphol. Embryol. 13: 65-76.
Lococo, D. J., and S. S. Tobe. 1984b. Retrograde and orthograde axon transport by brief-exposure to nickel chloride: methodology and parameters for success in the brain-retrocerebral complex of the cockroach Diploptera punctata. J. Insect Physiol. 30: 635-642.
Park J. H., Helfrich-Förster C., Lee G., Liu L., Rosbash M., and Hall J. C. 2000. Differential regulation of circadian pacemaker output by separate clock genes in Drosophila. Proc. Natl. Acad. Sci. USA 97: 3608-3613.
Petri B. and Stengl M. 1997.
Pigment-dispersing hormone shifts the phase of the circadian pacemaker of the cockroach Leucophaea
maderae. J. Neurosci. 17: 4087-4093.
Plautz, J. D., M. Kaneko, J. C. Hall, and S.A.Kay. 1997. Independent photoreceptive circadian clock throughout Drosophila. Science 278:1632-1635.
Renn S. C. P., Park J. H., Rosbash M., Hall J. C., and Taghert P. H. 1999. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause
severe abnormalities of behavioral circadian rhythms in Drosophila. Cell 99: 791-802.
Roller, L., Tanaka, Y., Tanaka, S., 2003. Corazonin and corazoninlike substances in the central nervous system of pterygote and apterygote insects. Cell and Tissue Research 312, 393–406.
Rosén, W. Q. ,G.-B. Han, and C. Lofstedt. 2004. The Circadian Rhythm of the sex-Pheromone-Mediated Behavioral Response in the Turnip Moth, Agrotis
segetum, Is Not Controlled at the
Peripheral Level. J. biol. Rhythm. 18: 402-08.
Saez, L. and M. W. Young. 1988. In situ localization of the per clock protein during development of Drosophila
melanogaster. Mol. Cell. Biol. 8:
5378-5385.
Sakai, T., and N. Ishida. 2001. Circadian rhythms of female mating activity governed by clock genes in
Drosophila. PNAS 98:9221-9225.
Sassone-Corsi, P. 1998. Molecular clocks: mastering time by gene regulation. Nature 392:871-874.
Sauman, I. and S. M. Repper. 1996. Circadian clock neurons in the silkmoth Antheraea pernyi: novel mechanisms of Period protein regulation. Neuron 17: 889-900. Stocker, A. J., J. M. Amabis, E. Gorab, C.
Elke, and M. Lezzi. 1997.
Antibodies against the D-domain of a
Chironomus ecdysone receptor
protein react with DNA puff sites in Trichosia pubescens. Chromosoma 106:456-464.
Takahashi, J. S. 1995. Molecular neurobiology and genetics ofcircadian rhythms in mammals. Annu. Rev. Neurosci. 18: 531-553.
Tanaka, S., Zhu, D.-H., Hoste, B., Breuer, M., 2002. The dark-color inducing
neuropeptide, [His7]-corazonin, causes a shift in morphometric characteristics towards the gregarious phase in isolatedreared (solitarious)
Locusta migratoria. Journal of Insect
Physiology 48, 1065–1074.
von Bartheld, C. S., D. E. Cunningham, and E. W. Rubel. 1990. Neuronal tracing with DiI: decalcification, cryosectioning, and photoconversion for light and electron microscopic analysis. J. Histochem. Cytochrm. 38: 725-733.
Wen, H. W., and H. J. Lee. 2000. Unequal coupling between locomotor pacemakers of the German cockroach,
Blattella germanica(L.). J. Insect
Physiol. 46: 89-97.
Wills S. A., T. L. Page, and C. S. Colwell. 1985. Circadian rhythms in the electroretinogram of cockroach. J. Biol. Rhythms 1: 35-37.