碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
初探山苦瓜萃物對 3T3-L1 脂肪細胞褐化及粒線體增生 相關基因表現之影響
An initial approach to explore the effects of Momordica charantia L. extracts on the mRNA expression of
browning and mitochondria biogenesis related genes in 3T3-L1 adipocytes
王思文 Si-Wen Wang 指導:黃青真博士
Advisor:Ching-Jang Huang, Ph.D.
中華民國 102 年 7 月
July, 2013
中文摘要
近年臺灣地區由於西方飲食的盛行及體能活動量不足,過重及肥胖盛行率在 不同性別及年齡層逐年增加。肥胖防治的主要策略是降低能量攝取、增加能量消 耗。最近研究顯示,刺激白色脂肪「褐化」可消耗熱量,抑制脂肪組織累積,防 止肥胖。本實驗室先前研究,小鼠攝食山苦瓜飲食可促進白色脂肪中「棕色脂肪」
及粒線體相關基因mRNA 表現,並增加能量代謝速率。本研究擬建立 3T3-L1 脂 肪細胞培養為試驗平台,觀察苦瓜萃物於培養之白色脂肪細胞是否能誘發褐化及 粒線體增生的特徵,並進一步觀察比較不同山苦瓜萃物/區分物對脂肪細胞褐化及 增加粒線體相關基因的活性。
首先以 5 μM troglitazone 處理,分別比較兩種分化法以及在 3 種處理時間,對 於成熟脂肪細胞褐化及粒線體增生相關基因表現的影響。根據實驗結果,選擇「分 化成熟前四天,樣品處理四天」,並以「分化法1」的脂肪細胞模式進行後續實驗。
試驗樣品包括:山苦瓜乙酸乙酯萃物 (EAE) 、皂化物 (S) 、不皂化物 (NS) 、山 苦瓜水萃物 (WE) 、經納豆菌作用後之水萃物 (WEn) 與自苦瓜純化分離的化合 物 (CLN、 phytol 和 leutin) 。結果顯示,低濃度的 EAE、S 及 NS 處理下,能觀 察到棕色脂肪相關基因表現增加;而EAE 也能增加脂肪細胞檸檬酸合成酶 (citrate synthase, CS) 的活性。而苦瓜中活化 PPARγ 的活性成分也能刺激脂肪細胞的褐 化。
綜合上述,3T3-L1 脂肪細胞可作為觀察褐化現象的試驗模式。而山苦瓜能活 化PPARγ 之區分物/化合物,皆可使脂肪細胞表現褐化相關基因,與文獻中 PPARγ agonist 可使白色脂肪細胞表現「褐化」之現象相符。
Abstract
Due to westernized diet and insufficient physical activities, the prevalence of overweight and obesity have been increased Taiwan and other parts of the world.
Managing energy balance is the principle of obesity prevention/treatment. Recent studies demonstrated that “browning”of white adipose tissue (WAT) might increase energy expenditure and prevent obesity. Previous study of our lab showed that mice fed the wild bitter gourd (Momordica charantia.L, WBG) diet had higher metabolic rate and some characteristics of “browning” WAT, i.e., higher expressions of brown-fat-selective and mitochondria biogensis genes. This study thus aimed to develop a cell model using cultured 3T3-L1 adipocytes for the examination of mRNA expressions of
brown-fat-selective and mitochondria biogensis genes as affected by various WBG extracts/fractions and compounds.
Two differentiation protocols and three treating stage/times were compared using 5 μM troglitazone as the positive control. The differentiation protocol 1 and treatment during the last 4 days of differentiation showed the best results. Ethyl acetate extract (EAE), its saponifable (S) and non-saponifable fraction (NS), water extract without (WE) or with pretreatment (WEn) as well as 3 PPAR active compounds of WBG were then tested in the system. Low concentrations of EAE、S and NS did induce brown-fat gene expression in 3T3-L1 adipocytes (p<0.05), and EAE can also increase the citrate synthase activity (p<0.05). The induction of brown-fat gene expressions were also observed in cells treated with the 3 PPAR active compounds of WBG (conjugated linolenic acid, phytol and leutin), implying the involvement of WBG PPARγ activity in the “browning” effect.
In conclusion, 3T3-L1 adipocytes can be used as a model to investigate “browning”
effect of WBG. The PPARγ active fractions and compounds of WBG induced mRNA
expressions of brown-fat-selective genes in this cell model.
keywords:obesity, Momordica charantia, adipocytes, browning, mitochondira
縮寫對照表
縮寫 全名
BGP
Bitter gourdCLA
Conjugated linoleic acidCLN
Conjugated linolenic acidCS
Citrate synthaseNS
Non- SaponifiablePPAR
Peroxisome proliferator-activated receptorRS
Rat serumS
SaponifiableTrog
TroglitazoneWE
Water extract of Bitter gourd freeze powderWEn
WE hydrolysis by bacillus subtilis natto culture in 5 % black soymilk總目錄
中文摘要………Ⅰ 英文摘要………Ⅱ 縮寫對照表………Ⅳ 總目錄……….Ⅴ 圖目錄……….Ⅷ 表目錄………...Ⅸ
第一章 緒論………...1
第一節 前言………...1
第二節 文獻回顧………...2
一、代謝症候群………2
二、過氧化體增殖劑活化受體……….2
(一) PPAR 活化劑與 ligands………...3
(二) PPARγ 與脂肪分化………...3
(三) PPAR 與代謝症候群………4
三、脂肪細胞褐化現象………4
(一) 脂肪組織……….…4
(二) 去偶聯蛋白……….5
(三) PPAR-γ co-activator-1 α (PGC1-α) ………6
(四) 白色脂肪「褐化現象」(browning) ……….8
(五) 基因及粒線體指標的篩選………...8
四、山苦瓜………11
第三節 實驗假說與實驗架構……….19
一、實驗假說……….19
二、實驗架構……….19
第二章 山苦瓜萃物對於脂肪細胞褐化相關基因表現之影響……….20
第一節 前言……….20
第二節 材料與方法……….21
一、白色脂肪細胞模式建立與最佳化………...21
二、山苦瓜萃物促進脂肪細胞褐化………...21
三、細胞株………...22
四、培養基與藥品試劑………...22
(一) 細胞培養與分化………...22
(二) 檸檬酸合成酶活性定………...…………. …...23
(三) quantitative Real-time PCR (qRT-PCR) 法分析基因表現……..…23
(四) 處理藥品………...24
五、儀器設備………...24
六、山苦瓜萃物之製備………...25
(一) 山苦瓜乙酸乙酯萃物暨其區分物………...25
(二) 山苦瓜水萃物………...…………. …...25
(三) 經 5 % 黑豆漿培養之納豆菌處理山苦瓜水萃物...25
七、實驗方法………...25
(一) 細胞培養………...25
(二) 細胞分化………...…………. ………...26
(三) 蛋白質測定……. …………...…………. ………...26
(四) 處理藥品………...…………. ………...27
(五) 檸檬酸合成酶活性測定………...27
(六) 基因表現………...…………. …...27
(七) 數據整理及統計分析……..……….28
第三節 實驗結果……….29
一、白色脂肪細胞褐化實驗模式建立與最佳化……….29
二、山苦瓜萃物處理 3T3-L1 脂肪細胞褐化相關基因表現………31
三、山苦瓜萃物處理 3T3-L1 脂肪細胞之檸檬酸合成酶 (CS) 活性…….33
四、山苦瓜中刺激 3T3-L1 脂肪細胞褐化基因表現的可能活性成分……34
第四節 討論……….55
一、 比較不同分化法及處理時間建立 3T3-L1 脂肪細胞褐化最佳模式…55 二、 山苦瓜萃物對於 3T3-L1 脂肪細胞褐化現象的影響………56
三、 山苦瓜中刺激 3T3-L1 脂肪細胞褐化基因表現的可能活性成……..58
第五節 結論……….60
第三章 綜合討論與總結論……….61
第一節 綜合討論………...…..…61
第二節 總結論……….63
第四章 參考文獻………...65
圖目錄
圖 1-1 成年人類中可偵測到 UCP1 positive 的棕色脂肪……….5
圖 1-2 棕色脂肪中 UCP1 之產熱作用機制………...6
圖 1-3 PPAR 受不同訊息分子所調控………...7
圖 1-4 PGC1 之調控機制………...7
圖 1-5 白色脂肪棕色化………....11
圖 1-6 山苦瓜之各種粗萃物之製備及區分過程……….…………...13
圖 1-7 共軛脂肪酸 CLA 和 CLN 之化學結構………...14
圖 2-1 分化成熟前 2 天以 troglitazone 處理 2 天對 3T3-L1 脂肪細胞褐化及粒線體 相關基因 mRNA 表現之影響…...36
圖 2-2 分化成熟前 2 天以 troglitazone 處理 4 天對 3T3-L1 脂肪細胞褐化及粒線體 相關基因 mRNA 表現之影響…...37
圖 2-3 分化成熟前 4 天以 troglitazone 處理 4 天對 3T3-L1 脂肪細胞褐化及粒線體 相關基因 mRNA 表現之影響…...38
圖 2-4 兩種分化方法及三種處理時間對 troglitazone 處理 3T3-L1 脂肪細胞之褐化 與粒線體生成作用相關基因 mRNA 表現之比較…...39
圖 2-5 山苦瓜乙酸乙酯萃物 (EAE) 及其皂化物 (S) 與不皂化物 (NS) 對 3T3-L1 脂肪細胞棕脂相關基因 mRNA 表現的影響…...40
圖 2-6 山苦瓜乙酸乙酯萃物 (EAE) 及其皂化物 (S) 與不皂化物 (NS) 對 3T3-L1 脂肪細胞粒線體相關基因 mRNA 表現的影響…...41
圖 2-7 山苦瓜乙酸乙酯萃物 (EAE) 及其皂化物 (S) 與不皂化物 (NS) 對 3T3-L1 脂肪細胞白色脂肪相關基因 mRNA 表現的影響…...42
圖 2-8 山苦瓜乙酸乙酯萃物 (EAE) 及其皂化物 (S) 與不皂化物 (NS) 對 3T3-L1 脂肪細胞轉錄因子相關基因 mRNA 表現的影響…...43 圖 2-9 山苦瓜乙酸乙酯萃物暨其區分物添加 forskolin 對分化中脂肪細胞 UCP1 及
C/EBPβ mRNA 的影響…...44 圖 2-10 山苦瓜水萃物及經納豆菌作用之水萃物對 3T3-L1 脂肪細胞棕脂相關基因 mRNA 表現的影響...45 圖 2-11 山苦瓜水萃物及經培養於 5 % 黑豆漿納豆菌作用之水萃物對 3T3-L1 脂肪 細胞粒線體生成相關基因 mRNA 表現的影響...46 圖 2-12 山苦瓜水萃物及經納豆菌作用之水萃物對 3T3-L1 脂肪細胞白色脂肪相關 基因 mRNA 表現的影響...47 圖 2-13 山苦瓜水萃物及經納豆菌作用之水萃物對 3T3-L1 脂肪細胞轉錄因子相關 基因 mRNA 表現的影響...48 圖 2-14 不同山苦瓜萃物對 3T3-L1 脂肪細胞檸檬酸合成酶活性之影響...49 圖 2-15 山苦瓜乙酸乙酯萃物暨其區分物與 CLN 之紫外光吸光圖譜...50 圖 2-16 山苦瓜 PPAR 活性成分對 3T3-L1 脂肪細胞棕脂相關基因 mRNA 表現之 影響...51 圖 2-17 山苦瓜 PPAR 活性成分對 3T3-L1 脂肪細胞粒線體生合成相關基因 Mrna 表現之影響...52 圖 2-18 山苦瓜 PPAR 活性成分對 3T3-L1 脂肪細胞白色脂肪相關基因 mRNA 表 現之影響...53 圖 2-19 山苦瓜 PPAR 活性成分對 3T3-L1 脂肪細胞轉錄因子相關基因 mRNA 表 現之影響...54
表目錄
表 1-1 PPAR agonist 刺激白色脂肪褐化之文獻整理………...15
第一章 緒論
第一節 前言臺灣地區近年來由於經濟成長及營養狀況的改善,加上速食文化的強力促銷 下,高熱量、高脂肪食品,不但成為國人的熱門食物,更成為外食族的主要食物 來源。除了熱量和脂肪的過度攝取,加上體能活動量不足及靜態活動增加,過重 及肥胖盛行率在不同性別及年齡層有逐年增加趨勢,也造成台灣地區第二型糖尿 病的 「年輕化」 ,主因是當身體脂肪組織越多,細胞越容易對胰島素出現抗拒。
根據衛生署國民健康局 「2012 台歐健康論壇」 統計發現,全台成年人三成男性 體重過重 (BMI 身體質量指數≧24),近兩成男性肥胖(BMI≧ 27),肥胖率是南 韓十二倍、日本八倍;而女性有近兩成過重,一成七者肥胖,肥胖盛行率為亞洲 之冠,也是全球代謝症候群發生率第二高的國家 (衛生署,2011) 。根據 100 年衛 生署公布十大死因中,有多項與肥胖有關,其中糖尿病 (死亡人數占率 6 %) 排名 第四,而高血壓 (占率 3 %) 也列於前十。這些調查顯示,肥胖及糖尿病正危及國 人健康。而在非手術性的治療肥胖方式中,健康的飲食及適量的運動無疑是最正 當的一種,但不易維持長久。根據最近的研究,以刺激白色脂肪細胞褐化 (Browning) 的方式來增加能量的消耗來減重,或許是研發更有效治療肥胖的方向。
本實驗室最近的研究指出,苦瓜除降血糖、血脂外,也能於長期餵食苦瓜的 小鼠中,觀察到白色脂肪組織中粒線體生成及棕色脂肪相關基因的表現增加,但 其有效成分及機制仍尚未被確認。另外也有研究顯示,體外培養的脂肪細胞經適 當處理亦可觀察到白色脂肪細胞褐化的現象。故本研究擬建立3T3-L1 脂肪細胞培 養為試驗平台,觀察苦瓜萃物於培養之脂肪細胞是否亦可誘發出褐化及粒線體增 生的相關基因表現,並進一步篩選出能促使脂肪細胞褐化及增加粒線體的活性之 苦瓜萃物/區分物。
第二節 文獻回顧 一、代謝症候群
代謝症候群 (metabolic syndrome, MetS),是指多種代謝異常的症狀集中於一 個體,使其未來發生心血管疾病及第二型糖尿病之風險增加。而這些慢性疾病,
皆年居台灣十大死因榜中,已成我國及世界重要上許多國家的公共衛生議 題。我國於2007 實施最新定義之代謝症候群判定標準 (行政院衛生署國民 健康局,2007) ,將 5 項危險因子中之腹部肥胖,單獨列為 1 項危險因子;
刪除身體質量指數(BMI),及下修空腹血糖異常值切點至 100mg/dl。以下 5 項危險因子中,若包含 3 項或以上即者可判定為代謝症候群:
(1)腹部肥胖 (腰圍) :男性≧90cm、女性≧80cm。
(2)高血壓:收縮血壓 (SBP) ≧130mmHg、舒張血壓 (DBP) ≧85mmHg。
(3)高血糖:空腹血糖值 (FG) ≧100mg/dl。
(4)高密度酯蛋白膽固醇 (HDL-C):男性<40mg/dl、女性<50mg/dl。
(5)高三酸甘油酯 (TG) ≧150mg/dl。
其中血壓、空腹血糖值等 2 危險因子之判定,包括依醫師處方使用降血壓
或降血糖等藥品 (中、草藥除外),導致血壓或血糖檢驗值正常者。
代謝症候群的核心異常因子分別為腹部肥胖和胰島素抗阻。肥胖主要是由於 攝取過多的熱量,造成能量的失衡,使多餘熱量以油脂型式長期累積在脂肪組織 中。脂肪細胞除了以三酸甘油脂的型式儲存能量外,也是一種內分泌細胞,過多 的脂肪會促使脂肪細胞分泌細胞激素 (adipocytokines) ,造成發炎反應以及胰島素 阻抗。而胰島素阻抗是指細胞上之胰島素受器對胰島素的刺激敏感性降低或產生 抗性,使細胞無法有效攝入及利用血液中的葡萄糖,造成代謝異常,最後導致第 二型糖尿病及心血管疾病的發生。
二、過氧化體增殖劑活化受體
(1) PPAR α 主要分布於肝臟、心臟、肌肉等脂肪酸氧化代謝活性較高的器官,調 控脂質代謝。
(2) PPAR β/δ 廣泛分布於各種細胞,參與胚胎發育。
(3) PPAR γ 主要調控脂肪細胞分化和胰島素敏感度。
其中PPAR γ 又可區分成 PPARγ1, PPARγ2, PPARγ3。PPARγ1 於心、肝、肌肉 及脂肪中均有表現;PPARγ2 專一表現在脂肪組織中,調控脂肪細胞之分化;而 PPARγ3 則表現在巨噬細胞及脂肪組織中。PPARγ 基因功能的獲得及遺失與人類的 肥胖問題很有關聯,且與代謝疾病也息息相關。
(一) PPAR 活化劑與 ligands
PPAR 為核受器家族的成員,受專一性結合的小分子 (ligand)所調控。當 ligands 與受器蛋白質結合後,會改變受器的構型,使其與目標基因的啟動子序列 上的特定DNA 序列結合,進而啟動目標基因的轉錄,促進該基因表現。所以 ligands 是本身能與受器蛋白質直接結合而產生轉錄效應的小分子;而活化劑是指可以活 化受器的分子。目前PPARs 的 ligands 可分為合成性與天然性:
(1) 合成性 (synthetic ligands) :fibrate 降血脂藥 (如 clofibrate 等)、過氧化體增殖 劑 (如 WY-14643)、非類固醇抗發炎藥 (如 indomethacin) 、抗糖尿病藥物,例 如TZDs (包含 Rosiglitazone、Pioglitazone、Troglitazone) 。
(2) 天然性 (nature ligands) :長鏈不飽和脂肪酸、支鏈、共軛與氧化脂肪酸,例如 conjugated linoleic acid (CLA) 等。
(二) PPARγ 與脂肪分化
脂肪細胞是由前驅脂肪細胞 (preadipocyte) 分化而成,一般脂肪組織中皆存有 前脂肪細胞。前脂肪細胞株如3T3-L1,為脂肪細胞之分化提供良好之體外模式,
於培養期間即可觀察到其分化過程,共可分為五期:細胞增生 (cell proliferation)、
細胞接觸生長停滯 (cell comtact, growth arrest)、細胞擴張 (clonal expansion)、生長 停滯 (growth arrest)及脂質堆積 (lipid accumulation),並且由數種轉錄音子共同調 控,包括CCAAT enhancer binding protein (C/EBPs)、PPAR 等。整個分化過程中,
先由C/EBPβ 與 C/EBPδ 共同啟動 PPARγ,之後再由 C/EBPα 繼續維持 PPARγ 的表 現,顯示兩者之間具有協同的作用,共同誘發脂肪細胞特定基因表現,如:aP2
(三) PPAR 與代謝症候群
PPARs 可調控一系列參與糖類與脂質代謝的基因和胰島 beta 細胞的功能 (Tobin et al., 2006) 。多數研究指出,許多 TZDs (thiazolidinediones) 藥物為 PPARγ ligands,如 pioglitazone 及 rosiglitazone 等,可調節血糖、增加胰島素敏感度 (Nawrocki et al., 2006) 、抑制細胞激素所引起的發炎反應 (Chinetti et al., 2000) , 也可作用在粒線體膜上的蛋白質,調節粒線體代謝的功能 (Colca et al., 2004) ,改 善代謝症候群引起的高血脂、高血壓及高血糖。但2007 年有研究報告指出
rosiglitazone 會增加心臟病發作和心臟病有關之死亡風險 (Nissen et al., 2007) 。女 性患者使用rosiglitazone 或 pioglitazone 也有較高的骨折發生率。而最近也有研究 指出,長 期 服 用 pioglitazone 可能會增加罹患膀胱癌之風險。
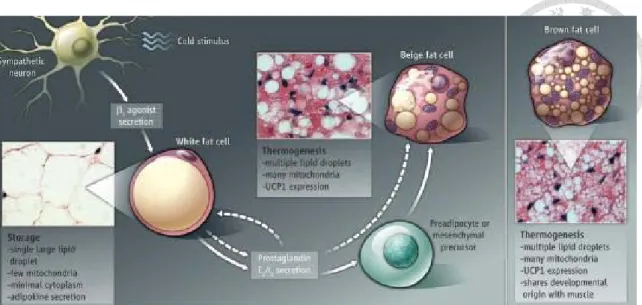
三 、 脂肪細胞褐化現象 (一) 脂肪組織
哺乳動物的脂肪組織可分成白色脂肪組織 (WAT) 及棕色脂肪組織 (BAT) 。 WAT 主要功能為儲存能量,將多餘的熱量將以三酸甘油酯的型式儲存,特色是細
胞內具有巨大的單一油滴,將細胞核及胞器擠至細胞邊緣。而BAT 主要功能是消
耗能量,以緊密摺疊的粒線體及表現UCP-1 (uncoupling protein-1) 著稱。近年研究,
發現棕色脂肪細胞與肌肉細胞皆源自myf-5 muscle-like 原始細胞 (progenitor) 系 (Seale et al., 2008) 。其中決定細胞往哪個方向發展,關鍵在於轉錄因子 PRDM16,
一旦開啟此基因則往BAT 分化;反之若移除 PRDM16,則分化為肌肉細胞 (Seale et al., 2011) 。有研究指出,人體內 BAT 的提高具有抵抗代謝症候群的治療潛力 (Nedergaard et al., 2010) 。雖然傳統認為人類在嬰兒時期之後,棕色脂肪的量會隨 著年齡的增加而逐漸減少。但最近研究顯示,以18F-FDG PET-CT (positron emission tomography and computed tomography,正子放射斷層掃瞄-電腦斷層攝影) 方式偵 測到成年人體內的棕色脂肪組織 (Nedergaard et al., 2007; Yeung et al., 2003)。而短 暫曝露在寒冷之下,也能觀察到UCP1 positive 的棕色脂肪,特別是在鎖骨及脖子
徵 (Cousin et al., 1992; Young et al., 1984) 。另外,有學者根據數量性狀基因座定 位 (quantitative trait loci,QTL mapping) ,提出 WAT 中的類棕色脂肪細胞和典型 棕色脂肪細胞,其生熱能力是受到不一樣的調節 (Coulter et al., 2003)。
圖
1-1 成年人類中可偵測到 UCP1 positive 的棕色脂肪
Fig 1-1 Adult humans seem to contain brown adipose tissue (BAT) primarily
behind the muscles of the lower neck and collarbone, and the spine of the chest and abdomen. (Farmer, 2009)(二) 去偶聯蛋白 (Uncoupling proteins,UCPs)
UCP 是存在於粒線體內膜上的氫離子通道蛋白,主要有三種型態,UCP1、
UCP2 及 UCP3。其中 UCP2 與 UCP3 主要調節 ROS (reactive oxygen species) ,而 UCP1 主要表現在棕色脂肪組織中,可進行去偶合作用,讓位於高能的氫離子梯度,
經由此氫離子通道回到粒線體內膜裡,過程中不產生ATP,而是把能量以熱的形
式釋出,被認為可避免低體溫症及肥胖的發生,又稱為產熱素 (thermogenin) 。文 獻指出,低溫的環境下或交感神經的刺激會導致棕色脂肪組織中的β-adrenergic receptor (β-AR) 活化,增加 UCP1 的表現來增加產熱作用 (Cousin et al., 1992;
Guerra et al., 1998) ,並可調控粒線體生合成、促進能量代謝 (Spiegelman et al., 2000) (圖 1-2、1-3) 。
圖
1-2 棕色脂肪中 UCP1 之產熱作用機制
Fig 1-2 Under cold conditions, placental mammals upregulate heat production in brown
adipose tissue and this regulatory thermogenesis is called nonshivering thermogenesis.(Saito et al., 2008)
(三) PPAR-γ co-activator-1 α (PGC1α)
PGC-1s 屬轉錄調控活化因子的家族,可誘導各種代謝基因的表現,家族成員 分別有PGC1-α、PGC1-β 以及 PRC (PGC-1-related co-activator) 。其中 PGC1-α 只 分布在高度耗氧的組織中,而且對粒線體的功能及能量代謝扮演重要的角色 (圖 1-3、1-4) 。PGC1-α 會受環境所調控,在低溫的刺激下,會使個體進行適應性生 熱作用 (adaptive thermogenesis) (Liang et al., 2006; Puigserver et al., 1998) 。此外,
PGC1-α 在肌肉中能經由運動誘導產生,進而影響粒線體生合成、提升粒線體呼吸 作用 (Handschin et al., 2008) 。當骨骼肌細胞中的鈣離子濃度上升,促使 CREB (cAMP response element binding protein) 受 cAMP 刺激而被活化,CREB 便與 PGC1-α 啟動子 (promoter) 上的 CRE 序列結合而促使 PGC1-α 的轉錄。也有研究 顯示,基因轉殖鼠只要稍微增加PGC1-α,就可抵抗因年齡所產生的肥胖和糖尿病,
圖
1-3 PPAR 受不同訊息分子所調控
Fig 1-3 Modulation of the actions of PPAR through phosphorylation by ERK, MAPK or
AMPK or through regulation of PGC-1 activity by various signaling events. (Schiff et al., 2012)圖
1-4 PGC1 之調控機制
Fig 1-4 (a) Effects of PGC1 expression on thermogenesis. (b)Cellular effects of ectopic
(四) 白色脂肪「褐化現象」 (browning)
早期研究發現,小鼠當處在寒冷環境或以 β-adrenergic pathways 刺激,可發現 棕色脂肪細胞出現在WAT 中 (Cousin et al., 1992; Guerra et al., 1998) 。這種細胞 (recruitable BAT,rBAT) ,外型呈現多房的特徵,顏色也較深,和典型棕色脂肪細 胞 (constitutive BAT,cBAT) 同樣含有大量細粒線體及高度表現 UCP1,並具有生 熱作用的潛力,又稱為 「beige cell」 (J. Wu et al., 2012) 、 「brite cell」 (brown in white) (Petrovic et al., 2010) 或 inducible BAT (Schulz et al., 2011) 。然而出現在 WAT 中這種類棕色細胞和典型棕色脂肪並非源自相同細胞系 (Seale et al., 2008;
Seale et al., 2011)。不管是棕色脂肪還是 WAT 中的褐化細胞,都以 Prdm16 基因轉 殖小鼠證實這兩種棕色脂肪都具有相當抗肥胖及抗糖尿病的能力 (Seale et al., 2011)。而目前也有研究指出,給予小鼠合成的 PPARγ agonists,可刺激 WAT 褐化 現象的產生,並抑制白色脂肪相關基因的表現 (Vernochet et al., 2009) 。類似的研 究指出,一種第二型糖尿病藥物噻唑烷二酮類(Thiazolidinediones, TZDs) ,除 了能增加對胰島素的敏感性,也能增強PRDM16 蛋白質的效果,使白色脂肪棕色 化 (Ohno et al., 2012) ,但確切機制尚不清楚。另外也有研究指出,大量存在於白 色脂肪組織中的視黃醛脫氫酶 (retinaldehyde dehydrogenase1, Aldh1a1) 受抑制時,
也會促使褐化現象的發生,將能量以熱的形式散失,使缺乏Aldh1a1 小鼠能耐受 低溫 (Kiefer et al., 2012) 。最近研究發現,經由運動誘導產生的一種天然荷爾蒙,
鳶尾素(irisin),促進胰島素生成,也能刺激皮下脂肪細胞中 UCP1 的表現及往棕 色脂肪細胞發展。只要稍微增加血液中的irisin,能使小鼠在不改變食物攝取量及 活動量下增加能量的消耗 (Bostrom et al., 2012; J. Wu et al., 2012) 。白色脂肪褐化 現象與PPAR 有關之相關文獻整理於表 1。
(五) 基因及粒線體指標的篩選 (1) 棕色脂肪相關基因
Cidea (Cell death-inducing DNA fragmentation factor α-like effector A)
減少脂肪囤積使體型變瘦,顯示Cidea 在生熱作用中扮演負調控的角色 (Zhou et al., 2003) 。雖然 Cidea 和 UCP1 在寒冷的刺激下有不同反應,但由於兩者皆位於粒線 體內膜上,也會受RIP140 (hermogenic repressors) 所調控,顯示 Cidea 和 UCP1 是 在相同的訊息傳遞 (cAMP-PKA pathway) 上是受到不同調控。而 Cidea 也會直接 或間接受到PPARγ 的調控,當給予 PPARγ agonists 刺激,在小鼠初代脂肪細胞或 分化成熟的 3T3-L1 脂肪細胞皆可觀察到 Cidea mRNA 表現增加 (Puri et al., 2008) 。
Prdm16 (PRD1-BF-1-RIZ1 Homologous Domain Containing Protein 16)
Prdm16 為轉錄調控因子,可調控棕色脂肪的分化,一般認為只會表現在 BAT 當中,但最近研究指出prdm16 也會高度表現在皮下白色脂肪組織中。aP2-PRDM16 基因轉殖小鼠,可以調控PGC1α、Cidea 及 UCP1 等基因表現增加,進而使 WAT 出現褐化的現象 (Seale et al., 2011) 。PRDM16 也能與許多轉錄因子 (如 PPARα、
PPARγ) 作用,增加基因的轉錄活性。
Tfam 與 NrF1
均為粒線體生合成重要的轉錄調節因子 (Z. Wu et al., 1999) 。Tfam (mitochondrial transcription factor A) 和 DNA 上的啟動子結合,進而調節粒線體基因的轉錄。而 NrF1 (nuclear respiratory factors) 則調控粒線體呼吸或粒線體 DNA 轉錄、複製等。
Cytochrome oxidase (Cox)
Cox 於粒線體氧化機轉中扮演很重要的角色,在粒線體電子傳遞鏈中,將電子 從cytochrome c 傳送給氧分子,最終形成水分子 (Boyer, 1997) 。而 Cox7a1 (cytochrome c oxidase polypeptide 7A1) 及 Cox8b (cytochrome c oxidase, subunit VIIIb) 均與粒線體電子傳遞鏈有關。
(2) 白色脂肪相關基因
Retn (Resistin)
是一種由脂肪細胞分泌的細胞激素,當脂肪細胞因油脂累積而增大,會促進 resistin 的合成及分泌,而造成胰島素抗性。Resistin 的合成與分泌會受到 PPARγ agonists 的抑制 (Patel et al., 2003) 。
Chemerin
是新發現的脂肪因子,由脂肪細胞分泌,可調節成熟脂肪細胞的脂肪生合成
及代謝相關作用,也會受到PPARγ 的調控 (Muruganandan et al., 2011) 。
(3) 轉錄因子相關基因
C/EBPα
屬於 CCAAT/Enhancer Binding Protein (C/EBP) 家族,在脂肪細胞分化過程中 扮演重要的角色,會與 PPARγ 共同調控脂質生成。有研究指出,PPARγ agonists troglitazone 抑制白色脂肪相關基因表現需要 C/EBPα 的存在 (Vernochet et al., 2009) 。
C/EBPβ (CCAAT/Enhancer Binding Protein β )
在 BAT 中表現量高於 WAT,能調節 UCP1 的轉錄。在寒冷刺激下,也能增加 C/EBPβ mRNA 在 BAT 中的表現 (Manchado et al., 1994) 。在 cAMP 刺激下,過度 表現的C/EBPβ 會和 CRE (cAMP responsive element) 結合,增加白色及棕色脂肪細 胞中PGC1α 及 UCP1 的基因表現 (Karamanlidis et al., 2007) 。先前研究指出,給 予正常鼠及ob/ob 鼠 non-TZD PPARγ agonist 後,對於 β-adrenergic 的刺激,都有相 當大的生熱潛力 (thermogensis) (Sell et al., 2004) ,顯示這兩種機制彼此有關。
(4) 檸檬酸合成酶 (Citrate Synthase,CS)
存在於真核細胞的線粒體中,是催化三羧酸循環 (TCA cycle) 第一步的限速 酶 (Wiegand et al., 1986) 。CS 由核 DNA 編碼而成,而非線粒體 DNA,在細胞質 中合成後,再運送至粒線體基質 (matix) 中,故常作為完整粒線體的定量指標 (Holloszy et al., 1970; Hood et al., 1989) 。檸檬酸合酶的組氨酸殘基作為親核試劑,
使乙醯輔酶A (acetyl CoA) 中的甲基去質子化而形成碳負離子,後者親核攻擊草醯 乙酸 (oxaloacetate,OAA) 的羰基碳,縮合生成檸檬醯輔酶 A (citroyl-CoA) 中間 體,將高能硫酯鍵水解後即得檸檬酸 (Citrate)。
圖
1-5 白色脂肪棕色化
Fig 1-5 White to Beige fat cell conversion (Ishibashi et al., 2010) .
四、 山苦瓜 (一) 苦瓜之背景
苦瓜,學名 Momordica charantia L.,英文名 bitter gourd 或 bitter melon,又名 涼瓜、錦荔枝、癩瓜、癩葡萄,屬於葫蘆科 (Cucurbitaceae) 、苦瓜屬 (Momordica) , 一年生蔓性草本植物,原產於亞洲熱帶地區,包括印度、東南亞、中國大陸與台 灣,台灣南部一年四季皆可生產。果實為漿果,表面有瘤狀突起,呈綠色、淺綠 色或是白色,成熟後會逐漸呈橘黃色,種子呈紅色,其中果實與葉子部分常作為
日常蔬菜用。【本草綱目】中,記載苦瓜有清暑解熱、解毒、清心明目與補腎潤脾
等功效,現今亦有抗腫瘤、抗病毒與抗發炎之功效受重視,而許多研究則著重於 苦瓜降血糖與降血脂等代謝症候群指標異常之治療潛力。
(二) 苦瓜與肥胖相關研究
Chen 等人給予高油飲食誘發肥胖的小鼠補充苦瓜仔油 (BMSO) ,能增加 WAT 中UCP1 的表現 (Chen et al., 2012) 。而本實驗室先前研究中,也觀察到給予 C57BL/6J 公鼠山苦瓜全果凍乾粉末,能增加粒線體生合成相關基因及白色脂肪組 織中UCP1 mRNA 的表現,顯示具有棕色脂肪的特性 (Lu et al., 2013);除此之外,
餵食苦瓜組的PGC-1α 基因表現量,在腓腸肌及棕色脂肪組織中也顯著高於對照組。
而根據先前研究,給予飲食誘導肥胖之大鼠苦瓜果汁凍乾物,能增加肌肉中PGC-1α 的表現 (Chan et al., 2005) 。另一方面,給予 lean 或 ob/ob 小鼠 PPARγ 的 agonist,
不但能顯著降低血糖,也能增加鼠蹊部皮下脂肪處之中UCP1 的表現量 (Sell et al., 2004) 。本實驗室也有研究證實,苦瓜乙酸乙酯萃取物為 PPARα 與 PPARγ 的雙效 活化劑 (Chao et al., 2003) (萃取流程如圖 1-6) 。以上結果顯示,苦瓜中內含活化 PPARγ 的成分,推測可降血糖、血脂,並進而促使白色脂肪組織棕色化。相關文 獻整理於表2。
(三) 苦瓜中 PPAR 活性成分 (1) CLN
共軛亞麻油酸 (conjugated linoleic acid, CLA) 在多種動物實驗中證實有降體 脂的功效,作用機制包含改變脂質代謝、抑制脂肪細胞分化及促進 (前) 脂肪細胞 凋亡等 (Domeneghini et al., 2006) 。但市售 CLA 商品係將亞麻油酸鹼處理而得,
只要含
c9,t11 及 t10,c12 兩種異構物。許多實驗 (包含人體試驗) 發現 t10,c12-CLA
會促進胰島素阻抗,血液中也有發炎介質及過氧化指標有上升的情形 (Riserus et al., 2002) ,有安全上的顧慮。然而共軛三烯-次亞麻油酸 (CLN) 則多存在於植物 種籽中,而本實驗室先前經由分離純化,鑑定出苦瓜中此種特殊脂肪酸-c9,t11,t13-CLN (α-eleostearic acid) ,為可活化 PPARs 之化合物。
(2) 植醇 (Phytol)
文獻顯示,小鼠餵食富含 phytol 的飲食,能增加體內的 phytenic、phytanic 及 pristanic acid 等代謝物,並活化 PPARα 或其他轉錄因子,影響脂肪酸的代謝機制 (Gloerich et al., 2005) 。phytol 的代謝產物 phytanic acid 能刺激棕色脂肪細胞的分 化,並增加棕色脂肪細胞中UCP1 及 Cytochrome c oxidase subunit II (CO II ) 的基 因表現 (Schluter et al., 2002) 。
(3) 葉黃素 (lutein)
屬於類胡蘿蔔素 (carotenoids) ,能與 PPARγ 作用。先前文獻指出,lutein 能 在不影響棕色脂肪細胞增生下,提高UCP1 蛋白質的表現 (Serra et al., 1999) ,也 能增加白色脂肪細胞PPARγ 及 PGC1α mRNA 基因表現 (Garcia-Rojas et al., 2010) 。
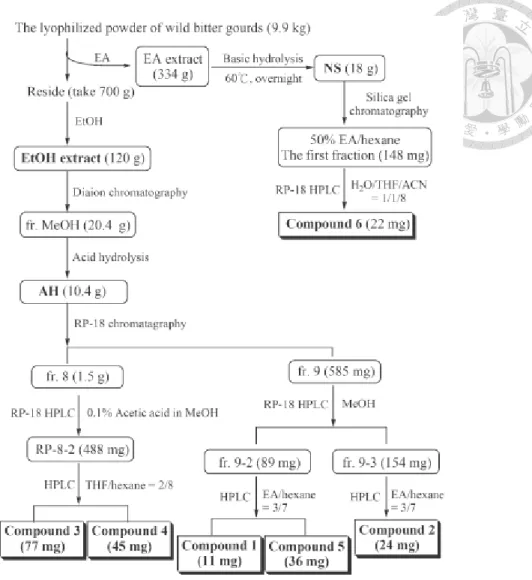
圖 1-6 山苦瓜之各種粗萃物之製備及區分過程
Fig 1-6 The scheme of separation and puri fi cation of Momordica charantia extracts.
(Hsu et al., 2011)
圖 1-7 共軛脂肪酸 CLA 和 CLN 之化學結構
Fig 1-7 Structure of conjugated fatty acid CLA and CLN.
表 1-1 PPAR agonist 刺激白色脂肪褐化之文獻整理
Table 1-1 Literature reviews of browning occur in white adipose tissue or adipocytes induced by PPAR agonist.
模式 樣品 實驗方法 結果 (文獻)
male C57Bl/6 mice
SV cells
Recombinant FGF21 protein
小鼠給予 FGF21 或 saline 處理 72 小時
分化第六天,以 50 nM FGF21 處理 SV 細胞 兩天,於第八天收細胞
in vivo
inguinal WAT (IWAT):UCP1, DIO2, PGC1α, Cidea, Cox7a↑
epididymal WAT (EWAT):UCP1,DIO2↑
perirenal adipose tissue (PRWAT):UCP1, DIO2, PGC1α, Cox7a↑
primary WAT and BAT adipocytes
UCP1, Cidea↑ (Fisher et al., 2012)
C57BL/6J lean and obese ob/ob female mice
COOH
(non-TZD PPAR agonist)
將 PPARγ agonist (COOH) 混於飼料中,每 天給予小鼠30 mg/kg 體重的刺激物,連續 14 天
UCP1 protein expression
BAT:ob/ob group↑
IWAT:lean, ob/ob group↑
(Sell et al., 2004)
primary cells of brown and white adipocytes
Rosiglitazone (PPARγ agonist)
初代脂肪細胞以含有 1 μM rosiglitazone 培養 基培養5 days
白色及棕色初代脂肪細胞
Brown-selected genes (UCP1, PGC1α, Cidea, Elovl3)↑
(Petrovic et al., 2010)
Male C57BL/6J mice
Primary adipocytes
Rosiglitazone (PPARγ agonist)
小鼠於 6-8 週齡,以腹腔注射方式給予 10 mg/kg 體重的 rosiglitazone 連續 10 天
初代脂肪細胞於分化期間以含有 1 μM rosiglitazone 培養基培養
in vivo
BAT and visceral WAT:UCP1↑
subcutaneous WAT:UCP1, Cidea ↑
primary WAT adipocytes
IWAT:UCP1, Cidea, PGC1α, prdm16↑
EWAT:UCP1, Cidea, PGC1α↑
(Ohno et al., 2012)
3T3-L1 preadipocytes
mouse embryo fibroblasts (MEFs)
male FVB mice
Troglitazone (PPARγ agonist)
3T3-L1 在分化成熟前兩天給予 troglitazone 刺激兩天
MEFs 於整個分化期間給予 troglitazone 刺激
小鼠以灌食方式給予 10 mg/kg 體重的 PPARγ agonist (COOH)及 vehicle 連續 14 天
3T3-L1, MEFs and in vivo
White genes (chemerin, resistin)↓
Brown-selected genes (UCP1, PGC1α)↑
(Vernochet et al., 2009)
表 1-2 苦瓜增加 UCP1 表現之文獻整理
Table 1-2 Literature reviews of increasing UCP1 in adipose tissue or muscle by feeding Momordica charantia
模式 樣品 實驗方法 結果 (文獻)
Male
Sprague-Dawley rats
經冷凍乾燥 之苦瓜汁
大鼠經高脂飲食三週後,苦瓜樣 品添加於飼料中,飼養6 週
HF
HF + 1 % BM
HF + 1.25 % BM
mRNA expression in BAT and gastrocnemius muscle (G)
HF+1 % BM:UCP1, UCP3, PGC1α↑
HF+1.25 % BM:UCP1, UCP3, PGC1α↑
protein contain in BAT and G
HF+1 % BM:UCP1 (BAT), UCP3 (G)↑
HF+1.25 % BM:UCP1 (BAT), UCP3 (G)↑ (Chan et al., 2005)
male C57BL/ 6J mice
苦瓜籽油 (BMSO)
小鼠給予餵食添加不同比例苦瓜 籽油的高脂飼料,飼養10 週
HS:30 % SBO (soybean oil)
LBM:25 % SBO + 5 % BMSO
MBM:20 % SBO + 10% BMSO
HBM:15 % SBO + 15 % BMSO
mRNA expression in white adipose tissue (compare to HS )
LBM:PGC1α↑
MBM:PGC1α, UCP2↑
HBM:PGC1α, UCP1, UCP2↑
(Chen et al., 2012)
male C57BL/ 6J mice
苦瓜全果凍 乾粉末
小鼠給予餵食經調整的 AIN-93G 飼料,分成Basal 組及添加 5 %苦 瓜凍乾粉末(BGP)組,餵養 22 週
mRNA expression
BAT:PGC1α, NrF1↑
epididymal WAT:UCP1, PGC1α, NrF1↑
gastrocnemius muscle:PGC1α, tfam↑ (Lu et al., 2013)
表 1-3 自苦瓜分離出具活化 PPARs 之化合物整理
Table 1-3 PPARs activity of compounds from Momordica charantia.
compounds Structure PPARs activity
Lutein C
40H
56O
2PPARα + PPARγ +
Loliolide C
10H
16O
3PPARα + PPARγ +
Phytol C
20H
40O
PPARα + PPARγ +
Compound 1
C
30H
44O
3PPARα + PPARγ +
Compound 2 C
30H
46O
3PPARα + PPARγ +
Compound 3 C
30H
44O
2PPARα + PPARγ -
Compound 4 C
32H
50O
3PPARα + PPARγ -
Compound 5 C
29H
42O
PPARα + PPARγ +
Compound 6
PPARα +第三節 實驗假說與架構 一. 實驗假說
先前研究顯示,小鼠餵食山苦瓜,能使白色脂肪細胞中UCP1 及粒線體相關 基因的表現量增加。另外,本實驗室也發現苦瓜中乙酸乙酯萃物能有效活化PPARγ,
且從中找出活化成分,推測其能更進一步刺激下游PGC-1α 基因表現量增加。所以 本實驗假說為:山苦瓜中具有刺激
3T3-L1 脂肪細胞褐化及粒線體相關基因增生的
成份。以此脂肪細胞做為試驗模式,給予分化過程中的脂肪細胞不同山苦瓜萃取 樣品,觀察樣品是否影響脂肪細胞UCP1 以及粒線體相關基因 mRNA 表現與酵素 活性,初探山苦瓜中可能之活性成分。二. 實驗架構
以3T3-L1 脂肪細胞模式,確立一個最佳評估樣品對於脂肪細胞棕色化的平台 以PPARγ agonists 刺激成熟的脂肪細胞,比較兩種不同分化法及三種處理時間 點對於脂肪細胞褐化及粒線體增生之相關基因表現變化。確立模式後,評估不同 山苦瓜萃物/區分物對於增加脂肪細胞褐化及粒線體增生之基因相關表現的潛力。
第二章 山苦瓜萃物對於脂肪細胞褐化相關基因表現之影響
第一節 前言當能量攝取量大於能量消耗量,多餘的熱量以三酸甘油酯的型式儲存於脂肪 組織中。脂肪可分成白色脂肪 (WAT) 和棕色脂肪組織 (BAT) ,WAT 主要功能為 儲存能量;棕色脂肪則以熱的形式消耗能量,維持體溫。文獻指出,低溫環境下 或交感神經的刺激會導致棕色脂肪組織中的β-adrenergic receptor (β-AR) 活化,而 這樣的調控是由Peroxisome proliferator-activated receptor γ (PPARγ) coactivator-1α (PGC-1α) 所刺激,可調控粒線體生合成、能量代謝 (Spiegelman et al., 2000) 、 UCP1 的表現來增加產熱作用 (Bostrom et al., 2012) 。另一方面,早期研究發現,
在低溫或交感神經的刺激下,會導致白色脂肪組織出現較深色的類棕色脂肪 (Cousin et al., 1992; Guerra et al., 1998) 。此種細胞,和棕色脂肪細胞同樣含有大量
粒線體及高度表現UCP1,並具生熱作用的潛力。目前也有研究指出,給予小鼠合
成的PPARγ agonists,可刺激 WAT 中出現此種類棕色脂肪細胞,並抑制白色脂肪 過多時的相關基因表現 (Vernochet et al., 2009) 。同時投予 β-adrenergic receptor (β-AR) 致效劑,則可使小鼠代謝速率增加、脂肪組織變小 (Sell et al., 2004) 。 本實驗室先前的研究證實,苦瓜乙酸乙酯萃取物為 PPARα 與 PPARγ 的雙效活 化劑 (Chao & Huang, 2003) 。經由分離純化,鑑定出 9c,11t,13t-CLN 為可活化 PPARs 之化合物 (Chuang et al., 2006) 。而山苦瓜乙酸乙酯萃物之不皂化物亦具有 活化 PPAR 之能力 (徐瑨,2011) 。先前研究也觀察到 C57BL/6J 公鼠給予餵食山 苦瓜全果凍乾粉末,能增加粒線體合成基因及白色脂肪組織中UCP1 mRNA 的表 現 (Lu et al., 2013) 。此外,Chao 等人給予高油飲食誘發肥胖的小鼠補充苦瓜仔油 (BMSO) ,能增加 WAT 中 UCP1 的表現以及促進 PKA 的活化 (P. H. Chen et al., 2012) 。以苦瓜汁凍乾物餵食 Sprague-Dawley 大鼠,發現其血清中正腎上腺濃度 較高 (Chan et al., 2005) 。這些文獻暗示,苦瓜中有會影響 β-AR 之訊息傳遞的成 分,配合PPARγ 的活化,進而導致白色脂肪褐化。本實驗以 PPARγ agonists 刺激 分化中之3T3-L1 脂肪細胞株為正對照組,觀察山苦瓜萃物是否能誘發出白色脂肪
第二節 材料與方法
一、白色脂肪細胞褐化實驗模式建立與最佳化
二、山苦瓜萃物促進白色脂肪細胞褐化
三、細胞株
3T3-L1 細胞株 (ATCC CL-173) 組織來源:embryo, Mus musculus
購自食品工業發展研究所 (BCRC 60159)
四、培養基與藥品試劑 (一) 細胞培養與分化 1. 培養基
藥品
Dulbecco’s modified Eagle’s medium (DMEM, Gibco 11995) Bovine serum (BS, Gibco 16170)
L-glutamine (Gibco 25030)
2. 分化培養基Ⅰ
藥品
Stock conc. Working conc.
Dexamethasone (DEX, Sigma 4902) 0.25 mM 0.25 μM 3-Isobytyl-methyl-xanthine (IMX, Sigma I5879) 0.5 M 0.5 mM Insulin (Sigma I9278) 10 mg/mL 10 μg/mL Biotin (Sigma B4501) 400 mg/mL 100 ng/mL 分化方法一:10 % BS/DMEM (4 mM Gln)
分化方法二:10 % FBS/DMEM (4 mM Gln) 於使用前配置。
3. 分化培養基Ⅱ
藥品
Stock conc. Working conc.
Insulin (Sigma I9278) 10 mg/mL 10 μg/mL Biotin (Sigma B4501) 400 mg/mL 100 ng/mL
4. 0.25 % Trypsin-EDTA (GIBCO 25200) 5. 0.4 % Trypan blue solution (Sigma T8154)
(二) 檸檬酸合成酶活性測定 (Citrate Synthase activity assay)
藥品
Stock conc. Working conc.
Tris [Tris-(hydroxymethyl)aminomethan] 1 M 0.1 M
Triton X-100 10 % 0.25 %
Oxalacetic acid (Sigma O4126) 10 mM 0.5 mM
DTNB 2 mM 0.1 mM
Acetyl CoA, lithium salt (Sigma A2181) 2 mM 0.31 mM Tris 以 HCl 調製 pH=8.0,為 Tris-HCl buffer。
Oxalacetic acid 及 DTNB 於使用前配置。
(三) quantitative Real-time PCR (qRT-PCR) 法分析基因表現 1. TRIZOL® Reagent (Invitrogen)
2. Choloform (Merck) 3. Isopropanol (Merck) 4. DEPC-H2O
5. 反轉錄試劑套組 High-capacity cDNA Reverse Transcription Kit (Applied Biosystems , P/N:4368814 )
6. 基因表現分析套組 Probe/primer assay ID:
Ppargc1a (peroxisome proliferator activated receptor, gamma, coactivator 1
α) (Mm01208835_ml)Ucp1 (uncoupling protein 1) (Mm01244861_ml)
Cidea (cell death-inducing DNA fragmentation factor, alpha subunit-like effector
A) (Mm00432554_m1)Prdm16 (PR domain containing 16) (Mm00712556_m1) Nrf1 (nuclear respiratory factor 1) (Mm01135606_ml)
Tfam (transcription factor A, mitochondrial) (Mm447485_ml)
Cox7a1 (cytochrome c oxidase subunit VIIa 1) (Mm00438296_m1)
Cox8b (cytochrome c oxidase subunit VIIIb) (Mm00432648_m1)
Cebpα (CCAAT/enhancer binding protein, alpha) (Mm00514283_s1)
Cebpβ (CCAAT/enhancer binding protein, beta) (Mm00843434_s1) Retn (resistin) (Mm00445641_m1)
Rarras (retinoic acid receptor responder (tazarotene induced) 2)
(Mm00503579_m1)Ppara (peroxisome proliferator activated receptors α) (Mm00440939_ml) Pparr (peroxisome proliferator activated receptors γ) (Mm00440940_ml) Ppard (peroxisome proliferator activated receptors δ) (Mm00803184_ml)
Actb (actin, beta) (Mm00607939_s1)Gadph (glyceraldehydes-3-phosphate dehydrogenase) (Mm99999915_gl)
7. 2× Taqman® Univeral PCR Master Mix (Applied Biosystems, K00861)(四) 處理藥品
藥品
Stock conc. Working conc.
Troglitazone (Enzo 3-H2315r) 5 mM 5 μM Dimethylsulfoxide (Sigma D2650)
Forskolin (Sigma F6886) 10 mM 10 μM 配置於10 % BS/DMEM (分化方法一) 或 10 % FBS/DMEM (分化方法二) 。
五、儀器設備
1. 二氧化碳培養箱 (REVCO RCO, Thermo Forma 3100) 2. 無菌操作台 (ESCO AC2-4S2)
3. 倒立式顯微鏡 (Nikon DIAPHOT) 4. 細胞計數器 (Hausser Science) 5. 微盤分析儀 (BioTek EL311) 6. 離心機 (KUBOTA 6500)
7. 基因定量分析儀 (ABI PRISM 7000 Sequence Detection) 8. 冷凍乾燥機 (SFD-25)
六、山苦瓜萃物之製備
(一) 山苦瓜乙酸乙酯萃物暨其區分物
本試驗使用花蓮縣吉安鄉改良場所提供之花蓮四號山苦瓜,經清洗、去蒂切 片、冷凍乾燥後,經研磨後取得山苦瓜全果凍乾粉末置-20 ℃儲存。山苦瓜凍乾粉 末以20 倍乙酸乙酯攪拌進行萃取隔夜,以 Whatman No.2 濾紙進行抽氣過濾,收 得乙酸乙酯苦瓜萃取液。將萃取液經減壓濃縮後即製成山苦瓜全果凍乾粉末乙酸 乙酯萃物 (ethyl acetate extract ,EAE) ,萃率約 5 %。取山苦瓜乙酸乙酯萃物,
溶於五倍體積之四氫呋喃 (THF) 中,再加入同體積之 40 % 氫氧化鈉溶液,以 60
℃ 加熱攪拌 16-20 小時。反應液先經減壓濃縮去除有機溶劑 THF 後,改加等量 之正己烷,與水層進行3-5 次萃取。收集上層之正己烷,以水洗至中性後,經減壓 濃縮後收得不皂化物 (non-saponification,NS) ,萃率為約 5 %。而水層部份加入 鹽酸,使其酸化至pH=2 後,加入等量乙酸乙酯萃取兩次,取上層乙酸乙酯層,以 水洗至中性後,減壓濃縮即可得皂化物 (saponification,S) ,萃率為 67 %。樣品 stock 皆配置於絕對酒精,EAE 及 S 濃度為 50 mg/mL,NS 濃度為 25 mg/mL,分 裝後存放於 -20℃。
(二) 山苦瓜水萃物
將100 g 山苦瓜全果凍乾粉末添加 20 倍 (w/v) 去離子水,於 60℃ 攪拌萃取 隔夜,以5000 g 離心 5 分鐘後取上清液,以 Whatman No.2 濾紙過濾,所得濾液為 山苦瓜全果水萃物 (water-soluble extract;WE) 。
(三) 經 5 % 黑豆漿培養之納豆菌處理山苦瓜水萃物
納豆菌 (Bacillus subtulis natto) 在 3 ml LB 於 37 ℃,125 rpm 活菌 12 小時後,
取適量菌液培養於5 % 黑豆漿,37 ℃,125 rpm,12 小時,離心去除上清將納豆 菌移至山苦瓜果汁中培養24 小時。以 5000 g 離心 5 分鐘後取上清液,以 Whatman No.2 濾紙過濾,所得濾液為納豆菌作用之山苦瓜全果水萃物 (WEn) 。
七、實驗方法 (一) 細胞培養
3T3-L1 前脂肪細胞株以 10 % BS/DMEM 培養於 37 ℃、5 % CO2中。當細胞 長至八分滿,移除培養基,以 PBS 清洗細胞兩次,加入 0.25 % trypsin-EDTA 均
下。取少許懸浮細胞以trypan blue solution 染色,以細胞計數器於倒立式顯微鏡計 算細胞數,將2.5×105 細胞數種入新 75 cm2 培養瓶進行繼代培養,每 3 日繼代培 養一次。
(二) 細胞分化
當脂肪細胞於 75 cm2培養瓶生長至八分滿,以0.25 % trypsin-EDTA 將細胞脫 附後,以10 % BS/DMEM 將細胞洗下,經細胞計數後,以 6×104細胞數種入12 孔 平底培養盤,培養於37 ℃、5 % CO2中,3 日後可達全滿 (confluence) ,更換培 養基,繼續培養兩日 (post-confluence) 後,改以分化培養基Ⅰ (DMⅠ) 培養 3 日。
3 日後更換為分化培養基Ⅱ (DMⅡ) ,每 2 天更換一次 DMⅡ。分化法一:培養 8 天後換回不含胰島素之basal medium,繼續培養 4 天,即分化完成;分化法二:培 養4 天後換回不含胰島素之 basal medium,繼續培養 4 天,即分化完成 (如圖示)。
(三) 蛋白質測定 1. 原理
BCA 法蛋白質使 reagent 中銅離子由二價轉為一價,再以 BCA 與一價銅離子 結核並呈紫色,在波長540 nm 具吸光。本實驗法對於陰離子、非離子性及二價離 子的清潔劑 (anionic, non-ionic, zwitterionic) 、guanidine-HCl 和尿素較具容忍性,
且較不受干擾。
2. 實驗步驟
將溶於1 N NaOH 之細胞溶液,取 25 μL 於 96 孔盤中,並將 kit 中的 A 液與 B
(四) 樣品處理
脂肪細胞分化完成前二日或前四日,加入含 5 μM troglitazone 或苦瓜樣品的 10 % BS/DMEM (或 10 % FBS/DMEM) 培養 2 天或 4 天,並以加入相同量的 dimethylsulfoxide (DMSO) 處理者為空白組。進行基因表現分析或粒線體活性測試 前,以10 μM forskolin 刺激 8 小時。
(五) 檸檬酸合成酶活性測定 (Citrate Synthase activity assay) 1. 樣品製備
每 1-2×106 cells/mL 細胞溶液取 110 μL 於液態氮結凍,供日後分析。
2. Reaction mixture
將 750 μL ddH2O、25 μL triton X-100 (0.25 %) 、50 μL DTNB (0.1 mM) 、50 μL oxalacetic acid (0.5 mM) 、25 μL acetyl CoA (0.31 mM) 混合均勻後,於 30℃ 水浴 5-10 分鐘後,加入解凍後之 100 μL 細胞液迅速混合均勻,取 1000 μL 於比色管中,
以未加入acetyl CoA 的溶液作為 blank,測定三分鐘內每 15 秒在波長 412 nm 的吸 光值,以蛋白質濃度校正後,測得檸檬酸合成酶活性。
(六) 基因表現 1. 總 RNA 抽取
樣品處理後之脂肪細胞吸除培養基,以PBS 潤洗細胞一次。加入 1 mL TRIZOL® reagent,吸放數次,移至 1.5 mL 微量離心管。12000 g,4℃ 離心 10 分 鐘,取上清液至新1.5 mL 微量離心管,靜置 5 分鐘。加入 0.2 mL choloform,震盪 15 秒後靜置 2-3 分鐘。12000 g,4℃ 離心 20 分鐘,取上清液至新的 1.5 mL 微量 離心管,加入0.5 mL isopropanol,上下倒置數次使之均勻,室溫 10 分鐘。12000 g,
4℃ 離心 10 分鐘,去除上清液,加入 75 % 酒精清洗一次 (7500 g,4℃ 離心 5 分鐘) 。室溫晾乾後以 10 μL DEPC-H2O 回溶,即得 RNA 溶液。以 Nano Drop 測 RNA 濃度,當 A260/A280比值接近2 時即為純化 RNA 樣品。
2. RNA 反轉錄成 cDNA
使用反轉錄試劑套組High-capacity cDNA Reverse Transcription Kit (Applied Biosystems , P/N:4368814 ) ,取 10 μL 0.5 μg/mL RNA 溶液加入 10 μL 2× Reverse Transcription Master Mix (2 μL 10× RT buffer, 0.8 μL 25× dNTP mix, 2 μL 10× RT
Random Primers, 1 μL 10× MulitiscibeTM RTase, 4.2 μL ddH2O) 進行反轉錄,轉錄條 件為25 ℃ 10 分鐘、37 ℃ 120 分鐘、85 ℃ 5 分鐘,最後冷卻至 4 ℃。轉錄完 成之cDNA 儲存於-20℃ 冰箱,待日後基因表現分析。
3. Real-Time PCR 分析脂肪細胞基因表現
採用市售2× Taqman® Univeral PCR Master Mix (Applied Biosystems, K00861) 分析細胞基因表現,以β-actin 作為 internal control (Mm00607939_s1) 。將 cDNA 稀釋成10 ng/μL,取 10 μL cDNA 加入 12.5 μL 2× TaqMan Master Mix, 1.25 μL 20×
probe/primers AssayMix 及 1.25 μL ddH2O,總體積為 25 μL。注入 ABI PRISMR Optical Strip 後蓋上蓋 (MicroAmp Optical Cap) ,離心去除氣泡後即可進基因定量 分析,條件為50℃ 10 分鐘、95℃ 10 分鐘、(95℃ 15 秒鐘、60℃ 1 分鐘) 共 40 個循環,所得之Ct 值進行換算以分析基因表現量。
算式:ΔCt = 目標基因 Ct 值-internal control (β-actin) Ct 值 ΔΔCt = 實驗組 ΔCt-空白組 ΔCt
Fold of change = 2 (-ΔΔCt)
(七) 數據整理及統計分析
重複三次以上獨立實驗,而每次獨立實驗皆有2-3 重複,數值以平均值 ± 標 準差 (mean ± S.D.) 表示。利用 excel 進行統計分析,以 Student's t test 檢定各組與 vehicle 組間差異之顯著性,當 p < 0.05 視為兩組在統計上具有顯著差異,以 * 做 標示。
第三節 實驗結果
一、白色脂肪細胞褐化實驗模式建立與最佳化
為建立最合適之細胞模式,本實驗比較兩種已建立之脂肪細胞培養方式,差 異在於分化培養基的使用,以及分化時間長短。
分化法一 (protocol 1):分化培養基配置於 10 % BS/DMEM,分化時間約十五天 分化法二 (protocol 2):分化培養基配置於 10 % FBS/DMEM,分化時間約十一天
(一) 分化成熟前 2 天,troglitazone 處理 2 天
↑/↓:5 μM troglitazone treatment;☆:harvest
(1) 分化法一:5 μM troglitazone 處理下,棕色脂肪相關基因 PGC1α、UCP1 與 Cidea 皆有顯著增加 (p<0.05 ~ p<0.005) ,其中 UCP1 顯著高於空白處理組,約 1.7 倍;粒線體生合成作用相關基因Tfam 有增加的趨勢,而 NrF1 和空白組並無顯著 差異 (p>0.05) ;白色脂肪相關基因表現上,Retn 顯著低於空白組 (0.6 倍) (p<0.005),
而Chemerin 卻有增加 (1.3 倍) 的現象 (p<0.05) 。PPARγ 雖受抑制,但和空白組 並無顯著差異 (p>0.05) (圖 2-1 (A))。
(2) 分化法二:5 μM troglitazone 處理下,棕色脂肪 PGC1α、UCP1、Cidea 及粒 線體生合作用相關基因NrF1、Tfam mRNA 表現雖有增加的現象,但和空白處理組 並無顯著差異 (p>0.05) 。白色脂肪相關基因表現上,Retn 顯著低於空白組 (0.4 倍) (p<0.005) ,而 Chemerin 顯著增加 (1.2 倍) (p<0.005) ,PPARγ 則顯著受到抑 制 (0.6 倍) (p<0.005) (圖 2-1 (B)) 。
(二) 分化成熟前 2 天,troglitazone 處理 4 天
↑/↓:5 μM troglitazone treatment;☆:harvest
(1) 分化法一:5 μM troglitazone 處理下,棕色脂肪相關基因 UCP1 及 Cidea 皆顯 著增加 (分別為 3.3 及 4.4 倍) (p<0.05 ~ p<0.005) ,而 PGC1α 基因表現雖有增加的 趨勢,但和空白處理組並無顯著差異 (p=0.06) 。粒線體生合作用相關基因 Tfam 與NrF1 也顯著高於空白組 (分別為 1.4 及 1.2 倍) (p<0.05) 。白色脂肪相關基因 Chemerin 有顯著增加 (2.4 倍) (p<0.05) ,而 PPARγ 和空白組並無差異 (p>0.05) (圖 2-2 (A)) 。
(2) 分化法二:5 μM troglitazone 處理下,棕色脂肪相關基因 PGC1α (p<0.01) 及 cidea (p<0.05) 皆有顯著增加,粒線體生合作用 Tfam, NrF1 mRNA 表現也有顯著增 加 (分別為 1.4 及 1.2 倍) (p<0.05) 。白色脂肪相關基因 Retn 與 PPARγ 皆顯著低於 空白處理組,分別為0.4 及 0.5 倍 (p<0.005) (圖 2-2 (B)) 。
(三) 分化成熟前 4 天,troglitazone 處理 4 天
↑/↓:5 μM troglitazone treatment;☆:harvest
(p<0.01) 都有顯著增加。白色脂肪相關基因 Retn 顯著低於空白處理組 (0.6 倍) (p<0.005) ,而 Chemerin 卻顯著增加 (1.9 倍) (p<0.01) ,PPARγ 也顯著低於空白 組 (0.8 倍) (p<0.01) (圖 2-3 (A)) 。
(2) 分化法二:以 5 μM troglitazone 處理 4 天後,棕色脂肪 PGC1α、UCP1、Cidea 及粒線體生合作用等相關基因NrF1、Tfam 都有顯著增加 (1.4-1.9 倍) (p<0.05 ~
p<0.005) 。白色脂肪相關基因表現 Retn 與 PPARγ 皆都顯著受到抑制 (p<0.005)
(圖 2-3 (B)) ,而 Chemerin 則無顯著差異 (p>0.05)。根據上述結果,分化方法一培養之脂肪細胞在棕色脂肪相關基因表現上,和 空白處理組相比,增加幅度都較分化方法二高。白色脂肪基因和轉錄活化因子 PPARγ 的基因表現,分化方法一也都稍高於分化法二。批次實驗結果也顯示,分 化方法一的重複性較高 (圖 2-4 (A) (B) (C)) 。整體而言,在分化成熟前 4 日開始,
處理4 天 (圖 2-4 (C)) 的脂肪細胞在各個目標基因表現之改變量的重複性較高。
後續山苦瓜樣品處理實驗皆以此模式進行試驗:以分化法一 (分化培養基配置 於10 % BS/DMEM) 培養 3T3-L1 脂肪細胞,樣品處理時間點為分化成熟前 4 天開 始處理
4 天。
二、山苦瓜萃物處理
3T3-L1 脂肪細胞褐化相關基因表現
(一) 山苦瓜乙酸乙酯萃物暨其區分物EAE (乙酸乙酯萃物) :脂肪細胞在低濃度 EAE 處理下,棕色脂肪相關基因 PGC1α、
UCP1 及 cidea 均高於空白處理組。其中 PGC1α 在濃度 5 及 0.2 μg/mL 下皆有顯著 增加 (p<0.05 ~ p<0.005) ,UCP1 在濃度 5 μg/mL 及 1 μg/mL 下有顯著增加 (p<0.05) ,增加幅度皆為空白組的 1.6 倍。Cidea 則是在濃度 5 μg/mL 及 0.2 μg/mL 下顯著較高 (p<0.05 ~ p<0.01) (圖 2-5 ) 。然而 Prdm16、Tfam、 NrF1 及 cox7a1 等其他棕色脂肪及粒線體生合成作用相關基因和空白組並無顯著差異 (圖 2-5、圖 2-6 ) 。cox8b mRNA 表現在各濃度處理下,皆顯著高於空白組 (p<0.05) 。白色脂 肪相關基因表現上,Retn 與 Chemerin 和空白組相比並無顯著差異 (p>0.05) (圖 2-7 ) 。在轉錄調控因子基因表現上,PPARα 在濃度 5 及 0.2 μg/mL 下有顯著增加 (1.4 倍、1.5 倍) (p<0.05~ p<0.005) ,PPARγ 及 C/EBPα 表現雖高於空白組,卻無 顯著差異 (p>0.05) ,C/EBPβ 則在 0.2 μg/mL 下顯著增加高於空白組 (1.2 倍)
S(皂化物):脂肪細胞在低濃度 S 處理下,棕色脂肪相關基因 PGC1α、UCP1 及 Cidea 均高於空白處理組,其中PGC1α 在濃度 0.2 μg/mL 有顯著增加 (1.4 倍) (p<0.01) , UCP1 基因表現在 5μg/mL 濃度下有顯著增加 (幅度分別為 1.5 倍) (p<0.05) 。Cidea 在5 μg/mL 濃度下,有最大增加幅度 1.7 倍 (p<0.05) (圖 2-5 ) 。而 prdm16、Tfam、
NrF1 及 cox7a1 等其他棕色脂肪及粒線體生合成作用相關基因和空白組並無顯著差 異 (圖 2-5、圖 2-6 ) 。雖然 cox8b 有高於空白組之趨勢,但並無顯著差異 (濃度 5 μg/mL 時,p=0.052) 。白色脂肪相關基因表現,Retn 與 Chemerin 和空白組相比並 無顯著差異 (p>0.05) (圖 2-7 ) 。轉錄調控因子基因表現,PPARα 在濃度 0.2 μg/mL 下有顯著增加 (1.2 倍) (p<0.01) ,C/EBPβ 則在山苦瓜皂化物處理濃度 1 μg/mL 下 有顯著增加 (1.3 倍) (p<0.005) ,而 PPARγ 及 C/EBPα 和空白組相比並無顯著差異 (p>0.05) (圖 2-8 ) 。
NS(不皂化物) :脂肪細胞在 NS 處理下,棕色脂肪相關基因 PGC1α、UCP1 及 cidea 均高於空白處理組。其中PGC1α 在濃度 1、0.2 μg/mL 有顯著增加 (皆為 1.3 倍) (p<0.005) 。UCP1 mRNA 基因表現於濃度 5、1、0.2 μg/mL 處理後,增加幅度分 別為1.2 倍、1.5 倍、1.8 倍,在 1 μg/mL 濃度下有顯著差異 (p<0.005)。在 1 及 0.2 μg/mL 濃度 NS 處理下,Cidea mRNA 顯著高於空白組 (1.4 倍及 1.3 倍) (圖 2-5 ) 。 然而Tfam 在各濃度皆顯著低於空白組 (0.9 倍) (p<0.01 ~ p<0.005) 。Cox7a1 表現 量在濃度5 μg/mL 處理下有顯著減少 (p<0.005) ,Prdm16 及 NrF1 mRNA 基因表 現和空白組相比並無顯著差異 (p>0.05) (圖 2-5、圖 2-6 ) 。Cox8b 在濃度 5 μg/mL 處理下顯著增加 (2.1 倍) (p<0.05) 。白色脂肪相關基因表現,Retn 與 Chemerin 和 空白組相比並無顯著差異 (p>0.05) (圖 2-7 ) 。轉錄調控因子基因表現,PPARα 在 濃度0.2 μg/mL 下顯著較高 (1.2 倍) (p<0.05) ,而 PPARγ 和空白處理組相比並無 顯著差異。C/EBPα mRNA 表現在濃度 1、0.2 μg/mL 分別增加 1.2 倍及 1.4 倍,有 顯著高於對照組的現象, (p<0.05) ,而 C/EBPβ 則在濃度 5 μg/mL 下有顯著增加 (1.2 倍) (p<0.05) (圖 2-8 ) 。
表現有增加的現象,但和對照組相比 (vehicle + forskolin) 並無顯著差異。1 μg/mL 皂化物 (S) 處理組在 forkolin 刺激 8 小時後,UCP1 表現有顯著高於對照組 (p<0.05) ,而 C/EBPβ 表現則無顯著差異 (p>0.05) 。1 μg/mL 不皂化物 (NS) 以 forkolin 刺激 8 小時後,UCP1 表現則是低於對照組,C/EBPβ 表現則較高,但皆無 顯著差異 (p>0.05) 。
(二) 山苦瓜水萃物及納豆菌作用後之水萃物
WE (60℃水萃物):在棕色脂肪相關基因 PGC1α、UCP1、Cidea、Prdm16 及粒線體 生合成作用相關基因Tfam、NrF1、cox7a1、cox8b 的表現,和空白處理組相比皆 無顯著差異 (p>0.05) 。但值得注意的是,濃度 1 μg/mL 的 WE 處理下,脂肪細胞 在各個棕色脂肪相關基因表現增加的幅度都是最大 (圖 2-10、圖 2-11) 。白色脂肪 相關基因表現上,Retn 在濃度 0.2 μg/mL WE 處理下,會顯著受到抑制 (0.7 倍) (p<0.05) ,而各濃度下,Chemerin 和空白組相比並無顯著差異 (p>0.05) (圖 2-12 ) 。 轉錄調控因子基因表現,PPARα、PPARγ、C/EBPα 及 C/EBPβ 的基因表現和空白 組相比,皆無顯著差異 (p<0.05) (圖 2-13 ) 。
WEn (納豆菌作用後之水萃物):在濃度 5 μg/mL 處理下,PGC1α、Cidea、Cox7a1 及Cox8b 的基因表現,都顯著受到抑制 (p<0.01 ~ p<0.005) ,UCP1 在濃度 0.2μg/mL 處理下也顯著受到抑制 (p<0.05) (圖 2-10、圖 2-11) 。白色脂肪相關基因表現,Retn 在濃度5 μg/mL 處理下顯著受到抑制 (0.6 倍) (p<0.005) ,而 Chemerin 在濃度 0.2 μg/mL 處理下有受到抑制的趨勢 (p=0.057) (圖 2-12) 。轉錄調控因子基因表現,濃 度5 μg/mL 處理下,PPARγ 及 C/EBPα 會顯著受到抑制 (p<0.05 ~ p<0.005) ,而相 同濃度下,C/EBPβ 則是顯著上升 (p<0.01) 。PPARα 則是在 0.2 μg/mL 濃度下受到 顯著抑制 (p<0.05) (圖 2-13) 。
三、山苦瓜萃物處理
3T3-L1 脂肪細胞之檸檬酸合成酶 (CS) 活性
(一) 山苦瓜乙酸乙酯萃物暨其區分物濃度5 及 1 μg/mL 乙酸乙酯萃物 (EAE) 處理下,能顯著增加 CS 的活性 (約 1.2 倍) (p<0.05 ~ p<0.01) 。但另外給予 10 μM forskolin 刺激 8 小時,CS 活性並無
forskolin 刺激,皂化物 (S) 處理組與空白處理組 (vehicle)
及對照組 (vehicle + forskolin) 相比皆無顯著差異 (p>0.05) 。各濃度不皂化物 (NS) 處理下,CS 活性和空白組相比皆無顯著差異,以 forskolin 刺激,CS 活性也無顯 著增加 (p>0.05) (圖 2-14 (A)) 。
(二) 山苦瓜水萃物 (WE) 及納豆菌作用後之水萃物 (WEn)
在各濃度WE 處理下,CS 的活性有增加的趨勢,但只有在 1 μg/mL 濃度下和 空白處理組相比,達顯著差異水平 (p=0.05) 。而各濃度 WEn 處理下,CS 的活性 和空白組相比皆無顯著差異 (p>0.05) (圖 2-14 (B)) 。
四、山苦瓜中刺激
3T3-L1 脂肪細胞褐化基因表現的可能活性成分
由前面實驗結果,山苦瓜乙酸乙酯萃取物有刺激脂肪細胞棕色脂肪相關基因 表現的潛力,乃進而探討其中可能的活性成分。先前實驗室經分離純化,鑑定出 苦瓜中9c,11t,13t-CLN 為可活化 PPARs 之化合物。CLN 由於具有共軛三烯的結構,
故在268 nm 有最大吸光值 (而 CLA 具有共軛雙烯結構,在 235 nm 有最大吸光) (Igarashi et al., 2000) 。根據紫外光吸光圖譜結果顯示,EAE 與 S、NS 相比,在 270 nm 吸光值最高,顯示 EAE 中 CLN 含量較高,S 其次,NS 最低 (圖 2-15) 。
(一) 共軛次亞麻油酸 CLN
脂肪細胞在CLN 處理下,棕色脂肪相關基因 PGC1α、UCP1、Cidea 及 Prdm16 mRNA 表現量有高於空白處理組的趨勢,皆在濃度 100 μM 有最大增加幅度 (幅度 分別為1.5 倍、2.4 倍、3.1 倍及 4.1 倍) 。其中 PGC1α 基因表現在濃度 25 μM 有 顯著增加 (1.2 倍) (p<0.005) (圖 2-16) 。粒線體生合成作用相關基因 Tfam 在濃度 50 μM 顯著高於空白組,但幅度並不大 (1.1 倍) (p<0.05) ,而 NrF1 和空白組並無 顯著差異 (p>0.05)。整體而言,Cox7a1 及 Cox8b 在各濃度下皆高於空白組,但只 有濃度100 μM 下,Cox7a1 基因表現顯著高於空白組 (p<0.05) (圖 2-17 )。白色脂 肪相關基因表現上,Retn 與 Chemerin 和空白組相比並無顯著差異 (p>0.05) (圖
(二) 植醇 phytol
脂肪細胞在 phytol 處理下,棕色脂肪相關基因 PGC1α 及 cidea 和空白處理組 相比,雖表現較高,但並無顯著差異 (p>0.05) 。而 UCP1 在各濃度下的表現量皆 顯著增加 (700、350 及 175 μM 分別增加 1.8 倍、1.5 倍及 1.5 倍) (p<0.05 ~ p<0.005) 。 在高濃度phytol 處理下 prdm16 表現較低,但隨著濃度減少有增加的趨勢,但並無 差異 (p>0.05) (圖 2-16) 。粒線體生合成作用相關基因 Tfam 在各濃度下皆顯著高 於空白組,但幅度並不大 (1.1 倍) (p<0.05) ,而 NrF1 mRNA 表現在濃度 350 及 175 μM 則顯著被抑制 (0.9 倍) (p<0.05) 。整體而言,Cox7a1 及 Cox8b 在各濃度 phytol 下皆高於空白組,唯因誤差較大,統計差異並不顯著 (p>0.05) (圖 2-17 ) 。白色脂 肪相關基因表現,Retn 和對照組相比並無顯著差異,Chemerin 則是在 350 μM 濃 度下,顯著較高 (p<0.05) (圖 2-18) 。PPARα、PPARγ 及 C/EBPβ 等轉錄調控因子 基因表現,phytol 處理組和空白組相比並無顯著差異,而 C/EBPα 在濃度 700 μM 處理下表現顯著升高 (1.2 倍) (p<0.05) (圖 2-19) 。
(三) 葉黃素 lutein
脂肪細胞在 lutein 低濃度 8.75 μM 處理下,UCP1 mRNA 表現顯著高於空白組 (p<0.05) 。而棕色脂肪相關基因 PGC1α、Cidea 及 prdm16 和空白組相比雖無顯著 差異,但隨著濃度降低有升高的趨勢 (p>0.05) ,在濃度 8.75 μM 皆有最大增加幅 度 (幅度分別為 1.2 倍、1.3 倍、2.4 倍及 1.5 倍) (圖 2-16) 。粒線體生合成作用相 關基因Tfam, NrF1 在各濃度處理下,和空白處理組並無顯著差異 (p>0.05) 。特別 的是,不同於其他棕色脂肪相關基因,在較高濃度 (35 μM) 處理下,Cox7a1 及 Cox8b 有最大增加幅度 (2.0 倍及 1.9 倍) ,其中除了 Cox7a1 基因表現在 8.75 μM 濃度處理下有顯著增加 (1.6 倍) ,其餘皆無顯著差異 (p>0.05) (圖 2-17 ) 。白色脂 肪相關基因表現上,Retn 與 Chemerin 和空白組相比並無顯著差異 (p>0.05) (圖 2-18) 。轉錄調控因子基因表現在 lutein 處理下,PPARα、C/EBPα 及 C/EBPβ mRNA 表現和空白組相比並無顯著差異,而濃度0.2 μg/mL 處理下,PPARγ 表現有顯著增 加 (1.3 倍) (圖 2-19) 。