國立臺灣大學植物病理與微生物學研究所 碩士論文
Department of Plant Pathology and Microbiology College of Bioresources and Agriculture
National Taiwan University Master Thesis
木瓜輪點病毒兩系統在不同木瓜品種中之病理性差異 與交互作用
Pathological variation and interaction between two Papaya ringspot virus (PRSV) strains in different papaya
cultivars
邱獻廣
Hsien-Kuang Chiu
指導教授:洪挺軒 博士 Advisor: Ting-Hsuan Hung, Ph.D.
中華民國 101 年 1 月
January, 2012
誌謝
將近三年的研究生生涯,隨著本篇論文完成而結束。感謝洪挺軒老師的耐心 指導,不僅在實驗、生活議題或是未來人生發展上均獲益良多,張龍生老師提供 詴驗所需種子與栽培上協助,以及田間經驗分享,葉信宏老師口詴時的指導使論 文更趨完整。感謝植物病毒室的融洽環境與所有成員,使我能順利完成實驗,雅 智學姊在我實驗遇到瓶頸時提供想法與解決之道,以及其它人在實驗過程中或生 活中的協助。感謝涵均準備所有詴驗的木瓜植株,使實驗能夠進行與完成;十分 感激伴隨多年的 B94 同學們,從大學到碩士班,一路上的陪伴幫助我解決難題,
每次的開心聚會讓我宣洩壓力,鼓勵我勇敢向前邁進;碩士班生活有苦有樂,是 有記憶以來印象深刻的旅程,身邊有人休學有人畢業,即使讀得很徨恐,終究還 是畢業了,在前往下個目標前,收拾好心情準備面對新挑戰,企盼大家也朝著各 自目標邁進,最後仍要感謝曾經、現在或未來幫助我的人,沒有你們就沒有現在 的我,謝謝!
2012 / 02/ 15
中文摘要
木瓜輪點病毒(Papaya ringspot virus = PRSV)是木瓜栽培中主要的限制因子,
番木瓜罹病植株的葉片會產生嵌紋、黃化等病徵,果實出現輪點病斑,影響果實 外觀、產量與品質。依據葉片上所引起病徵做區分,PRSV 可分為畸形系統
(deformation , DF strain)、嚴重嵌紋壞疽系統(severe mottling with necrosis, SMN strain)與嚴重嵌紋系統(severe mottling, SM strain)。畸形系統為目前田間最常見 的系統,但嚴重嵌紋壞疽系統則最具摧毀性。本實驗擬透過病徵表現與植株內病 毒增殖情形作為依據,綜合病理性與分子性的詴驗證據,建立一套完整的木瓜寄 主對 PRSV 抗(耐)感病性的評估方法,實驗過程之中配合應用即時定量反轉錄 聚合酶連鎖反應(real-time RT-PCR)的追蹤,比較受詴品系與對照的常用品系(如 台農一號、台農二號、紅妃等)之間病毒增殖動態之差異,以協助新育成木瓜品 系之篩選。此外,以同時接種 DF 與 SMN 系統的病毒感染方式、或兩者一前一後 的感染方式進行詴驗,藉由具有系統特異性的 real-time RT-PCR 定量偵測法來追蹤 不同病毒系統的增殖狀態,探討兩系統在木瓜寄主中的交互作用,裨益瞭解受詴 品系對兩個病毒系統複合感染的抗(耐)病性。由本論文的挑戰接種詴驗結果得 知受詴之三新品系(10A、10B 與 10F)皆對 DF 系統有耐病性,但只有 10A 品系 對 SMN 系統有耐病性,另兩品系對 SMN 則較感病。RT-PCR 均偵測到受詴木瓜 三品系均可被 PRSV 感染,而 real-time RT-PCR 偵測結果更發現新品系木瓜體內病 毒量高於一般常見的木瓜品種約 10 倍以上,顯示病毒在新品系中增殖狀況更佳,
但是外表病徵輕微而病徵指數低,表示其耐病性相當良好,PRSV 的大量複製並未 導致嚴重的病徵產生。另一方面,詴驗發現 SMN 系統對商業品種台農二號與紅妃 適應力好,體內病毒量高,田間栽培二品種有利於 SMN 系統生存。病毒系統之交 互作用的詴驗部份,同步與非同步接種兩系統病毒後,發現先接種的病毒主導病 徵發展,以台大八號(NTU 8)進行先接種 DF、後接種 SMN 感染詴驗中,先接種 SMN 系統會被 DF 系統追上或拉近差距;反之,先接種 DF、後接種 SMN 時,後 接種的 SMN 系統無法追上原先 DF 系統病毒量;同步接種二病毒系統時寄主體內 兩病毒系統互相競爭,後期 DF 系統取得優勢。由此看出 DF 系統在田間似乎仍較 SMN 系統優勢,但仍需注意未來 PRSV 系統之間的演化發展。本研究應用 real-time
RT-PCR 進行 PRSV 的定量偵測,綜合病毒增殖與病徵發展的兩項資訊,可提供評 估新品系木瓜抗(耐)病性時之重要參考資訊;複合感染詴驗的結果也顯示,台 灣木瓜新品種台大八號目前耐病性仍很穩定,唯因應 PRSV 系統的長期演化,長 期的耐抗病育種工作仍應持續。
關鍵字:木瓜輪點病毒;即時定量反轉錄聚合酶連鎖反應;病毒系統;耐病性
Abstract
Papaya ringspot virus (PRSV) is one of the limiting factors in the papaya industry.
The infected papayas produce several symptom such as mosaic, yellowing, ringspot in fruit, which causes tremendous economic losses and poor quality of fruits. According to the symptoms developed on leaves, PRSV was categorized into three strains: SM(severe mottling), DF(deformation),and SMN(severe mottling with necrosis)strains. The DF strain is predominated in the field at present time, and the SMN strain induce most destructive symptom in papayas. This study we dedicated to investigate the correlation between the severity of symptom and replication of virus, and develop a evaluation method to evaluate the resistance or tolerance of new papaya cultivars. The RT-PCR and real-time RT-PCR assays were used in this study to monitor the fluctuation dynamic of virus replication for comparative study of tolerance between new developed papaya lines and commercial cultivars. In addition, the simultaneously or asynchronously inoculation experiment test with the DF and SMN strains to were also conducted to investigate the interaction between these 2 strains in either new line or commercial cultivars of papaya based on the real-time RT-PCR assays with train-specific primers.
The result might be able to know the co-infection effect of the DF and SMN strains in various papaya cultivars(lines). The inoculation tests showed that all of 3 newly bred papaya lines(10A, 10B, and 10F) were tolerant to the DF strain without appearing apparent symptoms. However, only the 10A line is tolerant to the SMN strain. The real-time RT-PCR assays demonstrated the amounts of replicated virus in newly bred lines was 10 times higher than those in commercial cultivars. It means revealed that these newly bred papaya lines are actually better host for PRSV-replication though they do not produce symptoms showing their tolerance. The data also indicated that the SMN strain replicated very well in either Tainung 2 (TN 2) and Red-Lady (RL) cultivars, which means TN 2 and RL probably provide better conditions for survival of
PRSV-SMN. In the asynchronously inoculation tests with both DF and SMN strains, all of infected papayas by PRSV-DF before PRSV-SMN (2-week interval) developed the symptoms similar to those induced by PRSV-DF whereas all of infected papayas by PRSV-SMN before PRSV-DF developed the symptoms similar to those induced by PRSV-SMN. It presented that the earlier invading virus strain determines the
were co-infected by the DF and SMN simultaneously. In the inoculation tests of the NTU8 papaya, the real-time RT-PCR assays demonstrated that the later invading PRSV-DF could replicate well and reach or exceed the amount of earlier infecting PRSV-SMN whereas the later invading PRSV-SMN could not reach the amount of earlier infecting PRSV-DF. These two strains competed each other when they simultaneously co-infected a papaya host, and the DF strain gradually became
prevailing. This study reveals that the DF trains seemed to be more predominant than the SMN strain in the field, and we should pay more attention to the evolution of various PRSV strains. The investigation of symptomatology and quantitative detections of PRSV based on real-time RT-PCR assays provided important data to evaluate the resistance or tolerance against PRSV for newly bred papaya lines. The newly developed papaya cultivar tolerant to PRSV, NTU 8, showed its stable tolerance in the co-infection trials with both DF and SMN strains. However, the continuous breeding of papaya should be conducted to cope with the evolution of various strains of PRSV.
Keyword: Papaya ringspot virus ; real-time RT-PCR ; virus strain ; tolerance
目錄
口詴委員會審定書 ...……i
誌謝 ... ii
中文摘要 ... iii
Abstract ... v
目錄 ... vii
表目錄 ...ix
圖目錄 ... x
壹、前言 ... 1
貳、前人研究 ... 3
一、 木瓜輪點病毒之發現 ... 3
二、 木瓜輪點病毒之分類與型態 ... 3
三、 木瓜輪點病毒之寄主範圍與病徵 ... 4
四、 木瓜輪點病毒之傳播方式 ... 5
五、 木瓜輪點病毒之防治方法 ... 5
六、 台灣木瓜輪點病毒之偵測方法 ... 6
七、 台灣木瓜輪點病毒之系統 ... 6
八、 即時定量聚合酶連鎖反應(Real-Time PCR) ... 8
九、 不同植物病毒系統之交互作用 ... 9
參、材料與方法 ... 10
一、 詴驗植物之準備 ... 10
二、 木瓜輪點病毒(PRSV)之來源與接種 ... 10
三、 PRSV 核酸抽取 ... 10
四、 PRSV 接種詴驗 ... 11
(1) 單獨接種 PRSV ... 11
(3) 病徵分級 ... 12
五、 PRSV 之偵測 ... 12
六、 即時定量聚合酶連鎖反應 ... 13
(1) RNA 反轉錄 ... 13
(2) 標準品與標準曲線之建立 ... 13
(3) Taqman Primer / Probe 之設計 ... 14
(4) Real-time RT-PCR 反應 ... 14
肆、結果 ... 15
一、 不同木瓜輪點病毒系統在不同木瓜品種上病徵表現 ... 15
二、 不同木瓜輪點病毒系統在不同木瓜品種上病毒增殖情形 ... 16
三、 非同步接種不同木瓜輪點病毒系在不同木瓜品種上病徵表現 ... 16
四、 同步接種不同木瓜輪點病毒系在不同木瓜品種上病徵表現 ... 18
五、 非同步接種不同木瓜輪點病毒系在不同木瓜品種上病毒增殖動態 ... 18
六、 同步接種不同木瓜輪點病毒系在不同木瓜品種上病毒增殖動態 ... 20
伍、討論 ... 21
陸、參考資料 ... 25
柒、圖與表 ... 33
表目錄
表 一、以反轉錄聚合酶連鎖反應(RT-PCR)與即時定量反轉錄聚合酶連鎖反應
(REAL-TIME RT-PCR)偵測木瓜輪點病毒(PRSV)所用之引子與探針。 ... 34
表 二、木瓜輪點病毒感染不同品種(系)木瓜寄主所產生的病徵級數鑑別 ... 35 表 三、以反轉錄聚合酶鏈鎖反應(RT-PCR)偵測木瓜輪點病毒畸形系統(PRSV-DF)
與嚴重嵌紋系統(PRSV-SMN)於不同品種中之增殖情形。 ... 38
圖目錄
圖 一、木瓜輪點病毒畸形系統(PRSV-DF)感染 6 種不同品種或品系(10A、10B、
10F、TN1、TN2、RL)的木瓜在 28 天、56 天、70 天後之病徵表現。 ... 36 圖 二、木瓜輪點病毒嚴重嵌紋壞疽系統(PRSV-SMN)感染 6 種不同品種或品系
(10A、10B、10F、TN1、TN2、RL)的木瓜在 28 天、56 天、70 天後之病徵 表現。 ... 37 圖 三、以即時定量反轉錄聚合酶連鎖反應(REAL-TIME RT-PCR)偵測木瓜輪點病毒
畸形系統(PRSV-DF)在不同的木瓜品種(台農一號、台農二號與紅妃)中之增殖 動態。 ... 39 圖 四、以即時定量反轉錄聚合酶連鎖反應(REAL-TIME RT-PCR)偵測木瓜輪點病毒
嚴重嵌紋壞疽系統(PRSV-SMN)在不同的木瓜品種(台農一號、台農二號與紅 妃)中之增殖動態。 ... 40 圖 五、木瓜輪點病毒畸形系統(PRSV-DF)接種在不同木瓜品種(NTU8、TN2、RL)
木瓜後 14 天、28 天、56 天與 70 天後之病徵表現。 ... 41 圖 六、木瓜輪點病毒嚴重嵌紋壞疽系統(PRSV-SMN)接種在不同品種(NTU8、
TN2、RL)木瓜在 14 天、28 天、56 天與 70 天後之病徵表現。 ... 42 圖 七、非同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)(先接種 PRSV-DF/ 後接種 PRSV-SMN)於不同品種(NTU8、
TN2、RL)的木瓜在 14 天、28 天、56 天與 70 天後之病徵表現。 ... 43 圖 八、非同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)(先接種 PRSV-SMN/ 後接種 PRSV-DF)於不同品種(NTU8、
TN2、RL)的木瓜在 14 天、28 天、56 天、70 天後之病徵表現。 ... 44 圖 九、同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)於不同品種(NTU8、TN2、RL)的木瓜在 14 天、28 天、56 天、
70 天後之病徵表現。 ... 45 圖 十、非同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)(先接種 PRSV-DF/ 後接種 PRSV-SMN)於不同品種木瓜(NTU8、
TN2、RL)的病毒增殖動態。 ... 46 圖 十一、非同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)(先接種 PRSV-SMN/ 後接種 PRSV-DF)於不同品種木瓜(NTU8、
TN2、RL)的病毒增殖動態。 ... 47 圖 十二、同步接種木瓜輪點病毒畸形系統(PRSV-DF)與嚴重嵌紋壞疽系統
(PRSV-SMN)於不同品種木瓜(NTU8、TN2、RL)的病毒增殖動態。 ... 48
壹、前言
木瓜(Carica papaya L.),番木瓜科(Caricaceae),番木瓜屬,為世界上重要熱帶 經濟果樹,原產於熱帶美洲,17 世紀傳入亞洲,清朝末年再由中國大陸引入台灣,台 灣栽培歷史達百年之久,1940 年後因技術與品種之改良,產量與品質逐漸提升,栽種 面積持續增加,由於木瓜栽培容易,生長快且產量高,且外銷日本,為台灣重要經濟果 樹。1975 年台灣南部發現木瓜輪點病毒入侵(陳等,1976),此病最早在 1945 年於夏 威夷發現(Lindner et al, 1945),短短數年內蔓延全台且為害嚴重(王等,1978),摧毀 台灣木瓜產業,木瓜由多年生變為一年生,單位面積產量下降,品質降低,影響生產者 收益。
木瓜輪點病毒(Papaya ringspot virus, PRSV)分類於 Potyviridae 科,Potyvirus 屬(馬 鈴薯病毒 Y 屬)的長絲狀病毒,基因體為(+)ssRNA,約由 10326 個核甘酸組成,
病毒顆粒長約 780 nm(De la Rosa and Lastra, 1983)。依據其是否能感染番木瓜,區分 為 P 型與 W 型(Purcifull et al, 1984),P 型可感染番木瓜科、葫蘆瓜科及藜科,在台灣 依照 PRSV 在番木瓜葉片上產生病徵的不同分為數種系統(strain):輕微嵌紋系統(mild mottle, M)、嚴重嵌紋系統(severe mottle, SM)、嚴重嵌紋壞疽系統(severe mottle with necrosis, SMN)、畸形系統(deformation, DF)(翁,1981;卲井,1986;林,1980;關,
1990;廖,2004)。
木瓜輪點病毒可藉由機械方式傳播,田間則是透過媒介蚜蟲以非永續性(non-
persistent)方式傳播病毒,其中桃蚜(Myzus persicae)傳播能力最強,由於媒介昆蟲獲 毒與傳毒時間在數分鐘內即可完成(Jensen, 1949b; Singh, 1971),因此不易以藥劑防治 此病害。台灣早期以間作玉米和鋪設黑色反光塑膠布閃避媒介昆蟲(王,1991),或是 利用弱毒系統進行交叉保護作用,但效果均不佳(林,1980;翁,1981; Wang et al, 1987;
Yeh et al, 1988),爾後發展出網室栽培方式(徐,1989),雖然有效地隔絕媒介蚜蟲,
但也衍生新問題,如成本高、通風不良及易有白粉病和紅蜘蛛等等,因此並非長久良方。
近年來,中興大學葉錫東教授團隊,以基因靜默(gene silencing)作為理論基楚,將病 毒的鞘蛋白(Coat protein)轉殖至木瓜基因中,成功開發出抗木瓜輪點病毒轉基因木瓜,
在田間詴驗後期被木瓜畸葉嵌紋病毒(Papaya leaf distortion mosaic virus, PLDMV)感
染,於是又開發出雙抗病毒轉基因木瓜(包,2000;包等,2006;龔,2004),但礙於 食物安全與生態環境等因素,加上未被政府許可在田間生產,所以現今栽培方法仍以網 室栽培為主流,隔除媒介昆蟲達到防治目的。對轉基因作物有疑慮情況下, 仍需以傳 統雜交育種方式選拔抗病或耐病作物,台大園藝系遺傳育種實驗室利用傳統育種方法,
從雜交後代中成功選拔出新的耐病品系─台大八號(王,1997;王,2001; 郭,2001),
對木瓜輪點病毒與木瓜畸葉嵌紋病毒均有耐病性(吳,2007),為未來防治木瓜輪點病 一大利器。
現今偵測木瓜輪點病毒方法以反轉錄聚合酶連鎖反應(reverse-transcription polymerase chain reaction, RT-PCR)為主(廖,2004;李 2006),反應快速且靈敏度高,
但無法定量樣品中病毒量,近年來即時定量聚合酶連鎖反應(Real-time PCR)廣泛應 用於病原偵測(Schaad and Frederick, 2002; Schaefers, 1969; Agindotan et al, 2007),概念 為進行聚合酶連鎖反應時,同步定量核酸產物,再透過螢光偵測系統與後續資料處理,
精確地測量出樣品中原始模板濃度,再推算出樣品中病毒量,此法靈敏度高,準確性也 高(Valasek and Joyce, 2005),成功應用於偵測木瓜輪點病毒(李,2006;梁,2009)。
台灣田間木瓜輪點病毒系統變異快速且複雜,早期以嚴重嵌紋系統為流行系統,原 先族群較小的畸形系統,現今和嚴重嵌紋系統普遍存在田間(廖,2004),造成嚴重病 徵之嚴重嵌紋壞疽系統族群也有增加的趨勢,可見田間病毒系統間有複雜的交互作用,
以致於病毒系統族群間有消長行為。本研究應用 real-time RT-PCR 追蹤木瓜輪點病毒之 增殖,綜合病毒增殖與病徵發展的兩項資訊,以評估新品系木瓜的耐病性;並以複合感 染詴驗測詴其耐病的穩定性,觀察其耐病性是否被不同系統破壞其耐病機制,有助於瞭 解寄主被兩病毒系統感染時族群間消長情形,作為木瓜輪點病毒田間發病生態之參考,
對未來田間木瓜輪點病毒系統的變化趨勢有更進一步的瞭解甚至預測其變化,以便儘早 研擬因應對策或再進行新耐病品系的選拔。
貳、前人研究
一、木瓜輪點病毒之發現
木瓜輪點病毒(Papaya ringspot virus, PRSV)最早在牙買加被報導(Smith, 1928),病徵為嵌紋、黃化、心葉變小、老葉下垂枯死;而後夏威夷也發現同一病 害(Lindner et al, 1945)並定名,1949 年證實此病害由病毒造成(Jensen, 1949a),
病徵為葉片上產生嵌紋、帅葉捲曲、植株矮化、生長受阻,果實上有中央綠色黃緣 輪點。木瓜輪點病自 1945 年開始快速蔓延至熱帶與亞熱帶國家的木瓜產區,造成 嚴重危害,如美國佛羅里達州(Conover, 1962)、多明尼加(Story and Haliiwell, 1969)、
印度(Singh, 1969; Khurana and Bhargava, 1970)、夏威夷(Cook and Milbrath, 1971;
Ishii and Holtzamann, 1972)、波多黎各(Adsuar, 1972)、法屬加勒比海瓜德羅普島
(Quiot-Douine, et al. 1986)、日本(王等,1978)、委內瑞拉(De la Rosa and Lastra, 1983),以及墨西哥、德國、義大利、中東國家等皆受其害(Brunt et al, 1990)。
二、木瓜輪點病毒之分類與型態
依 International Committee for Taxonomy of Virus(ICTV)(2010)的病毒命名 分類法,將木瓜輪點病毒(Papaya ringspot virus, PRSV)歸入馬鈴薯 Y 病毒科
(Potyviridae),馬鈴薯 Y 病毒屬(Potyvirus),病毒顆粒長約 780 nm,寬約 12 nm
(Cook, 1972)。基因體是單股正譯股 RNA(De la Rosa and Lastra, 1983; Yeh and Gonsalves, 1985),由 10326 個核甘酸組成,轉譯得到 3344 個胺基酸之多蛋白再裂 解成多個小蛋白(Yeh et al, 1994),如內含體(Yeh et al, 1992)、鞘蛋白(Gonsalves, and Ishii, 1980; Yeh and Gonsalves, 1985)。多蛋白自 N 端起依序為 5‟UTR
(untranslated region),P1(RNA–binding helper protein & proteinase), HC–Pro
(helper component for insect transmission), P3(unknown function), 6K1(unknown function), NIa–VPg(5‟terminus genome–linked viral protein), NIa–Pro(nuclear inclusion component a & proteinase), Nib–Pro(nuclear inclusion component b &
polymerase), Coat protein, 3‟UTR(unrelated resion)等(Yeh et al, 1992; Yeh, 1994)。
以電子顯微鏡觀察木瓜輪點病毒顆粒為長絲狀病毒,長度約在 620-830 nm 之 間(陳等,1976;陳,1984),不同系統之病毒顆粒長度也不同,約在 660-726 nm 之間(林,1990)。以電子顯微鏡觀察木瓜病葉組織切片,在葉肉組織的細胞質中 有束狀(bundles)、環狀(circulars)、風車狀(pinwheels)之內含體(inclusion)(Zettler et al, 1968)。依其圓柱狀內含體的形式,將 Potyvirus 分成四個亞群:(1)捲軸狀
(scrolls)、(2)長直層狀聚集(long straight laminated aggregates)、(3)捲軸狀與 短彎曲層狀聚集、(4)捲軸狀與長直層狀聚集(short curve laminated aggregates),
並將木瓜輪點病毒歸入第一亞群(Edwardson, 1974; Edwardson et al, 1984)。台灣木 瓜輪點病毒不同系統的內含體組成也有差異,發現在嚴重嵌紋壞疽系統(severe mottling with necrosis strain, SMN)感染的病葉肉細胞中,有風車狀、捲軸狀、短直 層狀與類似短彎曲層狀之內含體,將此系統歸類於第三亞群(王,1988);畸形系 統(severe mottling with deformation strain, DF)感染的細胞中,具有典型風車狀、
大量捲軸狀及長直層狀之內含體,並將其歸入於第三亞群(廖,2004);嚴重嵌紋 系統(severe mottling strain, SM)感染的細胞中,具有少量捲軸狀與大量的短直層 狀內含體,亦有大量短直層狀內含體的聚合,將其歸類於第三亞群;嵌紋系統
(mottling strain, M)分類於第四亞群(王,1988),除以上差異外,各系統皆在感 染細胞中造成澱粉粒增加、葉綠餅變形及細胞核膨大,周圍有大量內含體累積(王,
1988;廖,2004)。
三、木瓜輪點病毒之寄主範圍與病徵
木瓜輪點病毒寄主範圍包括番木瓜科(Caricaceae)、藜科(Chenopodiaceae)
及葫蘆科(Cucubitaceae)。依據 PRSV 是否能感染番木瓜,區分為可感染番木瓜、
藜科及葫蘆科的 P 型(P-type)與感染藜科及葫蘆科的 W 型(W-type)(Purcifull et al, 1984)。PRSV 感染番木瓜後,初期葉片上產生葉脈透化,後期有嵌紋、黃化 及畸形病徵,葉面積變小,葉柄與莖部出現油浸狀條斑,帅葉皺縮畸形(Purcifull et al, 1984)。被害植株矮化、生長受阻及不易開花著果,花瓣上出現油浸狀輪紋,果 實發育不良或畸形,表面出現水浸狀輪斑(Conover, 1964)。
四、木瓜輪點病毒之傳播方式
木瓜輪點病毒無法藉由種子傳播(王等,1978;Capoor and Varma, 1948; Capoor and Varma, 1958; Ridings et al, 1978),但可經由機械磨擦、針刺、嫁接(Adsuar, J.
1946)及蚜蟲傳播,目前證實有傳播能力之蚜蟲有桃蚜(Myzus persicae)(Jensen, 1949b; Capoor and Varma, 1948; Capoor and Varma, 1958; Conover, 1962; Ishii and Holtzamann, 1963; Zettler et al, 1968)、棉蚜(Aphis gossypii)(Jensen, 1949b; Capoor and Varma, 1948; Capoor and Varma, 1958; Conover, 1962)、綠桔蚜(A. spiraecola)
(Adsuar, J. 1946; Cook, 1972)、苜蓿蚜(A. medicaginis)(Jensen, 1949b; Capoor and Varma, 1958)、夾竹桃蚜(A. nerii)(Schaefers, 1969; Khurana and Bhargava, 1970)、
豆蚜(A. rumicis)(Jensen, 1949b)、豇豆蚜(A. caeccivora)(Cook, 1972)、天竺葵 蚜(A. malvae)(Capoor and Varma, 1948; Capoor and Varma, 1958)、大戟長管蚜
(Macrosiphum euphorbiae)(Jensen, 1949b)等,其中以桃蚜和棉蚜效率最高。田 間媒介昆蟲以非永續性方式傳毒,蚜蟲獲毒只需 2 分鐘(Jensen, 1949b),在健康 植株上停留 5 分鐘即可傳毒,達 15 分鐘時具有最大帶毒力(Jensen, 1949b; Singh, 1971),病毒在蟲體內停留時間因蟲不同而異(Jensen, 1949b; Namba and Higa, 1977),
若獲毒前蚜蟲飢餓處理 30 分鐘(Namba and Higa, 1977)或 4 小時後可提高傳毒能 力(Singh, 1971),且蟲數量越多傳播率越強,所以蚜蟲族群高峰期時病害嚴重
(Jensen, 1949b ),蚜蟲帶毒後可傳播距離達 350 英呎,因此病害蔓延極快
(Wolfenbarger, 1966)。
五、木瓜輪點病毒之防治方法
田間蚜蟲傳播木瓜輪點病毒效率高,木瓜感染後無法以化學藥劑治癒,殺蟲劑 對蚜蟲傳,因此以逃避媒介昆蟲為主要防治策略。早期以田間鋪設反光布或間作玉 米,蚜蟲不易接觸到木瓜寄主,但成效不如預期(王,1991);人工誘變木瓜輪點 病毒產生弱毒系統,以交叉保護(Cross-protection)方式進行防治(林,1980;
翁,1981; Wang et al, 1987; Yeh et al, 1988),但因技術門檻高、接種數量少及系統 專一性等問題,效果不佳(Yeh and Gonsalves, 1994);夏威夷於 1985 年研發抗木 瓜輪點病基因轉殖木瓜,1998 年合法上市且順利生產(Fuchs and Gonsalves, 2007),
台灣葉錫東教授團隊自 1988 年開始研發基因轉殖抗病植株,1996 年進行田間評估,
詴驗結束前被另一木瓜病毒─木瓜畸葉嵌紋病毒(Papaya leaf-distortion mosaic virus, PLDMV)感染,2004 年再育成雙抗病毒基因轉殖木瓜(包,2000。包等,
2006);台灣 1985 年後採用網室栽培木瓜,阻絕蚜蟲傳播病毒(徐,1989),但網 室內通風不良,易受到白粉病與紅蜘蛛為害,且搭建網室成本高,收益有限;臺大 園藝系張龍生教授利用傳統雜交育種,自雜交後代中選拔出耐病品系─台大八號
(原台大選系十號)(王,1997;王,2001; 郭,2001)。
六、台灣木瓜輪點病毒之偵測方法
木瓜輪點病毒常用偵測法為抗血清法或由單株抗體進行酵素連結免疫分析
(Enzyme-linked immunosorbent assay, ELISA)(陳,1986;關,1990;廖,2004;
Gonsalves and Ishii, 1980; Thomas and Dodman, 1993; Pourrahim et al, 2003; Davis et al, 2005),由機器測定吸光值,依其讀值判斷是否罹病;其二鑑定方法為利用 Potyvirus 廣效性與病毒專一性引子製備木瓜輪點病毒 cDNA 探針,與 DIG 標定之 待測核酸物質,進行逆墨點雜合分析法(reverse dot-blot hybridization)(徐,2001;
葉,2004);近年來,以反轉錄聚合酶鏈鎖反應(reverse-transcription polymerase chain reaction, RT-PCR)配合廣效性引子對(Gibbs and Mackenzie, 1997)進行增 幅,產物經電泳分析後得到結果,鑑定效果不錯。本研究室更開發出系統專一性引 子對,對不同木瓜輪點病毒系統進行偵測,所設計的引子對有 DF-703、SM-517 及 SMN-467,RT-PCR 增幅後電泳分析,可準確偵測並區分不同病毒系統(廖,
2004 )。
七、台灣木瓜輪點病毒之系統
早在 1975 年,台灣南部─高雄大樹、阿蓮及燕巢鄉等地發現木瓜疑似罹患輪 點病,以電子顯微鏡觀察後細胞內有長約 700 nm 的長絲狀病毒顆粒,判斷此木瓜 病害是由木瓜輪點病毒所引起(陳等,1976)。爾後 2-3 年內,木瓜輪點病迅速蔓 延至全省木瓜產區(王等,1978),對此病害至今已有三十多年研究歷史。依據葉 片上病徵分為五個系統:一、輕微嵌紋系統(mild mottle, M):低溫下無病徵,高
溫時葉片產生黃斑或輕微嵌紋(翁,1981;卲井,1986;林,1980);二、嚴重嵌 紋系統(severe mottle, SM):葉片嚴重嵌紋,低溫時出現畸形葉,莖、葉及果實有 水浸狀斑(翁,1981;卲井,1986;林,1980; Lin et al, 1989);三、嚴重嵌紋壞疽 系統(severe mottle with necrosis, SMN):葉片嚴重嵌紋及壞疽,帅葉萎凋(翁,1981;
卲井,1986;林,1980);四、嚴重嵌紋畸形系統(severe mottle with deformation, SMD):
葉面黃化、葉片嚴重嵌紋及萎縮變形(翁,1981;卲井,1986;林,1980);五、
畸形系統(deformation, DF):葉片嚴重畸形(關,1990)。1975 年台灣發現木瓜輪 點病毒,1978 年調查報告指出,田間多為造成葉片嵌紋的輕微嵌紋系統(M),少 數嚴重嵌紋(SM)及嚴重嵌紋壞疽系統(SMN)(王等,1978);1982 年時,田間 流行系統轉為 SM 系統,其次為 SMN 系統與 M 系統(林,1980);1986 年田間發 現嵌紋畸形系統(SMD)(卲井,1986),1990 出現畸形系統(DF)(關,1990);
1995 年仍以 SM 系統為主,畸形系統(DF)數量升高及少數 SMN 系統(蔡,1995); 原本夏天較少的 DF 系統,2003 年時轉變為 SM 系統與 DF 系統各半,冬季 SM 系 統會出現 DF 系統之病徵(廖,2004)。台灣 PRSV 田間系統變異快速且複雜,早 期以 M 系統為主,隨即被 SM 系統取而代之,DF 系統出現後,直至今日 DF 系統 族群已與 SM 系統族群相當,可見系統間之消長變化難測,SMN 系統族群雖小,
但會造成嚴重病徵並使寄主快速萎凋,為極具摧毀性之潛在系統,不容忽視其存 在。
八、即時定量聚合酶連鎖反應(Real-Time PCR)
在 1990 年代即發現 Taq DNA polymerase 具有的 5‟nuclease 活性可將螢光標定 的探針水解,間接測量 PCR 增幅產物(Holland et al, 1991);同時也發現利用螢光 染劑標定之核苷酸進行 PCR,配合光學或雷射偵測系統同步監測增幅情形(Higuchi et al, 1992; Higuchi et al, 1993)。此技術之概念為進行 PCR 反應時,同步偵測增幅 物產量,因此又稱定量聚合酶連鎖反應(quantitative PCR, qPCR),儀器由 PCR 反應器與光學偵測系統,加上數據處理系統,即為 Real-time PCR 定量技術。主 要分為兩種系統:一為 SYBR Green I dye chemistry(Wittwer et al, 1997),特色是 利用可嵌入雙股 DNA 的螢光物質─SYBR Green I ,它會結合至雙股 DNA minor groove,在有激發光時釋放固定波長的光,偵測系統可監測 dsDNA 變化,非目標 PCR 產物(如 primer-dimer)也不會被系統排除,因此實驗結束後仍頇進行解離 曲線分析(dissociation curve)確定 PCR 反應之專一性(Valasek and Joyce, 2005)。
二為 TaqMan Chemistry(Gibson et al, 1996; Holland et al, 1991),原理是利用一對 引子對(reverse / forward primer pair)與一段專一性探針(probe),探針兩端為不 同螢光物質,5‟為 reporter dye,3‟為 quencher dye(Livak et al, 1995),當探針未被 水解前,由於兩螢光物質位於探針上距離近,激發後能量會由 reporter dye 轉移至 quencher dye 上(fluorescence resonance energy transfer, FRET)(Walker, 2002;
Lakowicz, 2006),產生遮蔽效應(Quench),機器只能偵測到一種波長的光。PCR 反應進行後,Taq DNA polymerase 的 5‟exonuclease 活性將探針水解,遮蔽效應消 失,能量由 quencher dye 轉移至 reporter dye 上並發出螢光(Heid et al, 1996),其 螢光強度與 PCR 產物量成正比,軟體分析後可得到增幅曲線。若在同一反應中加 入不同螢光物質的 reporter dye,可同時偵測不同目標 DNA(multiplex real-time PCR)(Agindotan et al, 2007; Mumford et al, 2000; Vingano and Stevens, 2007)。此外 定量方式有兩種,一為相對定量(relative quantitation),即是參考某樣品(reference sample)作不同樣本之比較;另一為絕對定量(absolute quantitation),需有已知濃 度之樣品作為標準品,同時進行偵測並定量(Mason et al, 2008)。此技術(real-time PCR 或 real-time RT-PCR)靈敏度較傳統 PCR 偵測方式高(Valasek, M.A. and Joyce, J.R. 2005.),可廣泛應用於基因表現等研究(Overbergh et al, 2003)與病原菌
之偵測(Schaad and Frederick, 2002; Schaefers, 1969)。
九、不同植物病毒系統之交互作用
依先前報導可知,木瓜輪點病毒系統在不同品種的番木瓜寄主上有不同的致病 能力(王,1982;Cook and Zettler, 1970; Provvidenti and Gonsalves, 1982; Sanchez et al, 1975),不同木瓜輪點病毒系統在相同品種之番木瓜寄主上也有不同感病能力
(Conover, 1976; Conover and Litz, 1983; Holmes, 1965; Tennant, et al 1994.),因此 可知病害嚴重度會受到病毒系統與寄主品種兩因素影響。近年來報導發現大豆嵌紋 病毒(Soybean mosaic virus, SMV)系統在同一寄主內有拮抗作用(Shi et al, 2008),
且嵌紋系統較壞疽系統更具優勢,以被兩系統感染的寄主作為接種源,被接種的寄 主只會產生嵌紋病徵(Chen et al, 2004)。梨李痘病毒(Plum pox virus, PPV)不同 系統(PPV-M, PPV-D)在不同品種梨樹(Black Diamond, Sun Gold)上有交互 作用,將 PPV-M 接種至已被 PPV-D 感染之 Black Diamond 中,PPV-M 會逐步 取代 PPV-D 之族群,反之在 Sun Gold 中則會共存;PPV-D 無法侵染被 PPV-
M 感染之 Black Diamond 和 Sun Gold 中,兩系統間無協同作用與交叉保護能力
(Capote et al, 2006)。番椒嵌紋病毒(Pepper mosaic virus, PepMV)不同分離株混 合接種同一寄主時,病徵表現與病毒量比單獨接種分離株輕微與少,感染初期病徵 近 似 健 康 值 株 , 因 此 田 間 觀 察 到 病 株 時 易 產 生 誤 判 而 造 成 防 治 上 漏 洞 , 且 PepMV-SP13(CH2 type)拮抗 PepMV-PS5(EU type),後期時兩者勢力相當,競 爭情形不影響最終病毒量與接種原數量,同時也提到病毒量與病徵嚴重度之間無明 顯關係(Gomez et al.,2009)。
參、材料與方法
一、詴驗植物之準備
詴驗中使用之番木瓜品種為台農一號(Tainung No.1 , TN1)、台農二號(Tainung No.2 , TN2)、紅妃(Red Lady , RL)、台大八號(National Taiwan University Hybrid No.8 , NTU8)及三個未定名雜交後代品系(10A, 10B, 10F)。所有詴驗植株由台大 園藝系遺傳育種實驗室提供,種子發芽後依其生長情形換盆,栽培介質為農友公司 滿地王 2 號,並混入適量蛇木、珍珠石、有機肥及好康多一號,植株高約 5 公分時 移入五吋盆中,達 35 公分時即可進行接種,詴驗地點分別為高雄區農業改良場溫 室與台大農場溫室。
二、木瓜輪點病毒(PRSV)之來源與接種
PRSV 系統有畸型系統(DF)與嚴重嵌紋壞疽系統(SMN),使用之接種源來 自實驗室保存之純系罹病木瓜植株。為維持病毒良好活性,接種前才將感染 PRSV 之木瓜葉片取下,秤取 1 克之罹病葉片,加上四倍體積的 0.05M 磷酸緩衝溶液(K-P buffer),置於玻璃板上以接種棒將其磨碎,所得汁液與金剛砂混合,以接種棒沾取 病汁液塗抹在木瓜葉片上,接種位置在木瓜的第三、四及五片本葉,作為病毒感染 來源,接種完以去離子水沖洗葉片,再以衛生紙沾乾葉表面。每隔一周記錄病徵表 現與採集系統葉(位於莖頂之完全展開葉)作為抽取核酸之樣品。
三、PRSV 核酸抽取
參考自:Hung (2000)所發表之方法。取 0.3 g 的植物組織置於研缽中,以液態 氮磨成粉末狀,加入 2 ml 核酸萃取緩衝液〔100 mM Tris–HCl(pH 8.0), 100 mM EDTA–2Na, 0.25 M NaCl, 1% N–Lauroylsarcosine 〕混合均勻,移至 1.5 ml 離心管 中,55℃乾浴 1 小時再以 6000 rpm 離心 5 分鐘,取上清液 800 μl 至新離心管中,
並加入 100 μl 5M NaCl 與 100 μl 10% CTAB(溶於 0.7M NaCl),混勻後於 65℃下 乾浴 10 分鐘,再加入 600 μl CI〔Chloroform:Isoamyl alcohol=24:1〕,混合至乳 狀再以 12000 rpm 離心 5 分鐘,取上清液 850 μl 移至新離心管中,並加入 600 μl PCI
〔Phenol:Chloroform:Isoamyl alcohol=25:24:1〕,混合至乳狀再以 12000 rpm 離心 5 分鐘,取 700 μl 上清液至新離心管中,並加入 420 μl Isopropanol 輕輕混勻,
置於-20℃冰箱 30 分鐘,於 4℃下 12000 rpm 離心 10 分鐘,去除上清液並以 500 μl 70% Ethanol 潤洗沉澱物,倒乾 70% Ethanol 再抽氣乾燥 10 分鐘,沉澱物以 150 μl TE buffer〔10 mM Tris–HCl(pH 8.0), 1 mM EDTA〕溶解。
四、PRSV 接種詴驗 (1) 單獨接種 PRSV
受詴木瓜為台農一號(Tainung No.1, TN1)、台農二號(Tainung No.2, TN2)、
紅妃(Red Lady , RL)及三新雜交品系─10A, 10B, 10F。詴驗於 2010 年 3 月初進 行接種,取病葉加緩衝液以四倍稀釋,在同一品系不同植株上分別接種不同病毒 系統(PRSV-DF, PRSV-SMN),四重複,接種葉片為第三片、第四片、第五片 本葉,接種方式參照前述“木瓜病毒之來源與接種“,接種後每隔七天取系統葉(離
莖頂最近之完全展開葉),作為核酸抽取之樣品,拍照記錄病徵及病勢發展,詴驗
時間為 10 週。
(2) 同步與非同步接種 PRSV
受詴木瓜為受詴木瓜為台農二號(Tainung No.2, TN2)、紅妃(Red Lady , RL)、 台大八號(National Taiwan University Hybrid No.8 , NTU8)。接種前以 Real-time PCR 測量病葉中病毒含量,再以緩衝液將兩系統病毒量分別稀釋至相同濃度,同 一品系共六種處理,分別是 Mock(健康對照組)、DF /-(單獨接種 DF)、DF / SMN
(先接 DF 二週後再接 SMN)、SMN /-(單獨接種 SMN)、SMN / DF(先接 SMN 二週後再接 DF)、及 DF+SMN(同時接種 DF 與 SMN)。每個處理三重複,接種 葉片為第三片、第四片及第五片本葉,接種方式參照前述“木瓜病毒之來源與接種
“,接種後每隔七天取系統葉(離莖頂最近之完全展開葉),作為核酸抽取之樣品,
拍照記錄病徵及病勢發展,詴驗時間為 12 週,接種日為 2011 年 9 月 6 日。
(3) 病徵分級
記錄之病徵分為 5 級:0 級,植株健康;1 級,30%系統葉出現輕微嵌紋、葉 脈透化等現象;2 級,70%系統葉出現輕微嵌紋、葉脈透化等現象;3 級,系統葉 出現明顯嵌紋、葉脈透化及黃化現象;4 級,葉片出現嚴重嵌紋、壞疽斑、捲曲、
畸形等病徵;5 級,葉片出現嚴重嵌紋、葉脈透化、捲曲、畸形、嚴重黃化或植 株死亡。
五、PRSV 之偵測
本實驗使用單一步驟的反轉錄聚合酶鏈鎖反應(one–step Reverse
Transcription –Polymerase Chain Reaction;one–step RT–PCR),該反應可同時將 RNA 反轉錄成 cDNA 並進行 PCR 反應。反應總體積為 25 μl,其成份最終濃度為:45 mM Tris–HCl(pH 8.3), 87.5 mM KCl, 4 mM MgCl2, 5 mM 1,4–Dithiotgreitol(DTT), 0.2 mM dNTPs(dATP, dTTP, dGTP, dCTP), 0.4 nM Forward primer, 0.4 nM Reverse primer, 0.25 units of Taq Polymerase(Super–Therm DNA Polymerase), 50 units of Reverse Transcriptase (Superscript®II, InvitrogenTM), 200 ng RNA template. 溫度循 環條件依序為:50℃,35 分鐘;94℃,2 分鐘;10 個循環〔94℃,30 秒;56℃,
30 秒;68℃,45 秒〕;25 個循環〔94℃,30 秒;56℃,30 秒;68℃,45 秒*
(*表示每個循環增加五秒)〕;68℃,7 分鐘。使用之引子對為實驗室所設計的 通用型引子對(PRSV–476)與系統專一性引子對(PRSV–DF598,PRSV–SMN467)
(李,2006)。增幅產物以 1.4 % Agarose 膠體進行電泳分析。
取 10 μl RT–PCR 產物加入 2 μl 6X Loading Dye 混勻,以 1.4% Agarose Gel 作 為膠體,電泳緩衝液為 TAE buffer(40 mM Tris–acetate; 1 mM EDTA,pH 8.0),
電泳以 100 V 進行 30 分鐘並以 100 bp DNA Ladder(Basic Life, Taiwan)作為產 物分子量大小判定標準。完成後以 Ethidium Bromide 染色,再使用電泳影象處裡 系統(Alpha Imagener)觀察拍照。此偵測方法修改自文獻(Hung et al, 2000)
六、即時定量聚合酶連鎖反應 (1) RNA 反轉錄
首先以 NanoDrop® ND-1000 Spectrophotometer(Thermo Fisher Scientific Inc., USA)測量樣品中 RNA 總濃度,再以 ddH2O 稀釋至 500 ng/μl,利用 PRSV-R829 primer 將稀釋後的 RNA 樣品反轉錄成 cDNA。總反應過程如下:取 4 μl 1 pmole Reverse primer(R829), 2 μl 2.5 mM dNTPs(dATPs, dTTPs, dCTPs, and dGTPs), 7 μl 500ng/μl RNA template 於 PCR 反應管中,65℃下作用 5 分鐘,完成後冰浴,再 加入 4 μl 5X RT buffer, 2 μl 0.1M DTT(1,4-Dithiothreitol), 1 μl reverse transcriptase
(200 U/ μl M-MLV, InvitrogenTM),42℃下作用 50 分鐘,得到互補 DNA(cDNA), 以 ddH2O 將 cDNA 稀釋 25 倍(6 μl cDNA+144 μl ddH2O),作為 real-time PCR 之 模板。
(2) 標準品與標準曲線之建立
將反轉錄後得到的 cDNA 以 R829 / F5end primer pair 進行 PCR 反應〔溫度循 環條件:94℃,3 分鐘; 30 個循環〔94℃,1 分鐘;60℃,1 分鐘;72℃,2 分 鐘〕;72℃,10 分鐘〕,PCR 產物轉殖至 TOPO TA cloning® Vector(pCR® 2.1-
TOPO®, InvitrogenTM)中,將 4 μl PCR 產物與 1 μl salt solution, 1 μl pCR®2.1-
TOPO® vector 混勻,室溫下作用 30 分鐘,再加入到大腸桿菌勝任細胞(JM109)
中混合,以 42℃水浴 heat shock 45 秒鐘,置於冰上 5 分鐘,預先將 40 μl X-gal
(40 mg X-gal in DMF)與 40 μl 100 mM IPTG 塗佈在含有 50 μg/ml ampicillin 的 LB 固態培養基上,再將已轉殖好的大腸桿菌,以 4℃預冷的三角玻棒均勻塗在 LB 固態培養基上,在 37℃下培養 8-12 小時。挑選白色菌落至新的 LB 固態培養基 上繼代培養,或挑至 1 ml 含 50 μg/ml ampicillin 的 LB 液態培養基中,於 37℃下 培養 12 小時,培養好之菌液以 Plasmid miniprep purification Kit(Protech
Gene-spinTM)抽取質體 DNA〔步驟如下:室溫下將 1 ml 菌液以 12000 rpm 離心 1 分鐘,去除上清液後加入 200 μl solution I 懸浮菌塊,再加入 200 μl Solution II 混 合,5 分鐘後再加入 300 μl solution III 並輕輕混勻,以 12000 rpm 離心 5 分鐘,將 上層液移至 spin colunm 中再以 12000 rpm 離心 1 分鐘,去除濾液,加入 600 μl Wash
Buffer 再以 12000 rpm 離心 1 分鐘,重覆此步驟一次,再以 12000 rpm 離心 3 分鐘 去除殘留液體,將 spin colunm 移至新離心管中,加入 50 μl TE buffer,靜置 5 分 鐘後以 12000 rpm 離心 3 分鐘,即可得到純化質體 DNA。〕。質體 DNA 以 R829/F5end primer pair 進行 PCR 反應,確認有將目標片段轉入質體 DNA 中,以 ABI 3730 自 動化核酸螢光定序儀(Applied Biosystems International, CA)進行核酸序列分析。
質體 DNA 經限制酶酵素─BamHI 切割,得到線形質體 DNA,純化後使用 mMESSAGE mMACHINE® Kit(Ambion)以質體上 T7 promoter 進行 in vitro transcription ,得到目標片段的 RNA,並以 NanoDrop ND-1000 Spectrophotometer
(Thermo Fisher Scientific Inc., USA)測量濃度,將 RNA 稀釋成 334 ng/μl,再使 用 reverse primer(R829)反轉錄成 cDNA,稀釋 25 倍後,再進行序列稀釋,即為 上機之標準品,機器會以標準品之 Ct 值與 Copy 數作圖得標準曲線。
(3) Taqman Primer / Probe 之設計
利用本實驗室前人所設計之通用型 (楊,2008)與專一性(李,2006)TaqMan® Primer / Probe 進行偵測,其內容物有 18 M forward primer, 18 M reverse primer, 5 M TaqMan® MGB probes(FAMTM dye-labeled)(Applied Biosystems International, ABI, USA)序列如表一。
(4) Real-time RT-PCR 反應
反應條件參考 ABI 公司之標準方法並稍加修改:50℃,2 分鐘;95℃,10 分 鐘;40 個循環〔95℃,15 秒;50℃,1 分鐘〕。反應總體積為 20 μl,包含 10 μl 2X TaqMan® Gene Expression Master Mix, 1 μl 20X TaqMan® Assay Mix(900 nM Forward primer、900 nM Reverse primer 及 250 nM TaqMan® MGB probes),及 9 μl 稀釋 cDNA。
肆、結果
一、不同木瓜輪點病毒系統在不同木瓜品種上病徵表現
木瓜輪點病毒畸形系統(PRSV-DF)接種在新的木瓜雜交品系(10A, 10B 10F)
時,莖部在接種後兩週出現水浸狀斑點,但上位葉在接種後十週內無病徵出現(圖 一),台農二號(TN2)、台農一號(TN1)、紅妃(RL)等對照組品系,出現程度 不一的病徵,接種後二週植株莖部產生水浸狀斑點,四週後 TN2 與 TN1 的上位葉 有輕微嵌紋與葉脈綠化等病徵,RL 的上位葉在接種後六週才出現輕微嵌紋的病徵,
十週後仍是輕微嵌紋(圖一)。木瓜輪點病毒嚴重嵌紋壞疽系統(PRSV-SMN)接 種在新雜交品系(10A, 10B, 10F)時,莖部兩週後出現水浸狀斑點,上位葉產生輕 微嵌紋,四週後轉成斑駁與葉脈透化,八週後 10A 與 10B 品系葉片尖端扭曲或呈 絲狀,部份葉片有嵌紋病徵,10F 品系的葉脈週圍葉肉組織白化,十週後三新品系
葉片褪色且嚴重嵌紋(圖二),老葉黃化掉落。對照組在接種後二週,莖部有水浸
狀斑點,葉片尚無異狀,三週後葉片有輕微嵌紋病徵,四週後葉片黃化與嵌紋,八 週後葉片嚴重嵌紋,以 TN1 最嚴重,十週後,葉片嚴重嵌紋,新生葉小且黃化(圖 二),TN2 葉扭曲,葉脈白化周圍有綠島,台農二號葉片黃化褪色且變形,RL 病 徵較前述兩品種輕微,但仍有斑駁與葉脈白化等病徵。三新品系接種 DF 系統或 SMN 系統後不產生病徵或出現輕微病徵,抗病表現均比對照組商業品種佳。
將上述病徵進行分級,接種 PRSV-DF 的新雜交品系(10A, 10B, 10F)病徵等 級均為零,顯示三者均未產生明顯病徵;對照組病徵由輕微到嚴重依序為:RL(病 徵等級為 2)、TN1(病徵等級為 2)、TN2(病徵等級為 2.33);接種 PRSV-SMN 後,新雜交品系中以 10A 病徵最輕微(病徵等級為 2)、10F 最嚴重(病徵等級為 2.67),對照組依其嚴重程度依序為 RL(病徵等級為 3)、TN1(病徵等級為 3)及 TN2(病徵等級為 3.67)(表二)。
二、不同木瓜輪點病毒系統在不同木瓜品種上病毒增殖情形
RT-PCR 偵測結果如表三,電泳分離產物,以軟體(Alpha Inotech)比較條帶 亮度再給予訊號強度。病毒接種兩週後,所有品種均可以 RT-PCR 偵測到病毒的訊 號,但在四週後才有強度明顯的增加,持續至十週強度仍無變化。
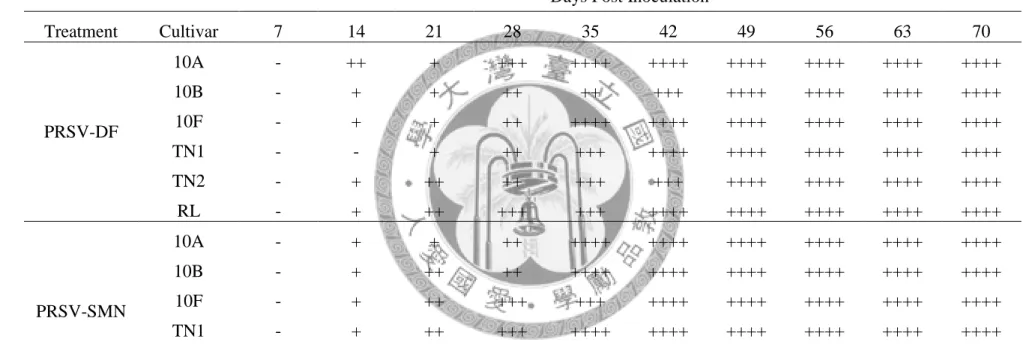
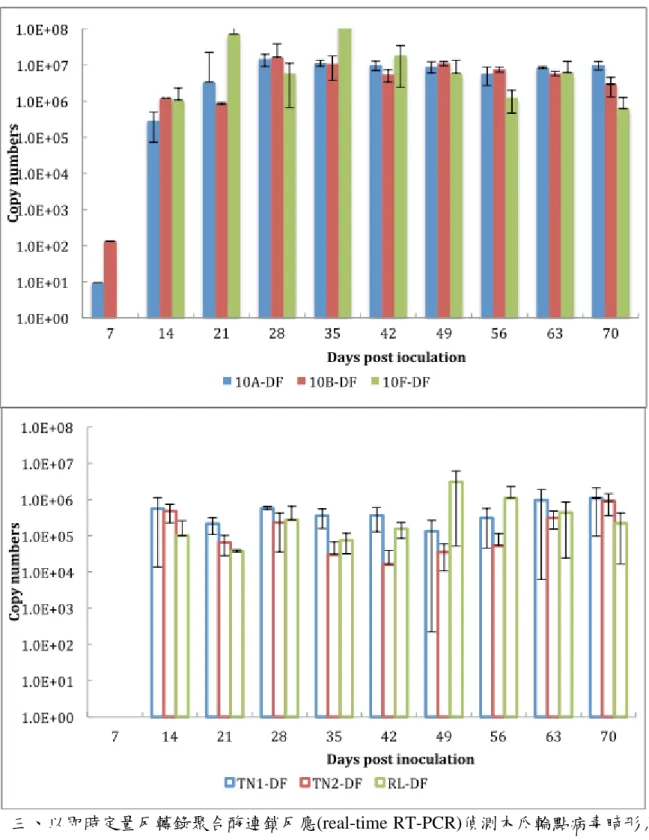
木瓜寄主接種 PRSV-DF 後,以 real-time RT-PCR 進行定量追蹤,發現六個品 種(10A, 10B, 10F, TN1, TN2, RL)在兩週後可明顯偵測到大量病毒,約有 105 copy numbers(CNs),伴隨感染時間增加,新品系體內病毒量上升至 107 CNs,對照組 品系仍維持在 105 CNs。整個感染過程中,唯有 10F 在第三週與第五週達到最高病 毒量:108 CNs,10A 與 10B 體內病毒量則無太大波動,維持在 107 CNs,TN2 體 內病毒量是對照組中最少的,但接種後六週病毒量開始增加,從 2x104 CNs 上升至 十週後的高峰:9x106 CNs,病毒在 TN1 體內穩定維持在 105~106 CNs,而 RL 介 於兩者之間,接種四週後病毒量為 2.7x104 CNs,七週後達到高峰:106 CNs,十週 後降到 2.2x105 CNs(圖三)。在新品系體內的 DF 系統病毒量高於對照組約 10 倍,
TN1 體內病毒增殖穩定,TN2 體內病毒量高於 TN1 與 RL。
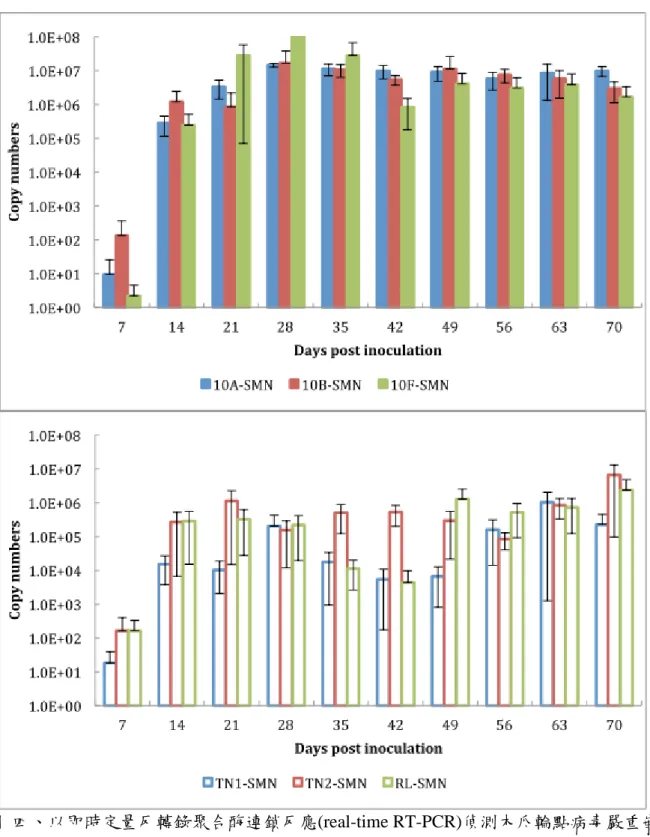
接種 PRSV-SMN 後,各品種皆在兩週後明顯偵測到病毒,病毒量為 105 CNs,
十週感染過程中,新品系的 10A 與 10B 體內病毒量維持在 106~107 CNs,10F 體 內病毒量有較大的變化,接種四週後上升至 108 CNs,隨後降至 5x105 CNs,在接 種八週後增加至 106 CNs;對照組的 TN1 體內病毒量在接種兩周後病毒量為 104 CNs,
接種四周後增殖至 2x105 CNs,六週時下降至 104 CNs,九週後上升至 CNs,TN2 體內的病毒量一直維持在 105~106 CNs,RL 體內的病毒量在接種後兩週為 2.8x105 CNs,五週時下降至 104 CNs,七週時增加到 1.2x106 CNs,穩定存在至十週(圖四)。 新品系體內 SMN 系統病毒量高於對照組品種,TN2 體內 SMN 系統病毒增殖情況 較另二商業品種佳。
三、非同步接種不同木瓜輪點病毒系在不同木瓜品種上病徵表現
單獨接種木瓜輪點病毒畸形系統的感染詴驗(DF / -)中,台大八號(NTU 8)
接種兩週後,莖部有水浸狀斑點出現,葉部無明顯病徵,四週後葉片斑駁、葉脈綠 化但葉子維持正常形態,八週後葉片嵌紋並產生綠島、末端扭曲且瘦長,十週後葉
片褪色、無畸形仍有綠島;TN2 接種兩週後,莖部有水浸狀斑,葉片斑駁,四週後 葉脈綠化、葉片變小、葉片變薄且較硬,八週後葉片畸形並朝上捲曲、葉肉組織上 有綠島,十週後葉子嚴重畸形、末端呈絲狀、葉肉細胞有綠島。RL 接種二週後,
莖部有水浸狀斑、葉片斑駁,四週後,葉脈綠化、葉片變薄且硬化,八週後葉片末 端產生綠島,十週後,葉肉細胞褪色且有綠島,末端呈絲狀,葉子變小但無畸形(圖 五)。
單獨接種木瓜輪點病毒嚴重嵌紋壞疽系統的感染詴驗(SMN / -)中,NTU 8 在接種二週後葉片斑駁,四週後葉片有明顯嵌紋與褪色情形,八週時葉片變小、褪 色、葉脈透化與嵌紋,十周後部份葉片轉為絲狀葉;TN2 在接種後二週,葉片褪色,
四週後嚴重嵌紋,葉片末端葉脈硬化,八週後葉片黃化、部份葉片絲狀化,完整的 葉肉細胞上有綠島,十週後新生葉呈絲狀,無葉肉細胞;RL 在接種後二週,葉脈 綠化、葉片斑駁,四週後葉片褪色、嚴重嵌紋等病徵,八週時葉片變小、褪色與嚴
重嵌紋、部份葉片呈絲狀葉,十週時新生葉為絲狀葉(圖六)。
先接種木瓜輪點病毒畸形系統,二週後再接種嚴重嵌紋壞疽系統的感染詴驗
(DF / SMN)中,NTU 8 接種二週時,莖部有水浸狀斑但葉片無病徵,四週後葉 片輕微嵌紋,八週後葉片褪色且帶有綠島,十週時葉片變小、褪色且產生綠島;
TN2 接種二週後,除了莖部水浸狀斑之外,葉片出現斑駁,四週後葉片嵌紋且畸形,
八週後葉片褪色並帶有綠島,十週時葉片變小、褪色,末端呈絲狀;RL 接種二週 後,莖部產生水浸狀斑點且葉片輕微嵌紋,四週後葉脈綠化、葉片褪色,八週後,
葉脈綠化、葉片褪色且有綠島,十週後葉片畸形、褪色、變小且末端呈絲狀,葉片 上有綠島(圖七)。
先接種木瓜輪點病毒嚴重嵌紋壞疽系統,二週後再接種畸形系統的感染詴驗
(SMN / DF)中,NTU 8 在接種後二週葉片斑駁,四週後葉片褪色、葉脈透化,
八週後葉片嚴重嵌紋、變小且部份呈絲狀,十週時,葉片變小、褪色與出現綠島;
TN2 在接種後二週葉片出現斑駁,四週後葉片褪色、變小、葉脈透化,八週後葉片 變小、嚴重嵌紋、部份葉片呈絲狀,十週時新生葉為絲狀葉;RL 在接種二週後葉 片黃化、嵌紋,四週後葉片嚴重嵌紋,八週後葉片變小與嚴重嵌紋,十週後葉片呈 絲狀(圖八)。
單獨接種 DF 系統後產生之病徵與先接種 DF 系統後接種 SMN 系統產生之病 徵相似,單獨接種 SMN 系統後產生之病徵與先接種 SMN 系統後接種 DF 系統產 生之病徵相似,先接種之病毒系統決定寄主之病徵發展。
四、同步接種不同木瓜輪點病毒系在不同木瓜品種上病徵表現
同時混合接種木瓜輪點病毒畸形系統與嚴重嵌紋壞疽系統的感染詴驗(DF+
SMN)中,NTU 8 在接種後兩週產生斑駁,四週後轉成嚴重嵌紋、葉脈透化,八 週時維持嚴重嵌紋且有綠島產生,十週時仍為嚴重嵌紋、綠島與葉脈透化,葉片大 小不變;TN2 二週後葉片褪色且斑駁,四週時葉片嚴重嵌紋且有綠島,葉脈周圍綠 化,八週時葉片畸形且有綠島,十週時葉片褪色、畸形與綠島,RL 在接種後二週,
葉片輕微嵌紋,四週後葉片褪色、嚴重嵌紋,八週後葉片嚴重嵌紋、褪色、畸形且 沿葉脈形成綠島,十週時葉片畸形、褪色情形較八週時嚴重(圖九)。同時混合接 種二病毒系統之病徵與單獨接種 DF 系統後產生之病徵類似。
五、非同步接種不同木瓜輪點病毒系在不同木瓜品種上病毒增殖動態
非同步接種 PRSV-DF 與 PRSV-SMN 的感染詴驗(先接種 DF / 再接種 SMN)
中,台大八號體內之 DF 系統病毒量,二週後為 106 CNs,第八週上升至 107 CNs 之後再降至 5x106 CNs;SMN 系統之病毒量在第四週為 102 CNs、第六週為 104 CNs、
第八週為 106隨後第九週降至 102 CNs,第十週增加至 103 CNs;單獨接種 DF 系統 之病毒量均維持在 106~107 CNs,單獨接種 SMN 系統之病毒量維持在 107~108 CNs
(圖十 A)。台農二號體內 DF 系統之病毒量在二週後為 106 CNs 並穩定存在 106~ 3x106 CNs 間,SMN 系統在第三週即出現,病毒量為 8x103 CNs,第四週無偵測到,
第五週至第七週約為 105 CNs,第八週無偵測到,第九週為 104 CNs,第十週為 103 CNs;單獨接種 DF 系統之病毒量十二週間於 7x105~6x106 CNs 間變動;單獨接種 SMN 系統之病毒量在第五週升至 106 CNs 並持續至第十二週(圖十 B)。紅妃體內 之 DF 系統病毒量二週後升至 106 CNs,第八週突增至 4x107 CNs,第九週降至 2.6x106 CNs,第十週升至 107 CNs;SMN 系統之病毒量在第三週為 105 CNs,第四 週無偵測到,第五至第七週為 105 CNs,第八週升至 5x105 CNs,第九週降至 104 CNs,
第十週再升至 5x105 CNs,第十二週無法偵測到;單獨接種 DF 系統之病毒量在 106~107 CNs 間變動;單獨接種 SMN 系統之病毒量再第四週為 3x104 CNs,第六週 升至 106 CNs 並維持至第十二週(圖十 C)。後接種的 SMN 系統病毒增殖量與先接 種 DF 系統病毒增殖量有百倍以上差距。
非同步接種 PRSV 兩系統(先接種 SMN /再接種 DF)的感染詴驗中,台大八 號體內 SMN 系統之病毒量在第二週為 105 CNs,第三週後上升至 106 CNs 直至第八 週為 107 CNs,第十週再下降為 4x106 CNs,DF 系統之病毒量在第四週為 6x104 CNs 隔週降至 103 CNs,第六週升至 105 CNs 後隔週再減低為 104 CNs,第九週升至 104 CNs,隔週達最高點 7x105 CNs,單獨接種 DF 系統之病毒量在 105~106 CNs 間變動 最高是第七週的 107 CNs,單獨接種 SMN 系統之病毒量在 105~106 CNs 間變動(圖 十一 A)。台農二號體內 SMN 系統之病毒量,第二週為 105 CNs,第三週後增殖為 107 CNs,直至第八週均在 5x106~107 CNs 間變動,第九週後降至 106 CNs;DF 系 統病毒量在在第三週即偵測到少許病毒量,但第五週與第七週無法測得,第九週為 104 CNs 隔週升上 105 CNs,第十二又降至 104 CNs;單獨接種 SMN 系統之病毒量 均在 4x106~107 CNs 間變動;單獨接種 DF 系統之病毒量在 104 ~106 CN 間變動(圖 十一 B)。紅妃體內 SMN 系統病毒量於第二週增殖至 105 CNs,第四週升至 106 CNs 維持至第九週再上升至 107 CNs,第十週降至 106 CNs;DF 系統病毒量從第三週至 第八週均維持在 102~6x103 CNs,第九週突增至 105 CNs 直至第十二週;單獨接種 SMN 系統之病毒量保持在 106~2x107 CN 間;單獨接種 DF 系統之病毒量在 106 ~107 CNs 間變動(圖十一 C)。後進入之 DF 系統病毒量會逐漸趕上先接種之 SMN 系統 病毒量。
六、同步接種不同木瓜輪點病毒系在不同木瓜品種上病毒增殖動態
同步接種 PRSV-DF 與 PRSV-SMN 的感染詴驗(DF+SMN)中,台大八號體內 SMN 系統的第一週病毒量為 104 CNs,第二週上升至 106 CNs 且維持到第六週,第八週 時降至 5x105 CNs,第九週再增加至 106 CNs,第十與十二週降至 105 CNs;DF 系 統在接種後一週達到 5x105 CNs,第四週時上升至 106 CNs,第七週時增加至 107 CNs,
至十二週期間病毒量在 5x106~107 CNs 間變化(圖十二 A)。台農二號體內 SMN 系 統病毒量在第一週為 7x104 CNs,第二週上升至 5x105 CNs 並維持至第八週,第九 週後無法偵測到病毒量;DF 系統首週為 3x105 CNs,第二週增加至 106 CNs 且維持 至第五週,第六週增殖為 107 CNs 直至第十二週(圖十二 B)。紅妃體內 SMN 系統 病毒量在第一週為 4x103 CNs,第二週升至 106 CNs 後至第九週均維持在 5x105~106 CNs,第十週下降為 105 CNs,第十二週則偵測不到病毒量;DF 系統首週病毒量為 8x102 CNs,第二週升至 106 CNs,第八週增高為 107 CNs,第九週反降至 106 CNs,
第十週與第十二週再回到 107 CNs(圖十二 C)。同時接種兩病毒系統五週後,DF 系統病毒量開始領先 SMN 系統病毒量。
伍、討論
木瓜輪點病毒(PRSV)是台灣木瓜產業一大限制因子,被害果樹由多年生轉為一 年生,產量減少且品質下降,嚴重影響木瓜栽培產業。台灣發展出網室栽培避免 PRSV 之危害,雖然有效防範蚜蟲傳病,同時也提高栽培成本,影響農民獲益。美國夏威夷與 台灣均已成功研發出基因轉殖抗 PRSV 之木瓜(包,2000;包等,2006;龔,2004),
但基因改造的農產品仍有風險疑慮,各國接受程度不一,而我國目前仍限於法令遲遲無 法推廣栽種。2009 年以傳統的抗病育種方法育成的台大八號(NTU 8)抗 PRSV 木瓜正式 通過國內的品種命名(王,1997;王,2001; 郭,2001),證明基因轉殖並非唯一抗 PRSV 的育種方式。然而,PRSV 進入台灣近 35 年,過去多次調查顯示田間系統複雜多 變(翁,1981;卲井,1986;林,1980; Lin et al, 1989;關,1990;廖,2004),十年前 與現今流行系統相差極大,危害程度不同。倘若要應用抗病育種來防治 PRSV,需探討 抗(耐)病品系在複雜 PRSV 系統的感染下是否仍具備持久的抗耐病能力,或瞭解 PRSV 系統間之交互作用關係,將有助於預測田間系統間消長趨勢,並評估新品種的抗病穩定 性。
三新品系(10A, 10B 與 10F)接種 PRSV 畸形系統(DF)後,皆無明顯病徵產生,
對照組品種則是出現輕微嵌紋等病徵。顯示三品系對 DF 系統有很好的抗病能力;三新 品系接種 PRSV 嚴重嵌紋壞疽系統(SMN)後出現嵌紋,對照組品種則是出現嚴重嵌 紋與黃化等病徵,表示新品系抗 SMN 系統的能力比對照組品種好,而在三新品系中又 以 10A 品系對 SMN 的病徵最輕微,因此在本詴驗中,10A 品系視為有潛力的耐病品種,
且接種後八週會耐病與感病品種之病徵會出現明顯差異,故第十週或更早的第八週皆可 視為病徵判讀的適合時間點。
三新品系接種 DF 系統後,病毒量在三週後均為 107 CNs,對照組則是只有 106 CNs,
新品系木瓜體內病毒量高於對照組 10 倍但無病徵產生,表示病毒在新品系中增殖較佳,
但不表現病徵,皆可視為對 PRSV-DF 的耐病品系,對照組台農一號與台農二號木瓜體 內病毒量無差異,病徵指數則以台農二號較嚴重,表示台農二號比台農一號感病,但 PRSV-DF 對兩者的親合性卻是相同。三品系接種 SMN 系統三週後病毒量為 107 CNs,
至第十週間病毒量在 106~107 CNs 間變動,對照組品種病毒量常在 106 CNs 以下,尤其
是台農一號和紅妃早期低於 104 CNs 後期才爬升至 106 CNs,台農二號體內病毒量保持 在 105~106 CNs 間,表示三個對照組仍以台農二號對 SMN 系的親合性最佳且病徵指數 最高,再次證實台農二號為感病品種的特性,即栽種台農二號對 PRSV-SMN 系統增殖 較有利;由新品系與對照組結果,病毒增殖量差十倍以上,仍需以統計方法證明是否有 差異,另外病徵嚴重度與病毒增殖情形無明顯直接關係,與前人結果(Gomez et al.,2009)
相同。本次詴驗之新品系為 F2 子代,未來可挑戰接種其親本與 F1 子代比較抗病能力,
辨明此耐病能力是由父本或母本繼承而來,爾後作為雜交耐病育種之親本,應用於未來 育種與選拔新耐病品系。將雜交後子代進行挑戰接種,十週後觀察病株株高、所產生之 病徵與體內病毒增殖情形,再與對照組比較結果,若病株生長勢同健康株、病毒量高於 感病品種且病徵比感病品種輕微,即篩選出有潛力之耐病品系;由同步與非同步接種之 病徵,可知兩系統間並無協力或拮抗等情形,因此挑戰接種時分別接種單一病毒系統即 可;至於進行定量 PCR 時的詴驗植株數目,利用統計方法訂出所需樣本數,取樣時間 點則是十週內取三個得知趨勢與最高增殖量。
不論木瓜寄主是否為耐病或是感病,本論文的詴驗結果發現先接種 DF、再接種 SMN,其病徵的展現與單獨接種 DF 者相同;若先接種 SMN、再接種 DF,其病徵的展 現則與單獨接種 SMN 者相同;同步混合接種二系統之病徵與單獨接種 DF 系統之病徵 類似但程度輕微,此結果與先人研究(Gomez et al.,2009)有相同趨勢。因此在不同步
(一前一後)的複合接種詴驗中,寄主病徵表現決定於首先接種的病毒系統,同時混合 接種 DF 與 SMN 之結果顯示病徵發展上 DF 比 SMN 強勢,此結果與前人研究(Chen et al, 2004)報告中指出多病毒系統複合感染後,最終其中一系統會成為優勢族群有相同 結果。此部份病徵結果與挑戰接種病徵結果,嚴重度差異頗大,且詴驗中台大八號比田 間感染結果嚴重,研判是受到栽培環境影響,包括光度不足、溫度過低、養份不足與盆 缽限制,使得植株在不正常狀態下生長,病徵因而嚴重許多。
非同步接種詴驗(先接種 DF 後接種 SMN)中,病毒在不同木瓜寄主中增殖情形 不同。台大八號體內後接種的 SMN 系統的進入後,不影響原先 DF 系統的病毒量,且 SMN 系統不易被偵測到且量遠少於 DF 系統;但單獨接種 SMN 病毒飽和量達 7x107 CNs 比單獨接種 DF 高十倍,表示 SMN 系統對台大八號體內增殖能力強,此結果與前人研 究(梁,2009)相似;台農二號的結果與台大八號結果相似,但單獨接種 DF 病毒飽和
量比單獨接種 SMN 高;紅妃體內 DF 系統與 SMN 系統有相同的變動情形,但 SMN 病 毒量低於 DF 一百倍。上述結果顯示,後進入的病毒系統難以取代已存在的病毒系統,
系統間沒有明顯的協力或拮抗的情形,這與前人研究同屬 Potyvirus 的 Plum pox virus 兩 系統的交互作用獲得相近之結果(Capote et al, 2006)。
非同步接種詴驗(先接種 SMN 後接種 DF)中,台大八號體內 DF 系統病毒量初期 變動很大,後期與 SMN 系統病毒量差異不大,有後來居上的情形;台農二號體內的 DF 系統增殖至 104 CNs 後 SMN 系統降至 6x105 CNs;紅妃和台農二號結果相同,但 DF 病毒量會增至 105 CNs 且 SMN 降為 106 CNs,二系統差距較台農二號小,表示台農 二號與紅妃體內二系統間彼此似有干擾作用。
同步混合接種二病毒系統,台大八號與紅妃體內二病毒系統病毒量一開始互有領先,
直至第五週後 DF 系統拉開差距,取得優勢地位,紅妃則在第十二週後就無法測到 SMN 系統;台農二號體內一直都是 DF 病毒量高於 SMN,差距在 10 倍以內,直至第七週才 明顯領先 SMN 100 倍,第九週即無法測到 SMN。二系統總合病毒量不會高於單獨接種 其中一個系統,顯示寄主體內可容納的病毒量有所限制。DF 系統與 SMN 系統二者間 為競爭關係,DF 系統在感染後期取得優勢,與 SMN 系統病毒量有極大差距,同時病 徵表現類似單獨接種 DF 系統,與病毒量之結果相符合,意即 DF 系統是強勢系統。DF 將主導病徵發展與族群優勢,因此 DF 系統成為田間優勢系統。
綜合以上結論,非同步感染時,先接種的系統有優勢,同步獲毒時 DF 系統有優勢,
Desbiez 報告(Desbiez et al. 2009)中提到,優勢的病毒系統會在數年內取代原先病毒 系統,本研究中 DF 系統為強勢病毒系統,但 SMN 多年來仍持續存在,未來 SMN 究 竟會逐漸消逝?或與 DF 繼續共存甚至演化成強勢系統?值得未來再予追蹤。在目前 DF 仍處強勢情況下,以台大八號木瓜為例,它對 DF 的耐病性明顯優於 SMN,而且 DF 在其植株體內增殖情況良好的同時病徵輕微,可視為台大八號的弱毒系統,依本論 文的詴驗結果推論,在 DF 保持優勢的情況下,即使台大八號受 SMN 感染,仍可保護 台大八號不產生嚴重病徵,因此其耐病性將可穩定持續。
PRSV 寄主除了番木瓜外,葫蘆科(Cucurbitaceae)作物是重要中間寄主、替代寄 主與接種源,預測田間病毒系統之變化需考慮葫蘆科作物複合感染情形,包括胡瓜嵌紋 病毒(Cucumber mosaic virus, CMV)、西瓜嵌紋病毒(Watermelon mosaic virus, WMV)、
矮南瓜黃化嵌紋病毒(Zucchini yellow mosaic virus, ZYMV),CMV 與 Potyvirus 間有協 力作用(Wang et al., 2002, Wang et al., 2004, Zeng et al., 2007),WMV、ZYMV、PRSV 同屬於 Potyvirus,彼此間交互作用尚不清楚,但可能會影響 PRSV 二系統之生態消長,
這方面值得未來做進一步之研究。
本研究以 real-time RT-PCR 作為分析病毒增殖之工具,綜合病徵發展與病毒增情形 二項資訊,在新耐(抗)病品系之選拔過程中,提供更詳實的評良參考。同步與非同步 的接種詴驗,探討不同病毒系統的交互作用關係,長時間觀察病徵與病毒量,測詴新品 種(如台大八號)耐病性之穩定,探討系統間之交互作用與族群趨勢,藉以預測田間病 毒系統之變化,研擬新防治策略並持續助益新耐病品種之開發。