Characterization and Immunomodulatory Activity of Polysaccharides Derived from Dendrobium tosaense
Li-Chan Yanga, Ting-Jang Lub, Chang-Chi Hsiehc, Wen-Chuan Lina*
aSchool of Medicine, Department of Pharmacology , China Medical University, Taichung, Taiwan b Institute of Food Science and technology, National Taiwan University, Taipei, Taiwan
c Department of Animal Science and Biotechnology, Tunghai University, Taichung, Taiwan
*Corresponding author: Department of Pharmacology, China Medical University, No. 91 Hsueh Shih Road, Taichung, Taiwan, R.O.C. Tel +886 4 22053366; fax +886 4 22053764
e-mail address: [email protected] (W.C. Lin)
Keywords:
Glucomannan, Innate immune, Dendrobium tosaense, Natural killer cells, Macrophages, Cytotoxicity, Phagocytosis
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Abstract
Dendrobium tosaense is a medicinal Dendrobium species widely used in traditional medicine. This study demonstrated some structural characterizations and immunomodulatory activity of the water-soluble polysaccharides derived from the stem of D. tosaense (DTP). DTP was fractioned using DEAE-650M anion-exchange gel filtration chromatography, producing one neutral polysaccharide fraction (DTP-N), which was investigated for its structural characteristics, using HPAEC-PAD, HP-SEC, GC-MS, and NMR spectroscopy. DTP and DTP-N consisted of galactose, glucose, and mannose in ratios of 1:9.1:150.7 and 1:12.2:262.5, respectively. DTP-N comprised (1→4)Man as its main backbone, and its average molecular weight was 220 kDa. We also investigated the immunomodulatory effects of DTP administered orally to BALB/c mice for 3 weeks. DTP substantially boosted the population of splenic natural killer (NK) cells, NK cytotoxicity, macrophage phagocytosis, and cytokine induction in splenocytes. This is the first study to demonstrate the structural characteristics of an active polysaccharide derived from D. tosaense and its immunopharmacological effects in vivo.
17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33
1. Introduction
Herba dendrobii (Shi-Hu), a medical orchid, is widely used in Asia and is prepared from several Dendrobium species. Herba dendrobii is used in both traditional Chinese and folk medicine for antipyretic, ophthalmic, and tonic purposes (Wu et al., 2013). Dendrobium tosaense is one of medicinal Dendrobium species present in Taiwan (Wu et al., 2013). In contrast to other herba dendrobii species, such as D. huoshanese and D. nobile, which are mainly distributed in China, D. tosaense is native to Taiwan and is mass produced through tissue cultivation (Ding, Xu, Wang, Xu & Zhou, 2002; Lo, Nalawade, Kuo, Chen & Tsay, 2004). Several studies have reported that quercetin is one of the bioactive compounds of D. tosaense, which exhibits antioxidant, antihypertensive, antiinflammatory, and anticarcinogenic activities (Chiang et al., 2012; Lo, Mulabagal, Chen, Kuo & Tsay, 2004). However, there are few reports on the polysaccharides of D. tosaense, even though the polysaccharides from other Dendrobium species have been thoroughly studied. The polysaccharides of other species of herba dendrobii are regarded as valuable compounds exhibiting pharmacological activities (Ng et al., 2012). Many studies have reported the structural characteristics of polysaccharides from other Dendrobium species, as well as their pharmacological properties, which include antitumor, antihyperglycemic, antioxidant, and immunomodulatory activities (Wang, Luo, Zha & Feng, 2010; Zha, Luo, Luo & Jiang, 2007; Zhao, Son, Kim, Jang & Lee, 2007).. However, no pharmacological studies on the polysaccharides of D. tosaense have been reported thus far.
Immunomodulatory agents can stimulate immune responses, and maintain the immune balance (Volman, Ramakers & Plat, 2008). Numerous studies have reported that polysaccharides are ideal immune modulators. Immunomodulatory 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
polysaccharides can not only improve host defense against pathogens, but also modulate adaptive immunity (Schepetkin & Quinn, 2006; Volman et al., 2008). The regulation of the levels of lymphocytes, cytokines, chemokines, and antibodies are the results of polysaccharide immunomodulatory (Schepetkin & Quinn, 2006; Yuan et al., 2008; Zhao, Hu, Wang, Liu & Guo, 2013) .
In this study, we reported the structural characteristics of the indigestible and purified polysaccharides isolated from the stem of D. tosaense, and investigated the immunomodulatory effects of the polysaccharides of D. tosaense, administered orally in vivo.
2. Materials and methods
2.1. Preparation of Polysaccharides
2.1.1. Purification and fractionation of polysaccharides
The dry stem of D. tosaense was provided by the Taiwan Seed Improvement and Propagation Station (Taichung, Taiwan). The plant materials were extracted using hot distilled water, and then evaporated under reduced pressure. A 4-fold volume of 95% ethanol was added to the aqueous extracts to precipitate the crude polysaccharides. The crude polysaccharides were collected, and the proteins and starches were removed according to Method 991.43 of the Association of Official Analytical Chemists, using a total dietary fiber assay kit (Megazyme, Wicklow, Ireland). Briefly, the crude polysaccharides were sequentially digested by α-amylase, protease and amyloglucosidase. After enzymatic treatment, the indigestible polysaccharides were precipitated by adding a 4-fold volume of 95% ethanol at 4 °C, and allowing the 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85
solution to stand overnight. The indigestible polysaccharides of D. tosaense (DTP) were dissolved in hot distilled water and filtrated for additional analyses and experiments.
2.1.2. Characterization of indigestible polysaccharides and the neutral polysaccharide fraction (DTP-N)
DTP was fractionated using anion-exchange chromatography on a DEAE-650M column (Toyopearl, Tokyo, Japan). The DEAE column was eluted with 20 mM Tris-HCl (pH 7.8), followed by a sodium chloride gradient (0–0.3 M). The carbohydrate elution profile of DTP was analyzed according to the phenol-sulfuric acid method, with glucose as the standard, and measured at 490 nm, using a TRAID LT ELISA reader (Dynex Technology, VA, USA) (Dubois, Gilles, Hamilton, Rebers, & Smith, 1956). The relevant fractions were collected, concentrated, and precipitated by adding ethanol to get the DTP-N. For the chemical analysis, the protein content of the pooled polysaccharide fractions was measured using a modified Bradford method assay with bovine serum albumin as the standard (Compton & Jones, 1985). The carbohydrate content of the DTP and DTP-N was determined using the phenol-sulfuric acid method, as described previously. The content of uronic acid was determined using the m-hydroxydiphenyl method, with galacturonic acid as the standard (Blumenkrantz & Asboe-Hansen, 1973). For nuclear magnetic resonance (NMR) analysis, DTP-N (5 mg) were dissolved in deuterium oxide (0.5 mL), and 1H NMR spectra were recorded on a Bruker DRX-600 spectrometer (Bruker BioSpin, Billerica, MA, USA) at 20 °C, using 3-(trimethylsilyl)-propionic 2,2,3,3,-d4 acid sodium salt as an internal reference (δ 0.00 ppm). The relative molecular mass (Mr) was determined using high performance-size exclusion chromatography (HP-SEC), using a TSKgel PWH guard 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111
column, and PWH (75 × 7.5 mm i.d.; TOSOH, Tokyo, Japan), TSKgel G4000PWXL, and G2500PWXL (300 × 7.8 mm i.d., TOSOH) columns connected in series. The columns were eluted with 0.3 N of sodium nitrate solution containing 0.02% sodium azide, with a flow rate of 0.8 mL/min at 65 °C. Peaks were detected using an Interferometric Refractometer (Wyatt, CA, USA). The average Mr was estimated by comparison with the retention time of the pullulan standard P-82 kit (molecular weights of standards: P-800, 708 kDa; P-400, 375 kDa; P-200, 200 kDa; P-100, 107 kDa; P-50, 47.1 kDa; P-20, 21.1 kDa; P-10, 9.6 kDa; and P-5, 5.9 kDa; Shodex, Kawasaki, Japan). For monosaccharide composition analysis, samples were hydrolyzed with 2 M trifluoroacetic acid (Sigma-Aldrich, St. Louis, MO, USA) at 100 °C for 4 h, and the resulting samples were analyzed using high-performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD). The peaks were detected using a 817 Bioscan PAD detector (Metrohm, Zofingen, Switzerland) and a CarboPac PA1 column (Dionex, Sunnyvale, CA, USA). The eluent consisted of 10 mM sodium hydroxide and 1 mM barium acetate (Sigma-Aldrich), pumped at a flow rate of 1.0 mL/min. Linkage analysis was performed by methylation, as described by Liane et al. (Laine, Tamminen, Vikkula, & Vuorinen, 2002). Methylation was performed using methyl iodide (Sigma-Aldrich) in dimethyl sulfoxide (Sigma-Aldrich) and sodium hydroxide (Wako, Osaka, Japan). After methylation, samples were hydrolyzed with 2 M trifluoroacetate, followed by reduction and acetylation, using the alditol acetate method. Inositol was used as an internal control. GC-MS analysis of the partially methylated alditol acetates was conducted on a DB-5 column (0.2 μm i.d. × 30 m, OV-1701, Agilent), applying the following temperature program: maintain at 100 °C for 2 min, then increase the temperature to 180 °C at a rate of 8 °C/min, maintain at 180 °C for 2 min, then further increase to 240 °C at a rate of 2 °C/min, and maintain at 240 °C for 5 min. The carrier 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137
gas was helium (1.0 mL/min, constant flow). For the mass spectrometer, an ionization potential of 70 eV and source temperature of 300 °C was used.
2.2. Innate immunomodulatory of DTP in BALB/c mice
2.2.1. Animals
Eight-week-old male BALB/c mice were obtained from the National Laboratory Animal Center (Taipei, Taiwan). The experimental animals received appropriate care and humane treatment, and all the animal experimental protocols followed the institutional guidelines of China Medical University for the use of laboratory animals. The animals were housed in an air-conditioned room (21–24 °C) and received humane care under a 12-h light (8:00 a.m.–8:00 p.m.), and were allowed free access to food pellets and water throughout the study. The experiment was performed on 36 BALB/c mice having a body weight of 24–26 g. The mice were randomly divided into three groups, and were orally administrated distilled water (control) or DTP (100 and 300 mg/kg of body weight), every day for 6 weeks. The body weight of the mice was measured once a week until the end of experiment. At the end of the experiment, the animals were euthanized using CO2. The spleen was removed, and weighted under sterile conditions for splenocytes preparation. The peritoneal cells were prepared by applying the method of Zhang et al. (Zhang, Goncalves, & Mosser, 2008), and used for phagocytosis assays.
2.2.2. Flow cytometry analyses 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162
To prepare the splenocytes, the spleens from mice were removed aseptically, placed in cold Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco, NY, USA), and then teased apart and passed through a nylon mesh (BD Falcon, CA, USA). The spleen cell suspensions were hemolyzed using 0.1- and 2-M HBSS to lyse the red blood cells, and then added to 5 mL of RPMI 1640 medium containing 10% FBS, 2 mM L-glutamine (Gibco), 100 μM 2-mercaptoethanol (Gibco), 1.0 mM sodium pyruvate (Gibco), and gentamycin (Gibco). The single spleen cell suspensions were centrifuged at 4 °C and 300 g for 10 min prior to collection. The cell viability of splenocytes was determined using a FACSCalibur™ flow cytometer (Becton-Dickinson, CA, USA) with a propidium iodide (PI)/RNAase (Invitrogen) stain. All antibodies used for flow cytometry analyses were purchased from eBioscience (CA, USA). The cells (5 × 105 cells/25 μL) from splenocytes were stained with 10 μL of FITC-labeled antimouse CD4, PE-labeled CD8, PE-Cy5-labeled antimouse CD3, labeled antimouse CD19, PE-labeled CD3, PE-Cy5-labeled CD45, , and FITC-labeled antimouse Pan NK (CD49b) at 4 °C for 30 min in the dark. After incubation, unlabeled antibodies were washed with 3 mL of PBS containing 0.05% sodium azide, and then resuspended in 200 μL of FACS buffer containing 2.0% FBS and 0.05% sodium azide (Sigma-Aldrich, MO, USA). Cells were then analyzed using a FACscan, and the data were analyzed using CellQuest software (BD Biosciences). The results of flow cytometry were presented as the percentage of positive fluorescent cells.
2.2.3. Determination of splenocyte proliferation
Splenocytes were isolated from the mice in the control and DTP groups (100 and 300 mg/kg). The splenocytes were seeded into a 96-well plate at 5 × 106 cell/mL in 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188
100 μL of RPMI 1640 medium (Gibco), and stimulated with Con A (5 μg/mL), LPS (10 g/mL), or RPMI 1640 medium in a final volume of 200 L. After 48 h of incubation, splenocyte proliferation was determined using the MTS assay (Wang et al., 2010), and the plate was then read at 492 nm, using a TRAID LT ELISA reader. The stimulation index (SI) was calculated as the absorbance value of stimulated cultures, and divided by the absorbance value of nonstimulated cultures.
2.2.4. Cytotoxicity assay
YAC-1 lymphoma cells were obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). These cells were cultured in RPMI-1640 medium (Gibco) supplemented with 10% (v/v) fetal bovine serum (FBS, Gibco), 100 μg/mL of streptomycin, and 100 U/mL of penicillin (Gibco) at 37 °C in a humidified atmosphere containing 5% CO2. For the cytotoxicity assay, the YAC-1 (Target-T) cell line was incubated with BCECF-AM (Molecular Probe) at 37 °C for 30 min, agitating gently. BCECF-AM labeled YAC-1 cells were washed twice with RPMI-1640 medium. Splenocytes harvested from the mice treated with distilled water and DTP were used as Effector (E) cells in the cytotoxicity experiment. For the cytotoxicity assays of splenocytes, a previously described method (Kimura & Sumiyoshi, 2004) was modified. The splenocytes and BCECF-AM-labeled YAC-1 cells were placed in V-bottom 96-well plates at effector (E): target (T) ratios of 25:1; 50:1, and 100:1, and incubated for 4 h. These cell mixtures were then centrifuged at 410 g for 10 min. The total fluorescence intensity of the BCECF-labeled YAC-1 cells was determined after adding 1.0% Triton X-100 for lysing. The fluorescence intensity of the supernatant was measured using a TRAID LT ELISA reader with excitation at 485 nm and emission at 535 nm. The cytotoxicity activity of splenocytes was calculated as 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214
follows: percent of specific cytotoxicity = (fluorescence intensity of target cells treated with splenocytes isolated from the experimental group − fluorescence intensity of the spontaneous release of target cells)/(total fluorescence intensity of target − fluorescence intensity of the spontaneous release of target cells) × 100.
2.2.5. Phagocytosis
Peritoneal cells were isolated from the mice in each treated group. The populations of macrophages and neutrophils were estimated using flow cytometry determination with size (FCS-H) and granularity (SSC-H) coordinates. The isolated peritoneal cells (1 × 105 cells/well) and FITC-labeled Escherichia coli (BioParticles, OR, USA) were cocultured in round-bottom 96-well plates at 37 °C and 150 rpm for 2 h at multiplicities of infection (MOI) of 1:1, 1:5, or 1:25. After incubation, the peritoneal cells and bacteria were washed three times with PBS. Extracellular fluorescence was quenched with trypan blue (Gibco). Quenching prevented the interference of fluorescence from uningested bacteria. Fluorescence was determined using a TRAID LT ELISA reader at 485 nm excitation and 535 nm emission.
2.2.6. Analysis of cytokine production
Separated splenocytes were isolated from BALB/c mice in the control and DTP (100 and 300 mg/kg) groups. The splenocytes were seeded into 24 well-plates at a density of 2 × 106/mL, and stimulated with Con A (5 μg/mL), LPS (10 μg/mL), or RPMI 1640 medium for 24 and 48 h of incubation. After incubation, the supernatants were harvested and used for the cytokines assays (IL-2, IL-4, IL-6, and IFN-γ) with commercial ELISA kits (eBioscience, CA, USA).
215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240
2.3. Statistical analysis
The results were expressed as the mean ± SD. All experimental data were analyzed using a one-way analysis of variance, applying the Dunnett test. Values of P < 0.05 were considered to be statistically significant.
3. Results
3.1. Preparation and characterization of D. tosaense polysaccharides
The yield of the aqueous extracts from dry D. tosaense was 24.7%. The yield of crude polysaccharides and DTP were 17.4% and 9.7%, respectively, on a dry basis. DTP consisted of 97% carbohydrates and less than 1% protein. The content of uronic acid in DTP was 4.1% on a carbohydrate basis. The protein and uronic acid content in DTP-N was both undetectable. DTP was fractioned using the DEAE-650M column to obtain one major (neutral polysaccharide, 87%) and one minor (acid polysaccharides) fraction, which were collected according to the total carbohydrate elution profile (Fig. 1). The content of uronic acid was determined from the elution, and distributed only in the acid polysaccharide fractions. 1H NMR (500 MHz) revealed the structural characteristics of DTP-N (Fig. 2). The strong signal observed at 4.79 ppm corresponded to D2O, whereas the signals at 4.56 and 5.55 ppm were consistent with the presence of β- and α-anomeric protons, respectively. The signal at 4.16 ppm might correspond to mannose (Hsieh et al., 2008).
According to the calibration curves derived from the pullulan standards, the results of HP-SEC analysis revealed two peaks, indicating that the molecular weight distribution of DTP ranged from 15 kDa to 305 kDa (Fig. 3). By contrast, only one 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266
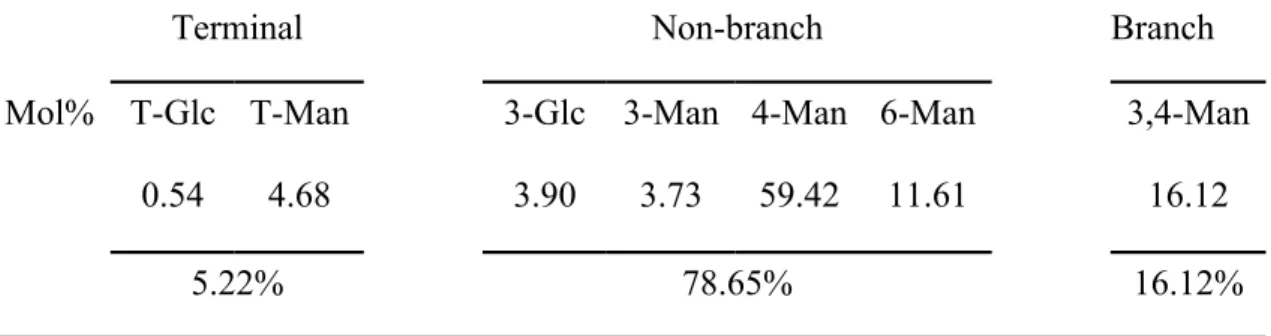
peak appeared in the molecular distribution of DTP-N. The average molecular weight of and DTP-N was 221 kDa. HPAEC-PAD analysis of the monosaccharide composition revealed that DTP was composed of galactose, glucose, and mannose at a ratio of 1:9.1:150.7. DTP-N was composed of galactose, glucose, and mannose at a ratio of 1:12.2:262.5. GC-MS analysis revealed the glycosidic linkage composition of DTP-N (Table 1). The results of GC-MS analysis indicated that the ratio of non-branch linkages to non-branched linkages was 4.9:1. The predominant linkage component in DTP-N was 4-mannose, and the other major component was 3,4-mannose.
3.2. Innate immunomodulatoryion of DTP in BALB/c mice
After treatment for 6 weeks, the body weight and spleen weight of mice exhibited no significant differences among groups. In control group, the body weight and spleen weight of mice were 26 ± 0.9 g and 0.14 ± 0.08 g, respectively.
3.2.1. Flow cytometry assays on splenocytes
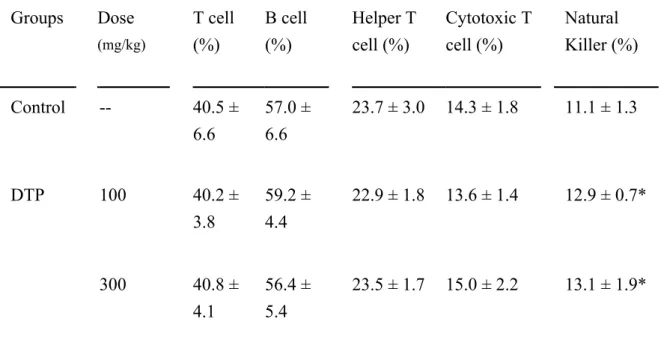
Flow cytometry analyses revealed the effects of DTP on the population of splenic T cells (CD45+CD3+), B cells (CD45+CD19+), T helpers (Th, CD3+CD4+), cytotoxic T cells (Tc, CD3+CD8+), and NK cells (CD49b+CD3-) and is showed in Table 2. The results revealed that neither the populations of splenic T cells or B cells nor their ratio was affected by DTP. Moreover, the Th and Tc populations exhibited no significant differences among groups. However, the population of splenic NK cells was substantially upregulated by DTP. Oral administration of DTP led to a 16.9% (100 mg/kg) and 18.2% (300 mg/kg) increase in the population of splenic NK cells, compared with the control group.
267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292
3.2.2. Splenocyte proliferation assays
The effects of DTP on the proliferation of splenocytes through mitogen (LPS or Con A) stimulation are displayed in Fig. 5. Splenocyte proliferation was substantially enhanced by LPS stimulation in a dose-dependent manner in both DTP-treated groups (100 and 300 mg/kg), compared with the control group. In addition, the DTP treatments (100 and 300 mg/kg) significantly upregulated the proliferation of splenocytes through Con A stimulation, compared with the control group.
3.2.3. Cytotoxic activity of DTP
Figure 5A showed the cytotoxic activities of splenocytes from mice treated with DTP for 3 weeks. No difference was observed in the cytotoxicity of splenocytes among the groups at an E/T ratio of 5. Mice treated with DTP exhibited an enhanced splenocyte cytotoxicity at an E/T ratio of 10. Moreover, DTP also promoted the cytotoxic activity at an E/T ratio of 25. At an E/T ratio of either 10 or 25, the cytotoxic activity was enhanced by DTP in a dose-dependent manner. However, the splenic NK cytotoxicity in the high-dose DTP treatment (300 mg/kg) exhibited significant differences only when compared with the control at E/T ratios of 10 and 25.
3.2.4. Effects of DTP on phagocytic activity
We investigated the phagocytosis of extracellular FITC-labeled E. coli, using peritoneal neutrophils and monocytes isolated from BALB/c mice treated with DTP. The phagocytic activity of DTP was observed at various MOI ratios. Figure 5B 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318
showed that phagocytic activity increased by DTP with an increase in engulfed pathogens. Moreover, DTP significantly improved the phagocytic activity compared with the control group under the conditions of MOI= 1, 5, and 25. These results demonstrated that the phagocytic activity was upregulated by either a low-dose administration of DTP (100 mg/kg) or a high dose (300 mg/kg), compared with the control.
3.2.5. Cytokine production
Figure 6 showed the cytokines production by the separated splenocytes, which were incubated with or without mitogen (ConA and LPS) from 24 to 72 h. After 48 h of incubation, the concentration of secreted IL-2 in the supernatants was determined. The results revealed that both DTP groups (100 and 300 mg/kg) exhibited an improvement in the secretion of IL-2 after Con A stimulation. IL-4 and IL-6 were collected in the supernatants after 72 and 24 h of incubation, respectively. However, the levels of IL-4 and IL-6 exhibited no significant differences among groups, with or without mitogen stimulation. The level of IFN-γ in the supernatants after 72 h of incubation demonstrated that secreted IFN-γ was significantly increased only after ConA stimulation. In addition, splenocytes isolated from DTP-treated mice secreted significantly more IFN-γ than did the control.
4. Discussion
Herba dendrobii has been used as a medicinal orchid in Asia since ancient times. Previous studies have identified the polysaccharides of herba dendrobii from several species other than D. tosaense, and these polysaccharides exhibited antioxidant, immunostimulating, and antitumor activities (Zhao et al., 2007). A major problem in 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344
the medicinal herb market is that the populations of several species of medicinal plants are rare and under protection. D. tosaense exhibited an excellent growth rate compared with other Dendrobium species, and a mass production process was recently established (Chiang et al., 2012). Therefore, this study investigated the structural characteristics and immunomodulatory properties of D. tosaense, and suggested that these properties might contribute to the therapeutic potential of this plant.
In this study, the polysaccharides were isolated from the stem of D. tosaense, and the primary structural characterization was achieved. Fractioning of DTP using a DEAE-650M column revealed that DTP was mainly composed of a neutral polysaccharide fraction (87%). Zha et al. (2007) reported that the neutral polysaccharide fraction represented 81% of the crude polysaccharides of D. huoshanense. Furthermore, previous studies (Hua, Zhang, Fu, Chen & Chan, 2004; Wang, Luo, Zha & Feng, 2010) reported that the neutral polysaccharide fraction represented 75% and 65.5% of the water-extracted polysaccharides of D. nobile and D. officinale, respectively. This result indicated that DTP-N represents the major portion of DTP, which might explain why the Mr of DTP and DTP-N were similar to each other. The monosaccharide composition analyses revealed that DTP and DTP-N were composed of galactose, glucose, and mannose in different ratios. Medicinal Dendrobium species exhibit various monosaccharide compositions, but that of D. huoshanense comprised galactose, mannose, and glucose, being the same as that of D. tosaense. Linkage analysis indicated that non-branched (1→4)-Man was the backbone of DTP-N. Hua et al. (2004) reported that the molar ratio of (1→4)-Man (the major backbone of D. officinale) was 26.1%. In addition, Hsieh et al. (2008) reported that the stem mucilage of D. huoshanense contains glucomannan in (1→4)-Glcp and β-(1→4)-Manp linkages. This evidence suggests that the existence of β-(1→4)-Man 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370
linkages might be common in medicinal Dendrobium species.
In traditional medicine, herba dendrobii is administered orally as an aqueous extract. Therefore, this study investigated the pharmacological effects of the oral administration of DTP to mice to elucidate its immunomodulatory effects. In this study, oral administration of DTP did not exert any effect on the spleen or body weight of mice. In addition, this study demonstrated the effects of DTP on the population of splenic lymphocytes; DTP boosted the population of splenic NK cells, but exerted no effects on the populations of T cells, B cells, cytotoxic T cells, or helper T cells. In contrast with the adaptive immunity of cytotoxic T cells, NK cells play a critical role in innate immunity, being involved in the first line of defense against various invasions (Lai, Wang, Chung, Hwang & Huang, 2011). NK cells display spontaneous cytotoxicity against pathogen-infected or malignant cells, without antigen-specific recognition (Lai et al., 2011). In our previous study, we reported that the anticancer activity of arabinogalactan derives from the stimulation of NK cells (Yang, Hsieh, Lu & Lin, 2013).
Moreover, DTP not only enhanced the population of splenic NK cells, but also promoted their cytotoxic activities. Generally, macromolecules such as polysaccharides are thought to be unable to be absorbed in the gastrointestinal tract. However, the oral administration of DTP for 3 weeks affected the population and cytotoxicity of splenic NK cells. A possible mechanism for this could be that orally-administrated DTP mediated interactions with microfold (M)-cells in the intestine, in the form of 1,3 β-glucans, which resulted in an immunomodulatory response (Suzuki et al., 1990). M cells are specialized epithelial cells specialized in transporting macromolecules in the Peyer's patches, which are involved in the intestinal immune system. Volman et al. (2008) suggested that orally-administrated polysaccharides might directly affect immunity after being taken by intestinal macrophages and 371 372 373 374 375 376 377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396
transported to the lymph nodes, spleen, and bone marrow.
In addition to the cytoxicity of NK cells, phagocytosis of macrophages is also a critical barrier for the innate immunity of host defense. Macrophages are crucial antigen-presenting cells (Schepetkin & Quinn, 2006). In the phagocytosis process, macrophages digest the pathogens and present the antigens of these pathogens to the T lymphocytes, representing a regulation of adaptive immunity (Schepetkin & Quinn, 2006). This study demonstrated that DTP promoted the phagocytic activity of macrophages isolated from mice. Based on the enhancement of the cytotoxicity of NK cells and phagocytosis of macrophages, we consider that DTP is a potent immunomodulator for innate immunity.
Even though the flow cytometry analyses showed that DTP did not influence the population of T cells or B cells of splenocytes, the splenocyte proliferation assays revealed that DTP is a mitogen for splenocytes. We investigated the mitogenic effect on splenocytes isolated from mice treated with DTP, as well as the comitogenic effect of DTP on ConA- or LPS-activated splenocytes. ConA and LPS are mitogens to T lymphocytes and B lymphocytes, respectively. Orally-administered DTP could promote the proliferation of separated splenocytes under the stimulation of mitogens. Oral administration of DTP increased the reaction levels of immune cells to mitogens. In this study, DTP stimulated the proliferation of T cells and B cells of splenocytes. T cells and B cells are involved in cellular and humoral immunity, respectively, playing a critical role in host defense (Volman et al., 2008). The ability to develop an effective T cell and B cell immunity can be regarded as a stimulation of the lymphocyte proliferation response. Our previous study revealed that a different polysaccharide, arabinogalactan, stimulated only the LPS-activated splenocytes (Yang et al., 2013). These results demonstrated that polysaccharides with different structural characteristics might affect the immune 397 398 399 400 401 402 403 404 405 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422
system through various mechanisms or pathways.
Several polysaccharides derived from plants have been reported to induce production of cytokines including tumor necrosis factor alpha (TNF-α), interleukin (IL)-1h, IL-6, IL-8, IL-12, IFN-γ, and IFN-β2 (Schepetkin & Quinn, 2006). Cytokine secretion is regarded as an immunomodulatory ability. In this study, the splenocytes isolated from DTP-treated mice were activated by LPS and ConA to investigate the secretion of cytokines such as IL-2, IL-4, IL-6, and IFN-γ. The induction of IL-2 and IFN-γ was significantly increased in the DTP group exhibiting ConA-activated splenocytes. IL-2 is part of the innate immunity, particularly against pathogen infection (Malek, 2008). IL-2 is essential for the growth, proliferation, and differentiation of T cells, and is produced by T cells normally during an immune response (Malek, 2008). ConA could trigger the activation and proliferation of T cells through interaction with their receptors (Palacios, 1982).. This study revealed that oral administration of DTP promoted the activation of T cells in splenocytes. However, the mechanism of IL-2 induction by DTP is not yet clear. IFN-γ, also called type II interferon, is a key cytokine against pathogen infections and tumors in innate and adaptive immunity. IFN-γ is mainly secreted by NK cells and T cells as part of innate immunity and antigen-specific immunity (Boehm, Klamp, Groot & Howard, 1997). One of the prominent characteristics of IFN-γ is its ability to activate macrophages (Herbst, Schaible & Schneider, 2011). The induction of IFN-γ in DTP-treated splenocytes might explain the potent cytotoxic and phagocytic activities of DTP reported here. The polysaccharides and aqueous extracts of D. officinale were reported to markedly increase IFN-γ production by murine splenocytes (Liu et al., 2011). Moreover, the polysaccharides of D. huoshanense were reported to significantly enhance the production of IFN-γ (Hsieh et al., 2008; Zha et al., 2007). The stimulation of IFN-γ secretion might be a common immunomodulatory 423 424 425 426 427 428 429 430 431 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447 448
characteristic in medicinal Dendrobium species.
In summary, the polysaccharides of D. tosaense were mainly composed of neutral polysaccharides. The major polysaccharide had a (1→4)Man backbone, and exhibited a potent immunomodulatory activity in mice, suggesting that this compound is a novel immunomodulatory agent.
Acknowledgements
This study was supported by grants from the Department of Health, Excutive Yuan of the Republic of China (CCMP 100-RD-108).
References
Blumenkrantz, N., & Asboe-Hansen, G. (1973). New method for quantitative determination of uronic acids. Analytical Biochemistry, 54, 484-489.
Boehm, U., Klamp, T., Groot, M., & Howard, J. C. (1997). Cellular responses to interferon-gamma. Annual Review of Immunology, 15, 749-795.
Chiang, C. H., Yu, T. A., Lo, S. F., Kuo, C. L., Peng, W. H., & Tsay, H. S. (2012). Molecular Authentication of Dendrobium Species by Multiplex Polymerase Chain Reaction and Amplification Refractory Mutation System Analysis. Journal of the American Society for Horticultural Science, 137, 438-444.
Compton, S. J., & Jones, C. G. (1985). Mechanism of dye response and interference in the Bradford protein assay. Analytical Biochemistry, 151, 369-374.
Ding, X. Y., Xu, L. S., Wang, Z. T., Xu, H., & Zhou, K. Y. (2002). Molecular authentication of Dendrobium chrysanthum from its allied species of Dendrobium. Zhongguo Zhong Yao Za Zhi, 27, 407-411.
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956).
Colorimetric Method for Determination of Sugars and Related Substances. Analytical Chemistry, 28, 350-356.
Herbst, S., Schaible, U. E., & Schneider, B. E. (2011). Interferon gamma activated macrophages kill mycobacteria by nitric oxide induced apoptosis. PLoS One, 6, e19105.
Hsieh, Y. S., Chien, C., Liao, S. K., Liao, S. F., Hung, W. T., Yang, W. B., Lin, C. C., 449 450 451 452 453 454 455 456 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475 476 477 478 479 480
Cheng, T. J., Chang, C. C., Fang, J. M., & Wong, C. H. (2008). Structure and bioactivity of the polysaccharides in medicinal plant Dendrobium huoshanense. Bioorganic & Medicinal Chemistry, 16, 6054-6068.
Hua, Y. F., Zhang, M., Fu, C. X., Chen, Z. H., & Chan, G. Y. (2004). Structural characterization of a 2-O-acetylglucomannan from Dendrobium officinale stem. Carbohydrate Research, 339, 2219-2224.
Kimura, Y., & Sumiyoshi, M. (2004). Effects of various Eleutherococcus senticosus cortex on swimming time, natural killer activity and corticosterone level in forced swimming stressed mice. Journal of Ethnopharmacology, 95, 447-453.
Lai, F. L., Wang, Y. H., Chung, Y. W., Hwang, S. M., & Huang, L. S. (2011). Cell docking, movement and cell-cell interactions of heterogeneous cell suspensions in a cell manipulation microdevice. Sensors, 11, 9613-9627.
Laine, C., Tamminen, T., Vikkula, A., & Vuorinen, T. (2002). Methylation analysis as a tool for structural analysis of wood polysaccharides. Holzforschung, 56, 607-614. Liu, X. F., Zhu, J., Ge, S. Y., Xia, L. J., Yang, H. Y., Qian, Y. T., & Ren, F. Z. (2011). Orally administered Dendrobium officinale and its polysaccharides enhance immune functions in BALB/c mice. Natural Product Communications, 6, 867-870. Lo, S. F., Mulabagal, V., Chen, C. L., Kuo, C. L., & Tsay, H. S. (2004). Bioguided fractionation and isolation of free radical scavenging components from in vitro propagated chinese medicinal plants Dendrobium tosaense Makino and Dendrobium moniliforme SW. Journal of Agricultural and Food Chemistry, 52, 6916-6919. Lo, S. F., Nalawade, S. M., Kuo, C. L., Chen, C. L., & Tsay, H. S. (2004). Asymbiotic germination of immature seeds, plantlet development and ex vitro establishment of plants of Dendrobium tosaense Makino - A medicinally important orchid. In Vitro Cellular & Developmental Biology-Plant, 40, 528-535.
Malek, T. R. (2008). The biology of interleukin-2. Annual Review of Immunology, 26, 453-479.
Ng, T. B., Liu, J., Wong, J. H., Ye, X., Wing Sze, S. C., Tong, Y., & Zhang, K. Y. (2012). Review of research on Dendrobium, a prized folk medicine. Applied microbiology and biotechnology, 93, 1795-1803.
Palacios, R. (1982). Concanavalin A triggers T lymphocytes by directly interacting with their receptors for activation. Journal of Immunology, 128, 337-342.
Schepetkin, I. A., & Quinn, M. T. (2006). Botanical polysaccharides: macrophage immunomodulation and therapeutic potential. International Immunopharmacology, 6, 317-333.
Suzuki, I., Tanaka, H., Kinoshita, A., Oikawa, S., Osawa, M., & Yadomae, T. (1990). Effect of orally administered beta-glucan on macrophage function in mice.
International Journal of Immunopharmacology, 12, 675-684. 481 482 483 484 485 486 487 488 489 490 491 492 493 494 495 496 497 498 499 500 501 502 503 504 505 506 507 508 509 510 511 512 513 514 515 516 517 518
Volman, J. J., Ramakers, J. D., & Plat, J. (2008). Dietary modulation of immune function by beta-glucans. Physiology & behavior, 94, 276-284.
Wang, J. H., Luo, J. P., Zha, X. Q., & Feng, B. J. (2010). Comparison of antitumor activities of different polysaccharide fractions from the stems of Dendrobium nobile Lindl. Carbohydrate Polymers, 79, 114-118.
Wang, P., Henning, S. M., & Heber, D. (2010). Limitations of MTT and MTS-based assays for measurement of antiproliferative activity of green tea polyphenols. PLoS One, 5, e10202.
Wu, C. T., Huang, K. S., Yang, C. H., Chen, Y. C., Liao, J. W., Kuo, C. L., Chen, C. L., Lo, S. F., Hsieh, C. C., & Tsay, H. S. (2013). Inhibitory effects of cultured Dendrobium tosaense on atopic dermatitis murine model. International journal of pharmaceutics, [Epub ahead of print]
Yang, L. C., Hsieh, C. C., Lu, T. J. & Lin, W. C. (2013). Structurally characterized arabinogalactan from Anoectochilus formosanus as an immuno-modulator against CT26 colon cancer in BALB/c mice. Phytomedicine. [Epub ahead of print]
Yuan, C., Pan, X., Gong, Y., Xia, A., Wu, G., Tang, J., & Han, X. (2008). Effects of Astragalus polysaccharides (APS) on the expression of immune response genes in head kidney, gill and spleen of the common carp, Cyprinus carpio L. International Immunopharmacology, 8, 51-58.
Zha, X. Q., Luo, J. P., Luo, S. Z., & Jiang, S. T. (2007). Structure identification of a new immunostimulating polysaccharide from the stems of Dendrobium huoshanense. Carbohydrate Polymers, 69, 86-93.
Zhang, X., Goncalves, R., & Mosser, D. M. (2008). The isolation and characterization of murine macrophages. Current Protocol Immunology, Chapter 14, Unit 14.11. Zhao, X., Hu, Y., Wang, D., Liu, J., & Guo, L. (2013). The comparison of immune-enhancing activity of sulfated polysaccharidses from Tremella and Condonpsis pilosula. Carbohydrate Polymers, 98, 438-443.
Zhao, Y., Son, Y. O., Kim, S. S., Jang, Y. S., & Lee, J. C. (2007). Antioxidant and anti-hyperglycemic activity of polysaccharide isolated from Dendrobium
chrysotoxum Lindl. Journal of biochemistry and molecular biology, 40, 670-677. 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547 548 549 550
Figure legends
Figure 1 Chromatographic fractionation of polysaccharides isolated from Dendrobium tosaense (DTP). DTP were separated on DEAE-650M and eluted with 20 mM tris buffer (pH 7.8) followed by a sodium chloride gradient (0-0.55M), and total carbohydrate content of the fractions was determined by a phenol-sulfuric acid method (detected at 492 nm).
Figure 2 1H NMR spectra of neutral polysaccharide fraction.
Figure 3 High performance size exclusion chromatography (HPSEC) of polysaccharides of D. tosaense. The indigestible polysaccharides (DTP) and neutral polysaccharide (DTP-N) of D. tosaense were presented as solid line and dotted line, respectively. HPSEC was carried out using a HPLC system equipped with RI detector and TSKgel guard column PWH (75 × 7.5 mm i.d.; TOSOH, Tokyo, Japan), TSKgel G4000PWXL, and G2500PWXL (300 × 7.8 mm i.d., TOSOH) connected in series. The molecular weight of pullulan standards were 708 kDa, 375 kDa, 200 kDa, 107 kDa, 47.1 kDa, 21.1 kDa, 9.6 kDa and 5.9 kDa. The standard curve axis is right off the figure.
Figure 4 Effects of splenocyte proliferations of DTP in BALB/c mice. The
proliferation of splenocytes was induced by mitogen, Con A or LPS. All values were mean ± SD (n=12). Values were significantly different compared with the control group by Dunnett’s test: * P< 0.05.
Figure 5 Innate immunity effects of AG in BALB/c mice. (A) Splenic NK cytotoxicity against YAC-1 cells in DTP-treated mice (B) phagocytic activity of splenocytes from DTP-treated or control mice. All values were mean ± SD (n=12). Values were significantly different compared with the control group by Dunnett’s test: * P< 0.05, **P< 0.01 and *** P<0.001
Figure 6 Effects of DTP in cytokines production (A) IL-2 (B) IL-4 (C) IL-6 (D) IFN-γ. The cytokines were secreted by separated splenocytes from DTP-treated or control mice, and induced by mitogen, ConA or LPS. All values were mean ± SD (n=12). Values were significantly different compared with the control group by Dunnett’s test: * P< 0.05. 551 552 553 554 555 556 557 558 559 560 561 562 563 564 565 566 567 568 569 570 571 572 573 574 575 576 577 578 579 580 581 582 583 584 585 586 587
Table 1 Linkage analysis of the neutral polysaccharide (DTP-N) of D. tosaense. The linkage of DTP-N was determined by GC-MS. The Methylation of DTP-N was done before analysis.
Terminal Non-branch Branch
Mol% T-Glc T-Man 3-Glc 3-Man 4-Man 6-Man 3,4-Man
0.54 4.68 3.90 3.73 59.42 11.61 16.12
5.22% 78.65% 16.12%
Mol%, the mole percent 588 589 590 591 592 593
Table 2 Immunostimulating effects of indigestible polysaccharide of D. tosaense (DTP) on the population of splenic lymphocytes in BALB/c mice.
Groups Dose (mg/kg) T cell (%) B cell (%) Helper T cell (%) Cytotoxic T cell (%) Natural Killer (%) Control -- 40.5 ± 6.6 57.0 ± 6.6 23.7 ± 3.0 14.3 ± 1.8 11.1 ± 1.3 DTP 100 40.2 ± 3.8 59.2 ± 4.4 22.9 ± 1.8 13.6 ± 1.4 12.9 ± 0.7* 300 40.8 ± 4.1 56.4 ± 5.4 23.5 ± 1.7 15.0 ± 2.2 13.1 ± 1.9*
All values were mean ± SD (n=12). Values were significantly different compared with the control group by Dunnett’s test: * P< 0.05.
594 595 596 597 598 599