國立台灣大學生命科學院動物學研究所 博士論文
Institute of Zoology College of Life Science National Taiwan University
Doctoral Thesis
草蝦硫醇氧化還原蛋白與蝦白點症病毒極早期表現蛋白 IE1 蛋白質交 互作用之機制研究
Study of the interaction mechanisms between white spot syndrome virus (WSSV) immediate early gene #1 (IE1) and P. monodon
thioreodxin
黃俊諺 Jiun-Yan Huang
指導教授:郭光雄 博士 Advisor: Guang-Hsiung Kou, Ph.D.
指導教授:羅竹芳 博士 Advisor: Chu-Fang Lo, Ph.D.
中華民國 101 年 7 月
July, 2012
I
誌謝
七年
這本論文是敘述一個關於蛋白質跟空氣接觸久了會壞掉的故事
這個故事讓一個在高中因為太多學科不及格而被留級的少年完成他的博士論文
首先要謝謝我的父母親及家人,自幼以來一直縱容我的任性,總是放手讓我 去做我想做的事情。雖然同住一個屋簷下,但慚愧的是七年來平日晚上從未在家 一起用餐過。謝謝您們的包容,也希望沒有辜負您們的期待。
謝謝兩位指導教授郭光雄老師與羅竹芳老師,兩位老師總是可以一針見血的 點出我的不足之處,學生自認資質駑鈍,但從兩位老師身上深深感受到的是沒有 不夠聰明、只有還不夠努力的道理。謝謝張震東老師,張老師強大的背景知識與 細膩的方法學讓學生突破許多研究上的瓶頸,更帶領著白點症病毒朝向一個新的 領域來發展。謝謝蔡孟勳老師,在蔡老師的幫忙下進行數據分析及比對,讓我們 鎖定了最初的研究方向。謝謝呂健宏老師及王涵青老師,學生一路上看著兩位前 輩的身影,從兩位前輩身上發現太多值得我效法與學習的事情,亦師亦友的情感,
我很珍惜。
謝謝 818 這個大家庭,感謝在這段時間當中我所認識的每一個人,我可以從 你們每一個人身上看到我所沒有的優點, 並且經由你們的提醒與叮嚀注意到自 己需改進的地方,在這幾年的朝夕相處之中,謝謝你們對我的包容,請原諒我不 擅長表達內心的情感,若不是有每一個人盡心盡力的用自己的方式相互支援及扶 持,不可能會有今天的我,在此深深的謝謝你們所有人。
謝謝佳薇,這幾年來一直都是支持著我不要停下腳步的最大動力來源,總是 讓我忍不住的開懷大笑,但也虧欠妳最多,這幾年辛苦妳了,往後的日子還請多 多指教。: )
II
中文摘要:
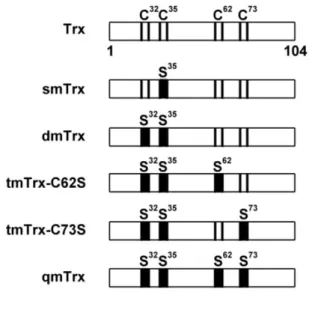
硫醇氧化還原蛋白為生物內參與許多氧化還原反應之蛋白質,利用硫醇-二硫 化物間的轉換提供氫離子及電子以進行氧化還原反應。在本研究中發現到當利用 本實驗室所產製的草蝦 cDNA 微陣列晶片觀測草蝦感染白點症病毒後基因表現 量的變化時,發現草蝦硫醇氧化還原蛋白基因的表現量有升高之趨勢,在佐以即 時定量核酸聚合酶鏈鎖反應確認此一結果後,接續利用西方點墨法得知在轉譯層 次上草蝦硫醇氧化還原蛋白在草蝦感染白點症病毒後其蛋白表現量也有上升的 現象,顯示草蝦硫醇氧化還原蛋白在白點症病毒感染草蝦過程中應該有重要的角 色。硫醇氧化還原蛋白的目標蛋白通常含有 Cys-x-x-Cys motif,故我們對白點症 病毒基因進行序列比對,共發現有 70 個白點症病毒開放轉譯區皆含有此 motif,
其中一個即是白點症病毒極早期表現基因 IE1。硫醇氧化還原蛋白會幫助轉錄因 子維持其還原態構型,使轉錄因子保有可以與目標 DNA 啟動子區域結合的能力,
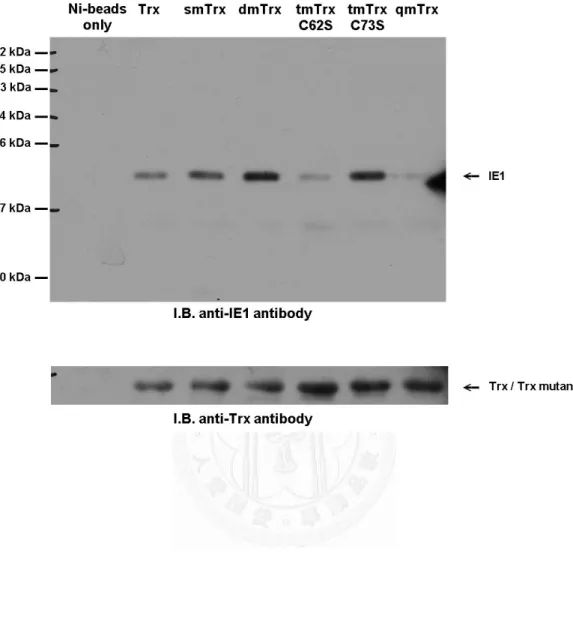
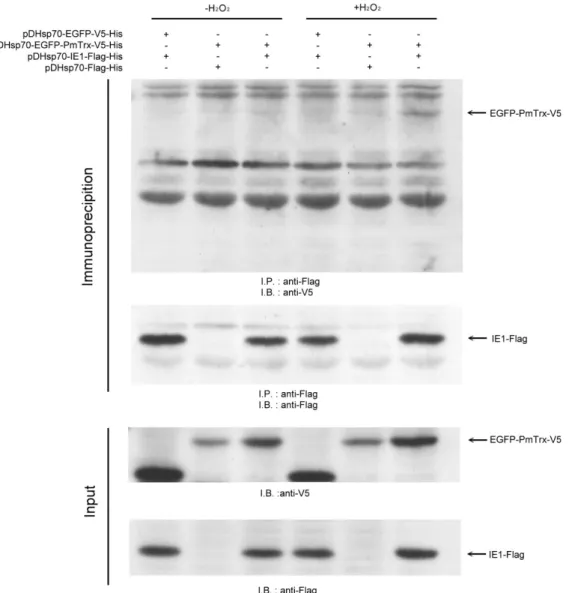
故我們先針對草蝦硫氧化還原蛋白是否會與白點症病毒極早期表現基因 IE1 有 蛋白交互作用進行試驗分析。我們利用體外試驗的 His pull down 分析法証實當 在有硫醇氧化劑 Diamide 的處理下,草蝦硫醇氧化還原蛋白會與白點症病毒極早 期表現基因、同時也是轉錄因子的 IE1 有蛋白交互作用的產生,並且鑑定出在草 蝦硫醇氧化還原蛋白藉由 Cys62半胱氨酸與 IE1 結合。接續以昆蟲細胞株 SF9 進 行共免疫沉澱法的體內試驗當中發現到,當細胞株受到氧化壓力如過氧化氫的處 理時,草蝦硫醇氧化還原蛋白會與白點症病毒 IE1 有蛋白交互作用的現象產生,
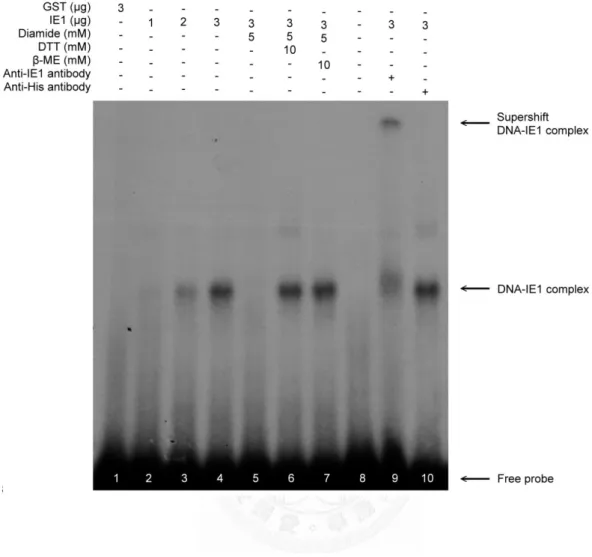
我們進一步的推測白點症病毒 IE1 其蛋白活性與功能會受到氧化還原的調控,進 而我們利用 EMSA 証實了當白點症病毒 IE1 受到硫醇氧化劑 Diamide 處理過後 會喪失與 DNA 結合的能力,而硫醇還原劑如 dithiothreitol 與 beta-mercaptoethanol 皆可以使其恢復活性、使白點症病毒 IE1 重獲與 DNA 結合的能力。進一步我們 也證實了 IE1 蛋白其 Cys55、Cys189與 Cys192 (CXXC motif) 半胱氨酸對 IE1 的 DNA 結合活性有重要的影響,我們証實了草蝦硫氧化蛋白可以直接幫助白點症病毒的
III
轉錄因子 IE1 維持或恢復其與 DNA 結合的能力,而以 LC/MS/MS 分析法發現氧 化壓力指標之一的 GSH/GSSG 比值在蝦體感染白點症病毒 24 小時後有上升的趨 勢,在感染後 48 小時會有顯著性的下降;而在 Trx 基因靜默化的蝦體中,白點 症病毒的複製套數以及蝦體感染白點症病毒的死亡率都會有顯著性的下降,本研 究結果讓我們對硫氧化蛋白所扮演的功能有更多的了解之外,對於白點症病毒的 致病機制與在宿主細胞中生存策略提供不同的視野與新思維。
IV
Abstract
A shrimp cDNA microarray and immunoblotting were used to confirm that white sopt syndrome virus (WSSV) infection up-regulated expression of the important redox regulator thioredoxin (Trx). WSSV immediate early gene #1 (IE1) was identified as a possible target protein of Trx and selected for further study. In a pull down assay, we found that recombinant Trx bound to IE1 under oxidizing conditions, and a co-immunoprecipitation assay showed that Trx bound to WSSV IE1 when the transfected cells were subjected to oxidative stress. A pull down assay with Trx mutants showed that no IE1 binding occurred when cysteine 62 was replaced by serine. EMSA showed that the DNA binding activity of WSSV IE1 was down-regulated under oxidative conditions, and that PmTrx restored the DNA binding activity of the inactivated, oxidized WSSV IE1. Another EMSA experiment showed that IE1’s Cys-X-X-Cys motif and cysteine residue 55 were necessary for Trx binding. Measurement of the ratio of reduced glutathione to oxidized glutathione (GSH/GSSG) in WSSV-infected shrimp showed that oxidative stress was significantly increased at 48 hpi. The biological significance of Trx was also demonstrated in a dsRNA Trx knockdown experiment where suppression of shrimp
Trx led to significant decreases in mortality and viral copy numbers. Based on all of
these results, we hypothesize that Trx uses the same binding sites and redox control mechanism in vitro and in vivo, and that Trx increase WSSV’s pathogenicity by rescuing IE1’s DNA binding ability in vivo.
V
目錄
誌謝...I 中文摘要...II 英文摘要...IV
第一章 文獻回顧...1
第二章 草蝦硫醇氧化還原蛋白與蝦白點症病毒極早期表現蛋白 IE1 蛋白質交互 作用之機制研究...32
文獻探討...32
研究方法...38
結果...48
討論...57
圖...69
表...88
參考文獻...94
個人相關著作...101
研討會論文...102
1
第一章 文獻回顧
1. 氧化還原反應(Redox reaction)
廣義的氧化還原反應的定義為:失去電子為氧化反應,接受電子為還原反應;
而當一分子接受一個電子時(被還原),同時間一定有另一分子失去電子(被氧化),
是故氧化反應必定伴隨著還原反應的發生,故總體來說稱為氧化還原反應。進一 步的說,氧化劑/還原劑是一種相對的概念,氧化劑本身會接受電子(被還原)並讓 與之反應的物質失去電子(被氧化);還原劑則會將本身的電子傳遞出去(被氧化) 並讓與之反應的物質得到電子(被還原) (40)。
依照電化學標準狀態所測出的還原電位,兩個物質在一起,則:
標準還原電位大者當氧化劑 標準還原電位小者當還原劑 以下是一些生物體中常見分子的標準還原電位(3,36,40)
Half-reaction
E0’=
Stand reduction potential (V)
low oxidizing agent DTT-oxi / DTT-red -0.33
O2 / O2.−
-0.33
GSSG, 2 H+ / 2GSH
-0.24
Trx-S2, 2H+ / Trx[SH]2 (Thioredoxin)
-0.24
Grx-S2, 2H+ / Grx[SH]2 (Glutaredoxin)
-0.22
H2O2, H+ / H2O, HO.
+0.32
O2.−, H+ / H2O2
+0.94
Strong oxidizing agent HO. , H+/ H2O +2.33
2
2. 氧化壓力(Oxidative stress)
對於所有的耗氧生物體(aerobic organisms)而言,生物體必需進行有氧呼吸以 產生能量來源--ATP,藉由 ATPase 幫浦讓氫離子進入粒腺體內以產生 ATP,而 粒腺體內的過多的氫離子需要靠電子傳遞鏈(electron transport chain)再將氫離子 送出粒腺體外,在進行電子傳遞的反應時,NADH 作為電子的提供者(electron donor)、,而氧氣作為電子的接受者(electron receptor),在行有氧呼吸時,約有 90%的 O2會走入電子傳遞鏈,以 cytochrome oxidase 作為電子的攜帶者(electron carrier)將四個電子給予氧氣以生成水(H2O);但約有 10%的氧分子會分別形成不 同的氧衍生物(oxygen derived),這些氧衍生物又稱為活性氧 (Reactive oxygen species;ROS)
生物體內氧催化代謝路徑如圖所示
造成生物體內氧化壓力的物質主要由 ROS 所造成,ROS 其成員為氧衍生物 形成的活性代謝物(Reactive metabolites),其種類繁多,最廣泛被研究的如屬於自 由基分子的超氧陰離子(superoxide anion, O2
‧–)與氫氧自由基(hydroxyl radical,–‧
OH),另外是非自由基分子的過氧化氫(H2O2) 。細胞內氧化物的產生與代謝為一 種動態平衡;而氧化壓力意指在生物體內氧化物的生成與消去之速度處於一種失 衡狀態(imbalance),當過量的 ROS 產生、又缺少足夠的抗氧化物質(anti-oxidants) 時,就會破壞生物體內的平衡狀態,造成氧化強化劑(Pro-oxidants)的累積,進而 有氧化壓力的產生(54),而不正常氧化壓力的累積,會使得細胞受到損傷之外,
也會伴隨著許多疾病的產生(39)。
3
2.1 自由基與活性氧族之介紹 2.1.1 自由基
分子由原子所組成,而原子又由質子、中子與電子所構成,一般而言,帶有 偶數電子對的分子才會達到安定的化學狀態。自由基(Free radical)是指具有不成 對電子的原子、離子或分子,故自由基是一種不安定的物質,並且會主動想要再 獲得一個電子讓自己達到穩定狀態,也因如此,自由基物質在生物體中具有高度 的生物反應性,而與之反應的分子因為其奇數電子的失去(氧化反應)而形成一個 新的自由基物質,並同時也可能改變了分子原有的功能與特性,往往造成了不可 逆的損傷;而新生成的自由基物質接續又會與別的分子進行反應,導致產生一連 串的放大連鎖反應並對細胞或整體組織造成傷害,在生物體當中為了要停止這種 連鎖反應,必需藉由體內的抗氧化物質(anti-oxidants),讓這種帶有不成對電子的 自由基跟另外一個也具有不成對電子的自由基物質進行反應,以消去帶有不成對 電子的自由基。
自由基的生成主要有外在因子與內生性因子(exogenous and endogenous factors)兩種來源,外在因子主要為 UV 光、香煙燃燒不完全所產生的煙霧、游離 輻射射源、除草劑或是殺蟲劑等;內生性因子 主要為粒腺體電子傳遞系統 (mitochondrial electron transport system)進行有氧呼吸時所產生電子逸漏的現象,
另外如細胞內進行氧化磷酸化(oxidative phosphorylation)、核苷酸的代謝與細胞 內啟動防禦機制時皆會有自由基的產生(23,56,81),生物體內自由基的種類繁多,
本論文後續僅回顧在氧氣代謝過程當中所產生的自由基分子。
2.1.1 超氧陰離子(superoxide anion, O2
‧–)
超氧陰離子與其它 ROS 物質相較之下並不是一個具活潑性質的 ROS,缺少 可以自由通透脂膜 (lipid membrane)的能力,雖然可以藉由擴散作用穿透生物膜,
但其速率極為緩慢(2.1×10-6 cm s-1)(11,27)。超氧陰離子大多受到細胞內的生物因
4
子所產生,如在粒腺體內膜上的電子傳遞系統中,大量電子的存在會造成電子洩 漏(leakage),致使氧氣自發性的生成超氧陰離子。超氧陰離子也是生物體當中許 多生化反應中不可避免的副產物,如在 ATP 代謝的過程當中會形成嘌呤的衍生 物次黃嘌呤 (hypoxanthine) ,而次黃嘌呤的代謝需經黃嘌呤氧化酶 (xanthine oxidase)氧化,隨後即會形成黃嘌呤(xanthine)與副產物--超氧陰離子和過氧化氫,
而黃嘌呤氧化酶可繼續將黃嘌呤氧化形成尿酸排出體外,但此時一樣會有副產物 超氧陰離子和過氧化氫的產生(58)。
超氧化物歧化酶(Superoxide dismutase, SOD) 可幫助超氧陰離子的代謝,藉 由金屬離子 Cu/Zn 或 Mn 的參與下可將超氧陰離子轉換成過氧化氫與氧氣 (77,117)。
2 O2
‧– + 2H+ → H2O2 + O2
2.1.2 過氧化氫(H2O2)
過氧化氫並不是自由基分子,是形成自由基過程中的一個中間分子,但也歸 類為 ROS (85)。生物體內除了由超氧陰離子還原形成過氧化氫之外,過氧化氫 本身為電中性(uncharged),並有著可以自由進出生物體細胞膜的特性,讓過氧化 氫成為細胞內訊息傳遞的一個重要分子,參與細胞凋亡調控機制(40),並在防禦 機制上扮演著重要的角色(86,91)。如白血球之中性粒細胞(neutrophil)內的骨髓過 氧化酶(Myeloperoxidase)可將過氧化氫轉變為次氯酸(hypochlorous acid, HOCl),
進而利用次氯酸殺死外來物如細菌等(116)。
H+ + Cl- + H2O2 →HOCl + H2O
在有金屬離子的存在下,當過氧化氫接受一個電子後,有兩種方式會形成最 不穩定的自由基分子--氫氧自由基,一者為著名的 Fenton’s reaction (26)。
H2O2 + Cu+/Fe2+ → ‧OH + OH- + Cu2+/Fe3+
另一種可能性是超氧陰離子與過氧化氫反應以生成氫氧自由基,此為 Haber-Weiss reaction (55)
5
O2‧- + H2O2 → ‧OH + OH- + O2
而受到 UV 光的照射也會使過氧化氫均裂(homolytic fission)為氫氧自由基 H2O2 → ‧OH + ‧OH
有三種酵素能在細胞內直接將過氧化氫代謝為水,分別為過氧化氫酶 (catalase)、麩胱甘肽過氧化酵素(glutathione peroxidase , GPx) 與過氧化物還原酶 (peroxiredoxin, Prx),這些機制在後續有論述。
2.1.3 氫氧自由基(hydroxyl radical,‧OH)
當過氧化氫本身得到一個電子被還原後即會形成氫氧自由基,而氫氧自由基 在 ROS 當中具有最活躍的生物反應性,標準還原電位為 E=2.59,在體內極端的 不穩定,並會與細胞內各種分子/巨分子進行氧化還原反應,雖然單一氫氧自由 基在體內存在的週期(life time)只有 10-9秒(11,27,98),並當氫氧自由基再獲得一
個電子會形成氫氧根離子(OH-),隨後與氫離子反應下即會完成氧氣在生物體內
代謝的最終產物—水。但氫氧自由基其對生物體所造成的的傷害確最為重大,可 以跟核酸(DNA and RNA)、蛋白質(protein)、脂質(lipid)或是小分子代謝物等進行 氧化還原反應,並可能進而改變氧化目標的結構與分子特性。除了由超氧陰離子 或過氧化氫轉變成氫氣自由基之外,游離輻射能的照射亦可使體內的水均裂 (homolytic fission)為氫氧自由基與氫自由基。
H2O → ‧OH + ‧H
2.2 活性氧在生物體中造成的傷害(Cellular damages by ROS)
ROS 在生物體中是細胞內各種生化反應下無法必免的副產物(by-product),
例如當進行有氧呼吸、或是活化嗜中性白血球時必定伴隨著 ROS 的產生,而 ROS 對於生物體本身主要攻擊的目標為核酸(DNA and RNA)、蛋白質(protein)與脂質 (lipid),所造成的影響分述如下。
6
核酸(DNA and RNA)
核酸(DNA and RNA)為受到 ROS 攻擊的主要目標之一,且被認為是一種核 酸誘變劑(mutagenic),因為 ROS 會造成單股或雙股的核酸骨架斷裂,並產生多 餘的含氮鹼基,而這些受損的含氮鹼基會與正常的含氮鹼基或其它分子配對形成 共價鍵結(99);在體外(in vitro)與體內(in vivo)試驗的研究結果顯示當 DNA 受到 ROS(特別是氫氧自由基)攻擊後會造成核酸構型異常扭曲,進而與蛋白質有架橋 連結(thymine-tyrosine cross-link)的現象產生(22,75)。
蛋白質(Protein)
在體外試驗的結果指出,ROS 會與胺基酸產生反應並造成電性的改變,
使蛋白質產生胜肽鏈斷裂,蛋白的聚集(aggregation)與雙硫鍵異常的架橋作用 (disulfide bonds cross-linking) 或 是 對 蛋 白 質 產 生 碳 醯 化 修 飾 作 用 (carbonyl modified proteins) (66),而生物體當中的 ROS 也被認為可調控帶有硫醇蛋白之活 性,利用 ROS 所產生的 氧化壓力對不同的蛋白質其活性產生不同的影響(68);
生物體中容易被氧化的胺基酸有四個,分別為 cysteine、methionine、 tyrosine 與 tryptophan (92),而在這四個胺基酸中最廣範被研究的即是以 Cysteine 為主的 氧化反應,Cys 也是唯一擁有數種不同氧化方式的胺基酸,主要藉由 Cys 本身的 硫氫基(Sulfhydryl group)氧化形成多種不同氧化狀態(25),大致上分成四種氧化 型態: 雙硫鍵的形成 (inter or intra disulfide bond formaton)、與過氧化氫的氧化反 應(sulfenic acid、sulfinic acid or sulfonic acid formation) 、與 GSH 鍵結的氧化反 應 (glutathiolation) 以 及 與 一 氧 化 氮 (nitrogen monoxide, NO) 相 關 的 氧 化 反 應 (S-Nitrosylation),反應示意圖如下:
7
在細胞外(extracellular space)與位於內質網(endoplasmic reticulum)的蛋白大 多藉由雙硫鍵的形成以維持自身蛋白正確的折疊構型(6),相較之下,細胞質內 是一個充滿還原力的環境,在細胞質內的蛋白其酵素活性作用區位、與金屬離子 的結合或是催化反應等皆是藉由還原態的 Cys (61),生物體當中並不是所有蛋白 質上的 Cys 皆與氧化還原相關,在蛋白質不同折疊構型的情況下大部份的 Cys 無法進行氧化還原反應,但 Cys 所受到的氧化還原反應確實是生物體內很重要的 調控機制(95),許多 Chaperone (45) 、代謝相關的酵素活性(21)、轉錄因子的活 性(48,89,121)以及激酶或磷酸酶 (kinase and phosphatase)(35,62)的活性皆與氧化 壓力所造成的氧化還原反應有相關。
脂質(Lipid)
在脂質方面,ROS 會攻擊在膜上的多元不飽和脂肪酸(Polyunsaturated fatty acid)並造成脂質的過氧化(lipid peroxidation),當不飽和脂肪酸被氫氧自由基攻擊 失去一個電子後會形成脂質自由基(lipid radical),與氧氣和其它的不飽和脂肪酸
8
接續反應後即會形成脂質過氧化物(lipid peroxide),整個代謝路徑也會產生脂質 氧化的連鎖反應(lipid peroxidation chain reaction),並在連鎖反應的情況下使整體 細胞膜的流動性降低,進而改變細胞膜的特性並造成膜蛋白的不穩定,需要藉由 Chain Breaking 的機制中止連鎖的氧化反應(82)。若脂質的氧化發生在粒腺體時,
更會同時對在粒腺體中的多種酵素產生不同的影響, 導致對細胞產生毒性 (cytotoxic)進而啟動細胞凋亡反應 (37)。
2.3 抗 氧 化 系 統 -- 硫 醇 氧 化 還 原 蛋 白 與 榖 氧 還 蛋 白 系 統 (Thioredoxin and Glutaredoxin system)
生物體當中有兩個主要針對硫醇(thiol)的抗氧化系統,分別為硫醇氧化還原 蛋白系統(Trx system)與榖氧還原蛋白系統(Grx system),這兩個系統會在生物體 內執行相類似的氧化還原反應,並且是細胞質中供給受質蛋白還原力的主要來 源;讓細胞質內維持還原的環境(reducing redox environment)(95),主要機制有 三種,第一是透過雙硫醇與雙硫鍵的轉換(dithiol-disulfide exchange)將受質蛋白 其雙硫鍵還原為硫醇,第二是受質蛋白質與 GSH 鍵結的麩胺基硫反應或去麩胺 基硫反應(glutathionylation or de-glutathionylation),第三是參與蛋白異構化反應 (isomerization),將受質蛋白的雙硫鍵鍵結位置改變以形成異構蛋白(38,41,78)。
硫醇氧化還原蛋白系統 (Thioredoxin system)
硫醇氧化還原蛋白系統主要由 Thioredoxin (Trx)與 Trx reductase (TR)所組成,
Trx 主要維持生物體內硫醇與雙硫鍵(thiol: disulfide)比例的衡定,例如可以促進 胰島素雙硫鍵的還原等,最早由 Laurent 等人發現在 DNA 合成過程當中需要硫 醇氧化還原蛋白的參與(59),核醣核酸(ribonucleotide, rNTP)在核醣核酸還原酶 (ribonucleotide reductase, RR) 催 化 下 可 被 Trx 還 原 為 去 氧 核 醣 核 酸 (deoxy ribonucleotide, dNTP) (42),反應式如下
9
Trx-[SH]2 + rNTP → Trx-S2 + dNTP + H2O
氧化態的 Trx 藉由 Trx 還原酶(thioredoxin reductase, TR)利用 NADPH 可將氧化態 的 Trx 進行還原作用,反應式如下
Trx-S2 + NADPH + H+ → Trx-(SH)2 + NADP+ (1) Protein-S2 + Trx-(SH)2 → protein-(SH)2 + Trx-S2 (2)
2.3.2 榖氧還蛋白系統 (Glutaredoxin system)
榖胱甘肽(Glutathione, GSH) ,穀氧還蛋白(Glutaredoxin, Grx)與麩胱甘肽還 原酶(Glutathione reductase, GR) 這三個蛋白質共同組成榖氧還蛋白系統,GSH 是 由 glutamate、cysteine 與 glycine 三種胺基酸組合而成的三胜肽,細胞內 GSH 含 量很高,可以達 milli-molar (mM) (14),Glutaredoxin 分子量在不同物種之間略有 差異,分子量約為 9-12 kDa,其作用活性區位亦為 CxxC motif(通常為 CPYC motif),
目前至少有約 30 種不同的 Grx 被鑑定出,除了像 Trx 系統一樣可以對雙硫鍵進 行 Dithiol 的還原反應之外,還可以對單硫醇進行 glutathiolation 相關的 Monothiol 氧化還原反應,Grx 系統的反應式如下
10
另外 Glutathione-S transferase (GST)可以將 GSH 與其它分子進行接合反應,為生 物體當中 de-toxic 反應的中間步驟,而在缺少 GR 酵素活性的果蠅品系之中,氧 化態的 GSH 可以作為 Trx 的受質,由 Trx 直接將 GSSG 還原為 GSH (50)。
2.4 其它的抗氧化酵素
超氧化物歧化酶(Superoxide dismutase, SOD)
超氧化物歧化酶是第一個被鑑定出其酵素功能為參與自由基的代謝(77),在 真核生物體中有兩種超氧化物歧化酶,分別在不同位置執行任務,而這兩種超氧 化物歧化酶在其酵素活性區位上皆需要金屬離子的參與才能保酵素活性(8,57);
在細胞質中為 Cu/Zn-SOD 執行清除超氧陰離子的工作,而 Mn-SOD 在粒腺體內 則扮演著很重要的角色,主要將有氧呼吸中電子傳遞鏈所產生的超氧陰離子轉換 成過氧化氫(77,117)。
2 O2
‧– + 2H+ → H2O2 + O2
過氧化氫酶(Catalase)
過氧化氫酶是含鐵的酵素(heme-containing enzymes)並以四聚體的形式聚合,
以致於以超過 220 kDa 的蛋白大小存在生物體中 (93),過氧化氫酶普遍存在於細 胞內各處,包含細胞質粒腺體與過氧化氫體(peroxisome)。NADPH 會與過氧化氫 酶結合並對其產生保護作用之外,更能促進其酵素活性(51-52)。過氧化氫酶最主 要的功能就是將 H2O2轉變為氧氣和水,對過氧化氫有相當高的 Km 值,生物體 中而 Gpx/Prx 與 Catalase 兩者都可以直接將過氧化氫轉變為水,但是在生物體中 以 Gpx 對過氧化氫的親合力較佳(46) 。
2 H2O2 →O2 + H2O
11
麩胱甘肽過氧化酵素(glutathione peroxidase , Gpx)
於哺乳動物中發現至少有四種不同的 Gpx,分別為 Gpx1-Gpx4,並在其酵素 活 性 區位 上皆 有硒 氨酸 (selenocysteine)的存 在,故 Gpx 必需要藉助 硒元素 (selenium-dependent )來保有其酵素活性(106)。Gpx1 為細胞質中表現量很高的酵 素,並幾乎在所有的組織皆可以偵測到 Gpx1 的存在(28), Gpx4 在粒腺體中的 含量最高並可代謝脂質過氧化物(lipid hydroperoxides),可將脂類過氧化物還原為 醇類(alcohols)(31), Gpx2 與 Gpx3 在高等生物體中主要為細胞外酵素,分別存 在於消化道與血漿當中(19-20,97,105)。
Gpx 可直接將過氧化氫還原為水,自己則形成帶有 sulfinic acid 的 Gpx-SOH,
Gpx-SOH 必需再藉著前述之機制由 Grx 系統內 GSH、GR 與 NADPH 的幫忙下 回復成還原態的 Gpx。
過氧化物還原酶 (peroxiredoxin, Prx or Tpx)
過氧化物還原酶 Prx 亦是含鐵的酵素(heme-containing enzymes),在生物體內 分子大約為 30 kDa,可以還原過氧化物(peroxides),例如過氧化氫(H2O2)或是烷 基過氧化物(ROOH)。Prx 為 Trx 的受質,氧化態的 Prx 必需要藉由 Trx 的幫助之 下回復為還原態的 Prx-SH,Prx-SH 可以與過氧化氫反應將過氧化氫代謝為水,
而 Prx 自己形成 sulfenic acid 的 Prx-SOH, Prx-SOH 除了可以再代謝一個過氧
12
化氫形成帶有 sulfinic acid 的 Prx-SOOH 之外,Prx-SOH 也可以跟還原態的 Prx-SH 自發性形成雙硫鍵鍵結的 Prx-S-S-Prx,再依賴 Trx 將 Prx-S-S-Prx 還原為 Prx-SH (94),而 Prx-SOOH 必需藉由 sulfiredoxin (Srx)與消耗 ATP 的情況下還原 為 Prx-SOH (7)。
整體反應路徑如下圖所示
綜合第二章所談到的 ROS 與抗氧化分子,用以下這張圖來同整體內氧氣代 謝的過程中所產生的 ROS,以及生物體內如何代謝這些 ROS 作個整體,首先是 當細胞接受到外界的 Stress(如 UV 光或是游離輻射)、或是進行有氧呼吸等的生 物反應時,細胞內會產生 superoxide,superoxide 不能自由通透生物膜 (或說通 透的速度極為緩慢),主要會在胞內流竄,而 SOD 是主要代謝 superoxide 的酵素,
SOD 可將 superoxide 轉換成過氧化氫,過氧化氫的分子量很小,可以自由的通 透生物膜,生物體內至少有三種酵素可以將過氧化氫直接代謝為水,這三種酵素 分別為 Gpx (與 Grx 系統相關)、Prx(與 Trx 系統相關)以及 Catalase (CAT)這三種 酵素,其中又以 CAT 對過氧化氫的具有最高的生物反應性;而過氧化氫有兩種
13
主要的方式自發性的形成氫氧自由基,一種是藉助生物體內 Fe+2或是 Cu+的催化 下進行 Fenton reaction 反應以產生氫氧自由基;另一種是 Fe+3 金屬離子先與 superoxide 進行反應生成 Fe+2後、Fe+2再與過氧化氫發生 Fenton reactoin 以產生 氫氧自由基(兩步驟合併起來稱為 Haber-Weiss reaction),氫氧自由基在生物體當 中具有極高的生物反應性,相較之下有很強的還原電位,對生物體而言是極強的 氧化劑,幾乎可以隨意的奪取周遭分子的電子,讓周圍原本穩定的分子變成不穩 定的自由基分子,也因這種極強的反應性,氫氧自由基在生物體中存在週期很短,
只有 10-9妙,但是卻會讓細胞內不論是核酸、蛋白或是脂質生物膜等發生不可逆 轉的損傷。
14
3 蝦類抗氧化相關基因之研究現況
超氧化物歧化酶(Superoxide dismutase, SOD)
對於真核生物來說,包含線蟲、昆蟲、貝類、魚類、或是人類而言,在細胞 質內都是由 Cu/Zn-SOD 作為清除自由基的酵素,而在粒腺體中是由 Mn-SOD 來 清除超氧陰離子,但非常讓人意外的是,各種在十足目中的蝦類與水生甲殼類動 物,其細胞質內並沒有 Cu/Zn-dependent SOD 的存在,對於蝦類而言,竟然是有 兩種的 Mn-SOD 分別在細胞質內與粒線體當中執行清除自由基的任務(9-10),後 來的研究者將細胞質中的 SOD 命名為 cytMn-SOD 或是 cMn-SOD,其蛋白分子 量為 25 kDa,並且不論在中國對蝦或是南美白蝦之中都已經將此基因選殖與作 功能上的鑑定分析,並且在全身性器官包含血球、心臟、肝胰臟、腸道、神經、
肌肉、泳足與鰓皆可偵測到有 SOD 基因的表現(18,29,32,67) 。
免疫刺激物如 beta-glucan 與 sulfated polysaccharide 可以增加 SOD 的活性(12),
在等足目(Probopyrus ringueleti)所寄生感染的蝦體(Palaemonetes argentinus)中其 SOD 酵素活性會有顯著性的下降(83)。在將 cMn-SOD 基因靜默化後佐以缺氧與 缺氧後覆氧試驗當中,蝦體內的鰓與肝胰臟其 SOD 活性與超氧陰離子的含量會 因為缺氧而下降;而缺氧後覆氧會造成 SOD 活性與超氧陰離子的含量的增加(29)。
另外在受到白點症病毒感染的蝦體中,SOD 基因的表現在感染後一小時會有顯 著性的上升,而隨著感染時間增加其基因表現會降低,並在感染後 12 小時會有 顯著性的下降(33)。
過氧化氫酶(Catalase, CAT)
過氧化氫酶在蝦類的研究當中相對較少,目前的文獻指出在蝦體中肝胰臟、
血球、淋巴組織、腸道、卵巢、肌肉與鰓皆可偵測到 Catalase 的基因表現(120),
並在肝胰臟可以偵測到其蛋白質的表現(101);在受到白點症病毒感染的蝦體中,
血球中 Catalase 基因的表現在感染後 14 小時有顯著性的上升;肝胰臟中 Catalase
15
基因的表現則在感染後 37 小時有顯著性的升高(120)。經微囊藻毒(Microcystin) 攻毒的蝦體其 Catalase 基因的表現與活性皆會升高(34),急性 pH 值改變的處理 也可以造成蝦體內 Catalase 基因表現量的變化,在酸性(pH5.4, 24 hpi; pH6.7, 72 hpi)或是鹼性處理(pH9.3, 48 hpi)其 Catalase 基因表現量會上升;但當酸/鹼性處理 超過 72 小時後其基因表現量皆有顯著性的下降(113)。
麩胱甘肽過氧化酵素(glutathione peroxidase , Gpx)
蝦類 Gpx 在 2007 年由 Liu 等人進行選殖與鑑定,在南美白蝦與草蝦其 Gpx 蛋白分子量大小約為 20 kDa,全身性的組織如血球、鰓、心臟、肝胰臟、腸道 與肌肉組織等皆有 Gpx 的基因表現,在親源關係方面蝦類 Gpx 比較近似於人類 的 Gpx1 與 Gpx2,在弧菌與白點症病毒的攻毒試驗之中,Gpx 基因的表現階會 有顯著性的上升,而酵素活性則是波動較大,雖有顯著性差異但真實情況還需更 多的文獻報告來証實(69-70)。
過氧化物還原酶 (peroxiredoxin, Prx)
蝦類的 Prx 直到 2007 年才由 Zhang 等人進行選殖與鑑定,在中國對蝦中 Prx 其開放轉譯區序列有 594 bp,蛋白質分子量大小為 22 kDa,與果蠅的蛋白質序 列相似度有 72%,並全身性的組織皆有 Prx 基因的表現,在弧菌攻毒試驗中發現 肝胰臟與血球中 Prx 基因的表現會有顯著性的變化,在肝胰臟中 8 hpi 其 Prx 基 因的表現量會有顯著性的下降,而 37 hpi 後則會有顯著性的上升;在血球中 5 hpi 與 37 hpi 其 Prx 基因表現量會有顯著性的上升,而 14 hpi 則會有顯著性的下降 (119)。對班節蝦施打免疫刺激物 peptidoglycan 過後心臟及淋巴器官之 Prx 基因 表現量會有顯著性的下降(4);而當草蝦施打免疫刺激物 lipopolyssacharide 後肝 胰臟之 Prx 基因表現量會有顯著性的上升(88);至於感染白點症病毒的蝦體其 Prx 基因有下降的趨勢(53)。
16
硫醇氧化還原蛋白(Trx)系統
直至 2008 年在蝦類才首度有 Trx 的相關研究報告出現,其 Trx 開放轉譯區有 315 bp,蛋白質分子量大小為 12 kDa,其基因並不具有入核訊號,並且含有四個 Cysteine ,其 中兩 個 Cysteine 為 在 所 有 物種 具 有高 度 保守 性的 CxxC motif (-Cys32-X-X-Cys35-),並且在全身各器官都可以測得 Trx 的基因表現而主要在鰓、
肝胰臟與泳足有高量表現(1,90),但令人意外的是,不論對蝦體進行缺氧或是缺 氧後復氧的處理,在鰓組織中的 Trx 其基因表現皆沒有達到顯著性的差異(1),
榖氧還蛋白(Grx)系統
要分析 Grx 系統中 GSH 或 GSSG 基因特性是一件困難的事,因為 GSH 是 由三個胺基酸所組成,幾乎不太可能用分生技術(如 RT-PCR 或是 Western Blot) 對其基因/蛋白的表現量做測定,大部份的研究者目前仍是就市售的 GSH 活性測 定試劑組對 GSH 進行量測,在蝦類研究當中,針對 GSH/GSSG/GR/Grx 此系統 之文獻報告相較之下仍非常稀少,一般認為 GSH 會對各種不同的逆境壓力(Stress) 作出反應,在對長臂蝦(Palaeomonetes pugio)進行不同的處理試驗發現到,高濃 度金屬離子氯化鎘(Cadmium chloride)、高溫(30℃)、養殖環境中含有不同濃度的 原油或柴油(Bunker or Diesel Fuel)與養殖環境中含有不同濃度的除草劑(Atrazine) 處理後,皆會造成 GSH 活性會有顯著性的上升(24),在 2012 年我們等人以 LC/MS/MS 的分析法測定 GSH/GSSG 的比值,指出當蝦體感染白點症病毒過後,
在 24 hpi 時 GSH/GSSG 比值相較控制組而言有上升的趨勢,48 hpi 時則會有顯 著性的下降(44)。這些研究報告代表逆境壓力會使正常蝦體的抗氧化系統啟動,
造成抗氧化活性上升進而抵抗氧化壓力。
17
4. 蝦類與氧化壓力之關聯性 (Aquatic animals and oxidative stress)
對生物體而言,各種逆境(Stress)的環境即往往會伴隨著不同的氧化壓力產生,
對於不同的水生生物而言,共通的環境逆境約可區成為溫度、水中含氧量、鹽度、
pH 質等,這些分別論述如下。
溫度
溫度對於所有的水生生物而言都是一個重要的環境因子,不論溫度的升高或 是降低對於許多的水生生物皆會造成氧化壓力,就熱力學而言溫度的升高會加速 各種代謝的反應,理論上會造成副產物—ROS 的累積;溫度對於青蛙(5)、魚(76)、
貝類(107)以及蟹類(80)等皆會造成氧化壓力;而當將蝦類養殖溫度從 15℃急速升 高至 28℃時,讓蝦體受急性熱刺激(acute heat stress)的處理後,逆境指標(stress marker)之一的熱刺激蛋白(heat shock protein, HSP)--HSP60 與 HSP70 其基因與蛋 白表現量會有顯著性的上升之外,蝦體內各抗氧化蛋白(SOD, Catalase, GPx 與 GST)其基因表現量亦會有顯著性的上升 (122)。雖然降低溫度可能會減少氧化壓 力的產生,但也有研究指出溫度的降低亦會對生物體造成氧化壓力(74,84),對蝦 類而言若將養殖水溫溫度從 25℃急遽降低至 12℃,蝦血球內 ROS 的測量數值在 處理後 3 小時與 6 小時會有顯著性的升高,並會造成 DNA 的損傷與脂膜滲透壓 的降低(87)。
水中含氧量
水中含氧量的濃度為造成氧化壓力的主要因素之一,在缺氧環境 (1 mg O2 ml-1) 的情況下處理 24 小時後,蝦體中的總血球數量(total haemocyte count)會有 明顯的降低之外,血球中的透明球(hyaline cell)及半顆粒球(semi-granular cell)數 量也會有顯著性的降低;此外,對於凝血系統(prophenoloxidase system)的酵素活 性也會減弱,而對蝦體進行弧菌(Vibrio alginolyticus or Vibrio parahaemolyticus)
18
的攻毒試驗當中,在缺氧的條件下對弧菌的死亡耐受性明顯的會降低(60,79),由 這些試驗結果可以推論出缺氧可能會造成蝦體的免疫機制下降,進而造成對外來 入侵物抵抗性的降低。而若當經過缺氧處理(1 mg O2/L)的蝦體、再以覆氧 (Reoxygenation)處理後,在蝦體的肌肉及肝胰臟的組織皆觀測到超氧陰離子的含 量有顯著性的上升以及抗氧化酵素活性的下降,並且在肌肉組織中還可以測量到 脂質氧化的比例降低(118),而在此篇報告當中值得注意的是,此缺氧條件的處 理或是缺氧後的覆氧並不會造成鰓組織當中有 ROS 升高或是抗氧化酵素的活性 上升,作者推論是鰓組織對於含氧量的變動有較高的耐受能力,但是肌肉或肝胰 臟所增加的 ROS 可能會在不同的組織之間傳遞並對不同組織造成損傷。
鹽度
一般而言蝦類對水質鹽度的耐受性較為廣泛,幼蝦或成蝦對於高/低滲透壓 (hyper/hypo-osmoregulators)皆具有較高的容忍度 (13),但不同濃度的鹽度緊迫試 驗(salinity stress test)一直是測試蝦苗健康與否的方法之一,在 PL1 之幼蝦以 10 ppt 鹽濃度浸泡處理 2 小時的情況下即達到過半致死劑量(2,96),而不論在急性高 鹽(從 30 ppt 至 50 ppt)或是急性低鹽(從 30 ppt 至 5 ppt)處理的情況下,SOD 與 GPx 的酵素活性會有顯著性的下降(71),雖然在本篇報告當中 Liu 等人亦有測量 Catalase 與 Na+/K+-ATPase 的酵素活性,但不論在急性高/低鹽的條件處理下,皆 沒有達到統計上的顯著差異,但由 SOD 與 GPx 有明顯變動的結果來看,還是暗 示了雖然蝦類為廣鹽性的水生生物,但在環境因子--鹽度快速且劇烈改變的情況 下,蝦體內仍會有 ROS 的堆積。
pH 質
相較於中性(pH=7.5-7.7)的環境下,在酸性(pH=4.6 – 5.0)與鹼性(pH=9.0-9.5) 的環境之中蝦體內總血球數量皆會有顯著性的下降(17), 而當蝦子處於鹼性環
19
境下 12 小時後,在血淋巴液中 GST 的酵素活性會有顯著性的下降,但在肝胰臟 與鰓當中並不會有異常的變動;而在酸性環境 12 小時後,在肝胰臟與鰓中 GST 的酵素活性則會有顯著性的上升,但血淋巴液中 GST 的酵素活性卻不會有異常 的變動(123)。
20
5 蝦白點症病毒
對於所有的蝦類而言蝦白點症病毒(White spot syndrome virus, WSSV)是一 個危害相當巨大的病毒,它具有雙股 DNA,外套膜蛋白,並有尾狀突起的構型,
其基因體大小約為 300 kbp,有 532 個長度大於 60 個胺基酸的開放轉譯區(open reading frame, ORF) (16,108)。宿主範圍極為廣泛,所有的甲殼類生物皆會感染白 點症病毒(72-73),而白點症病毒超過 90 %的 ORFs 皆與目前已知的蛋白毫無相 似之處(65),也因白點症病毒這種獨特的特性,國際病毒分類學會 ICTV 於 2004 年正式讓白點症病毒在分類學上單獨自成一個科(Nimaviridae)與屬(Whispovirus) (http://ictvonline.org/virusTaxonomy)。目前 WSSV 已鑑定出超過 40 個結構性蛋白 (102-103),在非結搆性蛋白的部份大致上分為早/中/晚期表現基因、泛素化作用 (ubiquitination)相關基因、抗細胞凋亡相關基因、DNA 擬態基因以及與複製相關 基因如 viral DNA polymerase 及 RR (ribonucleotide reductase)等(65,110);白點症 病毒其宿主範圍很廣,所有的十足目與甲殼類都會受到白點症病毒的感染,其致 病歷程極為快速,對蝦類蝦種(penaeid shrimps)感染白點症病毒後三到七天即可 造成百分之百的死亡率,如此獨特的病毒但我們對其詳細的致病機制仍有太多的 疑團未解 (65)。
目前在白點症病毒的感染機制的研究中,在早期除了對白點症病毒中具較高 保守性之功能性基因進行鑑定之外,也發現到白點症病毒會有其獨特的對抗宿主 細胞凋亡的機制(63-64)與利用 IRES (internal ribosome entry site)的轉譯機制(49);
亦發現白點症病毒高量表現蛋白 ICP11 為 DNA 擬態蛋白,會與宿主的組蛋白 (histone protein)的 DNA 結合區位進行結合,形成宿主體內 DNA 的競爭性抑制作 用(competitive inhibition),並造成組蛋白無法與目標 DNA 結合,進而導致宿主 DNA 的受損以及細胞瓦解(109);而在利用蛋白質體學技術進行白點症病毒感染 前後的蛋白表現差異分析結果出可以發現,具差異性表現的蛋白主要會與粒腺體 相關路徑(mitochondrial pathway)、能量產生(energy production)、核酸生成(nucleic
21
acid synthesis) 、 氧氣 傳遞 (oxygen carrier/transportation) 、 鈣離 子 調控 (calcium homeostasis)、訊息傳導(signaling/communication)以及與類泛素蛋白質修飾(small ubiquitin-like modifier, SUMO)相關的反應(111);其中以粒腺體相關調控機制已經 有初步的成果,VDAC (voltage dependent anion channel)是粒腺體外膜的主要蛋白,
與粒腺體所產生的細胞凋亡有高度正相關性,除了在白點症病毒感染的蝦體中發 現 VDAC 基因表現量的上升之外(111),對於 VDAC 基因靜默化的蝦體更會造成 白點症病毒感染流程的延遲(112),這些皆暗示了蝦體內之粒腺體與白點症病毒 的致病機轉有著密不可分的關係。
近年來也發現白點症病毒會引起蝦體產生瓦式效應(Warburg effect),瓦式效 應為諾貝爾獎得主 Otto Heinrich Warburg (October 8, 1883 - August 1, 1970)所提 出的理論,通常在探討癌細胞與正常細胞兩者間最主要的差別之一在於利用能量 的方式不同,癌細胞有別於一般正常細胞進行的有氧呼吸,其偏好利用醣解作用 (glycolysis)以取得能量(30,114);而在白點症病毒感染的蝦體中,感染早期(12 hpi) 蝦體內 glucose 消耗速率會有顯著性的增加、以及在五碳醣磷酸鹽路徑(Pentose phosphate pathway)中的關鍵酵素 glucose-6-phosphate dehydrogenase 活性的上升,
皆透露出白點症病毒會造成蝦體代謝的異常(15)。
目前對於白點症病毒的研究已經漸漸的朝向全面化進行探討,不論是宿主- 病毒之間的交互作用、或病毒本身的致病機轉方面近年來都有重大進展,另外也 亦有研究者的興趣是試圖找尋出與病毒作第一線接觸的膜蛋白分子(43,100,115);
也不斷的有新病毒檢疫平台出現(104)或是白點症病毒的體外試驗系統(47),這些 都讓研究者們對於病毒-宿主之間這場永無止盡的鬥爭提供全新視野與思維,持 續的發現生物學上的新知識。
22
參考文獻
1. Aispuro-Hernandez E, Garcia-Orozco KD, Muhlia-Almazan A, Del-Toro-Sanchez L, Robles-Sanchez RM, Hernandez J, Gonzalez-Aguilar G, Yepiz-Plascencia G, Sotelo-Mundo RR. Shrimp thioredoxin is a potent antioxidant protein. Comp
Biochem Physiol C Toxicol Pharmacol 148: 94-9, 2008.
2. Alvarez AL, Racotta IS, Arjona O, Palacios E. Salinity stress test as a predictor of survival during growout in Pacific white shrimp (Litopenaeus vannamei).
Aquaculture 237: 237-249, 2004.
3. Aslund F, Berndt KD, Holmgren A. Redox potentials of glutaredoxins and other thiol-disulfide oxidoreductases of the thioredoxin superfamily determined by direct protein-protein redox equilibria. Journal of Biological Chemistry 272:
30780-30786, 1997.
4. Bacano Maningas MB, Koyama T, Kondo H, Hirono I, Aoki T. A peroxiredoxin from kuruma shrimp, Marsupenaeus japonicus, inhibited by peptidoglycan.
Dev Comp Immunol 32: 198-203, 2008.
5. Bagnyukova TV, Storey KB, Lushchak VI. Induction of oxidative stress in Rana ridibunda during recovery from winter hibernation. Journal of Thermal
Biology 28: 21-28, 2003.
6. Bardwell JC. Building bridges: disulphide bond formation in the cell. Mol
Microbiol 14: 199-205, 1994.
7. Biteau B, Labarre J, Toledano MB. ATP-dependent reduction of cysteine-sulphinic acid by S. cerevisiae sulphiredoxin. Nature 425: 980-4, 2003.
8. Bordo D, Djinovic K, Bolognesi M. Conserved patterns in the Cu,Zn superoxide dismutase family. J Mol Biol 238: 366-86, 1994.
9. Brouwer M, Brouwer TH, Grater W, Enghild JJ, Thogersen IB. The paradigm that all oxygen-respiring eukaryotes have cytosolic CuZn-superoxide dismutase and that Mn-superoxide dismutase is localized to the mitochondria does not apply to a large group of marine arthropods. Biochemistry 36:
13381-8, 1997.
10. Brouwer M, Hoexum Brouwer T, Grater W, Brown-Peterson N. Replacement of a cytosolic copper/zinc superoxide dismutase by a novel cytosolic manganese superoxide dismutase in crustaceans that use copper (haemocyanin) for oxygen transport. Biochem J 374: 219-28, 2003.
11. Cadenas E. Biochemistry of oxygen toxicity. Annu Rev Biochem 58: 79-110, 1989.
12. Campa-Cordova AI, Hernandez-Saavedra NY, Ascencio F. Superoxide dismutase as modulator of immune function in American white shrimp
23
(Litopenaeus vannamei). Comp Biochem Physiol C Toxicol Pharmacol 133:
557-65, 2002.
13. Castille FL, Lawrence AL. The Effect of Salinity on the Osmotic, Sodium and Chloride Concentrations in the Hemolymph of Euryhaline Shrimp of the Genus Penaeus. Comparative Biochemistry and Physiology a-Physiology 68:
75-80, 1981.
14. Chang TC, Chang MJ, Hsueh S. Glutathione concentration and distribution in cervical cancers and adjacent normal tissues. Gynecol Obstet Invest 36: 52-5, 1993.
15. Chen IT, Aoki T, Huang YT, Hirono I, Chen TC, Huang JY, Chang GD, Lo CF, Wang HC. White Spot Syndrome Virus Induces Metabolic Changes Resembling the Warburg Effect in Shrimp Hemocytes in the Early Stage of Infection. Journal of
Virology 85: 12919-12928, 2011.
16. Chen LL, Leu JH, Huang CJ, Chou CM, Chen SM, Wang CH, Lo CF, Kou GH.
Identification of a nucleocapsid protein (VP35) gene of shrimp white spot syndrome virus and characterization of the motif important for targeting VP35 to the nuclei of transfected insect cells. Virology 293: 44-53, 2002.
17. Cheng W, Chen JC. Effects of pH, temperature and salinity on immune parameters of the freshwater prawn Macrobrachium rosenbergii. Fish &
Shellfish Immunology 10: 387-391, 2000.
18. Cheng W, Tung YH, Liu CH, Chen JC. Molecular cloning and characterisation of cytosolic manganese superoxide dismutase (cytMn-SOD) from the giant freshwater prawn Macrobrachium rosenbergii. Fish Shellfish Immunol 20:
438-49, 2006.
19. Chu FF. The Human Glutathione-Peroxidase Genes Gpx2, Gpx3, and Gpx4 Map to Chromosome-14, Chromosome-5, and Chromosome-19, Respectively.
Cytogenetics and Cell Genetics 66: 96-98, 1994.
20. Chu FF, Esworthy RS, Ho YS, Bermeister M, Swiderek K, Elliott RW. Expression and chromosomal mapping of mouse Gpx2 gene encoding the gastrointestinal form of glutathione peroxidase, GPX-GI. Biomedical and
Environmental Sciences 10: 156-162, 1997.
21. Cotgreave IA, Gerdes R, Schuppe-Koistinen I, Lind C. S-glutathionylation of glyceraldehyde-3-phosphate dehydrogenase: role of thiol oxidation and catalysis by glutaredoxin. Methods Enzymol 348: 175-82, 2002.
22. Dizdaroglu M, Gajewski E, Reddy P, Margolis SA. Structure of a Hydroxyl Radical Induced DNA Protein Cross-Link Involving Thymine and Tyrosine in Nucleohistone. Biochemistry 28: 3625-3628, 1989.
23. Dowell FJ, Hamilton CA, McMurray J, Reid JL. Effects of a xanthine
24
oxidase/hypoxanthine free radical and reactive oxygen species generating system on endothelial function in New Zealand white rabbit aortic rings. J
Cardiovasc Pharmacol 22: 792-7, 1993.
24. Downs CA, Fauth JE, Woodley CM. Assessing the health of grass shrimp (Palaeomonetes pugio) exposed to natural and anthropogenic stressors: a molecular biomarker system. Mar Biotechnol (NY) 3: 380-97, 2001.
25. Eaton P. Protein thiol oxidation in health and disease: techniques for measuring disulfides and related modifications in complex protein mixtures.
Free Radic Biol Med 40: 1889-99, 2006.
26. Fenton HJH. Oxidation of tartaric acid in presence of iron. J. Chem. Soc., Trans., 65: 899-910, 1894.
27. Fridovich I. Oxygen toxicity: a radical explanation. J Exp Biol 201: 1203-9, 1998.
28. Fujii T, Endo T, Fujii J, Taniguchi N. Differential expression of glutathione reductase and cytosolic glutathione peroxidase, GPX1, in developing rat lungs and kidneys. Free Radical Research 36: 1041-1049, 2002.
29. Garcia-Triana A, Zenteno-Savin T, Peregrino-Uriarte AB, Yepiz-Plascencia G.
Hypoxia, reoxygenation and cytosolic manganese superoxide dismutase (cMnSOD) silencing in Litopenaeus vannamei: effects on cMnSOD transcripts, superoxide dismutase activity and superoxide anion production capacity. Dev
Comp Immunol 34: 1230-5, 2010.
30. Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis? Nature
Reviews Cancer 4: 891-899, 2004.
31. Godeas C, Sandri G, Panfili E. Distribution of Phospholipid Hydroperoxide Glutathione-Peroxidase (Phgpx) in Rat Testis Mitochondria. Biochimica Et
Biophysica Acta-Biomembranes 1191: 147-150, 1994.
32. Gomez-Anduro GA, Ascencio-Valle F, Peregrino-Uriarte AB, Campa-Cordova A, Yepiz-Plascencia G. Cytosolic manganese superoxide dismutase genes from the white shrimp Litopenaeus vannamei are differentially expressed in response to lipopolysaccharides, white spot virus and during ontogeny. Comp
Biochem Physiol B Biochem Mol Biol 162: 120-5, 2012.
33. Gomez-Anduro GA, Barillas-Mury CV, Peregrino-Uriarte AB, Gupta L, Gollas-Galvan T, Hernandez-Lopez J, Yepiz-Plascencia G. The cytosolic manganese superoxide dismutase from the shrimp Litopenaeus vannamei:
molecular cloning and expression. Dev Comp Immunol 30: 893-900, 2006.
34. Goncalves-Soares D, Zanette J, Yunes JS, Yepiz-Plascencia GM, Bainy AC.
Expression and activity of glutathione S-transferases and catalase in the shrimp Litopenaeus vannamei inoculated with a toxic Microcystis aeruginosa
25
strain. Marine Environmental Research 75: 54-61, 2012.
35. Gopalakrishna R, Jaken S. Protein kinase C signaling and oxidative stress. Free
Radic Biol Med 28: 1349-61, 2000.
36. Green DE. The reduction potentials of cysteine, glutathione and glycylcysteine.
Biochem J 27: 678-89, 1933.
37. Green DR, Reed JC. Mitochondria and apoptosis. Science 281: 1309-12, 1998.
38. Greetham D, Vickerstaff J, Shenton D, Perrone GG, Dawes IW, Grant CM.
Thioredoxins function as deglutathionylase enzymes in the yeast Saccharomyces cerevisiae. Bmc Biochemistry 11, 2010.
39. Halliwell B. Free radicals, antioxidants, and human disease: curiosity, cause, or consequence? Lancet 344: 721-4, 1994.
40. Halliwell B, Gutteridge, J.M.C. . Free radicals in Biology and Medicine. New
York: Oxford Univ. Press., 1999.
41. Hill BG, Bhatnagar A. Role of glutathiolation in preservation, restoration and regulation of protein function. IUBMB Life 59: 21-6, 2007.
42. Holmgren A. Thioredoxin catalyzes the reduction of insulin disulfides by dithiothreitol and dihydrolipoamide. J Biol Chem 254: 9627-32, 1979.
43. Huang HT, Leu JH, Huang PY, Chen LL. A putative cell surface receptor for white spot syndrome virus is a member of a transporter superfamily. PLoS
ONE 7: e33216, 2012.
44. Huang JY, Liu WJ, Wang HC, Lee DY, Leu JH, Tsai MH, Kang ST, Chen IT, Kou GH, Chang GD, Lo CF. Penaeus monodon Thioredoxin Restores the DNA Binding Activity of Oxidized White Spot Syndrome Virus IE1. Antioxid Redox Signal, 2012.
45. Jakob U, Muse W, Eser M, Bardwell JC. Chaperone activity with a redox switch.
Cell 96: 341-52, 1999.
46. Jones DP, Eklow L, Thor H, Orrenius S. Metabolism of hydrogen peroxide in isolated hepatocytes: relative contributions of catalase and glutathione peroxidase in decomposition of endogenously generated H2O2. Arch Biochem
Biophys 210: 505-16, 1981.
47. Jose S, Jayesh P, Sudheer NS, Poulose G, Mohandas A, Philip R, Singh ISB.
Lymphoid organ cell culture system from Penaeus monodon (Fabricius) as a platform for white spot syndrome virus and shrimp immune-related gene expression. Journal of Fish Diseases 35: 321-334, 2012.
48. Kang JG, Paget MS, Seok YJ, Hahn MY, Bae JB, Hahn JS, Kleanthous C, Buttner MJ, Roe JH. RsrA, an anti-sigma factor regulated by redox change. EMBO J 18:
4292-8, 1999.
49. Kang ST, Leu JH, Wang HC, Chen LL, Kou GH, Lo CF. Polycistronic mRNAs and
26
internal ribosome entry site elements (IRES) are widely used by white spot syndrome virus (WSSV) structural protein genes. Virology 387: 353-363, 2009.
50. Kanzok SM, Fechner A, Bauer H, Ulschmid JK, Muller HM, Botella-Munoz J, Schneuwly S, Schirmer R, Becker K. Substitution of the thioredoxin system for glutathione reductase in Drosophila melanogaster. Science 291: 643-6, 2001.
51. Kirkman HN, Gaetani GF. Catalase: a tetrameric enzyme with four tightly bound molecules of NADPH. Proc Natl Acad Sci U S A 81: 4343-7, 1984.
52. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Mechanisms of protection of catalase by NADPH. Kinetics and stoichiometry. J Biol Chem 274: 13908-14, 1999.
53. Kiruthiga C, Rajesh S, Rashika V, Priya R, Narayanan RB. Molecular cloning, expression analysis and characterization of peroxiredoxin during WSSV infection in shrimp Fenneropenaeus indicus. J Invertebr Pathol 109: 52-8, 2012.
54. Koharyova M, Kolarova M. Oxidative stress and thioredoxin system. Gen
Physiol Biophys 27: 71-84, 2008.
55. Koppenol WH. The Haber-Weiss cycle--70 years later. Redox Rep 6: 229-34, 2001.
56. Kristal BS, Chen J, Yu BP. Sensitivity of mitochondrial transcription to different free radical species. Free Radic Biol Med 16: 323-9, 1994.
57. Kroll JS, Langford PR, Wilks KE, Keil AD. Bacterial [Cu,Zn]-superoxide dismutase: phylogenetically distinct from the eukaryotic enzyme, and not so rare after all! Microbiology 141 ( Pt 9): 2271-9, 1995.
58. Kuppusamy P, Zweier JL. Characterization of free radical generation by xanthine oxidase. Evidence for hydroxyl radical generation. J Biol Chem 264:
9880-4, 1989.
59. Laurent TC, Moore EC, Reichard P. Enzymatic Synthesis of Deoxyribonucleotides. Iv. Isolation and Characterization of Thioredoxin, the Hydrogen Donor from Escherichia Coli B. J Biol Chem 239: 3436-44, 1964.
60. Le Moullac G, Soyez C, Saulnier D, Ansquer D, Avarre JC, Levy P. Effect of hypoxic stress on the immune response and the resistance to vibriosis of the shrimp Penaeus stylirostris. Fish & Shellfish Immunology 8: 621-629, 1998.
61. Leichert LI, Jakob U. Protein thiol modifications visualized in vivo. PLoS Biol 2:
e333, 2004.
62. Leslie NR, Bennett D, Lindsay YE, Stewart H, Gray A, Downes CP. Redox regulation of PI 3-kinase signalling via inactivation of PTEN. EMBO J 22:
5501-10, 2003.
63. Leu JH, Chen LL, Lin YR, Kou GH, Lo CF. Molecular mechanism of the
27
interactions between white spot syndrome virus anti-apoptosis protein AAP-1 (WSSV449) and shrimp effector caspase. Developmental and Comparative
Immunology 34: 1068-1074, 2010.
64. Leu JH, Wang HC, Kou GH, Lo CF. Penaeus monodon caspase is targeted by a white spot syndrome virus anti-apoptosis protein. Developmental and
Comparative Immunology 32: 476-486, 2008.
65. Leu JH, Yang F, Zhang X, Xu X, Kou GH, Lo CF. Whispovirus. Curr Top Microbiol
Immunol 328: 197-227, 2009.
66. Levine RL. Carbonyl modified proteins in cellular regulation, aging, and disease. Free Radic Biol Med 32: 790-6, 2002.
67. Lin YC, Lee FF, Wu CL, Chen JC. Molecular cloning and characterization of a cytosolic manganese superoxide dismutase (cytMnSOD) and mitochondrial manganese superoxide dismutase (mtMnSOD) from the kuruma shrimp Marsupenaeus japonicus. Fish Shellfish Immunol 28: 143-50, 2010.
68. Linke K, Jakob U. Not every disulfide lasts forever: disulfide bond formation as a redox switch. Antioxid Redox Signal 5: 425-34, 2003.
69. Liu CH, Tseng MC, Cheng W. Identification and cloning of the antioxidant enzyme, glutathione peroxidase, of white shrimp, Litopenaeus vannamei, and its expression following Vibrio alginolyticus infection. Fish Shellfish Immunol 23: 34-45, 2007.
70. Liu KF, Yeh MS, Kou GH, Cheng W, Lo CF. Identification and cloning of a selenium-dependent glutathione peroxidase from tiger shrimp, Penaeus monodon, and its transcription following pathogen infection and related to the molt stages. Dev Comp Immunol 34: 935-44, 2010.
71. Liu Y, Wang WN, Wang AL, Wang JM, Sun RY. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei (Boone, 1931) exposed to acute salinity changes. Aquaculture 265: 351-358, 2007.
72. Lo CF, Ho CH, Peng SE, Chen CH, Hsu HC, Chiu YL, Chang CF, Liu KF, Su MS, Wang CH, Kou GH. White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Diseases of
Aquatic Organisms 27: 215-225, 1996.
73. Lo CF, Kou GH. Virus-associated white spot syndrome of shrimp in Taiwan: A review. Fish Pathology 33: 365-371, 1998.
74. Malek RL, Sajadi H, Abraham J, Grundy MA, Gerhard GS. The effects of temperature reduction on gene expression and oxidative stress in skeletal muscle from adult zebrafish. Comparative Biochemistry and Physiology
C-Toxicology & Pharmacology 138: 363-373, 2004.
28
75. Margolis SA, Coxon B, Gajewski E, Dizdaroglu M. Structure of a Hydroxyl Radical Induced Cross-Link of Thymine and Tyrosine. Biochemistry 27:
6353-6359, 1988.
76. Martinez-Alvarez RM, Morales AE, Sanz A. Antioxidant defenses in fish: Biotic and abiotic factors. Reviews in Fish Biology and Fisheries 15: 75-88, 2005.
77. Mccord JM, Fridovic.I. Superoxide Dismutase an Enzymic Function for Erythrocuprein (Hemocuprein). Journal of Biological Chemistry 244: 6049-&, 1969.
78. Meyer Y, Buchanan BB, Vignols F, Reichheld JP. Thioredoxins and glutaredoxins:
unifying elements in redox biology. Annu Rev Genet 43: 335-67, 2009.
79. Mikulski CM, Burnett LE, Burnett KG. The effects of hypercapnic hypoxia on the survival of shrimp challenged with Vibrio parahaemolyticus. Journal of
Shellfish Research 19: 301-311, 2000.
80. Moresino RDH, Helbling EW. Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds. Marine Drugs 8: 1681-1698, 2010.
81. Munday R. Toxicity of thiols and disulphides: involvement of free-radical species. Free Radic Biol Med 7: 659-73, 1989.
82. Neale TJ, Kerjaschki D, Witztum J, Davis P, Ruger B. Reactive Oxygen Species (Ros) and Lipid-Peroxidation (Lpo) in Proteinuric Experimental Renal-Disease.
Kidney International 46: 926-926, 1994.
83. Neves CA, Santos EA, Bainy AC. Reduced superoxide dismutase activity in Palaemonetes argentinus (Decapoda, Palemonidae) infected by Probopyrus ringueleti (Isopoda, Bopyridae). Dis Aquat Organ 39: 155-8, 2000.
84. Niyogi S, Biswas S, Sarker S, Datta AG. Seasonal variation of antioxidant and biotransformation enzymes in barnacle, Balanus balanoides, and their relation with polyaromatic hydrocarbons. Marine Environmental Research 52:
13-26, 2001.
85. Nordberg J, Arner ES. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic Biol Med 31: 1287-312, 2001.
86. Orozco-Cardenas ML, Narvaez-Vasquez J, Ryan CA. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin, and methyl jasmonate. Plant Cell 13: 179-91, 2001.
87. Qiu J, Wang WN, Wang LJ, Liu YF, Wang AL. Oxidative stress, DNA damage and osmolality in the Pacific white shrimp, Litopenaeus vannamei exposed to acute low temperature stress. Comparative Biochemistry and Physiology
C-Toxicology & Pharmacology 154: 36-41, 2011.
29
88. Qiu L, Ma Z, Jiang S, Wang W, Zhou F, Huang J, Li J, Yang Q. Molecular cloning and mRNA expression of peroxiredoxin gene in black tiger shrimp (Penaeus monodon). Mol Biol Rep 37: 2821-7, 2010.
89. Rainwater R, Parks D, Anderson ME, Tegtmeyer P, Mann K. Role of cysteine residues in regulation of p53 function. Mol Cell Biol 15: 3892-903, 1995.
90. Ren Q, Zhang RR, Zhao XF, Wang JX. A thioredoxin response to the WSSV challenge on the Chinese white shrimp, Fenneropenaeus chinensis. Comp
Biochem Physiol C Toxicol Pharmacol 151: 92-8, 2010.
91. Reth M. Hydrogen peroxide as second messenger in lymphocyte activation.
Nat Immunol 3: 1129-34, 2002.
92. Rinalducci S, Murgiano L, Zolla L. Redox proteomics: basic principles and future perspectives for the detection of protein oxidation in plants. J Exp Bot 59: 3781-801, 2008.
93. Romantsev FE, Prozorovskii VN, Andrianova LE. [Isolation from Micrococcus sp. n. of a homogeneous heme-containing catalase and a crystalline protein with catalase activity]. Biokhimiia 48: 2023-7, 1983.
94. Roos G, Messens J. Protein sulfenic acid formation: from cellular damage to redox regulation. Free Radic Biol Med 51: 314-26, 2011.
95. Ruddock LW, Klappa P. Oxidative stress: Protein folding with a novel redox switch. Curr Biol 9: R400-2, 1999.
96. Samocha TM, Guajardo H, Lawrence AL, Castille FL, Speed M, McKee DA, Page KI. A simple stress test for Penaeus vannamei postlarvae. Aquaculture 165:
233-242, 1998.
97. Schwaab V, Faure J, Dufaure JP, Drevet JR. GPx3: The plasma-type glutathione peroxidase is expressed under androgenic control in the mouse epididymis and vas deferens. Molecular Reproduction and Development 51: 362-372, 1998.
98. Sies H. Strategies of antioxidant defense. Eur J Biochem 215: 213-9, 1993.
99. Sies H, Menck CFM. Singlet Oxygen Induced DNA Damage. Mutation Research 275: 367-375, 1992.
100. Sritunyalucksana K, Wannapapho W, Lo CF, Flegel TW. PmRab7 is a VP28-binding protein involved in white spot syndrome virus infection in shrimp. Journal of Virology 80: 10734-10742, 2006.
101. Tavares-Sanchez OL, Gomez-Anduro GA, Felipe-Ortega X, Islas-Osuna MA, Sotelo-Mundo RR, Barillas-Mury C, Yepiz-Plascencia G. Catalase from the white shrimp Penaeus (Litopenaeus) vannamei: molecular cloning and protein detection. Comp Biochem Physiol B Biochem Mol Biol 138: 331-7, 2004.
102. Tsai JM, Wang HC, Leu JH, Hsiao HH, Wang AHJ, Kou GH, Lo CF. Genomic and
30
proteomic analysis of thirty-nine structural proteins of shrimp white spot syndrome virus. Journal of Virology 78: 11360-11370, 2004.
103. Tsai JM, Wang HC, Leu JH, Wang AHJ, Zhuang Y, Walker PJ, Kou GH, Lo CF.
Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. Journal of Virology 80: 3021-3029, 2006.
104. Tsai YL, Lin YC, Chou PH, Teng PH, Lee PY. Detection of white spot syndrome virus by polymerase chain reaction performed under insulated isothermal conditions. J Virol Methods 181: 134-7, 2012.
105. Tsuji T, Watanabe Y, Katoh H, Sato K, Kunieda T. Cloning and mapping of the mouse Gpx2 gene encoding gastrointestinal glutathione peroxidase. Journal
of Veterinary Medical Science 60: 651-654, 1998.
106. Ursini F, Maiorino M, Brigeliusflohe R, Aumann KD, Roveri A, Schomburg D, Flohe L. Diversity of Glutathione Peroxidases. Biothiols, Pt B 252: 38-53, 1995.
107. Verlecar XN, Jena KB, Chainy GB. Biochemical markers of oxidative stress in Perna viridis exposed to mercury and temperature. Chem Biol Interact 167:
219-26, 2007.
108. Wang CH, Lo CF, Leu JH, Chou CM, Yeh PY, Chou HY, Tung MC, Chang CF, Su MS, Kou GH. Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Diseases of Aquatic
Organisms 23: 239-242, 1995.
109. Wang HC, Ko TP, Lee YM, Leu JH, Ho CH, Huang WP, Lo CF, Wang AH. White spot syndrome virus protein ICP11: A histone-binding DNA mimic that disrupts nucleosome assembly. Proc Natl Acad Sci U S A 105: 20758-63, 2008.
110. Wang HC, Wang HC, Ko TP, Lee YM, Leu JH, Ho CH, Huang WP, Lo CF, Wang AHJ. White spot syndrome virus protein ICP11: A histone-binding DNA mimic that disrupts nucleosome assembly. Proceedings of the National Academy of
Sciences of the United States of America 105: 20758-20763, 2008.
111. Wang HC, Wang HC, Leu JH, Kou GH, Wang AHJ, Lo CF. Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Developmental and Comparative Immunology 31: 672-686, 2007.
112. Wang KCHC, Kondo H, Hirono I, Aoki T. The Marsupenaeus japonicus voltage-dependent anion channel (MjVDAC) protein is involved in white spot syndrome virus (WSSV) pathogenesis. Fish & Shellfish Immunology 29: 94-103, 2010.
113. Wang WN, Li BS, Liu JJ, Shi L, Alam MJ, Su SJ, Wu J, Wang L, Wang AL. The respiratory burst activity and expression of catalase in white shrimp, Litopenaeus vannamei, during long-term exposure to pH stress. Ecotoxicology,
31
2012.
114. Warburg O. On the origin of cancer cells. Science 123: 309-14, 1956.
115. Watthanasurorot A, Jiravanichpaisal P, Soderhall I, Soderhall K. A gC1qR prevents white spot syndrome virus replication in the freshwater crayfish Pacifastacus leniusculus. J Virol 84: 10844-51, 2010.
116. Winterbourn CC, Vissers MC, Kettle AJ. Myeloperoxidase. Curr Opin Hematol 7: 53-8, 2000.
117. Yost FJ, Jr., Fridovich I. An iron-containing superoxide dismutase from Escherichia coli. J Biol Chem 248: 4905-8, 1973.
118. Zenteno-Savin T, Saldierna R, Ahuejote-Sandoval M. Superoxide radical production in response to environmental hypoxia in cultured shrimp.
Comparative Biochemistry and Physiology C-Toxicology & Pharmacology 142:
301-308, 2006.
119. Zhang Q, Li F, Zhang J, Wang B, Gao H, Huang B, Jiang H, Xiang J. Molecular cloning, expression of a peroxiredoxin gene in Chinese shrimp Fenneropenaeus chinensis and the antioxidant activity of its recombinant protein. Mol Immunol 44: 3501-9, 2007.
120. Zhang Q, Li F, Zhang X, Dong B, Zhang J, Xie Y, Xiang J. cDNA cloning, characterization and expression analysis of the antioxidant enzyme gene, catalase, of Chinese shrimp Fenneropenaeus chinensis. Fish Shellfish Immunol 24: 584-91, 2008.
121. Zheng M, Aslund F, Storz G. Activation of the OxyR transcription factor by reversible disulfide bond formation. Science 279: 1718-21, 1998.
122. Zhou J, Wang L, Xin Y, Wang WN, He WY, Wang AL, Liu YA. Effect of temperature on antioxidant enzyme gene expression and stress protein response in white shrimp, Litopenaeus vannamei. Journal of Thermal Biology 35: 284-289, 2010.
123. Zhou J, Wang WN, Wang AL, He WY, Zhou QT, Liu Y, Xu J. Glutathione S-transferase in the white shrimp Litopenaeus vannamei: Characterization and regulation under pH stress. Comparative Biochemistry and Physiology