國立臺灣大學生物資源暨農學院昆蟲學系暨研究所 碩士論文
Department and Graduate Institute of Entomology College of Bioresources and Agriculture
National Taiwan University Master Thesis
臺灣地區新入侵煙草粉蝨 Q 型生物小種族群遺傳結 構與入侵源之研究
Population genetics and the origin of new invasive biotype Q of Bemisia tabaci (Hemiptera: Aleyrodidae) in
Taiwan
蔣宜弦 Yi-Hsien Chiang
指導教授:柯俊成 博士 Advisor: Chiun-Cheng Ko, Ph.D.
中華民國 98 年 6 月
June, 2
誌謝
本論文之完成首先要感謝指導教授柯俊成老師,兩年來在學術上的指導與生 活上的關照,在實驗遇到困難時的教導與指引,使我的研究能順利進行,並在論 文寫作上給予建議,使我可以排除萬難,順利完成這本論文。
同時感謝張慧羽老師對於遺傳理論提供許多寶貴的意見,並在論文的寫作上 給予多方的建議。感謝徐堉峰老師於親緣地理關係觀念的釐清,以及王弘毅老師 對於分子演化與資料的分析指導,使我受益匪淺。並感謝及辛竹英老師對於論文 的指正與建議,使論文內容更加完整。
感謝謝佳宏博士兩年來,在實驗設計、分析方法和採集所提供的協助,使我 的論文內容更加縝密。亦要感謝楊景程學長在資料的分析上提供建議。以及東華 大學黃國靖老師帶領我走進昆蟲的領域。
煙草粉蝨樣本的獲得,感謝 Dr. S. J. Suh (National Plant Quarantine Service, Goyang, South Korea ) 在韓國採集所提供的協助。粉蝨樣本採集上的協助,亦要 感謝國立臺灣大學昆蟲學系陳俊宏、吳復生、方華德、黎淑芬等人於採集時的協 助,同時也要感謝盆花協會理事長莊瀛正,以及聖誕紅業者鍾侑煌、吳亦炳、黃 國鏘和林先生提供採集的協助。在此由衷表達謝意。
感謝黃馨論、李仁耀、洪玉靜、洪惠方、Dr. Dubey、張德斌、楊世綵、施圓 通、曹昌旺和張奕元等人於論文期間的關懷與幫助。
最重要的是要感謝我的父母與家人多年來的支持與鼓勵,使我可以順利完成 學業。感謝所有幫助過我的人。
中文摘要
煙草粉蝨 (Bemisia tabaci) 被列為世界百大入侵生物種之一,造成農業 和經濟上嚴重的危害。目前世界上共訂名超過 26 型生物小種。最近研究指 出 Q 型生物小種近年來隨著聖誕紅迅速的擴散至世界各地。入侵臺灣的 Q 型生物小種於 2006 年 2 月,首度於臺南的亞洲蔬菜研究發展中心發現。
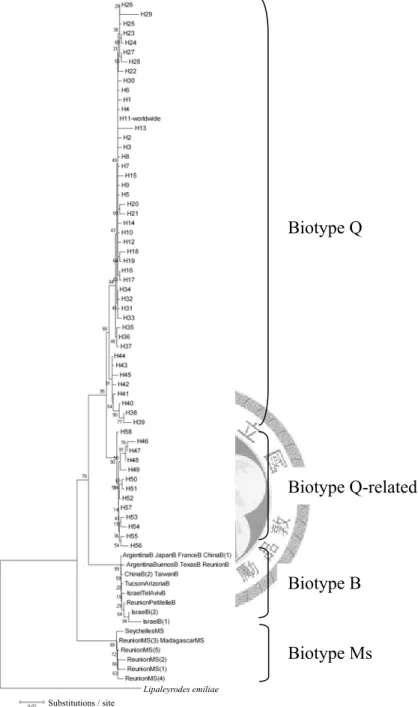
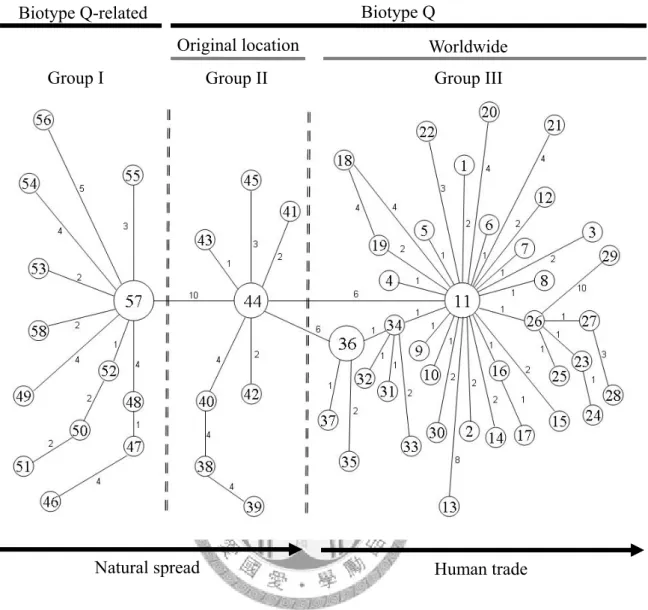
在 791 筆臺灣地區的採集資料中,僅在 7 個聖誕紅的溫室內發現到 Q 型 生物小種,推測目前還是屬於入侵的初期。本研究探討入侵來源具有重要的 意義,可做為檢疫工作上的依據,加強檢測由特定管道所進口的花卉,預防 Q 型生物小種的再次入侵。親緣地理學之研究,利用粒線體 COI 序列做為 遺傳標記,探討煙草粉蝨的 Q 型生物小種,在世界各地的遺傳差異及擴散 途徑。建構最小跨度網狀圖,發現 Q-related 型生物小種起源於中、西非洲,
而 Q 型 生 物 小 種 起 源 於 歐 洲 和 亞 洲 交 界 處 , 並 以 自 然 擴 散 的 方 式 進 入 歐 洲,而入侵世界各地的 Q 型生物小種源自於歐洲,推測是藉由人類貿易行 為擴散。利用 12 個微衛星基因座 (microsatellite loci),研究煙草粉蝨 Q 型 生物小種族群的遺傳結構和親緣關係,發現臺灣地區的 Q 型生物小種族群 可能來自兩次以上的入侵。桃園復興的族群呈現獨特的遺傳結構,顯示溫室 內的族群來自一個單一的入侵源。根據隔離分化模式的分析,臺灣地區其他 溫 室 內 各 族 群 基 因 交 流 頻 繁 , 推 測 是 隨 著 種 苗 交 換 , 而 在 各 個 溫 室 之 間 擴 散,使遺傳分化差異小。因此防治策略上,要加強歐洲進口花卉的檢疫,亦 要注意種苗交換的問題,防止 Q 型生物小種在臺灣地區的溫室之間迅速擴 散。
關鍵字:害蟲、入侵種、聖誕紅、粒線體 DNA、微衛星 DNA、親緣地理
Abstract
The sweetpotato whitefly, Bemisia tabaci, is among the top 100 invasive pests in the world. This species causes serious agricultural damages in many countries. More than 26 biotypes have been detected worldwide. Recently, biotype Q was reported as a new invasive pest globally which has spread throughout the world with the trade of poinsettia (Euphorbia pulcherimma). In 2006, biotype Q was first recorded at the World Vegetable Center in Tainan, Taiwan. After wards, 791 samples were collected in Taiwan.
Biotype Q was found in 7 poinsettia greenhouses, but not in the field. I hypothesized that it is in the early stages of invasion. Determining the origin of this invasion is important for importing quarantine. This study provides information for eradication program, and further introductions and reinvasions will hopefully be avoided by applying intensive quarantine regulations on possible invasion routes. Using mitochondrial cytochrome oxidase I (COI) sequences as a molecular marker to study phylogeography, I explored genetic differences and dispersal patterns of B. tabaci biotype Q in the world by reconstructing a minimum spanning network. The origin of biotype Q-related biotypes was in Central and West Africa. However, the origin of biotype Q was somewhere between Europe and Asia, and then it spread into Europe by natural dispersal. Recent invasion events in the world of biotype Q were from Europe.
This suggests that it spread by human trade. Moreover, I applied 12 microsatellite loci by means of multiplex polymerase chain reaction (PCR) to investigate phylogenetic relationships and the population structure of B. tabaci biotype Q populations. Results suggest that there were at least 2 invasions of Taiwan. The genetic structure of the Fusing population (Taoyuan County, northwestern Taiwan) possesses molecular characteristics, suggesting that the population initiated from an independent invasion.
Furthermore, great gene flow and slight genetic differentiation among populations in
Taiwan were shown by violating the isolation by distance expectation. The migration of individuals between greenhouses may be caused by the movement of poinsettias between greenhouses and other human activities. The program of pest management should prescribe the plantlet trade regulation to avoid further dispersal of B. tabaci Q biotype among greenhouses in Taiwan.
Keywords: pest, invasive species, poinsettia, COI sequence, microsatellite markers, phylogeography
目 錄
口試委員審定書
誌謝 I 中文摘要 II 英文摘要 IV 目錄 ……… V 表次 ……… VII 圖次 ……… VIII
壹 、 前 言 ……… 1
貳、往昔研究 ……… 4
一、煙草粉蝨種群與鑑定 ……… 4
二、煙草粉蝨 Q 型生物小種 ……… 5
三、Q 型生物小種的危害 ……… 6
四、生物入侵 ……… 8
五、粒線體 DNA 在煙草粉蝨之研究 ……… 9
六、微衛星 DNA 在煙草粉蝨之研究 ……… 11
七、臺灣地區 Q 型生物小種之研究 ……… 13
參、材料與方法 ……… 15
一、標本的採集及保存 ……… 15
二、萃取基因組 DNA ……… 15
三、測定生物小種 ……… 16
四、粒線體 (Mitochondrial) COI 基因序列之資料分析 ……… 16
五、微衛星基因座的增幅與資料分析 ……… 22
肆、結果 ……… 31
一、臺灣地區煙草粉蝨 Q 型生物小種採集和分布情形 ……… 31
二、粒線體 (Mitochondrial) COI 基因序列之資料分析 ……… 33
三、微衛星基因座的增幅與資料分析 ……… 40
伍、討論 ……… 52
一、臺灣地區煙草粉蝨 Q 型生物小種分布情形 ……… 52
二、親緣關係之分析 ……… 54
三、親緣地理學之探討 ……… 56
四、遺傳結構之探討 ……… 57
五、入侵模式和擴散方式之探討 ……… 58
六、監測防治之建議 ……… 60
陸、結論 ……… 63
柒、參考文獻 ……… 65
捌、附錄 ……… 74
附錄一、煙草粉蝨 Q 型生物小種採集記錄 ……… 74
附錄二、臺灣地區煙草粉蝨 Q 型生物小種粒線體 COI 序列 比 對 結 果 ……… 77
附錄三、利用 ∆K 選出最適合分群數 K 值為 8 ……… 78
表 次
表一 供試煙草粉蝨 Q 型生物小種樣本 ……… 17
表二 煙草粉蝨生物小種專一性引子組序列及其黏合溫度 ……… 20
表三 供試之煙草粉蝨微衛星基因座之引子組 ……… 25
表四 臺灣地區煙草粉蝨 Q 型生物小種採集記錄 ……… 32
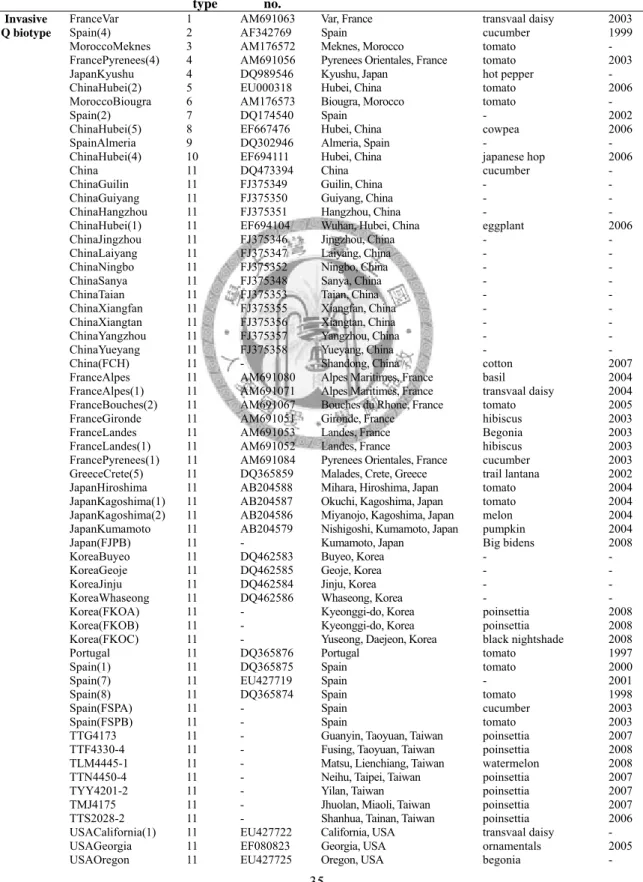
表五 本研究供試之煙草粉蝨 COI 序列 ……… 35
表六 族群遺傳結構分析 ……… 42
表七 煙草粉蝨族群間遺傳分化指數 ……… 43
表八 瓶頸效應檢測 ……… 45
圖 次
圖一 臺灣地區煙草粉蝨 Q 型生物小種採集位置圖 ……… 18 圖二 國外煙草粉蝨 Q 型生物小種採集位置圖 ……… 19 圖三 煙草粉蝨粒線體 COI 序列,以鄰近法所建構之親緣關係樹 ………… 38 圖四 煙草粉蝨 Q 型生物小種之最小跨度網狀圖 ……… 39 圖五 煙草粉蝨微衛星基因座,以鄰近法建構之親緣關係樹 ……… 46 圖六 鄰近法建構臺灣地區 4 個族群間微衛星基因座之親緣關係樹 ……… 48 圖七 微衛星基因座之貝氏分群分析 ……… 51
壹、前言
煙 草 粉 蝨 (Bemisia tabaci) 為 半 翅 目 (Hemiptera) , 胸 喙 亞 目 (Sternorrhyncha),粉蝨科 (Aleyrodidae),粉蝨亞科 (Aleyrodinae),伯粉蝨 屬 (Bemisia) 的昆蟲。歷史記錄上,在希臘的煙草上首度被描述,並命名 為 Aleyrodes tabaci (Gennadius, 1889),往後亦有許多命名事件被討論,其 中最重要的是 Takahashi (1936) 將 A. tabaci 併入伯粉蝨屬中,此乃現今 Bemisia tabaci (Gennadius) 的由來。煙草粉蝨性別決定為單雙套機制,單 套 (haploid) 為雄蟲,雙套 (diploid) 為雌蟲。雌蟲產下的卵無論受精與否 皆可孵化,受精卵產生雌蟲,未受精則產生雄蟲。煙草粉蝨至今被發表超 過 26 型 生 物 小 種 , 區 分 成 12 個 遺 傳 群 (Perring, 2001; Boykin et al., 2007; Hsieh et al., 2008),是農業及花卉上重要的害蟲。危害的方式包括銀 葉病及傳播多種植物病毒,在墨西哥、印尼和巴西等國家,嚴重危害樹薯 和紅蕃椒等農作物,使大批農民失去工作機會,造成嚴重的經濟及糧食危 機,成蟲和若蟲以刺吸式口器攝食植物葉片汁液,大量的攝食容易導致植 物葉片衰弱,蟲體分泌的蜜露,容易引發煤煙病,密度高時葉片呈現黑色,
嚴重影響花卉的美觀與價格 (Boykin, 2007; De Barro et al., 2008)。
煙草粉蝨種群中,以 B 和 Q 型生物小種最受到各國的關注,兩者除 了有相近的親緣關係和生存環境,也都演化出較高的抗藥性,因此成為世 界上重要的農業害蟲。B 型生物小種自從 1985 年大發生以來,已擴散至 世界各地,主要是因為國際間觀賞性作物聖誕紅盆栽的貿頻繁所傳播。Q 型 生物小種相對於 B 型生物小種,在繁殖力、發育速度和生殖行為上均位居 劣 勢 , 但 是 因 為 Q 型 生 物 小 種 對 於 百 利 普 芬 (pyriproxyfen) 、 益 達 胺 (imidacloprid) 等化學殺蟲劑有較高的抗藥性,而且對於極端的溫度和乾燥 的環境有較高的耐受度,所以在溫室的環境中是優勢的生物小種 (Horowitz et al., 2005)。荷蘭及地中海地區國家是溫室花卉的培育中心,對外輸出的
花卉佔有率達 50% 以上,隨著花卉及蔬果貿易日漸頻繁,原本只分布在 地中海區域的 Q 型生物小種,正隨著花卉及種苗的貿易迅速的擴散至世界 各地 (Dalton, 2006)。
煙草粉蝨目前臺灣地區共記錄有 4 型生物小種,其中 An 和 Nauru 為 本 土 的 生 物 小 種 , 而 B 和 Q 為 入 侵 的 生 物 小 種 (Hsieh et al., 2005, 2006, 2007, 2008)。Q 型生物小種採集記錄,最早發現於臺南的亞洲蔬菜研 究發展中心,隨著盆栽的來源亦在苗栗卓蘭的溫室中,調查到 Q 型生物小 種的煙草粉蝨 (Hsieh et al., 2007)。生物入侵的狀態可分為四個主要時期;
進口 (import)、引入 (introduction)、立基 (establishment) 和危害 (pest),
根據入侵生物學之十分之一法則 (Williamson and Fitter, 1996),每一個時期 成功進入另一個時期的成功機率為 10%,所以外來生物從入侵而能造成危 害是機率很低的事件,通常需要一段時間內多次的入侵才有可能成為有害 生物。入侵之煙草粉蝨生物小種常藉著多次入侵來達到成功立足,東亞地 區之入侵 B 型生物小種存在多次入侵的現象,西北太平洋地區之 Q 型生 物小種亦有多次入侵的現象 (Hsieh et al., 2006, 2007)。最近的研究顯示,Q 型生物小種在臺灣地區,目前介於進口和引入階段,尚未建立野外族群亦 未成為有害生物 (Hsieh et al., 2008),臺灣地區之新入侵 Q 型生物小種,
臺灣地區目前僅在聖誕紅溫室內偵測到新入侵 Q 型生物小種,而且樣本間 之粒線體 COI 序列相似度為 100%,推測為單一的入侵事件 (Hsieh et al., 2007, 2008)。然而 COI 序列的變異度不高,解析力不夠,臺灣地區 Q 型 生物小種是否也存在多次入侵的現象,仍然有討論的空間。
近年來分子生物學與生物技術快速發展,族群分子遺傳學在近十年來 得以快速發展。紅火蟻的入侵與偵測中,透過微衛星 DNA 的遺傳結構檢 視,可推測不同的入侵來源、擴散模式,廣泛於應用檢測入侵生物的族群 遺傳結構,推測入侵的歷史事件 (Yang et al., 2008)。此外,選擇適當的分
子標記,對煙草粉蝨做親緣地理學的研究,可以重建族群的擴散與分化,
並探討人為事件對族群入侵之影響。微衛星基因座由於多型性高、序列長 度短、共顯性遺傳且多為中性演化,常被用來研究族群內個體間或族群間 的遺傳結構,個體之間可偵測親子關係或評估親緣關係,在族群內檢視遺 傳多樣性及遺傳結構,可用來推測交配模式 (Winters and Waser, 2003)、推 判族群是否遭受近期的瓶頸效應 (Cornuet and Luikart, 1996),族群間可評 估基因交流的程度 (Hampton et al., 2004),推測族群擴散的模式 (Dalmon et al., 2007)。
本研究從族群遺傳及遺傳多樣性的角度,以粒線體 COI 序列,探討 Q 型生物小種的起源與擴散,並利用微衛星 DNA 基因座做為分子標記,判 定目前臺灣地區入侵事件的情形,包含入侵次數以及在臺灣地區溫室之間 的遷移。希望可以提供防疫檢疫及蟲害管理的建議,避免 Q 型生物小種再 次入侵臺灣,減少在野外建立族群的機會。
貳、往昔研究
煙草粉蝨寄主植物範圍廣泛,除了吸取植物汁液,同時能傳播超過 110 種的植物病毒 (Jones, 2003),引起多種植物的病徵,造成糧食及經濟作物 上嚴重的損失,近年來由於農業和花卉貿易頻繁而迅速擴散至世界各地,
目前廣泛分布於熱帶、亞熱帶及溫帶地區,是重要的農業害蟲之一。
一、煙草粉蝨種群與鑑定
煙草粉蝨為一種群,由於各群之間有遺傳上的差異,但是形態上無法 區分,所以分類的方式仍然沒有共識,現今以生物小種 (biotype) 和遺傳群 (genetic group) 這兩種方式,最廣受研究學者的使用。世界各地的煙草粉蝨 最早被分為 24 型生物小種 (Perring, 2001)。後來由於陸續的發表新型生 物小種,以及刪除同物異名的情況,增加及刪改後分為至少 26 型生物小 種 (Hsieh et al., 2008), 最 新 的 研 究 報 告 則 認 為 約 有 41 型 的 生 物 小 種 (Rabello et al., 2008)。 最 近 許 多 的 研 究 傾 向 分 為 12 個 遺 傳 群 (genetic group) (Boykin et al., 2007)。在農學方面目前仍然以生物小種這種說法為主 (Hsieh et al., 2008)。
生 物 小 種 的 鑑 定 早 期 以 RAPD-PCR (random amplified polymorphic DNA-polymerase chain reaction) 的方式進行 (Perring et al., 1993; De Barro and Driver, 1997; Guirao et al., 1997),但是重現性上比較不穩定,且在結果 的 判 讀 耗 時 費 工 。 粒 線 體 DNA 的 16S 及 COI 序列與核糖體 DNA 的 ITS 序列分析比對,亦可應用於煙草粉蝨生物小種的分析 (Frohlich et al., 1999; De Barro et al., 2000; Hsieh et al., 2005),但是定序的過程相當耗時且 成本高。害蟲的鑑定講求快速鑑定,因此最近利用粒線體 DNA 進行序列 特徵化增幅區域 (sequence characterized amplified region, SCAR) 引子的設 計,直接由 PCR 的結果,可以更快速而且準確的測定不同的生物小種 (Ko et al., 2007)。多對引子聚合酶連鎖反應 (Multiplex PCR) 和即時定量聚合
酶連鎖反應 (real-time PCR),將可提供更快速的生物小種鑑定 (Hsieh and Ko, unpublished)。
二、煙草粉蝨 Q 型生物小種
所有生物小種中,遺傳結構與 Q 型生物小種接近的,有 Q-related、
Ms 和 B 型生物小種 (Boykin, 2007)。2004 年辛巴威 (Zimbabwe) 及喀麥 隆 (Cameroon) 所發現的三個單倍型,以及 2006 年在奈及利亞 (Nigeria) 和迦納 (Ghana) 所發現的九個單倍型 (Berry et al., 2004; De La Rua et al., 2006),由於遺傳結構最接近 Q 型生物小種,但在南瓜葉片上會引發銀葉 病 , 生 理 上 不 同 於 Q 型生物小種,所以訂名為 Q-related 型生物小種。
Ms 型生物小種分布於,印度洋的西南方東非留尼旺島 (Reunion)、馬達加 斯加 (Madagascar) 和塞席爾群島 (Seychelles) 等地,經過實驗發現不但能 引發南瓜銀葉病,同時也能傳播蕃茄黃化捲葉病毒 (tomato yellow leaf curl virus, TYLCV) (Delatte et al., 2005)。現今農業和經濟嚴重損失的包括 B 和 Q 型生物小種,B 型生物小種就是銀葉粉蝨 (Bemisia argentifolii),因為寄 主植物廣泛,又能傳播許多植物病毒最受到重視,並且經由人類的貿易活 動,快速的擴散到全世界,對於熱帶、亞熱帶、乾燥及地中海型氣候地區 作物造成嚴重危害 (Brown et al., 1995; Frohlich et al., 1999; De Barro et al., 2000; Perring, 2001); Q 型生物小種分布於地中海、中東、北非地區 (Brown, 2000) , 最 早 在 伊 比 利 半 島 地 區 發 現 , 於 1997 年 首 度 在 西 班 牙 被 發 表 (Guiraro et al., 1997),2005 年之前環地中海的國家,陸續發表有關 Q 型 生物小種的相關報告,後來研究陸續證實 Q 型生物小種,已遍佈黎巴嫩半 島、薩丁尼亞、突尼西亞、克羅埃西亞、阿爾及利亞、意大利、德國、以 色列、塞普路斯、埃及等國家 (Horowitz et al., 2003, 2005; Simon et al., 2007),在 2005~2009 年間,中國、日本、韓國、美國、瓜地馬拉和墨西 哥,也相繼發表及報導有關 Q 型生物小種的採集記錄 (Chu et al., 2005;
Dennehy et al., 2005; Lee et al., 2005; Zhang et al., 2005; Ueda and Borwn, 2006; Borwn, 2007; Martinez-Carrillo and Brown, 2007; Bethke et al., 2009)。
三、Q 型生物小種的危害
Q 型生物小種相較於 B 型生物小種,雌蟲繁殖力、雌蟲的存活率及 發育的速度,都位居劣勢,且在實驗室的環境中,B 型生物小種可以競爭 並取代 Q 型生物小種,成為優勢小種 (Pascual and Callejas, 2004)。但是 在少數野外地區 Q 型生物小種仍具有生存優勢,以西班牙地區為例,Q 型 生物小種在部分的雜草和農作物上,比 B 型生物小種更具有競爭優勢,不 但沒有被取代,反而在蕃茄、辣椒和萵苣等作物上比 B 型生物小種造成更 嚴重的危害,因為在三種雜草 Malva parviflora L.、Capsella bursa-pastoris L.、Brassica kaber (DC) 上,Q 型生物小種雌蟲產卵量、化蛹數、成蟲羽 化率等,顯著高於 B 型生物小種; 此外,在 26℃ 的情況下,Q 型生物 小種在雜草曼陀羅
(
Datura stramonium) 和龍葵 (Solanam nigrum) 上,從卵發育到成蟲歷期 (分別為 22 天、20 天) 明顯短於 B 型生物小種所 需 的 時 間 (分別為 23 天、22 天),這些雜草是 Q 型生物小種的重要寄 主 , 為 該 型 生 物 小 種 為 害 農 作 物 提 供 大 量 蟲 源 , 使 其 具 有 更 強 的 入 侵 性 (Muniz, 2000)。部分的農作物也發現 Q 型生物小種的競爭優勢,在甜椒上 17℃、33℃ 時平均世代歷期 (分別為 46 天、17 天) 比 B 型生物小種的 (分別為 49 天、18 天) 要短 (Muniz and Nombela, 2001); 在部分品種的蕃 茄上,Q 型生物小種比 B 型生物小種具有更強的危害性 (Nombela et al., 2001)。Q 型生物小種還具有很強的傳毒能力,使蕃茄黃葉捲葉撒丁病毒 (Tomato yellow leaf curl Sardinia virus, TYLCSV-ES) 可以在蕃茄和兩種雜 草上 (D. stramonium、S. nigurm) 相互傳播 (Jiang et al., 2004)。
Q 型生物小種最大潛在的危害,是對於部分化學殺蟲劑具有很高的抗 性,而 Q 型生物小種對化學殺蟲劑的抗性,對於族群的增長相當有利,研
究 已 發 現 Q 型 生 物 小 種 對 於 百 利 普 芬 (pyriproxyfen) 、 益 達 胺 (imidacloprid) 等化學殺蟲劑有較高的抗藥性 (Horowitz et al., 2005)。已有 不 少 實 驗 證 明 B 型 生 物 小 種 很 難 控 制 和 防 治 , 但 北 美 植 物 保 護 組 織 (North American Plant Protection Organizations, NAPPO) 植物檢疫預警系統 (Phytosanitary Alert System) 的資料認為 Q 型生物小種的危害性超過 B 型生物小種。最近的研究發現,在沒有殺蟲劑的選汰壓力下,Q 型生物小 種對於新尼古丁類 (neonicotinoid) 殺蟲劑的抗性比較穩定,而 B 型生物 小種對新尼古丁類的抗性則會急遽下降 (Rauch and Nauen, 2003)。在意大 利和德國採集的樣本,都可以發現對於新尼古丁類具有很高的抗性,在西 班牙 Almeria 地區所採集的樣本尤其突出,而且在沒有殺蟲劑的選汰下,
仍然可以保持至少 2 年的抗性 (Nauen et al., 2002)。此外,對 Q 型生物 小種進行新尼古丁類殺蟲劑的選汰,亦能夠對於百利普芬產生抗藥性,由 於上述兩種殺蟲劑的使用,均可以引起 Q 型生物小種產生抗性,導致 Q 型生物小種能夠在這些使用殺蟲劑的地區存活 (Horowitz et al., 2005)。
聖誕紅溫室環境中,因為 Q 型生物小種,具有較高的殺蟲劑抗性,對 於熱和乾燥也有較好的耐受度,所以在溫室中歷經殺蟲劑,以及環境的人 為選汰(artificial selection),亦可以競爭取代 (competitive displacement) B 型生物小種,成為優勢小種 (Horowitz et al., 2005; Dalton, 2006)。長期的施 用農藥,容易使煙草粉蝨產生抗藥性,在去除農藥選汰的壓力後,B 型生 物小種的抗藥性則會急遽下降,然而 Q 型生物小種對於部分農藥,具有穩 定的抗藥性,如新尼古丁類藥劑,即使不再持續使用,Q 型生物小種仍然 可以延續數個世代具有抗性基因 (Rauch and Nauen, 2003)。此外,Q 型生 物小種相對於 B 型生物小種,對於極端的溫度有較高的耐受性,在歐洲有 逐 漸 向 北 擴 散 的 現 象 , 在 野 外 也 有 較 高 的 存 活 率 (Bonato et al., 2007;
Dalmon et al., 2008)。
四、生物入侵 (biological invasion)
外來入侵物種 (alien invasive species) 是指從自然分布區通過有意或 無意的人類活動而被引入、在當地的自然或半自然生態系統中建立族群、
給當地的生態系統或景觀造成損害或影響的物種。生物的入侵事件,有三 種主要的因素:(一) 人為的活動,導致生物的遷入,如:貿易和放生。(二) 入侵者本身遺傳條件改變,更適合入侵的環境。 (三) 環境的改變,造成生 物的遷移,如:全球暖化 (global warming) 和溫室 (glasshouse),使得原本 只 出 現 在 熱 帶 地 區 的 生 物 , 可 以 往 北 遷 移 至 亞 熱 帶 或 溫 帶 (Kiritani, 2006)。目前 Q 型生物小種,也隨著聖誕紅花卉,快速的在各個國家之間 擴散,由於人為的活動和貿易,成為一個聖誕節的入侵害蟲 (the Christmas invasion) (Dalton, 2006)。此外,人類為了花卉良好的培育環境所興建的溫 室,使 Q 型生物小種可以擴散到緯度更高的地方 (Dalmon et al., 2008)。
選 汰 壓 下 , 物 種 適 應 環 境 的 方 式 分 為 K- 策略者 (K-strategist) 與 r- 策略者 (r-strategist),煙草粉蝨屬於 r- 策略者,具有生殖率高、發育快速、
個體小、生活史短的特性,族群的密度變動大,即使少數個體,因有迅速 增殖的能力,只要經過少數世代,即可達高密度的族群數量。煙草粉蝨同 時具有隨著寄主植物擴散和遷移能力。這些適應生境的對策,使煙草粉蝨 成為成功的入侵生物。
以入侵生物危害程度而言,中國是目前世界上受害最嚴重的國家。根 據統計入侵中國的外來物種有 400 多種,其中造成嚴重危害的有 100 多 種,而國際自然保育聯盟所公布的百大入侵種中,中國地區調查記錄已有 50 多種,外來入侵種的問題已經受重視,尤其近年來,已經觀察到入侵生 物種的入侵數量增多、入侵頻率加快、蔓延範圍擴大、發生的危害加劇以 及經濟損失加重等趨勢,且中國在加入 WTO 後,所面對的外來生物種的 入侵問題更加嚴厲 (chu et al., unpublished)。
入侵成功的生物通常具有比較高的環境適應力,分布廣泛的結果導致 難以推測地理位置及入侵來源 (Lodge, 1993),因此,外來種入侵後的生物 學研究,著重在物種的入侵來源、傳播途徑、寄主,選擇有效的化學殺蟲 劑、天敵,以及了解入侵後生物學特性的變化,上述的研究對於提出有效 的控制策略均有重要的意義。分子生物學,近年來快速的發展,除對演化 生物學提供有力的證據,對於入侵生物的管理亦能提供防治策略 (Le Roux and Wieczorek, 2009)。紅火蟻的入侵與偵測中,透過遺傳結構的檢視,嘉 義和桃園的族群最有可能從美國入侵,但由於族群間分子遺傳變異相當顯 著,推測臺灣地區至少發生兩次獨立之入侵事件,入侵來源最有可能是由 美國入侵,但來自於美國境內的不同入侵源 (Yang et al., 2008)。
煙草粉蝨 B 型生物小種,在 1980 年代即藉由聖誕紅的貿易擴散到全 世界,並造成嚴重的危害,在入侵的過程中,B 型生物小種存在著非對稱 性的交配互動,使煙草粉蝨可以很快速在新棲地建立族群,煙草粉蝨的性 別決定機制為單雙套系統,入侵的生物小種會增加交配頻率,使卵受精率 增加以產生大量雌性子代,同時入侵生物小種的雄性,亦會干擾本土生物 小種的交配行為,使本土生物小種的交配率和卵受精率下降,使後代雌性 減少,並在 3~5 年的時間完成競爭取代 (Liu et al., 2007)。國際自然保育 聯 盟 (International Union for the Conservation of Nature and Natural Resources, IUCN) 亦 將 煙 草 粉 蝨 列 為 世 界 百 大 入 侵 生 物 種 之 一 (http://www.iucn.org)。
五、粒線體 DNA 在煙草粉蝨之研究
DNA 序 列 是 研 究 生 物 演 化 歷 史 的 重 要 工 具 之 一 (Nei and Kumar, 2000)。昆蟲的粒線體 DNA 為環狀的 DNA 分子,長度約為 16~20kb 左 右。粒線體 DNA 具有以下特點:(一) 母系遺傳無重組過程,常做為親緣 關係的分析;(二) 單套染色體,無基因重組的過程,遺傳差異僅來自於基
因突變;(三) 某些類群物種,突變的速率比核 DNA 快,適合做為生物小 種及近緣種群的研究。
粒線體 DNA 常用來計算族群之間的遺傳差異推測親緣關係。針對烏 干達所發現的兩個煙草粉蝨族群做研究,發現與 B 型生物小種之間有 8%
的遺傳差異,並推測族群為非 B 型的生物小種,而兩個族群也分別與尚比 亞 (Zambia) 和喀麥隆 (Cameroon),有高度的遺傳相關性,藉由此項證據 推論兩個族群之間有不同的來源 (Legg et al., 2002)。此外,粒線體 DNA 也 被利用來研究煙草粉蝨的種群,結果發現煙草粉蝨是一個複合群,全球可 分為 8 大群,分別為近撒哈拉沙漠區、新世界區、地中海區、中東區、亞 洲一區、亞洲二區、澳洲區和大洋區,並推論 B 型生物小種由舊世界起源,
發 源 地 自 於 乾 燥 區 之 中 東 、 阿 拉 伯 半 島 或 是 北 非 等 地 區 (Frohlich et al., 1999; De Barro et al., 2000)。最近的研究,利用粒線體 COI 序列進行貝氏 分 析 , 將 全 世 界 的 煙 草 粉 蝨 分 成 12 個 遺 傳 群 (genetic groups), 分 別 是 Mediterranean/Asia Minor/Africa、Mediterranean、Indian Ocean、sub-Saharan Africa silverleafing (Q-related)、Asia I、Australia、China、Asia II、Italy、
New World、sub-Saharan Africa non-silverleafing 和 Uganda sweet potato (Boykin et al., 2007)。希臘的 Q 型生物小種雖然屬於 Mediterranean/Asia Minor/Africa 的遺傳群,但是粒線體的 DNA 可以發現,希臘的族群和摩 洛哥、西班牙和葡萄牙的族群之間,有比較接近的親緣關係,而和東歐的 國家土耳其、塞普路斯和以色列之間,則有比較遠的親緣關係 (Tsagkarakou et al., 2007)。此外,日本最近對於本土採集的煙草粉蝨,進行粒線體的 COI 序列研究,並提出新的遺傳群,由於採集點分布於日本,相對於其他遺傳 群有相當大的遺傳距離,因此將遺傳群命名為 JpL (Ueda et al., 2009)。
由分子的角度,可從事親緣關係的探討,並且做為分類上的依據,解 決 形 態 上 無 法 鑑 定 的 問 題 。 此 外 ,COI 序 列 可 做 為 分 子 標 記 (DNA
marker),設法解開物種起源、族群擴散和歷史入侵事件。利用 COI 序列,
建立最小跨度網狀圖,比對單倍型與地理位置關聯性,探討擴散的過程,
推論起源中心及分化的過程 (De Barro et al., 2005)。以 Q 型生物小種為 例,從遺傳多樣性的角度,發現法國、亞洲和美國的遺傳相似度很高,可 能是短時間之內,經由人類的活動,而快速的擴散 (Dalmon et al., 2008)。
以粒線體 DNA 探討 Q 型生物小種的入侵事件,可以發現中國、日本和 韓國之間,雖然存在地理隔離,但是卻同時具有相同的兩種單倍型,推測 亞洲區的 Q 型生物小種是屬於近期多次入侵事件 (Hsieh et al., 2007)。此 外,由 GenBank 下載 Q 型生物小種的 COI 序列,也可提供不少資訊,
利用鄰接法繪製親緣關係樹,研究中顯示可分為兩群; 第一群分布在環地 中海國家,包含摩洛哥、西班牙、阿爾及利亞、葡萄牙、希臘和法國,以 及非地中海國家,包含荷蘭、中國、日本、美國和韓國。第二群分布在東 地中海地區,包含以色列、塞普路斯和美國。中國的 Q 型生物小種和第一 群有較相近的親緣關係,而美國的 Q 型生物小種序列,有分別和第一群及 第二群相近的親緣關係,顯示中國的 Q 型生物小種來自於環地中海國家,
而美國的 Q 型生物小種,則有來自於地中海國家和東地中海地區的兩個不 同入侵源 (Chu et al., 2008)。日本對於 Q 型生物小種的研究,從 2004 年 的調查結果只在 3 個縣有採集記錄,但是至 2008 年為止,已經在 36 個 縣有採集記錄,顯示 Q 型生物小種在日本已經迅速的在野外建立族群,並 擴散至本州和九州各地,根據粒線體 COI 序列的研究,初步推斷是經由國 際間的貿易行為入侵日本,但是入侵的來源及路徑尚不清楚 (Ueda et al., 2009)。
六、微衛星 DNA 在煙草粉蝨之研究
由於分子遺傳學快速發展,以及分子生物技術的廣泛運用,已經可以 應用在昆蟲生態保育和蟲害的管理,微衛星 DNA 廣泛分布在真核生物基
因組中,它是短小的 DNA 重複片段,通常重複 1~6bp,因為其具有高度 多型性,且符合孟德爾遺傳定律的特性,所以成為探討遺傳結構、親緣地 理及族群擴散的重要工具。而且由於突變速率快、具有多型性,所以常用 於入侵物種的研究 (Le Roux et al., 2009)。
微衛星 DNA 在煙草粉蝨的研究,開始於兩組研究人員,分別從各種 不同生物小種的煙草粉蝨基因組中,籂選出 15 個和 10 個引子用來增幅 微衛星基因座 (De Barro et al., 2003; Tsagkakou and Roditakis, 2003),以及 從 B 和 Q 型生物小種籂選出來的 8 個引子對 (Delatte et al., 2006)。接 著 (De Barro et al., 2005) 利用 15 對引子增幅微衛星 DNA 研究亞洲太 平洋地區煙草粉蝨的遺傳結構,發現這個地區的煙草粉蝨可區分成 6 群,
相鄰的群之間,因為長時間的地理隔離和生殖隔離,已有遺傳差異存在。
此外,對於地中海地區的 Q 型生物小種進行研究,由 De Barro 所設計的 15 組引子對中,挑選其中 6 組適用於 Q 型生物小種的引子對,以微衛星 DNA 進行親緣關係的分析,發現地中海地區的伊比利亞半島、義大利和以 色列的族群之間,都存在非常小的遺傳差異,所以難以推論入侵來源以及 擴散方向。北非加那利群島 (Canarian Island) 和相鄰的摩洛哥 (Morocco) 由 於 海 洋 的 地 理 隔 離 , 限 制 基 因 交 流 (gene flow) 而 有 較 大 的 遺 傳 距 離 (Simon et al., 2007)。顯示地中海地區 Q 型生物小種,並不符合隔離分化 的模式,可能透過人為的貿易或活動,而迅速的向其他國家擴散。
儘管煙草粉蝨經由人為的方式,可以迅速的擴散到全世界,但自然的 擴散能力卻很小。利用 Tsagkakou and Roditakis 所設計的 10 組引子對,
挑選其中 4 組,再加上新設計的 2 組引子對,檢測希臘南方島嶼 (Crete) 上的兩個族群,基因流動 (gene flow) 的方式,即使相距只有數公里遠,基 因流動卻很低,有遺傳上的差異,結果顯示兩個族群間,並沒有雜交的事 件,且在 2002 年對於島嶼上的 14 個族群做檢測,發現全部都為 Q 型
生物小種,即使只有數公里的距離,都有很大的遺傳距離 (FST > 0.2),顯 示 Q 型 生 物 小 種 在 希 臘 南 方 的 Crete 島 , 已 經 有 長 時 間 的 演 化 歷 史 (Tsagkarakou et al., 2007)。
最近,針對法國溫室裡的煙草粉蝨族群做研究,大部分溫室皆可調查 到 Q 型生物小種,對於 7 個 Q 型生物小種的族群,由 De Barro 等人、
Tsagkakou and Roditakis 和 Delatte 等人所發表的引子對,挑選 8 對做遺 傳分化的探討,發現雖然溫室之間的距離相距 600 公里,但是卻只有很小 的遺傳差異存在,而且各個溫室之間的遺傳差異,並沒有隨著地理距離相 關,反而都只有很小的遺傳差異,由遺傳結構來看,亦無法將各個族群做 分群,而是呈現混合的狀況,顯然各個溫室之間有頻繁的基因交流,由於 冬季法國溫室外的環境並不適合煙草粉蝨的生長,所以各個溫室之間,基 因交流顯然不是由自然的擴散所造成,成因可能是因為花卉的買賣,導致 煙草粉蝨在各個溫室之間擴散 (Dalmon et al., 2008)。
七、臺灣地區 Q 型生物小種之研究
自 2001 年,針對臺灣地區煙草粉蝨展開調查工作,到 2005 年為止,
僅偵測到 B、An 和 Nauru 三型生物小種,根據族群遺傳的研究,利用親 緣關係樹的建立,發現 B 型生物小種是近期隨著人為活動入侵臺灣,且有 多次入侵的現象,An 和 Nauru 型生物小種,由遺傳歧異度推測,為早期 的自然擴散進入臺灣地區 (Hsieh et al., 2006)。
2006 年 2 月,臺南的善化亞洲蔬菜中心被偵測到 Q 型生物小種,
根據 COI 序列的定序和比對,臺南善化的族群中只有一個單倍型,由於在 2005 年 以 前 並 無 Q 型 生 物 小 種 的 採 集 記 錄 , 根 據 單 倍 型 的 粒 線 體 DNA,推測是近期單一的入侵事件 (Hsieh et al., 2007)。在 2006 年 2 月 到 2008 年 7 月,全面採集臺灣地區煙草粉蝨 481 筆資料,在宜蘭、桃 園、苗栗、南投、臺南的 9 個聖誕紅溫室內發現 6 筆未知的生物小種,
經專一性引子鑑定後確認為 Q 型生物小種 (Hsieh et al., 2008)。臺灣地區 之 Q 型生物小種目前僅在溫室發現,尚未在野外偵測到,屬於入侵初期尚 未建立可自我維持的族群,因此目前定位為外來生物,預期有高風險性成 為入侵有害生物 (Hsieh et al., 2008)。
參、材料與方法 一、標本的採集及保存
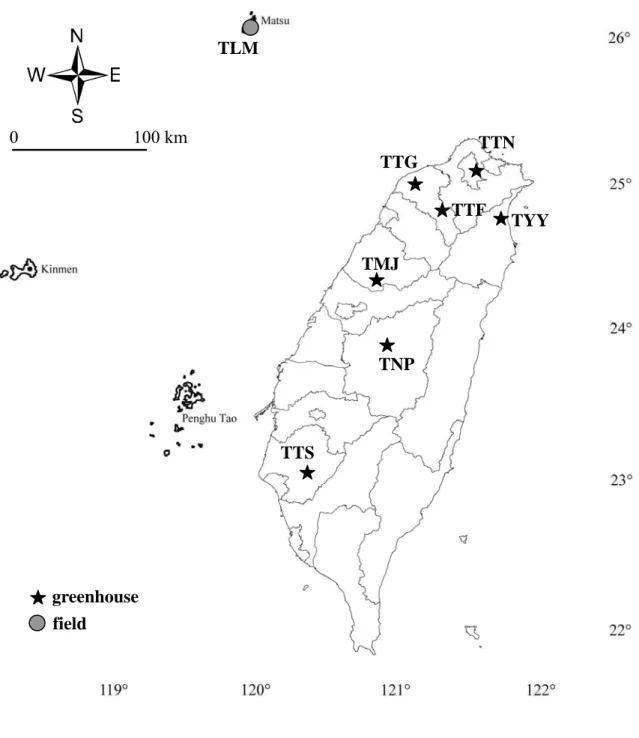
臺灣地區聖誕紅的產地主要分布在桃園縣、苗栗縣卓蘭、南投縣埔里 等地,採集點選定臺灣 7 個溫室族群,包括桃園縣復興 (TTF)、桃園縣觀 音 (TTG)、苗栗縣卓蘭 (TMJ)、南投縣埔里 (TNP)、臺南縣善化 (TTS)、
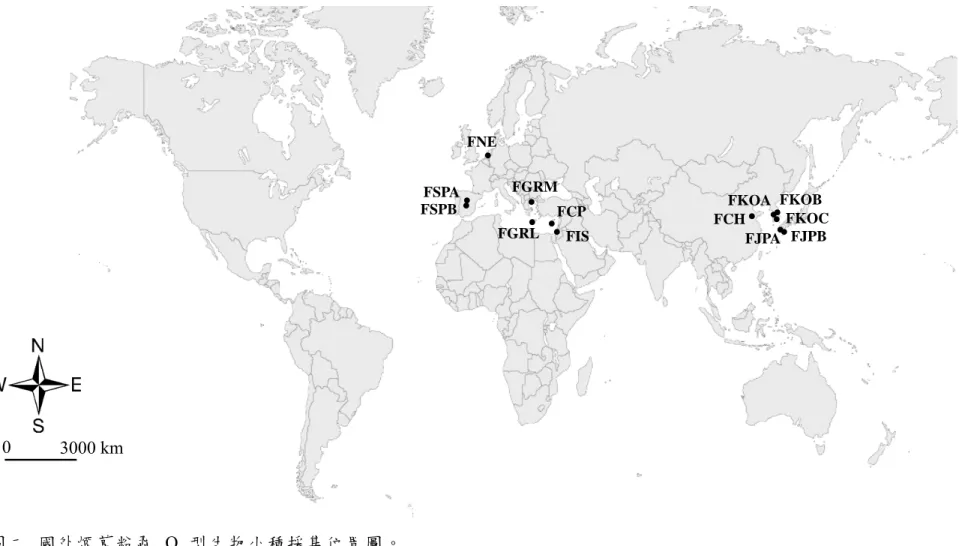
宜 蘭 縣 三 星 、 宜 蘭 市 (TYY) 等 地 點 (圖 一 )。 由 於 臺 南 縣 善 化 的 溫 室 在 2007 年 12 月時採集時已無發現 Q 型生物小種,其餘桃園縣復興、桃園 縣觀音、苗栗縣卓蘭、南投縣埔里、宜蘭縣三星、宜蘭市各族群,在 2007 年 12 月、2008 年 4 月、2008 年 8 月和 2008 年 12 月分別做 4 次 採 集 調 查 , 將 採 集 到 的 煙 草 粉 蝨 浸 泡 於 95% 酒精,置於 -4℃ 冷凍櫃保 存 。 這 段 期 間 同 時 做 野 外 及 臺 北 內 湖 花 市 的 採 集 調 查 , 偵 測 Q 型生物小 種,是否有擴散至野外。並向國外專家交換或索取煙草粉蝨 Q 型生物小種 95% 酒精浸液樣本 (表一),國外樣本採集分布於環地中海地區,包含荷蘭 採 集 自 觀 賞 花 卉 的 溫 室 族 群 (FNE)、 西 班 牙 兩 個 農 作 物 上 的 族 群 (FSPA 和 FSPB)、希臘野外的族群 (FGRM)、以及自希臘南方島嶼 Crete 島上採 回自實驗室飼養族群 (FGRL),另外尚有塞普路斯 (FCP) 和以色列 (FIS) 的兩個族群。東北亞的樣本,有採集自中國山東省棉花作物的族群 (FCH),
以及日本九州的兩個野外族群,一個是國外學者所提供 (FJPA),另一個族 群是實驗室在九州熊本市的採集調查 (FJPB),韓國的樣本採集自首爾附近 的二處聖誕紅溫室 (FKOA 和 FKOB),以及在大田市野外雜草採集到的族 群 (FKOC) (圖二)。
二、萃取基因組 DNA
將粉蝨自酒精中取出,以二次蒸餾水清洗,將酒精去除,並取 1 雌性 煙草粉蝨個體置於 1.5 ml 離心管中,加入 10µl lysis buffer (50mM KCl, 10mM Tris pH 8.4, 0.45% Tween 20, 0.2% gelatin, 0.45% NP40, 60µg/ml
proteinase K),利用磨蟲棒將蟲體磨碎,再加入 15µl lysis buffer 混和均勻。
以 65℃ 水浴 40 分鐘,離心將溶液集中,再至入 100℃ 水浴 10 分鐘,
冷 卻 至 室 溫 後 , 加 入 25µl 二 次 蒸 餾 水 混 和 均 勻 , 置 於 -20℃ 保 存 (De Barro and Driver, 1997)。
三、測定生物小種
根據 Ko et al. (2007) 所使用之專一性引子組 (表二),測定煙草粉蝨 之生物小種。
PCR 的 反 應 條 件 中 , 包 含 DNA 萃 取 液 1μl , 2U Taq DNA polymerase、10X Taq buffer 2.5μl、2.5mM dNTP 2μl、10mM forward primer 1μl、10mM reverse primer 1μl,最後再加入 ddH2O,使總體積為 25μl。
PCR 反應條件為 (1) 預熱 (prewarm) 94℃/5 分鐘,(2) 進行 35 個循環之 變 性 (denaturing) 94℃/1 分 鐘 、 黏 合 (annealing) 56-64℃/1 分 鐘 、 延 伸 (extension) 72℃/1 分鐘,(3) 再經延伸 (final extension) 72℃/5 分鐘完成反 應。將 PCR 增幅的產物,以 2% 的瓊脂電泳膠片進行電泳檢視,Q 型生 物小種 DNA 序列 可增幅出 892bp、700bp、400bp 片段產物,而 B、An、
Naura 型生物小種可以分別增幅出 649bp、564bp 和 666bp。用以判定為 何種型生物小種之煙草粉蝨。
四、粒線體 (Mitochondrial) COI 基因序列之資料分析
對於 Q 型生物小種進行 COI 序列分析,採集與交換所獲得的 13 個 國外族群,以及在臺灣地區採集到的 23 個族群 (包含馬祖),隨機挑選三 隻進行粒線體 COI 基因序列之定序,定序的方式根據 Frohlich et al. (1999) 所 使 用 之 引 子 組 ,C1-J-2195 (5`-TTGATTTTTTGGTCATCCAGAAGT-3`) / L2-N-3014 (5`-TCCAATGCACTAATCTGCCATATTA-3`)。增幅出大約 820bp COI 序列。PCR 反應條件為 94℃/5 分鐘,然後進行 35 個循環之 94℃/1
表一 供試煙草粉蝨 Q 型生物小種樣本
Table 1. Sample of Bemisia tabaci biotype Q used in this study
Sample sources Name Location Host plant Date
Global
FNE Netherlands Hibiscus, greenhouse 2002
FSPA Spain Cucumber 2003
FSPB Spain Tomato 2003
FCP Cyprus - 2004
FJPA Kyushu Island, Japan Sweet pepper 2005 FJPB Kumamoto, Kyushu Island, Japan pilose beggarticks 2008
FIS Israel - 2006
FCH Shandong, China Cotton 2007 FGRM Agrinio, Mainland Greece Cucumber 2004 FGRL Crete, Greece Cucumber 2006 FKOA Goyang-Si, Kyeonggi-do, Korea Poinsettia 2008 FKOB Goyang-Si, Kyeonggi-do, Korea Poinsettia 2008 FKOC Yuseong, Daejeon, Korea Black nightshade 2008 Taiwan
TTSA Shanhua, Tainan Poinsettia Feb-2006
TTSB Shanhua, Tainan Poinsettia Jul-2007
TMJA Jhuolan, Miaoli Poinsettia Apr-2006
TMJB Jhuolan, Miaoli Poinsettia Dec-2007
TMJC Jhuolan, Miaoli Poinsettia Apr-2008
TMJD Jhuolan, Miaoli Poinsettia Aug-2008
TMJE Jhuolan, Miaoli Poinsettia Dec-2008
TTGA Guanyin, Taoyuan Poinsettia Dec-2007
TTGB Guanyin, Taoyuan Poinsettia Aug-2008
TTGC Guanyin, Taoyuan Poinsettia Dec-2008
TNPA Puli, Nantou Poinsettia Dec-2007
TNPB Puli, Nantou Poinsettia Apr-2008
TNPC Puli, Nantou Poinsettia Aug-2008
TNPD Puli, Nantou Poinsettia Dec-2008
TYYA Yilan City Poinsettia Jan-2008
TYYB Yilan City Black nightshade Apr-2008
TYYC Yilan City Poinsettia Aug-2008
TYYD Yilan City Poinsettia Dec-2008
TTFA Fusing, Taoyuan Poinsettia Arp-2008
TTFB Fusing, Taoyuan Poinsettia Aug-2008
TTFC Fusing, Taoyuan Poinsettia Dec-2008
TTN Neihu, Taipei Poinsettia Oct-2008
TLM Matsu, Lienchiang Watermelon Nov-2008
圖一 臺灣地區煙草粉蝨 Q 型生物小種採集位置圖。
Fig 1. Sampling locations of Bemisia tabaci biotype Q in Taiwan.
TTS
TNP TMJ
TYY TTG
TTN
TTF TLM
greenhouse field
0 100 km
圖二 國外煙草粉蝨 Q 型生物小種採集位置圖。
Fig 2. Sampling locations of Bemisia tabaci biotype Q in the world.
.. .. . .. ...
.. .
FSPA FSPB
FNE
FGRL FGRM
FIS
FCP FKOA
FCH
FKOB FKOC FJPA FJPB
0 3000 km
表二 煙草粉蝨生物小種專一性引子組序列及其黏合溫度
Table 2. Primers and annealing conditions for identifying different biotypes of Bemisia tabaci
Biotype Primer Annealing temp (℃ )
Q BaQF
BaQR 5`-GAAGCAACGCACTACTTACAA-3`
5`-TTCTCGGCGTTTTTACCAA-3` 56℃
B BaBF
L2-N-3014 5`-CCACTATAATTATTGCTGTTCCCACA-3`
5`-TCCAATGCACTAATCTGCCATATTA-3` 60℃
An BaAnF L2-N-3014
5`-GGTTATTGCTGTTCCAACTGGG-3`
5`-TCCAATGCACTAATCTGCCATATTA-3` 64℃
Naura BaNaF
L2-N-3014 5`-GGCTTTGGTTTACTGGATTCTTTT-3`
5`-TCCAATGCACTAATCTGCCATATTA-3` 62℃
分鐘、52℃/1 分鐘、72℃/1 分鐘,最後再經 72℃/5 分鐘完成反應。將 PCR 增幅的產物,以 2% 的瓊脂電泳膠片進行電泳檢視。PCR 增幅出的產物送 至生技公司,利用自動定序儀 (ABI PRISMTM) 進行雙股定序。
1.序列之比對與排序
將 定 序 所 得 的 正 反 股 序 列 , 利 用 GeneDoc 軟體進行序列校正,整合 成完整的正股序列。並使用 NCBI 網頁之 GenBank 中的 BLAST 功能進 行序列比對,再次確認生物小種。另外由 GenBank 下載,相關的 Q、B、
Ms 型 生 物 小 種 的 COI 序 列 , 與 臺 灣 所 採 集 的 Q 型 生 物 小 種 COI 序 列,利用 Tsagkarakou et al. (2007) 建議使用的 Clustalx 1.81 軟體進行比 較分析。紫背草唇粉蝨 (Lipaleyrodes emiliae) COI 序列做為外群 (outgroup) (Hsieh et al., 2007)。
2.親緣關係樹之建立與信賴程度的統計檢驗
族群遺傳結構會受到突變 (mutation)、遺傳漂變 (genetic drift) 與選汰 (selection) 的作用而改變,並反映在分子的層次。族群內的遺傳變異度有 許 多 種 估 算 方 式 , 如 基 因 多 樣 性 (gene diversity) 、 異 型 合 子 的 比 例 (heterozygosity)。採用粒線體 DNA COI 序列的基因多樣性,計算族群間的 遺傳變異度,由同一原始族群經相同演化歷史形成遺傳結構,中性的遺傳 變異受中性突變及隨機遺傳漂變之影響,因不受環境選汰作用而可用來推 測分歧時間。本研究利用距離方法 (distance matrix method) 建立親緣關係 樹,首先獲得所有分類群間的遺傳距離,親緣關係樹的建構則基於這些距 離值之間的關係。鄰接法 (Neighbor-joining method, NJ) 基於最小遺傳距離 原理,假設物種進化速率不一致 (Saito and Nei, 1987)。將所有樣本序列兩 兩比對,計算序列核苷酸位點替代數之實際值為遺傳距離值後,將序列間 之遺傳距離排列成矩陣,依照此矩陣建立一棵星狀樹。根據星狀樹,將所 有分類群做潛在的鄰聚分析,選出分枝總合最短的群聚。計算群聚所有分
類群之間的距離,將同一分類群鄰接在一起,將所有分類群聚合成一棵樹。
利用自展檢驗 (bootstrap test) 進行統計檢驗,由自展重複抽樣技術,計算 每個分枝 clade 之 bootstrap 值,評估信賴程度 (Felsenstein, 1985)。NJ tree 以 MEGA4 (Tamura et al., 2007) 軟體建構。
3.COI 序列在親緣地理學 (Phylogeography) 研究
親緣地理學為歷史性生物地理學 (Avise et al., 1987),主要是在探討基 因 譜 系 與 地 理 分 布 以 及 與 時 間 的 關 聯 性 , 尤 其 是 親 緣 關 係 接 近 的 物 種 (Avise, 2000)。製作最小跨度網狀圖 (Minimum Spanning Network),將單倍 型間核苷酸突變數,由差異小至大,依序連接串聯成一網狀圖,並結合地 理分布資訊,可知單倍型關聯性與地理分布之關係。利用 Arlequin ver 2.0 軟體分析所得資訊,並使用繪圖軟體製作最小跨度網狀圖。
五、微衛星基因座的增幅與資料分析
微衛星 DNA 運用在族群遺傳的研究上,具有以下幾個特點:(一) 在 真 核 生 物 基 因 組 中 , 存 在 大 量 的 微 衛 星 DNA 。 ( 二 ) 高 度 多 型 性 (Polymorphism) 且突變速率快,微衛星 DNA 基因座,同時包含數十個的 等位基因,這樣的多型性有利於進行演化與族群遺傳研究,特別是近期入 侵的生物 (Le Roux and Wieczorek, 2009)。(三) 突變速率快。微衛星 DNA 序列包含重複序列 (repeat sequences) 與兩端非重複序列 (flanking regions) 兩部分。重複序列多由 1~ 6bp 大小的重複單元,重複 5 次以上所組成,
且由於減數分裂期間,容易在 DNA 複製時出現滑動 (slippage) 現象,使 微衛星 DNA 序列的突變率比一般序列高,且不同基因型之間的差異,主 要來自重複序列的重複次數不同 (Goldatein and Schlotterer, 2000)。在突變 速率的差異上,酵母菌每一個基因座的每一次複製,其微衛星 DNA 的突 變率介於 10-4~10-5 之間 (Strand et al., 1993),人類的微衛星 DNA 的突 變率約為 10-2 (Weber and Wong, 1993),而昆蟲中的果蠅 (Drosophila) 的
突變率約為 6.3×10-6 (Schug et al., 1997)。在時間的尺度上,粒線體可以研 究 105~102 年之間的演化事件,相較於粒線體 DNA 無法有效區別的族 群,微衛星 DNA 可以研究的範圍約在 103~101 年之間的演化事件,在 時間尺度上具有較大的優勢 (Grant and Waples, 2000)。(四) 等位基因表現 出共顯性遺傳:微衛星 DNA 遵守孟德爾遺傳定律,來自於父方及母方的 微衛星 DNA 都可同時被偵測到 (Hearne et al., 1992),所以可以做為親緣 分析的分子標誌。
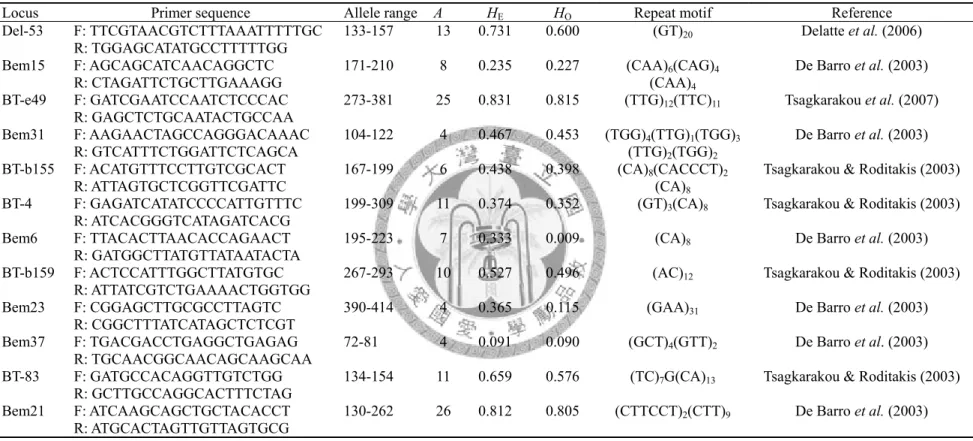
根據 De Barro et al. (2003)、Tsagkarakou and Roditakis (2003)、Delatte et al. (2006) 以及 Tsagkarakou et al. (2007),所使用之引子組,挑選出 12 組並分別標定 4 種不同的螢光 (Del-53、Bem15、BT-e49) FAM、(Bem6、
BT-b159、Bem23) NED、(Bem37、BT-83、Bem21) PET 和 (Bem31、BT-b155、
BT-4) VIC (表三),進行 Multiplex PCR 增幅出微衛星 DNA 基因座。PCR 的 反 應 條 件 中 , 包 含 DNA 萃 取 液 1μl , 0.5U hot start Taq DNA polymerase,10X Taq buffer 1μl,2.5mM dNTP 1μl 及 10mM forward primer 2.4μl、10mM reverse primer 2.4μl,最後再加入 ddH2O,使總體積為 10μl。
Multiplex PCR 反 應 條 件 為 95℃/10 分 鐘 , 然 後 進 行 35 個 循 環 之 95℃/40 秒、54℃/40 秒、72℃/40 秒,最後再經 72℃/30 分鐘完成反應。
Multiplex PCR 增幅的產物,以 2.5% 的瓊脂電泳膠片進行電泳檢視是否有 PCR 的增幅產物。Multiplex PCR 增幅的產物,送至生技公司,以 ABI3100 DNA 分析儀進行基因型判定 (genotyping),檢視 12 個基因座長度大小。
本研究依序列長度挑選 12 組引子對,利用 4 種不同的螢光進行標記,使 基因座的增幅可使用 Multiplex PCR,基因型的判讀也可一次完成,降低實 驗的成本。判讀的方式利用 GENESCAN 3.1.2 軟體進行。
1.遺傳變異 (Genetic variation) 之分析
計算聖誕紅溫室族群在不同時期,以及各國樣本之間基因座上的對偶
基因頻率分布,並估算變異度;即對偶基因數 (number of alleles)、異型合 子比例 (heterozygosity)、以及基因歧異度 (gene diversity)。異型結合子的 比例計算分為異型合子觀測值 (HO, Observed heterozygosity) 以及異型合 子期望值 (HE, Expected heterozygosity) 的估算。觀測值可直接由樣本異型 合 子 比 例 算 出 。 異 型 合 子 期 望 值 則 採 用 Nei (1978) 所 提 出 的 無 偏 估 值 (Unbiased heterozygosity) 算出單一基因座異型合子期望值,再由各基因座 之異型合子期望值相加平均後,求得平均異型合子期望值。公式之計算以 GENEPOP Version 3.4 程式進行運算 (Yeh and Boyle, 1997)。單一基因座異 型合子期望值 (h) 之計算如下 :
n:樣本數;
Pi:族群內第 i 種等位基因型之頻率。
平均異型合子期望值 (HE) 之計算如下:
r:基因座總數;
hi:平均異型合子期望值。
此外,檢測各個基因座是否獨立於另一個基因座,避免不同基因座來 自相同染色體所產生的偏估,利用 GENEPOP Version 3.4 程式進行基因座 間連鎖不平衡 (linkage disequilibrium) 測試。
2.族群哈溫平衡 (Hardy-Weinberg equilibrium) 檢測
一個隨機交配的族群,除非有外力的作用,否則其等位基因出現的頻 h = 2n (1 – ∑ Pi2)
2n – 1
n i =1
∑ hi
r
r i =1
HE =
表三 供試之煙草粉蝨微衛星基因座之引子組
Table 3. Characteristics of microsatellite loci in biotype Q of Bemisia tabaci
Locus Primer sequence Allele range A H
EH
ORepeat motif Reference Del-53 F: TTCGTAACGTCTTTAAATTTTTGC
R: TGGAGCATATGCCTTTTTGG 133-157 13 0.731 0.600 (GT)
20Delatte et al. (2006) Bem15 F: AGCAGCATCAACAGGCTC
R: CTAGATTCTGCTTGAAAGG
171-210 8 0.235 0.227 (CAA)
6(CAG)
4(CAA)
4De Barro et al. (2003) BT-e49 F: GATCGAATCCAATCTCCCAC
R: GAGCTCTGCAATACTGCCAA 273-381 25 0.831 0.815 (TTG)
12(TTC)
11Tsagkarakou et al. (2007) Bem31 F: AAGAACTAGCCAGGGACAAAC
R: GTCATTTCTGGATTCTCAGCA
104-122 4 0.467 0.453 (TGG)
4(TTG)
1(TGG)
3(TTG)
2(TGG)
2De Barro et al. (2003) BT-b155 F: ACATGTTTCCTTGTCGCACT
R: ATTAGTGCTCGGTTCGATTC 167-199 6 0.438 0.398 (CA)
8(CACCCT)
2(CA)
8Tsagkarakou & Roditakis (2003) BT-4 F: GAGATCATATCCCCATTGTTTC
R: ATCACGGGTCATAGATCACG
199-309 11 0.374 0.352 (GT)
3(CA)
8Tsagkarakou & Roditakis (2003) Bem6 F: TTACACTTAACACCAGAACT
R: GATGGCTTATGTTATAATACTA 195-223 7 0.333 0.009 (CA)
8De Barro et al. (2003) BT-b159 F: ACTCCATTTGGCTTATGTGC
R: ATTATCGTCTGAAAACTGGTGG
267-293 10 0.527 0.496 (AC)
12Tsagkarakou & Roditakis (2003) Bem23 F: CGGAGCTTGCGCCTTAGTC
R: CGGCTTTATCATAGCTCTCGT 390-414 4 0.365 0.115 (GAA)
31De Barro et al. (2003) Bem37 F: TGACGACCTGAGGCTGAGAG
R: TGCAACGGCAACAGCAAGCAA
72-81 4 0.091 0.090 (GCT)
4(GTT)
2De Barro et al. (2003) BT-83 F: GATGCCACAGGTTGTCTGG
R: GCTTGCCAGGCACTTTCTAG 134-154 11 0.659 0.576 (TC)
7G(CA)
13Tsagkarakou & Roditakis (2003) Bem21 F: ATCAAGCAGCTGCTACACCT
R: ATGCACTAGTTGTTAGTGCG
130-262 26 0.812 0.805 (CTTCCT)
2(CTT)
9De Barro et al. (2003)
A: total number of alleles found among all populations; H
E: expected heterozygosity; H
O: observed heterozygosi
率,在不同世代間維持不變,遵守此定律的族群須符合下列前提:(1) 逢機 交 配 (random mating) (2) 族 群 數 量 必 須 夠 大 (large population size) (3) 沒有個體的移入或移出 (no migration) (4) 沒有突變發生 (no mutation) (5) 沒有天擇的作用 (no selection)。但事實上,大部分的生物族群都會受到上 述不同演化機制的影響而發生改變。
為了檢測煙草粉蝨族群是否處於哈溫平衡的狀態,計算隨機交配下的 基 因 型 頻 率 期 望 值 (Expected genotypic frequencies) , 再 以 GENEPOP Version 3.4 程式進行運算,檢定族群內各基因座是否符合哈溫平衡的狀態 (Yeh and Boyle, 1997) 。 若 基 因 座 有 偏 離 哈 溫 平 衡 的 狀 態 , 再 利 用 micro-checker v.2.2.0 軟體 (Van Oosterhout et al., 2004) 檢測各個基因座是 否有無效對偶基因 (null allele),或基因型判讀錯誤的情況。
3.微衛星 DNA 之族群分化指數 (FST)
自然族群中,環境的隔離,使得原始族群分化成好幾個次族群。若各 次族群間沒有基因互相交流,就會造成之間遺傳組成的不同,而分化指數 的值也會越高。Q 型生物小種,目前在臺灣只生長在溫室的環境下,計算 族群分化指數,可以知道各個溫室之間族群,是否因為環境的隔離而有分 化的情況。根據 (Weir and Cockerham, 1984) 所修正的 Wright’s F statistics 來表示族群間分化的指數。利用 GENEPOP Version 3.4 程式進行以下公式 之檢測 (Yeh and Boyle, 1997)。
其中,δP2:族群間對偶基因頻率均方期望值;
δG2
:對偶基因間對偶基頻率均方期望值;
δI 2:族群內對偶基因頻率均方期望值。
族群分化指數 (FST)= δP2
δP2 + δG2 + δI2
4.微衛星 DNA 之族群近親交配指數 (FIS)
近親交配指數分析所有族群內異型合子偏離逢機交配預期的情況。使 用 Weir and Cockerham (1984) 的 方 法 , 計 算 近 親 交 配 指 數 (inbreeding coefficient) , FIS 指 數 顯 著 偏 離 , 表 示 族 群 內 的 個 體 交 配 不 是 隨 機 發 生 (non-random mating),族群的近親交配,可能使族群的遺傳多樣性降低,近 親交配指數 (FIS) 公式如下,並利用程式 FSTAT Version 2.9.3.2 進行分析 (Goudet, 1995)。在不符合哈溫的情況下,可進一步確認是否有近親交配的 可能。
近親交配指數 (FIS ) =
5.瓶頸效應 (Bottleneck effect) 檢測
當一個族群在經過一個巨大環境改變,造成大多數個體的死亡或限制 了正常的生殖行為之後的遺傳結果,稱之為瓶頸效應,可能導致物種基因 多樣性喪失。檢驗一個族群是否曾經經歷瓶頸,可利用下列二種方式。第 一、眾位數偏移指標 (Mode-shift indicator): 在遺傳突變與漂變的長期作用 下,穩定族群中存在較多的稀有對偶基因,因此微衛星基因座對偶基因的 頻度分布會呈現 L 型分布,而當有效族群量下降時,稀有對偶基因會大量 消失,造成基因座上對偶基因頻度分布偏離 L 分布 (Cornuet and Luikart, 1996)。第二、異型合子觀測值 (HO) 與期望值 (HE) 檢驗: 當近期內有效 族群的數量減低時,對偶基因的數目及基因多樣性皆會減少,但對偶基因 數 目 減 少 的 幅 度 又 比 異 型 合 子 比 例 減 少 的 快 (Cornuet and Luikart, 1996;
Luikart and Cornuet, 1998),所以可以利用現在族群的對偶基因數目和頻 度,利用族群異型合子比例 (HO) 是否顯著小於符合哈溫平衡的族群所預 期具有的異型合子比例 (HE),以推測族群是否遭遇瓶頸效應。
對 偶 基 因 突 變 有 「Infinite allele model; IAM」、「Step-wise mutation δI 2
δG2 +δI 2
model; SMM」 和 「Two phased model; TPM」 三種模式,其中 Infinite allele model 是針對同功異構酶而提出的突變模式,認為對偶基因的突變率低,
突變產生的對偶基因未曾出現在族群中,而且每一種新的對偶基因,都和 突變前的對偶基因形式無關,但是微衛星 DNA 的突變速率快,每一種新 的對偶基因是由原本的對偶基因增減一個至數個重複單元,突變後的對偶 基 因 長 度 可 能 和 族 群 內 原 有 的 對 偶 基 因 相 同 , 因 此 不 符 合 Infinite allele model。隨後有學者提出以 Step-wise mutation model 較符合微衛星 DNA 的突變模式,每一次的突變增加或減少一個重複單元,並提出估算族群間 分 化 程 度 或 遺 傳 距 離 的 方 法 (Slatkin, 1995),但這項計算方式忽略微衛星 DNA 突變方式可能包含兩個以上重複單元的突變 (Multistep mutations)。
因此,最近的學者則建議採用 Two phased model,以差距一個重複單元突 變 (Single-step mutations) 為基本模式,並同時考慮差距兩個以上重複單元 的 突 變 , 用 來 估 算 族 群 是 否 經 歷 近 期 的 瓶 頸 效 應 (Garza and Williamson, 2001; Simon et al., 2007)。
入侵生物在入侵的過程中,經歷瓶頸作用,可能導致遺傳多樣性降低。
本研究計算各個族群內異型合子的比例 (HO),是否顯著小於穩定族群之異 型合子比例的期望值 (HE),同時檢測對偶基因頻度是否偏離 L 型分布。
微衛星的演化模式設定,將差距一個重複單元的突變的比率設為 90%,差 距兩個以上重複單元突變的比率設為 10%,重複運算 1000 次建立異型合 子的期望值,並以 Wilcoxon test 來檢定異型合子是否過量。當有效族群在 近期內大規模縮小,具有多型性的微衛星基因座,對偶基因的數量會顯著 下降,使異型結合子的觀測值小於期望值 (HO<HE),若台灣地區內各煙草 粉蝨族群,偶基因的多樣性無明顯下降,且異型結合子的觀測值和期望值 並無差異,則可推測煙草粉蝨族群皆無族群量顯著下降的狀況。瓶頸效應 檢測利用 BOTTLENECK (Cornuet and Luikart, 1996; Piry et al., 1999) 軟體
進行。
6.計算遺傳距離 (Genetic distance) 與建構親緣關係樹 (Phylogenetic tree) 遺 傳 距 離 (Genetic distance) 可 表 示 不 同 族 群 間 遺 傳 結 構 的 差 異 程 度 與親疏關係。計算兩兩不同族群間樣本的遺傳距離 (DAS),以了解各個聖誕 紅溫室之間,以及國外煙草粉蝨的遺傳相似度。遺傳距離 (DAS) 計算方式 (Chakraborty and Jin, 1993) 如下:
其中,PSAkm:族群間個體分享對偶基因的平均比例;
PSAk:k 族群內個體間分享對偶基因的平均比例;
PSAm:m 族群內個體間分享對偶基因的平均比例。
DAS 計 算 之 後 , 以 鄰 接 法 (Neighbor-joined method) (Saitou and Nei, 1987) 的原理利用 Populations 1.2.30 軟體建構親緣關係樹,並進行 1000 次 bootstrapping (Felsenstein, 1985),建構最理想親緣關係樹。
7.貝氏族群遺傳結構分析
檢視族群內之分群狀況,基本假設是同一分群內各基因型的頻度符合 哈溫平衡,且各個基因座之間屬於連鎖平衡 (linkage equilibruim) 利用貝氏 分群分析 (Bayesian clustering analysis),研究族群之間遺傳組成的相似度,
利用 12 個微衛星基因座的變化和差異,將個體分群,遺傳結構相似度高 的 , 會 被 歸 為 同 一 個 分 群 (cluster), 並 利 用 Markov Chain Monte Carlo (MCMC) 演算法重複分群,統計個體分配到各群的機率,推估最可能的結 果。
進行分群檢驗時,假設所有樣本皆由同一群祖先繁衍而來,在 Structure 2.1 程 式 中 , 之 先 祖 模 式 (Ancestry model) 及 對 偶 頻 度 模 式 (Allele
2PSAkm
PSAk +PSAm
DAS = 1-
m|L (K+1) - 2L (K) + L (K-1)|
s[L ( K )]
frequency model) , 分 別 選 取 混 合 模 式 (Admixture model) 及 相 關 模 式 (Correlated model),進行 7×105 次 MCMC 演算,先 Burnin 3×105 次,之 後開始收集 4×105 次 MCMC 演算的結果,分群值設為 1 到 10,每個分 群值重覆演算 4 次 (Pritchard et al., 2000)。根據演算的結果,選出 ∆K 值 最大,且 α1<1 為可信度最高的分群方式 (Evanno et al., 2005),計算公式 如下:
∆K=
L (K):MCMC 演算的結果取 log-likelihood 值;
m:重覆數;
s[L ( K )]:標準差估算值。
肆、結果
一、臺灣地區煙草粉蝨 Q 型生物小種採集和分布情形
全 面 採 集 臺 灣 地 區 煙 草 粉 蝨 , 時 間 自 2008 年 8 月 至 2008 年 12 月,包括臺灣本島、馬祖和金門,共採集 310 筆,在記錄中僅有 7 個地 點 檢 查 到 Q 型 生 物 小 種 , 1 筆 在 野 外 , 6 筆 在 溫 室 。 其 中 桃 園 縣 復 興 (TTF)、桃園縣觀音 (TTG)、苗栗縣卓蘭 (TMJ)、南投縣埔里 (TNP)、宜蘭 市 (TYY) 等 5 個聖誕紅溫室有持續的採集記錄 (附錄一)。在 2008 年 10 月臺北內湖花市 (TTN) 亦調查到 Q 型生物小種,業者稱植株聖誕紅 的來源為南投的埔里,但由於南投埔里的聖誕紅溫室有多處,所以不一定 是論文中樣本的採集點。對於桃園縣復興、桃園縣觀音、苗栗縣卓蘭、南 投縣埔里、宜蘭市和宜蘭三星等 6 個溫室,進行為期 1 年 (2007 年 12 月、2008 年 4 月、2008 年 8 月、2008 年 12 月) 的 4 次採集調查 (表 四)。桃園復興 (TTF) 的聖誕紅溫室,在 2007 年 12 月的採集調查尚未 發現 Q 型生物小種煙草粉蝨,但是在 2008 年 4 月的採集調查,顯示溫 室出現有入侵的 Q 型生物小種,而且採集的 16 隻樣本皆為 Q 型生物小 種,在 2008 年 8 月以及 12 月的採集調查,發現 Q 型生物小種對於 B 型生物小種的比例,有逐漸降低的趨勢 (VIII-2008 Q : B = 15 : 6; XII-2008 Q : B = 15 : 65)。宜蘭市 (TYY) 溫室 4 次的採集中, 2008 年 8 月的 23 隻樣本中,B 型生物小種佔 7 隻,其餘為 Q 型生物小種,其餘三個時間 點所採集的樣本 46 隻個體皆為 Q 型生物小種。苗栗卓蘭 (TMJ) 的溫室 在 2008 年 4 月時,有 Q、B、An 和 Na 型生物小種共存的現象,其餘 4 個採集時間點 (包含 IV-2006) 皆為 100% 的 Q 型生物小種。桃園觀音 和南投埔里的溫室,多數的採集樣本為 Q 型生物小種,在部分的採集時間 出現少數的 B 型生物小種個體,桃園觀音 (TTG) 分別在 2008 年 8 月 和 2008 年 12 月的 17 隻和 18 隻樣本中,各發現 1 隻和 3 隻的 B
表四 臺灣地區煙草粉蝨 Q 型生物小種採集記錄
Table 4. Sample records of Bemisia tabaci biotype Q in Taiwan
Location Date Name Host plant Numbers of different biotypes
Q B An Na
Shanhua, Tainan Feb-2006 TTSA Poinsettia 15 1
Jul-2007 TTSB Poinsettia 16
Jhuolan, Miaoli Apr-2006 TMJA Poinsettia 16
Dec-2007 TMJB Poinsettia 16
Apr-2008 TMJC Poinsettia 8 3 5 2
Aug-2008 TMJD Poinsettia 16
Dec-2008 TMJE Poinsettia 15
Guanyin, Taoyuan Dec-2007 TTGA Poinsettia 16 Aug-2008 TTGB Poinsettia 16 1
Dec-2008 TTGC Poinsettia 15 3 Puli, Nantou Dec-2007 TNPA Poinsettia 16 Apr-2008 TNPB Poinsettia 16 2
Aug-2008 TNPC Poinsettia 16
Dec-2008 TNPD Poinsettia 15
Yilan City Jan-2008 TYYA Poinsettia 16 Apr-2008 TYYB Black nightshade 15
Aug-2008 TYYC Poinsettia 16 7
Dec-2008 TYYD Poinsettia 15
Fusing, Taoyuan Arp-2008 TTFA Poinsettia 16
Aug-2008 TTFB Poinsettia 15 6
Dec-2008 TTFC Poinsettia 15 65
Neihu, Taipei Oct-2008 TTN Poinsettia 15
Matsu, Lienchiang Nov-2008 TLM Watermelon 3 9
型生物小種,而南投埔里 (TNP) 在 2008 年 4 月的 18 隻樣本中,發現 2 隻 B 型生物小種個體。宜蘭三星的採集時間有二次,分別為 2007 年 12 月以及 2008 年 4 月,各測試 16 隻的樣本,結果溫室全為 B 型生 物小種,無 Q 型生物小種的記錄。野外採集記錄 1 筆,2008 年 9 月連 江 縣 馬 祖 (TLM) 西瓜作物上採集到的煙草粉蝨,經專一性引子鑑定,發 現 12 隻個體中有 3 隻個體為 Q 型生物小種,其餘為 B 型生物小種。
二、粒線體 (Mitochondrial) COI 基因序列之資料分析 1.序列之比對與排序
利用 PCR 擴增粒線體 COI 序列約 820bp。臺南善化 (TTS)、桃園觀 音 (TTG)、桃園復興 (TTF)、苗栗卓蘭 (TMJ)、南投埔里 (TNP)、宜蘭市 (TYY)、臺北內湖 (TTN) 和連江縣馬祖 (TLM) 的 8 個族群 (由 Q 型生 物小種的族群中任意挑選 3 隻樣本) 的 Q 生物小種 COI 序列,經過整 理和排序 (Alignment) 後,可以得到 739bp 的序列,分為三個單倍型,之 間 有 3 個變異的點位 (附錄二)。此外將核苷酸的序列轉譯為胺基酸序列 時,顯示三個單倍型轉譯出相同的胺基酸序列。
臺 灣 地 區 族 群 與 GenBank 下 載 之 煙 草 粉 蝨 粒 線 體 COI 序 列 ( 表 五),經過排序和比對後,選取 638bp 的序列,進行分析。臺南善化 (TTS)、
桃園觀音 (TTG)、桃園復興 (TTF)、苗栗卓蘭 (TMJ)、南投埔里 (TNP) 和 宜蘭市 (TYY) 的樣本為一個單倍型 (H11),此外宜蘭市的溫室中發現另一 個單倍型 (H12),有 3 個點位的差異,連江縣的馬祖 (TLM) 也自成一個 單倍型,與臺灣地區所採集到的 H11 有一個點位的差異,但是與世界各國 煙草粉蝨做 COI 序列比對時,只使用 739bp 序列中的 638bp,差異的點 位亦被切除,所以 TLM 族群的 COI 序列為 H11 (表五)。國外的 13 個 族群,每個族群 3 隻的定序結果,韓國 (FKOA、FKOB 和 FKOC)、日本 (FJPA 和 FJPB)、中國 (FCH) 和西班牙 (FSPA 和 FSPB) 皆為 H11,以