國立臺灣大學生物資源暨農學院園藝學系 碩士論文
Department of Horticulture
College of Bioresources and Agriculture
National Taiwan University Master Thesis
矮牽牛品種及其自交或雜交後代之 耐熱性與花朵性狀表現
Heat-tolerance and Flower Characteristics of Petunia Cultivars and Their Selfed or Cross Hybrid Progenies
劉子瑄 Tzu-Hsuan Liu
指導教授:葉德銘 教授 Advisor: Prof. Der-Ming Yeh
中華民國九十七年六月
June, 2008
口試委員審定書
誌 謝
首先感謝 葉德銘老師,於修業期間悉心指導與鼓勵,在健康與生活上的關心,
並於論文撰寫期間詳加批閱指證,本論文方能順利完成。感謝朱建鏞老師鼓勵我多方 嘗試,並提供許多實驗想法,感謝楊雯如老師指導遺傳育種觀念和論文撰寫方法,並 感謝沈榮壽老師於論文初稿上的指教,提供許多寶貴意見。
感謝大觀園藝林文欽先生與童明月小姐提供矮牽牛,並指導我許多矮牽牛的栽培 小撇步,和農友種苗李進雄先生於我剛開始接觸矮牽牛耐熱育種時,提供許多寶貴意 見。感謝任伯伯、韶妤學姐、草菇學姊、倩妤學姊、昭儀學姐、昱均學姐和花卉館所 有陣容堅強的助理學長姐們。謝謝錦木學長、朝卿學長、元聰學長還有搞笑的阿雄學 長,雖然難得在花卉館遇到你們,但卻喜歡和你們聊天。還有謝謝王大學長不時的叨 叨唸、小白總是一臉正經的騙我、小幼很厲害的軟硬兼施法,祈男很可憐的老是被我 們欺負!也謝謝小范和我一起當脾氣應該要很好的褓姆、很高興老能在冬天的分場中 遇到睿娟,還要謝謝穎君、荷惠和文秋,很開心能和你們坐在一起。
還要感謝來自各地不同系所,有緣能夠認識的同學們,開起車來很man 的班代、
老被我囉嗦的俊宏要顧身體阿!步調一點都不慢的佳盛、常會用老人語氣說話的怡 嘉、一點都不白目的秀婷、幸福的人妻旻宜要好好學做便當喔!還有記憶體容量很大 的筱媁和很會做麵包、蛋糕的伊婷,感謝這兩年來有你們的陪伴。當然還有感謝歷屆 小幫手和可愛的學弟妹們,屁屁、文茹、建佑、佳妏、揚昕、俊甫、維倩、瀞頤、一 霖、韻如、翊婷,謝謝你們在我忙不過來時的一臂之力。還有我的好姐妹們丹筑、慧 宜、小猴、阿秀、娜娜、慈慧和新婚師父倉海,謝謝你們在我失落時的陪伴,供我一 個休息的地方。特別感謝胖丁的陪伴、包容與照顧,能擁有你的支持,是我的幸福。
最後要感謝親愛的爸爸和媽媽對我的照顧和包容,讓我能無後顧之憂的順利完成 學業,還有很不低調的哥哥、工作繁忙的姊姊,辛苦大家容忍任性脾氣差的我,謹以 此論文獻給你們。
目 錄
口試委員審定書... i
誌 謝 ... ii
目 錄 ... iii
表目錄... v
圖目錄... ix
中文摘要... vii
Summary... ix
前 言 (Introduction)... 1
前人研究 (Literature Review) ... 4
一、 矮牽牛之簡介... 4
(一) 親源及育種史... 4
(二) 植株型態及繁殖方式... 5
二、 溫度對矮牽牛生長及開花之影響... 6
三、 高溫對草本花卉生長及開花之影響... 6
(一) 高溫對淨光合作用速率及植株乾重之影響... 6
(二) 高溫對分枝數之影響... 7
(三) 高溫對花芽創始與發育之影響... 8
四、 常用之耐熱選拔指標... 10
(一) 花粉選拔... 10
(二) 細胞膜熱穩定性... 13
五、 細胞膜熱穩定性之遺傳... 18
六、 影響花朵性狀之基因... 20
(一) 影響矮牽牛花朵型態之基因... 20
(二) 影響矮牽牛花色之基因... 20
(三) 影響花瓣斑紋表現之基因... 21
材料與方法 (Materials and Methods)... 23
植物材料... 23
栽培介質... 23
試驗一、溫度對F1種子系矮牽牛品種細胞膜熱穩定性、生長及花粉萌芽 之影響... 23
試驗二、溫度對營養系矮牽牛品種細胞膜熱穩定性、生長及花粉萌芽之影 響... 26
試驗三、矮牽牛品種授粉之結實率、結種數及蒴果成熟時間... 28
試驗四、溫度對矮牽牛品種自交或雜交後代細胞膜熱穩定性之影響... 29
試驗五、矮牽牛品種雜交後代花色、花瓣脈紋與花藥顏色之表現... 30
統計分析... 31
結 果 (Results)... 32
試驗一、溫度對F1種子系品種矮牽牛細胞膜熱穩定性及生長之影響... 32
(一) 溫度對 F1種子系矮牽牛品種細胞膜膜熱穩定性之影響... 32
(二) 溫度對 F1種子系矮牽牛品種營養生長之影響... 32
(三) 培養溫度對 F1種子系矮牽牛品種花粉萌芽之影響... 33
試驗二、溫度對營養系矮牽牛品種細胞膜熱穩定性、生長及花粉萌芽之影 響... 34
(一) 溫度對營養系矮牽牛品種細胞膜熱穩定性及生長之影響... 34
(二) 蔗糖濃度與培養溫度對營養系矮牽牛品種花粉萌芽之影響... 35
試驗三、矮牽牛品種授粉之結實率、結種數及蒴果成熟時間... 36
(一) F1種子系矮牽牛品種... 36
(二) 營養系矮牽牛品種... 37
試驗四、溫度對矮牽牛品種自交或雜交後代細胞膜熱穩定性之影響... 38
試驗五、矮牽牛品種自交或雜交後代之花色、脈紋與花藥顏色之表現... 39
討 論 (Discussion)... 44
試驗一、溫度對F1種子系矮牽牛品種細胞膜熱穩定性及生長之影響... 44
(一) 溫度對 F1種子系矮牽牛品種細胞膜膜熱穩定性之影響... 44
(二) 溫度對 F1種子系矮牽牛品種生長之影響... 45
(三) 培養溫度對 F1種子系矮牽牛品種花粉萌芽之影響... 46
試驗二、溫度對營養系矮牽牛品種花粉萌芽及生長之影響... 47
(一) 溫度對營養系矮牽牛品種細胞膜熱穩定性及生長之影響... 47
(二) 蔗糖濃度與培養溫度對營養系矮牽牛品種花粉萌芽之影響... 49
試驗三、矮牽牛品種授粉之結實率、結種數及蒴果成熟時間... 52
試驗四、溫度對矮牽牛自交或雜交後代細胞膜熱穩定性之影響... 52
試驗五、矮牽牛品種雜交後代花瓣脈紋與花色之表現... 53
(一) 矮牽牛品種自交或雜交後代花冠顏色之表現... 53
(二) 矮牽牛品種自交或雜交後代脈紋之表現... 54
(三) 矮牽牛品種自交或雜交後代花藥顏色... 54
參考文獻...119
表目錄
表 1. 栽培溫度對種子系矮牽牛‘Primeteme Carmine’、‘Wave Blue’ 及 ‘Tidal
Wave Silver’ 剛成熟完全展開葉 48 ℃水浴 20 分鐘後之相對熱傷害值。 .. 56
表 2. 溫度對營養系矮牽牛 ‘Cascadias Soft Pink’、‘Cascadias Pink Spark Improved’、‘Sunray’ 及 ‘Petitunia Blue Dream’ 植株分枝數、葉長、葉 寬、花數及花徑之影響。... 57
表 3. 溫 度 對 營 養 系 矮 牽 牛 ‘Cascadias Pink Spark Improved’ 、 ‘Petitunia Bordeaux Dream’、‘Sunray’ 及 ‘Petitunia Blue Dream’ 花粉萌芽率之影 響。... 58
表 4. 種子系矮牽牛不同自交或雜交組合之結實率、種子數及蒴果成熟時間。 ... 59
表 5. 營養系矮牽牛不同自交或雜交組合之結實率、種子數、稔實率及蒴果成 熟時間。... 60
表 6. 不同矮牽牛親本自交或雜交後代於 25/20 ℃下之葉片相對熱傷害值分 布。... 61
表 7. 不同矮牽牛親本自交或雜交後代於 30/25 ℃下之葉片相對熱傷害值分 布。... 61
表 8. 矮牽牛雜交組合在不同栽培溫度下之葉片相對熱傷害值之變方分析。 ... 62
表 9. 矮牽牛 ‘Sunray’ (S) 自交後代花冠顏色之分離情形。... 82
表 10. 矮牽牛 ‘Sunray’ (S) 自交後代花瓣脈紋之分離情形。... 82
表 11. 矮牽牛 ‘Sunray’ (S) 自交後代花藥顏色之分離情形。... 82
表 12. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Soft Pink’ (CSP) 雜交後代花冠顏色之 分離情形。... 84
表 13. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Soft Pink’ (CSP) 雜交後代花瓣脈紋之 分離情形。... 84
表 14. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Soft Pink’ (CSP) 雜交後代花藥顏色之 分離情形。... 84
表 15. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Pink Spark Improved’ (CPSI) 雜交後 代花冠顏色之分離情形。... 86
表 16. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Pink Spark Improved’ (CPSI) 雜交後 代花瓣脈紋之分離情形。... 86 表 17. 矮牽牛 ‘Sunray’ (S) × ‘Cascadias Pink Spark Improved’ (CPSI) 雜交後
代花藥顏色之分離情形。... 86 表 18. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Bordeaux Dream’ (PBoD) 雜交後代花
冠顏色之分離情形。... 88 表 19. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Bordeaux Dream’ (PBoD) 雜交後代花
瓣脈紋之分離情形。... 88 表 20. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Bordeaux Dream’ (PBoD) 雜交後代花
藥顏色之分離情形。... 88 表 21. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Blue Dream’ (PBlD) 雜交後代花冠顏
色之分離情形。... 90 表 22. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Blue Dream’ (PBlD) 雜交後代花瓣脈
紋之分離情形。... 90 表 23. 矮牽牛 ‘Sunray’ (S) × ‘Petitunia Blue Dream’ (PBlD) 雜交後代花藥顏
色之分離情形。... 90 表 24. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Sunray’ (S) 雜交後
代花冠顏色之分離情形。... 92 表 25. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Sunray’ (S) 雜交後
代花瓣脈紋之分離情形。... 92 表 26. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Sunray’ (S) 雜交後
代花藥顏色之分離情形。... 92 表 27. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Cascadias Soft Pink’
(CSP) 雜交後代花冠顏色之分離情形。 ... 94 表 28. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Cascadias Soft Pink’
(CSP) 雜交後代花瓣脈紋之分離情形。 ... 94 表 29. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Cascadias Soft Pink’
(CSP) 雜交後代花藥顏色之分離情形。 ... 94 表 30. 矮牽牛‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Bordeaux
Dream’ (PBoD) 雜交後代花冠顏色之分離情形。... 96 表 31. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Bordeaux
Dream’ (PBoD) 雜交後代花瓣脈紋之分離情形。... 96 表 32. 矮牽牛‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Bordeaux
Dream’ (PBoD) 雜交後代花藥顏色之分離情形。... 96 表 33. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Blue Dream’
(PBlD) 雜交後代花冠顏色之分離情形。... 98
表 34. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Blue Dream’
(PBlD) 雜交後代花瓣脈紋之分離情形。... 98 表 35. 矮牽牛 ‘Cascadias Pink Spark Improved’ (CPSI) × ‘Petitunia Blue Dream’
(PBlD) 雜交後代花藥顏色之分離情形。... 98 表 36. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Sunray’ (S) 雜交後代花
冠顏色之分離情形。... 100 表 37. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Sunray’ (S) 雜交後代花
瓣脈紋之分離情形。... 100 表 38. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Sunray’ (S) 雜交後代花
藥顏色之分離情形。... 100 表 39. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Soft Pink’ (CSP)
雜交後代花冠顏色之分離情形。... 102 表 40. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Soft Pink’ (CSP)
雜交後代花瓣脈紋之分離情形。... 102 表 41. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Soft Pink’ (CSP)
雜交後代花藥顏色之分離情形。... 102 表 42. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花冠顏色之分離情形。 ... 104 表 43. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花瓣脈紋之分離情形。 ... 104 表 44. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花藥顏色之分離情形。 ... 104 表 45. 矮牽牛‘Petitunia Bordeaux Dream’ (PBoD) 自交後代花冠顏色之分離情
形。... 106 表 46. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) 自交後代花瓣脈紋之分離
情形。... 106 表 47. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) 自交後代花藥顏色之分離
情形。... 106 表 48. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Petitunia Blue Dream’
(PBlD) 雜交後代花冠顏色之分離情形。... 108 表 49. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Petitunia Blue Dream’
(PBlD) 雜交後代花瓣脈紋之分離情形。... 108 表 50. 矮牽牛 ‘Petitunia Bordeaux Dream’ (PBoD) × ‘Petitunia Blue Dream’
(PBlD)雜交後代花藥顏色之分離情形。... 108 表 51. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Sunray’ (S) 雜交後代花冠顏
色之分離情形。...110 表 52. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Sunray’ (S) 雜交後代花瓣脈
紋之分離情形。...110 表 53. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Sunray’ (S) 雜交後代花藥顏
色之分離情形。...110 表 54. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Soft Pink’ (CSP) 雜
交後代花冠顏色之分離情形。...112 表 55. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Soft Pink’ (CSP) 雜
交後代花瓣脈紋之分離情形。...112 表 56. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Soft Pink’ (CSP) 雜
交後代花藥顏色之分離情形。...112 表 57. 矮 牽 牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花冠顏色之分離情形。 ...114 表 58. 矮 牽 牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花瓣脈紋之分離情形。 ...114 表 59. 矮 牽 牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Cascadias Pink Spark
Improved’ (CPSI) 雜交後代花藥顏色之分離情形。 ...114 表 60. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Petitunia Bordeaux Dream’
(PBoD) 雜交後代花冠顏色之分離情形。...116 表 61. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Petitunia Bordeaux Dream’
(PBoD) 雜交後代花瓣脈紋之分離情形。...116 表 62. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) × ‘Petitunia Bordeaux Dream’
(PBoD) 雜交後代花藥顏色之分離情形。...116 表 63. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) 自交後代花冠顏色之分離情
形。...118 表 64. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) 自交後代花瓣脈紋之分離情
形。...118 表 65. 矮牽牛 ‘Petitunia Blue Dream’ (PBlD) 自交後代花藥顏色之分離情
形。...118
圖目錄
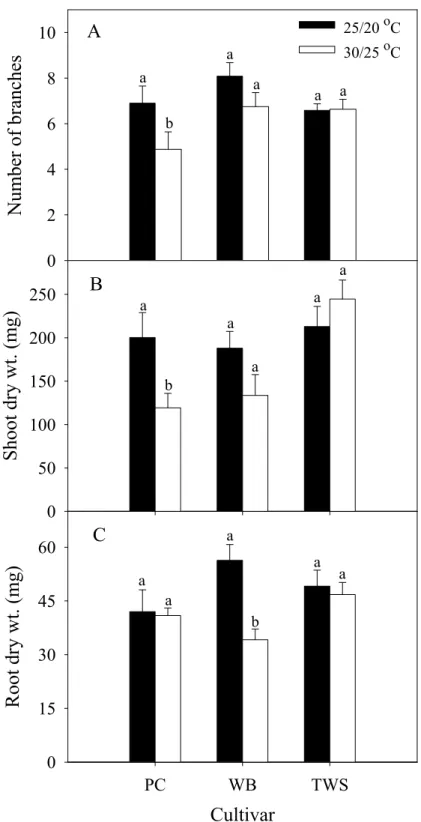
圖 1. 水浴溫度對種子系矮牽牛 ‘Primetime Carmine’、‘Wave Blue’ 及 ‘Tidal Wave Silver’ 葉片相對熱傷害值之影響。 ... 63 圖 2. 溫度對種子系矮牽牛‘Primetime Carmine’ (PC)、‘Wave Blue’ (WB) 與
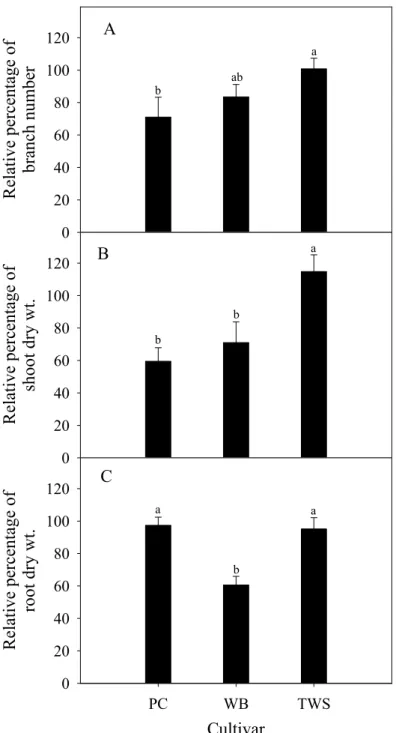
‘Tidal Wave Silver’ (TWS) 分枝數 (A)、枝梢乾重 (B) 與根乾重 (C)之 影響。... 64 圖 3. 相較於 25/20 ℃,種子系矮牽牛‘Primetime Carmine’ (PC)、‘Wave Blue’
(WB) 與‘Tidal Wave Silver’ (TWS) 於 30/25 ℃之相對分枝數 (A)、相對 枝梢乾重 (B) 與相對根乾重 (C)。 ... 65 圖 4. 相較於 25/20 ℃,種子系矮牽牛‘Primetime Carmine’ (PC)、‘Wave Blue’
(WB) 與‘Tidal Wave Silver’ (TWS) 於 30/25 ℃之相對根:地上部乾重。
... 66 圖 5. 栽培溫度及培養溫度對種子系矮牽牛 ‘Primetime Carmine’、‘Wave Blue’
與 ‘Tidal Wave Silver’ 花粉萌芽率之影響。... 67 圖 6. 水浴溫度對營養系矮牽牛 ‘Cascadias Soft Pink’、‘Cascadias Pink Spark
Improved’、‘Sunray’ 及 ‘Petitunia Blue Dream’ 葉片相對熱傷害值之影 響。... 68 圖 7. 溫 度 對 矮 牽 牛 ‘Cascadias Soft Pink’ (CSP) 、 ‘Cascadias Pink Spark
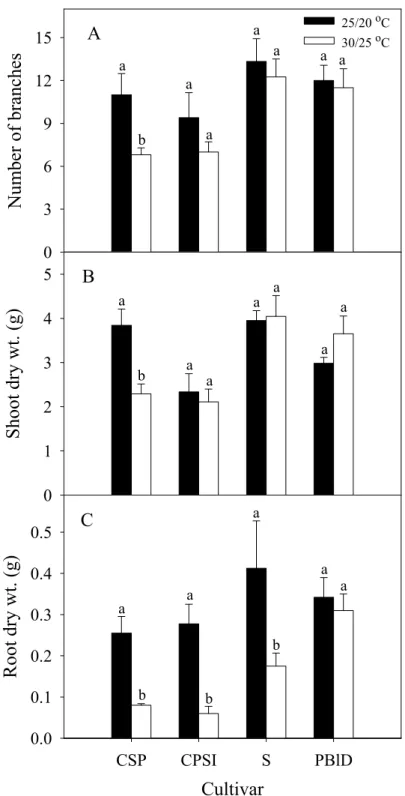
Improved’ (CPSI)、‘Sunray’ (S) 及 ‘Petitunia Blue Dream’ (PBlD) 分枝數 (A)、枝梢乾重 (B) 與根乾重 (C) 之影響。... 69 圖 8. 相較於 25/20 ℃,營養系矮牽牛‘Cascadias Soft Pink’ (CSP)、‘Cascadias
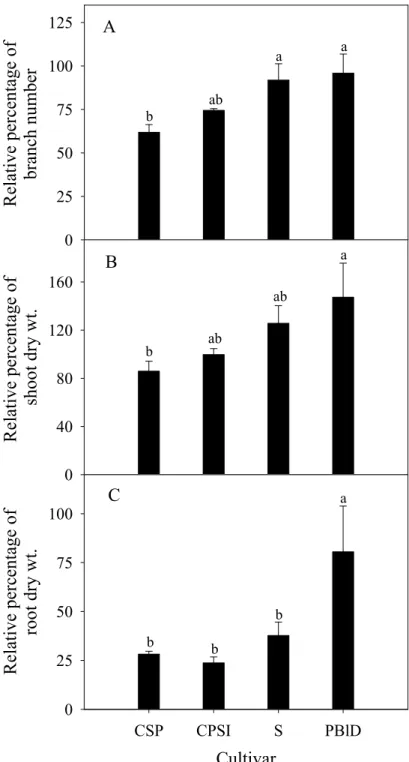
Pink Spark Improved’ (CPSI)、‘Sunray’ (S)、‘Petitunia Blue Dream’ (PBlD) 於30/25 ℃之相對分枝數 (A)、相對枝梢乾重 (B)與相對根乾重 (C)。.... 70 圖 9. 溫度對營養系矮牽牛 ‘Cascadias Soft Pink’ (CSP)、‘Cascadias Pink Spark
Improved’ (CPSI)、‘Sunray’ (S) 與 ‘Petitunia Blue Dream’ (PBlD) 根:地 上部乾重之影響。... 71 圖10. 相較於 25/20 ℃,營養系矮牽牛 ‘Cascadias Soft Pink’ (CSP)、‘Cascadias
Pink Spark Improved’ (CPSI)、‘Sunray’ (S)、‘Petitunia Blue Dream’ (PBlD) 於30/25 ℃之相對根:地上部乾重。 ... 72 圖 11. 蔗糖濃度對矮牽牛 ‘Petitunia Bordeaux Dream’ 和 ‘Petitunia Blue
Dream’ 花粉萌芽率之影響。 ... 73 圖 12. 溫度對矮牽牛‘Cascadias Pink Spark Improved’、‘Petitunia Bordeaux
Dream’、‘Sunray’、‘Petitunia Blue Dream’ 花粉萌芽率之影響。... 74
圖 13. 矮牽牛之雜交授粉步驟。 ... 75
圖 14. 分別以矮牽牛‘Cascadias Pink Spark Improved’ (A)、‘Petitunia Bordeaux Dream’ (B)、‘Sunray’ (C) 及 ‘Petitunia Blue Dream’ (D) 為種子親所得之 種子。... 76
圖 15.矮牽牛‘Primetime Carmine’、‘Wave Blue’及‘Tidal Wave Silver’為母本或 父本之雜交後代於栽培溫度25/20 ℃及 30/25 ℃下之相對熱傷害值頻度 分布。... 77
圖 16. 矮牽牛‘Sunray’ (A)、‘Cascadias Soft Pink’ (B)、‘Cascadias Pink Spark Improved’ (C)、‘Petitunia Bordeaux Dream’ (D)及‘Petitunia Blue Dream’ (E) 之花色表現。... 78
圖 17. 矮牽牛後代花色之區別標準。 ... 79
圖 18. 矮牽牛後代花瓣脈紋程度區別標準。 ... 80
圖 19. 矮牽牛 ‘Sunray’自交之後代花色表現。 ... 81
圖 20. 矮牽牛 ‘Sunray’ × ‘Cascadias Soft Pink’ (♀ × ♂) 之後代花色表現。... 83
圖 21. 矮牽牛 ‘Sunray’ × ‘Cascadias Pink Spark Improved’ (♀ × ♂) 之後代花 色表現。... 85
圖 22. 矮牽牛 ‘Sunray’ × ‘Petitunia Bordeaux Dream’ (♀ × ♂) 之後代花色表 現。... 87
圖 23. 矮牽牛 ‘Sunray’ × ‘Petitunia Blue Dream’ (♀ × ♂) 之後代花色表現。 ... 89
圖 24. 矮牽牛 ‘Cascadias Pink Spark Improved’ × ‘Sunray’ (♀ × ♂) 之後代花 色表現。... 91
圖 25. 矮牽牛 ‘Cascadias Pink Spark Improved’ × ‘Cascadias Soft Pink’ (♀ × ♂) 之後代花色表現。... 93
圖 26. 矮牽牛 ‘Cascadias Pink Spark Improved’ × ‘Petitunia Bordeaux Dream’ (♀ × ♂) 之後代花色表現。 ... 95
圖 27. 矮牽牛 ‘Cascadias Pink Spark Improved’ × ‘Petitunia Blue Dream’ (♀ × ♂) 之後代花色表現。... 97
圖 28. 矮牽牛 ‘Petitunia Bordeaux Dream’ × ‘Sunray’ (♀ × ♂) 之後代花色表 現。... 99
圖 29. 矮牽牛 ‘Petitunia Bordeaux Dream’ × ‘Cascadias Soft Pink’ (♀ × ♂) 之 後代花色表現。... 101
圖 30. 矮牽牛 ‘Petitunia Bordeaux Dream’ × ‘Cascadias Pink Spark Improved’ (♀ × ♂) 之後代花色表現。 ... 103
圖 31. 矮牽牛 ‘Petitunia Bordeaux Dream’自交之後代花色表現。... 105 圖 32. 矮牽牛 ‘Petitunia Bordeaux Dream’ × ‘Petitunia Blue Dream’ (♀ × ♂) 之
後代花色表現。... 107 圖 33. 矮牽牛 ‘Petitunia Blue Dream’ × ‘Sunray’ (♀ × ♂) 之後代花色表現。 .... 109 圖 34. 矮牽牛 ‘Petitunia Blue Dream’ × ‘Cascadias Soft Pink’ (♀ × ♂) 之後代
花色表現。...111 圖 35. 矮牽牛 ‘Petitunia Blue Dream’ × ‘Cascadias Pink Spark Improved’ (♀ ×
♂) 之後代花色表現。...113 圖 36. 矮牽牛 ‘Petitunia Blue Dream’ × ‘Petitunia Bordeaux Dream’ (♀ × ♂) 之
後代花色表現。...115 圖 37. 矮牽牛 ‘Petitunia Blue Dream’ 自交後代花色表現。 ...117
中文摘要
進行耐熱育種時,若能建立耐熱指標和瞭解耐熱性狀之遺傳,可提高育種效 率。本研究分別比較高溫對矮牽牛 (Petunia ×hybrida Hort. Vilm. Andr.) F1種子系品 種或營養系品種之細胞膜熱穩定性、生長特性及花粉萌芽之影響,並進一步進行 雜交,比較耐熱程度不同之親本雜交所得後代細胞膜熱穩定性表現,和其花朵型 態之分離情形。
將F1種子系品種 ‘Primetime Carmine’、‘Wave Blue’ 及 ‘Tidal Wave Silver’ 分 別置於25/20 ℃及 30/25 ℃栽培 18 天後,取剛成熟之完全展開葉,利用不同水浴 溫度25、42、44、46、48、50、52、54、56℃加熱其葉圓片 20 分鐘,相對熱傷害 值 (relative injury, RI) 隨水浴溫度之上升而呈 S 形曲線關係。參試 F1種子系矮牽 牛RI 值 S 形曲線之反曲中點溫度約為 48 ℃,因此可利用 48 ℃水浴溫度鑑別參試 F1種子系矮牽牛品種之耐熱性。栽培於30/25 ℃之‘Wave Blue’及‘Tidal Wave Silver’
者之RI 值較低,且 S 形曲線較 25/20 ℃者有向右移之趨勢,顯示兩品種可被熱馴 化,表現較高之耐熱性,而 ‘Primetime Carmine’則為熱敏感品種。高溫 30/25 ℃造 成分枝數與枝梢乾重下降情形,與細胞膜熱穩定性有類似之表現,耐熱品種 ‘Tidal Wave Silver’ 之分枝數與枝梢乾重較不易因高溫而下降,而熱敏感之 ‘Primetime Carmine’ 下降較顯著。
營養系矮牽牛‘Cascadias Soft Pink’、‘Petitunia Bordeaux Dream’、‘Sunray’及
‘Petitunia Blue Dream’,經高溫 30/25 ℃栽培後細胞膜熱穩定性亦可被熱馴化,耐 熱程度佳,且以50 ℃為 S 形曲線之反曲點,較參試 F1種子系矮牽牛品種者之48 ℃
高。顯示參試營養系矮牽牛品種之耐熱性較參試 F1種子系矮牽牛品種佳。營養系
矮牽牛‘Sunray’ 及‘Petitunia Blue Dream’之花粉於 30 ℃培養者萌芽率較高,而
‘Cascadias Soft Pink’ 及‘Petitunia Bordeaux Dream’則以 25 ℃培養者萌芽率較高。高 溫30/25 ℃造成 Cascadias Soft Pink’及‘Petitunia Bordeaux Dream’分枝數、枝梢及根 乾重之下降較 ‘Sunray’及‘Petitunia Blue Dream’顯著。結果顯示參試營養系矮牽牛
品種間之耐熱程度仍有差異。
經30/25 ℃熱馴化 18 天後,參試 F1種子系矮牽牛雜交後代之葉圓片利用48 ℃ 水浴加熱 20 分鐘後,以矮牽牛 ‘Primetime Carmine’ 為花粉親者之後代 RI 值較 高,而以 ‘Wave Blue’ 及 ‘Tidal Wave Silver’ 為花粉親者之 RI 值較低。以 ‘Tidal Wave Silver’ 為花粉親者,後代 RI 值分布高峰較低,介於 30% - 40%,而以
‘Primetime Carmine’ 為花粉親者之 RI 分布高峰則較高,介於 45% - 55%間,顯示 雜交後代細胞膜熱穩定性可能受花粉親之影響較大。
調查參試營養系矮牽牛經自交或雜交後,後代之花冠顏色、花瓣脈紋表現程 度與花藥顏色之分離情形,結果顯示白花品種 ‘Sunray’ 及藍花品種 ‘Petitunia Blue Dream’ 之自交後代花色與親本相同,而桃紫色花之‘Petitunia Bordeaux Dream’後代花色會分離為粉紅色、桃紫色及紫色。不同花色之矮牽牛雜交後,後 代之花色分布介於兩親本間。花冠上具脈紋之品種,與花冠上沒有脈紋之品種
‘Cascadias Soft Pink’雜交所得後代,大部分為具有脈紋者,而有少量後代沒有脈紋 之表現。具黃色花藥之矮牽牛‘Sunray’及‘Cascadias Soft Pink’進行自交或雜交後,
其後代之花藥顏色大部分為黃色或白色。而具紫色花藥之品種‘Cascadias Pink Spark Improved’、‘Petitunia Bordeaux Dream’、‘Petitunia Blue Dream’自交或雜交,
或與黃色花藥品種雜交時,後代之花藥顏色較多,分別有白色、黃色與紫色。
Summary
Establishing heat-tolerant indicators and understanding the inheritance of the associated characteristics for heat-tolerance breeding of petunia are desirable. The objectives of this study were 1) to determine the high temperature effects on cell membrane thermostability (CMT) as measured with electrolyte leakage technique, growth and pollen germination of seed- or vegetatively-propagated petunias, and 2) to compare the CMT and flower characteristics of selfed or cross progenies obtained from petunia parents with differential heat-tolerance.
Plants of F1 seed-propagated cultivars Primetime Carmine, Wave Blue, and Tidal Wave Silver were treated at 25/20 and 30/25 for 18 days. The recently developed ℃ leaves of these cultivars were evaluated for CMT measured at water bath temperatures of 25, 42, 44, 46, 48, 50, 52, 54, and 56℃ for 20 mins. The relationship between the relative injury (RI) value occurring in leaf tissue discs of petunia cultivars and treatment temperature was sigmoidal. The RI values near the midpoint of the sigmoid response curve occurred at the 48 ℃ treatments. The cultivars Wave Blue and Tidal Wave Silver exhibited heat-acclimation and had lower RI values than the ‘Primetime Carmine’.
Reduced branch number and shoot dry weight were more apparent in the heat-intolerant
‘Primetime Carmine’ than the heat-tolerant ‘Tidal Wave Silver’ after grown at 30/25 ℃,
as compared with those at 25/20 .℃
All the vegetatively-propagated cultivars Cascadias Soft Pink, Petitunia Bordeaux Dream, Sunray, and Petitunia Blue Dream exhibited heat-acclimation as shown with reduced RI under the 30/25 ℃ treatment. The RI values of the vegetatively- propagated cultivars near the midpoint of the sigmoid response curve occurred at water bath temperature of 50 , higher than the 48 ℃ ℃for seed-propagated cultivars tested in this study. Most pollens germinated at 30 in ‘Sunray’ and ‘Petitunia Blue Dr℃ eam’, as compared with 25 in ‘Cascadias Soft Pink’ and ‘Petitunia Bordeaux Dream’. High ℃ temperature at 30/25 decreased more branch number, shoot and root dry weights in ℃
‘Cascadias Soft Pink’ and ‘Petitunia Bordeaux Dream’ than ‘Sunray’ and ‘Petitunia Blue Dream’. Results suggested that vegetatively-propagated cultivars had higher heat-tolerance than seed-propagated cultivars. However, differential heat-tolerance existed in these vegetatively-propagated cultivars.
Progeny seedlings were raised and moved to 30/25 ℃ for 18 days, and then leaf RI of these progenies was measured at water bath temperature of 48 ℃ for 20 mins. Higher RI vales were measured in progenies obtained from ‘Primetime Carmine’as pollen parent, as compared with lower RI vales in progenies from ‘Wave Blue or ‘Tidal Wave Silver’ as pollen parents. RI ranged between 30 % and 40 % in most progenies obtained from ‘Tidal Wave Silver’ as the pollen parent, while RI values were between 45 % and
55 % in most progenies from ‘Primetime Carmine’ as the pollen parent. Results suggested that CMT of petunia progenies at 30/25 °C were largely affected by the pollen parents.
Corolla color, conspicuousness of vein on the upper corolla, and anther color segregation were recorded in the selfed or cross progenies from vegetatively-propagated cultivars. Selfed progenies and the parents had the same corolla color in white-flowered
‘Sunray’ and blue-flowered ‘Petitunia Blue Dream’. Corolla color exhibited pink, violet red or purple in the selfed progenies of the violet red-flowered ‘Petitunia Bordeaux Dream’. Corolla color regimes of cross progenies were between those parents with different colors. Conspicuousness of vein was observed in most progenies from the vein less ‘Cascadias Soft Pink’ crossed with those with veins on the upper corolla. Anther color almost exhibited yellowish white or yellow in the selfed or cross progenies from
‘Sunray’ and ‘Cascadias Soft Pink’ with yellow anthers. However, anther color exhibited yellowish white, yellow, or violet in the selfed or cross progenies from
‘Cascaidas Pink Spark Improved’, ‘Petitunia Bordeaux Dream’, and ‘Petitunia Blue Dream’ with violet anthers.
前 言 (Introduction)
矮 牽 牛 (Petunia ×hybrida Hort. Vilm. Andr.) 原 生 於 南 美 洲 , 為 茄 科 (Solanaceae) 矮牽牛屬植物,栽培品種多,且花形、株形及花色變化多,為世界 重要之花卉類作物,在美國可供花壇、盆花與吊盆類花卉作物之生產。經美國農 部統計,矮牽牛於2007 年之總生產值達 11 億美金 (USDA, 2008),而在臺灣,矮 牽牛之銷售量為花壇植物中之第三位,每年可生產164 萬株 (林,2002)。
矮牽牛在臺灣一般作為秋播冬春開花之ㄧ年生草本花卉,因台灣夏季平均溫 度高,最高平均溫度於7 月可達 34.1 ℃,且台灣之雨季集中在夏季,較少於夏季 進行生產。近年來,日本Suntory 公司、美國 PanAmerican 公司及以色列 Danziger 公司陸續育出號稱可耐高溫之品種,使矮牽牛漸漸有潛力成為夏季之新興花卉作 物 (Craig, 2003)。因此希望利用已育出之耐熱品種,將耐熱性導入觀賞性佳但熱 敏感之品種,育出適合台灣夏季生產之矮牽牛品種。
進行耐熱育種工作時,常耗費大量時間與空間,因此若能建立耐熱指標,將 有助於品種耐熱性之鑑定,並增加後代耐熱性之篩選效率。利用高溫對於植株生 長反應作為耐熱鑑定標準較費時,因此若能利用花粉萌芽或細胞膜熱穩定性 (cell membrane thermostability, CMT) 等耐熱生理指標,可節省空間、加快篩選速度、
為非破壞性測試且不需昂貴儀器或是困難技術 (Marcum, 1998; Hormaza and Herrero, 1992; 1996)。
孢子體和配子體世代之基因表現有重疊性、面對逆境時,孢子體與配子體會 有相同之反應、而提高花粉間之競爭力,亦可提高其後代於種子數、種子重量及 株高之表現,因此可利用花粉作為耐熱篩選指標 (Hormaza and Herrero, 1992;
1996)。耐熱程度不同之品種,花粉可萌芽之最適溫度不同,可萌芽之最高與最低 溫度亦不同,耐熱品種之花粉萌芽最適溫度較高 (Lyakh et al., 1991),且經高溫預 處理後之萌芽率仍佳 (Reddy and Kakani, 2007)。顯示可利用花粉萌芽溫度作為品
種間耐熱性之鑑定標準。
細胞膜熱穩定性已證實與許多作物之耐熱程度有相關 (Chen et al., 1982;
Howarth et al., 1997; Li et al., 1991; Srinivasan et al, 1996; Yeh and Lin, 2003)。高溫 逆境會破壞植物蛋白質穩定性,使細胞膜的流動性與滲透性改變 (Ismail and Hall, 1999; Sangwan et al., 2002),因此可藉測定葉片經短暫高溫處理後之相對熱傷害值 (relative injury, RI),來檢測植物之細胞膜熱穩定性,以提供快速且有效之作物耐 熱性鑑定方法。
林 (2006) 指出,不同栽培溫度會影響矮牽牛之營養生長情形,且品種間有 顯著差異。隨栽培溫度由 16 ℃升高至 27 ℃,耐熱品種 ‘Tidal Wave Silver’ 與
‘Wave Blue’ 之分枝數下降較少,枝梢乾重與根乾重則上升,也沒有根系變弱的情 形;而熱敏感品種 ‘Primetime Carmine’ 之分枝數與根乾重則會因高溫處理而顯著 下降。
林 (2006) 亦指出,栽培於 27 ℃之矮牽牛經高溫水浴後,葉片相對熱傷害值 依品種耐熱程度而異,可作為品種間之耐熱指標。耐熱品種 ‘Tidal Wave Silver’ 與
‘Wave Blue’ 葉圓片經 50 ℃水浴 20 分鐘後,可誘導其耐熱性,其 RI 值較熱敏感 品種 ‘Primetime Carmine’ 低。但林 (2006) 並未以不同水浴溫度處理測試矮牽牛
於不同栽培溫度下之RI 值,因此若欲利用單一溫度區別品種間之差異,並確定矮
牽牛不同品種之熱馴化程度,仍需進行進一步之試驗。
矮牽牛一般利用 F1種子繁殖,於臺灣為秋冬季草本花卉,自 90 年代育出利 用營養繁殖者後,可利用於夏季之生產,因此利用牽插繁殖之矮牽牛耐熱性可能 較佳。而林 (2006) 並無使用營養系作為試驗材料,因此本論文將探討高溫對參
試F1種子系品種與參試營養系矮牽牛品種生長之影響,期能確認其耐熱指標之再
現性,並應用於耐熱育種上,探討後代耐熱性之遺傳。並調查雜交後代花朵性狀 之表現,以供育種參考。主要研究內容如下:
一、 從高溫對細胞膜熱穩定性、植株生長及花粉萌芽之影響,探討參試 F1種子 系矮牽牛品種間耐熱性之差異,並尋找耐熱指標。
二、 以高溫對細胞膜熱穩定性、植株生長及花粉萌芽之影響,探討耐熱指標於參 試營養系矮牽牛品種之應用。
三、 利用細胞膜熱穩定性探討矮牽牛自交或雜交後代之耐熱性遺傳。
四、 矮牽牛自交或雜交後代之花色與花瓣脈紋之表現。
前人研究 (Literature Review)
一、 矮牽牛之簡介
(一) 親源及育種史
矮牽牛 (Petunia ×hybrida Hort. Vilm. Andr.) 為茄科 (Solanaceae) 一年生草本 花卉,原生於南美洲阿根廷、巴西及烏拉圭,大部分之原生種原生於巴西東南部,
由 P. axillaris Lam. 與 P. integrifolia Hook. 雜交而來。於 1823 年在烏拉圭採集到 矮牽牛 P. axillaries Lam.,並送至法國開始栽種,P. axillaries Lam.之株形為直立株 型、花朵為黃白色花,而花筒較細且長,花朵之開張角度較平,於下午會散發出 濃郁之香味;於1831 年在布宜諾斯艾利斯採集到 P. integrifolia Hook.,為懸垂型 株型、葉片橢圓至披針形、花色則為紫羅蘭色花 (Griesbach, 2006;Sink, 1984)。
於 1838 年育出花喉有脈之品種 P. vittata;於 1844 年時育出花瓣鑲邊之品種 P.
nixemi,德國人藉由改良此品種,於 1855 年育出重瓣之品種。接著由多花型
(multiflora) Burpee’s Defiance 品系之突變株中選拔出大花型 (grandiflora) 矮牽 牛,此大花型由一對顯性基因GG 所調控。而於 1988 年,利用基因體突變育出四 倍體之大花型矮牽牛,Shepherd 女士再利用此品系於 1888 年育出風靡 1930 年代 之 ‘California Giant’ 品種,此時之矮牽牛以盆花生產為主,因其生長習性與植株 活力尚不足以作為花壇植物之生產 (Griesbach, 2006;Sink, 1984)。
在1930 年代之前,矮牽牛之育種方式以集團選拔為主,並利用營養繁殖方式
繁殖優良品系。進入1930 年代之後,選拔方式改為單株選拔,並利用種子繁殖優
良品系,而1940 年代之後,則開始生產 F1種子,美國 PanAmerican Seed 公司也 推出一個利用種子繁殖之大花型矮牽牛。1950 年代之前,矮牽牛所育出之花色,
以粉紅、紫、桃紫以及白色為主,直至50 年代,才育出紅色之品種,如美國 Bodger 公司所育出之多花型紅色矮牽牛 ‘Fire Chief’ 與 PanAmerican Seed 公司創辦人 Claude Hope 所育出之大花型紅色矮牽牛 ‘Comanche’,日本 Sakata 育種公司股東
須田畯一郎先生曾稱讚此品種為 ‘魔性之花’ (Griesbach, 2006; Sink, 1984)。
進入1970 年代後,因長時間之育種,許多矮牽牛原生種之優良特性,如抗病
性、抗逆境及耐蟲性等,於育種過程中逐漸消失,且消費者偏好的開始改變,使 得矮牽牛之市場逐漸萎縮。直至育種家將一些舊有品種和野生種加入,並成為新 的育種親本,矮牽牛市場萎縮之情形才得以改善 (Griesbach, 2006; Sink, 1984)。首 先於1989 年,日本 Suntory 公司,派出探險隊至原生地巴西採集原生種並用為育 種親本,育出 ‘Surfinia’ 品種,此品種為懸垂性品種,因其懸垂特性與親本之耐候 性,此品種號稱可耐熱。而於1993 年,美國 PanAmerican Seed 公司則育出同為懸 垂株形之 ‘Wave’ 系列,此系列依株型又可分為 ‘Easy Wave’、‘Wave’ 及 ‘Tidal Wave’,同樣號稱具耐熱性。於 2004 年,荷蘭 HEMGENETICS 公司則育出直立株 形、大花型之 ‘Limbo’ 系列與多花型之 ‘Membo’ 系列,除具耐熱性外,此兩品 種遇高溫時,節間較不易抽長,為矮牽牛之重要特性之ㄧ。目前市面上堪稱可耐 熱之品種尚有美國Bodger 公司所育出之 ‘Tiny Tunias’ 及 ‘Trumpets’ 系列,以色 列Danziger 公司所育出之 ‘Cascadias’ 及 ‘Petitunia’ 等系列 (Griesbach, 2006)。
(二) 植株型態及繁殖方式
接近 170 年之育種發展,矮牽牛依其植株型態和花朵大小,可分為大花型
(grandiflora)、重瓣大花型 (double grandiflora)、多花型 (multiflora)、重瓣多花型 (double multiflora)、懸垂型 (cascading) 及迷你型 (miniature)等六種,但大花型和 多花型矮牽牛因長時間之雜交,已難去分辨兩者之間之差異,因此又多一分類為 花較大之多花型 (floribunda) 矮牽牛。上述各型態之矮牽牛,以大花型矮牽牛之耐 候 性 較 差 , 而 迷 你 型 與 多 花 型 之 矮 牽 牛 耐 候 性 較 佳 , 經 逆 境 後 可 快 速 復 原 (Armitage, 2001)。
矮牽牛之繁殖方法,大致可分為兩種,分別為種子繁殖系統與營養繁殖系統,
前者為矮牽牛傳統上主要之繁殖方法,進入1900 年代以後,出現利用扦插繁殖之
品種,第一個出現之品種為 ‘Supertunia’,之後漸漸利用營養繁殖之品種作為矮牽
牛盆花與吊盆之使用 (Armitage, 2001; Craig, 2003)。
二、 溫度對矮牽牛生長及開花之影響
當溫度大於20 ℃,矮牽牛 ‘Dreams Red’ 之淨光合作用速率會開始下降,暗 呼吸作用速率則會上升 (van Iersel, 2003)。隨栽培日溫由 10 ℃提高至 30 ℃,矮牽 牛 ‘Snow Cloud’ 之株高亦會增加,日溫減去夜溫,即 DIF (different between day and night temperature) 對株高之影響較小,而夜溫則不會影響株高;矮牽牛之側枝 長度與分枝數則會隨栽培日溫增加而下降 (Kaczperski et al., 1991);當氣溫為 21 ℃ 時,根溫25 ℃可使側枝伸長,亦可促進地上部乾重之累積與葉面積之增加 (Merritt and Kohl., 1982)。
當溫度由15 ℃開始增加,矮牽牛 ‘Express Blush Pink’ 於 21 ℃下花芽分化最 快,最早進入花芽分化 (Adams et al., 1998),且當溫度大於 23 ℃時。花芽發育速 度則會開始趨於平緩,到花日數不再減少 (Adams et al., 1999)。Kaczperski 等人 (1991) 亦指出矮牽牛 ‘Snow Cloud’ 之到花日數隨與溫度上升而減少呈二次負相 關,溫度大於22 ℃後曲線會趨於平緩。
三、 高溫對草本花卉生長及開花之影響
(一) 高溫對淨光合作用速率及植株乾重之影響植物遇高溫逆境時,淨光合作用速率會下降,且為最敏感之生理反應之ㄧ,
因此可藉由量測高溫下之淨光合作用速作為植物對高溫適應性之優良指標,將天 竺葵 (Pelargonium ×hortorum L. H. Bail.)、萬壽菊 (Tagetes patula L.)、三色菫 (Viola ×wittrockiana Gams.) 與矮牽牛從 0℃開始,每 3 個小時將溫度升高 5℃,直 至溫度升至36℃,當溫度大於 15℃後,四種草本花卉之淨光合作用開始下降 (van Iersel, 2003);矮牽牛、藿香 [Agastache urticifolia (Benth.) O. Kuntze]、觀賞辣椒 (Capsicum annuum L.)、日日春 [Catharanthus roseus (L.) G. Don]、銀葉菊 (Cineraria maritime L.) 與藍雪花 (Plumbago auriculata Lam.) 等六種花壇植物種植於 20-40
℃下,當栽培溫度高於25 ℃,凈光合作用速率會開始下降,而矮牽牛於 40 ℃下 之淨光合作用率僅為25 ℃下之 25% (Niu et al., 2006)。
而不同物種或品種光合作用對溫度之適應性依植株之原生地氣候與其栽培溫 度所改變 (Berry and Björkman, 1980)。原生於歐洲之鼠尾草植物 (Salvia greggii A.
Gray Furman’s Red),遇逆境會發生簇生化,栽培日溫由 20 ℃升至 40 ℃,光合作
用速率下降情形較不明顯,但原生於美洲大陸鼠尾草屬植物,栽培於40 ℃之高溫
導致光合作用速率下降之情形較顯著,植株甚至無法存活;而高溫亦會造成鼠尾
草屬植物枝梢乾重下降,原生歐洲之鼠尾草屬植物,當栽培溫度高於 30℃後枝梢
乾重才會開始少量下降;而原生美洲之鼠尾草屬植物,當日溫大於22 ℃,枝梢乾
重及顯著下降 (Lasseigne et al., 2007);由此可知,高溫會降低植株淨光合作用速 率,進而影響植株乾物重之累積 (Lasseigne et al., 2007; Niu et al., 2006; Warner and Erwin, 2005)。高溫造成花卉類作物乾重與乾重累積速率降低情形尚出現於金魚草 (Antirrhinum majus L.)、金盞花 (Calendula officinalis L.)、非洲鳳仙花 (Impatiens wallerana Hook. F.) 與龍頭花 (Minulus ×hybridus Hort. ex Siebert & Voss) (Warner
and Erwin, 2005)、桔梗 (Platycodon grandiflorus (Jacq.) A. DC.) (Park et al., 1998) 與 三色菫 (Viola ×wittrockiana Gams.) (Warner and Erwin, 2006)。隨栽培溫度由 16 ℃ 增加至33 ℃,矮牽牛耐熱品種 ‘Tidal Wave Silver’ 之枝梢乾重與根乾重皆會顯著 上升,不易因高溫對根系造成不良影響,但熱敏感品種 ‘Primetime Carmine’ 之根 乾重則會顯著下降,顯示高溫造成植株枝梢乾重下降之情形會依其耐熱程度不同 而異 (林,2006)。
(二) 高溫對分枝數之影響
高溫會導致作物分枝數下降,聖誕紅 (Euphorbia pulcherrima Willd.) 栽培溫度 高於27 ℃時,會抑制腋芽之分化與發育,導致盲芽 (blind leaf axils) 之形成,最 後使分枝數下降,且品種間對高溫之敏感度不同 (Faust and Heins, 1996)。於高溫 33/27 ℃栽培之菊花 (Dendranthema ×grandiflorum
(Ramat.) Kitamural
) 腋芽會喪失分化能力,因腋芽之分生組織會提早分化,且剩餘的分生組織 (residual meristem) 會不正常分化,導致不正常腋芽之產生,品種間耐熱程度之不同會影響分枝數下 降情形 (Schoellhorn et al., 2001);高土溫會抑制菊花熱敏感品種腋芽之萌芽數 (Schoellhorn et al., 1996)。而高日溫 35 ℃則會導致菊花側芽數減少 (Faust and Heins, 1992);但如於腋芽分化完成後始給予高溫處理,則不會降低分枝數 (Faust and Heins, 1996; Schoellhorn et al., 1996)。高溫導致花卉類作物分枝數下降之情形亦見 於 四 季 秋 海 棠 (Begonia ×semperflorens-cultorum hybrids) (Schoellhorn et al., 2001)、非洲菊 (Gerbera jamesonii H. Bolus. ex Hook. F.) (陳,2007)、非洲鳳仙花 (Schoellhorn et al., 2001)、矮牽牛 (林,2006; Kaczperski et al., 1991)、桔梗 (Park et al., 1998) 與三色菫 (Warner and Erwin, 2006)。
當栽培溫度由25 / 20 ℃升至 35 / 30 ℃,非洲菊耐熱品種 ‘Monte Crysto’ 之分 枝數並未因高溫而下降,但熱敏感品種 ‘Rosalin’ 之植株分枝數則會減少 (陳,
2007)。矮牽牛 ‘Snow Cloud’ 之側枝數量與側枝長度隨栽培溫度由 10 ℃上升至 30
℃而下降 (Kaczperski et al., 1991);將栽培日夜溫度由 20/15 ℃升至 30/25 ℃,矮牽 牛耐熱品種 ‘Tidal Wave Silver’ 和 ‘Wave Blue’ 減少 3 個分枝數,而較不耐熱之矮 牽牛 ‘Fantasy Red’ 與 ‘Primetime Carmine’ 則分別減少了 8.0 及 4.7 個分枝 (林,
2006) 。綜上所述,高溫會導致矮牽牛分枝數下降,且品種間會有差異,因此可作 為品種間耐熱程度之鑑定標準。
(三) 高溫對花芽創始與發育之影響
栽培高溫30/26 ℃會延遲菊花之花芽創始與分化,造成開花熱延遲 (heat delay) (Whealy et al., 1987),可能因開花時生殖生長與營養生長會競爭光合作用同化產物 (Dinar and Rudich, 1985);溫度由 20 ℃升至 25 ℃,朱槿屬之植物 (Hibiscus sp.) 之 花下葉片數會增加 (Warner and Erwin, 2001);而金盞花、非洲菊、三色菫於 32 ℃ 處理之花下葉片數會較20 ℃下者多,亦有開花熱延遲 (Warner and Erwin, 2005) 之 現象發生。顯示因高溫導致花葉片數增加且不利於花芽之發育;而矮牽牛於日均
溫10 ℃至 25 ℃間,到花日數會顯著下降,而當日均溫大於 25 ℃時,因花苞消蕾 束增加,導致開花時間延遲 (Kaczperski et al., 1991)。
花朵數目與花朵大小為花卉類作物之重要觀賞品質,但兩者常因栽培溫度過 高而下降 (Niu et al., 2001),維持花芽發育所需之能量為植物生殖生長時最主要之 積儲 (sink) 來源,因此植物養分之吸收分配是引響花朵形態之重要因素 (Aloni et al., 1999);月見草 (Oenothera fruticosa L.) 於 15.2 ℃栽培者之花數為 29.8 ℃者之 六倍 (Clough et al., 2001);桔梗栽培於 14 ℃之花芽數為栽培於 29 ℃者之兩倍 (Park et al., 1998);栽培溫度由 16 ℃升至 26 ℃,金雞菊 (Coreopsis grandiflora)、
西洋濱菊 (Leucanthemum ×superbum) 與金光菊 (Rudbeckia fulgida)之花數分別下 降80%、75%與 55% (Yuan et al., 1998);三色菫之花數隨栽培溫度由 20 ℃升至 30 ℃而減少 (Warner and Erwin, 2006);風鈴草 (Campanula carpatica Jacq.) 則於可 見花苞出現後,花朵數目會隨栽培溫度提高而減少 (Niu et al., 2001)。
提高溫度由15 ℃至 25 ℃,會使三色菫之花徑變小 (Niu et al., 2000; Warner and Erwin, 2006),而高溫 31 ℃亦會使三色菫之花朵面積變小,呈負相關 (r2 = 0.72),
隨高溫之處理時間與溫度升高之強度增強,花朵直徑下降越顯著,可能因為營養 生長時期變短,導致細胞分裂 (cell division) 與細胞伸長 (cell expansion) 減少,
且可見花苞出現後,花朵直徑受高溫之影響更劇 (Pearson et al., 1995)。風鈴草亦 有相同之情形,可見花苞出現後,花徑更易受高溫影響而顯著下降 (Niu et al., 2001)。栽培於 35/30 ℃之非洲鳳仙花之花徑會較栽培於 30/24 ℃與 24/18 ℃者小,
可能受植物賀爾蒙與水分潛勢所影響,且高溫會造成光合作用同化產物無法分配 至花芽 (Lee et al., 1990)。當溫度由 21 升至 25 ℃時,菊花耐熱品種 ‘Iridon’ 與
‘Dark Broze Charm’ 之花徑會減少 4%,而不耐熱品種 ‘Yellow Mandalay’ 與 ‘Coral Charm’ 之花徑則會下降 9%,因此可利用高溫造成花徑變小作為菊花之耐熱指標 (Willits and Bailey, 2000)。高溫導致花卉類作物花朵直徑尚出現於金雞菊、西洋濱 菊與金光菊 (Yuan et al., 1998);金盞花、非洲鳳仙花、龍頭花與夏菫 (Torenia
fournieri Linden ex E. Fourn) (Warner and Erwin, 2005)、月見草 (Clough et al., 2001)
等。
高溫會影響花卉類作物營養生長與生殖生長之表現,可利用於花壇植物品種 間耐熱程度之比較,耐熱之三色菫栽培於高溫時,花會較小但花色較濃、分枝數 多、枝梢乾重較重,且根/枝梢比例低 (Warner and Erwin, 2006);較耐熱之非洲鳳
仙花栽培於35/30℃時,植株覆蓋範圍會較大、枝梢乾重較高,且花徑較不易因高
溫而變小,可作為非洲鳳仙花之耐熱指標 (Lee et al., 1990)。
林 (2006) 指出,不同栽培溫度會影響矮牽牛之營養生長,隨栽培溫度由 16 ℃ 升高至27 ℃,耐熱品種 ‘Tidal Wave Silver’ 與 ‘Wave Blue’ 之分枝數下降較少,
熱敏感品種 ‘Primetime Carmine’ 則顯著下降;栽培溫度由 16 ℃升高至 27 ℃,耐 熱品種 ‘Tidal Wave Silver’ 之枝梢乾重與根乾重皆會顯著上升,不易因高溫對根系 造成不良影響,但熱敏感品種 ‘Primetime Carmine’ 之根乾重則會顯著下降,由此 可知,對高溫適應性不同之矮牽牛品種於高溫之表現隨其耐熱性而不同,且可利 用分枝數、枝梢乾重與根乾重作為矮牽牛品種間耐熱程度之鑑定。栽培溫度之提 高亦會影響矮牽牛之生殖生長,栽培溫度由16 ℃升高至 33 ℃,不同品種矮牽牛 之花朵直徑皆會降低,因此並不是可靠的耐熱篩選指標。因此將利用高溫造成矮 牽牛分枝數、枝梢乾重與根乾重之影響,來鑑別矮牽牛不同品種之耐熱性
四、 常用之耐熱選拔指標
(一) 花粉選拔花粉篩選具有下列優點:體積小,可同時進行多項性狀鑑定;數量多,可節 省育種空間 (Frankel and Galun, 1977; Heslop-Harrison, 1979);花粉為單倍體,隱性 基 因 與 突 變 基 因 不 易 受 顯 性 基 因 所 影 響 (Murphy and Mulcahy, 1987; Zamir, 1983);篩選速度快 (Ottaviano and Mulcahy, 1986) 、離體培養容易、非破壞性測試、
不需昂貴儀器或是困難技術 (Hormaza and Herrero, 1992; 1996),因此可有效利用於
育種。而花粉篩選之理論基礎在於孢子體和配子體世代之基因表現有重疊,提高 花粉之競爭力,可提高後代之活力,且孢子體和配子體對許多外在因子之反應相 同 (Hormaza and Herrero, 1992; 1996),因此可利用花粉形態、花粉釋放量、花粉活 力與萌芽率等重要特性進行作物耐逆境之選育。而其測定方法又可分為化學染色 法、花粉之萌芽測定可於體內 (in vivo) 或體外 (in vitro) 測定 (Shivanna and Rangaswamy, 1992)。目前將花粉篩選利用於作物對溫度逆境育種之研究已不少,
例如利用於耐寒育種之作物有小藜豆 (Cicer arietinum L.) (Clarke et al., 2004) 與番 茄 (Lycopersicon esculentum Mill.) (Domíınguez et al., 2005),而在耐熱育種方面,
則已利用於玉米 (Zea mays Linn.) (Frova et al., 1995; Petolino et al., 1990)。
不同物種之花粉進行體外萌芽測定時,適合其萌芽之培養基所需添加之成分 會不同,而大部分之物種之花粉於添加硼酸 (H3BO3) 與蔗糖之培養基中即可萌芽 (Shivanna and Rangaswamy, 1992);而有少數物種之花粉置於蒸餾水中即可萌芽,
如菸草 (Nicotinana tabacun L.) 與山茶 (Thea sinensis L.),萌芽率與置於添加有蔗 糖者相同 (Loo and Hwang, 1944)。
於 花 粉 萌 芽 培 養 基 中 添 加 蔗 糖 , 可 作 為 滲 透 壓 調 節 物 質 (Adhikari and Campbell, 1998; Martin, 1913; O’Kelly, 1955),當培養基之滲透壓低於花粉之滲透 壓,培養基中之去離子水會進入花粉粒,當花粉粒之細胞壁無法承受此多餘之壓 力時,會導致花粉管爆裂 (bursting),而當培養基中之蔗糖濃度過高或過低時,皆 會因滲透壓而導致花粉管爆裂 (Adhikari and Campbell, 1998; Brink, 1924);蔗糖亦 可作為花粉管於體外萌芽時之養分提供,花粉管萌芽時,會進行自我催化作用 (autocatalytic reaction),因此必須消耗蔗糖作為碳源之提供 (Brink, 1924),但若於 培養基中添加大量存在於植物體中之碳水化合物,如果糖,則會抑制花粉管生長。
O’Kelly (1955) 利用 C14追蹤技術,指出花粉管萌芽時,會吸收蔗糖利用於呼吸作 用,產生大量之二氧化碳釋放至培養基中。培養基中添加蔗糖是某些作物花粉萌 芽之必要條件 (Shivanna and Rangaswamy, 1992),不同物種所需之蔗糖濃度未必相
同 (Shivanna and Rangaswamy, 1992),適合添加 5%蔗糖者有彩葉芋 (Caladium
×hortulanum Birdsey) (Deng and Harbaugh, 2004);適合添加 10%蔗糖者有袋鼠爪花 (Anigozanthos manglesii) (Sukhvibul and Considine, 1993) 與 帝 王 花 (Protea magnifica) (van der Walt and Littlejohn, 1996);適合添加 15%蔗糖者有山龍眼科植物
(Banksia menziesii) (Maguire and Sedgley, 1997)、飛燕草屬植物 (Delphinium) (Honda et al., 2002)、與美國石竹 (Dianthus barbatus L.) (彭,2007);適合添加 20%蔗糖者 則有大白菜 (Brassica rapa L. ssp. Pekinensis) (Sato et al., 1998)、觀賞鳳梨科植物 (Bromeliaceae) (Parton et al., 2002)、大花山茱萸 (Cornus florida L.) (Craddock et al., 2000)、巴陵石竹 (D. palinensis S.S. Ying) (彭,2007) 與玫瑰 (Rosa hybrida) (Voyiatzi, 1995)。不同品種之花粉所適合添加之蔗糖濃度亦不同,玫瑰花 ‘Ferry Porche’、‘ Ferry Porche’、‘ Queen Elizabeth’ 與 ‘John FKennedy’ 品種之花粉於添 加15%蔗糖者萌芽率最好,而玫瑰花 ‘ Lady X ’ 之花粉則以添加 20%蔗糖者最適 合其萌芽 (Voyiatzi, 1995),因此在進行溫度逆境對花粉萌芽之影響時,須先尋找 花粉萌芽所適合之蔗糖濃度,以利之後選拔。
不同物種間之基因差異會影響其花粉萌芽之最適溫度 (Luza and Polito, 1985),原生於北溫帶 (North temperate zone) 至北寒帶 (North frigid zone) 地區之 飛燕草屬植物,其花粉培養於15-20 ℃間萌芽率最高,平均萌芽率可達 58%,但當 培養溫度為30 ℃時,萌芽率則降至 15% (Honda et al., 2002);而原生於熱帶亞熱帶 台灣之鵝鑾鼻燈籠草 (Kalanchoe garambiensis Kudo) 及其紫葉變種,花粉則於 20-25 ℃萌芽率最好,當培養溫度升至 30 ℃,花粉萌芽率仍可達 27% (侯,2003)。
同屬不同種植物之花粉對溫度之適應性可能不同 (Stern, 1998),原生台灣之鵝 鸞鼻燈籠草及其紫葉變種之花粉於 35 ℃水浴中培養 2 小時後,萌芽率可維持在 20%左右,而由丹麥 Kund Jepsen a/s 公司所育出之長壽花 (Kalanchoe blossfeldiana Poelln. ‘Isabella’) 於 35 ℃培養者之萌芽率僅剩 3% (侯,2003);當水浴培養溫度大
於35 ℃時,由日本福花園種苗公司所生產耐熱性差之美國石竹花粉已無法正常萌
芽,但耐熱性佳且於台灣可周年開花之巴陵石竹與瞿麥 (Dianthus superbus L. var.
superbus (Masamune) Lin & Ying) 在水浴溫度為 40 ℃時仍具有萌芽能力 (彭,
2007)。
不 同 耐 熱 程 度 的 品 種 , 其 花 粉 萌 芽 對 溫 度 的 反 應 會 有 差 異 , 耐 熱 辣 椒 (Capsicum annuum L.) 品種花粉之最適萌芽溫度較高,因此可藉此作為辣椒屬植株 耐熱性之鑑別標準 (Reddy and Kakani, 2007);玉米耐熱品系 ‘A641’ 之花粉經高溫 35 ℃短暫處理 3-5 分鐘後,仍具萌芽能力且花粉管生長正常,但熱敏感品 ‘MK386’
與teosinte 經 35 ℃處理 3 分鐘後,萌芽率即顯著下降,且花粉管長度變短 (Lyakh et al., 1991)。
品種特性與栽培溫度亦會影響花粉之萌芽,番茄熱敏感品系 ‘NC 8288'栽植 於32/26℃時,因高溫栽培導致絨氈層 (tapetum) 提早崩解,無法繼續供應養分至 花粉發育,花粉呈不規則形,因此無法撐開花藥囊正常釋放花粉,導致花粉釋放 量下降,且花粉萌芽率亦下降 (Sato and Peet, 2005)。草莓 (Fragaria ×ananassa Duch.) 耐熱品種 ‘Nyoho’ 栽培於 23/18 ℃和 30/25 ℃,利用體內培養觀察花粉管 於柱頭內之生長,皆可觀察到花粉粒於柱頭上萌芽、穿過花粉管並抵達胚珠;而 不耐熱品種 ‘Toyonoka’ 栽培於 30/25 ℃之花粉無法在花柱內正常生長、亦無法到 達胚珠受精成功 (Ledesma and Sugiyama, 2005)。花生 (Arachis hypogaea L.) 栽培 於28 ℃下之花粉萌芽率可達 75%,但若於開花前 6 天給予 39 ℃之高溫,花粉萌 芽率有顯著下降之情形,僅剩20%-40%,尤以開花前 4 天與開花當天給予高溫所 造成之萌芽率降低最顯著;栽培高溫除造成花粉萌芽率下降,亦會使結果率下降,
栽培於 28 ℃之結果率平均 60%,但栽培於 39 ℃之高溫會使結果率降至 20%
(Prasad et al., 2001)。顯示在花粉培養溫度與植株栽培溫度提高下,耐熱品種之花 粉除能正常表現,且於高溫下具授粉優勢,結種率可提高,综上所述,花粉之耐 熱特性可供為高溫適應性之篩選指標。
(二) 細胞膜熱穩定性
高溫逆境會植物破壞蛋白質穩定性,並使活性氧族 (activated oxygen species, AOS) 生 成 , 進 而 破 壞 細 胞 膜 之 半 透 性 (Basaga, 1989) , 使 細 胞 膜 之 流 動 性 (membrane fluidity) 與 滲 透 性 (permeability) 會 改 變 (Ismail and Hall, 1999;
Sangwan et al., 2002),因細胞膜由雙層磷脂質、蛋白質與醣類構成,遇高溫時,雙 層磷脂質之鍵結會改變,因此細胞膜之滲透性會增加,進而導致電解質之滲漏,
因此可藉測定葉片經短暫高溫處理後之電解質滲漏率或相對熱傷害值 (relative injury, RI) 來檢測植物之細胞膜熱穩定性 (cell membrane thermostability, CMT),以 提供快速且有效之作物耐熱性鑑定方法。細胞膜熱穩定性為植物遇高溫逆境時最 敏感之反應之ㄧ (Marcum et al., 1998; Raison et al., 1980),利用細胞膜熱穩定性檢 測植物之耐熱性,具檢測快速、便宜、敏感、空間需求小,可有效反應植株之耐 熱性且可同時檢測大量物種或品種 (Marcum et al., 1998)。
利用高溫會造成細胞膜滲透性改變,造成電解質滲漏之原理,將葉片置於高 溫水浴中處理,測定高溫處理後溶液之電導度 (Ti),並測定葉片經高溫高壓完全 破壞後之電導度 (Tf),代入下列公式後即可得相對熱傷害值 (Martineau et al., 1979;
Yeh and Lin, 2003)。
RI (%) = Ti / Tf × 100
為避免每次測量環境不同所造成之變因,可利用下列公式經室溫25 ℃水浴後
所得之電導度 (Ci),即其經完全破壞後所得之電導度 (Cf) 做校正 (Martineau et al., 1979; Yeh and Lin, 2003)。
RI (%) = { 1 - [ ( 1 – Ti / Tf ) / ( 1 – Ci / Cf ) ] }
相對傷害值會受高溫水浴溫度上升或高溫水浴時間增加而呈 S 形曲線上升,
因此可藉達到相對熱傷害值 50%所需之溫度及時間作為評估細胞膜熱穩定性之方
法,分別為熱致死溫度 (heat-killing time, Temp50) 及熱致死時間 (heat-killing time, Time50),利用熱致死溫度與熱致死時間可將細胞膜熱穩定性量化 (Marcum et al.,
1998)。
不同物種間之細胞膜對高溫之適應性而不同,豆科植物中,較耐熱之落花生 (Arachis hypogaea L.) 以 54 ℃為其熱致死溫度,熱敏感之小黎豆 (Cicer arietinum L.) 則於 44.3 ℃即達熱致死之溫度;而不同品種間對高溫之適應性亦會影響其細 胞膜熱穩定性,耐熱性佳之維吉尼亞系統落花生經45 ℃水浴加熱 15 分鐘後,相 對熱傷害值約為50%,而西班牙系統熱敏感之落花生經高溫 45 ℃水浴加熱 15 分 鐘後之相對熱傷害值則約 70%;小黎豆原生於南印度之 ‘Annigeri’ 品種與原生於 敘利亞之 ‘ILC 482’ 品種較耐熱,經 45 ℃水浴加熱 15 分鐘後,相對熱傷害值約 45%,而原生於中印度之 ‘K 850’ 品種之相對熱傷害值則高達 60% (Srinivasan et al, 1996)。
植物除具有其原有之耐熱性 (basal thermotolerance) 可供其渡過高溫逆境,亦 也 可 藉 由 逐 漸 提 高 栽 培 溫 度 , 但 不 可 超 過 其 致 死 溫 度 之 溫 度 以 誘 導 耐 熱 性 (acquired thermotolerance),所利用之機制有產生熱休克蛋白 (heat shock protein, HSP) (Vierling, 1991) 或使蛋白質變性 (denature) (Lee and Vierling, 2000),但此熱 馴化之溫度範圍較小,大於植物所能接受的熱馴化溫度,將不會再提高其耐熱性,
反而將造成傷害 (Chen et al., 1982; Li et al., 1991)。
菜豆 (Phaseolus vulgaris L.)耐熱品種‘BBL-415-1’與熱敏感品種 ‘BBL-47’ 栽 培於20/15 ℃時,經 50 ℃水浴處理葉圓片,熱致死時間皆為 5 分鐘,但若栽培於 40/30 ℃,耐熱品種 ‘BBL-415-1’ 之熱致死時間則顯著增加為 85 分鐘,而熱敏感 品種則僅增加至50 分鐘 (Chen et al., 1982),顯示不同品種間可誘導出之耐熱性依 其本身對高溫之適應性而不同。珍珠栗 (Pennisetum glaucum (L.) R. Br.) 熱敏感品 種 ‘ICMV 155’ 實生小苗未經 40 ℃熱馴化兩小時,熱致死溫度為 47 ℃,但若經 馴化後之熱致死溫度仍未提高;而耐熱品種 ‘HHB 67’ 與 ‘IP 3201’ 經高溫 43 ℃ 熱馴化兩小時後,熱致死溫度則顯著上升,由未經馴化者之47 ℃上升至 52 ℃;
高粱 (Sorghum bicolor L. Moench) 亦有相同之情形,耐熱品種 ‘SPV 386’ 經高溫
43 ℃馴化兩小時後,熱致死溫度上升 4 ℃ (Howarth et al., 1997),顯示耐熱品種經 較高溫之馴化溫度,才可達到熱誘導之效果。
大豆 (Glycine max Merr.)、番茄 (Lycopersicon esculentum Mill.)、菜豆與馬鈴 薯 (Solanum tuberosum L.) 之細胞膜熱穩定性亦可利用栽培之高溫所誘導 (Chen et al., 1982);糠穗草 (Agrostis palustris Huds.) 經高溫 40 ℃誘導 14 天後,其相對 熱傷害值為未經誘導者減少33%,若於 40 ℃誘導 28 天後,相對熱傷害值則為未
經誘導者減少 57%,顯示康穗草經高溫栽培誘導,其細胞膜耐熱性提高,且經高
溫馴化誘導耐熱性者,會有57 kDa 與 54 kDA 之熱休克蛋白產生,未經馴化者則 否 (He et al., 2005),顯示經高溫馴化後可產生熱休克蛋白以提高其耐熱性。
細胞膜之耐熱性除可利用栽培溫度誘導,亦可直接從葉圓片誘導出,辣椒葉 圓片經38 ℃水浴加熱 4 小時後,熱致死時間為 34.7 分鐘,但葉圓片若不經高溫水 浴,經8.4 分鐘後即可達到熱致死溫度 (Anderson et al., 1990)。番茄、大豆與馬鈴 薯之葉圓片經水浴 35-40 ℃熱馴化後,亦可提高其細胞膜之穩定性 (Chen et al., 1982);而甜瓜 (Cucumis melo L.) 之葉片經 35 ℃加熱 2-4 小時後,即可誘導其耐 熱性 (Lester, 1986)。
栽培於22/20 ℃之辣椒經不同水浴溫度 15 分鐘後,熱致死溫度為 51.1 ℃,但 若將辣椒置於38/30 ℃環境 24 小時後,熱致死溫度可升至 54.1 ℃,顯示辣椒可經 高溫栽培誘導,而提高細胞膜熱穩定性,但若再將辣椒放置回22/20 ℃兩天後,熱 致 死 溫 度 則 會 降 至 52 ℃ , 顯 示 原 本 經 誘 導 出 之 耐 熱 性 , 也 可 能 會 逆 馴 化 (deacclimation),顯示需要持續的高溫以持續維持所誘導出之耐熱性 (Anderson et al., 1990; Chen et al., 1982)。
Li (1991) 指出,菜豆經熱馴化所誘導出之耐熱性較其原所具有之耐熱性重 要,耐熱品種 ‘GO 3689’ 未經馴化時,經 50 ℃水浴加熱葉圓片 15 分鐘後,會達 50%相對熱傷害值,熱敏感品種 ‘BBL 47’ 則於 10 分鐘時就已達到 50%熱傷害值,
胞膜熱穩定性較耐熱品種高,但若將三品種經37 ℃熱馴化 24 小時後,耐熱品種
‘GO 3689’ 經後天誘導之耐熱性顯著提高,其熱致死時間增加為 130 分鐘,耐熱性 中等之 ‘G 6-6’ 則為 92 分鐘,顯示馴化所誘導出之耐熱性為大豆耐熱育種時較佳 之選拔指標,而非品種原具有之耐熱性 (Li et al., 1991)。
細胞膜熱穩定性目前已廣泛運用於農藝作物、草坪與蔬菜,於花卉作物之應 用尚少。耐熱性較佳之長春花 (Catharanthus roseus (L.) G. Don) 與不耐熱之香豌 豆 (Lathyrus odoratus L.) 葉片於不同水浴溫度處理 15 分鐘後之電解質滲漏率亦隨 水浴溫度提高而呈S 形曲線,香豌豆之滲漏率於 47-48 ℃開始急速上升,而長春花 則於50 ℃才開始上升,且兩物種電解質滲漏率上升之溫度與其抗氧化酵素 Catalase 和Ascorbate peroxidase 活性開始下降之溫度相似 (Anderson and Padhye, 2004);當 水浴溫度為50℃時,菊花耐熱品種之相對熱傷害值介於 32-35%,而不耐熱品種之 傷害值則為65% - 68%,顯著高於耐熱品種,且菊花之相對熱傷害值與夏季定植之 開花熱延遲天數有高度正相關 (Yeh and Lin, 2003);利用不同水浴溫度亦可區分菊 花不同品種之耐熱性,夏菊以高溫水浴50 ℃加熱 30 分鐘可有效作為品種間之耐 熱指標,而秋冬菊則以45 ℃水浴溫度為良好之耐熱鑑別溫度 (王,2005)。常春藤 (Hedera helix L.) 利用於水浴溫度為 50 ℃時為 RI 值形曲線之轉折點,因此可利用 50 ℃水浴加熱 30 分鐘鑑別不同品種對高溫之敏感度 (Yeh and Hsu, 2004)。栽培於 35/30 ℃之非洲菊品種 ‘Monte Crysto’、‘Rosella’、‘Lisa’ 及 ‘Rosalin’ 葉片經高溫 水浴加熱15 分鐘後,熱致死溫度分別為 54.1 ℃、53.2 ℃、53.1 ℃與 52.2 ℃,因 此可以 53 ℃水浴區別品種間細胞膜熱穩定性之差異,且 ‘Lisa’ 品種栽培於高溫 35/30 ℃相對熱傷害值之 S 形曲線,會較栽培於 25/20 ℃者右移,顯示 ‘Lisa’ 品種 可被熱馴化之能力較高 (陳,2007)。
林 (2006) 亦 於 不 同 之 栽 培 溫 度 測 定 矮 牽 牛 不 同 品 種 之 相 對 熱 傷 害 值 (relative injury, RI),隨栽培溫度由 16 ℃升高至 33 ℃,矮牽牛之葉圓片經 50 ℃水 浴 20 分鐘後,相對熱傷害值皆會下降,其中以耐熱品種 ‘Tidal Wave Silver’ 與
‘Wave Blue’ 之下降最多,且栽培於 27 ℃下之相對熱傷害值隨品種之耐熱程度而 不同,耐熱品種 ‘Wave’ 與 ‘Tidal Wave Silver’ 之相對熱傷害值較低,分別為 9.7%
與19.7%,熱敏感品種 ‘Primetime Carmine’ 之相對熱傷害值則高達 42.4%,顯示 耐熱品種於27 ℃可誘導其耐熱性,作為耐熱指標,但林 (2006) 並無利用不同水
浴溫度測試矮牽牛於不同栽培溫度下之相對熱傷害值,建立 S 形曲線之轉折點,
因此若欲利用單一溫度區別品種間之差異,並確定矮牽牛不同品種之熱馴化程 度,仍需進行進一步之試驗。
五、 細胞膜熱穩定性之遺傳
進行耐熱育種時,如能暸解耐熱性狀之遺傳行為,可有效應用於選拔,且效 率 較 僅 由 外 表 型 態 作 為 判 斷 者 高 且 可 靠 (Ibrahim and Quick, 2001a) 。 菜 豆 (Phaseolus vulgaris L.) 耐熱程度不同之品種經雜交所得之後代經 37 ℃熱馴化 24 小時後測定其相對熱傷害值,結果主由多基因控制其耐熱表現,且環境對耐熱表 現基因之影響大 (Marsh et al., 1985)。
數量性狀 (quantitative character) 之遺傳表現除受基因所控制,亦受環境因子 之影響,因此進行選拔時需考慮耐熱性狀之遺傳力,即其性狀可遺傳至後代之能 力 。 小 麥 (Triticum aestivum L.) 細胞膜熱穩定性之廣義遺傳力 (broad-sense heritability) 高 (Blum et al., 2001),顯示小麥之細胞膜熱穩定性易遺傳至子代,且 較不易受環境因子之影響。
自交作物利用雜種優勢進行育種時,會依據自交系之性狀表現,即其組合力 (combining ability) 作為挑選育種親本之標準,以選出最優良之育種親本,而組合 力又可分為一般組合力 (general combining ability, GCA) 及特殊組合力 (specific combining ability, SCA)。若干自交系進行雜交時,當某一自交系與其他自交系雜交
所得 F1之組合力皆高,顯示此為一般組合力高之自交系;若某一自交系與另一雜
交組合之組合力高,顯示此組合為具高特殊組合力之組合。控制一般組合力之基
因為加性基因 (additive gene),而特殊組合力則由顯性基因 (dominant gene) 所影 響,因此若能暸解耐熱性狀受一般組合力或特殊組合力所影響,將能有效應用於 耐 熱 親 本 選 擇 。 小 麥 之 細 胞 膜 熱 穩 定 性 與 其 一 般 組 合 力 經 相 關 係 數 分 析 (correlation coefficient analysis),兩者間具顯著正相關,顯示小麥經熱馴化所誘導 出之細胞膜熱穩定性由加性基因所控制,利用具於細胞膜熱穩定性具高一般組合 力之親本 ‘WH 730’ 及 ‘Raj 3765’ 進行雜交,其後雜交後代亦具有較高之細胞膜 熱穩定性,但如親本之一般組合力為負值者,後代之細胞膜熱穩定性表現則會較 差,顯示細胞膜熱穩定性可遺傳至子代,且可利用具高一般組合力之品種作為優 良耐熱育種親本 (Dhanda and Munjal, 2006)。
Ibrahim and Quick (2001b) 亦認為小麥經高溫 39 ℃熱馴化 48 小時所誘導出之 細胞膜熱穩定性遺傳是由加性基因所控制,由具高一般組合力之品種 ‘Kauz’ 為種 子親時,其後代之細胞膜熱穩定性較高,但若由一般組合力低之熱敏感品種 ‘Arlin’
為種子親者則相反;小麥膜熱穩定性之遺傳主要受母本效應 (maternal effect) 之影
響,在全互交試驗中,種子親之影響佔 67%,即其後代之耐熱表現主受種子親細
胞核基因型所影響。但豇豆 (Vigna unguiculata (L.)Walp.) 細胞膜熱穩定性之遺傳 則不是由種子親所影響 (Thiaw and Hall, 2004)。
小麥利用TTC 還原法 (2,3,5-Triphenyl tetrazolium chloride)和葉綠素螢光值評 估其耐熱性,結果顯示耐熱性之遺傳主要受種子親之影響且廣義遺傳力高,因此 小麥之耐熱性由加性基因所控制,且可利用一般組合力高之品種作為雜交親本,
以選拔並增加後代之耐熱性(Fokar et al., 1998; Moffatt et al., 1990; Porter et al., 1995)。
小麥耐熱品種進行雜交,雜交組合為 ‘WH 533’ בWH 730’ 者之細胞膜熱穩定 性、細胞活力與產量於高溫逆境下表現皆較佳,但如為 ‘WH 533’ בRaj 3765’ 或
‘WH 533’ בHUW 234’ 之雜交組合,耐熱表現並不如 ‘WH 533’ בWH 730’,顯示 小麥耐熱品種 ‘WH 533’ 與 ‘WH 730’ 兩者之基因於耐熱表現上較相似,因此後