國立臺灣大學生命科學院漁業科學研究所 碩士論文
Graduate Institute of Fisheries Science College of Live Science
National Taiwan University Master Thesis
飼料中添加不同濃度與來源之類胡蘿蔔素對珍珠鱗 金魚增豔效果之影響
Effects of Dietary Different Carotenoids Supplements on Pigmentation on the pearlscale goldfish ,
Carassius auratus
張嘉育 Chia-Yu Chang
指導教授:廖文亮 博士 Advisor: Wen-Liang Liao, Ph.D.
中華民國 102 年 7 月
July, 2012
I
誌謝
本論文得以順利完成,首先得感謝恩師 廖文亮博士。廖老師總是充滿仁慈 並無私的指導每位學生,無論在學業、工作或是為人處事上,廖老師的指導與恩 情學生將永記在心。在論文審查期間承蒙 丁雲源所長、葉信利主任以及張繼堯 老師在百忙之中抽空前來審閱與指教論文的缺失,讓學生得以更正,使本論文得 更趨完善。另外感謝實驗室的夥伴和其他實驗室的朋友們,相輔相成、亦師亦友,
也謝謝學弟妹在實驗期間的幫忙。感謝在就學期間提供實習機會給我的台灣微軟 公司,RSM 部門、Xbox 戰隊、Grace 媽以及提攜人 Jack,謝謝你們無論在工作 或是生活學業上為我加油打氣,2 年的研究所生涯因為有你們讓我充滿幹勁。感 謝我的好姊妹-台客幫的每個人,這麼多年的友誼,感謝你們的相伴,謝謝你們 總是聽我發牢騷、讓我解悶,使我有勇氣有動力再衝下去。謝謝威廷這 7 年來的 陪伴,你的愛護和疼惜我真的很感謝,接下來的日子還是要請你多多指教。
最後能成功畢業,絕對要感謝的是我的家人,謝謝我的媽媽含辛茹苦,每個 月都很準時的給我生活費,讓我得以專心在功課上,也謝謝媽媽給我的一切。謝 謝我的外婆、外公、阿姨們、兄弟姊妹們,你們是我歡樂的來源,也謝謝你們所 給的建議和人生的經歷。謝謝我的奶奶、爺爺、爸爸、姑姑們,謝謝你們的支持 和照顧,沒齒難忘。
II
摘要
珍珠鱗金魚 (以下簡稱珠鱗 ),英文俗名 pearlscale goldfish 為金魚的一種,
只需保持水體清潔即可飼養,因此在觀賞魚市場具有相當高的經濟價值 。
本研究主要探討在飼料中添加不同種類及濃度的類胡蘿蔔素對於珍珠鱗金 魚的呈色和色素代謝之影響。在飼料中採用兩種不同的類胡蘿蔔素:玉米黃素 (Zeaxanthin,以下簡稱 Z),合成蝦紅素(Synthesis Astaxanthin,以下簡稱 S),其 添加在飼料中的濃度與種類分別為 0(control)、Z-5、Z-10、Z-15、Z-20、S-5、S-10、
S-15、S-20(mg/kg diet)共計九組。試驗用魚種珍珠鱗金魚,馴養四星期,馴養期 間投餵不含類胡蘿蔔素的飼料,以免在馴養期間攝入類胡蘿蔔素而影響實驗,並 於分組試驗前一天絕食。隨機取樣魚體平均體重 2.69±0.23g 的每組 8 尾珍珠鱗金 魚 , 每 組 固 定 有 一 隻 白 色 體 色 的 珠 鱗 為 盲 點 對 照 組 , 飼 養 於 長 * 寬 * 高 為 45*30*30cm 配有上部過濾的水缸共計 9 組。
經 7 星期飼養珍珠鱗金魚後各組間的活存率及平均增重 0.94g 於成長上無顯 著差異。結果顯示呈色添加合成蝦紅素的組別與添加玉米黃素的組別有顯著差異,
隨著合成蝦紅素的濃度升高而有增高的趨勢。合成蝦紅素的添加影響珍珠鱗金魚 體色的呈色效果較玉米黃素為佳,並以 15mg/kg 為最適添加量。
III
Abstract
This study focused on the different kinds and concentrations of carotenoid in diet, for the effects of pigmentation of pearlscale goldfish (Carassius auratus), assessment of pigment metabolism, and raise fish pigmentation in the suitable amount.
Experimental feeds are supplemented with two different carotenoid: zeaxanthin (hereinafter referred to as Z), synthesis astaxanthin (hereinafter referred as S).
Experiment period was total 7 weeks, mean weight of pearlscale goldfish was about 2.69g. Experimental diets were divided into 9 groups: control, Z5, Z10, Z15, Z20, S5, S10, S15 and S20 (mg/kg diet). Fish skin were tested with tristimulus colorimeter, and further quantified for integument pigment.
Results shows that the addition of carotenoids have no effects on growth and feed efficiency, but improve the pigmentation by enhanced a and b values in pearlscale goldfish. The coloration of both S groups and Z groups were increase by the raising of concentrations of carotenoid in diet. Therefore, we can say that synthesis astaxanthin is more effective than zeaxanthin in the addition of carotenoids in diets for pearlscale goldfish, and the optimal concentration is 15mg/kg diet.
KEY WORDS : pearlscale goldfish, growth performance, integument pigmentation, carotenoid, dietary supplements
IV
目錄
頁次
誌謝……….. I
中文摘要……….. II
英文摘要……….. III
目錄………... IV
第一章 前言………. 1
第二章 材料與方法………. 7
第三章 結果………. 11
第四章 討論………. 17
第五章 結論………. 24
參考文獻………... 25
表與圖………... 31
1
第一章 前言
珍珠鱗金魚(Carassius auratus auratus),俗名 pearlscale goldfish、珠鱗,是一 種原產於中國的觀賞魚類,是野生鯽魚演化而來的彩色變種-金魚的變種。和觀 賞性的金魚和食用的野生鯽魚,學名同為 Carassius auratus。在分類上屬脊索動 物 門 (Chordata) 、 輻 鰭 魚 綱 (Actinopterygii) 、 鯉 形 目 (Cypriniformes) 、 鯉 科 (Cyprinidae)、鯽屬(Carassius)、鯽魚種(C.auratus)、金魚亞種(C.a.auratus)。
原產於亞洲中部,中國和日本,由於它是受歡迎的池塘和水族館觀賞魚,已 被引入全世界各地。金魚生活在河流,湖泊,池塘,潟湖和水溝等水溫冷、水流 慢且有水生植物的水域,能夠承受長期暴露在鹽度高於 15 ppt,並能容忍低溶氧 量。金魚是一種小型到中型的魚,身體縱高,橫截面圓形,頭部和眼睛很大,嘴 小,尾巴分叉;鱗大,單一背鰭,前面有 3~4 根粗鰭,背面顏色從橄欖銅色到深 金色,腹面淡色到銀白色,可以長到 41 厘米長,2 公斤重,圈養環境下可活 30 年;攝食各種水生植物(包括藻類),有機碎屑,甲殼動物,蠕蟲,昆蟲和小蝸牛,
在底部吸食的習性會攪動養分供給於藻類使用,有助於藻類大量繁殖 (Morgan &
Beatty, 2004),以及提高水濁度和增生耗氧的水生植物(Richardson et al, 1995) 。 在淺海雜草之間產卵,每次可產下多達幾十萬顆小型的卵(直徑 1 ~ 2 mm),一隻 魚可產卵 3~10 批,產卵間隔 8~10 天,利用選擇性繁殖,許多具有各種各樣的 色彩和鰭不同形狀的金魚品種被生產出來,這些魚如果被釋放到野外,通常會恢 復到橄欖青銅色和正常形狀的鰭(FishBase, 2004; McDowall, 2000)。
魚的體色因應不同的生長階段與環境改變,其表現受到多重因子影響,包含 色素細胞的型態與數量以及環境的變化。位於為真皮層中的色素細胞是構成體色 的基本單元,一般包含三種細胞,第一種為黃色素細胞(xanthophores),包含蝶啶 (pteridine)、類胡蘿蔔素(carotenoid)色素,第二種為黑色素細胞(malanophores),
包含了黑色素(melanin),第三種為彩虹色素細胞(iridophore),由結構變化影響體 色。其中類胡蘿蔔素與黑色素,是魚類中比較常研究的二種色素,而魚體呈現的 各種紅色、橙色、黃色則會隨著與黃色素結合類胡蘿蔔素種類與數量所影響(Price
2
et al., 2008)。除此之外,魚體可經由感知環境後,由神經及內分泌系統操控色素 細胞內色素的聚散達到快速而短期的體色變化;另外,透過色素細胞的種類更替 消長與色素細胞內的色素蓄積與代謝之變化,則較為緩慢而長期(Fujii, 2000)。
體色對魚類具有多樣的生物功能,其中在行為上比較廣泛被認識的擇偶選擇 (sexual selection),是藉由個體較醒目的體色爭取更多與異性交配機會的種內競爭 (Berglund et al., 1996),除此之外,其它行為上功能的還包括物種的辦識(species recognition)、警戒色彩(warning colouration)、侵略展示(aggressive displays)以及偽 裝作用(camouflage) (Svensson and Wong, 2011)。
目前在自然界中己經被發現 750 種以上的類胡蘿蔔素。類胡蘿蔔素為廣泛存 在自然界中的脂溶性化合物,可由植物、藻類或微生物合成雖然發現動物身上也 被發現到存在類胡蘿蔔素,但由於動物無法自身合成,僅能由食物中攝取,或是 經代謝轉換攝取到的類胡蘿蔔素(Goodwin, 1986)。
依結構大致可分成二大類,一為結構中不含氧,僅由碳與氫原子所構成的胡 蘿蔔素(carotenes),例如: β-胡蘿蔔素(β-carotene)、蕃茄紅素(lycopene);另一類則 為結構中含氧原子的葉黃素(xanthophylls),例如:玉米黃素(zeaxanthin)、蝦紅素 (astaxanthin)。
由於類胡蘿蔔素含有的多個共軛雙鍵結構,容易吸收波長 400-500 nm 的光,
呈現出各種黃色、橙色或紅色(Bjørnland, 1997)。而也因為其結構穩定性不高,
容易受到氧、光、熱和酸的影響產生異構化的現象。
類胡蘿蔔素在生物體中能與脂質、蛋白質結合,或單獨的以游離的型式存在。
在皮膚上的類胡蘿蔔素經常與脂肪酸結合而以酯化的型式存在。其中蝦紅素 (astaxanthin)為水生生物中最常見的類胡蘿蔔素,在魚類經常以此種方式在魚皮 蓄積蝦紅素,如嘉鱲(Pagrus pagrus);及在魚肉上蓄積,如鮭鱒科的魚類(salmonid)。
此外,類胡蘿蔔素也可能與蛋白質結合,形成類胡蘿蔔素蛋白(carotenoproteins)。
甲殼類生物體內的蝦紅素常與外骨骼中的蛋白質結合形成類胡蘿蔔素蛋白,因色 素分子結構與蛋白質結合後使得色素穩定性增加,又因結合的蛋白質不同,造成 甲殼類呈現出如紫色、藍色甚至綠色等不同的顏色(Zagalsky et al., 1970),所以當
3
經過熱處理、使類胡蘿蔔素蛋白變性後,甲殼類會顯現出原始蝦紅素的紅色 (Cianci et al., 2002)。除此之外,出現在卵中的類胡蘿蔔素一般則以游離態的方式 聚集,並且溶於脂質當中(Parker, 1996)。
除了生態行為上的意義之外,體色也具有許多生理功能,在影響體色的類胡 蘿蔔素中也有愈來愈多的作用被發現。例如在鬥魚(Betta splendens)的藍、紅二個 品系的研究中發現,類胡蘿蔔素在紅色雄魚身上,可以增強紅色程度來吸引雌魚 增加繁殖的機會,而在藍色雄魚身上,類胡蘿蔔的含量則與呈現較高的免疫能力 具較高的相關性(Clotfelter et al., 2007)。在孔雀魚(Poecilia reticulata)的研究也指 出,雄魚攝食較多的類胡蘿蔔素,會提升身上橘色的鮮豔度,並可增加吸引雌魚 的機會(Kodricbrown, 1989),而且也具有比較好的免疫能力來抵抗外部的寄生蟲 (Kolluru et al., 2006);另外剛治癒完寄生蟲疾病的孔雀魚,身上橘色的鮮豔度會 下降,削弱了吸引雌魚的機會(Houde and Torio, 1992)。綜合以上研究,可以了解 魚的體色不僅在生態與生理上皆具有一定的重要性,在不同魚種利用類胡蘿蔔素 的策略的考量上亦有所差異。
類胡蘿蔔素提供卵黃色澤(Blount et al., 2000;Craik,1985),有研究指出在敏感 的胚胎發育時期,需要類胡蘿蔔素的抗氧化功能避免細胞遭受自由基的損害 (Blount et al., 2000; Surai and Speake, 1998),另外於卵中的多元不飽合脂肪酸容 易氧化以及快速的被氧化劑所破壞,但在胚胎發育時期的快速代謝卻會不斷的產 生氧化劑,由於類胡蘿蔔素具有能夠降低氧化壓力的功能,因此顯得相當重要 (Miki, 1991)。
水生動物外表的呈色除了和攝入的色素種類有關之外,也可能經由自身的酵 素系統轉換,代謝成其它種類的色素,也因為各個物種轉換吸蓄積色素的能力不 盡相同,因此呈色結果也有所差異。蝦紅素為多數水生動物蓄積的色素,將類胡 蘿蔔素轉換成蝦紅素之代謝模式分為以下三種(Simpson and Kamata, 1979):
1. 紅鯉型(Red carp type)
可以利用 Lutein、Zeaxanthin 或中間的代謝物轉換成蝦紅素,大部分的淡水魚屬 於此類型。例如:錦鯉(Cyprinus carpio)、金魚(Carassius auratus)
4
2. 鯛魚型(Sea bream type)
不能由其它的類胡蘿蔔素轉換成蝦紅素,但可以直接將色素吸收蓄積在體內,大 部分的海水魚以及鮭鱒科的魚類屬於此類型。
例如:嘉鱲(Pagrus major)、鱒魚(Onchorhychus kisutch)、鮭魚(Salmo gairdoeri) 3. 蝦型(Prawn type)
可以利用 β-carotene、Zeaxanthin 或中間的代謝物轉換成蝦紅素,大部分的甲殼 類屬於此類型。
在人工養殖環境下,往往因為缺乏適當或足夠的類胡蘿蔔素來源,造成水產 動物的呈色不如野生的色彩鮮豔。因此在飼料中添加類胡蘿蔔素來增色。提升水 生動物的色澤與價值,尤其是鮭鱒科的魚類以及觀賞魚,魚體的呈色常常是直接 影響市場價值的一項重要指標也影響消費者選購的意願。
在增色的研究中,在 1966 年開始有將辣椒紅素(paprika)、小龍蝦(crawfish) 的萃取物、金盞花(marigold)的花瓣,添加在養殖虹鱒(Oncorhynchus mykiss)的飼 料中的研究。由於期望能夠提升魚隻色素蓄積的效率,一直以來仍有相當多的研 究探討鮭鱒魚類的增色,促使魚肉能呈現賣相較佳的橘紅色。目前為了使大西洋 鮭魚獲得較佳的呈色,一般會在飼料中添加 30-60mg/kg 的蝦紅素。在鮭魚的養 殖中,添加的色素約在飼料總成本的 10-15%,是飼料中的成分最昂貴的部分 (Buttle et al., 2001)。由此可知,類胡蘿蔔素在水產養殖上的需求量相當可觀。
近 年 來 亦 有 愈 來 愈 多 的 研 究 朝 向 觀 賞 魚 的 增 色 , 包 括 孔 雀 魚 (Poecilia reticulata)(Karino and Haijima, 2004),金魚(Xu, 2006; Yanar et al., 2008)、鬥魚 (Betta splendens)(Clotfelter et al., 2007)、紅旗(Hyphessobrycon callistus)(Wang et al., 2006)、小丑魚(Amphiprion ocellaris)(Yasir and Qin, 2010)、紅魔鬼(Cichlasoma citrinellum)(Pan and Chien, 2009)、金波羅(Cichlasoma severum)(Kop and Durmaz, 2008)。除了魚類之外,亦有研究致力於蝦類(Boonyaratpalin et al., 2001; Liao et al., 1993; Paibulkichakul et al., 2008)及海膽(Psammechinus miliaris)生殖腺(McBride et al., 2004; Shpigel et al., 2006; Suckling et al., 2011)的增色效果。
5
由於各個物種蓄積、轉換色素的能力不盡相同,而在飼料中添加色素將造成 本負擔,因此色素在種類的選擇上與劑量的考量上都是值得研究的問題。隨著類 胡蘿蔔素有愈來愈多的生理功能被發現,除了增豔效果,有研究開始探討類胡蘿 蔔素對於水生生物更多層面的影響,其中包括維他 A 轉換能力(Hu et al., 2006)、
抗氧化(Pan et al., 2011)、免疫能力(Amar et al., 2004; Clotfelter et al., 2007)、抗緊 迫(Pan et al., 2011)。
顏色的測定方法係根據國際照明委員會(Munsell Color Science Lab)依據模 擬人眼感官所能感受的色彩空間 CIE L*a*b*系統。此套系統將顏色定義出顏色 參數(a*,b*和 L*值),a*值越高代表紅色程度越高,而 a*值越低代表綠色程 度越高;b*值越高代表黃色程度越高,而 b*值越低代表藍色程度越高;L*值代 表亮度,其值越高代表亮度越高。其應用方法為,利用色差計(Color difference meter)直接偵測魚體,並可利用色差公式進一步計算顏色差異值(△E)比較色差。
當△E>6 則表示已達人眼感官可明顯分辨的標準。依此系統,前人的研究發現,
天然環境生長的魚隻,其體色與人工養殖的魚隻,其體色有很明顯的差異,其中 天然的嘉鱲(Pagrus major)與養殖的魚隻其△E 可達 13.5,天然的香魚(Plecoglossus altivelis)與養殖的魚隻其△E 可達 13.5,皆達人眼感官可以明顯感受到差異的程度 (Aoki et al., 1991)。亦有許多研究應用此套系統做為評斷色彩變化的客觀依據:大 西 洋 鮭 魚 (Salmo salar L.)(Buttle et al., 2001) 、 虹 鱒 (Oncorhynchus mykiss)(Storebakkena et al., 2004)、嘉鱲 (Pagrus auratus)(Booth et al., 2004;
Kalinowski et al., 2005;Doolan et al., 2008)、海膽(Psammechinus miliaris)(Suckling et al., 2011)等。
6
研究動機與目的
本實驗室已針對不同濃度與來源的類胡蘿蔔素在不同魚種增豔效果的影響 有過多次的探討:飼料中添加不同類胡蘿蔔素對赤鰭笛鯛之影響(曾, 2004)、 飼 料中添加不同濃度與來源之類胡蘿蔔素對紅色吳郭魚體表增豔效果之影響(黃, 2009)、 飼料中添加不同類胡蘿蔔素對紅頭金鯛之呈色影響(劉, 2012)。因此本實 驗將探討類胡蘿蔔素在珍珠鱗的呈色影響,使金魚或是其他觀賞魚種的增豔飼料 開發更加完善。本實驗目的主要分成兩個部分:
1. 利用在珍珠鱗金魚之飼料中添加不同濃度的蝦紅素,期望能找出蝦紅素的色 素蓄積飽和曲線,在飼料成本及時效性的考量上,研究出最適的添加量。
2. 為評估珍珠鱗金魚之類胡蘿蔔素代謝模式,在飼料中添加玉米黃素,測試其 是否能轉換成蝦紅素,替代蝦紅素添加在珠鱗的飼料中使用。
7
第二章 材料與方法 一、 實驗魚種及試驗條件
試驗用魚種珍珠鱗金魚,馴養四星期間投餵不含類胡蘿蔔素的飼料以免在馴 養期間攝入類胡蘿蔔素而影響實驗,並於分組試驗前一天絕食。隨機採樣分組於 長*寬*高為 45*30 *30cm*30cm 配有上部過濾的水缸,每組固定有一隻白色體色 的珍珠鱗金魚為對照,共分 9 組,每組 8 尾,魚體平均重 2.69±0.23 克。試驗預 計進行 7 週,分別於第 5 星期和第 7 星期秤重一次。
實驗進行期間,每日依攝食狀況,投餵總量為魚體體重 2-4 %的實驗飼料,
每日投餵二次,並於每次餵食完一小時後清除殘餌與換水,每日換水量約為總水 體的 3/4,定期清洗過濾棉及刷洗內壁,避免藻類生長。實驗於室內進行,飼養 期間水溫範圍 19~29℃。
實驗共進行 7 週,各組皆投餵無添加類胡蘿蔔素的對照組飼料。實驗於第 5、
7 週秤重一次和測量體表呈色(L*,a*,b*),7 週後秤重並隨機採樣 4 尾(摒除白色的 個體),冰存於-20 ℃的冰箱內,以供日後魚體之類胡蘿蔔素分析。
二、 實驗飼料配製
實驗用之飼料配方如 Table 1 所示,以 25%魚粉及豆粉 45%做為蛋白質主 要來源,脂肪以魚油 1.2%及大豆油 1.8%添加,配置出蛋白質含量約為 25%,
粗脂肪含量約為 9.05%的九組飼料,對照組飼料無添加類胡蘿蔔素為對照組;
第二組到第四組分別添加濃度為 5、10、15、20mg/kg 的玉米黃素(Zeaxanthin);
第六組到第九組分別添加濃度為 5、10、15、20mg/kg 的合成蝦紅素(Synthesis Astaxanthin)。
將各種原料攪拌均勻後,加入原料 30~35%的水混合均勻,通過孔徑直徑 2mm 之擠粒機,再分成適當大小,送入鼓風式乾燥機,以 40℃烘 15 小時,冷卻 後保存在-20 ℃冰箱。
三、 飼料成分分析 (一) 水分與灰分
8
依據 AOAC ( 1984 ) 方法分析。首先將坩鍋與坩鍋蓋以 110 ℃烘乾 2 小時,
再將坩鍋移到乾燥器中冷卻半小時後,秤坩鍋重量(W0)。
1. 水分分析方法
秤取飼料 0.5 g 置於坩鍋,以烘箱 110℃烘乾,每隔 2 小時秤重一次,秤重 前先坩鍋移到乾燥器中冷卻半小時,直到所測坩鍋重量(W1) 達恆定為止。將 W1 減去 W0,可得水分重量;將水分重量再除以一開始所秤的飼料重量,即可 求得飼料中的水分。
2. 灰分分析方法
秤取飼料 0.5 g 置於坩鍋並加蓋坩鍋蓋,以灰化爐 600℃持續加熱 15 小時,
秤重前先坩鍋移到乾燥器中冷卻半小時,測坩鍋重量(W2)。將 W2 減去 W0,可 得灰分重量;灰分重量再除以一開始所秤的飼料重量,即可求得飼料中的灰分。
(二) 粗蛋白
各組飼料蛋白質之含量,皆依照 Micro-Kjeldahl 的分析方法( AOAC,1984 ) 來分析。取樣品( 0.2 g )以 90 mm 濾紙( Toyo,Japan )包覆,放入凱氏氮分解瓶中,
加入 K2SO4 : CuSO4 = 9 : 1 的催化劑 3 g,隨後再加入 10 ml 18 N 的濃硫酸,將凱 氏氮分解瓶放入粗蛋白質分解裝置( Büchi 435 ),先以 210 ℃預熱 30 分鐘,接著 再提高溫度至 560 ℃,約二小時之後分解完畢,此時溶液為淡藍綠色且澄清,
使其於室溫下冷卻,將此分解完畢的樣品,以凱氏氮自動測定儀( Büchi K355 ) 根據硫酸滴定量,再計算樣本中粗蛋白質含量。
自動分析裝置使用的溶劑及溶液分別為:30 ml 蒸餾水、75 ml 氫氧化鈉 ( NaOH, 32% )、50 ml 硼酸( H3BO3, 2% )及滴定用硫酸( H2SO4, 0.1 N )。換算公式 如下:
粗蛋白(%) = [6.25x 0.1x 1.0x 硫酸滴定量(ml) x 14 ÷ 樣本重(mg)]x 100
9
(三) 粗脂質
依照 Folch 等人( 1957 )的方法。秤取飼料 2 克,研磨成粉狀,加入 50 ml 氯 仿/甲醇 ( choloroform / methanol, 2:1 v/v )的溶液,以均質機 ( Nissei AM-3, Tokyo Japan ) 5000 rpm 攪拌 5 分鐘,再以 Büchner funnel 過濾,並以氯仿/甲醇 ( 2:1 v/v ) 50 ml 洗滌,將過濾液完全移入分液漏斗中,並加入 0.03 M 氯化鎂 ( MgCl2 ) 20 ml 強力混合 1 分鐘,靜置一夜。取濃縮瓶加入氯仿,以減壓回轉濃 縮機濃縮後,將空濃縮瓶秤重(W0)。取分液漏斗中含脂質之下層液至濃縮瓶中,
以減壓回轉濃縮機濃縮,將含萃取物之濃縮瓶秤重(W1)。將 W1 減去 W0,可得 粗脂質重量;將粗脂質重量除以一開始所秤的飼料重量,即可求得飼料中的粗脂
質 。
四、 體表呈色測定
利用色差計(Minolta CR-10 hand held chromameter)直接偵測魚體的各個測 量點(Fig.1),測量出顏色參數(a*,b*和 L*值),並比較顏色差異(△E)。顏 色參數係依據國際照明委員會(Munsell Color Science Lab)制定之色彩空間 CIE Lab 系統,a*值越高代表紅色程度越高,而 a*值越低代表綠色程度越高;b*值越 高代表黃色程度越高,而 b*值越低代表藍色程度越高;L*值代表亮度,其值越 高代表亮度越高。當△E>6 則表示已達人眼感官可明顯分辨的標準。
五、 類胡蘿蔔素分析 類胡蘿蔔素總量
魚皮經剝取後秤重置於研缽,加入適量無水硫酸鈉共同研磨,利用丙酮萃取 組織中的類胡蘿蔔素,並將丙酮萃取液以 90 mm 濾紙( Toyo,Japan )過濾後收集 至濃縮瓶中,重覆加入丙酮萃取組織中的類胡蘿蔔素,直至丙酮萃取液出現的顏 色極淡。以減壓迴轉濃縮機(Yamato BM42、Yamato BM400)將收集的萃取液進 行濃縮。濃縮完後加入總量為 15 ml 乙醚與 5 ml 去離子水,各分 3 次加入濃縮 瓶中,輕輕搖晃後,將溶液倒入分液漏斗,並上下搖晃充分混合溶液,待溶液靜 置分層後收集乙醚層,水層再另外加入乙醚,同樣以分液漏斗萃取乙醚層,重覆 進行 3 次。將收集到的乙醚層再以減壓迴轉濃縮機進行濃縮。
10
濃縮後得到的產物以苯溶解,移至定量瓶中,定容至標線,以分光光度計 (Thermo AQUAMATE)測量樣本在波長 480 nm 的吸光值。總類胡蘿蔔素的含量 換算根據 McBeth’s formula。最後將樣本移入樣本瓶,保存於-20 度冰箱,待類 胡蘿蔔素組成分析。
六、 參數分析
(一) 增重百分比 Weight gain (%)
=[ Final body weight (g)-Initial body weight(g) ] / [ Initial body weight (g) ]×100
(二) 飼料效率 Feed efficiency (%)
=[ Body weight gain (g) / Feed intake(g) ]×100 (三) 存活率 Survival (%)
= ( Final fish number / Initial fish number ) ×100 (四) 色差值 ΔE
=〔(a-ac)2+(b-bc)2+(L-Lc)2〕1/2 ac, bc和 Lc為對照組的 a, b 和 L 值
(五) 類胡蘿蔔素含量 Carotenoids content (mg carotenoids / 100g tissue)
=[ ODλ×Vol.(ml) ×103 ] /[ E×weight of sample(g) ]
E1%1cm:類胡蘿蔔素在苯中 1%的 extinction coefficient value=1900 七、 統計分析
以 SAS(Statistical Analysis System) 套 裝 軟 體 對 測 量 參 數 進 行 one-way ANOVA 分析比較,若存在顯著差異時,續以 Tukey’s HSD (Honestly Significant Difference) test,做各組間兩兩比較(α=0.05)。
11
第三章 結果 一、 飼料分析
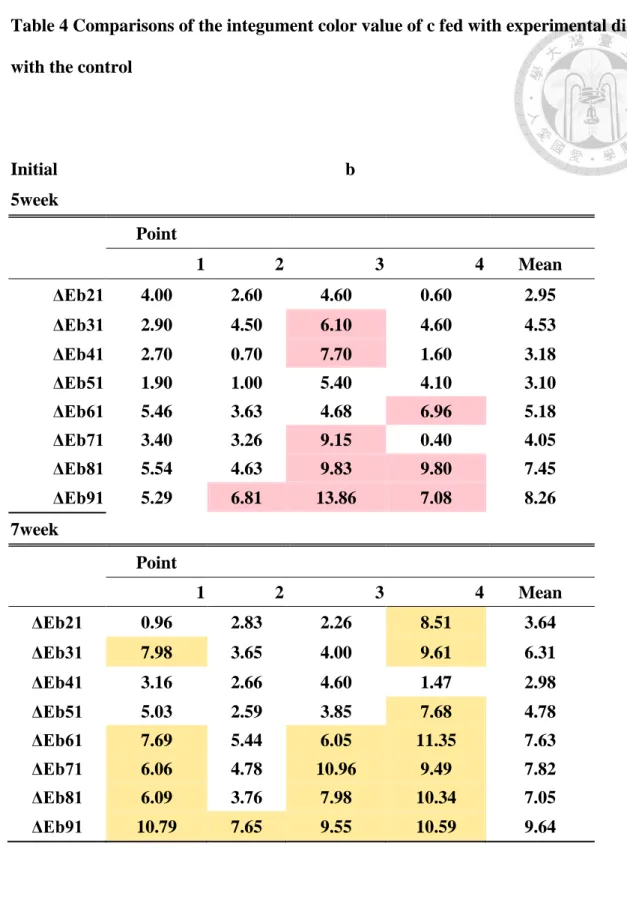
實驗飼料設計及成分分析如 Table 1 所示。配製的實驗飼料水分含量為 5.91-8.46 %,灰份含量大約為 11 %,蛋白質含量在 24.78-25.94%之間,粗脂質 含量為 9.5-11%。使用兩種類胡蘿蔔素來源,添加玉米黃素的 4 組 (Z5、Z10、
Z15、Z20)使用 5、10、15、20mg/kg 的玉米黃素;而添加合成蝦紅素的 4 組(S5、
S10、S15、S20)使用濃度為 5、10、15、20mg/kg 的合成蝦紅素。實際測量飼料 中的類胡蘿蔔素含量,Z5、Z10、Z15、Z20 為 0.0263、0.0263、0.0526、0.1056 mg/kg diet ,而 S5、S10、S15、S20 為 0.2105、0.1842、0.2368、0.2105 mg/kg diet。
二、 成長與活存
在珍珠鱗金魚的飼料中分別添加不同類胡蘿蔔素含量為玉米黃素 5、10、15、
20 mg/kg(Z5、Z10、Z15、Z20)與合成蝦紅素 5、10、15、20 mg/kg(S5、S10、
S15、S20),飼養 5 個星期及 7 個星期後,其成長結果如 Table 2 所示。首先各 組在飼養 5 個星期後,平均體重由 2.5-3.07g 增加到 3.18-4.28g,魚隻重量在各組 之間並無顯著差異,增重率為 27 - 43%,飼料效率為 15 - 17%。接著在飼養 7 個 星期後,平均體重由 2.5-3.07g 增加到 3.88-4.97g,魚隻重量在各組之間並無顯著 差異,增重率為 54-77%,飼料效率為 13-15%。由本實驗結果顯示,在飼料中有 無添加或添加種類的不同的類胡蘿蔔素並不影響珍珠鱗金魚的增重率與飼料效 率。除此之外,本實驗結果亦顯示飼養期間各階段之存活率除了第 8 組在第 7 星期死亡一隻外皆為 100%。
三、 體表顏色測定
由色差計偵測魚體的體表各測點 (Fig.2)顯示實驗進行至 5、7 週的呈色效 果。
(一)L 值(lightness 亮度,值介於 0-100,其值愈高代表亮度愈高) 1. 臉頰(Point 1)
12
第 5 星期的平均 L 值在 34.2-42.2,皆低於初始的 49.7。第 7 星期的平均 L 值在 39.9-47.9,其中以 Z5 與 Z15 組最高,有顯著差異。由臉頰的 L 值(Fig.3)結 果顯示,第 7 星期在類胡蘿蔔素含量較多的組別,其亮度也略高於其它組別,但 在第 5 星期時則沒有此趨勢。
2. 背部(Point 2)
第 5 星期的平均 L 值在 43.1-57.8;第 7 星期的平均 L 值在 41.0-52.0,兩次 測量皆以對照組的值最高,第五星期與 S15、S20 組存在顯著差異;第 7 星期與 大多的組別存在顯著差異。在背部的 L 值(Fig.4)結果顯示,第 5、7 星期各組的 亮度亦相差不大。
3. 尾部(Point 3)
第 5 星期的平均 L 值在 44.2-57.8,在各組之間並無顯著差異。第 7 星期的 平均 L 值在 43.0-49.6,其中皆以對照組的值最高,與 S10、S15 與 S20 組存在顯 著差異。在背部的 L 值(Fig.5)結果顯示,在第 5 星期時各組之間的亮度沒有差異,
而第 5、7 星期各組的亮度亦相差不大。
4. 腹部(Point 4)
第 5 星期的平均 L 值在 56.2-63.6,其中對照組的值較高,而 S10 及 S15 組 的值較低。第 7 星期的平均 L 值在 53.7-63.4,其中以對照組的值最高,而 S15 及 S20 組最低,與對照組存在顯著差異。在尾鰭的 L 值(Fig.6)結果顯示,第 5、
7 星期各組的亮度亦相差不大。
(二)a 值(redness,其值愈高代表紅色程度愈高) 1. 臉頰(Point 1)
第 5 星期的平均 a 值在 9.6-18.2,其中以對照組為最低、S10 組為最高,S15 組次之,存在組間顯著差異。第 7 星期的平均 a 值在 11.4-22.8,其中以對照組最 低,S15 及 S20 組最高,存在組間顯著差異。由臉頰的 a 值(Fig.7)結果顯示,添 加蝦紅素劑量較高的 S15、S20 組,在實驗期間其紅色的程度明顯比其它組高,
13
而添加玉米黃素的 Z10 組的紅色程度則介於 S5 及 S10 組的紅色程度之間,相當 接近,其中對照組在第 5 星期和第 7 星期皆最低。
2. 背部(Point 2)
在第 5 星期的平均 a 值在 4.8-14.7,其中以 S15 及 S20 組最高,a 值皆為 14.7;
而對照、Z5 組為最低,分別為 4.8 和 7.4,存在顯著差異。第 7 星期的平均 a 值 在 6.3-14.5,其中添加合成蝦紅素的 S15 及 S20 組的紅色程度最高,與對照組、
添加玉米黃素的 Z5 的紅色程度相差甚多,存在著顯著差異。在背部的 a 值(Fig.8) 結果顯示,自第 5-7 星期,都可以偵測到添加合成蝦紅素的各組別紅色程度明顯 高於對照組與添加玉米黃素的組別;另外在第 5 星期時,背部的紅色程度在添加 合成蝦紅素的四組為相當接近,但於第 7 星期時,S15 組明顯為各組最高。
3. 尾鰭(Point 3)
在第 5 星期的平均 a 值在 2.7-20.2,添加玉米黃素的組別和添加合成蝦紅素 的各組別紅色程度皆明顯高於對照組,具有顯著差異,而添加合成蝦紅素的組之 紅色程度也高於添加玉米紅素的各組別紅色程度。第 7 星期的平均 a 值在 8.7-18.5,
添加蝦紅素的組別已遠高於對照組與添加玉米黃素的組別,但對照組的 a 值有明 顯的增高的趨勢。由尾鰭的 a 值(Fig.9)結果顯示,添加蝦紅素的組別尾鰭紅色程 度較對照組與玉米黃素高。
4. 腹部(Point 4)
在第 5 星期的平均 a 值在 1.3-11.3,添加玉米黃素的組別和添加合成蝦紅素 的各組別紅色程度皆明顯高於對照組,具有顯著差異,但 Z15、Z20 和 S5、S10 這四組並無太大的差別。第 7 星期的平均 a 值在 3.9-11.6,S15 及 S20 組的 a 值 已遠高於其他的組別,但對照組的 a 值有明顯的增加,而 Z15 組的紅色程度不增 反減。由腹部的 a 值(Fig.10)結果顯示,添加蝦紅素的組別尾鰭紅色程度較對照 組與玉米黃素高。
(三)b 值(Yellowness,其值愈高代表黃色程度愈高)
14
1. 臉頰(Point 1)
第 5 星期的平均 b 值在 20.8-26.3,其中以添加合成蝦紅素的 S15 組的值較 高,對照組、添加玉米黃素的組別和添加合成蝦紅色的組別其 b 值相差並不大,
無顯著差異。第 7 星期的平均 b 值在 23.3-34.1,其中以 Z10、S5、S20 組較高,
而對照組最低,存在顯著差異。由臉頰的 b 值(Fig.11)結果顯示,第 7 星期,添 加玉米黃素的組別,除了 Z5 組外,黃色的程度都相對提高許多,Z10 及 S20 組 在第 5 星期和第 7 星期的 b 值變化上是有顯著差異的。
2. 背部(Point 2)
在第 5 星期的平均 b 值在 25.1-32,其中以 Z10、S15 及 S20 組為高,且兩種 不同的添加色素組之 b 值相差不大,無顯著差異。第 7 星期的平均 b 值在 23.5-31.2,
其中雖以 S20 的值為最高,但與第 5 星期一樣並無顯著差異。由背部的 b 值(Fig.12) 結果顯示,對照組、添加玉米黃素的組別和添加合成蝦紅色的各組 b 值相差並不 大。
3. 尾鰭(Point 3)
第 5 星期的平均 b 值在 19.7-33.5,以添加合成蝦紅素 S20 組的值為最高、
對照組為最低,存在組間顯著差異。第 7 星期的平均 b 值在 23.4-32.9,以添加合 成蝦紅素 S10 組的值為最高,存對照組依然為最低,在組間顯著差異。在尾鰭的 b 值(Fig.13)結果顯示,添加濃度較高的類胡蘿蔔素的各組尾鰭的黃色程度會比濃 度較低的組別相對也較高。
4. 腹部(Point 4)
第 5 星期的平均 b 值在 32.2-42,其中以 S15 組為高,且兩種不同來源的添 加色素組之 b 值相差不大,但與對照組存在顯著差異。第 7 星期的平均 b 值在 29.3-40.7,其中以 S5 的值為最高,但與第 5 星期一樣,各組間的 b 值相差不大,
但還是與對照組存在著顯著差異。由背部的 b 值(Fig.14)結果顯示,添加玉米黃 素的組別和添加合成蝦紅色的各組 b 值相差並不大,但在添加的類胡蘿蔔素之濃 度較高的組別和對照組存在著顯著差異的。
15
(四) ΔE、ΔEa、ΔEb(ΔE 可比較整體色差,另外 ΔEa、ΔEb 可分別檢視紅色與黃 色的差異程度)
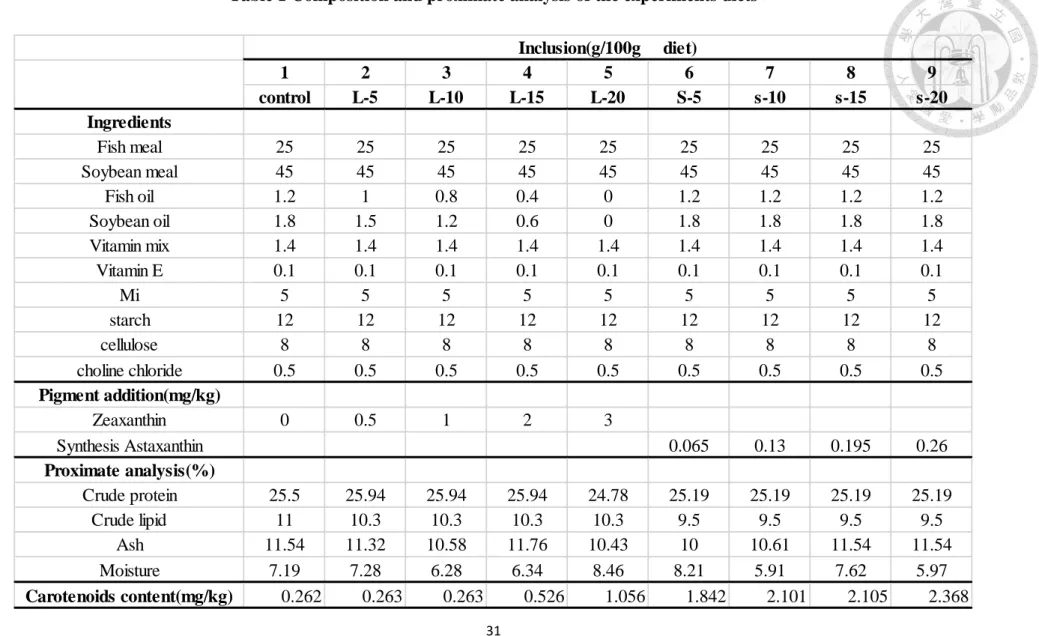
Z5、Z10、Z15、Z20、S5、S10、S15、S20 八組與對照組的色彩差異比較結果如 Table4 和 Fig.15-17 所示。
1. 各測點之平均ΔE 值(Fig.15)
在第 5 星期其整體色彩差異值(ΔE)為 5.96-14.33,除 ΔE2Ctrl 組為 5.96 外其 餘組別皆大於 6(己達人眼感官清楚辨別的水準),其中又以添加合成蝦紅素的四 組的平均ΔE 值最高;而在第 7 星期各組的整體色彩差異值(ΔE)為 5.27-15.22,
ΔE2Ctrl、△E4Ctrl、△E7Ctrl 三組皆為下降,其他組別的 ΔE 皆為上升,甚至 S20 組已高達 15.22,為標準的 2 倍之多。
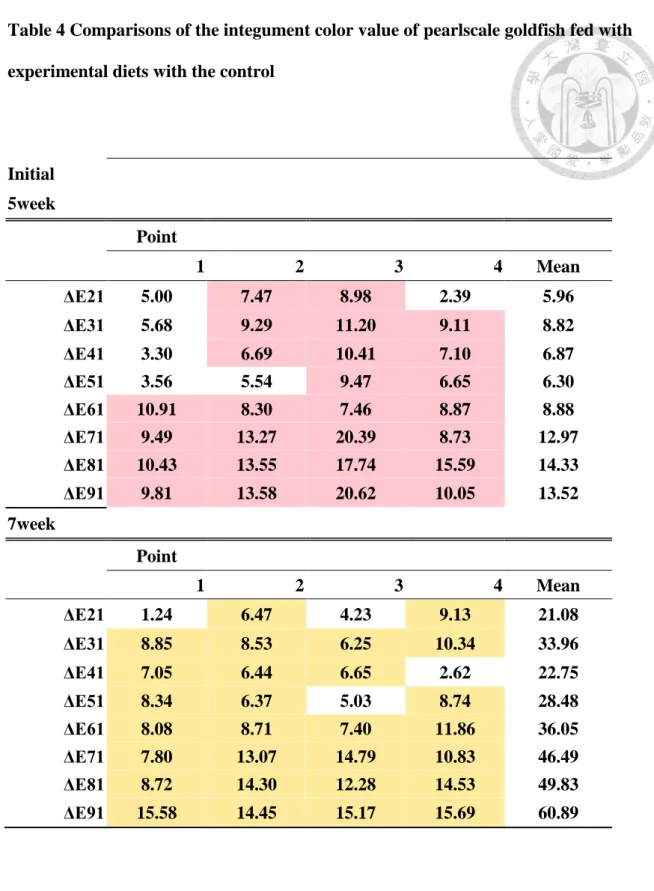
2. 各測點之平均ΔEa(Fig.16)
在第 5 星期,添加合成蝦紅素的 ΔEa 除 S20 組外皆高於添加玉米黃素的組 別,到第 7 星期,添加玉米黃色的組別的 ΔEa 與第 5 星期比較起來想對下降許 多,而添加合成蝦紅色的組別 ΔEa 有相對增加的趨勢,由其表現在 S20 組上,
表示添加合成蝦紅素的組別與對照組之紅色差異程度,明顯高於添加玉米黃素的 組別,其結果亦能清楚由第 7 星期的魚隻照片(Fig.1)印證。
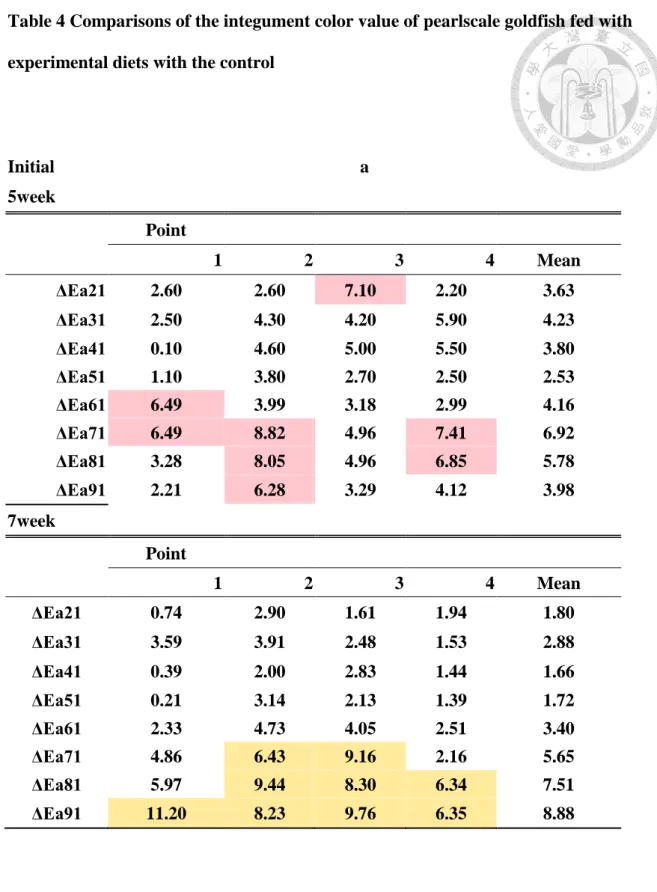
3. 各測點之平均ΔEb(Fig.17)
第 5 星期,添加合成蝦紅素的組別 ΔEb 高於添加玉米黃素的組別,但到第 7 星期,可以發現添加玉米黃素的 Z5、Z15、Z20 組和添加合成蝦紅紅素的 S5、
S10 組之 ΔEb 提升有明顯的成長且漲幅大。
四、 表皮總類胡蘿蔔素含量測定
表皮總類胡蘿蔔素測定的結果如 Table5 和 Fig.18 所示。各組魚皮所含有的 平均類胡蘿蔔素含量由對照組的 0.021 mg/100g 最低,其餘有添加類胡蘿蔔素的 組別皆比對照組高,介於 0.42-0.25 mg/100g,以 S10 組的 0.100 mg/100g、S15 組的 0.105 mg/100g 和 S20 組的 0.25 mg/100g 為最高的三組。平均總類胡蘿蔔素 量可達到 0.253 mg/100g;比較實驗前分析之魚皮中的總類胡蘿蔔素含量可以發
16
現,實驗前所分析的數據為 0.021 mg/100g,與其餘投餵含有類胡蘿蔔素添加的 各組之表皮所含總類胡蘿蔔素含量則明顯高於對照組。
17
第四章 討論 一、 成長效果
由本實驗結果指出,在平均體重 2.69g 的珍珠鱗金魚飼料中,添加玉米黃素、
合成蝦紅素做為類胡蘿蔔素來源以及不添加類胡蘿蔔素的對照組,其實驗魚隻的 增重率、飼料效率、存活率在各組中並無顯著差異。此結果顯示無論珍珠鱗金魚 的飼料中有無添加類胡蘿蔔素抑或是添加不同濃度與種類的類胡蘿蔔素,皆不影 響魚隻的成長與活存。
另外在其它魚種的實驗當中,亦出現相似的結果。在平均體重 5g 的金魚 (Carassius auratus) 以紅酵母(Xanthophyllomyces dendrorhous)做為蝦紅素來源,
添加 60mg/kg 含量的類胡蘿蔔素到飼料中,經 60 天飼養後,皆與對照組間的成 長沒有顯著差異(Xu, 2006);在平均體重 300g 的大西洋鮭(Salmo salar L.)的飼料 中分別添加 50mg/kg diet 的蝦紅素與不同濃度之葉黃素(lutein),飼養 138 天後,
皆與對照組間的成長沒有顯著差異(Olsen and Baker, 2006);在平均體重 88g 的嘉 鱲(Pagrus auratus) 分別在飼料中添加 60mg/kg diet 斑螫黃素(canthaxanthin)與蝦 紅素,飼養 50 天以後,皆與對照組間的成長沒有顯著差異(Doolan et al., 2008);
Sun et al., (2012)在飼料中添加葉黃素和合成蝦紅素(嘉麗紅, Carophyll® red) 投 餵平均體中約為 18.05g 的昭和錦鯉 99 天,其增重率沒有明顯差異。
利用不同大小的嘉鱲(Pagrus pagrus)所進行的研究,在平均體重 44g 的嘉鱲 (Pagrus pagrus)在飼料中分別添加斑螫黃素(canthaxanthin)與以蝦殼粉為來源的 蝦紅素,飼養 105 天後,皆與對照組間的成長沒有顯著差異(Kalinowski et al., 2005);平均體重 82g、150g 的嘉鱲(Pagrus pagrus)的飼料中分別添加 100mg/kg diet 的蝦紅素、β-胡蘿蔔素(β-carotene)、茄紅素(lycopene),飼養 10 週後,皆與對照 組間的成長沒有顯著差異(Chatzifotis et al., 2005);平均體重 223g 的嘉鱲(Pagrus pagrus)以蟹殼粉做為蝦紅素來源,添加 3-6mg/kg 含量的類胡蘿蔔素到飼料中,
經 165 天飼養後,皆與對照組間的成長沒有顯著差異(Garcia et al., 2010)。
雖然在水生生物的生理代謝過程中,類胡蘿蔔素扮演了許多重要的角色,但 在本篇的研究結果以及前人對於其它的魚種、不同的魚隻大小或者使用其它來源
18
的類胡蘿蔔素之研究中,皆顯示在飼料中添加類胡蘿蔔素並不會對魚隻的成長產 生影響。
二、 呈色
由實驗結果的照片(Fig.1)可以看出,對照組的體色偏淡,添加玉米黃素的組 別體色偏黃,而添加蝦紅素的組別體色偏紅。從第 7 星期的魚隻照片結果(Fig.1) 亦能客觀的由色彩差異值得到印證。
先初步將實驗組與對照組數值比較平均整體色彩差異值(ΔE),來檢視與各實 驗組與對照組之差異,結果指出添加類胡蘿蔔素的各組,在第 5 星期其整體色彩 差異值(ΔE)為 5.96-14.33,除 ΔE2Ctrl 組為 5.96 外其餘組別皆大於 6(己達人眼感 官清楚辨別的水準),顯示在飼料中添加的這二種色素都可以有效提升魚隻的體 色。而在第 7 星期各組的整體色彩差異值(ΔE)為 5.27-15.22,△E5Ctrl、△E6Ctrl、
△E9Ctrl 三組為上升,其他組別的 ΔE 皆下降,不過添加類胡蘿蔔素的 8 組當中 依然是以添加合成蝦紅色的四組之ΔE 為最高。顯示即使持續的投餵添加類胡蘿 蔔素的飼料也不一定仍能夠維持與對照組之顏色差異。接著分別再以平均紅色差 異值(ΔEa)與黃色差異值(ΔEb)來判斷與對照組之差異,由數據指出在添加合成蝦 紅素的組別,其紅色程度明顯比添加玉米黃素的組別高;相反的,在添加玉米黃 素的組別,其黃色程度在第 5 星期和在第 7 星期的成長上明顯比添加合成蝦紅素 的組別高,且造成紅色程度的下降。結果顯示添加合成蝦紅素主要能夠明顯提升 的為紅色色澤,而添加玉米黃素能夠明顯提升的為黃色色澤。
比較各組實際之色差值,紅色程度(a 值)在添加合成蝦紅素的組別均有隨添 加濃度而提升的趨勢,而在對照組與添加玉米黃素的組別之 a 值有下降的情形;
黃色程度(b 值)在對照組與實驗組間,皆有上升的現象,而對照組與添加合成蝦 紅素的組別的黃色程度較為接近;最後在亮度方面(L 值),則出現添加色素濃度 愈高, L 值愈低的趨勢。
各個測點的呈現出魚的體色並非平均分布,以頭部的呈色值最高,而尾鰭、
背部次之,除此之外,在第 7 週時,背部和尾鰭之測點在 S15、S20 的 a 值均無 顯著差異,故推論其紅色程度都己達到了飽合。
19
在赤鰭笛鯛(Lutanus erythropteru)的研究中,平均 35g 的魚隻的飼料中添加 50、 100mg/kg diet 的蝦紅素、螺旋藻,飼養 8 週後,可使平均 ΔE 達 8.6-13.3;
平均 2.32g 的魚隻飼料中添加 50、 100mg/kg diet 的蝦紅素、β-胡蘿蔔素、南極 蝦粉,飼養 6 週後,可使平均 ΔE 達 7.0-9.0,二次實驗結果皆達人眼感官能明顯 感受到差異的效果。另外,a*值的增加皆明顯較 b*、L*高,顯示與對照組的主 要呈現在紅色的差異(曾,2004)。另外在紅色吳郭魚(Oreochromis sp.)的研究中,
平均 2.6g 的魚隻的飼料中添加 50、 100mg/kg diet 的蝦紅素,飼養 10 週後,可 使平均ΔE 達 8.06-14.89;而平均 53g 的魚隻,飼養 60 天後,可使平均 ΔE 達 9.06-14.15,二次實驗結果皆達人眼感官能明顯感受到差異的效果,另外,a*值 皆明顯較 b*、L*高,顯示與對照組的主要呈現在紅色的差異(黃,2009)。
在先前的研究當中,嘉鱲(Pagrus pagrus)亦呈現魚體體色分布不均的現象,
在前側出現的紅色程度最高,接著是尾鰭和背部,色彩飽合度在 105 天的時候下 降,推測顏色己達到飽合,在 L 值方面則沒有影響(Kalinowski et al., 2005);同 樣以嘉鱲(Pagrus pagrus)為實驗魚種,以蟹殼粉做為蝦紅素來源,添加到飼料中,
經 165 天飼養後,對體色有促進的效果,但在 L*值在各組之間沒有差異(Garcia et al., 2010)。而在嘉鱲(Pagrus auratus)的研究當中指出,隨飼料中添加的蝦紅素,
可提高體表的紅色程度,在第 6 週以後,添加 36 與 72mg/kg diet 之組別,所呈 現之 a 值沒有顯著差異,由此推測己達到蝦紅素蓄積的飽合,另外在 L*值與 b*
值方面,並沒有影響(Booth et al., 2004);另外一篇研究也指出,嘉鱲(Pagrus auratus)隨飼料中添加的蝦紅素,可提高體表的紅色程度,在第 50 天以後,添加 30 與 60mg/kg diet 之組別,所呈現 a*、b*值沒有顯著差異,由此推測己達到蝦 紅素蓄積的飽合,另外在 L*值方面,並沒有影響(Doolan et al., 2008)。
除了添加玉米黃素的組別之外,在添加合成蝦紅素的組別中,珍珠鱗金魚體 表的黃色也有增加的現象,而無另外添加類胡蘿蔔素的對照組亦呈現出淡淡的黃 色色澤,可能是飼料所添加的魚粉也含有一些類胡蘿蔔素(0.018mg/kg diet)所造 成的,在添加玉米黃素組別當中的白色個體魚其體色亦呈現出淡淡的黃色色澤。
雖然對照組中的類胡蘿蔔素沒有確定其組成,但由在組織萃取出的色素在苯中所
20
測得的吸光值最大值落在 480nm 左右,推測對照組與添加蝦紅素組別之體表出 現黃色,是因為原始飼料中含有一些主要組成為黃色的類胡蘿蔔素所致。
由 L 值結果顯示,添加色素濃度愈高,而亮度 L 值愈低的趨勢。由照片結 果可以看出對照組的魚略為較黑,而添加類胡蘿蔔素的組別,魚體略為較白的現 象。而在赤鰭笛鯛(Lutjanus erythropterus)的研究中亦指出,平均體重 35g 的魚隻,
飼料中添加螺旋藻粉與蝦紅素做為色素來源,飼養 8 週之後,養殖魚隻體表的亮 度有下降的現象(曾,2004)。但在前述其它的研究當中,並沒出現這個趨勢。
不過亦有研究認為,添加也有光保謢作用的類胡蘿蔔素在飼料當中,可能可 以促使黑色素下降。相反的低劑量的類胡蘿蔔素可能會促使黑色素生成,因為類 胡蘿蔔素具有抗氧化壓力的效果(Chatzifotis et al., 2005)。因此推論,原來需要黑 色素保護的珍珠鱗金魚,魚隻成長的大小剛好正在經歷體色轉變過程,也許此時 在飼料中獲得也有保護效果的類胡蘿蔔素,能夠幫助魚皮組織褪去黑色素,造成 體表亮度的提高。
三、 色素蓄積效果
在總類胡蘿蔔素的蓄積量分析可以發現,在飼料中添加玉米黃素及合成蝦紅 素的魚隻,組織內的類胡蘿蔔素蓄積量都會隨飼養天數而增加,在第 7 週時,對 照組的類胡蘿蔔素含量為 0.021mg/100g,實驗組為 0.021-0.253 mg/100g,而添加 玉米黃素 Z15、Z20,在所取樣的各部位組織中己達到了色素蓄積的飽合程度,
此外由添加劑量相近的 Z5 與 S10 相比,其色素在魚體中蓄積的量並無顯著差異,
結果顯示在珍珠鱗金魚的身上,合成蝦紅素與玉米黃素皆能有效的被蓄積在體 內。
在赤鰭笛鯛(Lutanus erythropteru)的研究中,平均 35g 的魚隻的飼料中添加 50、 100mg/kg diet 的蝦紅素、螺旋藻,飼養 8 週後,類胡蘿素含量為 0.99-1.62 mg/100g;平均 2.32g 的魚隻飼料中添加 50、 100mg/kg diet 的蝦紅素、β-胡蘿蔔 素、南極蝦粉,飼養 6 週後,類胡蘿素含量為 0.36-1.74 mg/100g(曾,2004)。而 在紅色吳郭魚(Oreochromis sp.)的研究中,平均 2.6g 的魚隻的飼料中添加 50、
100mg/kg diet 的蝦紅素,飼養 10 週後,對照組的類胡蘿蔔素含量為 0.25mg/100g,
21
實驗組為 0.84 mg/100g;平均 53g 的魚隻,飼養 60 天後,對照組的類胡蘿蔔素 含量為 0.25mg/100g,實驗組為 4.08 mg/100g(黃,2009)。
在其它魚種中的研究當中,嘉鱲(Pagrus pagrus) 在飼料中添加 100mg/kg diet 的蝦紅素,飼養 10 週後,體表呈現與野生相近的紅色澤,而魚皮中的類胡蘿蔔 素明顯高於照組的含量,實驗中也嘗試添加β-胡蘿蔔素(β-carotene)、茄紅素 (lycopene),但結果只有蝦紅素能明顯提升背部魚皮類胡蘿素的含量,而 β-胡蘿 蔔素(β-carotene)、茄紅素(lycopene)並沒有明顯的效果 (Chatzifotis et al., 2005)。
金魚(Carassius auratus)以 60mg/kg 的濃度在飼料中添加蝦紅素,15 天後即與對 照組有顯著差異,不同部位蓄積的色素的能力不同,在色素濃度最高的出現在尾 鰭,接著是魚鱗、頭部(Xu et al., 2006)。
魚類因應不同時期的生理需求,可能發生蓄積的類胡蘿蔔素在體內產生轉移 到不同組織的現象。在紅鮭(Oncorhynchus nerka)的研究中發現,紅鮭在產卵時,
魚體肌肉中的游離蝦紅素僅剩產卵前的1%,其中雄魚以酯化的型式蓄積了身上 95%蝦紅素在皮膚,而在雌魚則以游離的型式將身上85%的蝦紅素蓄積在卵當中 (Crozier, 1970)。鮭魚在經歷性成熟時期,為了產卵而在溪流中上溯,此時肌肉 中的類胡蘿蔔素會開始下降,經由血液運送到魚皮和生殖腺(Ando et al., 1986)。
四、 代謝轉換能力
各個魚種對類胡蘿蔔素的代謝轉換能力不盡相同,在嘉鱲(Pagrus pagrus)的 研究指出,添加 25-50mg/kg 的蝦紅素到飼料中,經 4 個月飼養後,其魚皮中蝦 紅素的含量為 2.9-4.8mg/100g,組成佔總類胡蘿蔔素的 65-84%,其研究指出在飼 料中添加的蝦紅素較多時,體內蓄積的蝦紅素比例也會比較高(Tejera et al., 2007) 。
經飼養 7 週後珍珠鱗金魚於添加合成蝦紅色 5-20mg/kg 的組別中,魚皮中的 蝦紅素含量為 0.063-0.253mg/100g。結果相符與前述研究,也就是飼料中添加的 合成蝦紅素量會影響到體內蓄積色素的組成。此外,其體內蓄積的色素組成皆以
22
合成蝦紅素為主,並沒有發現轉換成其它種類胡蘿蔔素的現象。另外在添加玉米 黃素的組別,其組成雖以玉米黃素為主,但可以發現在紅色程度皆有成長的現像,
可以推測玉米黃素有轉換成蝦紅素的現象。
由以上結果推測珍珠鱗金魚,在水生動物代謝轉換類型中,確實是如紅鯉魚 型(red carp type)的代謝類型能夠將玉米黃素,或其中間產物轉換成蝦紅素。
五、 最適添加量
由第 5 星期呈色實驗的結果,以無添加類胡蘿蔔素的對照組與添加玉米黃素 的 Z5、Z10、Z15、Z20 四組及添加合成蝦紅素的 S5、S10、S15、S20 的四組做 評估。藉由色差計在魚體 4 點所測得之 a 值,可以看出玉米黃素之最適添加量為 15mg/kg diet 而合成蝦紅素亦為 15mg/kg diet。
在嘉鱲(Pagrus pagrus)的研究中指出,以體表呈色值做為評估依據,建議以 蝦殼粉做為色素來源時,蝦紅素在飼料中的最適添為 40 mg/kg diet,以期能夠有 效促進體色呈現紅色(Kalinowski et al., 2005);而在嘉鱲(Pagrus auratus)的研究中 建議,以體表呈色值做為評估依據,蝦紅素在飼料中的最適添為 30 mg/kg diet,
經 50 天在箱網中飼養後可以有效的增進紅色色澤(Doolan et al., 2008);另外在金 魚(Carassius auratus)的研究中建議,由體內的類胡蘿蔔素蓄積為評估依據,添加 100mg/kg diet 的類胡蘿蔔素到飼料中,經 60 天飼養後可以有效的增加組織內的 類胡蘿蔔素,進而增強色澤來提高市場接受度(Yanar et al., 2008)。亦有研究建議,
在上市前 120-180 天的嘉鱲(Pagrus pagrus)投餵以蝦殼粉做為蝦紅素來源,蝦紅 素濃度為 21mg/kg diet 的飼料,可以使體色接近野生飼養的魚隻,並能夠提升魚 皮中的類胡蘿蔔素含量。但是實驗組體表的 a*(7.8-9.0)、b*(11.2-12.2)值與野生 魚隻的 a*(8.7)、b*(13.1)值相近,但魚皮類胡蘿蔔素之含量(10.4-14.0mg/kg diet) 卻明顯低於野生魚隻(54.0mg/kg diet)。由相關性檢定發現,魚皮類胡蘿蔔素之含 量與 a*、b*值比較接近對數性相關,而非線性 ,故以人眼感官判斷組織內的色 素含量有其限制性(Kalinowski et al., 2007) 。
綜合以上各點,雖然魚隻體表呈色、組織內的類胡蘿蔔素含量皆可能隨攝取 的色素增進,若就消費者一般僅以人眼感官做為判斷的水產品的外觀呈色的依據
23
而論,其添加的色素劑量可以考慮以體表之呈色效果為標準,較能符合提升魚體 增色之目的實用性與飼料成本之經濟性,並非一定要滿足體內組織蓄積之類胡蘿 蔔素完全達到飽合程度,故建議珍珠鱗金魚在飼料中的添加量為 15mg/kg diet。
除此之外,每個人對於顏色的喜好有所不同,雖然許多東方人傾向選購代表 喜氣的紅色觀賞魚隻,不過也許有消費者偏好黃色魚隻,若有生產黃色魚隻的考 量時,由於實驗結果可以看出玉米黃素與合成蝦紅素的在組織中的蓄積能力相當,
且玉米黃素影響體色提升黃色程度的能力亦不亞於蝦紅素提升紅色程度的能力,
故欲添加玉米黃素至飼料當中,增進黃色的體色,其添加量可以參考合成蝦紅素 的用量。
24
第五章 結論
一、在珍珠鱗金魚的養殖過程中添加玉米黃素與合成蝦紅素可以增加魚體體表 之呈色作用,但對其成長表現沒有影響。
二、類胡蘿蔔素的添加中,玉米黃素與合成蝦紅素皆可明顯增強體色,其中玉米 黃素主要使色差計所測得的 b 值有上升的趨勢,而合成蝦紅素則對 a 值的上 升比較有明顯的作用,而若停止供給類胡蘿蔔素的添加,會使體色出現褪色 的現象。
三、珍珠鱗金魚在水生動物代謝轉換類型中,確實是如紅鯉魚型(red carp type) 的代謝類型能夠將玉米黃素,或其中間產物轉換成蝦紅素。
四、合成蝦紅素影響珍珠鱗金魚體色的呈色效果較玉米黃素為佳,並以 15mg/kg diet 為最適添加量。
25
參考文獻
曾偉誠(2004),飼料中添加不同類胡蘿蔔素對赤鰭笛鯛體色之影響。國立台灣大 學漁業科學研究所論文。
黃侑勖(2009),飼料中添加不同濃度與來源之類胡蘿蔔素對紅色吳郭魚體表增豔 效果之影響。國立台灣大學漁業科學研究所論文。
劉于溶(2012),飼料中添加不同類胡蘿蔔素對紅頭金鯛之呈色影響。國立台灣大 學漁業科學研究所論文。
Amar, E.C., Kiron, V., Satoh, S., and Watanabe, T. (2004). Enhancement of innate immunity in rainbow trout (Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish & Shellfish Immunology 16, 527-537.
Ando, S., Takeyama, T., and Hatano, M. (1986). Deterioration of Chun Salmon Muscle during Spawning Migration .8. Transport Associated with Serum Vitellogenin of Carotenoid in Chum Salmon (Oncorhynchus-Keta). Agricultural and Biological Chemistry 50, 557-563.
Aoki, T., Takada, K., and Kunisaki, N. (1991). On the study of proximate composititon, minteral, fatty acid, free amino acid, muscle hardness, and color difference of six species of wild and culture fishes. 57 10, 1927-1934.
Berglund, A., Bisazza, A., and Pilastro, A. (1996). Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biological Journal of the Linnean Society 58, 385-399.
Bjørnland, T. (1997). UV-vis spectroscopy of carotenoids. In Phytoplankton Pigments in Ceanography (Paris UNESCO), pp. 578-594.
Blount, J.D., Houston, D.C., and Moller, A.P. (2000). Why egg yolk is yellow. Trends in Ecology & Evolution 15, 47-49.
Boonyaratpalin, M., Thongrod, S., Supamattaya, K., Britton, G., and Schlipalius, L.E. (2001).
Effects of beta-carotene source, Dunaliella salina, and astaxanthin on pigmentation, growth, survival and health of Penaeus monodon. Aquaculture Research 32, 182-190.
Booth, M.A., Warner-Smith, R.J., Allan, G.L., and Glencross, B.D. (2004). Effects of dietary astaxanthin source and light manipulation on the skin colour of Australian snapper Pagrus auratus (Bloch & Schneider, 1801). Aquaculture Research 35, 458-464.
Buttle, L., Crampton, V., and Williams, P. (2001). The effect of feed pigment type on flesh pigment deposition and colour in farmed Atlantic salmon, Salmo salar L. Aquaculture Research 32, 103-111.
26
Chatzifotis, S., Pavlidis, M., Jimeno, C.D., Vardanis, G., Sterioti, A., and Divanach, P. (2005).
The effect of different carotenoid sources on skin coloration of cultured red porgy (Pagrus pagrus). Aquaculture Research 36, 1517-1525.
Cianci, M., Rizkallah, P.J., Olczak, A., Raftery, J., Chayen, N.E., Zagalsky, P.F., and Helliwell, J.R. (2002). The molecular basis of the coloration mechanism in lobster shell: beta-crustacyanin at 3.2-angstrom resolution. Proceedings of the National Academy of Sciences of the United States of America 99, 9795-9800.
Clotfelter, E.D., Ardia, D.R., and McGraw, K.J. (2007). Red fish, blue fish: trade-offs between pigmentation and immunity in Betta splendens. Behavioral Ecology 18, 1139-1145.
Craik, J.C.A. (1985). Egg quality and egg pigment content in salmonid fishes. Aquaculture 47, 61-88.
Crozier, G.F. (1970). Tissue carotenoids in prespawning and spawning sockeye salmon (Oncorhynchus-Nerka). Journal of the Fisheries Research Board of Canada 27, 973-&.
Doolan, B.J., Allan, G.L., Booth, M.A., and Jones, P.L. (2008). Effect of carotenoids and background colour on the skin pigmentation of Australian snapper Pagrus auratus (Bloch & Schneider, 1801). Aquaculture Research 39, 1423-1433.
FishBase, 2004. Species profile Carassius auratus Goldfish. #1758
Fujii, R. (2000). The regulation of motile activity in fish chromatophores. Pigment Cell Research 13, 300-319.
Goodwin, T.W. (1986). Metabolism, nutrition, and function of carotenoids. Annual Review of Nutrition 6, 273-297.
Houde, A.E., and Torio, A.J. (1992). Effect of parasitic infection on male color pattern and female choice in guppies. Behavioral Ecology 3, 346-351.
Hu, C.J., Chen, S.M., Pan, C.H., and Huang, C.H. (2006). Effects of dietary vitamin A or beta-carotene concentrations on growth of juvenile hybrid tilapia, Oreochromis niloticus x O-aureus. Aquaculture 253, 602-607.
Jimenez-Martinez, L.D., Alvarez-Gonzalez, C.A., Contreras-Sanchez, W.M., Marquez-Couturier, G., Arias-Rodriguez, L., and Almeida-Madrigal, J.A. (2009). Evaluation of larval growth and survival in mexican mojarra, Cichlasoma urophthalmus, and bay snook, Petenia splendida, under different initial stocking densities. Journal of the World Aquaculture Society 40, 753-761.
Josefa Romero García, Carmen Tatiana H. Kalinowski, María Soledad L. Izquierdo, Lidia Esther R. Robaina. (2010). Marine and freshwater crab meals in diets for red porgy
27
(Pagrus pagrus): effect on growth, fish composition and skin colour. pages 1759–
1769.
Kalinowski, C.T., Izquierdo, M.S., Schuchardt, D., and Robaina, L.E. (2007). Dietary supplementation time with shrimp shell meal on red porgy (Pagrus pagrus) skin colour and carotenoid concentration. Aquaculture 272, 451-457.
Kalinowski, C.T., Robaina, L.E., Fernandez-Palacios, H., Schuchardt, D., and Izquierdo, M.S.
(2005). Effect of different carotenoid sources and their dietary levels on red porgy (Pagrus pagrus) growth and skin colour. Aquaculture 244, 223-231.
Karino, K., and Haijima, Y. (2004). Algal-diet enhances sexual ornament, growth and reproduction in the guppy. Behaviour 141, 585-601.
Kodricbrown, A. (1989). Dietary carotenoids and male mating success in the guppy - an environmental component to female choice. Behavioral Ecology and Sociobiology 25, 393-401.
Kolluru, G.R., Grether, G.F., South, S.H., Dunlop, E., Cardinali, A., Liu, L., and Carapiet, A.
(2006). The effects of carotenoid and food availability on resistance to a naturally occurring parasite (Gyrodactylus turnbulli) in guppies (Poecilia reticulata). Biological Journal of the Linnean Society 89, 301-309.
Kop, A., and Durmaz, Y. (2008). The effect of synthetic and natural pigments on the colour of the cichlids (Cichlasoma severum sp., Heckel 1840). Aquaculture International 16, 117-122.
Liao, W.L., Nureborhan, S.A., Okada, S., Matsui, T., and Yamaguchi, K. (1993).
Pigmentation of cultured black tiger prawn by feeding with a spirulina-supplemented diet. Nippon Suisan Gakkaishi 59, 165-169.
Martinezpalacios, C.A., and Ross, L.G. (1988). The feeding ecology of the central American cichlid Cichlasoma-Urophthalmus (Gunther). Journal of Fish Biology 33, 665-670.
McBride, S.C., Price, R.J., Tom, P.D., Lawrence, J.M., and Lawrence, A.L. (2004).
Comparison of gonad quality factors: color, hardness and resilience, of Strongylocentrotus franciscanus between sea urchins fed prepared feed or algal diets and sea urchins harvested from the Northern California fishery. Aquaculture 233, 405-422.
McDowall, R. M. 2000. The reed field guide to New Zealand freshwater fishes. Auckland, Reed.Reed Books, Auckland. 224 p.
Miki, W. (1991). Biological functions and activities of animal carotenoids Pure Applied Chemistry 63, 141-14.
28
Morgan, D. L., Gill, H. S., Maddern, M. G., Beatty, S. J. 2004. Distribution and impacts of introduced freshwater fishes in Western Australia. New Zealand Journal of Marine and Freshwater Research 38: 511-523.6.
Olsen, R.E., and Baker, R.T.M. (2006). Lutein does not influence flesh astaxanthin pigmentation in the Atlantic salmon (Salmo salar L.). Aquaculture 258, 558-564.
Pérez-Vega, M.H., Uribe-Aranzabal, M.C., García-Lorenzana, M., Romero-Ramírez, M.C., and Arredondo-Figueroa, J.L. (2006). Description of the ovarian follicle growth of the neotropical cichlids Petenia splendida and Parachromis managuensis (Perciformes:
Cichlidae). Journal of Applied Ichthyology 22, 515-520.
Paibulkichakul, C., Piyatiratitivorakul, S., Sorgeloos, P., and Menasveta, P. (2008). Improved maturation of pond-reared, black tiger shrimp (Penaeus monodon) using fish oil and astaxanthin feed supplements. Aquaculture 282, 83-89.
Pan, C.H., and Chien, Y.H. (2009). Effects of dietary supplementation of alga Haematococcus pluvialis (Flotow), synthetic astaxanthin and beta-carotene on survival, growth, and pigment distribution of red devil, Cichlasoma citrinellum (Gunther). Aquaculture Research 40, 871-879.
Pan, C.H., Chien, Y.H., and Wang, Y.J. (2011). Antioxidant defence to ammonia stress of characins (Hyphessobrycon eques Steindachner) fed diets supplemented with carotenoids. Aquaculture Nutrition 17, 258-266.
Parker, R.S. (1996). Carotenoids .4. Absorption, metabolism, and transport of carotenoids.
Faseb Journal 10, 542-551.
Price, A.C., Weadick, C.J., Shim, J., and Rodd, F.H. (2008). Pigments, patterns, and fish behavior. Zebrafish 5, 297-307.
Richardson, M. J., Whoriskey, F. G., Roy, L. H. 1995. Turbidity generation and biological impacts of an exotic fish, Carassius auratus, introduced into shallow seasonally anoxic ponds. Journal of Fish Biology 47(4): 576-585.
Shpigel, M., Schlosser, S.C., Ben-Amotz, A., Lawrence, A.L., and Lawrence, J.M. (2006).
Effects of dietary carotenoid on the gut and the gonad of the sea urchin Paracentrotus lividus. Aquaculture 261, 1269-1280.
Storebakkena, T., Sørensena, M., Bjerkengb, B., and Hiuc, S. (2004). Utilization of astaxanthin from red yeast, Xanthophyllomyces dendrorhous, in rainbow trout, Oncorhynchus mykiss: effects of enzymatic cell wall disruption and feed extrusion temperature. Aquaculture 236.
29
Suckling, C.C., Symonds, R.C., Kelly, M.S., and Young, A.J. (2011). The effect of artificial diets on gonad colour and biomass in the edible sea urchin Psammechinus miliaris.
Aquaculture 318, 335-342.
Sun Xiangjun, Yu Chang, Yuantu Ye, Zhihong Ma, Yongjun Liang, Tieliang Li, Na Jiang, Wei Xing, Lin Luo. (2012). The effect of dietary pigments on the coloration of Japanese ornamental carp (koi, Cyprinus carpio L.). Aquaculture 342-343, 62–68.
Surai, P.F., and Speake, B.K. (1998). Distribution of carotenoids from the yolk to the tissues of the chick embryo. Journal of Nutritional Biochemistry 9, 645-651.
Svensson, P.A., and Wong, B.B.M. (2011). Carotenoid-based signals in behavioural ecology:
a review. Behaviour 148, 131-189.
Tejera, N., Cejas, J.R., Rodriguez, C., Bjerkeng, B., Jerez, S., Bolanos, A., and Lorenzo, A.
(2007). Pigmentation, carotenoids, lipid peroxides and lipid composition of skin of red porgy (Pagrus pagrus) fed diets supplemented with different astaxanthin sources.
Aquaculture 270, 218-230.
Treviño, L., Alvarez-González, C.A., Perales-García, N., Arévalo-Galán, L., Uscanga-Martínez, A., Márquez-Couturier, G., Fernández, I., and Gisbert, E. (2011).
A histological study of the organogenesis of the digestive system in bay snook Petenia splendida Günther, 1862 from hatching to the juvenile stage. Journal of Applied Ichthyology 27, 73-82.
Uscanga-Martinez, A., Perales-Garcia, N., Alvarez-Gonzalez, C.A., Moyano, F.J., Tovar-Ramirez, D., Gisbert, G.E., Marquez-Couturier, G., Contreras-Sanchez, W.M., Arias-Rodriguez, L., and Indy, J.R. (2011). Changes in digestive enzyme activity during initial ontogeny of bay snook Petenia splendida. Fish Physiol Biochem 37, 667-680.
Wang, Y.J., Chien, Y.H., and Pan, C.H. (2006). Effects of dietary supplementation of carotenoids on survival, growth, pigmentation, and antioxidant capacity of characins, Hyphessobrycon callistus. Aquaculture 261, 641-648.
Xu, X. (2006). Effect of astaxanthin from Xanthophyllomyces dendrorhous on pigmentaion of goldfish, Carassius auratus. Journal of the world aquaculture society 37, 282-288.
Yanar, M., Ercen, Z., Hunt, A.O., and Buyukcapar, H.M. (2008). The use of alfalfa, medicago sativa as a natural carotenoid source in diets of goldfish, Carassius auratus.
Aquaculture 284, 196-200.
Yasir, I., and Qin, J.G. (2010). Effect of dietary carotenoids on skin color and pigments of false clownfish, Amphiprion ocellaris, cuvier. Journal of the World Aquaculture Society 41, 308-318.
30
Zagalsky, P.F., Ceccaldi, H.J., and Daumas, R. (1970). Comparative studies on some decapod crustacean carotenoproteins. Comparative Biochemistry and Physiology 34, 579-&.
31
Table 1 Composition and proximate analysis of the experiments diets
1 2 3 4 5 6 7 8 9
control L-5 L-10 L-15 L-20 S-5 s-10 s-15 s-20
Ingredients
Fish meal 25 25 25 25 25 25 25 25 25
Soybean meal 45 45 45 45 45 45 45 45 45
Fish oil 1.2 1 0.8 0.4 0 1.2 1.2 1.2 1.2
Soybean oil 1.8 1.5 1.2 0.6 0 1.8 1.8 1.8 1.8
Vitamin mix 1.4 1.4 1.4 1.4 1.4 1.4 1.4 1.4 1.4 Vitamin E 0.1 0.1 0.1 0.1 0.1 0.1 0.1 0.1 0.1
Mi 5 5 5 5 5 5 5 5 5
starch 12 12 12 12 12 12 12 12 12
cellulose 8 8 8 8 8 8 8 8 8
choline chloride 0.5 0.5 0.5 0.5 0.5 0.5 0.5 0.5 0.5 Pigment addition(mg/kg)
Zeaxanthin 0 0.5 1 2 3
Synthesis Astaxanthin 0.065 0.13 0.195 0.26
Proximate analysis(%)
Crude protein 25.5 25.94 25.94 25.94 24.78 25.19 25.19 25.19 25.19 Crude lipid 11 10.3 10.3 10.3 10.3 9.5 9.5 9.5 9.5
Ash 11.54 11.32 10.58 11.76 10.43 10 10.61 11.54 11.54 Moisture 7.19 7.28 6.28 6.34 8.46 8.21 5.91 7.62 5.97 Carotenoids content(mg/kg) 0.262 0.263 0.263 0.526 1.056 1.842 2.101 2.105 2.368
Inclusion(g/100g diet)
32
Table 2 Growth performance of Petenia splendida fed with experimental diets
Average body weight(g)
Weight gain FCR
Exp.
Group Initial*1 5 weeks*2
7
weeks*3 5weeks 7werks 5weeks 7weeks 1 3.07±1.34 4.28±1.99 4.91±2.35 39.4% 54.2% 18.35% 18.09%
2 2.60±1.28 3.33±1.33 4.01±1.39 28.1% 48.3% 26.57% 19.78%
3 2.50±0.96 3.18±1.06 3.88±1.15 27.2% 49.4% 26.47% 19.56%
4 2.36±1.61 3.37±2.31 4.17±2.58 42.8% 66.5% 17.27% 14.17%
5 2.60±1.04 3.42±1.24 4.04±1.31 31.5% 49.7% 22.68% 19.38%
6 2.90±1.48 3.92±1.96 4.61±2.35 35.2% 52.9% 20.59% 18.42%
7 2.57±1.36 3.65±1.96 4.09±2.19 42.0% 53.9% 17.22% 18.48%
8 2.94±0.98 3.91±1.17 4.97±1.27 33.0% 60.3% 21.96% 15.74%
9 2.70±1.32 3.70±1.71 4.34±1.73 37.0% 54.3% 19.50% 17.51%
*1: Mean ± SD (n=8)
*2: Mean ± SD (n=8)
*3: Mean ± SD (n=8)