Department of Chemistry College of Science
National Taiwan University Master Thesis
Effect of Side Chain Structure of d-Position Residue on Coiled Coil Stability
YI-JEN WENG
Advisor: Richard Ping Cheng, Ph.D.
January, 2013
!
(5 timers at a time) paper
-- --
!
α
d
GCN4 d
(Leu12) IaLd
Es, MR, [L, B1, B5]
d
α
Abstract
Coiled coil is a superhelical twist formed by two to five wrapping α-helices. It is a
common structural motif that can be found in transcription factors, cytoskeletal systems,
contractile systems and etc. These biochemical roles rely on well-defined and stable
structures of coiled coils. Accordingly, the effect of side chain structure of d-position
residue, which is buried in the coiled coil interface, on coiled coil stability was
investigated. GCN4 coiled coil was employed, of which the 2nd d-residue (Leu12) was
substituted with various amino acids. IaLd coiled coils were used to obtain the coiled
coil propensities of these amino acids. Guanidinium denaturation of the coiled coils was
monitored by circular dichroism spectroscopy. Free energy of unfolding was derived
from the guanidinium denaturation data. The hydrophobicities of the amino acids were
measured by thin layer chromatography. Structural parameters Es, MR, [L, B1, B5], and
side chain volume were employed in the analysis. Results show that the shape and size
of the residue side chain contribute more than hydrophobicity to the coiled coil stability
in coiled coils with the d-position residue bearing aliphatic side chains.
Keywords: coiled coil, GCN4, helix propensity, amino acid hydrophobicity, side chain structure
!
Table of Contents
... I ... II ... III Abstract ... IV!
Table of Contents ... V!
List of Figures ... VII!
List of Tables ... IX!
Abbreviations ... X!
Chapter 1 Introduction ... 1!
1.1 Proteins ... 2!
1.2 Protein Structures ... 4!
1.3 Forces in Protein Folding ... 6!
1.4 α-Helix ... 8!
1.5 Coiled coil ... 10!
1.6 Thesis Overview ... 11!
1.7 References ... 12!
Chapter 2 Effect of Side Chain Structure of d-Position Residue on Coiled Coil Stability ... 15!
2.1 Introduction ... 16!
2.1.1 Coiled Coils in Biological Systems ... 16!
2.1.2 GCN4 Coiled Coil Motif ... 17!
2.1.3 Coiled Coil Sequence and Structure ... 18!
2.1.4 Knobs-Into-Holes Interactions of a Dimeric Coiled Coil ... 21!
2.1.5 Coiled Coil Stability ... 21!
2.1.6 Chapter Overview ... 26!
2.2 Results and Discussions ... 27!
2.2.1 Design of GCN4-Derived Peptides ... 27!
2.2.2 Peptide Synthesis of GCN4-Derived Peptides ... 30!
2.2.3 UV-Visible Spectroscopy (UV-vis) of GCN4-Derived Peptides ... 32!
2.2.4 Circular Dichroism (CD) Spectroscopy of GCN4-Derived Peptides .... 33!
2.2.5 Guanidinium Denaturation of GCN4-Derived Peptides ... 37!
2.2.6 ΔGunfold, H2O of GCN4-Derived Peptides ... 43!
2.2.7 Design of IaLd-Derived Peptides ... 47!
2.2.8 Peptide Synthesis of IaLd-Derived Peptides ... 49!
2.2.9 UV-Visible Spectroscopy (UV-vis) of IaLd-Derived Peptides ... 50!
2.2.10 Circular Dichroism (CD) Spectroscopy of IaLd-Derived Peptides ... 50!
2.2.11 Guanidinium Denaturation of IaLd-Derived Peptides ... 53!
2.2.12 ΔGunfold, H2O of IaLd-Derived Peptides ... 57!
2.2.13 Measuring Hydrophobicities of the Amino Acids ... 58!
2.2.14 Size and Shape Parameters ... 59!
2.2.15 Discussion ... 60!
2.3 Conclusion ... 65!
2.4 Acknowledgement ... 65!
2.5 Experimental Section ... 66!
2.5.1 General Materials and Methods ... 66!
2.5.2 Peptide Synthesis ... 70!
2.5.3 UV-Visible Spectroscopy ... 88!
2.5.4 Circular Dichroism Spectroscopy ... 88!
2.5.5 Guanidinium Denaturation ... 89!
2.5.6 Derivation of ΔGunfold, H2O ... 90!
2.5.7 Measurement of Hydrophobicity by Thin Layer Chromatography ... 91!
2.5.8 Calculation of Side Chain Volume ... 91!
2.6 References ... 92!
2.7 Appendix ... 97!
2.7.1 Guanidinium Denaturation Curves of GCN4-Xaa ... 97!
2.7.2 Guanidinium Denaturation Curves of IaLd-Xaa ... 100
2.7.3 NMR Spectrum of Fmoc-Allo Ile-OH ... 102
2.7.4 NMR Spectrum of Fmoc-Cpa-OH ... 103
2.7.5 NMR Spectrum of Fmoc-Nle-OH ... 104
2.7.6 ESI-MS Spectrum of Fmoc-Allo Ile-OH ... 105
2.7.7 ESI-MS Spectrum of Fmoc-Cpa-OH ... 106
2.7.8 ESI-MS Spectrum of Fmoc-Nle-OH ... 107
2.7.9 Elsevier License for Reprint of Figure ... 108
!
List of Figures
Figure 1-1. Dihedral angles φ and ψ ... 3!
Figure 1-2. Ramachandran plot. ... 3!
Figure 1-3. Four levels of protein structure. ... 4!
Figure 1-4. A hydrogen bond. ... 7!
Figure 1-5. The α-helix backbone hydrogen bond. ... 9!
Figure 1-6. Illustration of a coiled coil. ... 11!
Figure 2-1. GCN4-DNA complex. ... 18!
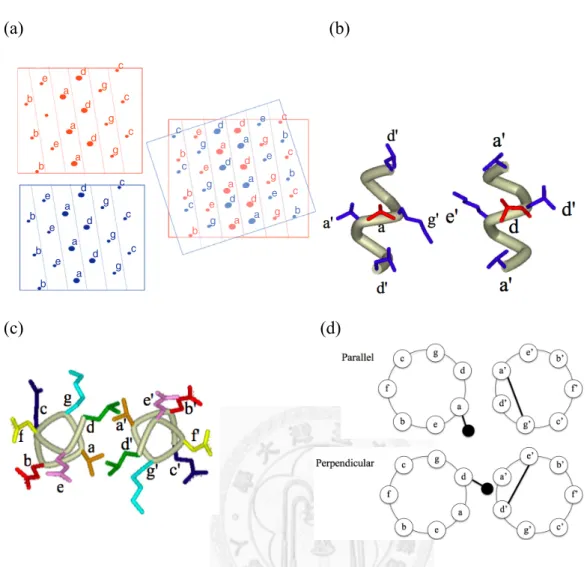
Figure 2-2. Illustration of knobs-into-hole interactions, parallel packing, and perpendicular packing. ... 22!
Figure 2-3. Structures of leucine, trifluoroleucine, and hexafluoroleucine. ... 25!
Figure 2-4. The sequence and the helical wheel diagram of GCN4.. ... 29!
Figure 2-5. CD spectrum of GCN4-Phe2. ... 29!
Figure 2-6. Chemical structures of the amino acids. ... 30!
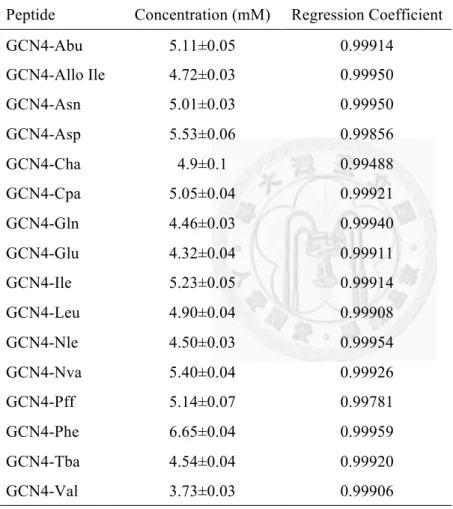
Figure 2-7. CD spectra of GCN4-Xaa peptides at 30 µM peptide concentration in 50 mM phosphate and 150 mM NaCl buffer at pH 7 and 4 °C ... 34!
Figure 2-8. A typical guanidinium titration curve with the proper folded and unfolded baselines depicted. ... 38!
Figure 2-9. Guanidinium denaturation curves for GCN4-Xaa peptides at 30 µM peptide in 50 mM phosphate, 150 mM NaCl, and 0 M to 6 M (with 0.1 M intervals) guanidinium chloride at pH 7 and 4 °C as monitored by CD at 222 nm reported in mean residue ellipticity. ... 39!
Figure 2-10. Fraction unfolded as a function of guanidinium concentration for GCN4-Xaa peptides as derived from the guanidinium denaturation curves. ... 40!
Figure 2-11. The plot of ΔGunfold against guanidinium concentration for GCN4-Tba .. 44!
Figure 2-12. The bar graph of ΔGunfold, H2O of GCN4 peptides. ... 44!
Figure 2-13. ΔGunfold of GCN4-Leu and GCN4-Tba. ... 46!
Figure 2-14. Sequence and helical wheel of IaLd. ... 48!
Figure 2-15. CD spectra of IaLd-Xaa at 20 µM peptide in 10 mM MOPS at pH 7.5 and 25 °C. ... 51!
Figure 2-16. Guanidinium denaturation curves for IaLd-Xaa peptides at 20 µM peptide concentration in 10 mM MOPS with 0 to 6 M (at 0.1 M intervals)
guanidinium chloride at pH 7.5 and 25 °C as monitored by CD at 222 nm reported in mean residue ellipticity ... 55!
Figure 2-17. Fraction unfolded as a function of guanidinium concentration for
IaLd-Xaa peptides as derived from guanidinium denaturation curves. ... 56!
Figure 2-18. The bar graph of ΔGunfold, H2O of IaLd peptides. ... 57!
Figure 2-19. Regressions between the residual energy and log Pow, Es, MR, and side chain volume. ... 62!
!
!
!
List of Tables
Table 2-1. Crude Yield of Peptide Synthesis, Molecular Formula, Calculated [MH+], and Observed m/z of GCN4 Peptides ... 32!
Table 2-2. Concentrations and Regression Coefficients of GCN4 Peptides ... 33!
Table 2-3. [θ]222 at 0 M Guanidinium, Melting Concentrations ([C]m), m values, and ΔGunfold, H2O of GCN4 Peptides ... 35!
Table 2-4. Crude Yield of Peptide Synthesis, Molecular Formula, Calculated [MH+], and Observed m/z of IaLd Peptides ... 49!
Table 2-5. Concentrations and Regression Coefficients of IaLd Peptides ... 50!
Table 2-6. [θ]222 at 0 M Guanidinium, Melting Concentrations ([C]m), m values, and ΔGunfold, H2O of IaLd Peptides ... 52!
Table 2-7. log Pow of the Amino Acids ... 58!
Table 2-8. Es, [L, B1, B5], MR, and the Volume of the Amino Acids Side Chains ... 60!
Table 2-9. Equations and R values of the Regressions Between Residual Energy, log Pow Es, MR, [L, B1, B5], and Side Chain Volume ... 62!
!
Abbreviations
Abu Aminobutyric aicd
Allo Ile Alloisoleucine
Arg Arginine
Asn Asparagine
Asp Aspartate
CaM Calmodulin
CD Circular dichroism
Cha Cyclohexylalanine
Cpa Cyclopentylalanine
DIEA Diisopropylethylamine
DMF Dimethylformamide
DNA Deoxyribonucleic acid
ESI-MS Electrospray ionization mass spectrometry Fmoc N-9-Fluorenylmethoxycarbonyl
Fmoc-Osu N-(9-Fluorenylmethoxycarbonyloxy) succinimide
Gln Glutamine
Glu Glutamate
Gly Glycine
HBTU O-1H-benzotriazol-1-yl-1,1,3,3-tetramethyluronium hexafluoro phosphate
Hfl Hexafluoroleucine
His Histidine
HOBt 1-Hydroxybenzotriazole
Ile Isoleucine
KIH Knobs-into-holes
Leu Leucine
Lys Lysine
MALDI-TOF MS Matrix-assisted laser desorption ionization time-of-flight
Met Methionine
MOPS 3-(N-morpholino)propanesulfonic acid
MR Molecular refractivity
MRE Mean residue ellipticity
MW Molecular weight
!
Nle Norleucine
NMR Nuclear magnetic resonance
Nva Norvaline
Pff Pentafluorophenylalanine
Phe Phenylalanine
QSAR Quantitative structure–activity relationship
RNA Ribonucleic aicd
RP-HPLC Reverse phase-high performance liquid chromatography
Ser Serine
SPPS Solid phase peptide synthesis
Tba tert-Butylalanine
TFA Trifluoroacetic acid
Tfl Trifluoroleucine
THF Tetrahydrofuran
Tyr Tyrosine
TLC Thin layer chromatography
UV-vis Ultraviolet-visible
Val Valine
VBP Vitellogenin binding protein
Chapter 1 Introduction
!
Proteins
Proteins are one of the most versatile and most abundant macromolecules in living
systems.1, 2 Proteins serve as the downstream terminus of the central dogma of
molecular biology, which describes the direction of the flow of genetic information.3
The genetic information stored in deoxyribonucleic acid (DNA) is passed down to
ribonucleic acid (RNA) by transcription.4 RNA then serves as the template in the
translation process, leading to the synthesis of proteins.5 These proteins then participate
in a variety of physiological processes, such as catalysis,6 immune response,7 signal
transduction,8 and construction and support of cellular structures.9 Proteins are
important to living systems, and studies to enhance fundamental knowledge on proteins
should facilitate our understanding of how Nature functions, and also lead to
unprecedented technological advancements.
A protein is a biopolymer composed of amino acids.2 There are twenty naturally
occurring amino acids, nineteen of which are L-α amino acids bearing different side
chains. These side chains have various functional groups and properties, and play
different roles in protein functions. The remaining amino acid is glycine, which has no
side chain and is therefore achiral. Amino acids are connected by an amide bond
! 3!
between the α-amino group of one amino acid and the α-carbonyl group of the other,
yielding a polypeptide chain. Due to resonance, the amide bond has considerable double
bond character, limiting bond rotation and therefore restricting the conformation of the
peptide backbone. Dihedral angles are defined along the peptide backbone. The dihedral
angle between Ccarbonyl - N plane and Cα - Ccarbonyl plane is defined as phi (φ), and the
dihedral angle between N - Cα plane and Ccarbonyl - N plane is defined as psi (ψ) (Figure
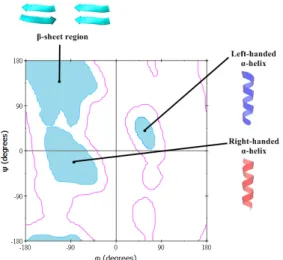
1-1).10 A plot of ψ against φ is called Ramachandran plot (Figure 1-2).10 The shaded
region in the plot shows the distribution of φ and ψ found in several structures (Figure
1-2).10
Figure 1-1. Dihedral angles φ and ψ.
Figure 1-2. Ramachandran plot.10
NH O
R ψ
φ
Figure 1-4. Ramachandran plot for various secondary structures.19 Protein Secondary Structure
The polypeptide chain can form local regularly repeating structures called secondary structure. These structures are formed by hydrogen bonds between NH and carbonyl groups in the backbone. Common secondary structure motifs include α-helix, β-sheet, β-turn, and Ω-loop.18, 20-24 An α-helix is a right-handed rod-like structure.18 The left-handed helix is very rare due to the unfavorable steric clash between the side chains and the backbone.18 A β-sheet is a relatively flat with some right-handed twist structure.22 A β-sheet is composed of two or more β strands in either antiparallel or parallel orientation.22 The compact and globular shape of most proteins can be attributed to the presence of reverse turns (aka: β-turn, hairpin bends) which promotes the chain reversal of polypeptide chain.24, 25 Reverse turns are stabilized by hydrogen bonds between the carbonyl group of residue i and the NH of residue i+3.
!
Protein Structures
The structure of a protein is critical for its biological function. A protein performs its
function only when it is correctly folded.11 Upon losing its structure, for instance, being
denatured by heat or chemical denaturant, a protein becomes inactive.12 The structure of
a protein is categorized into four hierarchical levels: primary, secondary, tertiary, and
quaternary structures (Figure 1-3). Primary structure refers solely to the amino acid
sequence of the protein, without consideration of the three dimensional structures of the
polypeptide chain.13
Figure 1-3. Four levels of protein structure. (Tertiary structure: calmodulin, protein data bank code 1CLL. Quaternary structure: fibrinogen, protein data bank code 3GHG.)
Secondary structures are local structural motifs composed of regularly repeating
spatial arrangement of the residues. Secondary structures can be defined by the pattern
of hydrogen bonds or backbone dihedral angles of the peptide chain.14, 15 For example,
α-helix, a common secondary structure, is characterized by the main-chain hydrogen
bond between the carbonyl oxygen and the amide hydrogen located four residues away Primary
LEDKVEE
Secondary
2ZTA
Page 1 of 1 Printed at 23:53:54 on Thu Mar 1 2012
Tertiary
1CLL
Page 1 of 1 Printed at 00:01:19 on Fri Mar 2 2012
Quaternary
in the sequence.16 The dihedral angles (φ, ψ) of an α-helix is defined to be (-57°, -47°)
(Figure 1-2).10 Another common secondary structure, β-sheet, is made of two or more β
strands that are connected by hydrogen bonds between backbone carbonyl oxygen and
the amide hydrogen.17 The dihedral angles (φ, ψ) of a β-sheet are (-119°, +113°) for
parallel sheet and (-139°, +135°) for antiparallel sheet.10
Tertiary structure refers to the overall structure of one polypeptide chain, containing a
combination of several secondary structures. A typical tertiary structure has nonpolar
residues buried in the interior, making the hydrophobic core.18 Polar and charged
residues are usually found on the surface, where the protein is in contact with the
aqueous surrounding.18 Calmodulin (CaM) is an example of tertiary structure, which is
a protein that binds calcium ion and evokes downstream biochemical reactions. CaM is
a 148-amino-acid polypeptide chain made up of several α-helices and turns.19
Quaternary structure is the spatial arrangement of several polypeptide chains. One
example is the fibrinogen, a protein found in blood and is involved in blood coagulation.
Fibrinogen consists of six chains, (αβγ)2, in which α, β, and γ are the differernt
subunits.20
!
Forces in Protein Folding
The forces that dictate protein folding include electrostatic interactions, hydrogen bond,
hydrophobic effect, and van der Waal’s interactions.21 The first two occur in charged
and polar groups, whereas the last two are important for nonpolar entities. Electrostatics
include charge to charge, charge to dipole, and dipole to dipole interactions. The
charges originate from the following amino acids: aspartic acid (Asp), glutamic acid
(Glu), lysine (Lys), arginine (Arg), and to a much lesser content, histidine (His). At
physiological pH (pH 7.4), side chains of Asp and Glu are deprotonated and are
negatively charged. On the other hand, Lys, Arg, and His are protonated and bear a
positive charge. Opposite charges attract one another, resulting charge-charge
interactions between these amino acids. A dipole is the asymmetric distribution of
election density along a chemical bond caused by the difference in electronegativity of
atoms. Dipoles can be attracted by ions or other dipoles on the protein, leading to
dipole-charge and dipole-dipole interactions. Electrostatic interactions may provide
extra stability to protein structures, as in the case of hyperthermophilic proteins.22 These
proteins are able to withstand high temperatures, and this stability originates from the
numerous electrostatic interactions between the charged residues on the protein
surface.22
A hydrogen bond is the consequence of the interaction between a hydrogen bond
donor (HX) and a hydrogen bond receptor (Y) (Figure 1-4). The atoms X and Y could
be nitrogen, oxygen, or fluorine atoms, which are all highly electronegative atoms. The
electronegativity of X causes uneven distribution of electron density along the H-X
bond, resulting in a partial positive charge on H. This charge is attracted to the
nonbonding electrons of Y, therefore forming a hydrogen bond. Hydrogen bond is the
dominant force for the formation of secondary structures. The deprival of these
hydrogen bonds (e.g., using solvents like trifluoroacetic acid or formic acid, which
protonate the peptide backbone) diminishes peptide structure.23, 24
Figure 1-4. A hydrogen bond.
Hydrophobic effect is the release of water molecules around a hydrophobic group
when two or more such groups approach one another.25 Water molecules that are in
contact with nonpolar solutes are, to some extent, orderly arranged.25 When the
nonpolar groups come into contact with one another, these ordered water molecules are
25
X H Y
δ
−δ
−δ
+!
example of the importance of the hydrophobic effect in protein folding is the coiled coil
motif. A coiled coil is composed of intertwining α-helices.26 The residues in helices
interface are mainly hydrophobic.26, 27 Substituting the hydrophobic residues with polar
residues destablizes the coiled coil.27
van der Waal's interaction (or London dispersion force) is the attractive force
between apolar molecules.21 The electron density distribution of the atoms in apolar
group may fluctuate, generating an instantaneous dipole. This dipole may induce an
opposite dipole in a nearby molecular entity. The attraction between these dipoles is
termed London dispersion force. This is a weak interaction and is significant only when
the apolar groups are close in distance. The huge number of London dispersion forces in
proteins make them an important driving force in protein folding.
α-Helix
α-Helix is a common secondary structure that constitutes about 30% of all protein
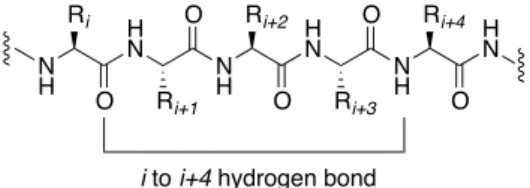
structures known to date.15, 28 The naturally occurring α-helix is a right-handed, rod-like
structure characterized by consecutive, main-chain, i i + 4 hydrogen bonds between
each carbonyl oxygen (i) and an amide hydrogen (i+4) on the adjacent helical turn
(Figure 1-5).16 A helix has backbone dihedral angles (φ, ψ) around (-57°, -47°) and has
3.6 residues per turn.10, 16 Many factors determine the stability of a helix, one of which
is the helix propensity of the constituting amino acids. Chou and Fasman analyzed the
frequency of appearance of each amino acid in helices.15 The statistics revealed that
different amino acids have different probabilities to be found in a helix. The
thermodynamic tendency to form a helix can be described in a statistical mechanical
manner in Lifson-Roig theory, in which helix propensity (w) of an amino acid is
employed to describe the equilibrium between α-helix and random coil.29 The greater
the helix propensity, the greater the probability to find an amino acid in the helical state.
Helices are often combined with other secondary structures to form motifs, which
usually exhibit certain functions.1 For example, the helix-turn-helix motif is a DNA
binding motif,30 and the helix-loop-helix ("EF hand") is a calcium ion binding motif.19
Figure 1-5. The α-helix backbone hydrogen bond.
NH HN
O
NH
O H
N O
O NH
O HN
Ri Ri+2 Ri+4
Ri+1 Ri+3
i to i+4 hydrogen bond
!
Coiled coil
Coiled coils are formed by 3-5% of all amino acids in proteins,31 and can be found in
transcription factors,32 cytoskeletal33 and contractile34 systems, viral envelope
proteins,35 and other systems.36 A coiled coil is formed by two to five α-helices
wrapping around each other to form a slight left-handed superhelical twist (Figure
1-6).26 The sequence of a coiled coil is characterized by the “heptad repeat”, abcdefg, in
which positions a and d are predominantly hydrophobic residues, such as leucine (Leu)
and isoleucine (Ile), leading to a hydrophobic interface between two wrapping helices.
In contrast, the e and g positions are frequently charged residues that form interhelical
ion pairs.26 The three dimensional architecture was first proposed by Crick in 1953 as
the “knobs into holes” model, which states that a coiled coil is stabilized by the packing
of hydrophobic side chains “knobs” into the “holes” that are the space between
hydrophobic side chains of the neighboring helix.37 According to the type and
orientation of the monomers, coiled coils can be further categorized into
homodimer/heterodimer and parallel/antiparallel coiled coils.38 A homodimer is made
up of two identical monomers, whereas a heterodimer consists of two different
monomers. A parallel coiled coil has the N-termini of the two helices located on the
!
same end, whereas an antiparallel coiled coil has the two N-termini located at opposite
ends.
(a) (b)
Figure 1-6. Illustration of a coiled coil. (a) A coiled coil. (GCN4 leucine zipper. Protein data bank code 2ZTA.) (b) The helical wheel of a coiled coil.
Thesis Overview
Coiled coil is a common structure in proteins. The factors contributing to coiled coil
stability include hydrophobicity of the coiled coil interface, coiled coil propensity of the
constituting amino acids, and the packing (or the van der Waal's interacitons) of the
residues at the interface.39 Studies involving natural amino acids have shown that
geometric properties of the buried amino acids can influence the stability of coiled
coils.39, 40 To further elucidate the relationship between structure, hydrophobicity, and
coiled coil stability, natural and non-natural amino acids with various side chain
structures and properties were introduced into different coiled coil systems. Also, the
hydrophobicities of the amino acids were measured. Several structural parameters were
Page 1 of 1 Printed at 23:44:37 on Thu Mar 1 2012
!
Reference
1.! Berg, J. M.; Tymoczko, J. L.; Stryer, L. Biochemistry 6th ed., W.H. Freeman:
New York, 2006.
2.! Nelson, D.; Cox, M. Lehninger Principles of Biochemistry 5th ed, W. H. Freeman:
New York, 2009.
3.! Crick, F. Central Dogma of Molecular Biology. Nature 1970, 227, 561-563.
4.! Conaway, J. W.; Shilatifard, A.; Dvir, A.; Conaway, R. C. Control of elongation by RNA polymerase II. Trends Biochem. Sci. 2000, 25, 375-380.
5.! Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 2002, 108, 557-572.
6.! Radzicka, A.; Wolfenden, R. A proficient enzyme. Science 1995, 267, 90-93.
7.! Aderem, A.; Ulevitch, R. J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782-787.
8.! Nishizuka, Y. The role of protein kinase C in cell surface signal transduction and tumour promotion. Nature 1984, 308, 693-698.
9.! Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509-514.
10.! Edsall, J. T.; Flory, P. J.; Kendrew, J. C.; Liquori, A. M.; Nemethy, G.;
Ramachandran, G. N.; Scheraga, H. A. A proposal of standard conventions and nomenclature for the description of polypeptide conformations. J. Mol. Biol. 1966, 15, 399-407.
11.! Wright, P. E.; Dyson, H. J. Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. J. Mol. Biol. 1999, 293, 321-331.
12.! Tanford, C. Protein denaturation. Adv. Protein Chem. 1968, 23, 121-282.
13.! Sanger, F. The arrangement of amino acids in proteins. Adv. Protein Chem. 1952, 7, 1-67.
14.! Ramachandran, G. N.; Ramakrishnan, C.; Sasisekharan, V. Stereochemistry of polypeptide chain configurations. J. Mol. Biol. 1963, 7, 95-99.
15.! Chou, P. Y.; Fasman, G. D. Conformational parameters for amino acids in helical, beta-sheet, and random coil regions calculated from proteins. Biochemistry 1974, 13, 211-222.
16.! Pauling, L.; Corey, R. B. The structure of synthetic polypeptides. Proc. Natl. Acad.
Sci. U. S. A. 1951, 37, 241-250.
17.! Pauling, L.; Corey, R. B. The pleated sheet, a new layer configuration of
polypeptide chains. Proc. Natl. Acad. Sci. U. S. A. 1951, 37, 251-256.
18.! Pace, C. N.; Shirley, B. A.; McNutt, M.; Gajiwala, K. Forces contributing to the conformational stability of proteins. FASEB J. 1996, 10, 75-83.
19.! Chattopadhyaya, R.; Meador, W. E.; Means, A. R.; Quiocho, F. A. Calmodulin structure refined at 1.7 A resolution. J. Mol. Biol. 1992, 228, 1177-1192.
20.! Kollman, J. M.; Pandi, L.; Sawaya, M. R.; Riley, M.; Doolittle, R. F. Crystal structure of human fibrinogen. Biochemistry 2009, 48, 3877-3886.
21.! Dill, K. A. Dominant forces in protein folding. Biochemistry 1990, 29, 7133-7155.
22.! Xiao, L.; Honig, B. Electrostatic contributions to the stability of hyperthermophilic proteins. J. Mol. Biol. 1999, 289, 1435-1444.
23.! Hanlon, S.; Klotz, I. M. Protonation of polypeptides in "helix-breaking" solvents:
spectral and optical-rotatory properties in solutions containing strong organic acids. Biochemistry 1965, 4, 37-48.
24.! Lotan, N.; Bixon, M.; Berger, A. Alpha-helix formation by solvent-solvent interaction. Biopolymers 1967, 5, 69-77.
25.! Southhall, N. T.; Dill, K. A.; Haymet, A. D. A View of the Hydrophobic Effect. J.
Phys. Chem. B 2002, 106, 521-533.
26.! Mason, J. M.; Arndt, K. M. Coiled coil domains: Stability, specificity, and biological implications. ChemBioChem 2004, 5, 170-176.
27.! Gonzalez, L., Jr.; Woolfson, D. N.; Alber, T. Buried polar residues and structural specificity in the GCN4 leucine zipper. Nat. Struct. Biol. 1996, 3, 1011-1018.
28.! Cheng, R. P.; Girinath, P.; Suzuki, Y.; Kuo, H. T.; Hsu, H. C.; Wang, W. R.; Yang, P. A.; Gullickson, D.; Wu, C. H.; Koyack, M. J.; Chiu, H. P.; Weng, Y. J.; Hart, P.;
Kokona, B.; Fairman, R.; Lin, T. E.; Barrett, O. Positional effects on helical Ala-based peptides. Biochemistry 2010, 49, 9372-9384.
29.! Lifson, S.; Roig, A. On the theory of helix - coil transition in polypeptides. J.
Chem. Phys. 1961, 34, 1963-1974.
30.! Brennan, R. G.; Matthews, B. W. The helix-turn-helix DNA-binding motif. J.
Biol. Chem. 1989, 264, 1903-1906.
31.! Wolf, E.; Kim, P. S.; Berger, B. MultiCoil: A program for predicting two- and three-stranded coiled coils. Protein Sci. 1997, 6, 1179-1189.
32.! O'Shea, E. K.; Klemm, J. D.; Kim, P. S.; Alber, T. X-ray structure of the GCN4 leucine zipper, a two-stranded, parallel coiled coil. Science 1991, 254, 539-544.
!
33.! Pauling, L.; Corey, R. B. Compound helical configurations of polypeptide chains:
structure of proteins of the alpha-keratin type. Nature 1953, 171, 59-61.
34.! Sodek, J.; Hodges, R. S.; Smillie, L. B.; Jurasek, L. Amino-acid sequence of rabbit skeletal tropomyosin and its coiled-coil structure. Proc. Natl. Acad. Sci. U. S. A.
1972, 69, 3800-3804.
35.! Wilson, I. A.; Skehel, J. J.; Wiley, D. C. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 A resolution. Nature 1981, 289, 366-373.
36.! Cohen, C.; Parry, D. A. Alpha-helical coiled coils: more facts and better predictions. Science 1994, 263, 488-489.
37.! Crick, F. H. C. The Packing of Alpha-Helices: Simple Coiled-Coils. Acta Cryst.
1953, 6, 689-697.
38.! Grigoryan, G.; Keating, A. E. Structural specificity in coiled-coil interactions. Curr.
Opin. Struct. Biol. 2008, 18, 477-483.
39.! Moitra, J.; Szilak, L.; Krylov, D.; Vinson, C. Leucine is the most stabilizing aliphatic amino acid in the d position of a dimeric leucine zipper coiled coil.
Biochemistry 1997, 36, 12567-12573.
40.! Harbury, P. B.; Zhang, T.; Kim, P. S.; Alber, T. A switch between two-, three-, and four-stranded coiled coils in GCN4 leucine zipper mutants. Science 1993, 262, 1401-1407.
!
Chapter 2 Effect of Side Chain Structure of d-Position
Residue on Coiled Coil Stability
!
Introduction
Coiled Coils in Biological Systems
Coiled coils are formed by 3-5% of all amino acids in proteins,1 and can be found in
transcription factors2, cytoskeletal3 and contractile4 systems, viral envelope proteins,5
and other systems.6 In particular, transcription factors are an excellent example to
illustrate the importance of coiled coils. Transcription factors are proteins that bind
specific DNA sequences and thereby regulate gene expression.2 Among the numerous
transcription factors, bZIP transcription factors constitute an important class of DNA
binding proteins.7, 8 bZIP transcription factors achieve specific DNA binding through
the dimerization of the coiled coil motifs.7, 8 Such dimerization results in the formation
of various homodimers and heterodimers, which enhance specific binding between the
proteins and DNA, generate a large number of variations for binding of different DNA
sequences, and therefore form a complex regulatory network in a variety of organisms.7,
8 This suggests an important role for coiled coils in living systems, and studies of this
structural motif should facilitate the understanding of protein functions and the design
of novel proteins.
GCN4 Coiled Coil Motif
GCN4 is a bZIP transcription factor in yeast. GCN4 contains a basic region and a
coiled coil region.9 The former is responsible for DNA recognition, and the latter
promotes dimerization (Figure 2-1).9 The crystal structure of the coiled coil region of
GCN4 was first solved in 1991.2 This achievement has turned coiled coils, which was
mainly considered to be structural for decades because coiled coils were first recognized
in fibrous proteins, into a popular research topic that has blossomed in the past twenty
years.6, 10, 11 There were two significances. First, it showed that coiled coils participate
in crucial interactions such as transcription. Second, the knowledge obtained from the
physical details of the coiled coil structure provided a means to recognize tertiary
structures by the inspection of primary structures, leading to de novo coiled coil design.6,
10, 11 The coiled coil region of GCN4 is a 33-residue, 4-heptad repeat leucine zipper (see
Chapter 1 for the general introduction of coiled coils). Leucine zipper bears its name
due to the conserved leucine at the d positions. The a positions are taken up by valine,
so that both a and d!positions are occupied by hydrophobic residues. One exception
occurs at the a position in the third heptad, where there is an asparagine.2 This buried
polar residue determines the oligomeric state of the coiled coil, making GCN4 a
!
homodimeric parallel coiled coil.12
Figure 2-1. GCN4-DNA complex (protein data bank code 1YSA).
Coiled Coil Sequence and Structure
The sequence of a coiled coil is characterized by the “heptad repeat”, usually
designated abcdefg. A regular α-helical turn is composed of 3.6 residues.10 However,
this value is lowered to 3.5 for helices in a coiled coil, leading to the heptad repeat every
two turns of a helix.10 Inspection of coiled coil sequences led to the concept of "peptide
Velcro", which points out three elements for the formation of a specific coiled coil.10, 13
First, the residues at a and d positions, which are located at the interface between a
coiled coil, need to be hydrophobic residues like leucine, valine, or isoleucine, so that
the coiled coil is stabilized through hydrophobics and van der Waals interactions.
Second, the e and g positions, which are located adjacent to the coiled coil interface,
have to be occupied by charge residues such as glutamate or lysine, which form
interhelical electrostatic interactions that also contribute to coiled coil stability. Last, the
1YSA
Page 1 of 1 Printed at 01:27:10 on Thu Apr 12 2012
remaining b, c, and f positions have to be hydrophilic residues, since these positions are
located at the solvent-accessing face of a coiled coil.
These features are insufficient to conclude the variety of coiled coils observed in
Nature and laboratory. More precisely, the features are insufficient to predict the
oligomerization (dimer, trimer, or tetramer, etc.), pairing specificity (homodimer or
heterodimer), and orientation (parallel or antiparallel). 20% of the residues at the a and
d positions are polar or charged.12 Buried polar residues affect the oligomeric state of a
coiled coil.10, 12, 14-16 For instance, the substitution of the two buried asparagines of
GCN4 coiled coil to valine resulted in change in oligomeric state from dimers to a
mixture of dimers and trimers.17, 18 These asparagines confer the dimer state through
interhelical hydrogen bonds between the Asn side chains.2, 10, 17 Besides, the structure of
the a and d residues also have affect on oligomerization.18 GCN4 coiled coil mutants,
with the 4 a residues replaced with isoleucine, valine, or leucine yielded dimers,
mixture of dimers and trimers, and trimers, respectively.18 This switch between
oligomeric states is a consequence of the packing interactions at the coiled coil
interface.18
Orientation and pairing specificity are greatly influenced by the electrostatic
!
interactions between the e and g residues.10, 19 For instance, placing only negatively
charged residue at the e and g positions in one strand of a coiled coil and only positively
charged residues at the e' and g' positions in another strand, favors the formation of a
heterodimer due to attractive interhelical Coulombic interactions.13 Also, deliberate
positioning of oppositely charged residues at g and g' led to the preference for the
antiparallel orientation as a result of the repulsive Coulombic interactions that would be
encountered if the coiled coil was parallel.20
There are still many undiscovered underlying rules for the structures of coiled coils.
Amazing examples include a GCN4 variant with all e positions mutated to valine gave a
parallel tetramer,21 whereas another GCN4 variant with all g residues substituted with
either valine or alanine resulted in an antiparallel tetramer.21 Still, when all e and g
residues of GCN4 were replaced with alanine, the coiled coil turned out to be a parallel
heptamer.21 Predicting the oligomeric state, pairing specificity, and orientation remains
an intriguing challenge.
Knobs-Into-Holes Interactions of a Dimeric Coiled Coil
Knobs-into-holes (KIH) interactions of a dimeric coiled coil was first described by
Crick in 1953.22 KIH states that a side chain from one helix, referred to as the knob, fits
into the hole surrounded by the side chains of the partner helix.22 For instance, in a
parallel, dimeric coiled coil, the a knob fits into a hole made by d', a', d', and g' of the
neighboring helix, whereas the d knob fits into a hole made by a', e', a', and d' residues
(Figure 2-2a and b).2, 11, 18 Furthermore, the a knob projects out of the interface; the
Cα-Cβ vector of the knob is parallel to the Cα-Cα vector at the bottom of the recipient
hole.2, 11, 18 The d knob projects directly into the interface; the Cα-Cβ vector of the knob
is perpendicular to the Cα-Cα vector at the bottom of the hole (Figure 2-2c and d).2, 11,
18
Coiled Coil Stability
Many factors contribute to the stability of a coiled coil, such as the coiled coil
propensities of the constituting amino acids,23 hydrophobic interactions,10, 12, 18, 24, 25
packing of the core residues,18, 26-30 and electrostatic interactions.10, 31, 32 Herein, coiled
coil propensities, hydrophobic interactions, and side chain packing will be discussed.
Amino acids have different conformational preferences that influence the stability of
!
(a) (b)
(c) (d)
Figure 2-2. Illustration of knobs-into-hole interactions, parallel packing and
perpendicular packing. (a) Cartoons showing knobs-into-holes interactions. On the left hand side are the projections of each helix strand in a coiled coil onto flat surfaces. On the right is the overlap of the two sheets which reflects the interdigitation of side chains in coiled coil. The angle between the two sheets is the angle between the two strands of helices, which is ~20°. (Reprinted from Woolfson et al. New currency for old rope:
from coiled-coil assemblies to α-helical barrels. Curr. Opin. Struct. Biol. 2012,
doi:10.1016/j.sbi.2012.03.002, with permission from Elsevier.) (b) KIH interactions in GCN4 coiled coil. On the left shows the a knob and the corresponding d'a'd'g' hole, the other is the d knob and the a'e'a'd' hole. (c) One heptad of GCN4 coiled coil. The a knob projects out of the interface, whereas the d knob projects into the interface. (d) Cartoons showing the paralle and perpendicular packing of the a and d knobs. Open circles represent Cα atoms, and solid circles represent Cβ atoms. (GCN4 PDB ID:
2ZTA.)
4 Engineeringanddesign COSTBI-976;NO.OFPAGES10
Figure2
(a)
b
b b b b b b
b e
a
a a
a a
a
d a
d
d d d d d d d d d d d d d d d d d d d d d d d
d c
c
c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c
c c
g g
g gggggggg g
g g g g g g g g g
g g g g g g g g g e e e
e e
b
b b
b b b b b e
e e e e e e e e e e e e e e e e e e
e a
a
a a a a a a a a
a d
d
a
a
a
g d
d d
d d d d d d d d d
d d
c
c c c c c c c c c c c c c c c c
c g
g g e
e
(d)
Type N (xHxxHxx)
Type I (HHxxHxx)
Type II (HHxxHHxx)
Type III (xHxHHxH)
(b) (c)
a = parallel
d = parallel a,d = acute
d = perpendicular
a = perpendicular
c g
f
b e
b f
e c
a d
g
51º
d a
c g
f
b e
d d
b
a e
g
c f
a
c g
b e
f c
103º
154º
a d
g e b
a
d g
f c f
b e
b e
f d
c g
a a
e b
f c g
g d a
e c
f b d
d a
a e
d b
f g c
Current Opinion in Structural Biology
Knobs-into-holes(KIH)interactions.(a)Crick’soriginalhelical-netconcept,whichledtotheideaofKIHpacking.Ontheleft-handside,theheptad repeatsfortwohelices(colouredredandblue)areprojectedontoflatsurfaces.Theseillustratehowthehydrophobica/dseamswindaroundthe helicesintheoppositesensetothatofaregular,right-handeda-helix.Ontheright-handside,thetwonetsareinterlacedtoshowhoweachresidueon
CurrentOpinioninStructuralBiology2012,22:1–10 www.sciencedirect.com
protein secondary and tertiary structure.23, 33 "Helix propensity" describes the
probability of an amino acid to be in a helical state.34 Many studies use monomeric
model helices to measure helix propensities.23 O'Neil et al. employed a
parallel, homodimeric coiled coil instead.23 They claimed that a monodimeric helix may
may not be representative of helices found in a protein because a helix in a protein
experiences non-uniform solvent accessibility and dielectric constant.23 The guest site,
which was substituted with the 20 natural amino acids, was situated at the f position of
the middle heptad. The f position is distant from the dimerization surface, therefore
interhelical packing interactions are avoided. Residues near the guest site were
designated to be small and neutral in order to exclude possible interactions between the
guest residue and nearby residues. As such, differences in coiled coil stabilities solely
depended on the different preferences for the coiled coil conformation of each guest
residue. In other words, this system measures the coiled coil propensity. The result
showed an overall good correlation with data obtained from various monomeric helix
models.23
Hydrophobic interactions arise from the exclusion of surrounding orderly-arranged
water molecules when hydrophobic moieties approach one another.35 The release of
!
water molecules leads to the increase in entropy and decrease in free energy.35
Hydrophobicity is quantified by measuring the partition coefficient of a solute between
water and a non-polar solvent, which is further transformed into transfer free energy.35
Hydrophobic interaction is one of the main determinants for coiled coil dimerizaiton.10
A great deal of work has shown that decrease in the hydrophobicity of the coiled coil
interface decreases coiled coil stability. For instance, the substitution of the buried
asparagine in GCN4 to valine increases the melting temperature (the temperature at
which 50% of the coiled coils are unfolded) by 40°C.18 The gain in stability is at the
expense of the specificity of oligomeric state (vide supra).10, 18 In another study, the
a-position valines and d-position leucines in GCN4 were replaced with asparagine,
threonine, serine, and glutamine, and all of the mutants exhibited lower melting
temperatures.12 Instead of substituting with polar entities, residues with higher
hydrophobicity were also employed, and coiled coil stability was enhanced. When the
d-position leucines in coiled coil A1 were replaced with trifluoroleucine or
hexafluoroleucine (Figure 2-3), the melting temperatures increased by 10 °C and 22 °C
respectively. The increase in thermal stability was a consequence of the superior
hydrophobicity of trifluoromethyl over methyl groups.24, 25
Figure 2-3. Structures of leucine, trifluoroleucine and hexafluoroleucine.
The hydrophobic effect is generally agreed to be one of the major determinant factors
in protein folding and stability,26, 35 but packing of the residue side chains also play a
significant role in protein stability.26, 30 In a typical experiment, a hydrophobic residue
in the core of a protein was substituted with a smaller hydrophobic residue (for example,
leucine to alanine), and the difference between the folding free energies of both the
mutant and wild-type protein was measured.26 This energy difference reflected the
different stabilities of the mutant and wild-type protein.26 Studies revealed that the
energy difference was larger than what would be expected for the transfer free energy,
which is a measure of hydrophobicity.26 The change in protein stability, in this case,
was not caused by hydrophobicity alone, and the discrepancy can be explained by the
difference in packing efficiency.26, 27 The formation of cavities within a protein is
destabilizing.26, 30 In coiled coils, the geometric properties of buried amino acids
influenced the overall structure and stability.18, 27 For example, substituting the 5th
H2N OH O
H2N OH O CF3
H2N OH O CF3 F3C
Leucine Trifluoroleucine (2S, 4R)-Tfl
Hexafluoroleucine (Hfl)
!
isoleucine and valine lowers the thermal stability, even though these amino acids are of
similar sizes.27 Leucine is the most favored amino acid at the d position, and is nearly 3
kcal/mole more stabilizing than the similarly sized amino acid isoleucine.27 β-Branched
amino acids are conformationally constrained in α-helices.27 Molecular modeling
indicates that at d positions, the favored rotamers of isoleucine and valine, which are
both β-branched, produce a steric clash between the two amino acids on opposite
helices.27 Recent studies from Gellman's group shows that a'-a-a' vertical interactions
are more prominent than d'-d-d' in antiparallel dimeric coiled coil.28, 29 Coiled coil
database mining reveals that the Cα-Cβ vectors of a and a' residues points together and
their trajectories appear to cross, leading to steric repulsion between side chains.29 On
the other hand, Cα-Cβ vectors of d and d' residues are nearly parallel but offset, and the
steric repulsion is avoided.29
Chapter Overview
Coiled coil stability has been intensively investigated over the past two decades, and
the hydrophobicity at the coiled coil interface is the most recognized factor that
determines the stability of this structural motif. Although packing interactions are
known to affect the stability of proteins and coiled coils, little has been carried out to
probe the relationship between packing and stability. Furthermore, most of these studies
only employed natural amino acids. To further elucidate the relationship between
hydrophobicity, side chain packing, and stability, both natural and non-natural amino
acids with various side chain structures and hydrophobicities were introduced into two
coiled coil systems.
Results and Discussions
Design of GCN4-Derived Peptides
The GCN4 coiled coil region is a well-studied model system for coiled coils,
therefore we chose to study the effect of altering the residues at the d position in this
particular system. There are 4 a positions and 4 d positions in a GCN4 coiled coil
(Figure 2-4a). Initially, all 4 d residues were mutated simultaneously (previously
performed by Hsien-Po Chiu in our lab), but some of these mutants exhibited no coiled
coil conformation by circular dichroism (CD) spectroscopy. This was probably due to
the profound changes in the interface constituents. Therefore, mutation sites were
narrowed down to the central 2 d positions. Still, the second-generation mutant did not
!
exhibit a coiled coil conformation (Figure 2-5). To enable significant formation of the
coiled coil conformation, only one of the two central d sites (positions 12 and 19) could
be mutated. The GCN4 coiled coil crystal structure showed that both sites were
positioned at the same distance from the center of the coiled coil (Figure 2-4b). In
addition, the side chain of a residue in an α!helix points towards the N-terminus. For
position 12 (Leu12), the side chain points towards position 5, which is a valine at an a
position. Position 19 (Leu19) points towards position 16, which is an asparagine at an a
position. Therefore, position 12 was chosen as the mutation site to exclude the
unfavorable interactions between aliphatic Leu19 and polar Asn16 (Figure 2-4c).
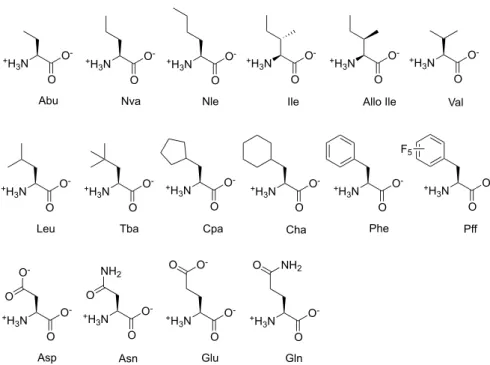
Sixteen different amino acids were incorporated individually at the mutation site (Figure
2-6). These amino acids can be divided into linear (Abu, aminobutyric acid; Nva,
norvaline; and Nle, norleucine), β-branched (Ile, isoleucine; Allo Ile, alloisoleucine; and
Val, valine), γ-branched (Leu, leucine; Tba, tert-butylalanine; Cpa, cyclypentylalanine;
Cha, cyclohexylalanine; Phe, phenylalanine; and Pff, pentafluorophenylalanine), and
polar (Asp, aspartic acid; Asn, asparagine; Glu, glutamic acid; Gln, glutamine).
(a)
(b) (c)
Figure 2-4. The sequence and the helical wheel diagram of GCN4. (a) Sequence of GCN4. The a and d positions are labeled. The d positions are numbered. (b) Ribbon representation of GCN4 with d-position leucines in space filling. Yellow: Leu12, blue:
Leu19. (c) Helical wheel diagram of GCN4. X denotes the mutation site.
Figure 2-5. CD spectrum of GCN4-Phe2. The central 2 d positions of GCN4-Phe2 were substituted with Phe.
Ac-RMKQ LEDKVEE LLSKNYH LENEVAR LKKLVGE R - OH
d a d a d a d a
5 12 19 26
2ZTA (GCN4 O'Shea d colored)
Page 1 of 1 Printed at 03:38:14 on Wed Apr 18 2012
a b
c
d
e f
g
f’
d’
b’
g’ c’
a’
e’
M V N V V E L E K K E Y A G KNSD
E R H E Q
L E K K
L L X L
L X L L
K K E L
Q E H R E V
V N V M
K E L E
G A Y E K
DSNK
!
Figure 2-6. Chemical structures of the amino acids.
Peptide Synthesis of GCN4-Derived Peptides
Peptides were synthesized using commercially available reagents by Fmoc-based
solid peptide synthesis (SPPS). However, Allo Ile, Nle, and Cpa were purchased in the
unprotected form, and the Fmoc group had to be incorporated for SPPS (Scheme 1).
The amino acid was dissolved in Na2CO3 solution. This aqueous solution was kept at
pH 10 over ice-water bath. A solution of Fmoc-OSu in tetrahydrofuran (THF) was
added dropwise into the amino acid solution, and the mixture was stirred for 3.5 hours.
For work up, the THF in the mixture was first removed under reduced pressure. The
remaining basic aqueous solution was extracted with diethyl ether. The aqueous fraction
+H3N O-
O Abu
+H3N O-
O Nva
+H3N O-
O Nle
O- O Ile
+H3N O-
O Allo Ile
+H3N O-
O Val
+H3N O-
O Tba
+H3N O-
O Cpa
+H3N O-
O Cha
+H3N O-
O Phe
+H3N O-
O Asp O- O
+H3N O-
O Asn NH2 O
+H3N O-
O Glu O O-
+H3N O-
O Gln O NH2
+H3N O-
O Pff
+H3N O-
O Leu
+H3N
F5
was then acidified with 1 M HCl until pH 2, followed by extraction with ethyl acetate.
The organic fraction was dried over Na2SO4, and concentrated in vacuo. The crude was
purified by flash chromatography.
Scheme 1. Incorporation of Fmoc Onto Amino Acid
The identity of the peptides was confirmed by matrix-assisted laser desorption
ionization – time of flight (MALDI-TOF) mass spectrometry. Upon confirmation of the
peptides, purification was carried out on reverse phase high performance liquid
chromatography (RP-HPLC) to greater than 95% purity. The crude yield of peptide
synthesis, molecular formula, calculated [MH+], and observed m/z of GCN4 peptides
are shown in Table 2-1.
H2N O
OH
R Fmoc-OSu, THF
Na2CO3 (aq), ice bath, 3.5 hrs
R= Allo Ile Cpa Nle
Fmoc-Allo Ile-OH, 90.1% yield Fmoc-Cpa-OH, 86.9% yield Fmoc-Nle-OH, 78.5% yield
O N
H O
OH O R


![Table 2-1. Crude Yield of Peptide Synthesis, Molecular Formula, Calculated [MH + ], and Observed m/z of GCN4 Peptides](https://thumb-ap.123doks.com/thumbv2/9libinfo/9607991.633653/44.892.120.768.209.738/table-peptide-synthesis-molecular-formula-calculated-observed-peptides.webp)