國立臺灣大學漁業科學研究所 碩士論文
Institute of Fisheries Science College of Life Science National Taiwan University
Master Thesis
呂宋鰻在台灣及菲律賓的生物地理分佈 Biogeographic distributions of Anguilla luzonensis
in Taiwan and Philippines
羅敏睿 Min-Jui Lo
指導教授:韓玉山 博士 Advisor: Yu-San Han, Ph.D.
中華民國 102 年 7 月
致謝
在這段碩士的學程當中,讓我學到了非常的多,由原本進實驗室像個愛玩 的小孩不知道長大,到後來拼命的量魚做實驗,轉變非常的大,從小就好動的 我,家人一直擔心我是否無法去面對實驗室的生活,我自己也非常的存疑,這 之中的轉變最大的感謝真的是韓老師,在月報的幾句簡單的訓誡,能讓一向坐 立難安好玩的我,真正的有去思考未來,進一步的去拚實驗,而接下來的每個 月報,也都是皮繃得緊緊的希望能得到認可,老師的幾句稱讚,能讓簡單的我 高興半個月,從懵懂的我,變成跟朋友聊天話題卻時常離不開鰻魚的我,真的 是個很大的轉變,學生敏睿是真心的謝謝這位專精與廣泛能二者唯一的恩師。
當然,在實驗室之中每一位都是不可或缺的,從剛進實驗室的時候,敬華 學姊一步一腳印的教我量魚、鑑定、歸檔,這些都歷歷在目,是非常盡責且不 吝指導的學姊,而婉之、何敏更是有任何的好東西,都與我們這些學弟妹分享,
讓我們能在實驗室有像在家一樣的感覺,明惠及小陶更是實驗室說說笑笑的好 夥伴,就連我碩士論文不懂的地方在網路上詢問小陶,他也是耐心的一一解答,
有這群這樣好的學姐們,也讓我們碩士過得更加的精彩。而現在碩一可愛的學 弟妹們,真的個個都是寶,做實驗跟雅棻聊的天花亂墜,昱如學妹每次的聽我 發牢騷,翔奕不但是個好球友,更是個好學弟,下午茶後門的行動,他總是不 會缺席,最有緣的是寧比學妹了,我們不但同樣是苗栗人,連國中高中也都是 讀同樣的學校,真的有一見如故稱兄道弟的感覺,讓人心曠神怡。新進實驗室 的一群,雖然相處時間不久,但我相信你們一定能非常順利的!而同屆的阿旗、
搞搞、阿梅、邦又、鳳媖、觀元,我們一起走過一起分享一起緊張一起開心,
這些不需要言語並且永遠忘不了的!最後我要謝謝一路陪著我不管是高興生氣吵 架的咪咪醬以及我可愛的奇丁跟桔子還有老媽,不用多語,沒有你們,我走不 到今天,也非常謝謝口委們熱誠熱心的幫助我們完成論文,順利畢業!
中文摘要
鰻魚為東亞國家重要的養殖魚種之一,但近年鰻魚的資源量都在急速的下 降,不論是鰻苗或是成鰻,數量都比以往來的少,而鰻魚養殖所需的鰻苗,完 全都是從漁民捕撈而來,在資源量減少的情況下,物種復育、新養殖鰻種的興 起甚至取代,都成為研究的目標之一,而呂宋鰻 (A.luzonensis,又名黃氏鰻 A.huangi) 於 2009 年被公布為第 19 種鰻魚,然而此新鰻種雖然已被公布,但 對於它的生物地理分布、資源量、生物特徵等相關的研究資料是少之又少,因 此本篇想針對呂宋鰻的生物地理分布與生物形態特徵,建立較完整的資料庫。
本篇的樣本來源分成兩個部分,以呂宋鰻被發現的時間 (2008) 做為分界 點,第一部分是 2008 年至 2011 年,採自菲律賓、台灣採集之鰻苗,第二部分 是 1984 年至 2008 年採自台灣之鰻苗。利用型態與 DNA 鑑定方法,鑑定出各採 樣點之鰻苗組成,由此建立出呂宋鰻的生物特徵及生物地理分布。結果指出,
呂宋鰻的高捕獲量主要出現在菲律賓呂宋島的 7、8、9 三個月,而在這三個月 中,其數量甚至超過鱸鰻,而在台灣方面,呂宋鰻非常稀少,捕捉量 8、9、10 月較其他月份為高。生物形態特徵方面,鱸鰻及呂宋鰻的鰭差比分布圖之比較,
可看出有少部分重疊的情況發生,而導致有可能誤判。呂宋鰻色素與體長的相
關變化,從 VA 時期至 VIA3時期呈現緩慢下降或是停滯,趨勢與日本鰻類似;月
份與體長之比較,趨勢並不明顯且無規律。呂宋鰻的生物地理分布於本篇中確 i
立出以菲律賓為主。
關鍵字: 呂宋鰻、生物地理分布、生物型態、鰭差比
ii
Abstract
Eels have been an important category in aquaculture in East Asian countries. But in recent years the stock size of eels has been in rapid decline. No matter it is eel fry or adult eels, their quantities are much less than the past. All the eel fry needed in eel farming come totally from the catches by fishermen. Under the circumstance of reduced stock size, things like species rehabilitation, emergence of newly farmed eel species or replacement by new species have become one of the targets of the researchers. A. luzonensis, also known as A. huangi, has been announced as the 19th species in 2009. Although this new species has been announced, the relevant research information on its biogeographic distributions, stock size, and biological features is
quite meager. As such, this paper aims at establishing a more comprehensive database of the biogeographic distributions and early life history of A. luzonensis.
Samples used in this paper are from two parts. The first part is collections of A.
luzonensis from 2008 to 2011. The second part is collections of A. marmorata from
1984 to 2008. The demarcation is the time (2008) when A. luzonensis was discovered.
Collections made after 2008 are mostly in places in the Philippines and Taiwan. Prior to 2008, they were mainly collected in Taiwan. After that, pattern identification is employed to establish the biological features and biogeographic distributions of A.
luzonensis. The results indicate that high catches of A. luzonensis are mainly in the iii
three months of July, August, and September in the Philippines. In the three months the amount of A. luzonensis caught is even more than that of A. marmorata. In Taiwan, A. luzonensis is extremely rare and from September to January more catches are seen. In terms of biological patterns and features, we can see that there is an overlap that leads to possible misidentifications from the comparison of fin difference ratio in A. marmorata and A. luzonensis. The associated change of pigmentation of A.
luzonensis and its total length shows resemblance in the run chart compared to that of
A. japonica in that there is a slow downward slope or even a standstill from VA to
VIA3 stages. The comparisons of months with total lengths showed no obvious upward or downward trend.
In this paper we have established the biogeographic distributions of A. luzonensis in the Philippines primarily.
Keywords: Anguilla luzonensis, biogeographic distribution, biological pattern, fin difference ratio
iv
Contents
中文摘要 ... i
Abstract ... iii
Introduction ... 1
Eel resource decline ... 1
Biogeographic distributions ... 2
Pigmentation stage ... 3
Materials and methods ... 6
Sample collection ... 6
Species identification based on morphometr : morphological measurement ... 6
Species identification based on morphometry:molecular methods ... 7
Results ... 9
Fin difference distribution of A. luzonensis and A. marmorata ... 9
Monthly glass eel composition by location ... 9
Correction of Past Misidentifications ... 10
Pigmentation stages of A. luzonensis and its changes with TL ... 10
Comparison of the biological patterns and features of A. luzonensis in Taiwan, Luzon Island and Mindanao ... 12
Discussion ... 13
Misidentification of A. celebesensis ... 13
Spawning area and Species composition of Anguilla glass eels ... 15
Pigmentation v.s Total-length ... 16
Conclusion ... 19
References ... 20
v
Figure legend
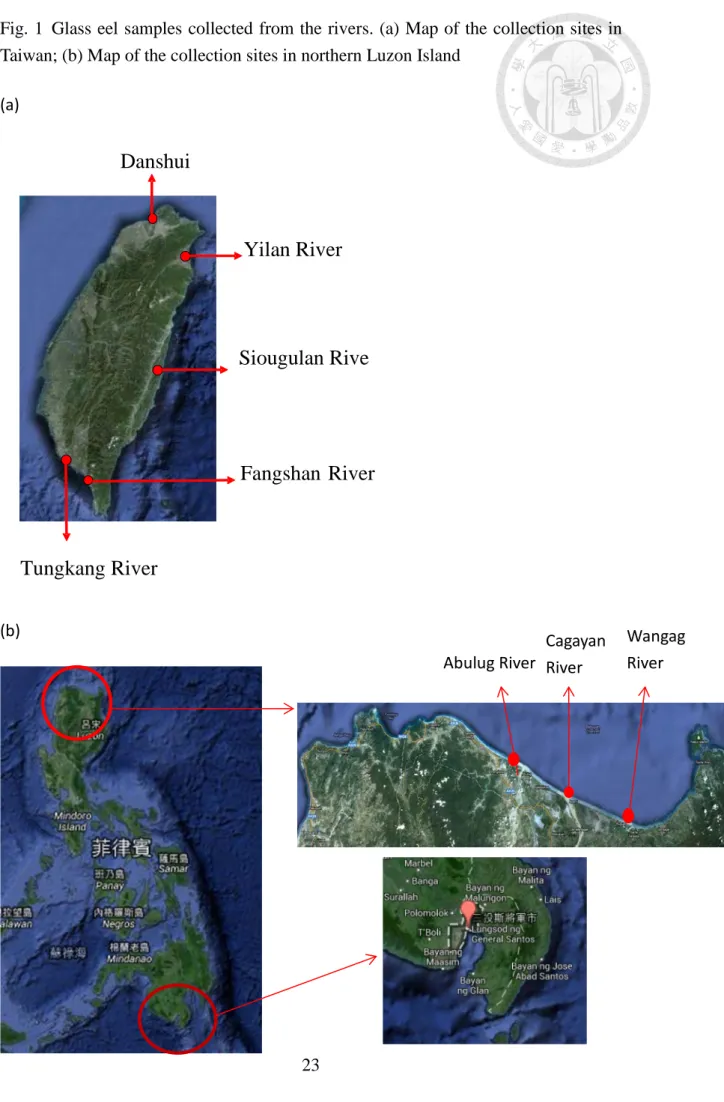
Fig. 1 Glass eel samples collected from the rivers. (a) Map of the collection sites in
Taiwan; (b) Map of the collection sites in northern Luzon Island ... 23
Fig. 2 The fishing tackle for capturing glass eel in the estuaries. (a) The traditional
triangle nets were used in Siouguluan and Fengshan River; (b) the fixed fyke
nets were used in Company Tian creek of Danshui, Cagayan, Wangag and
Abulug River; (c) the mobile fyke nets were used in Yilan and Tungkang River
... 23 Fig. 3 Identification of different development stages of pigmentation on the Anguilla
luzonensis glass eels. VA, VB, VIA1, VIA2, VIA3 and VIA4 refer to Strubberg
(1913). ... 27
Fig. 4 Identification of Anguilla glass eels based on morphology ... 28
Fig. 5 Fin difference distribution with AM (Anguilla marmorata) and AL (Anguilla
luzonensis) ... 29
Fig. 6 Sample collection from north Luzon Island from May.2008 to Sep. 2009 in (a)
Cagayan river (CR) (b) Abulug river (AR) (c) Wangag river (WR) ... 30
Fig. 7 Pigmentation stage v.s Total length (mm) of Anguilla luzonensis (a) Abulug
river (b) Cagayan river (c) Wangag river (d) Taiwan (e) Philippines (3 rivers) 33
Fig. 8 Means Total length of A.luzonensis (VA) from May.2008 to Sep.2009 (a)
vi
Abulug river (b) Cagayan river (c) Wangag river (d) Abulug river+Cagayan
river + Wangag river (VA+VB) ... 36
Fig. 9 Means Total length of A.luzonensis (VB) from May. 2008 to Sep. 2009 (a)
Abulug river (b) Cagayan river (c) Wangag river ... 40
vii
Table contents
Table 1 Sample collection from north Luzon island from 2008 May and 2009 Dec ... 43
Table 2 Monthly composition of glass eel samples of A. marmorata (AM) and A.
luzonensis (AL) collected from Taiwan ... 44
Table 3 Sample collection from Mindanao from 2009 Jan and 2010 Apr ... 45
Table 4 Pigmentation stage of A.luzonensis from Abulug River, Cagayan River,
Wangag River and Taiwan. ... 46
Table 5 Pigmentation and TL±SD of A.luzonensis from Abulug River,Cagayan River,
Wangag River and Taiwan. ... 47
Table 6 Morphology of Anguilla luzonensis from Luzon Island,Taiwan and Mindanao ... 48
Appendix table
Appendix table 1. Data of Anguilla luzonensis eels used for analysis in this thesis.
... 49
viii
Introduction
Eel resource decline
The eels (genus Anguilla) are important economic species in Southeast and Northeast Asia. Due to artificial reproduction techniques of eel fry (glass eel) haven’t been found yet, all of the eel fry needed for aquaculture depends on the wild catch.
In recent years the Japanese eel Anguilla japonica, European eel A. Anguilla and American eel A. rostrata stock sizes have been significantly declined owing to overfishing, habitat destruction, global climate change, dams, and other unknown factors (Tatsukawa 2003; Hitt 2012). How to restore the abundance of these eels or discover a new aquaculture species has become an important task.
There are 19 species of anguillids distributed around the world in the coastal areas of North Atlantic Ocean, Indian Ocean and West Pacific Ocean (Aoyama 2009).
All anguillids spawn in the tropical or subtropical oceans. After eggs are hatched, leaf-like bodies called leptocephali are transported by ocean currents from their spawning area to continental habitats then metamorphose into glass eels (Miller 2009;
Aoyama 2009). After metamorphosis, glass eels enter estuaries, rivers or lakes. They grow for years and then become silver eels to return to their birthplace to spawn and then die (Tsukamoto 2006).
1
Biogeographic distributions
In Taiwan, there are four species of freshwater eels: A. japonica, A. marmorata, A. bicolor pacifica (Tzeng 1983) and A. luzonensis have been reported (Han 2010).
The biogeographic distributions of tropical eel, Anguilla mamorata, larvae are transported by the North Equatorial Current (NEC), then into either the northward flow of Kuroshio or the southward flow of Mindanao Current and recruit to Taiwan, Southern Japan, the Philippines and northern Indonesia. In Taiwan, A. marmorata is found throughout the year and dominant between March and July. In the Philippines, A. marmorata is dominant between February and June (Han 2013) . Anguilla japonica
is transported by the NEC and then enters the northward flow of Kuroshio towards East Asia. In Taiwan, A. japonica glass eels mainly occur in winter in Yilan River, Danshui and Tungkang River. Aoyama (2009) suggested that the spawning areas of A.
bicolor pacifica and A. luzonensis were also located in the NEC region. The mid- and
large-size leptocephali of A. bicolor pacifica are collected in the east of New Guinea, Indonesia and west of the Mariana Islands. This finding indicates that A. bicolor pacifica may spawn in the western North Pacific (WNP) region and might have
multiple spawning areas in the western Pacific (Kuroki et al. 2006) .
The new species, A. luzonensis, also known as A. huangi, was found in Cagayann River, on the northern Luzon Island of the Philippines (Watanabe et al. 2009; Teng et
2
al. 2009) . The leptocephali of A. luzonensis were found in the west of the Mariana
Islands, which supported that A. luzonensis also spawned in the NEC (Aoyama 2009;
Watanabe et al. 2009; Kuroki et al. 2012). Although A. luzonensis has spawning sites in the NEC, the precise spawning area, migratory routes, biogeographic distributions and early life history of A. luzonensis are still poorly understood. The experiment of this study is to understand the biogeographic distributions of A. luzonensis in Taiwan and the Philippines.
Pigmentation stage
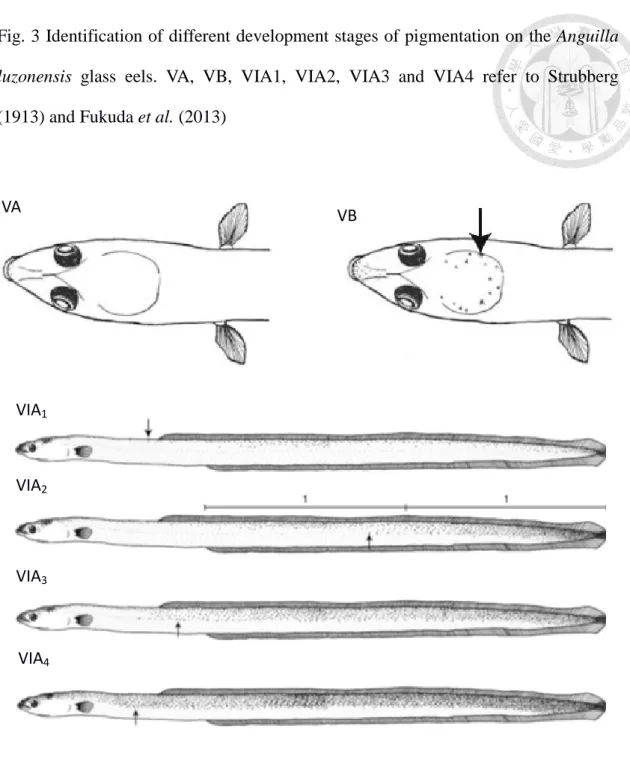
The progress of glass eel growth and development can be measured by pigmentation stages, which commence as water temperature rises and physiological changes take place (Strubberg 1913). Previous studies revealed that the changes in glass eel size and pigmentation proceed in parallel (Tesch 2003a). Water temperature determines the pigmentation and correspondingly TL decrease and later growth as well (Strubberg 1913). As the glass eel continues to grow, external pigments develop from sub-epidermal layers. Glass eel is defined in this paper as all developmental stages from the completion of the leptocephalus metamorphosis to full pigmentation.
The process includes stages VA to VIB. Aside from progressive pigmentation, the total length changes simultaneously to some extent. Knowing the stage of development of recently arrived recruits is also important since there is considerable
3
variation in the stages of glass eels as they arrive in coastal areas after their long migration in the ocean (Sugeha 2001; Aoyama 2009). Their stages and body condition when they arrive are important information for evaluating seasonal and interannual recruitment variations especially for anguillid species that have shown recruitment declines in recent decades. An alternative but much more laborious way to evaluate the development stage is to estimate the age using analysis of their otolith rings.
However, for young eels this method may not always be reliable because the deposition of otolith material can stop at water temperatures below 10ºC, which is a temperature that anguillid glass eels recruiting to the higher latitude regions would encounter during the winter months (Fukuda 2009). The experiment of this study is to understand the TL variation of all the pigmentation stages in detail for A. luzonensis.
Therefore the main purpose of this paper is to conduct a further research on the biogeographic distributions of A. luzonensis in Taiwan, Luzon Island, and Mindanao.
Their monthly catches would be used to do pattern identification and survey of pigmentation stages. Then clearer temporal and spatial distributions of A. luzonensis could be established. Furthermore a comparison is made between the relative abundance of A. marmorata and A. luzonensis to understand the differences between the two species in the peak season as well as the distinction between the places they are caught. Lastly a trend comparison of pigmentation stages and total lengths is made
4
in the hope of completely establishing the biogeographic distributions and biological features of A. luzonensisin Taiwan, the North Luzon Island and Mindanao.
5
Materials and methods
Sample collection
Glass eels in the northern Luzon Island were collected monthly from May 2008 to September 2009 of the Philippines of Cagayan River, Wangag River, and Abulug River (Fig.1), all in the province of Cagayan in northern Luzon. Samples were collected with the use of a fyke net (Fig. 2b) dipped to the bottom of the river. With glass eels from Taiwan (Fig.1), we can divide them into 2 parts. Part1: were caught using fyke nets (Fig. 2a, c) at night between February 2009 and December 2011 between 1 and 3 times monthly from the estuaries of Danshui River (Northwestern Taiwan), Donggang River (Southwestern Taiwan), Yilan River (Northeastern Taiwan), and Siouguluan River (Eastern Taiwan). All glass eels were purchased from local fishermen. Part 2: were caught using fyke nets between 1984 and 2008 from estuaries in Taiwan. Glass eels in areas around the Celebes Sea were collected using hand nets from January 2009 to April 2010 from Buayan River, General Santos City in southern Mindanao.
Species identification based on morphological measurement
Eel species were identified using morphological methods. The total length, pre-
6
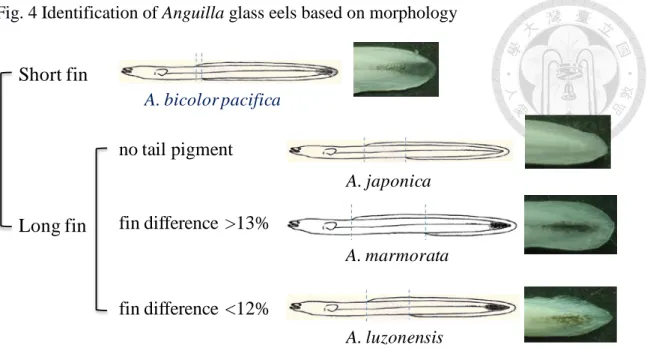
dorsal length, and pre-anal length (to the nearest 0.1 mm) of each glass eel were measured under a stereomicroscope. The following equation was used to calculate the fin difference as an index to classify whether an eel was long finned or short finned: Fin difference = AD/TL × 100%, where AD is the vertical distance from the origin of the dorsal fin to the anus and TL is the total length. Morphological measurement were also checked by the presence or absence of caudal cutaneous pigmentation (Strubberg 1913) (Fig. 3). A. japonica and A. bicolor pacifica were easily identified by using morphological methods. Samples with fin difference
<13% from Taiwan and Luzon Island were identified by molecular methods. All samples from Mindanao were identified by molecular methods.
Species identification based on molecular methods
The fin difference value of A. marmorata and A. luzonensis of 12-13% was usually ambiguous (Fig. 4), so molecular identification was used. A small piece of muscle was purified and extracted by a commercial DNA kit (Bioman Scientific Ltd.). The sample tissue was grounded and homogenized by using a micropestle and 200μl GT buffer. After reaching homogenity, 20μl Proteinase K was added and the sample was incubated at 60ºC for 30 min to lyse the sample. After that, 200μl GB buffer was added and the sample was incubated at 70ºC for 20 min until the sample
7
lysate was clear. Then 200μl ethanol was added to the sample lysate and vortex immediately to mix the sample. The mixture was then transferred to GD column and centrifuged at 13,000 rpm for 5 min. After centrifuge, the flow-through was discarded and 400μl W1 buffer was added for centrifugation at 13,000 rpm for 1 min. After 600μl wash buffer washing at 13,000 rpm for 1 min and again for additional 3 min to dry the column, the GD column was transferred into a new microcentrifuge tube and 100μl preheated Elution buffer was added. Wait 3-5 min for the elution buffer to be absorbed by the matrix, then the GD column was centrifuged at 13,000 rpm for 1 min to elute purified DNA. The extracted DNA was stored at -20ºC before polymerase chain reaction (PCR) occurred. PCR was performed in 25µl reaction volume consisted of 11µl of double distilled water (ddH2O), 12.5µl 2X Taq DNA Polymerase Mastermix-Red (Bioman Scientific Ltd.), 0.5µl forward primer, 0.5µl reverse primer, and 0.5µl of total genomic DNA for a total of 25µl solution. Amplification parameters were as follows: initial denaturation at 94ºC for 3 min, followed by 30 cycles of denaturation at 94ºC for 30s, annealing at 57ºC for 30s and extension at 68ºC for 30s, followed by final extension at 68ºC for 10min and 4ºC for 10min. The PCR products were
electrophoresed and sent for sequencing for direct species identification. Universal Anguilla primer set for cytochrome b (forward:5′-GATGCCCTAGTGGATCTACC-
8
3′; reverse: 5′TATGGGTGTTCTACTGGTAT-3′) was used for PCR.
Results
Fin difference distribution of A. luzonensis and A. marmorata
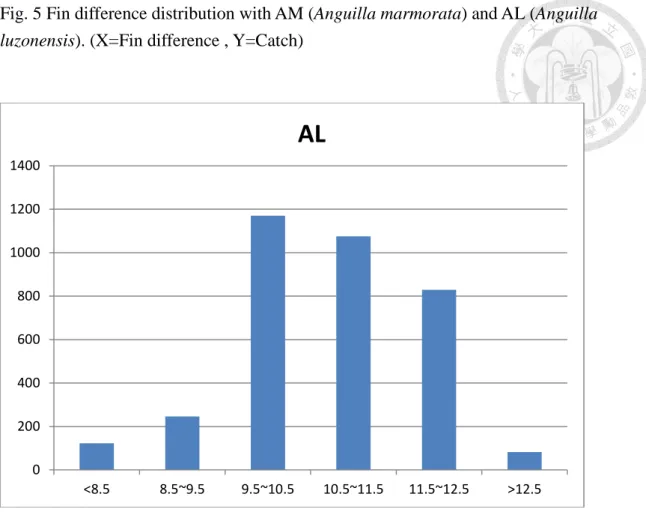
A distribution chart is made against the fin differences between A. luzonensis and A. marmorata in the three major rivers of Northern Luzon, namely Cagayann River, Wangag River, and Abulug River. Comparing the two species and the result after analysis shows there are 829 eels of A. luzonensis with fin differences in the range of 11.5~12.5, and 82 eels have a fin difference above 12.5 (Fig. 5). As to A.
marmorata, there are 38 eels with a fin difference below 12.5. There are another
494 eels in the sample with fin differences in the range 12.5~13.5. It clearly shows an existence of overlapped region in the distribution chart against the fin differences between A. luzonensis and A. marmorata. For this, molecular identification has become the most accurate way of differentiating between A. luzonensis and A.
marmorata for now (Fig. 5 ).
Monthly glass eel composition by location
4 Anguilla species were identified in Taiwan and Luzon Island: A. japonica, A.
marmorata, A. bicolor pacifica, and A. luzonensis. In Taiwan, glass eels of A.
9
marmorata are found all the year, and are much more dominant than A. luzonensis.
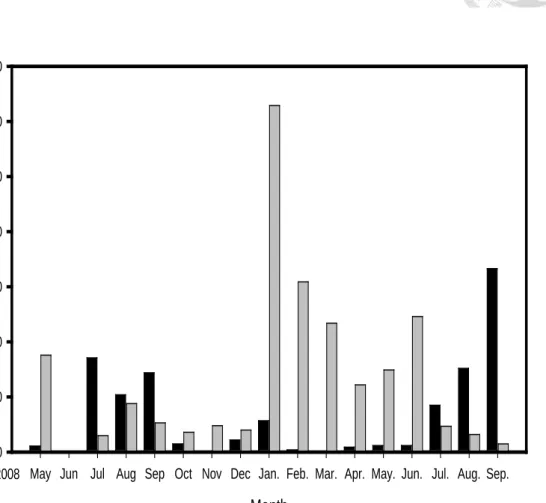
In the Philippines, A. marmorata is dominant between February and June in Luzon Island, A. luzonensis is dominant half of the year, mainly from July to September, in northern Luzon Island (Table 1, Fig 6a,6b,6c). In Taiwan, A. marmorata is dominant between March and July and A. luzonensis are rare all year round (Table 2). In contrast to the wild distributions of A. marmorata in Southeast Asia, A. luzonensis seems to be mainly distributed in Luzon Island but is rare in Taiwan and Mindanao (Table 3).
Correction of Past Misidentifications
Pattern identification of eels before 1984 and up to 2008 also determined 6 and 5 A. luzonensis separately in 2001 and 2000. It is quite obvious that even A.
luzonensis was discovered in 2009, Taiwan had been one of its habitats prior to that.
One thing worth mentioning is that A. celebesensis was also discovered.
Pigmentation stages of A. luzonensis and its changes with TL
After measurement of its total length and distinction of pigmentation stages, the result is a relationship chart of total lengths vs. pigments for A. luzonensis in the three rivers in Luzon Island (Table4). In these three rivers, we could find that the VA and VB stages of A. luzonensis showed a significant difference in total lengths in
10
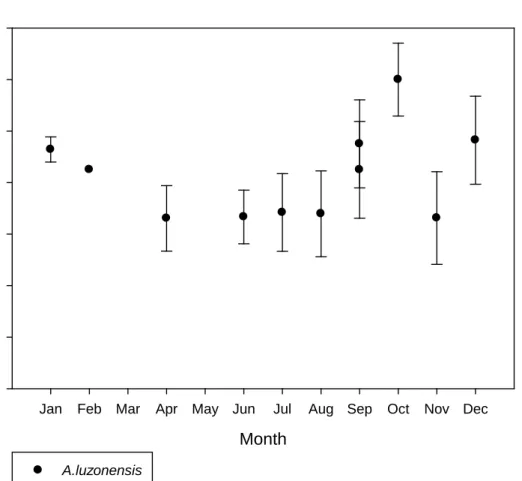
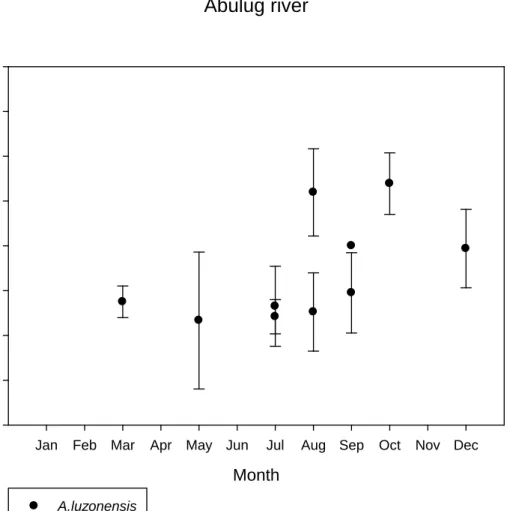
Cagayan river and Wangag river (P<0.05) (Table5). A. luzonensis in two of the rivers show a slow decrease of total lengths. The two rivers that are more significant are Cagayan River and Wangag River (Fig. 7a,7b). The analysis chart of Wangag River shows a slightly significant slow downward slope and then a gradual upward slope (Fig. 7c). As to Cagayan River and Abulug River, there is no clear indication of changes with relation to pigmentation stages and total lengths after VIA3 as there is no sample available.
The pigmentation analysis chart from Taiwan indicates a slow gradual decrease or gradual increase after the VA stage. Right after the VIA4 stage, it shows a more rapid growth of total lengths. However, the quantity of A. luzonensis is quite scattered in Taiwan (Fig. 7d).Yet if the analysis is conducted using a larger scale (Abulug River+Cagayan River +Wangag River) , the analyzed result would show that total lengths are actually shorter in stage VA than in stage VB and it would exhibit significant differences as well. However when viewed from VIA2 to VIA4, there is no significant change in total lengths as they either are at a standstill or decrease slightly. (Fig. 7e)
When we use the VA stages of A. luzonensis to do analysis of the variation in total lengths between the months of May 2008 and August 2009, we can see that there's no obvious pattern in the total length between each month (Fig. 8a,b). As to
11
Wangag river, there is no way to see if there is an upward or downward trend since the quantities collected in many of the months were too low or even none (Fig. 8c).
From the aggregated data of the three rivers, the condition is also level with no obvious upward or downward trend. (Fig. 8d). As to analysis for VB, it does not show any obvious pattern or change either (Fig. 9a, b, c). Generally speaking, there is no significant change in total lengths of A. luzonensis between each of the months.
Comparison of the biological patterns and features of A. luzonensis in Taiwan,
Luzon Island and Mindanao
From Table 6 we can see that A. luzonensis in the three major Philippine rivers do not show significant differences in pre-dorsal fin length, pre-anal fin length and fin difference ratio and so forth and if we do not take pigmentation stages into consideration. The same is true in Taiwan with differences shown only in pre-dorsal fin length and fin difference ratio. As to Mindanao, the sample was only 30 and they showed significant differences with those of Luzon Island in total length, pre-anal length, and pre-dorsal length. However as the sample was extremely small in size, more samples are needed to verify the accuracy of the analysis.
12
Discussion
Misidentification of A. celebesensis
The new anguillid species A. luzonensis was identified in the Cagayan River System (Watanabe et al. 2009). Meanwhile, Teng et al. (2009) also found this new species, named A. huangi. To identify this species, identification based on morphology such as variegated markings, maxillary bands, numbers of vertebrae, and the location of the origin of the dorsal fin were not reliable enough due to its similar and overlapping morphometric characteristics with those of A. celebesensis and A. marmorata. Molecular-based technique was a better method for species identification than morphometric characteristics but this method needs more time to be spent than identification on morphology and is expensive (Aoyama 2009; Teng et al. 2009; Watanabe et al. 2009). In the previous paper, A. luzonensis is misidentified
as A. celebesensis in 1976 and 1982, because they have similar fin difference ratio between 10%-12.5% and indistinguishable morphological characteristics, such as marbled dark brown on the back of adults and caudal cutaneous pigmentation for glass eels (Tabeta et al. 1976; Tzeng 1982 ; Teng et al. 2009; Watanabe et al. 2009) Glass eels of A. celebesensis could have come from a spawning area near the mouth of the Celebes Sea, recent data on the distribution of leptocephali of this species
13
indicate that spawning occurs in both the Celebes Sea and in Tomini Bay (Aoyama et al. 2003). However, the northern Philippines is further north than the known
distribution range of A. celebesensis (Ege 1939), so it now appears fairly certain that the glass eels originally identified as A. celebesensis from this area were actually A.
luzonensis, since no A. celebesensis has been identified using DNA analysis from
the northern Philippines, whereas A. luzonensis glass eels and yellow eels are clearly present there (Teng et al. 2009, Watanabe et al. 2009). Regardless of how often A. luzonensis leptocephali or glass eels may have been historically misidentified, there appear to be very few reports of “Anguilla celebesensis”
collected in the regions north of the Philippines (Ege 1939, Arai et al.2003, Teng et al. 2009, Watanabe et al. 2004).
One report from Taiwan (Tzeng 1982) and some previously reports of A.
marmorata leptocephali that were not genetically identified, could have actually
been those of A. luzonensis (Jespersen 1942, Miller et al. 2002, Miller and Tsukamoto 2004). Nevertheless in this paper, we could see that the total lengths, pre-anal lengths and pigments of the two types of glass eels are very similar from pattern identification of A. marmorata and A. luzonensis in the Philippine region and the distribution chart of their fin difference ratio. Furthermore the fin difference ratio also shows there is an overlap between the two species. Therefore we could
14
deduce that some A. marmorata identified in the past through pattern identification but not yet through molecular identification could have been A. luzonensis as well.
Spawning area and Species composition of Anguilla glass eels
In previous studies, the leptocephali of A. luzonensis were collected offshore or at the edge of the subtropical gyre of the western North Pacific. The offshore presence of these leptocephali supports the hypothesis that this species may have a spawning area similar to the three other anguilliform eels in the region (A. japonica, A. marmorata and C. myriaster) (Kuroki 2012). From the analysis of this paper, the
biogeographic distributions of A. luzonensis also coincide with previous studies by other scholars. The A. luzonensis collected in this study are from regions of Taiwan, Luzon Island, and Mindanao. Among them, most of the A. luzonensis are from Luzon Island and fewer are from Taiwan and Mindanao. This matches the paper of Han et al.(2013) in that their hatching grounds correspond to those of A. marmorata and A. japonica and that A. luzonensis are drifted with the NEC. When they reach the southern part of Luzon Island, they are split into two branches that are the Mindanao current (minor) and the Kuroshio (major) separately. Therefore the major distributions are found in Luzon Island. As to the rare sample in Mindanao, they are transported by the Mindanao current (minor). On the other hand, A. luzonensis
15
occurred dominantly during July, August and September but the total collection was rare in Taiwan and Mindanao. As to why most of the distributions of A. luzonensis are centered around Luzon Island and not like A. marmorata to be plenty in amount in Taiwan as well, the conjecture is that it could be related to the biological features of the leptocephalus stage of A. luzonensis. A likely guess is that the larva duration of A. luzonensis is not as long as that of A. marmorata. Therefore during the drifting process, they could not be transported to areas like Taiwan in large amounts as A.
marmorata due to its ability to move, leading to its distributions to be limited to
northern Luzon Island only and not to be drifted further to Taiwan. This is also why there are only small amounts of A. luzonensis in Taiwan and Mindanao.
Pigmentation v.s Total-length
In a previous study the body measurements of A. japonica indicate that the TL and BW decreases or stays about the same until VIA3 and VIA4 and then starts to increase rapidly, which is similar to observations of A. anguilla from Tunis Lake, Tunisia, whose TL and BW decreased until VIA3 before increasing rapidly during the VIA4 stage (Tesch 2003). A. japonica, the TL and BW slightly decreased until VIA2 and then increased. The TL–BW relationships of each pigmentation stage showed developmental changes in their body proportions. The body proportion
16
changed gradually to the slender body form from VA to VIA3 and VIA4, and became the thicker body form at VII. This was because the eels with larger TL had a thicker body form during VIB, suggesting that VIB was the transition time of the change from the slender body (VIA3 and VIA4) to the thick body (VII). (Fukuda et al. 2013).
The relationship of pigmentation stages and total lengths in this study shows a similar pattern to studies previously made. A. luzonensis in VA and VB stages did not show a significant increase in total lengths. Among them, total lengths in Wangag River and Cagayan River show a decrease from the VA to VB stages. In VIA2 to VIA3 stages, it is a situation of ups and downs with no significant increase. But as the lab does not have pigmentation stage A. luzonensis samples after VIA4, we could not make sure if there is a relevance to previous studies made by others in stages after VIA4 to VII.
The analysis of the total length and pigmentation of A. luzonensis in this study shows that even the total lengths from stages VA to VIA3 show a condition of slow decrease or even standstill, it is obvious the variation is not so much as those of A. japonica.
Among them, the total length in stage VB for those from Abulug River is even longer than that in stage VA. The explanation concerning this could be that as A. japonica are temperate eels and A. luzonensis are tropical eels, there is difference in the habitats they choose. Furthermore we usually see more production and resources in tropical
17
regions than in temperate regions. Therefore the conjecture that variation in the total lengths of A. luzonensis is not so noticeable is because they have more food choices and lead to little impact on their growth. The explanation for the total lengths of eels and change in the pigmentation stage is that the transition between body forms and growth may correspond to their ecological change from using passive transport in glass eels with tidal flow into rivers, to their settlement and further movements of elvers upstream (Fukuda et al. 2013). But it is still little known whether these changes in pigmentation stages and body growth are due to the effect of salinity change, a time-delay effect of recruitment, ecological change or all of them (Leander et al.2012). As to the fact that there is no significant change in total lengths of A.
luzonensis between each of the months, the conjecture in this study is that as Luzon
Island is situated in the temperate zone with a mean annual temperature of 27 degrees C. In tropical regions the latitudinal diversity gradient is higher than that of temperate regions. Furthermore as the climate in Luzon Island is not as changeable as in Taiwan, but rather is one with a constant high temperature. Therefore there is no significant change in total lengths of A. luzonensis as they keep a constant length when they live in an environment with abundant food and a higher growth rate.
18
Conclusion
Through analysis we could find that the Philippines is the major distribution region for A. luzonensis. The three months of July, August, and September are the time with a higher trend. As to Taiwan and Mindanao, there is only a small amount of A. luzonensis. In the identification between A. marmorata and A. luzonensis afterwards, we need to be more cautious as there is an overlapped morphometric index between them. Molecular identification is still the top choice nowadays. Even it is higher in cost, it has the best accuracy. As to pigmentation stages and changes in total lengths of A. luzonensis, we can see there is no significant different from VIA2 to VIA4 and stage VA to VB show a decrease that is similar to that of A.
japonica.
19
References
Aoyama J (2009) Life history and evolution of migration in catadromous eels (genus Anguilla). Aqua-BioScience Monographs 2:1-42.
Arai T, Miller MJ, Tsukamoto K (2003) Larval duration of the tropical eel Anguilla celebesensis from Indonesian and Philippine coasts. Marine Ecology Progress
Series 251:255-261.
Fukuda N, Kuroki M, Shinoda A, Yamada Y, Okamura A,Aoyama J, Tsukamoto K (2009) Influence of water temperatureand feeding regime on otolith growth in Anguilla japonica glasseels and elvers: does otolith growth cease at low
temperatures? Journal of Fish Biology 74:1915–1933.
Fukuda N, Miller MJ, Aoyama J, Shinoda A, Tsukamoto K(2013) Evaluation of the pigmentation stages and body proportions from the glass eel to yellow eel in Anguilla japonica.Fisheries Science 79:425-438
Han YS (2010) Study of production of potential aquaculture species – Anguilla marmorata. Project report of Council of Agriculture, Executive Yuan.
Haro AJ, Krueger WH (1988) Pigmentation, size, and migration of elvers (Anguilla rostrata (Lesueur)) in a coastal Rhode-Islandstream. Canadian Journal of
Zoology 66:2528–2533.
Kuroki M, Aoyama J, Miller MJ, Wouthuyzen S, Arai T, Tsukamoto K (2006) Contrasting patterns of growth and migration of tropical anguillid leptocephali in the western Pacific and Indonesian Seas. Marine Ecology Progress Series 309:233-246
Kuroki M, Miller MJ, Aoyama J, Watanabe S, Yoshinaga, Tsukamoto K (2012) Evidence of offshore spawning for the newly discovered anguillid species
20
Anguilla luzonensis (Teleostei: Anguillidae) in the western North Pacifc. (early
view) Pacific Science 66:4
Kuroki M, Aoyama J, Miller MJ, Yoshinaga T, Shinoda A, Hagihara S, Tsukamoto K (2009) Sympatric spawning of Anguilla marmorata and Anguilla japonica in the western North Pacific Ocean. Journal of Fish Biology74:1853-1865
Leander NJ, Shen KN, Chen RT and Tzeng WN (2012) Species Composition and Seasonal Occurrence of Recruiting Glass Eels (Anguilla spp.) in the Hsiukuluan River, Eastern Taiwan. Zoological Studies, 51: 59-71.
Luers DF, Love JW, Bath-Martin G (2011) Settlement and pigmentation
of glass eels (Anguilla rostrata Lesueur) in a coastallagoon.Environmental Biology of Fishes 90:19–27
Strubberg A (1913) The metamorphosis of elvers as influenced by outward conditions.
MeddrKommn Havunders. Serie Fiskeri 4: 1-11.
Sugeha H, Arai T, Miller MJ, Limbong DKT (2001) Inshoremigration of the tropical eels Anguilla spp. recruiting to thePoigar River estuary on north Sulawesi Island.
Marine Ecology Progress Series 221:233–243
Tatsukawa K (2003) Eel resources in East Asia.In : Aida K, Tsukamoto K, Yamauchi K (eds.) Eel biology, Springer, Tokyo. 293-298
Teng HY, Lin YS, Tzeng CS (2009) A new Anguilla species and a reanalysis of the phylogeny of freshwater eel. Zoological Studies 48:808-822
Tesch FW (2003a)The eel. Oxford, UK: Blackwell Science 130-131.
Tsukamoto K (2006) Spawning of eels near a seamount. Nature 439:929
Tzeng WN (1983) Species identifiecation and commercial catch of the anguillid elvers from Taiwan. China Fish Month. 366:16-23 (in Chinese)
Watanabe S, Aoyama J, Tsukamoto K (2009) A new species of freshwater eel Anguilla
21
luzonensis (Teleostei: Anguillidae) from Luzon Island of the Philippines. Fish Science 75:387-392
22
Fig. 1 Glass eel samples collected from the rivers. (a) Map of the collection sites in Taiwan; (b) Map of the collection sites in northern Luzon Island
(a)
(b)
Siougulan Rive Yilan River
Fangshan River Danshui
Tungkang River
Abulug River Cagayan River
Wangag River
23
Fig. 2 The fishing tackle for capturing glass eel in the estuaries. (a) The traditional triangle nets were used in Siouguluan and Fengshan River; (b) the fixed fyke nets were used in Company Tian creek of Danshui, Cagayan, Wangag and Abulug River;
(c) the mobile fyke nets were used in Yilan and Tungkang River (a)
(b)
24
(c)
25
26
Fig. 3 Identification of different development stages of pigmentation on the Anguilla luzonensis glass eels. VA, VB, VIA1, VIA2, VIA3 and VIA4 refer to Strubberg (1913) and Fukuda et al. (2013)
VA VB
VIA1
VIA2
VIA3
VIA4
27
Fig. 4 Identification of Anguilla glass eels based on morphology
A. bicolor pacifica
no tail pigment
A. japonica
fin difference <12%
A. luzonensis fin difference >13%
A. marmorata Short fin
Long fin
28
Fig. 5 Fin difference distribution with AM (Anguilla marmorata) and AL (Anguilla luzonensis). (X=Fin difference , Y=Catch)
0 200 400 600 800 1000 1200 1400
<8.5 8.5~9.5 9.5~10.5 10.5~11.5 11.5~12.5 >12.5
AL
0 200 400 600 800 1000 1200 1400 1600 1800
<12.5 12.5~13.5 13.5~14.5 14.5~15.5 15.5~16.5 16.5~17.5 >17.5
AM
29
Fig. 6 Sample collection from north Luzon Island from May.2008 to Sep. 2009 in (a) Cagayan river (CR) (b) Abulug river (AR) (c) Wangag river (WR)
(a)
Month
May Jun Jul Aug Sep Oct Nov Dec Jan. Feb. Mar. Apr. May. Jun. Jul. Aug. Sep.
Catch
0 100 200 300 400 500 600 700
CR:AL CR:AM 2008
30
(b)
Month
May Jun Jul Aug Sep Oct Nov Dec Jan. Feb. Mar. Apr. May. Jun. Jul. Aug. Sep.
Catch
0 50 100 150 200 250 300
AR:AL AR:AM 2008
31
(c)
Month
May Jun Jul Aug Sep Oct Nov Dec Jan. Feb. Mar. Apr. May. Jun. Jul. Aug. Sep.
Catch
0 100 200 300 400
WR:AL WR:AM
2008 2009
32
Fig. 7 Pigmentation stage v.s Total length (mm) of Anguilla luzonensis (a) Abulug river (b) Cagayan river (c) Wangag river (d) Taiwan (e) Philippines (3 rivers)
(a)
Pigmentation
5a 5b 6a1 6a2
TL(mm)
0 10 50 60
Abulug river
n=570 n=476
n=87 n=20
a a
b
b
(b)
Pigmentation
5a 5b 6a1 6a2 6a3
TL(mm)
0 10 50
Cagaya river n=465
n=505
n=154
n=54
n=3
a b
a
c 55
33
(c)
(d)
Pigmentation
5a 5b 6a1 6a2 6a3 6a4
TL(mm)
0 10 40 50
Taiwan n=53
n=79
n=7
n=2
n=2
n=4 55
34
(e)
Pigmentation stage
5a 5b 6a1 6a2 6a3 6a4
TL(mm)
0 10 40 50
1426 1565 362
170 44 11
a bc
ab
bc
c
abc 55
35
Fig. 8 Means Total length (mm) of A.luzonensis (VA) from May.2008 to Sep.2009 (a) Abulug river (b) Cagayan river (c) Wangag river (d) Abulug river+Cagayan river + Wangag river (VA+VB)
(a)
Abulug river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
42 44 46 48 50 52 54 56
A.luzonensis
36
(b)
Cagayan river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
42 44 46 48 50 52 54 56 58
A.luzonensis
37
(c)
Wangag river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
44 46 48 50 52 54 56 58 60
A,luzonensis
38
(d)
North Luzond Island
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
42 44 46 48 50 52 54 56 58 60
A.luzonensis(VA) A.luzonensis(VB)
39
Fig. 9 Means Total length (mm) of A.luzonensis (VB) from May.2008 to Sep. 2009 (a) Abulug river (b) Cagayan river (c) Wangag river
(a)
Abulug river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
44 46 48 50 52 54 56 58 60
A.luzonensis
40
(b)
Cagayan river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
42 44 46 48 50 52 54 56 58 60
A.luzonensis
41
(c)
Wangag river
Month
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
TL
42 44 46 48 50 52 54 56 58
A.luzonensis
42
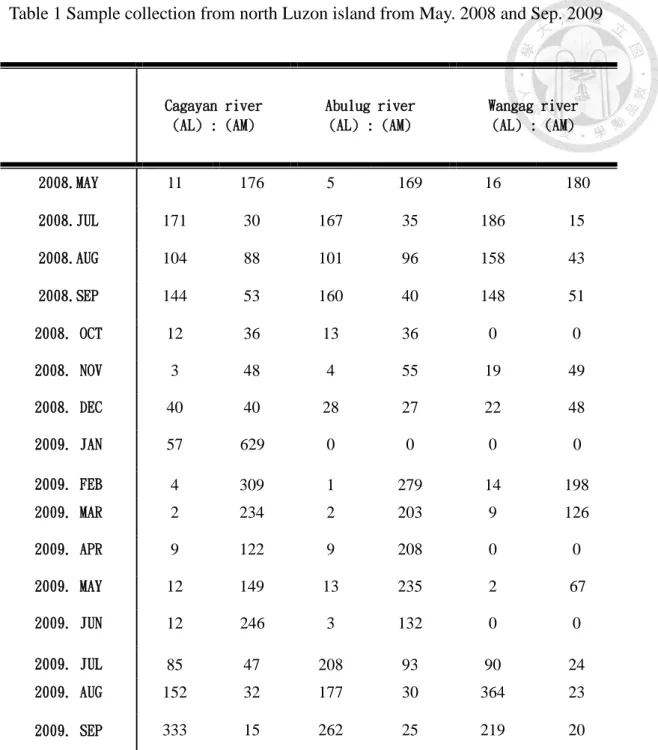
Table 1 Sample collection from north Luzon island from May. 2008 and Sep. 2009
Cagayan river (AL) : (AM)
Abulug river (AL) : (AM)
Wangag river (AL) : (AM)
2008.MAY 11 176 5 169 16 180
2008.JUL 171 30 167 35 186 15
2008.AUG 104 88 101 96 158 43
2008.SEP 144 53 160 40 148 51
2008. OCT 12 36 13 36 0 0
2008. NOV 3 48 4 55 19 49
2008. DEC 40 40 28 27 22 48
2009. JAN 57 629 0 0 0 0
2009. FEB 4 309 1 279 14 198
2009. MAR 2 234 2 203 9 126
2009. APR 9 122 9 208 0 0
2009. MAY 12 149 13 235 2 67
2009. JUN 12 246 3 132 0 0
2009. JUL 85 47 208 93 90 24
2009. AUG 152 32 177 30 364 23
2009. SEP 333 15 262 25 219 20
43
Table 2 Monthly composition of glass eel samples of A. marmorata (AM) and A.
luzonensis (AL) collected from Taiwan
Month AL AM
JAN 14 436
FEB 16 1247
MAR 3 1901
APR 1 1251
MAY 1 661
JUN 0 991
JUL 14 1439
AUG 11 872
SEP 19 1036
OCT 46 1177
NOV 20 1101
DEC 2 587
44
Table 3 Sample collection from Mindanao from Jan. 2009 and Apr. 2010
Month AL AM
JAN 0 195
FEB - -
MAR - -
APR 0 97
MAY - -
JUN - -
JUL - -
AUG - -
SEP 27 106
OCT 2 163
NOV 1 102
DEC 0 94
45
Table 4 Pigmentation stage of A.luzonensis from Abulug River, Cagayan River, Wangag River and Taiwan.
Abulug river Cagayan river
Wangag river Taiwan Pigmentation
5A 570 465 391 53
5B 476 505 586 79
6A1 87 154 121 7
6A2 20 54 96 2
6A3 0 3 41 2
6A4 0 0 11 4
6B 0 0 1 0
Total 1153 1181 1247 147
46
Table 5 Pigmentation and TL±SD of A.luzonensis from Abulug River,Cagayan River, Wangag River and Taiwan.
Abulug river Cagayan river Wangag river Taiwan
Pigmentation TL ± SD TL ± SD TL ± SD TL ± SD
5A 49.97±2.0a 50.49±1.8a 51.52±1.8a 50.27±2.8 5B 50.04±2.4a 49.79±2.2b 50.27±1.9b 50.94±2.9
6A1 51.8±3.4b 50.5±2.9a 49.27±1.7c 48.51±2.3
6A2 51.5±2.9b 51.46±3.1c 48.79±1.8c 46.6±3.3
6A3 - 50.5±3.0 49.15±2.3c 47.85±3.3
6A4 - - 50.22±2.6 53.12±1.6
6B - - 51 -
47
Table 6 Morphology of Anguilla luzonensis from Luzon Island,Taiwan and Mindanao
Morphology Abulug river Cagayan river Wangag river Taiwan Mindanao
TL(mm) 50.17±2.4a 50.24±2.3a 50.40±2.0a 50.38±3.5a 48.83±2.0b
Fin difference 10.77±1.0a 10.70±1.1a 10.61±1.1a 11.12±1.4b 10.36±2.1a
Total samples 1153 1181 1247 147 30
48
Appendix table 1. Data of Anguilla luzonensis eels used for analysis in this thesis.
Year Country Sampling sites Month n Total length Pigmentatoin
Mean ± SD VA VB VIA1 VIA2 VIA3 VIA4
2008 Philippne Abulug river May 5 48.4 ±2.3 0 3 1 1 0 0
2008 Abulug river Jul 167 49.2 ±1.7 0 165 1 1 0 0
2008 Abulug river Sep 160 51.5 ±1.7 159 1 0 0 0 0
2008 Abulug river Oct 13 54.5 ±1.3 4 9 0 0 0 0
2008 Abulug river Nov 4 48.6 ±1.7 4 0 0 0 0 0
2008 Abulug river Dec 28 51.6 ±1.6 24 4 0 0 0 0
2009 Abulug river Feb 1 50.5 1 0 0 0 0 0
2009 Abulug river Mar 2 49.5 ±0.7 0 2 0 0 0 0
2009 Abulug river Apr 9 48.6 ±1.2 9 0 0 0 0 0
2009 Abulug river May 13 49.4 ±0.6 13 0 0 0 0 0
2009 Abulug river Jun 3 48.6 ±1.0 3 0 0 0 0 0
2009 Abulug river Jul 208 48.8 ±1.4 205 3 0 0 0 0
2009 Abulug river Aug 177 48.9 ±1.7 67 109 1 0 0 0
2009 Abulug river Sep 262 49.9 ±1.9 81 131 40 10 0 0
2008 Cagayan river May 11 50.7±3.0 0 8 0 0 0 0
2008 Cagayan river Jul 171 48.0±1.6 0 111 52 8 0 0
2008 Cagayan river Aug 104 54.0±1.8 0 34 44 25 1 0
2008 Cagayan river Sep 144 51.2±1.6 133 10 1 0 0 0
2008 Cagayan river Oct 15 51.7±2.0 15 0 0 0 0 0
2008 Cagayan river Nov 2 48.5±0.7 2 0 0 0 0 0
49
Appendix table 1. Data of Anguilla luzonensis glass eels used for analysis in this thesis.
Year Country Sampling sites Month n Total length Pigmentatoin
Mean ± SD VA VB VIA1 VIA2 VIA3 VIA4
2008 Philippne Cagayan river Dec 22 51.1±1.4 8 14 0 0 0 0
2009 Cagayan river Jan 57 49.6±1.7 28 27 0 0 0 0
2009 Cagayan river Feb 6 50.5±1.7 6 0 0 0 0 0
2009 Cagayan river Apr 9 50.0±0.9 9 0 0 0 0 0
2009 Cagayan river May 12 49.6±1.0 11 1 0 0 0 0
2009 Cagayan river Jun 9 48.7±2.4 7 2 0 0 0 0
2009 Cagayan river Jul 199 50.3±1.8 197 2 0 0 0 0
2009 Cagayan river Aug 153 49.4±2.0 39 114 0 0 0 0
2009 Cagayan river Sep 228 49.8±1.5 0 155 52 20 1 0
2009 Cagayan river Dec 39 51.2±1.8 9 28 0 0 0 0
2008 Wangag River May 16 50.1±2.0 0 15 1 0 0 0
2008 Wangag River Jul 186 49.4±1.7 0 177 7 2 0 0
2008 Wangag River Aug 158 50.3±1.6 0 153 4 1 0 0
2008 Wangag River Sep 148 50.7±1.7 78 70 0 0 0 0
2008 Wangag River Nov 19 51±1.4 4 3 2 3 4 3
2008 Wangag River Dec 22 49.6±1.3 0 4 15 2 1 0
2009 Wangag River Feb 14 50.5±2.7 0 0 0 4 6 3
2009 Wangag River Mar 9 49.5±3.5 0 0 0 1 3 5
2009 Wangag River May 2 45.5±2.1 0 1 0 0 1 0
50
Appendix table 1. Data of Anguilla luzonensis glass eels used for analysis in this thesis.
Year Country Sampling sites Month n Total length Pigmentatoin
Mean ± SD VA VB VIA1 VIA2 VIA3 VIA4
2009 Philipine Wangag River Jul 90 52.0±1.6 87 3 0 0 0 0
2009 Wangag River Aug 364 49.8±2.1 92 92 84 77 19 0
2009 Wangag River Sep 219 51.4±1.9 130 68 8 6 7 0
2009 Taiwan Danshui River Feb 4 46.1±1.6 3 1 0 0 0 0
2009 Danshui River Apr 1 51.5 1 0 0 0 0 0
2009 Danshui River Jul 1 48 0 1 0 0 0 0
2009 Danshui River Sep 3 45.5±2.7 3 0 0 0 0 0
2009 Donggang River Jul 5 47.5±1.4 5 0 0 0 0 0
2010 Donggang River Mar 1 53 1 0 0 0 0 0
2010 Fengshan River Oct 1 45 0 1 0 0 0 0
2011 Fengshan River Jul 2 52.5±0.7 0 0 0 0 0 2
2011 Fengshan River Sep 2 52.2±4.5 0 0 0 1 0 1
2011 Fengshan River Oct 1 52 0 0 0 0 0 1
2011 Fengshan River Nov 1 45.5 0 0 0 0 1 0
2010 Yilan River Jul 4 46.9±2.8 1 1 0 1 1 0
2010 2010
Yilan River Aug 3 53.0±3.0 0 3 0 0 0 0
Yilan River Nov 6 49.3±2.1 0 5 1 0 0 0
2011 Yilan River Jan 5 49.1±1.9 2 3 0 0 0 0
2011 Yilan River Feb 10 52.0±2.0 1 8 1 0 0 0
51
Appendix table 1. Data of Anguilla luzonensis glass eels used for analysis in this thesis.
Year Country Sampling sites Month n Total length Pigmentatoin
Mean ± SD VA VB VIA1 VIA2 VIA3 VIA4
2010 Taiwan Yilan River Sep 1 48.8 1 0 0 0 0 0
2011 Yilan River Aug 1 47 0 1 0 0 0 0
2011 Yilan River Nov 1 49 0 1 0 0 0 0
2009 Siouguluan River Feb 1 49.5 0 1 0 0 0 0
2009 Siouguluan River Mar 2 55.5±0.7 0 2 0 0 0 0
2009 2009
Siouguluan River May 1 47.9 0 1 0 0 0 0
Siouguluan River Jul 1 53 1 0 0 0 0 0
2009 Siouguluan River Aug 2 49.5±0.7 2 0 0 0 0 0
2009 Siouguluan River Sep 1 46 1 0 0 0 0 0
2009 Siouguluan River Oct 2 46.2±0.3 2 0 0 0 0 0
2009 Siouguluan River Dec 2 47.2±1.7 1 1 0 0 0 0
2010 Siouguluan River Jan 1 47.5 0 0 1 0 0 0
2010 Siouguluan River Jul 1 45.5 0 1 0 0 0 0
2010 2010
Siouguluan River Aug 5 50.7±3.1 2 3 0 0 0 0
Siouguluan River Sep 11 50.1±3.0 7 2 2 0 0 0
2010 Siouguluan River Oct 42 51.8±2.3 5 35 2 0 0 0
2010 Siouguluan River Nov 12 51.4±2.8 7 5 0 0 0 0
2011 Siouguluan River Jan 8 50.6±3.0 1 7 0 0 0 0
2011 Siouguluan River Feb 1 55 1 0 0 0 0 0
52