糖尿病小鼠肺部感染不同群落的肺炎克雷伯氏菌之研究; The Studies of Pulmonary Infection by Klebsiella pneumoniae of Divergent Clusters in Diabetic Mice Model
107
0
0
全文
(2) 主防禦方面,DM 小鼠的巨噬細胞對 KP 具有正常的吞噬能力,凋亡 現象也與正常小鼠一致。為進一步研究 AM 與 hypo-或 hyper-mucoid KP 之間是否存在著不同的交互作用(interaction),因此利用老鼠的巨 噬細胞株(RAW 264.7 cell line)感染 hypo-或 hyper-mucoid KP,比較 KP 對巨噬細胞的存活力(viability)與凋亡(apoptosis)之作用,同時也比 較 KP 對巨噬細胞的附著(adhesion)與在巨噬細胞內的 KP 存活量 (macrophage-associated KP),結果發現,hyper-mucoid KP 對巨噬細胞 具較高的黏著量,甚至可以進入巨噬細胞中存活約一小時;而利用轉 移子突變(mini-Tn10 transposon mutagensis)方式將 hyper-mucoid KP 的 黏性去除,再予以感染巨噬細胞,則失去其原先的高黏著量,也無法 在巨噬細胞內存活。經由本篇的研究結果,說明了黏性特質影響細菌 與巨噬細胞間的交互作用, KP 的黏性特質是一個獨立致病因子, hyper-mucoid KP 具有較高的致病力;然而,DM 此因子一旦存在, 便使 hypo-mucoid KP 的致病力加成,不僅如此,hypo-mucoid KP 可 能存在著有別於黏性特質的致病因子。. 2.
(3) Abstract Diabetic (DM) patients are susceptible to Klebsiella pneumoniae (KP) infection, resulting in community- and hospital-acquired pneumonia and a high mortality rate in Taiwan. In the lungs, alveolar macrophages (AM) are pivotal for the defense against KP invasion. To investigate the effect of mucoviscosity and diabetes on virulence of KP between divergent cluster, in this study, C57BL/6J male mice was intraperitoneally injected with streptozotocin at 6 wk and DM was successfully induced with a blood sugar of ≧ 300 ml/dl at 8 wk. At 30 wk, the diabetic mice were instilled intratracheally with hypo- or hyper-mucoid KP from divergent cluster, and sacrificed 20 hr or 40 hr later. To study the clearance of KP in the lung and blood, the KP burden was determined by culturing the homogenate of left lung and the intra-cardiac blood. The lung burden of hyper-mucoid KP, after 20 hr of infection, was greater than that of hypo-mucoid one in control mice. Besides, the hypo-mucoid KP burden in the lung or blood of infected diabetic mice was more than that of control ones. The phagocytic ability and apoptosis of AM from right lung was assessed by Gram stain and FACScan analysis, respectively. The results showed that AM from diabetic mice possess normal phagocytic ability and their percentage of apoptosis was similar to that of control mice. Furthermore, to study the interaction between hypo-/hyper-mucoid KP and macrophage cell line, RAW 264.7 cells were infected with hyper- or hypo-mucoid KP. It showed that there was no difference in the percentages of cell apoptosis. However, hyper-mucoid KP adhere and invade into RAW 264.7 cells easier than hypo-mucoid one, and it survived longer in the host cell. Additionally, mini-Tn10 transposon mutagensis was introduced into wild type KP and resulted in a mutant KP with hypo-mucoid phenotype. RAW 264.7 cells were infected by this mutant KP, and thereafter, the adhesion and invasion of the mutant KP was decreased. These results suggested that the mucoid property was not only a key factor for KP to adhere and to invade macrophages, but also an independent KP virulent factor. On the other hand, DM played a important role for the infection by hypomucoid KP possibly making animals susceptible to hypo-mucoid KP infection.. 3.
(4) 誌謝 2002 的初秋,茫然與軟弱的我進入醫研所;而 2004 的仲夏,我 用希望、勇氣與熱情為這兩年劃下逗點,並朝著下一個更高深的學術 殿堂邁進。一路走來,最要感謝我的指導教授盧敏吉老師與蔡宗統老 師,謝謝你們耐心的指導與幫助。盧老師對研究的基本精神與實驗過 程的重視,以及樂觀幽默地面對問題的態度,使我學會了如何有系統 地去探索問題、分析問題及解決問題,更重要的是讓我具有勇氣與熱 情。蔡老師對科學的美感與創新獨到的見解,啟發我更寬廣與多元的 思考模式,也讓我對大自然有更深刻的體會。 非常感謝生理科的全體師長,給予我研究和實驗方面的建議與提 示,以及生理書報討論的教導與指正,經由書報討論的訓練,除了專 業知識的成長之外,也使我能夠對研究設計進行批判與條理分明的表 達意見。很謝謝李妙蓉老師、謝長奇老師在百忙中熱心的解說流氏細 胞儀,協助我順利的分析資料,而林文川老師寶貴的糖尿病動物模型 經驗之提供,使我的實驗能步上正軌,還要感謝賴怡琪老師對於細菌 實驗詳細的教導與特別的觀點,使我的研究能更加深入,也引發我對 細菌行為與宿主防禦濃厚的興趣,此外,邱乾順主任與其他口試委 員:盧敏吉老師、蔡宗統老師、李妙蓉老師、賴怡琪老師對於我論文 完整而專業的指正與建議,在此致上最深的感謝。. 4.
(5) 追求真理的路上,幸好有伴同行,在我茫然與軟弱時,有親愛的 學長姐與同學扶我一把,給我力量。謝謝美蘭學姐、家欣學長對實驗 的指導,特別感謝亦師亦友的程姐與靜華學姐,在我茫然的時候,給 我精闢的建議與溫暖的關心。也很感謝與我一起成長、前進的同學瓊 文、湯姐、瓊瑤、致芬、佩蒨、竹莞、玲瑜、慧淇、昆瀚與郁婷,特 別是佩玉,謝謝你的”於心不忍”,特別是信忠,感謝你的熱心,以及 ”用在刀口上的朋友”慧芬,謝謝你的”捨命陪君子”,在我軟弱的時 候,你們給我最大的幫助與鼓勵,此外,還有學妹怡均、怡蒨、曉嵐, 謝謝你們。 最重要的,我要感謝一直為我付出心力的爸媽與哥哥,謝謝你們 讓我無後顧之憂的求學,還要感謝年底將與我步上紅毯的未婚夫碩 爵,謝謝你的陪伴與支持,讓我可以盡情任性的研究,真的謝謝你。 真的謝謝大家,我會帶著你們送給我的禮物--希望、勇氣與熱情, 朝著追求真理的路上邁進,一直在心裡祝福著你們。. 5.
(6) 目錄 中 文 摘 要 ---------------------------------------------------------------------------1 英 文 摘 要 ---------------------------------------------------------------------------3 誌 謝 ---------------------------------------------------------------------------------4 目 錄 ---------------------------------------------------------------------------------6 圖 目 錄 ------------------------------------------------------------------------------9 略 字 表 -----------------------------------------------------------------------------12 第 一 章 緒 論 ---------------------------------------------------------------------14 壹、前言-----------------------------------------------------------------------14 貳、糖尿病(diabetes mellitus, DM)概述----------------------------------14 參、糖尿病和感染-----------------------------------------------------------19 肆、肺炎克雷伯氏菌(Klebsiella pneumoniae, KP)----------------------22 伍、肺泡巨噬細胞(alveolar macrophage, AM)------------------------27 陸、細菌的致病與巨噬細胞凋亡 (apoptosis)------------------------32 柒、研究目的-----------------------------------------------------------------34 第 二 章 研 究 設 計 與 方 法 ------------------------------------------------------36 壹、研究設計------------------------------------------------------------------36 貳、研究方法-----------------------------------------------------------------38 第一節 體內試驗(in vivo) ----------------------------------------------38 6.
(7) 一、糖尿病動物模型的建立----------------------------------------38 二、小鼠肺部感染 Klebsiella pneumoniae (KP) ----------------41 三、感染後小鼠左肺、右肺肺泡巨噬細胞 (broachoalveolar lavage)及血液的取得-----------------------------------------44 四、小鼠肺部及血液中 Klebsiella pneumoniae 菌量-----------46 五、肺泡巨噬細胞吞噬 Klebsiella pneumoniae 菌量之計數--47 六、肺泡巨噬細胞凋亡之偵測--------------------------------------50 第二節 體外試驗(in vitro) ----------------------------------------------51 一、RAW 264.7 細胞株感染 Klebsiella pneumoniae----------51 二、Klebsiella pneumoniae 所造成之細胞毒性(cytotoxicity)與 所誘發之細胞凋亡(inducing-apoptosis)------------------53 三、Klebsiella pneumoniae 對 RAW cells 的附著(adhesion)及 KP 在巨噬細胞內的存活量評估---------------------------55 四、Mini-Tn10 transposon mutagenesis---------------------------56 參、統計方法-----------------------------------------------------------------59 第 三 章 結 果 ---------------------------------------------------------------------61 第一節 體內試驗( in vivo ) ---------------------------------------------61 1-1. DM 小鼠實驗模式之建立-------------------------------------61 1-2. KP1004n (hypo-mucoid) 與 KP1004m (hyper-mucoid) 之. 7.
(8) 肺部感染-------------------------------------------------------62 1-3. Cluster A 的 paired KP (hypo-mucoid : KP1084/ hyper-mucoid:KP1112) 肺部之感染-------------------63 1-4. Cluster B 的 paired KP (hypo-mucoid : KP1008/ hyper-mucoid:KP2002) 肺部之感染----------------------65 1-5. Cluster C 的 paired KP (hypo-mucoid : KP1283/ hyper-mucoid:KP1284) 肺部之感染----------------------66 第二節 體外試驗(in vitro)---------------------------------------------67 2-1. Hypo-/hyper-mucoid KP 所造成之細胞毒性(cytotoxicity) 與所誘發之細胞凋亡(inducing-apoptosis) ---------------67 2-2. Hypo-/hyper-mucoid KP 對 RAW cells 的附著(adhesion) 及 KP 在巨噬細胞內的存活量評估------------------------68 2-3. 突變株與野生株 KP (mutant/ wild type KP) 對 RAW cells 的附著 (adhesion) 及 KP 在巨噬細胞內的存活量 評估--------------------------------------------------------------69 第 四 章 討 論 ---------------------------------------------------------------------71 第 五 章 結 論 ---------------------------------------------------------------------82 第 六 章 參 考 資 料 ---------------------------------------------------------------83 作 者 簡 歷 -----------------------------------------------------------------------107. 8.
(9) 圖目錄 圖一、Klebsiella pneumoniae 相關致病因子------------------------------92 圖二、Alveolar macrophage 之殺菌機制------------------------------------93 圖三、 糖尿病小鼠血糖值比較。控制組小鼠(con)與糖尿病組小鼠(DM) 第八週齡與第二十八週齡之血糖值--------------------------------94 圖四、糖尿病小鼠體重變化。控制組(con)小鼠與糖尿病組(DM)小鼠 各週齡體重紀錄--------------------------------------------------------95 圖五、 正常與 DM小鼠肺部感染 KP1004n (hypo-mucoid) 與 KP1004m (hyper-mucoid) 20 小時後,在肺部(A)及血液(B)中的 KP 菌落 數--------------------------------------------------------------------------96 圖六、 糖尿病小鼠肺泡巨噬細胞(AM)對 hyper-mucoid KP (KP1004m) 的吞噬量-----------------------------------------------------------------97 圖七、 正常與 DM 小鼠肺部感染 cluster A 的 paired KP (hypo-mucoid: KP1084/hyper-mucoid:KP1112) 20 小時後,在肺部(A)及血液 (B)中的 KP 菌落數-----------------------------------------------------98 圖八、Cluster A的 paired KP (hypo-mucoid:KP1084/hyper-mucoid: KP1112)對正常與 DM 小鼠肺泡巨噬細胞(AM)引發之凋亡--99 圖九、 正常與 DM 小鼠肺部感染 cluster A 的 paired KP (hypo-mucoid: KP1084/hyper-mucoid:KP1112) 40 小時後,在肺部(A)及血液. 9.
(10) (B)中的 KP 菌落數---------------------------------------------------100 圖十、 正常與 DM 小鼠肺部感染 cluster B 的 paired KP (hypo-mucoid: KP1008/hyper-mucoid:KP2002) 20 小時後,在肺部(A)及血液 (B)中的 KP 菌落數---------------------------------------------------101 圖 十 一 、 正 常 與 DM 小 鼠 肺 部 感 染 cluster C 的 paired KP (hypo-mucoid:KP1283/hyper-mucoid:KP1284) 20 小時後, 在肺部(A)及血液(B)中的 KP 菌落數--------------------------102 圖十二、 Cluster A 的 paired KP (hypo-mucoid:KP1084/hyper-mucoid: KP1112)感染老鼠巨噬細胞株 (RAW 264.7)所誘發之細胞凋 亡(inducing- apoptosis)-------------------------------------------103 圖十三、 Cluster A 的 paired KP (hypo-mucoid:KP1084/hyper-mucoid: KP1112) 感染老鼠巨噬細胞株 (RAW 264.7),KP 對細胞株 的 附 著 (adhesion) 與 KP 在 巨 噬 細 胞 內 存 活 的 菌 落 數 ------------------------------------------------------------------------104 圖十四、突變株 KP (mutant:KPG6)與野生株 KP (wild type:KP1112) 依照 MOI=1 與 MOI=10 分別感染老鼠巨噬細胞株 (RAW 264.7),KP 對細胞株附著(adhesion)之菌落數---------------105 圖十五、突變株 KP (mutant:KPG6)與野生株 KP (wild type:KP1112) 依照 MOI=1 與 MOI=10 分別感染老鼠巨噬細胞株 (RAW. 10.
(11) 264.7),KP 在巨噬細胞內存活的菌落數----------------------106. 11.
(12) 略字表 DM:diabetes mellitus KP:Klebsiella pneumoniae AM:alveolar macrophage BAL:bronchoalveolar lavage BALF:bronchoalveolar lavage fluid STZ:streptozotocin FACScan:flow cytometry NDDG:National Diabetes Data Group FPG:fasting plasma glucose IFG:impaired fasting glucose GAD:glutamic acid decarboxylase DKA:diabetic ketoacidosis NKHS:nonketotic hyperosmolar state PMNL, PMN:ploymorphonuclear leukocyte IL-1, IL-2, IL-6, IL-8 : interleukin-1, interleukin-2, interleukin-6, interleukin-8 TNF-a:Tumor necrosis factor a COPD:chronic obstructive pulmonary disease. 12.
(13) LPS:lipopolysaccharide CPS:capsular polysaccharide NADPH:reduced nicotine adenine dinucleotide phosphate MSR:macrophage scavenger receptor MMR:macrophage mannose receptor PFGE:pulse d-field gel electrophrosis PBS:phosphate buffer saline FBS:fetal bovine serum MOI:multiplicity of infection DAPI:4.6-dianmidin-2-phenylinodle INF-?:interferon gramma MIP-2:macrophage inflammatory protein-2. 13.
(14) 第一章 緒論 壹、前言 在台灣,糖尿病(diabetes mellitus, DM)盛行率逐年上升,住 院率更是居高不下(Wei et al., 2002),於民國 71 年至 90 年間, DM 一直位居國人前十大死因的第五名,根據行政院衛生署 91 年 6 月所公佈的資料中(行政院衛生署, 2002),DM 的死因順位已 由原先第五名晉升至第四名,其每千萬人口的死亡率高達 39.26,且其大多死於 DM 的併發症。DM 病人的感染問題一直 存在,尤其是 DM 病人很容易引發細菌性呼吸道感染,而且感染 期間很長 (Smitherman et al., 1995),特別是由肺炎克雷伯氏菌 (Klebsiella pneumoniae, KP) 所造成的肺部感染,其臨床病程進展 快速,且其死亡率高達 54 % (Ko et al., 2002)。由於台灣 DM 病 人容易感染 KP,因而這幾年國內的專家學者與感染科醫師紛紛 投入對 KP 的研究,以尋找 KP 的致病機制。 貳 、 糖 尿 病 (diabetes mellitus, DM)概 述 (一)定義 糖尿病(DM)是一種常見的內分泌疾病,其共同特徵是病人 具有高血糖值。導因於不同原因,造成病人體內胰島素相對或絕 對不足,或周邊組織器官對胰島素(insulin)產生抗性(resistance),. 14.
(15) 使細胞無法有效利用葡萄糖(glucose),進而造成高血糖。而長期 的高血糖,連帶影響許多器官及系統,產生腎臟病變,視網膜, 心臟血管及神經病變等(Carter et al., 1996)。糖尿病的定義,已經 由 National Diabetes Data Group (NDDG)及世界衛生組織(WHO) 在 1997 年修正如下:1)空腹血糖 (fasting plasma glucose, FPG) <110 mg/dl (6.1 mmol/L)是為正常 2)FPG 介於 110~126 mg/dl (6.1 mmol/L ~ 7.0 mmol/L), 是 為 impaired fasting glucose (IFG) 3)FPG ≧ 126 mg/dl (7.0 mmol/L),定義為 DM。另外,若隨機 血糖≧ 200 mg/dl (11.1 mmol/L),且合併糖尿病典型症狀(多 渴、多尿、體重減輕)也可診斷為 DM (Alberti and Zimimet, 1997)。 (二)分類 根據發病原因,可將 DM 分為 type 1、type 2 及其它特異型 三類,type 1 DM 可分為 type 1 A 及 type 1 B。 1. Type 1 A 此類病人以非裔美人或有家族史的亞洲人為主。他們出生時 體內胰臟β細胞的數目是正常的。但因不明原因,包括病毒感 染、環境等因素活化自體免疫系統,使β細胞受到破壞而逐漸減 少進而造成 insulin 分泌不足(Schranz et al., 1998)。Type 1A DM 涉及許多基因,他們大多有 HLA DR3 及 1 或 HLA DR40 而在. 15.
(16) HLA 基因位置 (locus)上 (Schranz., 1998),又以 DQA1*0301, DQB1*0302及 DQA1*501,DQB1*0201最為相關,可在 40 % type 1A DM 病人身上發現。胰島細胞自體抗體(ICAs)是由數種不同抗 體 組 成 , 他 們 會 攻 擊 胰 島 的 不 同 分 子 如 glutamic acid decarboxylase (GAD),insulin,IA-2/ICA512 等。而這些自體免 疫反應有:○ 1 產生自體免疫抗體○ 2 活化的淋巴球在胰臟胰島旁之 淋 巴 結 , 及 血 液 循 環 內 分 布○ 3 T- 淋 巴 球 的 增 生 ○ 4 釋出 cytokines。這些淋巴球在胰島細胞內浸的情況稱之為 insulitis。 雖然胰島內有包含β-cell 外的不同細胞(α-cell,δ-cell,PP-cell) 存在,但這些自體抗體似乎僅破壞β-cell 而已。 2. Type 1B 如同 type 1A,也是缺乏 insulin,但這些病人體內並沒有免 疫標誌 (immunology marker),因此造成β-cell 被破壞的原因並 不清楚。 3. Type 2 DM 導因於三大病態生理學特徵:周邊組織對 insulin 的抗性、 insulin 分泌減少,以及肝臟 glucose 製造增加(Cline et al., 1999)。 肥胖,尤其是軀幹性的肥胖,常會加重 insulin resistance (Gerich, 1998)。這是由於脂肪細胞會分泌許多生物性產物。如 leptin、TNF-. 16.
(17) α、游離脂肪酸改變了 insulin 的分泌、作用,體重增加甚至也 與 insulin resistance 相關。在疾病早期,血糖耐受性仍可維持正 常,這是因為 insulin 代償性的分泌增加。當這種代償機制無法 再維持下去時,血糖耐受性便開始下降。先是飯後血糖昇高,接 下來飯前血糖也會昇高。最終結果便是胰島細胞的衰竭。另外也 有研究發現,由於 GluT-4 transport 的缺損,造成了肌肉內 glycogen 合成異常,而成為 insulin 抗性的重要原因(Shulman, 1999)。Type 2 DM 有很強的基因遺傳性。雖然主要基因尚未被 確定,但可以肯定的是其為多基因多因子遺傳(Defornzo, 1997)。 4.其它型 DM 原因包括因基因缺損使 insulin 分泌減少或功能障礙,或個 體本身對 glucose 耐受性下降。胰島細胞若遭大量 (80 %) 破壞, 或其他內分泌疾病對 insulin 的作用產生拮抗,如肢端肥大症, 庫辛氏病 (Cushing’s disease) 等,也會使病人產生 DM。 (三)流行病學 在過去 20 年間,DM 盛行率大增。在 1976~1994 年間,在 美國的盛行率由 8.9 上升至 12.3 %。且 type 2 DM 增加的速度遠 比 type 1 DM 快(Rosenbloom et al., 1999)。Type 1 及 type 2 DM 的 分布有地域性。在 Scandinavia,有全世界最多的 type 1 DM 患. 17.
(18) 者,在芬蘭每年便有十萬分之三十五的發生機率;而太平洋地區 的 type 1 DM 少許多,在日本及中國,發生率約每年十萬分之一至 十萬分之三;北歐及美國的發生率約每年十萬分之七。Type 1 DM 的發生率反應了與其相關的 HLA 基因分佈的地域性,type 2 DM 的盛行率,在太平洋地區最高,印度及美國次之,而在蘇俄及中 國大陸最少。可能是由於遺傳、環境及種族不同有關。在台灣地 區,國人糖尿病盛行率隨年齡與年代的增加而升高。於 1996-2000 年之間,男女年齡別 DM 盛行率由 0-9 歲 0.4-1.2/1000 增加到≧ 70 歲 的 129-210/1000;五年間,所有年齡的男性盛行率介於 29-37/1000,女性則介於 38-46/1000。此外,男性歷年 DM 住院 率介於 18.6-20.0 %,女性則是 16.5-17.8 %。因此,台灣 DM 的 盛行率不僅逐年上升,住院盛行率更是居高不下,DM 是非常值 得大家正視的現代健康問題(Wei et al., 2002)。 (四)DM 併發症 DM 併發症可分為急性及慢性: 1.急性併發症 (1)糖尿酮酸中毒 (diabetic ketoacidosis, DKA):由於 insulin 的不 足及昇糖激素,兒茶酚胺,皮質醇,或生長激素過度分泌,使得 肝臟的糖質新生作用增加,且變成肝臟代謝途徑進而產生酮體。. 18.
(19) 其臨床表現為噁心、嘔吐、痛、多尿、呼吸困難及意識不清。 (2)高滲透壓、高血糖非酮酸中毒(nonketotic hyperosmolar state, NKHS):由於胰島素相對不足,且水份缺乏造成。由於 insulin 的不足,使糖質新生作用增加,組織利用 glucose 減少,加上高 血糖造成的利尿作用,使血管內容積嚴重缺乏造成。NKHS 的 insulin 缺乏程度較 DKA 為輕。其臨床症狀包括多尿、姿態性低 血壓,其中樞神經症狀,包括疲倦、嗜睡、遲鈍、癲癇,甚至昏 迷。 2.慢性併發症 DM 影響許多器官系統,且造成 DM 最常見到的死亡及罹病 原因,可分為血管性及非血管性。血管性又可分為巨血管性及微 血管性,巨血管性包括冠狀動脈疾病,週邊血管疾病(Grund et al., 1999);微血管性包括視網膜病變,神經病變與腎病變(Grundy et al., 1995)。非血管性,包括腸胃功能障礙、性功能障礙、皮膚病 史。除了上述合併症,容易受到感染也是 DM 病人常見問題。 參、糖尿病和感染 (一)DM 病人對感染的高感受性(high susceptibility) 有一些感染特別容易發生在 DM 的病人,甚至只有他們才會 得到;有些感染在 DM 病人身上會特別嚴重,其產生併發症的機 19.
(20) 會也較高。常見的感染有呼吸道感染、泌尿道感染、軟組織感染 等(Nirmal et al., 1999)。就呼吸道感染而言,DM 早期被視為獨立 的危險因子(Mackowiak et al., 1979),但近來有文獻提出不同看 法,認為 DM 並不是呼吸道感染的獨立致病因子,而是 DM 會增加 呼吸道對特定微生物的感受性,例如對金黃色葡萄球菌 (Staphylococcus aureus)、革蘭氏陰性菌等。使 DM 病人在某幾 種肺部的感染,發生率比一般人高,甚至致病率及死亡率也較增 加(Robbins et al., 1944; Helene et al., 2001)。在眾多微生物之中, 以細菌所引起的感染最為常見也持續較久 (Smitherman et al., 1995),例如在台灣因 KP 所引起的肝膿瘍(liver abscess)中便有 45 %至 75 %是 DM 病人(Wang et al., 1998)。因此,具有 DM 的病人 較容易感染某些特殊微生物,例如由 group B Streptococcus 所引 起的菌血症在 DM 的病人身上其盛行率便有 27.5 % (Farley et al., 1993);此外,有 30-60 %的 DM 病人會感染 KP 而引發菌血症 (Leibovici et al., 1991),肝膿瘍,眼內炎 (Chee et al., 1995)。. (二)糖尿病與免疫系統 DM病人對感染之所以具高感受性,乃是由於其免疫功能上 的缺損,目前認為高血糖會造成免疫方面的改變,如巨噬細胞在 高血糖的環境,其殺菌能力會下降(Nielson and Hindson., 1989), 20.
(21) 白血球抗微生物的能力亦降低,但良好的血糖控制卻可以改善免 疫功能,降低感染(Gallacher et al., 1995; McMahon et al., 1995)。 免疫系統的改變分成以下幾點討論: 1.先天性免疫(innate immunity) (1)多型核白血球(ploymorphonuclear leukocyte, PMNL, PMN): Type 1 和 type 2 DM 病人縱使其黏著因子表現增加,以及 產生較多的自由基,但是嗜中性白血球卻對自由基的刺激敏感 度 , 及 本 身 趨 化 能 力 (neutrophil chemotaxis)均 有 顯 著 下 降 (Delamaire et al., 1997)。此外,在吞噬作用、氧化呼吸作用 (oxidative burst) 和殺菌作用的功能也減弱;在細胞激素分泌方 面,細胞靜止狀態中分泌的 TNF-a 、 interleukin-6 (IL-6) 、 interleukin-8 (IL-8)皆會增加;一旦細胞受到刺激,嗜中性白血 球產生 IL-6 和 IL-8 的量就會比正常細胞少(Helene et al., 2001)。 (2)單核球(monocyte)及巨噬細胞(macrophage): 單核球與巨噬細胞的功能也有所改變。在早期的研究指 出 , 慢 性 糖 尿 病 大 鼠 (BB rat) 的 肺 泡 巨 噬 細 胞 (alveolar macrophage, AM )吞噬金黃色葡萄球菌的能力比正常的 AM 差,同時其殺菌力也減低(Sima et al., 1988)。 2.適應性免疫(Adaptive immunity). 21.
(22) 在體液免疫方面並沒有受到 DM 太大的影響,不僅具正常的 免疫球蛋白量,對於疫苗注射的反應也正常。但是,細胞免疫方 面卻有改變。在某些致病菌的刺激之下,例如金黃色葡萄球菌, 淋巴球的增生反應並沒有因此而增加,但對其他的致病菌,如白 色念珠菌 (Candida albicans)的反應卻很正常 (Deresinski et al., 1995)。 肆 、 肺 炎 克 雷 伯 氏 菌 (Klebsiella pneumoniae, KP) (一)一般特徵 肺炎克雷伯氏菌(Klebsiella pneumoniae, KP)是一種革蘭氏陰 性菌(Gram-negative),屬腸內桿菌科(Enterobacteriae),為伺機性 感染的致病原,從輕微的泌尿道感染到嚴重菌血症(bacteremia) 乃至具高死亡率的肺炎皆可由 KP 感染造成 (Bartlett et al., 1986)。KP 會造成急性且嚴重的肺炎與肺的破壞,由 KP 所引起 的肺部感染特色便是臨床病程進展快速,且經常合併肺膿瘍及多 肺葉的侵犯(Meyer et al., 1993),其痰液呈現紅醋栗色果醬(currant jelly)。KP 在革蘭氏染色(Gram stain)之下便有明顯表現,由於具 有莢膜(capsule),有些 KP 在培養皿中會形成黏性的菌株;KP 以 D 型 的 葡 萄 糖 (D-glucose) 和 檸 檬 酸 鹽 (citrate) 為 主 要 碳 源 (Podschun et al., 1998)。此外,R 因子(R factor)使其對 ampicillin、. 22.
(23) carbeuicillin 具抗藥性,並由於現今抗生素的廣泛使用,使得更 多具抗藥性的菌株出現。 (二)流行病學與高危險群 KP 是大葉型肺炎的重要致病菌,此外也會造成泌尿道感 染、肝膿瘍(liver abscess)、腦膜炎(meningitis)等,甚至導致菌 血症;有 9 %的泌尿道感染與 14 %的菌血症病人與 KP 相關, 僅次於大腸桿菌(E. coil; CDC, 1982)。KP 感染造成的死亡率平 均約在 35 % (Fine, 1996);而 KP 的感染常發生在免疫功能有問 題的病人,例如糖尿病者、正在服用抗代謝藥物或類固醇的病 人與酗酒者。此外,KP 造成的下呼道感染,除了上述危險因 子外,還包括慢性阻塞性肺病(chronic obstructive pulmonary disease, COPD),而且大多發生在 40 歲以上的病人(Pierce et al, 1974)。在台灣,社區感染(community-acquied infection)佔了 73 %;此與國外有所不同,在國外以院內感染(nosocomial infection) 居多,佔了 43-77 % (Yang et al., 2001)。根據美國疾病管制局統 計,克雷伯氏菌屬(Klebsiella spp.)佔了院內感染的 8 %及流行 性爆發的 3 %。在台灣,1920 年至 1960 年之間,KP 便被視為 造成社區性肺炎的重要原因(Carpenter et al., 1990),近十年來, KP 造成社區性感染的肝膿瘍與日俱增,由 30 %上升至 80 %,. 23.
(24) 同時有 70 %左右的病人具有 DM (Wang et al., 1998; Ko et al., 2002)。因此,有專家學者針對其 KP 族群研究,發現 Cluster A KP 是造成台灣肝膿瘍主要的 KP 族群,其相似度有 72.4 % ± 9.4 % (Lau et al., 2000)。由於台灣地區 DM 病人對 KP 具有很大的 感受性,而且會感染特定部位,使得 KP 感染被視為重要的公 共健康問題。 (三)重要致病因子(virulence factor) 關於克雷伯氏菌屬(Klebsiella spp.)的感染,經由體外與體內 實驗(in vitro and in vivo),得知某些細菌的因子可以引發疾病的 產生,這些因子有五種 (圖一),包括莢膜(capsule)、具黏附力的 纖毛(pili)、siderophores、血清抵抗(serum resistance)與脂多醣體 (lipopolysaccharide, LPS);而莢膜與脂多糖體在 KP 感染中是最 主要,也最被重視的致病因子(Williams et al., 1990)。 1.莢膜 (capsule) KP 通常會形成顯著的莢膜,其莢膜主要是由複雜的酸性多 醣體所組成稱之莢膜多醣體(capsular polysaccharides, CPS)。一般 而言,CPS 可被分為兩類,第一類富含尿酸(uronic acid),以其 為酸性組成,具有較大的分子量;第二類 CPS 的分子量低,其 酸性組成較具多樣性(Lai et al., 2003)。由於 CPS 上的某些酸性組. 24.
(25) 成使得 KP 具有黏性特質(mucoid),而黏性的程度則被認為與感 染的形成有關(Nassif et al., 1986),很多研究也證實 KP 的黏性 (mucoviscosity)是重要的致病因子,在臨床上所分離出具入侵性 的菌株大都具有黏性的特質(Fang et al., 2004)。在莢膜上造成黏 性特質的 colanic acid (M antigen),可由 rcsA 、rmpA 、rmpB 等 基因調控其表現,當活化 colanic acid 大量合成時,會使細菌表 現黏性(mucoid phenotype; Nassif et al., 1989; McCallum et al., 1991)。另外,K 抗原是莢膜上具有重複次單元的多醣類,其組 成大多是 4-6 個的糖類單元加上尿酸(uronic acid);目前,克雷伯 氏菌屬(Klebsiella spp.)K 抗原有 77 種(Ø rskov et al., 1984)。莢膜 包披在 KP 的最外層,有很多研究指出,莢膜對 KP 的致病力而 言是必要的,原因在於莢膜一方面可以保護 KP 免於受到 PMN 的吞噬(Podschun et al., 1992),另一方面又能使 KP 躲掉血清殺菌 因子 (bactericidal serum factors)的攻擊,成為 KP 重要的屏蔽 (Williams and Tomás, 1990)。在體內實驗發現,將大白鼠施打抗 KP 的 CPS 單株抗體,可以減少 KP 肺炎的出血與嚴重度,證明 了莢膜對於 KP 肺炎形成的重要性(Held et al., 1992);而在體外實 驗也發現 CPS 會抑制巨噬細胞的分化能力和功能(Yokochi et al., 1977)。有研究指出,造成呼吸道的感染,尤其是院內感染,可. 25.
(26) 能與莢膜上的第 1、3、4、5 型 K 抗原有所關聯;在另一實驗中 則認為第 1、2 型 K 抗原最具致病力,因為其缺乏 mannose-α -2/3-mannose 的結構,無法被 AM 辨認吞噬(kabha., 1995)。此外, 引起 DM 病人感染而造成肝膿瘍的 KP 莢膜血清型以 K1 為主, K2 次之(Fung et al., 2002)。 2.脂多醣體 (lipopolysaccharide, LPS) LPS 位在細胞壁上,組成包括脂質 A (lipid A)、核心糖脂 (core glycolipid)與 O 抗原 (O-polysaccharide antigen; Cortés et al., 2002);其中 O 抗原可以抵抗補體所調節的殺菌機制,因為 O 抗 原的側支鏈 (O side chain) 會伸展至莢膜外而曝露到外在環境 中,一旦遇上補體便會與之固定,阻止補體接近 KP 的細胞膜, 因而避開了補體的攻擊(Albertí et al., 1996)。另一個次單位脂質 A,進入宿主體內會引起發燒、改變白血球功能。一般而言,LPS 進入體內會活化補體及巨噬細胞,然而 KP 的 LPS 因 CPS 的屏 蔽而減弱了宿主免疫系統的活化。 除了上述二種致病因子外,KP 也可運用具黏附力的纖毛(pili) 黏附於宿主細胞表面而達到致病的第一步;此外,biofilm 此種外 層多醣體(exopolysaccharide)的形成亦可幫助 KP 抵禦抗生素與吞 噬細胞的攻擊。. 26.
(27) (四)KP 肺部感染 當 KP 入侵肺部時,會開始大量繁殖,在這同時,原始肺部 的防禦機制便是活化非特異性免疫,例如肺泡巨噬細胞(AM), 快速地將細菌由呼吸道清除掉(Lipscomb et al., 1995)。此外,KP 肺炎最明顯的臨床併發症便是 KP 會由肺泡散佈至血液中,宿主 一旦無法將 KP 由血液中清除,生物體內會累積大量內毒素,導 致多重器官功能失常與死亡(Thomas et al., 2002)。 伍 、 肺 泡 巨 噬 細 胞 (Alveolar macrophage, AM) (一)AM 簡介 肺泡巨噬細胞(AM)位在空氣和肺組織的界面間,為一種常 駐肺臟的單核吞噬細胞,也是下呼吸道對抗微生物入侵的第一道 宿主防禦。先天和後天免疫對於肺泡抗微生物而言,AM 才是最 主要的(William & Martin, 2000)。若將正常吸煙者進行肺泡灌流 (bronchoalveolar lavage),可發現沖出來的細胞大約 85 %是 AM, 7 %∼12 %是淋巴球,而嗜中性白血球只佔了不到 3 %,因此 AM 對於吸入性顆粒和微生物的清除,吞噬和殺菌是相當重要的 (Murray & Nadel, 2000)。除了吞噬和殺菌作用外,AM 也會分泌 一些化學物質,在發炎反應中扮演著調節的角色。大致上 AM 主 1 對抗入侵肺部的微生物和吸入性顆粒○ 2 清除 要的功能可包括:○ 27.
(28) 3 抑制不適當的免疫反應○ 4 抗原呈現及活化 肺部的表面張力素○ 發炎和免疫系統。 (二)AM 形態及來源 AM 的細胞直徑大小為 15-50 μm,細胞質核比例約為 3: 1,擁有許多質膜所形成的包含體(cytoplasmic inclusions);其中 最大特色便是具高密度的顆粒和粒線體,偶爾,有些 AM 會出 現板狀體的結構(lamellar bodies)。大多數的 AM 可以存活數週到 數個月,最後經由黏膜纖毛的清除或淋巴管離開肺部,甚至在肺 部死亡而被吞噬(Brain et al., 1977)。存在身體各組織器官的巨噬 細 胞 乃 由 骨 髓 幹 細 胞 (myeloid stem cell)衍 生 成 類 骨 髓 細 胞 (myeloid),再分化成單核球(monocyte)釋放到血液中循環(Furth et al., 1970)。這些單核球會進入到肺或其他器官,隨著不同器官 環境的刺激下,單核球分化成巨噬細胞;由於進入肺的單核球處 於具表面張力素和富含磷脂、高氧環境之下,而使 AM 的生物 特性有別於其他的巨噬細胞(Shellito and Kaltreider, 1984)。此 外,在重氫標定胸腺嘧啶的實驗中,可以得知 AM 在肺部雖有 複製能力,但卻只佔 0.35 %∼1.25 %的比例。由此可知,AM 的 來源大多是由單核球進入肺部分化而來(Golde et al., 1974)。 (三)代謝. 28.
(29) 多數的單核球和巨噬細胞是偏向厭氧的,然而 AM 卻是進 行需氧代謝(Simon et al., 1977)。一般巨噬細胞行吞噬作用和吞噬 後引發的呼吸性作用(respiratory burst),亦即經由 NADPH 氧化 ? 和己醣單磷酸代謝途徑(hexose monophosphate shunt)。由於對 於氰化物並不敏感,因此其能量消耗主要依賴醣解作用 (Cline and Lehrer, 1968)。但是,AM 的能量來源卻是依賴氧氣和氧化代 謝。在 AM 平時休息狀態便有很高的 H2O2 產生,而吞噬後所耗 費的氧氣及代謝更是明顯增加(Babior, 1984)。 (四)抗微生物的機制 1.活化 一般而言,單核球和巨噬細胞可以很快地被活化,而達到 毒殺微生物(microcidal)和毒殺腫瘤(tumorcidal)複雜的生物代謝 活性(Adams and Hamilton, 1984)。引發巨噬細胞活化的因子有 宿主本身產生的生理因子例如細胞激素(cytokines),以及外來環 境因子例如微生物,會經由不同的活化途徑造成巨噬細胞不同 的活化狀態。巨噬細胞的活化狀態可包括吞噬作用和產生細胞 毒性狀態,免疫和細胞抗原呈現狀態及特定性分化狀態 (Hoffman, 2000)。因此,一但吸入性顆粒和微生物進入肺部則 會引發 AM 的活化,將其清除消滅。. 29.
(30) 2.吞噬作用 AM 的吞噬作用對於宿主防禦入侵的微生物而言是很重要 的。行吞噬作用等吞噬粒子的理想大小則介於 1~3 μm 之間 (Camner, 1980)。在 AM 的表面具有許多接受器,與吞噬作用有 關的接受器有: (1)清道夫接受器(macrophage scavenger receptors, MSRs):可以 結合及胞攝低密度脂蛋白(low-density lipoproteins),有些 MSRs 則可和胞外基質或細菌的脂台酸(lipoteichoic acids)、 LPS 結 合,而引發吞噬作用。 (2)免疫球蛋白 Fc 接受器(immunoglobulin Fc-receptors):可與經 抗體調理過粒子與微生物結合並吞噬。一旦與其結合後,AM 會釋出許多細胞激素如 TNF 及引發呼吸作用產生超氧離子 (superoxide; Foreback et al., 1997)。 (3)辨識補體的接受器(complement receptors):會和補體、微生物 複合物的補體部位結合進而促進吞噬作用。 (4)甘露糖接受器(macrophage mannose receptors, MMRs):可調 節內攝,吞噬及細胞毒性。例如有些莢膜具有重複 mannose-α -2/3-mannose 結構的克雷伯氏菌 (Klebsiella pneumoniae)易被 AM 的 MMRs 辨識及結合,進而引發 AM 的吞噬(kabha, 1995)。. 30.
(31) (5) CD14:會和 LPS 結合,調節吞噬並引發訊息傳遞(signal transduction),形成引起發炎反應的細胞激素(pro-inflammatory cytokines)。 當外在分子結合到 AM 表面,在分子上的 ligands 會和 AM 上的接受器產生交互作用,AM 的細胞膜沿著此分子展開,藉 著膜下的肌動蛋白 (actin)及 微 絲 (microfilaments)將 此 分 子 噬 入,形成了吞噬小泡(phagosome),如由 Fc 接受器所產生的吞 噬小泡。吞噬作用可被分類成二種:需經調理及不需調理的吞 噬(opsonic and nonopsonic phagocytosis)。在人類及小鼠的 AM 中,其 C3b/iC3b 接受器相當小量,甚至小鼠的 AM 不具 IgG, Fcγ的接受器(Berger et al.,1994; Stokes et al., 1998);此外,在 下呼吸道出現抗體及補體的量也很少,因此 AM 消滅肺部的致 病原最有可能的方式便是利用非調理性吞噬 (Doroth et al., 2000)。 3. AM 的殺菌機制 有二種方式可以達到殺菌的效果,一為溶小體(lysosome)內 酵素活化,另一為超氧離子(superoxide)的產生,而最佳的殺菌 能力便是兩者的協調(Mandell, 2000)。當免疫複合物,細菌內毒 素或其它發炎因子與細胞表面接受器結合便會觸發 respiratory. 31.
(32) burst (Pizzorno, 1999),由訊息傳遞(signal transduction)引發質膜 上 NADPH 氧化? 進行氧化還原產生超氧離子,並釋放到吞噬 小泡內或細胞外將致病原殺死(圖二) 。另外,在吞噬小泡形成 後 , 會 很 快 地 與 溶 小 體 (lysosome)融 合 , 形 成 次 級 溶 小 體 (secondary lysosome or phagolysosome)。若 PH= 5 時,會使其內 43 種酵素活化而能殺菌及消化物質(Nyberg et al., 1994)。不僅 如此,在溶小體(lysosome)內的 myeloperoxidase 也可將 H2O2 和氯離子轉換成 hypochorite,更加強了殺菌能力。 4.分泌化學物質和抗原呈現 巨噬細胞一旦活化便會分泌引起發炎反應的細胞激素 (pro-inflammatory cytokines),例如 TNF-a、IL-1、IL-6、MIP-1/2、 INF-? 等,可誘導顆粒性白血球和單核球進入肺泡內以及活化其 它免疫系統;此外,也可由吞噬作用將抗原處理過,呈現給 T 細胞辨識,啟動進一步的免疫防禦。 陸 、 細 菌 的 致 病 與 巨 噬 細 胞 凋 亡 (apoptosis) 經由體外實驗(in vitro),很多細菌的致病因子已被認為是導 致細胞凋亡的調節物質(mediator);細菌經由內毒素,或其它致 病因子與宿主細胞死亡相關的基因交互作用而造成宿主細胞的 凋亡(Chen et al., 1994)。一般而言,宿主免疫細胞的凋亡,尤其. 32.
(33) 是單核吞噬細胞,如 PMN 或 AM 等凋亡,對細菌來說是有利的。 某些細菌會藉由誘發細胞凋亡的方式殺死巨噬細胞,克服巨噬細 胞的吞噬與殺菌機制,而達到細菌散播的目的(Hilbi et al., 1997)。除了藉由誘發凋亡作為破壞宿主及其防禦機制之外,有 些細菌,也會藉凋亡而觸發發炎反應,破壞宿主的組織,例如: 志賀氏桿菌(Shigella),其 IL-1ß 轉換? (interleukin 1 converting enzyme)的活化,不僅造成宿主細胞凋亡,同時會切割 IL-1,使 其 活 化 並 由 宿 主 細 胞 釋 出 ,大 量 破 壞 宿 主 細 胞 造 成 赤 痢 (Sansonetti et al., 1995)。另外,有一些胞內細菌 (intracellular bacteria),由於與巨噬細胞共存,因此會抑制細胞的凋亡(Colotta et al., 1992),此時,巨噬細胞的凋亡對宿主而言卻是防禦的機 轉。在分枝桿菌(Mycobacteria)感染的實驗發現,巨噬細胞凋亡 的數目愈多其存活的分枝桿菌數目便愈少(Molloy et al., 1994)。 有趣的是,另一種分枝桿菌的感染則會活化巨噬細胞的 Fas 接受 器而誘發凋亡,在這同時也使細胞的殺菌機制無法啟動,讓細胞 的凋亡不具防禦功能( Elkon and Kaplan, 1996)。因此,細菌誘發 巨 噬 細 胞 的 凋 亡 具 有 不 同 的 意 義 , 但 就 胞 外 菌(extracellular bacteria) 而 言 , 引 發 巨 噬 細 胞 的 凋 亡 是 一 種 有 效 的 武 器 (Zychlinsky et al., 1997)。. 33.
(34) 柒、研究目的 先前的研究發現,cluster A 的 KP 是造成台灣 KP 肝膿瘍的 主要族群(Lau et al., 2000);而我們實驗室由不同疾病的檢體,收 集近五百株 KP 進行 PFGE (pulse d-field gel electrophrosis)分群, 分析出相似的 cluster A KP,以及其他的 cluster B、C… 等,也有 不屬於 cluster 內的 KP。在台灣,KP 造成的社區性肺炎的死亡 率高達 54 % (Ko et al., 2002),然而造成 KP 呼吸道感染的致病機 制及 DM 病人對 KP 的高感受性和 KP 的致病因子,仍尚未清楚。 近年來,許多學者針對 KP 的黏性特質進行研究,發現經過 突變而成為 hypo-mucoid KP 便不具致病力 (Fang et al., 2004)。 由於目前皆以突變的 KP 菌株研究 KP 的黏性,為避免因突變而 影響 KP 某些未知的生物特性或功能,本實驗選擇以野生型(wild type)臨床分離菌株,亦即由 cluster A、B、C 中挑選出基因相似 度最高、黏性特質差異最大的成對 KP (paired KP)進行黏性的致 病研究。為探討不同 cluster KP 的黏性特質在糖尿病與正常血糖 動物的致病力,將來自不同 cluster 的成對 KP 分別感染正常與糖 尿病小鼠的肺部,觀察 20 或 40 小時後動物肺部 KP 的清除情形, 同時進一步探討 KP 是否有散佈至血液中;更進一步,也將成對 的 KP 分別感染老鼠的巨噬細胞株,探討 KP 與巨噬細胞之間的. 34.
(35) 交互作用(interaction);藉由來自不同 cluster 的成對 KP 進行體內 (in vivo)試驗與體外試驗(in vitro),比較黏性特質及糖尿病的致病 作用,試圖找出 KP 的致病機制。. 35.
(36) 第二章 研究設計與方法 壹、研究設計 此研究在 cluster A、B、C 與不在 cluster 之內的 KP 中,挑 選出經 PFGE 方式分析,其間 PFGE pattern 的相似度約 98 %, 以黏度測試 (string test)區分黏性,一為低黏性(hypo-mucoid),另 一為高黏性(hyper-mucoid)的成對 KP (paired KP),進行體內試驗 (in vivo)與體外試驗(in vitro)。 1. Cluster 的定義:以 PFGE 方式將 KP 基因進行分析,當其 PFGE pattern 相似度高達 80 %以上便成為一個 cluster。 2. 成對 KP 定義:在同一個 cluster 中所挑選出 PFGE pattern 有 98 % 相似度的 hyper-mucoid 和 hypo-mucoid KP 便稱為成對 KP。. cluster Paired KP: hypo-mucoid hyper-mucoid. 第 一 節 體 內 試 驗 (in vivo) 將低黏性(hypo-mucoid)與高黏性(hyper-mucoid)基因相似度. 36.
(37) 高達 98 % 成對的 KP,分別以菌量 1×104 cfu 感染正常與糖尿病 小鼠,於 20 或 40 小時將其犧牲。將小鼠右肺進行支氣管灌流術 (bronchoalveolar lavage),將肺泡巨噬細胞(AM)沖出,探討 KP 入侵肺部後引起 AM 的凋亡(apoptosis)與吞噬(phagocytosis)情 形;為了解 KP 在肺部清除的情形,將小鼠左肺均質,培養細菌; 同時收集小鼠心臟血液,進行細菌培養,可知 KP 是否自肺部散 佈至血液中。以此比較黏性特質與糖尿病的致病作用。 第 二 節 體 外 試 驗 (in vitro) 雖然 KP 被視為胞外菌,但有研究指出 KP 可以入侵肺部上 皮細胞內並引發其凋亡(Oelschlaeger et al., 1997;Cortés et al., 2002) 。因此將低黏性(hypo-mucoid)與高黏性(hyper-mucoid)基因 相似度高達 98 % 成對的 KP,分別以 MOI (multiplicity of infection;1 或 10) 感染老鼠巨噬細胞株(RAW 264.7 cell lines), 比較 KP 對巨噬細胞的存活力(viability)與凋亡 (apoptosis)之影 響,同時也比較 KP 對巨噬細胞的附著(adhesion)與 KP 在巨噬細 胞內的存活量(macrophage-associated KP)。探討黏性特質是否具 細胞毒性,以及黏性特質是否會影響 KP 對巨噬細胞的附著與 KP 在巨噬細胞內的存活量。 另外,有實驗提出,莢膜多糖體(CPS)厚度會阻礙 KP 對上. 37.
(38) 皮細胞的附著與入侵(Sahly et al., 2000),但有些學者持不同看 法,認為莢膜多糖體(CPS)是最早與宿主細胞互動的細菌成分, 也和 KP 群體間之聚集有關(Sabine et al., 1999);因此,為探討莢 膜多糖體(CPS)黏性特質對細菌與巨噬細胞的黏附、在巨噬細胞 內 的 存 活 是 否 重 要 , 利 用 轉 移 子 突 變 (miniTn10 transposon mutagenesis)的方式將 hyper-mucoid KP (KP1112)黏性特質去除, 將野生型(wild type)與突變型(mutant type) KP 分別感染老鼠巨噬 細胞株,比較兩者對巨噬細胞的附著與在巨噬細胞內的存活量。 貳、研究方法 第一節 體 內 試 驗 (in vivo) 一、糖尿病動物模型的建立 * 材料: 檸檬酸鹽 (citric acid; C6H5Na3O7; USB)、微量天平、二次水 (ddH2O)、1N & 12N 氫氧化鈉 (NaOH)、1N & 12N 鹽酸 (HCl)、 50 ml 離心管(5 ml centrifuge tube )、塑膠滴管(dropper)、小燒杯、 拭淨紙(wipes)、酸鹼側定儀(PH Vision, Jenco, Electronic; LTD)、 pH4、pH6.8 校正液,試管震盪器(vortex mixer)、PE 飼養盒、木 屑墊料、鼠用飼料、餵水用水瓶、一次水、磅秤、streptozotocin (N-(Methylnitrosocarbamoyl)-o-glucosamine; Sigma) 、 0.05 M ,. 38.
(39) pH4.5, 4℃的檬酸緩衝溶液(citrate buffer)、 吸 管 唧 筒 (pipette pump)、微量吸管套(blue tip)、1 ml 注射用針(1ml syringe)、5 ml 試管、葡萄糖分析儀(Model 1500 sidekick glucose analyzer; YSI)、 校正液(YSI 2747 standa rd;YSI)、25μl 鈍針加樣器(YSI 151; YSI)、膠帶、報紙、透明小塑膠袋、麻醉用玻璃鐘罩、麻醉用小 罩杯、乾棉球、止血小夾、70%酒精棉、水、小燒杯、乙醚(ethyl ether)、解剖台、解剖用剪刀。 * 步驟: 1-1. 實驗動物之品管 1.動物品系:C57BL/6J male mice 2.動物來源:國家科學委員會-國家實驗動物繁殖及研究中心 3.動物週? :5 週 4.動物分組:將 5 週大的 mice 分為對照組與糖尿病組 5.飼養環境:恆溫 22℃、空調房間,相對溼度 55%、12 小時照 明 1-2. 0.05M, pH 4.5 檸檬酸鹽緩衝溶液(citrate buffer)的配製 加入 10ml dd H2O,加入 2.5 mmole citric acid、vortex 均勻, 加入 ddH2O 至 30 ml,vortex 均勻,先後以 pH4、pH6.8 校正酸鹼 測定儀,以 ddH2O 清洗探針,以鏡紙擦乾探針,測量 sodium citrate. 39.
(40) buffer 的 PH 值,以 NaOH、 HCl 滴定之,直到 pH4.5,酸鹼互 換間,須以 ddH2O 清洗探針,以拭鏡紙擦乾探針,最後加入 ddH2O 至 50 ml,其 pH 應為 4.5。 1-3. 糖尿病的誘發 將 PE 盒底部鋪滿墊料,蓋子上放置飼料及裝水小瓶,把小 鼠放在 PE 盒中飼養 1 week,任其自由進食飲水,將 6 週大的小 鼠 秤 重 , 糖 尿 病 組 依 小 鼠 的 體 重 酌 取 0.055 mg/ml/g 的 streptozotocin (STZ)的量,注射先將 streptozotcin 溶於 0.2 ml 的 citrate buffer,稀釋後至少 2 分鐘後再讓小鼠接受注射(但是泡藥 後要盡快完成注射以防 streptozotocin 失去活性) ,小鼠接受連續 4 天腹腔的 streptozotocin 注射;而對照組小鼠則依照體重取與糖尿 病組相等量之 citrate buffer 腹腔注射 4 天。在連續四天腹腔注射 完成後的第七天測量小鼠血糖值,血糖值必須≧300 mg/dl 才視為 有意義之糖尿病誘發,若血糖值未超過 300 mg/dl,則再連續給予 小鼠兩天的 0.055 mg/g 的 streptozotocin 注射,同樣的,在兩天注 射 streptozotocin 完成後的第七天測量小鼠血糖值,血糖值必須≧ 300 mg/dl,糖尿病誘發後養育 6 個月至小鼠週齡為 30 週左右, 於小鼠週齡為 28 週時再進行一次血糖測量,每週秤一次體重並 記錄。. 40.
(41) 1-4. 血糖的測量 準備材料,鋪報紙,注入乙醚於玻璃鐘罩及麻醉小罩杯,校 正儀器,校正液值為 180 mg/dl,用水清洗 pipette 後,用拭淨紙 擦乾,將小鼠放入玻璃鐘罩中麻醉,小鼠昏迷後,將其平躺置於 解剖台上,小鼠四肢以膠帶固定,若小鼠有醒來的跡象,以麻醉 小杯蓋住其口鼻,70 %酒精棉消毒小鼠尾巴,用剪刀剪斷一小 段尾巴,擠出 5∼6 滴血(>25μl)於透明平滑塑膠帶上,以鈍針加 樣器吸收取 25μl mice 全血,將全血打入儀器測量血糖值,清洗 pipette 後,用拭鏡紙擦乾,以乾棉球及止血夾夾住尾小鼠尾巴止 血,記錄血糖值。 二 、 小 鼠 肺 部 感 染 Klebsiella pneumoniae (KP) * 材料: Klebsiella pneumoniae (KP)、酒精燈、70 %噴霧酒精、擦手 紙、裝 70 %酒精的小燒杯、微量離心管(1.5 ml micro centrifuge tube; eppendrof)、微量離心管放置架(eppendorff rack)、15ml 無菌 培養血清瓶、試管架、無菌 Tryptic Soy Broth (TSB broth; Becton Dickinson)、無菌 LB agar, Miller Plate (Becton Dickinson)、電動吸 取器(Electrical Pipette)、無菌吸管(5 ml pipettes)、無菌的一倍磷酸 鹽緩衝溶(1X PBS)、1000p、200p、20p吸管唧筒(pipette pump)、. 41.
(42) 吸管尖(blue & yellow tip)、白金 loop、三角玻璃棒、無菌通風櫥、 37℃培養箱(incubator),10 倍連續稀釋的 KP 菌液、廢物桶、廢 液桶、試管架、30 週 C57BL/6J 雄性小鼠、乙醚(ethyl ether)、麻 醉用玻璃鐘罩、無菌乾棉球、膠帶、縫線、縫針、持針器、解剖 盤、麻醉小罩杯、優碘、彎盆、剃毛機、試管震盪器(vortex mixer)、 手術燈、鈍頭鑷子、尖頭鑷子、尖頭小剪刀、圓頭剪刀、70 %噴 頭酒精、無菌生理食鹽水(normal saline)、50μl 微量注射針筒(50 μl syringe)、27 號針頭的 1ml 無菌注射針、104 cfu/30μl 濃度之 KP、冰塊,冰桶。 * 步驟: 2-1. Klebsiella pneumoniae (KP) 1.KP 來源:中國醫藥大學附設醫院之臨床病人分離之 KP 菌株(中 國醫藥大學附設醫院湯惠玲小姐提供) 2.KP 種類: (1) Cluster A: KP1084 與 KP1112 (2) Cluster B: KP1008 與 KP2002 (3) Cluster C: KP1283 與 KP1284 (4) 不在 cluster 之內的 KP: KP1004 因 plasmid 的有無,而有兩 種不同的表現型,一為高黏性另一是低黏性,KP1004n 與. 42.
(43) KP1004m 表列如下: cluster. Paired KP. String test. DM history. KP1004 (不在cluster 之內). KP1004n. -. +. KP1004m. +. +. A. KP1084. -. +. KP1112. +. -. KP1008. -. -. KP2002. +. +. KP1283. -. +. KP1284. +. +. B. C. 2-2. 定量 Klebsiella pneumoniae 菌液濃度 以下步驟必須在無菌通風櫥中操作,用 loop 取 1 個 colony 的 KP,將 KP 種入 3ml TSB broth,混合,放入 37 ℃培養箱, 培養 18 小時後,作 KP 菌液的 1:10 連續積釋,準備 10 支標記 號碼的微量離心管,各加入 900μl 無菌 PBS,取 100μl 含 KP 的 broth 置入 900μl 無菌 PBS,混合,重覆上述步驟,共 10 次 1:10 連續稀釋,準備 10 盤標記號碼的 LB agar plate,每管稀釋 菌液,混合均勻,各取 20μl 滴加在 LB agar plate 上,以三角 玻璃棒均勻塗抹,三角玻棒泡在 70 %酒精,取出後陰乾,在火. 43.
(44) 焰上反覆燒烤三次,並冷卻之,再塗抹下一盤,將塗抹好稀釋 菌液的 LB agar plate,放入 37 ℃培養箱,培養 18 小時後,計 數並記錄 plates 上的 colony 數目,經計算並回推每管稀釋菌液 的菌量,之後取 10-3 此管稀釋菌液,以 PBS 做進一步稀釋而達 到 104 cfu/30μl。 2-3. Klebsiella pneumoniae (KP) 肺部之感染 器皿以 70 %酒精消毒,以無菌 normal saline 沖洗殘餘酒精, 器械泡於無菌 normal saline,微量針以 KP 菌液沖洗一次,KP 菌 液 vortex 均勻,以微量針抽好 30μl KP 菌液,乙醚麻醉小鼠,不 可麻死,用膠帶將小鼠四肢固定在解剖板上,將小鼠門牙以線綁 住,拉直小鼠頭部再用膠帶固定,剃掉小鼠頸部的毛後,以優碘 消毒小鼠頸部皮膚,將小鼠頸部皮剪開,用鑷子將氣管上方的肉 和胸腺小心撥開,將氣管上的膜拉高,剪開膜露出氣管,暴露出 氣管後,用鑷子將氣管游離出並撐住,以 27 號針頭注射入 30μ KP,共 104 cfu,讓小鼠直立 2 min,將小鼠頸部的皮縫合,再以 優碘消毒小鼠頸部皮膚並放回飼養盒,使其自由進食進水,飼養 20 小時或 40 小時。 三、感染後小鼠左肺、右肺肺泡巨噬細胞(bronchoalveolar lavage)及血 液的取得. 44.
(45) * 材料: 感染 KP 20h 或 40 h 之後 C57BL/6J 雄性小鼠、乙醚(enthyl ether)、麻醉用玻璃鐘罩、無菌乾棉球、膠帶、縫線、止血鉗、解 剖盤、麻醉小罩杯、優碘、彎盆、鈍頭鑷子、尖頭鑷子、尖頭小 剪刀、圓頭剪刀、70 %噴頭酒精、無菌生理食鹽水(normal saline)、 無菌一倍磷酸鹽緩衝溶液(1X PBS)、含 EDTA 的抗凝管、離心管 (15ml centrifuge tube)、微量離心管(1.5ml micro-centrifuge tube)、 離心管(15ml & 50 ml centrifuge tube)、PE 管(polyethylene tube; PE-50),21 號針頭(21gauge needle)、針筒(1ml syringe)、26 號針 頭的 1ml 無菌注射針,冰塊、冰桶。 *步驟: 將事先滅菌的器皿以無菌 normal saline 倒入適量,器械泡於 無菌 normal saline 中,小鼠感染後 20 小時,乙醚麻醉小鼠,以下 手術部分皆在無菌通風桶中操作,將小鼠胸腔打開,將胸骨完全 剪開,以止血鉗將肋骨往左右扳開固定,,以 1 ml 空針插入心臟, 採集 0.2 ml 全血,將全血置入己含 EDTA 的抗凝管中並混合之, 全血靜置於冰上,剪開頸部皮膚,並用 2 隻尖鑷子撥開,找到甲 狀腺、氣管,小心仔細的扯開氣管上的肉,將氣管上的膜拉高, 剪開露出氣管,暴露出氣管後,用鑷子將氣管撐住,鑷子夾線,. 45.
(46) 從氣管下方穿線,將小鼠心臟拉高剪去,將其左肺以鈍端鑷子拉 出後,可看到左支氣管,把左肺拉出並用止血鉗將左支氣管夾 緊,左支氣管用縫線綁兩個死結,剪下左肺,以無菌 normal saline 沖過並放入含有 3 ml PBS 的高速離心管中,將左肺靜置於冰上, 接續進行支氣管灌流術(bronchoalveolar lavage;BAL),用尖剪刀 在鑷子下方(較靠近頭的方)替氣管剪一小洞,穿入 PE tube,PE tube 綁線固定,用膠帶在桌上固定 PE tube,插上抽好 0.6 ml PBS 的針筒,慢慢打入 PBS,看到右肺脹起來,再慢吸回 PBS,重複 灌洗 0.6 ml PBS,最後總共灌出 15 ml 灌洗液(BALF),肺泡灌洗 液靜置於冰上。 四、小鼠肺部及血液中 Klebsiella pneumoniae 菌量測定 * 材料: 均質機、滅菌的均質棒、70 %酒精(alcohol)、裝冰塊的小燒 杯、廢液桶、15 ml 離心管、左肺、全血、無菌的一倍磷酸鹽緩 衝溶(1X PBS)、電子天平秤、酒精燈、 70 %噴霧酒精、擦手紙、 裝 70%酒精的小燒杯、微量離心管(1.5ml micro-centrifuge tube)、 微量離心管放置架(eppendorff rack)、試管震盪器(vortex mixer)、 三角玻璃棒、無菌 PBS、無菌 LB agar plate、1000p、200p、20p 吸管唧筒(pipette pump)、吸管尖(blue & yellow tip)、廢物桶、無. 46.
(47) 菌通風櫥、37℃培養箱(incubator)、小鼠左肺均質液與 0.2 ml 全 血。 * 步驟: 4-1. 小鼠左肺及血液的處理 將左肺以電子天平秤秤重,之後以均質機在 10000 rpm、10 秒、4 ℃下共均質三次,均質液置於冰上,清洗質棒,15 ml 離 心管內裝 70 %酒精與無菌 1X PBS,順序為:70 %酒精二次,等 酒精乾燥後以無菌 1X PBS 清洗三次。清洗完均製棒後才能均質 下一管左肺;0.2 ml 全血置於冰上。 4-2. 左肺及血液的細菌培養 取老鼠之肺均質液與全血 100μl 用無菌 PBS 作 1:10 的連續 稀釋,各稀釋到 10-1、10-2 倍,取均質液及全血與其 10-1、10-2 倍 稀釋液各 100μl 均勻塗抹於 LB agar 上,將塗抹好的 LB agar 靜 置於 37℃培養箱中,培養 18 小時,18 小時後計算 LB agar 上的 菌落數。 五、肺泡巨噬細胞吞噬 Klebsiella pneumoniae 菌量之計數 * 材料: 15 ml 離心管(15 ml centrifuge tube)、滅菌一倍磷酸鹽緩衝溶 液(1X PBS)、滴管(dropper)、電動吸取器(electrical pipette)、無菌. 47.
(48) 吸管(5 ml pipettes)、細胞計數盤(hemacytometer; Hausser Scientific, USA)、計數器、顯微鏡(microscopy)、離心機、小鼠肺泡灌洗液、 玻片(micro slides glass; Daco)、玻片架、滴管(dropper)、cytospin 用固定架和 cytospin 用漏斗 (Shadon)、打洞濾紙(190005 Filter Cards; Shandon)、cytospin 離心機(cytospin 3; Shadon)、Giemsa 染 液(Meditech)、ddH2O、methanol、Gram stain Kit (Meditech)、光 學顯微鏡(Olympus)。 *步驟: 5-1. 細胞計數 (cell count) 15 ml 支氣管灌流液,離心 1200 rpm、4 ℃、5 min,倒掉 1X PBS,輕輕把管子底部細胞敲散,加入 1X PBS 至 5 ml,以 5 ml 電動 pipette 小心地混合細胞液,這一步是為了清洗細胞,離心 1100 rpm、4 ℃、5 min,倒掉 PBS、輕輕敲散底部細胞,加入 1X PBS 至 5 ml,電動 pipette 小心均勻地混合後,吸 20μl 細胞液, 前 2-3 滴去掉,輕輕打入已經蓋上蓋玻片細胞計數盤的凹槽中, 看到細胞液剛好蓋滿格子即可,再顯微鏡下算取井自四周四大格 之細胞總數,再以下列公式計算細胞的總數。 【公式】 (四格計算細胞總數/4)× 3 ml ×104=支氣管灌流液中 細胞數目. 48.
(49) 5-2. 細胞型態 A.) Cytospin 固定架上架玻片,貼濾紙,洞對固定的洞,放漏斗,夾住, 滴 40μl(約有 4×103 cells)支氣管灌洗液,開離心機 power,開 蓋子,架子放對稱,蓋內蓋,壓緊,蓋外蓋,按 set speed 300 rpm., 按 set time 5 min.,按 START。 B.) Giemsa stain 取出 cytospin 好的玻片,風乾,用滴管灑 1ml 甲醇(methanol) 於玻片上,固定細胞用,風乾,約 30 min.後,玻片放入玻片架, Giemsa 使用前搖勻,取 3 ml Giemsa 染液,加 ddH2O 到 90 ml, 使 Giemsa 染液與 ddH2O 的比例為 1: 30,玻片架放入裝好染液染 瓶中,稍微上下動一下,染勻,染 30 min,片架拿出,放入水盒 中,用水沖 1 min,風乾,在光學顯微鏡 400x 下看片,觀察細胞 型態。 5-3. 胞內細菌量計數 取出 cytospin 好的玻片,風乾,以火焰將玻片背面快速烘烤 一遍,以 Gram stain Kit (Meditech)第一劑染一分鐘,之後以自來 水將玻片背沖 30 秒,再染第二劑一分鐘便以自來水將玻片背沖 30 秒,接續第三劑脫色 15 秒,便以自來水將玻片背沖 30 秒後染. 49.
(50) 第四劑一分鐘,而後玻片背沖 30 秒,風乾,以光學顯微鏡在 1000 x 油鏡下看片,計數胞內的 KP 細菌數。 六、肺泡巨噬細胞凋亡之偵測 * 材料: 小鼠肺泡灌洗液、細胞凋亡偵測試劑組(cell death detection Kit;Roche)、電動 piptte、10ml 電動 piptte 吸管、微量離心管、 1000p、200p、20p 吸管唧筒(pipette pump)、吸管尖(blue & yellow tip)、15 ml & 50 ml 離心管(15ml & 50 ml centrifuge tube)、1X PBS、5ml 試管(Falcon;BD)、試管放置盒、離心機、流氏細胞 儀(Flow cytometer , FACScan; BD)、鋁箔紙、避光用冰桶、試管 震盪器(vortex mixer)。 * 步驟: 將小鼠肺泡灌洗液在 4℃、1100 rpm、5 分鐘離心後,倒去 上清液,並加入 10 ml PBS 以相同條件離心,將細胞清洗兩次, 之後加入 3 ml PBS,並以電動 piptte 混合,每組各取 20μl 至細 胞計數器進行細胞計數,並將各組之細胞以相同條件離心後,個 別加入 100μl HEPES buffer,每組分別取 1×105 個細胞數至 5 ml 試管(Falcon;BD)中分成四管,其餘每組細胞則混合分裝至 5ml 試管共三管以調條件用,依照需要將此七管加入螢光試劑各. 50.
(51) 是 1μl 分別如下:1st 皆不加,2nd 則加入 Annexin-V 螢光試劑, 3rd 加入 PI 螢光試劑,4th、5th、6th、7th 則加入 Annexin-V 螢光試 劑及 PI 螢光試劑,靜置室溫反應 20 分鐘後,每管各加入 400μl PBS 混合之,於一小時內需以流氏細胞儀收集資料分析。 第二節 體 外 試 驗 (in vitro) 一、RAW 264.7 細胞株感染 Klebsiella pneumoniae * 材料: RAW264.7 cell line (老鼠巨噬細胞株;ATCC) 、RPMI-1640 medium (GIBCO)、fetal bovine serum (FBS; GIBCO)、L-glutamine (GIBCO) 、 peniciline-streptomycin. (GIBCO) 、 fungizone. amphotericin B (250µg/ml) 、 sodium bicabernate (sigma) 、 trypsin-EDTA (GIBCO)、1X PBS、細胞培養皿(T75)、0.2μm filter、3 ml 玻璃吸管、電動 piptte、10 ml 電動 piptte 吸管、70 % 酒精、倒立顯微鏡( Nikon Ellipae TE300) 、細胞培養箱(37℃; 5 % CO2) 、6-well 細胞培養盤、細胞計數器、37℃水浴槽( water box) 、hypo-/hyper- mucoid Klebsiella pneumoniae 已定量菌液、蓋 玻片(2×2 mm2) 、1000p、200p、20p 吸管唧筒(pipette pump)、 吸管尖(blue & yellow tip) 、酒精燈、試管震盪器(vortex mixer)。 * 步驟:. 51.
(52) 1-1. 細胞培養 將第十一代 RAW264.7 cells 自-80℃冰箱取出一管(約 1×107 cells),於 37℃水浴槽回溫,而後加入含有 10 ml 培養液中(內含 RPMI-1640 medium 、 10% FBS 、 1% L-glutamine 、 1% peniciline-streptomycin、0.2 % fungizone amphotericin B),放入細 胞培養箱中,每天觀察細胞生長情形,若細胞長滿則須分盤,即 將培養皿中的培養液以連接抽吸幫浦(suction pump)玻璃吸管將 培 養 液 吸 淨 , 並 以 1X PBS 清 洗 細 胞 二 次 後 加 入 1 ml trypsin-EDTA 靜置細胞培養箱一分鐘,之後加入 3 ml 培養液並 以電動 piptte 上下抽吸以將貼於壁上之細胞沖下,並混合均勻取 0.5 ml 細胞液加入含有 10 ml 培養液,放置於細胞培養箱中。 1-2 Klebsiella pneumoniae 之感染 準備 6-well 細胞培養皿,標記組別(blank、hypomucoid、 hypermucoid)與 MOI(multiplicity of infection;1 或 10)並在每 一 well 加入 2 ml 細胞培養液或有些則再放入蓋玻片,經細胞分 盤與計數後,每一 well 分別放入第十五代 1×106 RAW264.7 cells, 而後靜置於細胞培養箱一小時使細胞貼壁,一小時將其取出並吸 淨原先之培養液,以 1X PBS 清洗細胞二次後,每一 well 加入 2ml 純 RPMI-1640 meium,再加入先前已定量好的 KP 菌量. 52.
(53) (MOI=1 則加入 1×106 KP/1×106 RAW cells,MOI=10 則加入 1× 107 KP/1×106 RAW cells),輕輕搖勻,而後再放入細胞培養箱四 十五分鐘。 二、Klebsiella pneumoniae 所造成之細胞毒性(cytotoxicity)與所誘發之 細胞凋亡(inducing- apoptosis) * 材料: 已感染四十五分鐘之 RAW264.7 cells 、RPMI-1640 medium (GIBCO)、fetal bovine serum (FBS; GIBCO)、gentamicine(sigma) 、 3 ml 玻璃吸管、10 ml 電動 piptte 吸管、電動 piptte、70 %酒精、 倒立顯微鏡( Nikon Ellipae TE300)、細胞培養箱(37℃; 5 % CO2) 、1X PBS、蓋玻片(2×2 mm2) 、4 % paraformaldehyde、0.4 % trypan blue (sigma)、DAPI( 4.6-dianmidin-2-phenylinodle; Sigma)、 37℃水浴槽(water box)、1000p、200p、20p 吸管唧筒(pipette pump) 、吸管尖(blue & yellow tip) 、15 ml 離心管(15 ml centrifuge tube)、膠帶、螢光顯微鏡(Olympus)、酒精燈。 *步驟: 2-1. Trypan blue 細胞存活力測試 將已感染四十五分鐘之 RAW264.7 cells 其培養液吸淨,再 以 1X PBS 清洗細胞三次,而後加入含有 gentamicine (100 µg/ml). 53.
(54) 的 RPMI-1640 medium (含有 10 % FBS) 2 ml 放置一小時,隨後將 其培養液吸淨,再以 1X PBS 清洗細胞三次便加入 1 ml 0.4 % trypan blue 靜置三分鐘,之後將 trypan blue 移除,加入 1 ml 1X PBS,置於倒立顯微鏡之下評估被 trypan blue 染上的細胞百分比。. 2-2. DAPI(4.6-dianmidin-2-phenylinodle)核染色 A.) 1μg/ml DAPI 配製 將 0.5 ml ddH2O 加至含有 5 mg 粉狀 DAPI 的 1ml 避光瓶 中並搖勻,儲存於-20℃冰箱中,使用時則取 1 µl 至 10 ml PBS 中稀釋並避光混合之後即可染色。 B.) DAPI 染色 將已感染四十五分鐘之 RAW264.7 cells(其 6-well 細胞培養 皿上放置蓋玻片可使細胞貼壁)其培養液吸淨,再以 1X PBS 清 洗細胞三次,而後加入含有 gentamicine (100µg/ml) 的 RPMI-1640 medium (含有 10% FBS) 2 ml 放置十二小時,之後將將其培養液 吸 淨 , 再 以 1X PBS 清 洗 細 胞 三 次 , 便 加 入 1 ml 4% paraformaldehyde 三十分鐘以固定細胞,而後吸除之,再以 1X PBS 清洗細胞三次,於每一 well 中再加入 l ml DAPI(1μg/ml)於室 溫下反應三十分鐘,再以 1X PBS 清洗細胞三次,之後將蓋玻片. 54.
(55) 移開轉置於玻片上,以膠帶將蓋玻片四周固定,於螢光顯微鏡 400x 下觀察(340/380 excitation)。 三、Klebsiella pneumoniae 對 RAW cells 的附著(adhesion)及 KP 在 巨噬細胞內的存活量(macrophage-associated KP)評估 * 材料: 已感染四十五分鐘之 RAW264.7 cells、 RPMI-1640medium (GIBCO)、fetal bovine serum (FBS; GIBCO)、gentamicine(sigma) 、 Triton- X 100(Merck)、細胞刮杓、3ml 玻璃吸管、電動 piptte、 10 ml 電動 piptte 吸管、70%酒精、倒立顯微鏡(Nikon Ellipae TE300) 、細胞培養箱(37℃;5 %CO2) 、1X PBS、37℃水浴槽(water box) 、1000p、200p、20p 吸管唧筒(pipette pump) 、吸管尖(blue & yellow tip)、15 ml 離心管(15 ml centrifuge tube)、LB agar 三 角玻璃棒、酒精燈、LB agar、微量離心管(1.5 ml micro centrifuge tube) 、微量離心管放置架( eppendorff rack) 、試管震盪器( vortex mixer)。 * 步驟: 將已感染四十五分鐘之 RAW264.7 cells 之培養液吸淨,再以 1X PBS 清洗細胞三次,而後不加(評估 adhesion)或加入(KP 在 巨 噬 細 胞 內 的 存 活 量 評 估 )含 有 gentamicine (100 µg/ml) 的. 55.
(56) RPMI-1640 medium(含有 10 % FBS) 2 ml 放置一小時,之後將其 培養液吸淨,再以 1X PBS 清洗細胞三次,分別加入 1 ml 0.5 % Triton-X 100 靜置 10 分鐘,而後以細胞刮杓將細胞刮下,以 piptte 重覆抽吸三次混合,並取 100μl 之細胞細菌混合液至 900μl PBS 稀釋之,再取 20μl 稀釋液至 LB agar 上塗盤,將塗抹好的 LB agar 靜置於 37℃培養箱中,培養 18 小時後,計算 LB agar 上的菌落 數。 四、Mini-Tn10 transposon mutagenesis. * 材料: M9 salt (Becton Dickinson) 、 agar (Becton Dickinson) 、 Glucose、MgSO4、CaCl2、kanamycine(sigma) 、E. coli( S17-1?pir, mini-Tn10;中山醫學大學微免科賴怡琪教授提供)、細菌培養 箱、1000p、200p、20p 吸管唧筒(pipette pump)、8 爪 piptte、 吸管尖(blue & yellow tip) 、微量離心管( 1.5 ml micro centrifuge tube;eppendrof)、微量離心管放置架(eppendorff rack)、無菌 normal saline、2×2 cm2 NC membrane、酒精燈、試管震盪器(vortex mixer)、塗盤用無菌滾珠、70%酒精、鑷子、無菌牙籤、96-well plate、3M 鋁箔膠帶、剪刀、擦手紙、保鮮膜、垃圾盤、牙籤丟. 56.
(57) 棄桶。 * 步驟: 4-1. M9 medium (agar and plate) 配製 取 7.5 g agar 加入 400 ml ddH2O 混合均勻,再取 5.64 g M9 salt 加入 100 ml ddH2O ,將上二者滅菌,當其溫度於 45-50℃時 將二者加在一起,並依序加入無菌 10 ml 20% glucose、無菌 1 ml 1M MgSO4、無菌 50 ml 1M CaCl2、500 µl kanamycin (50 g/ml), 當加入 CaCl2 會有白色物質產生,可搖晃之再加下者,之後將配 製好之培養基適量倒入無菌培養皿中待其冷卻凝固;若為 broth 則無須加入 agar,以相同條件配製。 4-2. Conjugation 試驗 取 2 ml LB broth,加入 2μl kanamycin (50 g/ml)使之為 50μ g/ml 的濃度,之後挑 KP1112 與 E. coli (miniTn10;已構築含 kanamycin 抗藥基因)單一 colony 種入 broth 中,斜放在 37℃、160 rpm 細菌培養箱,培養 18 小時。隔日,將已長菌的 broth 由培養 箱取出,並將 broth 分別取 1.5 ml 至 eppendrof,KP1112 則以 13000 rpm 離心 3~5 分鐘,E. coli 則以 6000 rpm 離心 3 分鐘, 之後移除上清液並個別加入 1 ml 無菌 normal saline (contain 10 mM MgSO4) , 以 piptte 將 其 重 覆 抽 吸 以 混 合 均 勻 , 洗 去. 57.
(58) kanamycin,再進行第二次相同條件的離心,之後吸去上清液, 加入 1 ml 無菌 normal saline 以 piptte 重覆抽吸混合之,而後 KP1112 不離心,E. coli 則需以相同條件再離心第三次,之後加 入 l ml normal saline (contain 10 mM MgSO4)並將之混合均勻。 KP1112 與 E. coli各取 500μl 到 eppendrof 並混合均勻,以 13000 rpm 離心 3 分鐘,移除上清液並加入 100μl 無菌 normal saline (contain 10 mM MgSO4);另一方面則以過火滅菌過的鑷子取 2×2 cm2 NC membrane 放到 M9 plate 上,將混合均勻的 KP1112 與 E. coli 菌液取 100μl 到 NC membrane 中心點,須小心避免溢出, 之後將其放置於 37℃細菌培養箱,培養 4 小時。 4 小時後,以 過火鑷子夾 NC membrane 兩對角將其置入 2 ml 的 M9 borth (contain 50μ g/ml kanamycin)培 養 基 中 , 並且以試管震盪器 (vortex mixer)將其上的細菌震盪至培養基中(可見到培養基由 清澈略變渾濁),之後放入 37℃、160 rpm 細菌培養箱中放置一 小時,再取 200μl 至 M9 plate 中,放入滾珠將 plate 塗乾,放於 37℃、160 rpm 細菌培養箱,培養 18 小時。 4-3. 細菌挑選與保存 隔日將長出的細菌(已 mutagenesis 成功) ,依照需要(分成 hypo-/hyper-mucoid)以牙籤挑出,並放置於每一 well 已有 100. 58.
(59) μl M9 broth(contain 50μg/ml kanamycin)之 96-well plate 中, 將牙籤在 broth 輕輕攪拌,在細菌挑選完畢後,以 3M 膠帶將 96-well plate 密封,蓋上外蓋,將 96-well plate 放入放入 37℃、 120 rpm 細菌培養箱,以膠帶固定之,培養 18 小時。 18 小時後,打開 96-well plate 撕開膠帶,可見每一 well 中 有白色的細菌沉澱,於每一 well 中加入 50μl 80 % glucose,以 3M 膠帶將 96-well plate 密封,蓋上外蓋,其外並以保鮮膜包住 以防水氣,放進-80℃冰箱保存,而感興趣的細菌則取 20μl 菌液 至 2 ml LB broth (contain 50μg/ml kanamycin)增菌,進行對 RAW cells 的附著(adhesion)及 KP 在巨噬細胞內的存活量評估 之實驗。 參、統計方法 本實驗正常與糖尿病小鼠其體重、血糖值及感染 KP 後,其 cfu+1 取 log 之後的值、凋亡的百分比、吞噬細菌數之比較以成 對 t 測試 (paired-t test; SPSS 10.0) 檢定;正常與糖尿病小鼠其 不同週數的血糖值的比較,及正常或糖尿病小鼠感染不同 KP 後,其 cfu+1 取 log 後的值、凋亡的百分比之比較,以獨立 t 測 試 (independent-t test; SPSS 10.0) 檢定。另外,成對 KP 感染巨 噬細胞株之細胞存活力、凋亡之百分比及 KP 附著、KP 在巨噬. 59.
(60) 細胞內的存活量之 cfu+1 取 log 後的值,皆使用獨立 t 測試 (independent-t test; SPSS 10.0) 檢定。實驗資料的統計數據以平均 值± 標準誤差 (mean±standard error of the mean, mean ±SE) 表 示。當 P 值小於 0.05 時(*P<0.05),代表具有統計學上顯著的差 異。. 60.
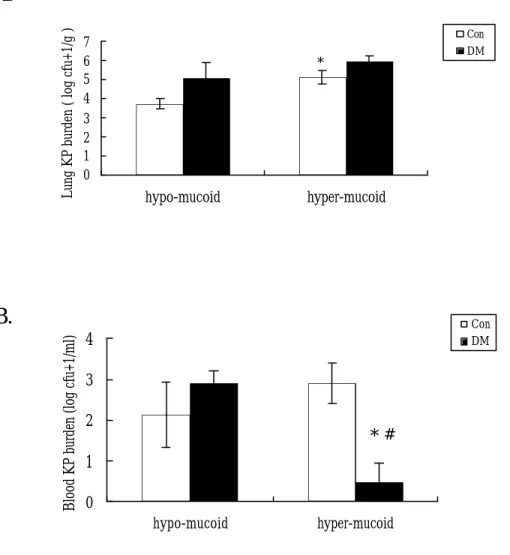
(61) 第三章 結果 第 一 節 體 內 試 驗 ( in vivo ) 1-1. DM 小鼠實驗模式之建立 為探討 KP 在肺部感染時,糖尿病(DM)是否為致病因子,故 須建立 DM 小鼠模式。將 6 週齡 C57BL/6J 公鼠分為兩組,控制 組 (Con) 則予檸檬酸鹽緩衝液注射;糖尿病組 (DM) 予連續四 天低劑量 STZ 腹腔注射,以破壞蘭氏小島之β-cell 而誘發糖尿 病,當第 8 週隨機血糖大於 300 mg/dl 時便定義其為糖尿病鼠。 1. 小鼠血糖 結果圖三顯示第八週時,控制組隨機血糖為 117.5 ± 2.1 mg/dl, DM 組則為 320.3 ± 5.8 mg/dl,符合 DM 定義且 DM 組血 糖明顯較控制組高 (P<0.05;n= 46)。為使 DM 小鼠長期處在高 血糖的生理情況下,將二組一起飼養至 30 週齡才進行實驗;於二 十八週時測其血糖,控制組隨機血糖為 118.9 ±1.9 mg/dl,DM 組 則為 411.3 ± 8.5 mg/dl,DM 組血糖明顯較控制組高並持續至 28 週(P<0.05;n= 46 )。此外,DM 小鼠其 28 週的血糖較第 8 週時 有顯著性的上升 (P<0.05)。 2. 小鼠體重 圖四結果顯示於第十週時,DM 小鼠體重為 20.2 ± 0.2 克,正. 61.
數據
Outline
相關文件
一、貴公司依據「多國多中心藥品臨床詴驗計畫 審查程序」申請之藥品臨床詴驗計畫,案內申請 人/詴驗委託者為羅氏大藥廠股份有限公司,本部 同意之計畫書版本日期為:Protocol WO30070
liraglutide 對於第二型糖尿病病患的療效與安全性之臨床 試驗。 一個 26 週並再延長 26 週、隨機分配、三個治療
評估以 S-649266 或最佳現有療法進行治療罹患抗 Carbapenem 革蘭氏陰性菌感染的患者的臨床結果 (包括詴驗用藥 S-649266
6 《中論·觀因緣品》,《佛藏要籍選刊》第 9 冊,上海古籍出版社 1994 年版,第 1
提高免疫力上有良好的功效。可治 療糖尿病血管併發症及扁平疣、傳染性疣。在動物 試驗中,利用環磷醯胺製出免
在現行的 99
比較(可與不同時期、不同藝術家,對同類型/主題創 作的處理進行比較。例:Donatello的《David》)、分
•給學生很多的機會嘗試 比較不同物件的重量,鼓 勵學生表達兩件物件相對 的重量。.
