國立交通大學
生化工程研究所
碩士論文
人類醯亞胺水解酵素之分子
選殖表現及其功能性質探討
Heterologous Expression, Purification and Functional
Characterization of Human Imidase
研究生:廖家煒
指導教授:楊裕雄 教授
人類醯亞胺水解酵素之分子選殖表現及其功能性質探討
Heterologous Expression, Purification and Functional Characterization of
Human Imidase
研 究 生:廖家煒 Student: Chia-Wei Liao
指導教授:楊裕雄 教授 Advisor: Prof. Yuh-Shyong Yang
國立交通大學
生化工程研究所
碩 士 論 文
中華民國九十七年七月
A Thesis
Submitted to Department of Biological Science and Technology
National Chiao Tung University
in Partial Fulfillment of the Requirements for the Degree of Master of
Science in Biologic Science and Technology
Hsinchu, Taiwan, Republic of China
博碩士論文授權書
本授權書所授權之論文為本人在_ 國立交通__大學(學院)_生化工程_系所 _______組_九十六 _學年度第_二_學期取得_碩_士學位之論文。 論文名稱:_ 人類醯亞胺水解酵素之分子選殖表現及其功能性質探討 指導教授:_楊裕雄 教授 _______________________ 1.□同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予行政院國家科學委員會科學技 術資料中心(或改制後之機構),得不限地域、時間與次數以微縮、光碟或數位化等各種 方式重製後散布發行或上載網路。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之一, 申請文號為:______________,註明文號者請將全文資料延後半年再公開。 2.□同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予教育部指定送繳之圖書館及國 立交通大學圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學 術研究之目的,教育部指定送繳之圖書館及國立交通大學圖書館得以紙本收錄、重製與 利用;於著作權法合理使用範圍內,不限地域與時間,讀者得進行閱覽或列印。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之一, 申請文號為:______________,註明文號者請將全文資料延後半年再公開。 3.□同意 □不同意 本人具有著作財產權之上列論文全文(含摘要),授予國立交通大學與台灣聯合大學系統 圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學術研究之目 的,國立交通大學圖書館及台灣聯合大學系統圖書館得不限地域、時間與次數,以微縮、 光碟或其他各種數位化方式將上列論文重製,並得將數位化之上列論文及論文電子檔以 上載網路方式,於著作權法合理使用範圍內,讀者得進行線上檢索、閱覽、下載或列印。 論文全文上載網路公開之範圍及時間 – 本校及台灣聯合大學系統區域網路: 年 月 日公開 校外網際網路: 年 月 日公開 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行權利。依本授 權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與不同意之欄位若未鉤選,本 人同意視同授權。 研究生簽名: 學號: (親筆正楷) (務必填寫) 日期:民國 年 月 日 1. 本授權書請以黑筆撰寫並影印裝訂於書名頁之次頁。國家圖書館博碩士論文電子檔案上網授權書
本授權書所授權之論文為本人在__國立交通_大學(學院)_生化工程_系所 _____ _組_九十六_學年度第_二_學期取得_碩_士學位之論文。 論文名稱:_人類醯亞胺水解酵素之分子選殖表現及其功能性質探討 指導教授:_ 楊裕雄 教授_____________________ □同意 □不同意 本人具有著作財產權之上列論文全文(含摘要),以非專屬、無償授權國家圖書館,不限 地域、時間與次數,以微縮、光碟或其他各種數位化方式將上列論文重製,並得將數位 化之上列論文及論文電子檔以上載網路方式,提供讀者基於個人非營利性質之線上檢 索、閱覽、下載或列印。 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行權利。 依本授權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與不同意之欄位 若未鉤選,本人同意視同授權。 研究生簽名: 學號: (親筆正楷) (務必填寫) 日期:民國 年 月 日 甲、 本授權書請以黑筆撰寫,並列印二份,其中一份影印裝訂於附錄三之 一(博碩士論文授權書)之次頁﹔另一份於辦理離校時繳交給系所助理,由圖 書館彙總寄交國家圖書館。人類醯亞胺水解酵素之分子選殖表現及其功能性質探討
學生: 廖家煒
指導教授: 楊裕雄 教授
國立交通大學生化工程研究所碩士班
摘要
醯亞胺水解酵素(DHP, dihydropyrimidinase, EC 3.5.2.2 )參與嘧啶的還原性分 解途徑。人類醯亞胺水解酵素基因座落於染色體位置8q22,含 1560 個鹼基所構 成的開放讀碼框,編碼著由519 個胺基酸所構成的多肽鏈。人類醯亞胺水解酵素
屬 於 cyclic amidohydrolase superfamily , 其 中 包 含 了 dihydropyrimidase 、
allantoinase、hydatoinase、dihydroorotase 等,皆參與嘌呤與嘧啶環的代謝。分析 人類醯亞胺水解酵素的胺基酸序列,其與大鼠及小鼠分別有 90%及 88%的相似 度。人類醯亞胺水解酵素缺乏症是一種體染色體隱性遺傳的疾病,不但可藉由檢 測病患尿液中dihydropyrimidine 含量來判斷是否患有醯亞胺水解酵素缺乏症,而 且與許多臨床病徵如癲癇、畸形、心理及生理發育不完全有關。然而截至目前為 止,在蛋白質層面,人類醯亞胺水解酵素仍有尚未被了解的生化特性及功能。為 了分析人類醯亞胺水解酵素的特性,我們解決在大腸桿菌內表現人類醯亞胺水解 酵素所遇到的蛋白質包涵體(Protein inclusion body)問題,在大腸桿菌內大量表現 可溶並具活性的人類醯亞胺水解酵素,並進一步比較不同物種來源,在酵素動力
學、pH 值及溫度對活性的影響、耐熱度、耐鹽度上的差異。除此之外我們還積
極地建立一個能穩定生產人類醯亞胺水解酵素的平台,以便後續能夠從蛋白質層 面來探討,在病理上如何因為單一個氨基酸突變而造成人類醯亞胺水解酵素失去 活性,導致人類醯亞胺水解酵素缺乏症的發生。
Heterologous Expression, Purification and Functional Characterization of
Human Imidase
Student: Chia-Wei Liao Advisor: Prof. Yuh-Shyong Yang Department of Biological Science and Technology and Institute of Biochemical Engineering,
National Chiao Tung University, Hsinchu, Taiwan, ROC
ABSTRACT
Imidase participates in the reductive degradation pathway of pyrimidine. The location of
Homo sapiens dihydropyrimidinase (hDHP, DPYS, NM_001385) gene is on the chromosome 8q22 and its cDNA clone contains 1560-bp open reading frames encoding polypeptides of 519 residues. hDHP belongs to the superfamily of cyclic amidohydrolase, including dihydropyrimidase, allantoinase, hydatoinase, dihydroorotase, all of which are involved in the metabolism of purine and pyrimidine rings. The deduced amino acid sequence of hDHP shows 90% and 88% identity with that of rat and mouse DHP, respectively. hDHP deficiency
is an automosomal recessive disorder characterized by dihydropyrimidinuria and associated with variable clinical phenotypes, such as seizures, mental retardation, growth retardation and dysmorphic features. However, little is known about the properties and biological functions of human imidase at protein level. In order to characterize the biochemical properties of human imidase, we expressed human imidase gene in E. coli and rendered the purified protein in soluble and active form. The characterizations in enzyme kinetics, the effect of pH and temperature to enzyme activity, thermostability and salt tolerance of human imidase and those from other species were compared. This provided a basis for the future preparation of imidase mutants to study the effect of single amino acid substitution on imidase that causes hDHP deficiency
Contents Page
Abstract (Chinese)
... i
Abstract (English)
...ii
Contents
...iiii
Content of table and figure
... iv
Abbreviation and Symbol
... vi
Introduction
... 1
Experimental Procedures... 6
Results... 11
Discussion... 15
Reference... 19
Appendixe... 40
Content of table and figure
Page
Table I Summary of purification of Nus-His-taged recombinant hDHP from E.
coli ………. 23
Table II Specific Activity of hDHP and a.r.HYD ………... 24
Table III Kinetcs constants of imidase from different sources with phathlimide … 25 Table IV Calibration curve determination of molecular weight on Sephacryl S-300 HR ……….. 26
Table V Physical and chemical parameter of hDHP ……….. 27
Figure 1 SDS-PAGE (10%) of purified recombinant hDHP ……….. 28
Figure 2 Purified recombinant hDHP analyzed by MADI-TOF after trypsin-in-gel digestion ………. 29
Figure 3 Temperature profile of hDHP ……… 30
Figure 4 pH profile of hDHP ………... 31
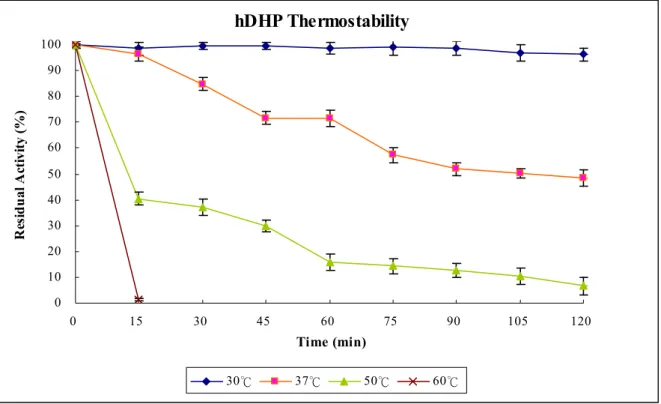
Figure 5 Thermostability of hDHP ……….. 32
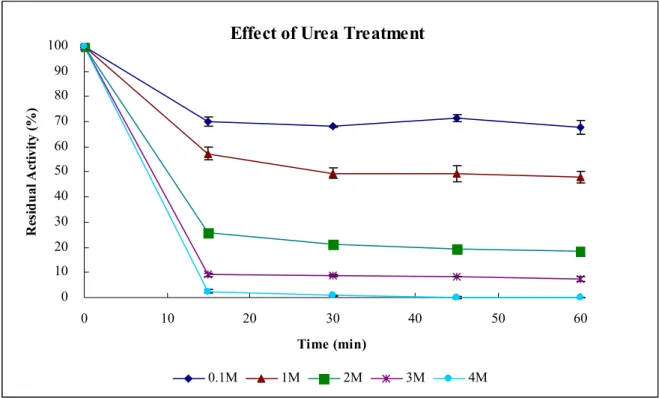
Figure 6 Effect of urea treatment of hDHP ……….. 33
Figure 7 Effect of urea concentration of hDHP ………... 34
Figure 9 hdhp-pET-43.1a (+) construct DNA sequencing ……….. 36
Abbreviation and Symbol
Abbreviation and Symbol Full name
ε Absorption (extinction) coefficient
A280 Absorption at 280 nm
A298 Absorption at 298 nm
Bis-Tris propane 1,3-bis[ tris( hydrohxymethyl) methylamino] propane
Tris-HCl tris( hdroxymethyl) aminomethane hydrochloride
SDS dodium dodecly sulfate
PAGE polyacrylamide gel electrophoresis
hdhp dihydropyrimidinase gene of Homo sapien hDHP dihydropyrimidinase of Homo sapien
Introduction
Imidase catalyzes the hydrolytic cleavage of imide bond. Imidase is also known as
dihydropyrimidinase (EC 3.5.2.2), hydantoinase, dihiydropyrimidine hydrase, and
dihydropyrimidine amidohydrolase due to its broad substrate specificity.
Figure.1 The typical reaction catalyzed by imidase
The first research of imidase activity in living organisms was found as far back as 1926.
Gaebler and Keltch investigated the metabolism of hydantoin derivatives by detecting that the
hydantoic acid was excreted after injection of hydantoin into dogs and called this kind of
imidase as hydantoinase [1]. Afterward, many studies show that imidase activity could be
found in some tissues of different plants and animals. Up to 1957, Wallach and Grisolia isolated
imidase from calf liver and used dihydropyrimidine as nature substrate, so they called the
hydantoinase found by Gaebler and Keltch, and therefore imidase participates in the pyrimidine
metabolism [2].
The cDNA clone enconding dihydropyrimidinase from human liver had been firstly
isolated in 1996 [3]. The deduced amino acid sequence of human DHP show 90% and 88%
identity with that of rat and mouse DHP, respectively (Appendix 3). The gene of Homo sapiens
dihydropyrimidinase (hDHP, DPYS, NM_001385) locates on the chromosome 8q22
(Appendix 1) and its cDNA clone contains 1560-bp open reading frames encoding
polypeptides of 519 residues (Appendix 2) [3]. Whilecloning the hDHP cDNA, there are three DHP related protein (DRP)-1, DRP-2 and DRP-3 have been found. All of them show high identity of amino acid sequence with hDHP (Appendix 3), but without imidase activity. DHP and DRPs display differential tissue distribution, i.e. human DHP in liver and kidney; human DRP-1 in brain; human DRP-2 ubiquitously expressed except for liver; human DRP-3 mainly
in heart and skeletal muscle. The function of DRPs and their relationships with DHP in
evolution are still not clear [3].
hDHP belongs to the superfamily of cyclic amidohydrolase, including
dihydropyrimidase, allantoinase, hydatoinase, dihydroorotase, all of which are involved in
metabolism of purine and pyrimidine rings. In the biological system, pyrimidines are
metabolized by either an oxidative or a reductive pathway. It is well-known that mammals,
Dihydropyrimidinase performs reversible hydrolytic ring-opening of dihydrouracil and
dihydrothymine to N-carbamoyl-β-alanine and N-carbamoyl-β-aminoisobutyric acid,
respectively. 3-ureidopropionase catalyzes the irreversible hydrolysis of
N-carbamoyl-β-alanine and N-carbamoyl-β-aminoiosbutyric acid to β-alanine,
β-aminoisogutyric acid, ammonium ions, and carbon dioxide [4].
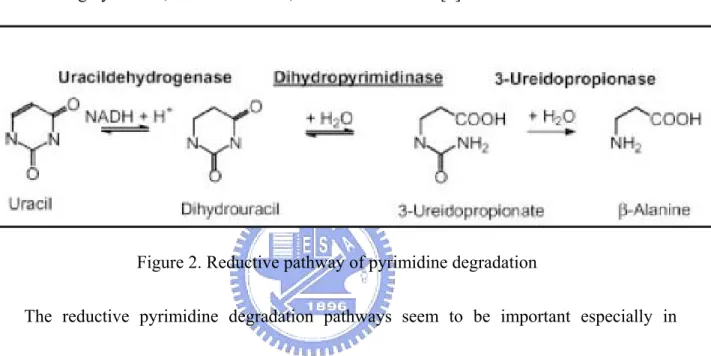
Figure 2. Reductive pathway of pyrimidine degradation
The reductive pyrimidine degradation pathways seem to be important especially in
mammalian cell. In mammalian tissue, it is now established that the degradation of uracil via
dihydrouracil and N-carbamoyl-β-alanine plays an important role in the synthesis of β-alanine
and β-aminoisobutyric acid. β-alanine is a structural analogue of γ-aminobutyric acid and
glycine, the important neurontransmitter in nerve system. Furthermore, β-aminoisobutyric acid
has been shown to be a potential agonist of the glycine receptor [5]. Pathologically, the defect
of dihydropyrimidine may cause the altered homeostasis of β-alanine in patients then accounted
DHP deficiency (MIM 222748) is an automosomal recessive disorder that is characterized
by dihydropyrimidinuria. To date, only nine cases of complete DHP deficiency have been
reported. These patients reveal variable clinical phenotypes, such as seizures, mental
retardation, growth retardation and dysmorphic features [6-15]. It is not a direct evidence to
infer the relationship between the homeostasis of β-alanine and the apparent clinical
abnormalities. DHP deficiency, therefore, is not only probably a necessary, but sufficient for
the onset of a clinical phenotype.
Analysis of the genotype of some patients with DHP deficiency reveals the missense at the
nucleotide location of open reading frame 1078, mutated from T to C (W360R) in exon 6 and a
novel missense mutation 1235G to T (R412M) in exon 7 [15]. Analysis of the structural effect on the DHP mutations, W360 and R412, by structure modeling indicates that W360 and R412 are located on the periphery of hDHP rather distant from the catalytic centre. This excludes a
direct effect of the amino acid exchanges on active site architecture and catalysis. Instead, the
lack of residual activity for the point mutants is likely based on global effects of the exchanges
on the protein structure [15]. However, all of these data are only based on bioinformatical
prediction without the evidence at protein level.
According to the analysis of imidase structures, all imidases are metalloproteins with
TIM-barrel architecture. All known mammalian imidases purified from the livers of bovine,
atoms, and one Zn atom per subunit. The variations are found, however, in structure, metal
content, substrate specificity and other requirement of similar enzyme from different organism.
Since then, little is known about the properties and biological functions of human imidase
at protein level. In our experience, the bottleneck is the problem of protein inclusion bodies
during expressing the hdhp gene in E. coli.In order to characterize the biochemical properties
of human imidase, we will express human imidase gene in E. coli in soluble and active form.
The differences in enzyme kinetics, the effect of pH and temperature to enzyme activity,
thermostability and salt tolerance of human imidase and those from other species will be
compared. We will also prepare imidase mutants, W360 and R412, to study the effect of single
Experimental Procedures
Materials― cDNA of hdhp gene on pCMV-sports6 vector, obtained from human gene
cDNA library in College of Biological Science and Technology, NCTU, pGEM-T
Vector(Promega), modified pET-43.1a(+) vector(Novagena) modified by Dr. LE, BL21
(DE3)pLyS competent cell(genotype: E. coli B F– dcm ompT hsdS(rB– mB–) gal λ(DE3)[pLysS
Camr](Novagen), Sephacryl S-300 HR column(amersham pharmacia biotech), High Molecular
Weight standard (code: 17-0041-01) Gel Filtration Calibration Kits(amersham pharmacia
biotech).
Molecular cloning hdhp gene into the pET-43.1a(+) vector― The PCR products of
cDNA of hdhp gene of Homo sapien on pCMV-sports6 vector was cloned into pGEM-T
Vector (Promega), and the termini of amplified DNA was added the restriction sites of BamH I
and Xho I with the pairing primers hdhp-F (the forward primer: 5'-CGGATCCATGGCGGC
GCCCTCGCG-3’) and hdhp-R (the reverse primer: 5’-CCTCGAGGGGGTG
GGCCTGTTTCCTGG-3’) by PCR. The DNA fragment containing the hdhp gene on pGEM-T
vector was subcloned into the modified pET-43.1a(+) vector by restriction enzyme, BamH I &
Xho I, and T4 DNA Ligase.
Bacterial expression, purification of hDHP― The flask culture (500 ml of LB medium
containing 50 μg/ml ampicillin) of the BL21 (DE3)pLyS cell containing the construct of hdhp
hours at 37℃ in a shaking incubator until cells reach mid-log growth (A600 of 0.8-1.0). The
culture was induced by adding 0.4mM IPTG and 1mM CoCl2, then incubated at 20℃ for
16 hours. The cells were harvested by centrifugation at 15000g for 30 min at 4℃. The cell
pellet was resuspended in 20 ml of Histrap Column Buffer A (pH 8.0) , consist of 50mM Tris,
500mM NaCl, 5mM Imidazole10% Glycerol, and the cell was lysed by sonication with 550
Sonic Dismembrantor (Fisher Scientific). The insoluble debris was removed by centrifugation
at 30000g for 30 minutes at 4℃, if necessary, repeat this step again to ensure removing the
insoluble debris completely. The sample should be filtered through a 0.45 mm filter before it
is applied to the Histrap ion exchange column(Pharmacia), equilibrated by Histrap Column
Buffer A. The column was washed with >1000 ml of Histrap Column Buffer B (pH 8.0) ,
consist of 50mM Tris, 500mM NaCl, 50mM Imidazole, 10% Glycerol, until no material
appears in the effluent. After changing the buffer in the column with Histrap Column Buffer A,
applied 40 unit of bovine thrombin to column and incubate it for >16 hours at 4 ℃ to cleave
the Nus-tag from the N-terminal of hDHP. The column was washed with 200 ml of Histrap
Column Buffer B, then The bounded protein was eluted with 50 ml of Histrap Column Buffer
C(pH 8.0), consist 50mM Tris, 500mM NaCl, 500mM Imidazole, 10% Glycerol.The product
was concentrated with Amicon Ultra-15 50K NMWL device (MILLIPORE) by centrifugation
In-gel-digestion and idetificaiton using MALDI-TOF― The spots of interest were excised
and digested in gel with trpsin according to Shevchenko’s method (Shevchenko et al., 1996).
The digested sample was taken up and analyzed using MADI-TOF. The results were
correlated with the sequence database using the NCBI and SwissProt database and analyzed
by Mascot software.
Enzyme assays― A rapid Spectrophotometric assay was used as the standard assay. the
decrease in absorbency at 298 nm was measured upon hydrolysis of phthalimide as the
substrate at 25 ℃. To start the reaction, the enzyme solution was added into 1 ml solution,
containing 1 mM phthalimide and 100 mM Bis-Tri propane at pH 7.0. Under these conditions,
a change in A298 of 2.26 represents the hydrolysis of 1 μmol of the substrate. The hydrolysis of
phthalimide was monitored with a UV/VIS spectrophotometer (Hitachi U3300).
Protein concentration determination― The protein concentration of enzyme solution
was determined by A280 or BCA protein assay (Bio-rad) using bovine serum albumin as a
standard. For the homogeneous imidase, 1 unit of A280 equals to 0.907 mg/ml imidase based
on the BCA protein assay.
Kinetic constants determination― Measurement of kinetic constant of each substrate was
performed by varying the concentration of one substrate, while keeping the other substrate at a
fixed and near saturating concentration. The apparent Km and Vmax were determined using
Temperature profile analysis― The specific activity of hDHP with phathalimide at given
temperature was measured with a UV/VIS spectrophotometer. To start the reaction, the enzyme
solution was added into 1 ml of solution containing 1 mM phthalimide and 100 mM Bis-Tri
propane to process enzyme assay at pH 7.0 and given temperaturature.
pH profile analysis― The specific activity of hDHP with phathalimide at given pH was
measured with a UV/VIS spectrophotometer. To start the reaction, the enzyme solution was
added into 1 ml of solution containing 1 mM phthalimide and 100 mM Bis-Tri propane to
process enzyme assay at given pH and 25℃
Thermostability analysis― The residual activity of hDHP with phathalimide was measured
with a UV/VIS spectrophotometer. Prior to start the reaction, the enzyme solution was
incubated in 1 ml of 100 mM Bis-Tri propane(pH 7.0) for time course at given temperature,
then added 1mM phtalimide to process enzyme assay.
Salt tolerance analysis―The residual activity of hDHP with phathalimide was measured
with a UV/VIS spectrophotometer. Prior to start the reaction, the enzyme solution was
incubated in 1 ml of the given concentration urea and 100 mM Bis-Tri propane(pH 7) for time
course at 25℃, then added 1mM phtalimide to process enzyme assay.
Size-exlusion chromatography and calibration curve preparation― Aliquots of 1 ml of
various samples were applied on a Sephacryl S-300 HR column that was equilibrated with 20
flow rate of 1.0 ml/min. Molecular weights were estimated against Gel Filtration Calibration
Kits(amersham pharmacia biotech) consisted of aldolase(178 KDa), catalase(228 KDa), ferritin
(437 KDa), thyroglobulin(692 KDa), Blue dextran(2000 KDa). Molecular weight of hDHP was
Result
Molecular Cloning of hdhp gene into the pET-43.1a(+) vector― we constructed hdhp gene on different vectors to screen an appropriated vector (Fig. 9, only show the hdhp-pET-43.1a (+) construct) to express soluble and active hDHP. We found pET-43.1a (+)
vector can overexpress the soluble and active hDHP.
Bacterial Expression and Purification of hDHP―The pET-43.1a(+) vector provides the hDHP for fusion to Nus-tag and His-tag on N-terminus and C-terminus, respectively. The
Nus-His-taged fusion protein was expressed in the BL21 (DE3)pLyS cell and purified by
Histrap column as describe in “Experimental Procedure”. The enzyme was finally purified
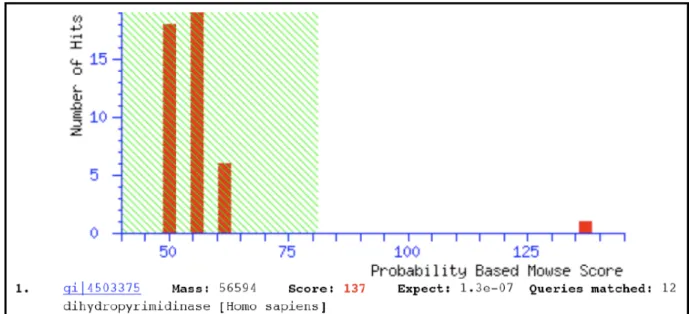
about 343.7-fold with 49% yield (Table I) and in a homogenous form according to the
identifications of SDS-PAGE (Fig. 1) and MADI-TOF data (Fig. 2). These data display that we
are the first one to not only purify hDHP in a homogenous and active form but also express the
mammalian imidase in heterolous system. We analyzed the biochemical characterization to
deduce the difference in enzyme kinetics, the effect of pH and temperature to enzyme activity,
thermostability and salt tolerance of human imidase between from human and other species.
Substrate specificity and kinetic constants of the cobalt-amended hDHP with different substrates― A number of xenobiotic and physiological substrates were tested as substrates for hDHP (Table II). The specific activities of the different substrates of hDHP were determined
concentration of substrates are 1 mM. The specific activities of hDHP with dihydrouracil,
hydantoin, phthalimide, and maleimide are 0.37, 3.48, 24.49, and 29.72 μmol/min/mg,
respectively. hDHP has higher specific activities with phthalimide, and maleimide in the
condition. The kinetic constants of hDHP with phathalimide were determined experimentally
by direct measurement with a spectrophotometer as indicated under “Experimental
Procedures”. Km, kcat, and kcat/Km of of hDHP with phathalimide are 0.65, 2615, and 4021
μmol/min/mg, respectively (Fig. 10).
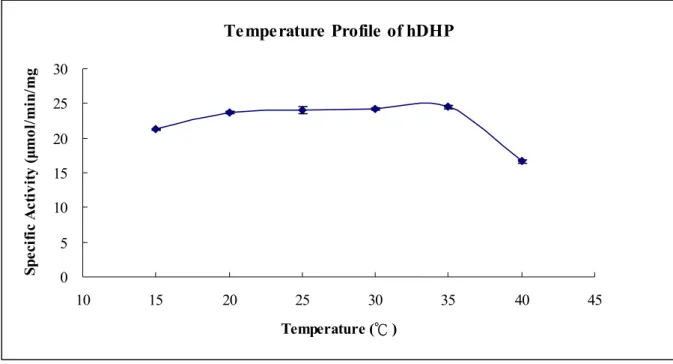
The temperature and pH profile of hDHP― The temperature profile of hDHP is displayed
by the specific activity of hDHP with phathalimide at given temperature measured with a
UV/VIS spectrophotometer as indicated under “Experimental Procedure” (Fig. 3). The specific
activities of hDHP with phathalimide from 20 ℃ to 35 ℃ are all about 24.0 μmol/min/mg
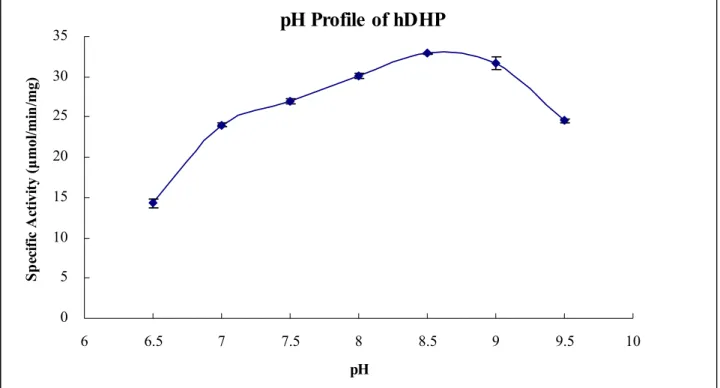
without significant different. The pH profile of hDHP is displayed by the specific activity of
hDHP with phathalimide at given pH measured with a UV/VIS spectrophotometer as indicated
under “Experimental Procedure” (Fig. 4). The specific activities of hDHP with phathalimide at
pH6.5, pH7.0~7.5, 8.0~9.0, and 9.5 are about 14.2, 25.4, 31.2, and 24.5 μmol/min/mg,
respectively. hDHP has higher specific activity at pH 8.0~9.0.
The thermostatbility of hDHP― The thermal stability of hDHP is displayed by the
residual activity of hDHP with phathalimide measured with a UV/VIS spectrophotometer as
(Fig. 5). The residual activies of hDHP with phathalimide after treating at 30 ℃ for 60, 120,
and 150 minutes are 98.53, 96.19, and 95.42 %, respectively. The residual activies of hDHP
with phathalimide after treating at 37 ℃ for 60, 120, and 150 minutes are 71.54, 48.53, and
45.67 %, respectively. The residual activies of hDHP with phathalimide after treating at 50 ℃
for 60, 120, and 150 minutes are 15.66, 6.57, and 2.19 %, respectively. The residual activity
of hDHP with phathalimide after treating at 60 ℃ for 15 minutes is 1.30 %. hDHP is much
unstable above 37℃.
The effect of urea treatment of hDHP― The effect of urea treatment of hDHP is displayed
by the residual activity of hDHP with phathalimide and measured with a UV/VIS
spectrophotometer as indicated under “Experimental Procedure” after treating in given
concentration of urea for time course (Fig. 6). The residual activity of hDHP with 0.1M, 1M,
2M, 3M, and 4M urea after treating for 15minutes are 70.01, 57.33, 25.87, 9.01, and 2.39%,
respectively. The residual activity of hDHP with 0.1M, 1M, 2M, 3M, and 4M urea after
treating for 30 minutes are 67.97, 49.34, 21.18, 8.76, and 0.71, respectively. After treating in
different concentration of urea for 30 minutes, the residual activity of hDHP performed stable
and didn’t change.
Analysis of the quaternary structure of hDHP by gel filtration― The subunit size and native
size of hDHP was estimated by its deduced amino acid sequence (A. 6) and gel filtration(Table
retention volume of the blue dextran 2000 was 37.28 ml and the Kav could be estimated by the
equation: Kav= (Ve-Vo)/(Vt-Vo). The calibration curve was plotted as the independent variable
was log(M.W.) versus the dependent variable Kav as show in Fig. 8. The retention volume of
hDHP and a.r.HYD were 54.31 ml and 60.40 ml, respectively (Table V). The corresponding
molecular weight according to the retention time was shown in Fig. 8 and the calculated values
were approximately equal to the theoretical molecular weight of hDHP and a.r.HYD (Table V).
Discussion
Since then, little is known about the properties and biological functions of human imidase
at protein level, in our experience, the bottleneck is the problem of protein inclusion bodies
during expressing the hdhp gene in E. coli. Here, the protein inclusion bodies mean the
cytoplasmic aggregates of misfolded protein. This phenomenon was often founded during
expressing the eukaryotic gene in heterologous system, especially in E. coli, and it was due to
the internal microenvironment of E. coli may differ from that original source of the gene. A
variety of methods have been published describing solving the problem of protein inclusion
bodies. The choice of vector and expression host can significantly increase the activity and
amount of target protein present in the soluble fraction. An appropriate vector can enhance
solubility and/or folding by providing for fusion to a polypeptide that itself is highly soluble
(e.g.,NusA , GST, Trx), or providing for fusion to an enzyme that catalyzes disulfide bond
formation (e.g., thioredoxin, DsbA, DsbC), or provide a signal sequence for translocation into
the periplasmic space (pelB, DsbA, DsbC). In this study, we expressed the hdhp gene on the
pET-43.1a(+) vector which provided for fusion to NusA (A. 4,5). Nus A, N utilization
substance A, is the transcription factor of E. coli naturally. Nus A not only is high soluble but
also can increase the solubility of the full-length fusion protein to solve the problem of protein
inclusion bodies during expressing the hdhp gene in E. coli.
Agrobacterium [22], Blastobacter [23], Arthrobacter [24], bovine liver[16], calf liver[17], pig
liver[18,25], and rat liver[11,19], and most purifications of these were conducted by three or
four chromatography steps. We solved the problem of protein inclusion bodies during
expressing the hdhp gene in E. coli and developed a time-and-effort-saving, inexpensive, high
quality and quantity platform to express and purify homogenous hDHP with only one
chromatography step for further biochemical characterization to deduce the difference of
specific activity and structure of imidase between human and other species.
A number of xenobiotic and physiological compounds were tested as substrates for
enzymes, and the specific activities of hDHP were compared with a.r.HYD (hydantoinase from
Agrobacterium radiobacter NRRL B1) referred to the thesis of Yi-Rong Chen (Table II)[25].
The catalytic efficiencies of hDHP with the substrates are higher than that of a.r.HYD except
dihydrouracil, the six-member ring substrate. The kinetic constants, Km, Kcat, and Kcat/Km,
of hDHP with phathalimide as substrate are 0.65mM, 2615min-1, and 4021mM-1min-1,
respectively. Comparing with imidases from fish liver, pig liver, and a.r.HYD, hDHP performs
better substrate affinity and catalytic efficiency (Table III).
The temperature profile of hDHP is displayed by the specific activity of hDHP with
phathalimide at given temperature (Fig. 3). hDHP performs stable catalytic efficiency from 20
℃ to 35 ℃. It is to deserve to be mentioned the limit of enzyme assay that the substrate is
we can not measure the initial rate of hDHP activity with phathalimide at the high temperature
and the specific activity at 40 ℃ is measured much lower than which at 35 ℃. The pH profile
of hDHP is displayed by the specific activity of hDHP with phathalimide at given pH
measured (Fig. 4). hDHP performs stable catalytic efficiency from pH 8 to pH 9.
The thermal stability of hDHP is displayed by the residual activity of hDHP with
phathalimide after treating at given temperature for time course (Fig. 5). hDHP performs stable
catalytic efficiency, the residual activity is 95.4% after treating at 30 ℃ for 150 minutes. The
residual activities of hDHP are 45.8% and 2.2% after treating at 37 ℃ and 50 ℃ for 150
minutes, respectively. After treating at 60 ℃ for 15 minutes, the residual activity of hDHP is
only 1.3%. Under the same treatment, 50 ℃ for 1 hour, the residual activities of hDHP,
a.r.HYD, and fish imidase are about 13.6%, 93%, and 90%, respectively. So, hDHP is more
thermolabile than a.r.HYD and fish imidase.
The effect of urea treatment of hDHP is displayed by the residual activity of hDHP with
phathalimide after treating in given concentration of urea for time course (Fig. 6). After treating
in different concentration of urea for 30 minutes, the residual activity of hDHP performed
stable and didn’t change, so we can simply determine the stability of urea treatment of hDHP by
measuring [Urea]50%, the concentration of urea at 50% residual activity of hDHP treated for 30
R412, with wild hDHP to study the effect of single amino acid substitution on hDHP that cause
human DHP deficiency. The [Urea]50% of wild hDHP is about 1 M.
The subunit size and native size of hDHP was estimated by its deduced amino acid
sequence (A. 6) and gel filtration (Table V). This data indicated that the native form of hDHP is
homopentameric different from other mamammalian imidases, such as calf [17], pig (5) rat [19],
yeast and slime mold DHPs [25] are homotetrameric.
In this study, it is the first one solved the problem of protein inclusion bodies during
expressing the hdhp gene in E. coli, then developed a time-and-effort-saving, inexpensive, high
quality and quantity platform to express and purify homogenous soluble and active hDHP with
only one chromatography step. We have analyzed the complete biochemical characterization of
hDHP. These results indicate that hDHP performs very unique biochemical characterization on
specific activity, thermal stability, and the quaternary structure compared with the imidases
from other species. It is worthy to know which reason caused it, maybe the sequence or
structure, and its significance on evolution. We will resolve and clarify that in the future. On the
other hand, we will prepare imidase mutants, W360 and R412, then analyzed the difference of
the structure stability and other biochemical characterization of them to study the effect of
References
[1]Gaebler OH, and Keltch AK (1926) On the metabolism of hydanoins and hydantoic acid. J
Bio, Chem 70:763-777.
[2]D.P. Wallach and S. Grisolia (1957) The purification and properties of dihydropyrimidine
hydrase. J Biol Chem. 226:277-288.
[3]Hamajima N, Matsuda K, Sakata S, Tamaki N, Sasaki M, and Nonaka M (1996) A novel
gene family defined by human dihydropyrimidinase and three related proteins with
differential tissue distribution. Gene 180:157–163.
[4]C. Wasternack (1980) Degradation of pyrimidines and pyrimidine and primidine
analogs—path ways and mutual influences. Pharmaco Ther. 8:629-651.
[5]A.B.P. van Kuilenburg, A.E.M. Stroomer, H. van Lenthe, N.G.G.M. Abeling, and A.H. van
Gennip (2004) New insights in dihydropyrimidinedehydrogenase deficiency: a pivotal role
for betaaminoisobutyric acid? Biochem. J. 379:119–124.
[6]S. Sumi, M. Imaeda, K. Kidouchi, S. Ohba, N. Hamajima, K. Kodama, H. Togari, and Y.
Wada (1998) Population and family studies of dihydropyrimidinuria: prevalence,
inheritance mode, and risk of fluorouracil toxicity, Am J Med Genet. 78:336–340.
[7]M. Duran, P. Rovers, P.K. de Bree, C.H. Schreuder, H. Beukenhorst, L. Dorland, and R.
Berger (1990) Dihydropyrimidinuria. Lancet 336:817–818.
Inherit Metab Dis. 14:367–370.
[9]M.J. Henderson, K. Ward, H.A. Simmonds, J.A. Duley, and P.M. Davies (1993)
Dihydropyrimidinase deficiency presenting in infancy with severe developmental delay. J
Inherit Metab Dis. 16:574–576.
[10]C.W. Putman, J.J. Rotteveel, R.A. Wevers, A.H. van Gennip, J.A. Bakkeren, and R.A. De
Abreu (1997) Dihydropyrimidinase deficiency, a progressive neurological disorder?
Neuropediatrics 28:106–110.
[11]Yang, Y.S., Ramaswamy, S. and Jakoby, W.B. (1993) Rat liver imidase. J Biol Chem.
268:10870-10875.
[12]S. Sumi, K. Kidouchi, K. Hayashi, S. Ohba, and Y. Wada (1996) Dihydropyrimidinuria
without clinical symptoms. J Inherit Metab Dis. 19:701–702.
[13]S. Ohba, K. Kidouchi, S. Sumi, M. Imaeda, N. Takeda, H. Yoshizumi, A. Tatematsu, K.
Kodama, K. Yamanaka, M. Kobayashi, and Y. Wada (1994) Dihydropyrimidinuria: the
first case in Japan. Adv Exp Med Biol. 370:383–386.
[14]N. Hamajima, M. Kouwaki, P. Vreken, K. Matsuda, S. Sumi, M. Imaeda, S. Ohba, K.
Kidouchi, M. Nonaka, M. Sasaki, N. Tamaki, Y. Endo, R.A. De Abreu, J. Rotteveel,
A.B.P. van Kuilenburg, A.H. van Gennip, H. Togari, and Y. Wada (1998)
Dihydropyrimidinase deficiency: structural organization, chromosomal localization, and
717–726.
[15]A B.P. van Kuilenburg J. Meijer , D. Dobritzsch, R. Meinsma, M. Duran, B. Lohkamp, L.
Zoetekouw, N.G.G.M. Abeling, H.L.G. van Tinteren, and A.M. Bosch (2007) Clinical,
biochemical and genetic findings in two siblings with a dihydropyrimidinase deficiency.
Mol Genetics and Metabolism 91:157-164.
[16]K. P. Brooks, E.A. Jones, B.D. Kim, and E.B. Sander (1983) Bovine liver
dihydropyrimdine amidohydrolase purification, properties and characterization as a zinc
metalloenzyme. Arch Biochem Biophys. 226:469-483.
[17]Kautz, J. and Schnackerz, K.D. (1989) Purification and properties of
5,6-dihydropyrimidine amidihydrolase from calf liver. Eur J Biochem.181: 431-435.
[18]T. M. Su and Y.S. Yang (2000) Identification, purification, and characterization of a
thermophilic imidase from pig liver. Protein Expr Purif. 19:289-297.
[19]Kikugawa, M., Kaneko, M., Fujimoto-Sakata, S., Maeda, M., Kawasaki, K., Takagi, T.
and Tamaki, N. (1994) Purification, characterization and inhibition of
dihydropyrimidinase from rat liver. Eur J Biochem. 219:393-399.
[20]J. Ogawa. J.M. Kim, W. Nirdonoy, Y. Amano, H. Yamada, and S. Shimizu (1995)
Purification and characterization of an ATP-dependent amidohydrolase,
N-methylhydantoin amidohydrolase from Pseudomonas putida 77. Eur J Biochem. 229:
[21]S.G. Lee, D.C. Lee and M.H. Sung (1994) Isolation of thermostable
D-hydantoinase-producing thermophilic Bacillus sp.SD-1. Biotech Lett. 16:461-466.
[22]S.M. Runser and P.C. Meyer (1993) Purification and biochemical characterization of the
hydantoin hydrolyzing enzyme from Agrobacterium species -A hydantoinase with no 5,6-
dihydropyrimidine amidohydrolase activity. Eur J Biochem. 213:1315-1324.
[23]C.L. Soong, J. Ogawa, M. Honda, S. and Shimizu (2000) Cyclic-imide- hydrolyzing
activit of D-hydantoinase from Blastobacter sp. strain A17p-4. Appl Environ Microbiol.
65:1459-1462.
[24]O. May, M Siemann, M. Pietzsch, M. Kiess, R. Mattes, and C. Syldatk (1998)
Substrated-dependent enantioselectivity of a novel hydantoinase from Arthrobacter
aurescens DSM 3745: purification and characterization as new member of cyclic
amidases. J Biotechnol. 26:1-13.
[25]C.Y. Huang, Y.P. Chao and Y.S. Yang (2003) Purification of industrial hydantoinase in one
step chromatography step without affinity tag. Protein Expr. Purif. 30: 134-139
[26]Gojkovic, Z., Rislund, L., Andersen, B., Sandrini, M. P. B., Cook, P. F., Schnackerz, K. D.,
and Piškur, J. (2003) Dihydropyrimidine amidohydrolases and dihydroorotases share the
Table I
Summary of purification of Nus-His-taged recombinant hDHP from E. coli.
a.“Histrap Elute After Thombin Digest” represented the step of purification of hDHP with Histrap ion exchange column as described in
Table II
Specific Activity of hDHP and a.r.HYD
29.72±0.54 4.7±0.1 Maleimide 24.49±0.50 0.76±0.00 Phthalimide 3.48±0.03 0.18±0.01 Hydantoin 0.37±0.00 5.08±0.06 Dihydrouracil hDHP a.r.HYDa Specific Activity(μmol/min/mg) b Substrate 29.72±0.54 4.7±0.1 Maleimide 24.49±0.50 0.76±0.00 Phthalimide 3.48±0.03 0.18±0.01 Hydantoin 0.37±0.00 5.08±0.06 Dihydrouracil hDHP a.r.HYDa Specific Activity(μmol/min/mg) b Substrate
a. a.r.HYD represented Agrobacterium radiobacter hydantoinase. The specific activities of the
different substrates of a.r.HYD were referred to the thesis of Yi-Rong Chen.
b. The specific activities of the different substrates of hDHP were determined with UV/VIS
spectrophotometer as indicated under “Experimental Procedure”, and all the concentration.of
Table III
Kinetc constants of imidase from different sources with phathlimide
41 33 ± 1 0.8 ± 0.04 Pig liver 790 1270 ± 80 1.6 ± 0.1 Fish liver 10 40 ± 15 3.8 ± 1.8 a.r.HYD 4021 2615 ± 75 0.65 ± 0.07 hDHP kcat/ km(mM-1 min -1) kcat(min -1) km(mM) 41 33 ± 1 0.8 ± 0.04 Pig liver 790 1270 ± 80 1.6 ± 0.1 Fish liver 10 40 ± 15 3.8 ± 1.8 a.r.HYD 4021 2615 ± 75 0.65 ± 0.07 hDHP kcat/ km(mM-1 min -1) kcat(min -1) km(mM)
a. a.r.HYD represented Agrobacterium radiobacter hydantoinase. The kinetc constants of
imidases from fish (Oreochromis niloticus) liver, pig liver, and were a.r.HYD referred to
the thesis of Cheng-Yang Huang
b.The kinetic constants of hDHP with phathalimide as substrate were determined with UV/VIS
Table IV
Calibration curve determination of molecular weight on Sephacryl S-300 HR
0.03 40.06 2.84 692.00 Thyroglobulin 0.15 49.58 2.64 437.00 Ferritin 0.26 59.06 2.36 228.00 Catalase 0.28 60.36 2.25 178.00 Aldolase Kavb Vea log(MW) MW 0.03 40.06 2.84 692.00 Thyroglobulin 0.15 49.58 2.64 437.00 Ferritin 0.26 59.06 2.36 228.00 Catalase 0.28 60.36 2.25 178.00 Aldolase Kavb Vea log(MW) MW a. V
e represente the retention volume of eluted protein under the condition as indicated under
“Experimental Procedures”.
b. K
av was calculated by the equation: Kav = (Ve-V0)/(Vt-V0). The Vt was the column volume as
120 ml and the V0 was the retention volume of blue dextran 2000 (2000 KDa), whose
Table V
Physical and chemical parameters of hDHP
60.40 5.46 49939 a.r.Hydantoinase 54.31 6.58 59439 hDHP Veb Theoretical pIa Molecular weight a 60.40 5.46 49939 a.r.Hydantoinase 54.31 6.58 59439 hDHP Veb Theoretical pIa Molecular weight a
a. The molecular weight and theoretical isoeletric point(pI) were determined by
the ProtParam on ExPASy server (http://tw.expasy.org/tools/protparam.html).
b. The Ve indicated the retention volume as described above and determined by
Figure 1. SDS-PAGE (10%) of purified recombinant hDHP. Lane 1, pellet; lane 2,
supernatant; lane 3, Histrap elute after thrombin on-column digest. The steps of purification of
Figure 2. Purified recombinant hDHP analyzed by MALDI-TOF after trypsin-in-gel digestion. The purified recombinant hDHP on SDS-PAGE were excised and digested in gel
with trpsin according to Shevchenko’s method (Shevchenko et al., 1996). The digested samle
was taken up and analyzed using MADI-TOF. The results were correlated with the sequence
Temperature Profile of hDHP 0 5 10 15 20 25 30 10 15 20 25 30 35 40 45 Temperature (℃ ) Spe ci fi c A cti vi ty (μ mo l/ m in /m g
Figure 3. Temperature profile of hDHP. The temperature profile of hDHP is displayed by the
specific activity of hDHP with phathalimide at given temperature measured with a UV/VIS
pH Profile of hDHP
0 5 10 15 20 25 30 35 6 6.5 7 7.5 8 8.5 9 9.5 10 pH S p ec if ic A ct iv it y ( μ mol /mi n /mg)Figure 4. The pH profile of hDHP. The pH profile of hDHP is displayed by the specific activity of
hDHP with phathalimide at given pH measured with a UV/VIS spectrophotometer as indicated under
hDHP Thermostability 0 10 20 30 40 50 60 70 80 90 100 0 15 30 45 60 75 90 105 120 Time (min) R es id u al A cti vi ty (%) 30℃ 37℃ 50℃ 60℃
Figure 5. Thermal stability of hDHP. The thermal stability of hDHP is displayed by the
residual activity of hDHP with phathalimide measured with a UV/VIS spectrophotometer as
Effect of Urea Treatment 0 10 20 30 40 50 60 70 80 90 100 0 10 20 30 40 50 60 Time (min) R es id u al A cti vi ty (% ) 0.1M 1M 2M 3M 4M
Figure 6. Effect of urea treatment of hDHP. The effect of urea treatment of hDHP is
displayed by the residual activity of hDHP with phathalimide measured with a UV/VIS
spectrophotometer as indicated under “Experimental Procedure” after treating in given
Urea-induced fractional unfolding of hDHP 0 10 20 30 40 50 60 70 80 90 100 0 0.5 1 1.5 2 2.5 3 3.5 4 Urea Concentration (M) R es id ua l A cti vi ty (% )
Figure 7. Urea-induced fractional unfolding of hDHP. Determine the stability of urea
treatment of hDHP by measuring [Urea]50%, the concentration of urea at 50% residual activity
Figure 8. Calibration curve of Sephacryl S-300 HR. A molecular weight calibration curve,
which defined the relationship between the elution volumes of a set standards and the logarithm
of their respective molecular weights, was determined with Sephacyl S-200 HR. Calibration
standard(crisscross) used were High Molecular Weight standard of Gel Filtration Calibration
Kit, such as aldolase (178 KDa), catalase (228 KDa), ferritin (437 KDa), thyroglobulin (692
KDa), blue dextran (2000 KDa). The hDHP and a.r.HYD were closed and open diamonds,
Figure 9. hdhp-pET-43.1a (+) construct DNA sequencing. The DNA sequence of
hdhp-pET-43.1a (+) construct was analyzed according to Senger Method by MISSION
Michaelis-Menten
Phthalimide (mM) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 Activity ( umol/min /mg ) 0 5 10 15 20 25 30Figure 10. Enzyme Kinetics of hDHP with Phthalimide. Measurement of kinetic constant
of each substrate was performed by varying the concentration of one substrate, while keeping
the other substrate at a fixed and near saturating concentration. The apparent Km and Vmax
were determined using nonlinear regression by Sigmaplot 2001,V7.0 and Enzyme Kinetics
Module, V1.1.
Km= 0.66mM
Vmax= 44.3 umol/min/mg Km= 0.66mM
Appendix
Appendix 1. The structure organization and chromosome localization of hdhp gene. (http://www.ensembl.org/Homo_sapiens/geneview?gene=ENSG00000147647)
Appendix 2. Nucleotide sequence of cDNA and deduced amino acid sequence of hdhp gene. Analyze the nucleotide sequence of cDNA and deduced amino acid sequence of hdhp
Appendix 3. The identity of the amino acid sequences of imidase from other species and Homo Sapien. Analyze the the amino acid sequences of imidase from other species and Homo Sapien by ClustalW2( http://www.ebi.ac.uk/Tools/clustalw2/index.html
Appendix 5. The map of hdhp-pET-43.1a (+) construct. Clone the PCR products of cDNA of
hdhp gene of Homo sapien on pCMV-sports6 vecto into pGEM-T Vector (Promega) and add
restriction sites of BamH I and Xho I to the termini of amplified DNA with the pairing primers hdhp-F and hdhp-R . Subclone the DNA fragment containing the hdhp gene on pGEM-T vector into the modified pET-43.1a(+) vector by restriction enzyme, BamH I & Xho I, and T4 DNA Ligase.
Appendix 6. Physico-chemical parameters of hDHP. Analyze the Physico-chemical
parameters of hDHP by ProtParam tool on the ExPASy sever
Appendix 7. Multiple amino acid sequences alignment of imidase from other species and Homo Sapien. Analyze the Multiple amino acid sequences alignment of imidase from other species and hDHP by ClustalW2 ( http://www.ebi.ac.uk/Tools/clustalw2/index.html). The labeled amino acids, such as His67, His 69, Lys159, His192, His 248, and Asp 326 are responsible for metal binding.