綠原酸降低高血脂兔子氧化壓力與發炎相關基因表現; Chlorogenic acid reduce oxidative stress and inflammatory-related gene expression in hyperlipidemic rabbits
154
0
0
全文
(2) 中文摘要 動脈粥狀硬化是造成心血管疾病的主因。許多研究都指出氧化修 飾的 LDL 對於動脈粥狀硬化的發展扮演重要角色危險因子之ㄧ。綠原 酸 (chlorogenic acid, CA)是一種酚類物質,廣泛存在於蔬果和飲品 如:蘋果、西洋梨、漿果、萱草(Hemerocallis spp.)、紅茶和咖啡中。 先前研究發現,綠原酸具有抗氧化、抗腫瘤與免疫調節的能力。本研 究先以捕捉 DPPH 自由基、抑制 LDL 氧化作用以及總抗氧化能力 (trolox equivalent antioxidant capacity, TEAC)等體外試驗,探討綠原 酸的抗氧化能力,接著以補充綠原酸對於高脂血症兔子體內氧化壓力 與發炎相關基因表達進行探討。將紐西蘭大白兔隨機分為四組且飼養 8 週後犧牲,組別分別為餵食一般 rabbit chow 的正常組(N);給予高油 高膽固醇飲食的控制組(C);高油高膽固醇飲食並添加 0.1 % (w/w) probucol 的 probucol 組(P),以及添加相同劑量的綠原酸組(CA)。體外 試驗顯示,綠原酸捕捉 DPPH 自由基以及抑制 LDL 氧化作用較 probucol 佳,綠原酸的總抗氧化能力與 probucol 相當。動物實驗部份, 控制組相較於正常組有明顯地內膜增厚情形,綠原酸組和 probucol 組 之內膜增厚情形皆較控制組低。綠原酸組和 probucol 組相較於控制 組,皆可顯著降低血清中總膽固醇(TC)、LDL-C、三酸甘油脂(TG)、. I.
(3) MDA 的產量,以及胸主動脈 luminol-CL 的產生,並延長低密度脂蛋 白的遲滯期(lag time);綠原酸組和 probucol 組相較於控制組,皆可顯 著降低 VCAM-1、MCP -1、腫瘤壞死因子-α (tumor necrosis factor-α, TNF-α) 和腫瘤壞死因子-β (tumor necrosis factor-β, TNF-β) 之 mRNA 表現。綜合以上結果,綠原酸可藉由降低高脂血症兔子血清脂質、氧 化壓力,以及降低發炎相關基因(TNF-α、TNF -β)、VCAM-1 與 MCP -1 之 mRNA 表現,達到降低初期動脈粥狀硬化的發展。. II.
(4) 英文摘要 Atherosclerosis is a major cause of coronary heart disease. Previous studies have indicated that oxidized LDL plays an important role in initiation and progression of atherosclerosis. Chlorogenic acid (CA) is a phenolic compound present in fruits, vegetables and some drinks including apples, pears, berries, Chinese day lily (Hemerocallis spp.), black tea and coffee. It has been found to have antioxidant, antimutagenic, and immunomodulatory properties. In this study, we investigated the antioxidative effects of CA using the DPPH radicals scavenging ability, inhibition of LDL oxidation and trolox equivalent antioxidant capacity (TEAC) in vitro, and the effects of chlorogenic acid on oxidative stress and anti-inflammatory gene expression in hyperlipidemic rabbits. Twenty-four New Zealand White (NZW) male rabbits were assigned randomly into four dietary groups and were scarified after eight weeks. The normal group was fed regular rabbit chow and the control group was fed a high fat and cholesterol diet. The probucol group and the CA group were fed the same diet as the control group plus 0.1 % (w/w) probucol or chlorogenic acid. The result indicated that CA had better antioxidant activity than probucol in DPPH radicals scavenging ability and inhibition of LDL oxidation, and as good as probucol in TEAC. The serum levels of total cholesterol (TC), LDL-cholesterol (LDL-C), triglyceride (TG), MDA, luminol-CL of thoracic aorta and intimal thickening was significantly increased in the control group compared with the normal group, and decreased in the probucol and the CA group. The lag time of LDL oxidation in the control group was shorter than the normal group, and longer in the. III.
(5) probucol and the CA group. The mRNA expression of VCAM-1 (vascular cell adhesion molecule-1), TNF-α (tumor necrosis factor-α), TNF-β (tumor necrosis factor-β) and MCP-1 (monocyte chemotactic protein-1) was significantly increased in the control group compared with the normal group, and decreased in the probucol and the CA group. In conclusion, chlorogenic acid suppresses the progression of atherosclerosis by lowering serum lipid, reducing oxidative stress and inflammatory-related gene expression including TNF-α, TNF-β, VCAM-1and MCP-1 in hyperlipidemic rabbits.. IV.
(6) 總目錄 中文摘要. I. 英文摘要. III. 總目錄. V. 表目錄. VIII. 圖目錄. IX. 縮寫對照表. XI. 第一章. 研究設計原理與目的. 1. 第二章. 文獻回顧. 第一節. 動脈粥狀硬化的成因. 4. 第二節. 動脈血管壁的基本構造. 6. 第三節. 動脈粥狀硬化的發展. 8. 第四節. 自由基. 11. 第五節. 氧化壓力與動脈粥狀硬化. 15. 第六節. 抗氧化劑與動脈粥狀硬化. 20. 第七節. 發炎與動脈粥狀硬化. 24. 第八節. 黏附因子與動脈粥狀硬化. 27. 第九節. 單核球趨附因子與動脈粥狀硬化. 29. V.
(7) 第三章. 實驗設計. 31. 第四章. 材料與方法. 34. 第五章 結果 第一節. 綠原酸的抗氧化力. 62. 第二節. 實驗期間各組生長狀況的觀察. 62. 第三節. 血液生化值的影響. 63. 第四節. 綠原酸對抗氧化指標:血清脂質過氧化物 MDA、 LDL 過氧化遲滯期的影響. 65. 第五節. 綠原酸對血管 Luminol-CL 的影響. 66. 第六節. 綠原酸對發炎性基因:腫瘤壞死因子-α(TNF-α) 、腫瘤壞死因子-β(TNF-β)mRNA 表現量的影響. 第七節. 第八節 第六章. 66. 綠原酸對血管黏附因子(VCAM-1)、單核球趨化 因子(MCP-1)mRNA 表現量的影響. 68. 觀察兔子胸主動脈形態學改變之組織切片圖. 69. 討論. 第一節. 動脈粥狀硬化誘發模式與飲食劑量探討. 第二節. 餵食高油高膽固醇飲食和添加綠原酸對血液生化值. 91. 92. 的影響. VI.
(8) 第三節. 綠原酸和 probucol 的抗氧化作用. 第四節. 綠原酸對抗氧化指標: MDA 和 LDL 氧化遲滯期. 95. 98. 之影響 綠原酸對血管中 Luminol-chemiluminescence(CL). 第五節. 的影響. 101. 第六節. 綠原酸對 TNF-α 和 TNF-β mRNA 表現量的影響. 102. 第七節. 綠原酸對血管黏附因子(VCAM-1)和單核球趨化 因子(MCP-1)mRNA 表現量的影響. 104. 第八節. 綠原酸對兔子主動脈弓形態學改變之影響. 105. 第九節. 以補充 probucol 作為本研究正控制組之原因. 106. 第十節. 綠原酸的吸收、消化代謝與其衍生物之探討. 109. 第十一節. 114. 未來研究發展. 第七章. 結論. 116. 第八章. 參考文獻. 117. VII.
(9) 表目錄 表 1-1 九十四年度國人十大死因排行. 3. 表 3-1 即時定量聚合脢鏈鎖反應引子序列表. 60. 表 4-1 綠原酸和 probucol 的抗氧化能力. 70. 表 4-2 紐西蘭大白兔體重紀錄表. 71. 表 4-3 八週紐西蘭大白兔血液生化值與脂質過氧化指標和 LDL 遲滯期. 72. VIII.
(10) 圖目錄 圖 2-1 動脈血管壁的構造. 7. 圖 2-2 脂質過氧化路徑. 18. 圖 2-3 氧化 LDL 在動脈粥狀硬化的調控. 20. 圖 2-4 抗氧化劑在動脈粥狀硬化的角色. 21. 圖 2-5. 22. Probucol 的化學結構. 圖 2-6 綠原酸的化學結構. 24. 圖 2-7 NF-κB 對動脈粥狀硬化的調控. 27. 圖 3-1 體外實驗流程簡圖. 32. 圖 3-2 體內實驗流程簡圖. 33. 圖 4-1 血清總膽固醇於實驗期間零週、四週、八週的變化圖. 73. 圖 4-2 血清三酸甘油酯於實驗期間零週、四週、八週的變化 74. 圖 圖 4-3 八週兔子胸主動脈 Luminol-CL 圖. 75. 圖 4-4 綠原酸對兔子胸主動脈腫瘤壞死因子-α(TNF-α)mRNA 76. 表現量的影響 圖 4-5 綠原酸對兔子胸主動脈腫瘤壞死因子-β(TNF-β)mRNA. 77. 表現量的影響. IX.
(11) 圖 4-6 腫瘤壞死因子-α(TNF-α)的同步定量曲線圖. 78. 圖 4-7 腫瘤壞死因子-α(TNF-α)的熔點溫度曲線圖. 79. 圖 4-8 腫瘤壞死因子-β(TNF-β)的同步定量曲線圖. 80. 圖 4-9 腫瘤壞死因子-β(TNF-β)的熔點溫度曲線圖 圖 4-10. .. 81. 綠原酸對兔子胸主動脈血管細胞黏附因子(VCAM-1) mRNA 表現量的影響. 82. 圖 4-11 綠原酸對兔子胸主動脈單核球黏附因子(MCP-1) mRNA 表現量的影響. 83. 圖 4-12. 血管細胞黏附因子-1(VCAM-1)的同步定量曲線圖. 84. 圖 4-13. 血管細胞黏附因子-1(VCAM-1)的熔點溫度曲線圖. 85. 圖 4-14. 單核球趨化因子-1(MCP-1)的同步定量曲線圖. 86. 圖 4-15. 單核球趨化因子-1(MCP-1)的熔點溫度曲線圖. 87. 圖 4-16. 紐西蘭大白兔主動脈弓組織切片圖(倍率:40 倍). 88. 圖 4-17. 紐西蘭大白兔主動脈弓組織切片圖(倍率:200 倍). 89. 圖 4-18 綠原酸對紐西蘭大白兔動主脈弓內膜增厚的影響. X. 90.
(12) 縮寫對照表 ABTS:2,2'-azino-bis(3-ethylbenzothiazoline -6-sulfonic acid) BSA:Bovine serum albumin CA:Chlorogenic acid CHCl3:Chloroform Luminol-CL:Luminol-chemiluminescence CuSO4:Cupric sulfate DEPC:Diethyl pyrocarbonate DDW:Distilled water DPPH:1,1-diphenyl-2-picrylhydrazyl EDTA:Ethylene diamine tetraacetic acid 乙烯二氨四乙酸 GK:Glycerokinase GOP:Glycerolphosphate oxidase C:High cholesterol diet CA:High cholesterol + chlorogenic acid diet HDL-C:How density lipoprotein-cholesterol HE:Hematoxylin-eosin KBr:Potassium bromide. XI.
(13) KCl:Potassium chloride KH2PO4:Potassium dihydrogenphosphate LDL-C:Low density lipoprotein-cholesterol LPL:Lipoprotein lipase MDA:Malondialdehyde MCP-1:Monocyte chemoattractant protein-1 N:Normal diet NaCl:Sodium chloride Na2HPO4:Disodium hydrogenphosphate NZW rabbits:New Zealand White rabbits P:High cholesterol + probucol diet PBS:Phosphate buffer saline PCR:Polymerase chain reaction ROS:Reactive oxygen species RT:Reverse transcription TBA:Thiobarbituric acid TBHP:turt-Butyl hydroperoxide TCA :Trichloroacetic acid). XII.
(14) TEAC:Trolox equivalent antioxidant capacity TG:Triglyceride Trolox:6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid TNF-α:Tumor necrosis factor-alpha TNF-β:Tumor necrosis factor-beta VCAM-1:Vascular cell adhesion molecule-1 VLDL:Very low density lipoprotein β-actin:beta-Actin. XIII.
(15) 第一章. 研究設計原理與目的. 台灣九十四年國人十大死因中如表 1-1 所示,腦血管疾病和心臟疾 病分別為第二位及第三位。許多研究都指出,動脈粥狀硬化是導致心 臟、腦血管疾病的主要原因。常見的心血管疾病包括:冠心病、中風、 高血壓、風溼性心臟病。動脈粥狀硬化是一種慢性的發炎疾病與體內氧 化壓力增加有關。當低密度脂蛋白被氧化時,氧化低密度脂蛋白 (oxLDL)會誘發發炎相關的基因(例如:IL-1、TNF-α、TNF-β)和黏 附因子、趨化因子(如:VCAM-1、MCP-1)等基因表現,而誘發動脈 粥狀硬化的形成。 近年預防醫學漸漸受到人們的重視,於是飲食中的有效成分如:多 酚類的研究風潮日益興盛。多酚類普遍具有抗氧化的特性,期望藉由平 日飲食攝入有效成分,而對生理機能產生調節功能,達到預防或降低疾 病發生的效果。流行病學的研究也發現,多酚類的攝取與心血管疾病和 癌症的發生率呈反相關性。 本研究所使用的綠原酸(chlorogenic acid, CA)是廣泛存在於植物 中的多酚類,在蔬菜水果和飲品中如:蘋果、西洋梨、藍莓、蕃茄、紅 茶、咖啡都富含綠原酸。先前的研究也指出,綠原酸具有抗氧化、抗腫 瘤的特性。因此本篇之研究目的以高油高膽固醇飲食誘發紐西蘭大白兔. 1.
(16) 高脂血症,進而發展動脈粥狀硬化的模式,探討綠原酸是否能降低血脂 質、氧化壓力,並進一步以分子生物的角度,探討發炎反應相關基因、 細胞黏附因子和單核球趨化因子調控動脈粥狀硬化的角色。. 2.
(17) 表 1-1 九十四年度國人十大死因排行 順位. 死. 亡. 原. 因. 1. 惡性腫瘤. 2. 腦血管疾病. 3. 心臟疾病. 4. 糖尿病. 5. 事故傷害. 6. 肺炎. 7. 慢性肝病及肝硬化. 8. 腎炎、腎徵候群及腎性病變. 9. 自殺. 10. 高血壓性疾病. (摘自 2006/7/1 行政院衛生署 衛生統計資訊網). -3-.
(18) 第二章. 文獻回顧. 第一節. 動脈粥狀硬化的成因. 動脈粥狀硬化是一種主要發生在大血管如:胸主動脈的疾病,其在 中型血管和肌肉附近的血管也會發生。動脈粥狀硬化形成的斑塊破裂, 容易引發血管栓塞,導致心臟、腦部與手腳四肢的缺血而壞死,是造成 中風和心臟疾病的主因【1,2】。流行病學和解剖學的研究都顯示,身體 老化的過程、血管平滑肌細胞與動脈粥狀硬化有相關性。在沒有確切的 危險因子如:高血脂症、發炎因子如:TNF、IL-1 存在之下,老化過程 仍然會改變血管壁的型態,尤其是增加血管內膜的增厚,而內膜增厚是 動脈粥狀硬化初期很重要的特徵,且發現血管壁也有脂肪的囤積【3】。 但也有研究顯示,人的一生都有可能伴隨著動脈粥狀硬化的發生。在出 生的嬰兒胸主動脈中,發現有動脈粥狀硬化的形成,尤其是高膽固醇血 症的母親,更會加強血管壁有內膜增厚與單核球囤積的情形【4】 。 動脈粥狀硬化不只是一種老化過程,也是一種慢性發炎的狀態;一 旦有斑塊破裂或栓塞的情形產生,亦可能轉變成急性的臨床症狀【1,2】 。 介白素-1(interleukin-1, IL-1)和腫瘤壞死因子(tumor necrosis factor, TNF)是一種前發炎細胞激素(pro-inflammatory cytokine) ,會誘發其他. -4-.
(19) 細胞激素如:IL-4、IL-6、IL-10,和趨化因子如:單核球趨化因子的產 生,並且會促進血管內皮細胞黏附因子的表現。在 Apo-E 剔除小鼠模式 的實驗顯示,由於 IL-1 接受器拮抗劑(IL-1 receptor antagonist, IL-1 Ra) 是一種急性時期蛋白質(acute phase protein) ,IL-1 Ra 會與 IL-1 相互競 爭接受器,但卻不會產生下游發炎反應作用;因此,IL-1/IL-1 Ra 的比 值對於血管發炎和動脈粥狀硬化上扮演重要的角色【5】 。 愈來愈多研究者想了解參與動脈粥狀硬化的危險因子、致病路徑與 訊息傳遞路徑。許多危險因子,如:抽菸、高醣血症(hyperglycemia)、 高血壓、胰島素抗性、循環血液中有過多的脂肪,以及不足的運動習慣, 都對動脈粥狀硬化的發展有正向的影響【6】 。 低密度脂蛋白(low density lipoprotein ,LDL)體積大小和血漿中高 密度脂蛋白(high density lipoprotein ,HDL)的濃度有正相關,而和三酸 甘油酯(triglyceride, TG)的濃度有負相關,即血漿中有較多體積大的 LDL 時,會有較高濃度的 HDL 以及較低濃度的 TG 存在。因此,體積 較小、濃度較高的 LDL 與較低的 HDL-cholesterol 濃度,以及較高的三 酸甘油酯濃度—以上三者合併稱為「致動脈粥狀硬化生成之脂蛋白的表 現型(atherogenic lipoprotein phenotype)」 【7】 ,此種表現型可能會造成 代謝性症候群的產生,亦可能增加罹患心血管疾病的危險【8,9】 。. -5-.
(20) 許多研究都證實氧化修飾的 LDL,會導致動脈粥狀硬化的生成: (1)在體內實驗或體外試驗,皆發現氧化 LDL(oxLDL)會促進動脈 粥狀硬化的生成【10,11】 。 (2)在動脈粥狀硬化損傷的斑塊中,可以分離出類似受到氧化傷害 (oxidative damage)的脂蛋白粒子(lipoprotein-like particles) 【12】 。 (3)脂肪氧化產生的脂質過氧化終產物如:丙二醛(malondialdehyde) , 在人類和動物的動脈粥狀硬化斑塊中,可以用免疫化學染色法測 到【12】 。 (4)在高膽固醇血症動物,或非人類的哺乳類動物實驗中,發現有多 種不同的抗氧化劑可減低初期動脈粥狀硬化損傷的形成【11-14】 。 由此可見,脂蛋白的氧化傷害在動脈粥狀硬化中,扮演非常重要的角色 【13】。. 第二節 動脈血管壁的基本構造. 正常的胸主動脈血管壁在組織學上主要分為三層如圖 2-1 所示,血 管橫切面由外而內分別為: (1) 外層(adventitia):外層為血管壁的最外層,由結締組織和 散佈的纖維母細胞(fibroblasts)與平滑肌細胞(smooth muscle cell, SMC). -6-.
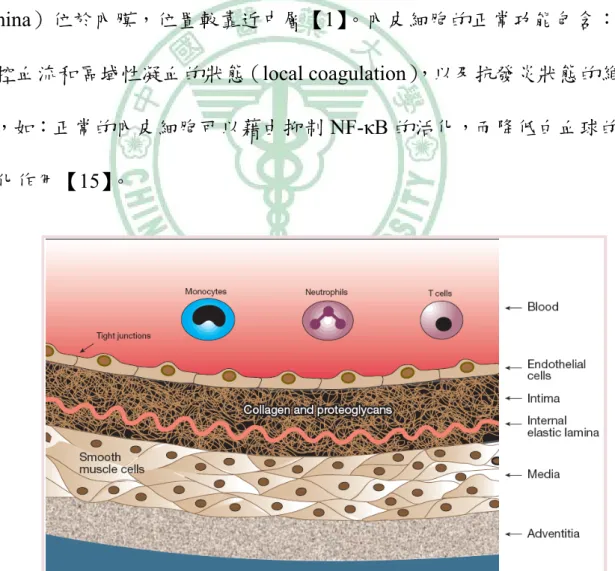
(21) 胞組成。 (2) 中層(media) :位於血管的中層,平滑肌細胞為其主要成分, 以及細胞外基質(extracellular matrix) 。 (3) 內膜(intima) :正常的內膜非常薄,包含接觸血流的內皮細 胞與細胞外結締組織基質(extracellular connective tissue matrix),如: 蛋白醣(proteoglycan)和膠原蛋白(collagen) 。內部彈力層(internal elastic lamina)位於內膜,位置較靠近中層【1】 。內皮細胞的正常功能包含: 調控血流和區域性凝血的狀態(local coagulation) ,以及抗發炎狀態的維 持,如:正常的內皮細胞可以藉由抑制 NF-κB 的活化,而降低白血球的 活化作用【15】。. 圖 2-1 動脈血管壁的構造. ( Lusis AJ. 2000 ). -7-.
(22) 第三節 動脈粥狀硬化的發展. 動脈粥狀硬化的進展主要分為四期【2】 : (1) 內皮功能不良(endothelial dysfunction). 早期的動脈粥狀硬化損傷發生在內皮組織,除了血流變化而導致的 shear stress 改變會造成內皮功能不良外【16,17】 ,可藉由一氧化氮(nitric oxide) 【17】、前列腺素(prostacyclin)、血小板衍生生長因子 (platelet-derived growth factor, PDGF)、血管收縮素Ⅱ(angiotensinⅡ) 和 endothelin 來增加血管的通透性【18】 ,而使得內皮層對血漿中脂蛋白 與其他組成物之通透性增加【19】 ;並且會促進白血球黏附因子(leukocyte adhesion molecules)、內皮黏附因子(endothelial adhesion molecules)的 表現【20-22】 ;接著,藉由 oxLDL 和一些趨化因子如:單核球趨化因子, 促使單核球進入血管內【23】。. (2) 脂肪條形成(fatty-streak formation). 脂肪條一開始是由充滿脂肪的單核球和巨噬細胞,以及 T 淋巴球所 組成,接著平滑肌細胞也會加入脂肪條形成的過程。此過程包括了平滑. -8-.
(23) 肌的遷移(migration) 、T 細胞的活化、泡沫細胞的形成,以及血小板的 黏附【24】。其中平滑肌的遷移是受到血小板衍生生長因子等生長因子 的刺激【25】 。T 細胞的活化則是因為腫瘤壞死因子 α(tumor necrosis factor-α, TNF-α) 【26】 、介白素 interlukin-2(IL-2) 、IL-15、oxLDL【27】 和顆粒球-巨噬細胞聚落刺激因子(granulocyte-macrophage colony-stimulating factor, GM-CSF)的刺激【28】。泡沫細胞的形成則是 受到 oxLDL【29】 、巨噬細胞聚落刺激因子(macrophage colony-stimulating factor, M-CSF) 【30】 、TNF-α 和 IL-1 的影響;而一些內皮細胞黏附因子, 如:intergrin、P-selectin,以及血小板凝集素 A2(thromboxane A2),被 認為和血小板的黏附有關【2】。. (3) 進一步複雜的動脈粥狀硬化損傷之形成(formation of an advanced, complicated lesion of atherosclerosis). 當脂肪條繼續發展成進一步的損傷時,就會形成纖維帽。纖維帽是 由平滑肌細胞、膠原蛋白與細胞外基質(extracellular matrix, ECM)所 組成【31】。此時,動脈粥狀硬化損傷的部份,仍有單核球不斷得黏附 至內皮細胞,並受到單核球趨化因子(monocyte chemotactic protein 1, MCP-1)的刺激而進入血管壁,接著受到 M-CSF 的刺激而分化成巨噬. -9-.
(24) 細胞,並且繼續吞噬 oxLDL 形成泡沫細胞而囤積於血管壁。泡沫細胞 受到細胞凋亡(Apoptosis)與壞死(necrosis)作用,導致充滿脂肪的壞 死核(necrotic core)形成;由於血小板衍生生長因子、TNF-α、IL-1 等 細胞激素的活化,以及降低結締組織的分解導致纖維帽的產生【32,33】 。. (4) 不穩定的纖維斑塊(unstable fibrous plaques in atherosclerosis)的 形成. 較薄的纖維帽通常會導致纖維帽破裂,而可能進一步引發血栓 (thrombosis)的情況。因為活化的巨噬細胞也會活化 T 細胞產生許多 -. 細胞激素如:IL-1、TNF-α、IFN-γ、Fas-L,以及 H2O2、O2• 等自由基, 進而導致平滑肌細胞凋亡,也降低細胞外基質(extracellular matrix)的 合成;再者,血管損傷處不斷有巨噬細胞的囤積,而活化的巨噬細胞會 放出基質金屬蛋白酶(matrix metalloproteinase, MMP)和蛋白質分解酵 素,基質金屬蛋白酶和蛋白質分解酵素均會分解細胞外基質,造成血管 壁細胞與細胞之間的接合(cell-to-cell connection)減弱,致使不穩定的 斑塊形成而使血管壁破裂導致血栓形成(thrombus formation),血栓透 過血流堵塞血管進而引起冠狀動脈栓塞(coronary embolism)或(腦動 脈栓塞(cerebral embolism),造成心肌梗塞(myocardial infarction)或. - 10 -.
(25) 腦中風的發生【34,35】 。血小板凝集(Platelet aggregation)在動脈粥狀 引起的栓塞(embolism)扮演重要的角色,血清脂質可以顯著地影響血 小板的凝集作用。血小板除了不會黏附到 VLDL 外,皆會黏附到其他的 脂蛋白上如:LDL、HDL【36】。除此之外,血中膽固醇的濃度會直接 影響血小板的凝集作用。有許多的研究都顯示:高膽固醇血症病患之血 小板凝集較敏感,可能是因為 oxLDL 的增加而造成血小板凝集敏感性 增加【37-40】 。. 第四節. 自由基. 任何含有一個或多個不成對電子的物質都可以稱為自由基(free radical)。 氧分子(oxygen, O2)是生命賴以存活的物質,常以雙分子的形式 穩定存在,卻同時也是許多活性物質的受質。目前有許多的研究都指 出,活性氧屬(reactive oxygen species, ROS)會參與一些病理過程,且 對於一些功能性反應的調節,為不可缺少的物質。概括來說,ROS 是氧 分子在不同環境下,與氫原子或氮原子進行的氧化作用或還原作用,所 形成的化合物,包含:可以直接由還原氧分子所產生的超氧陰離子 -. (superoxide, O2• )、具有生物活性的過氧化氫(hydrogen - 11 -.
(26) peroxide,H2O2) 、氫氧自由基(hydroxyl radicals, OH•) 、hypochlorite ion -. -. (OCl )和過氧亞硝基陰離子(peroxynitrite, ONOO )等含氧自由基 【41,42】 。 -. -. 氧分子 O2 接受一個電子而生成超氧陰離子(O2• )。O2• 在一般 代謝過程或身體接受陽光照射的時候,經由「活化(activation)」氧分 -. 子而產生,因此 O2• 為初級(primary)的 ROS;而初級 ROS 再藉由酵 素活性或過度金屬催化的過程,會進一步和其他分子產生次級 -. (secondary) ROS 如:H2O2。O2• 在正常生理環境的 pH 值下非常脆 -. 弱,因為 O2• 為水溶性,所以無法輕易的通過細胞膜的脂質雙層,但 -. 卻可以穿越離子通道進入細胞【43,44】 。但 O2• 在較低的 pH 值(pKa = 4.8)會和 H+變成 H2O2,而在正常生理 pH 值下,則需要超氧歧化酶 (superoxide dismutase, SOD)的協助,反應如下: -. 2 O2• + 2 H+. SOD. H2O2 + O2. 而且 dismutation 的速率非常快【45】 。身體的調控在正常情況下,細胞 內並沒有游離鐵離子(free iron)存在;但當體內呈現壓力狀態時(under -. stress conditions) ,O2• 就會刺激游離鐵離子從含鐵蛋白中游離出來, 且研究指出人在高膽固醇血症、糖尿病和高血壓的情況下,體內 O2• 濃度都會增加【46】。. - 12 -. -.
(27) 過氧化氫(H2O2)相較於超氧陰離子,則較易進出細胞膜脂質層, 這也使過氧化氫成為活性氧屬和細胞訊息傳導中,重要的生物活性物質 【47,48】 。超氧陰離子會刺激體內的抗氧化劑和酵素如:catalase、 peroxidase,將 H2O2 分解成水和氧分子,以維持細胞內 H2O2 的正常濃 度。當身體處於壓力狀態下,超氧陰離子也會刺激含鐵蛋白釋放游離鐵 (free iron) ;此時,H2O2 便可和這些過量的金屬離子如:Fe2+、Cu+會經 由 Fenton reaction 產生氫氧自由基,反應式如下【49】 : -. Fe2+ + H2O2. Fe3+ + OH•+ OH. 若超氧陰離子接著參與二價鐵離子的還原作用,反應式如下: Fe 2+ + O2•. -. Fe3+ + O2. 合併以上和 Fenton reaction 兩個作用,稱為 Haber-Weiss reaction,反應 式如下: -. O2• + H2O2. -. O2 + OH•+ OH. 氫氧自由基(hydroxyl radical, OH•)具有高度的反應性,在體內 的半衰期非常短,所以 OH•會就近與其他物質進行反應。氫氧自由基 的產生方式有以下幾種: (1)當水分子被輻射線照射後,即可使水分子 分解形成 OH•和 H+;(2)在體內主要透過 Fenton reaction 進行過度金 屬電子轉移而生。OH•也會和 DNA 的鹼基或 DNA 去氧核酸的骨幹. - 13 -.
(28) (backbone)反應,導致 DNA 鹼基或序列傷害或斷裂【50】。 氧 衍 生 的 自 由 基 還 有 過 氧 自 由 基 ( peroxyl radical 有 時 亦 稱 peroxide) ,過氧自由基是高能產物,依據 R group、所屬的環境、當時 氧分子的濃度和反應物的不同,可從+ 0.77V 還原電位至+ 1.44V。脂質 過氧化作用是目前最常被研究者偵測與觀察的項目,也代表了過氧自由 基參與人類多種疾病發病與進程的證據【50-52】。 超氧陰離子會迅速地與一氧化氮(nitric oxide, NO•)反應而產生 -. 過氧亞硝基陰離子(peroxynitrite, ONOO )反應式如下: NO• + O2•. -. -. ONOO. 藉由使 NO•對血管舒張的作用失活而導致內皮功能不良。另外,活性 氧屬(reactive oxygen species, ROS)也會和血管裡的多元不飽和脂肪酸 (polyunsaturated fatty acid, PUFA)反應,開始脂質過氧化的連鎖反應。 脂質過氧化所產生的氫過氧自由基(HOO•) ,也會和 NO•反應而產生 -. ONOO ,並直接降低內皮對 NO•合成【46;50-52】 。 由以上自由基的介紹中發現,自由基與人體生理機能調控、代謝調 控與疾病都息息相關。. - 14 -.
(29) 第五節. 氧化壓力與動脈粥狀硬化. 活性氧屬(reactive oxygen species, ROS) ,可以參與眾多生化反應, 有時候會對生物系統產生有害的影響。在哺乳類的細胞中,有許多的來 源都可以產生活性氧屬,如:NADH/NADPH oxidase【53,54】 、Xanthine oxidase【55】 ,代謝花生四烯酸路徑裡的脂氧酶(lipoxygenase) 【56】和 環氧酶(cyclooxygenase) 【57】 。在正常生理狀態下,細胞粒線體的氧化 代謝作用就會產生活性氧屬,如:超氧陰離子。哺乳類的細胞擁有超氧 -. 歧化酶(superoxide dismutase, SOD),可以將兩個超氧陰離子(O2• ) -. 轉化(dismutate)成一個過氧化氫(H2O2) 【50】 。而 H2O2 和 O2• 可藉 由 Fenton reaction 產生活性強大的氧化物如:氫氧自由基(hydroxyl radical, OH•)。在正常生理狀況下,體內自由基和抗氧化劑與抗氧化 酵素會相互作用已達到平衡狀態。例如:過氧化氫會被 catalase 分解成 ,或透過 peroxidase 將其他受質氧化而抑制 H2O2 的累 水和氧分子(O2) 積,反應如下: 2 H2O2. catalase. substrate-H2 + H2O2. 2 H2O + O2 peroxidase. substrate + 2 H2O. 另外,glutathione peroxidase 廣泛的存在於體內可以和 catalase 共同作 用,藉由還原 glutathione(GSH),移除體內的 H2O2,反應式如下:. - 15 -.
(30) 2 H2O2 + 2GSH. glutathione peroxidase. 2 H2O + GSSG. glutathione peroxidase 更可以和 H2O2 之外的過氧化自由基反應,最受矚 目的是其可以還原血管壁內的 LOOH(lipid hydroperoxide)成脂醇類, 反應式如下: LOOH + 2GSH. glutathione peroxidase. LOH + H2O + GSSG. 所以在正常的生理狀態下,超氧歧化酶、peroxidase 和 catalase 是主要控 -. 。 制細胞內 H2O2 和 O2• 平衡的內生性抗氧化酵素系統【49】 其他的酵素系統,NOS(nitric oxide synthase)也被認為會調控細胞 內活性氧屬的產生。此種酵素有三種異構型,有兩種為正常狀態就會表 現的 eNOS(endothelial NOS)和 nNOS(neuronal NOS) ,另一種為誘發 型的 iNOS(inducible NOS)【53,58】 。 當 ROS 的產生和抗氧化劑之間不平衡,而傾向有較多的活性氧屬 -. 如:O2• 、OH•和脂質過氧化自由基(lipid peroxyl radicals)存在時, 「氧化壓力」因此產生並會對細胞造成傷害。過多 ROS 累積於血管內 皮細胞和平滑肌與心血管疾病如:缺血或再灌流所引發的發炎傷害 (ischemia/reperfusion-induced inflammatory injury)、缺血性心臟病、動 脈粥狀硬化、高血壓和心臟衰竭有高度相關性【59,60】。活性氧屬也被 認為可直接或間接影響細胞的訊息傳遞,氧化還原敏感性的轉錄因子 如:NF-κB 和 AP-1 都是活性氧代謝物的受質【61】。透過活化 NF-κB. - 16 -.
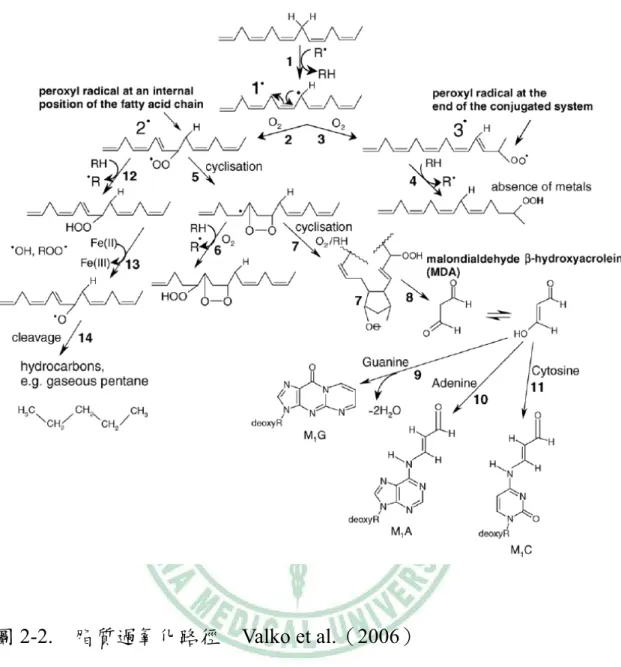
(31) 和 AP-1 則會導致血管的功能失調,包含:內皮功能不良、血管對 LDL 的通透性增加、促進內皮中 LDL 的氧化,也會增加血管內皮細胞上的 黏附因子表現與促進發炎反應【62,63】 ,而更高濃度的 ROS 則會導致細 胞死亡【61】 。 當氧化壓力升高時,體內常發生脂質過氧化(lipid peroxidation) , 尤其是 LDL 脂質過氧化作用和動脈粥狀硬化初期發展有著密不可分的 關係。巨噬細胞、血管平滑肌細胞或內皮細胞所產生的自由基會攻擊 LDL 內的不飽和脂肪酸(unsaturated fatty acid, LH),使其 α-methylene 產生脫氫化作用(hydrogen abstraction),而形成脂質烷基自由基(lipid alkyl radical, L•) ;由於在 lipid alkyl radical 上的電子非常不穩定,所以 會形成共軛雙烯(conjugated diene alkyl radical)反應如下: LH + OH•. L•+ H2O. 接著氧分子和 conjugated diene alkyl radical 反應則形成 lipid peroxyl radical (LOO•) ;而 lipid peroxyl radical 會繼續攻擊旁邊的不飽和脂肪 酸,形成 lipid hydroperoxide(LOOH)和 lipid alkyl radical(L•) ,而產 生一發不可收拾的脂質過氧化鏈鎖反應,如圖 2-2 所示,反應如下: L•+ O2 LOO•+ LH. LOO• L•+ LOOH. - 17 -.
(32) 圖 2-2. 脂質過氧化路徑. Valko et al.(2006). 當有金屬離子存在時,LDL 內的 lipid hydroperoxide(LOOH)會和 Fe2+ 或 Cu+進行 Fenton reaction,產生脂質烷氧基自由基(lipid alkoxyl radical, LO•)和 OH•;接著,再經過 β-scission 後產生帶有醛基的脂質過氧 化終產物如:malondialdehyde(MDA),這些產物會和 LDL 上的 Apo. - 18 -.
(33) B-100 結合而使 LDL 帶負電,因此,氧化修飾的 LDL 較易被巨噬細胞 上的清道夫接受器(scavenger receptor)辨識【64-68】 。 氧化修飾的 LDL(oxLDL)被認為可以透過下列機制,有效的影響 動脈粥狀硬化的生成,如圖 2-3 所示【166】 : (1) oxLDL 促進巨噬細胞的吞噬作用而產生泡沫細胞。 (2) oxLDL 具有細胞毒性。 (3) oxLDL 本身就會促進平滑肌細胞和巨噬細胞的有絲分裂,即具有促進 增生的作用。 (4) oxLDL 會促進巨噬細胞上清道夫接收器(scavenger receptor)的表現。 【69】 (5) oxLDL 具有免疫調節性,可以活化 T 細胞的功能。【27】 (6) oxLDL 會誘發細胞激素如:PDGF 的產生,造成血小板的凝集(platelet aggregation)。 若能抑制 LDL 的氧化作用,便能控制動脈粥狀硬化的發展【69】 。. - 19 -.
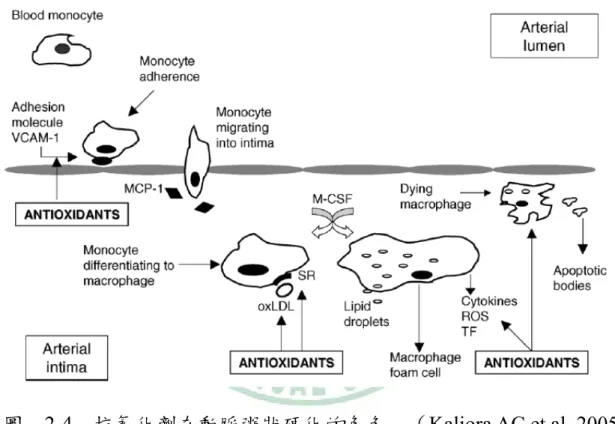
(34) 圖 2-3 氧化 LDL 在動脈粥狀硬化的調控. 第六節. (Diaz M et al., 1997). 抗氧化劑與動脈粥狀硬化. 多攝取蔬菜水果不只能降低油脂或其他高卡路里食物的攝取,其更 富含許多抗氧化物,如:維生素 C、E、β-carotene、類黃酮和多酚類, 可以移除或捕捉自由基【70】。類黃酮是廣泛存在於植物中的酚類代謝 產物,在一般飲食的蔬菜水果、咖啡豆和茶葉中也有。流行病學的研究 和動物模式的研究都顯示,大量攝取蔬果類食物中的酚類化合物有益健 康【71】 。紅酒裡的類黃酮物質 resveratrol,可以藉由抑制核酸因子 AP-1. - 20 -.
(35) 的活化和 endothelin-1 的基因表現,進而抑制金屬基質蛋白酶-9 (matrix metalloproteinase-9)的 mRNA 表現;且 resveratrol 透過減低活性氧屬形 成而部份影響 ERK1/2 路徑。因此,如圖 2-4 所示,補充抗氧化劑可能 具有降低動脈粥狀硬化的發展【72】 。. 圖. 2-4 抗氧化劑在動脈粥狀硬化的角色. (Kaliora AC et al.,2005). < 綠原酸(chlorogenic acid)> 綠原酸(chlorogenic acid, CA)是酚酸(phenolic acid)中桂皮酸 (cinnamic acid)的一種,結構如圖 2-4 所示,經由咖啡酸和奎寧酸(qunic acid)酯化作用而成【73】 ,。廣泛的存在於植物中,在下列的食物中也. - 21 -.
(36) 有發現,如:蘋果、西洋梨、漿果類、洋薊、茄子、咖啡【74】紅茶【75】 萱草(Hemerocallis) 【76】蕃茄【77】 ;CA 在蕃茄皮萃取物中佔總酚類 的 50.3 %,並且擁有抗氧化、抗微生物【77~79】 、抗腫瘤、抗癌的特性 【80,81】 ,是人類飲食中最豐富的多酚類之ㄧ。有研究顯示,CA 可以促 進細胞抗氧化酵素的活性,且抑制由 ROS 所媒介的 NFκB、AP-1 和 MAPK 的活化,對抗環境中的致癌物【82】 。CA 可以降低(fa/fa)Zucker 大鼠肝臟與血漿中膽固醇、三酸甘油酯的濃度,並且改善肝臟、血漿中 礦物質濃度和葡萄糖耐受情形【83】 ;但並未發現 CA 參與肝臟上胰島素 接受器(insulin receptor)和其 ligands 的調控,也未參與肝臟 insulin receptor 數量多寡的調控,只發現 CA 可改善 glucose-insulin index,且促 進肝臟中脂蛋白之蛋白質和 DNA 的濃度【84】 。在希臘香草植物貓薄荷 (Nepeta sibthorpii)的甲醇萃取物中,也發現有少量 CA 的成分,其萃 取物具有捕捉 DPPH 自由基的能力與降低水腫的能力【85】 。. 圖. 2-5 綠原酸的化學結構. (Azuma K et al., 2000). - 22 -.
(37) < Probucol > Probucol,4,4’-(isopropylidenedithio)bis(2,6-di-tertbutylphenol), 是一種廣泛被使用的降膽固醇藥物,且因此而被認為具有抗動脈粥狀硬 化的作用,其化學結構如圖 2-5。1986 年由 Parthasarathy 等人在細胞實 驗中發現,probucol 可以抑制銅離子所誘發的 LDL 氧化修飾作用【86】 。 餵食 2 %膽固醇飲食的紐西蘭大白兔的動物模式,萃出紐西蘭大白兔內 皮細胞與單核球進行培養,發現 probucol 可以降低單核球的黏附作用 【87】 。Probucol 也可以降低由自由基所引發的內皮功能不良情形【88】 , 且可以降低高血壓紐西蘭大白兔胸主動脈內膽固醇的氧化【89】。在 Apo-E 踢除的小鼠中,發現 probucol 可以降低胸主動脈之動脈粥狀硬化 生成,且會降低胸主動脈中巨噬細胞的囤積【90】 ;在 Apo-E 和 HDL 接 受器(scavenger receptor class B)基因雙重踢除的小鼠中,發現 probucol 可以藉由降低動脈粥狀硬化生成而有效地預防冠心病的發生和死亡 【91】 。以人類臍靜脈內皮細胞的細胞研究模式中,probucol 可以選擇性 抑制由 TNF-α 和 IL-1 所誘發的 VCAM-1 mRNA 的表現,但卻不會抑制 ICAM-1 mRNA 的表現【92】。研究指出,具有抗氧化效果的 probucol 除了可以降低 LDL-C 的濃度外,藉由促進透過 HDL-C 對膽固醇的傳 輸,幫助週邊組織過量膽固醇運回肝臟,因而具有抗動脈粥狀硬化的作. - 23 -.
(38) 用【93】 。. 圖. 2-6 Probucol 的化學結構. 第七節. (Kuzuya M and Kuzuya F. 1993). 發炎與動脈粥狀硬化. 動脈粥狀硬化是一種慢性的發炎疾病,其特徵包含前發炎細胞激素 的產生,如:腫瘤壞死因子(tumor necrosis factor, TNF) 、介白素-1(Interlu kin-1, IL-1)、介白素-6(Interlukin-6, IL-6)、介白素-10(Interlukin-10, IL-10) 、介白素-12(Interleukin-12, IL-12) ,和氧化脂質先質(pro-oxidative lipids)的形成,以及免疫反應的產生【94】 。氧化態的 LDL 具有發炎性, 會造成慢性的血管發炎【95】。在動脈粥狀硬化的損傷中發現有巨噬細 胞、T 細胞、前發炎細胞激素和黏附分子的出現,映證了免疫反應參與 動脈粥狀硬化發展的假說;顯示多元化的影響因子,如:發炎反應、感 染、免疫系統都參與了這個過程【94】. - 24 -.
(39) <腫瘤壞死因子(tissue necrosis factor, TNF)> 腫瘤壞死因子(TNF)是具有多功能性的蛋白質,不但可以作為促 發炎反應因子,也可以調節免疫功能。腫瘤壞死因子家族中的 TNF-α, 主要是由活化的巨噬細胞所分泌,在一些非專一性的機制,如:吞噬作 用或 LPS 刺激所導致的巨噬細胞活化作用,都會產生 TNF-α 和 IL-1 這 類的細胞激素【96】 ,除了巨噬細胞外,淋巴球【97】 、單核球、噬中性 球(neutrophils) 、自然殺手細胞(natural killer cells)和 T 細胞在 LPS 的刺激下,也會產生 TNF-α【98】。而腫瘤壞死因子-β(tumor necrosis factor-β, TNF-β)則主要是由 T 細胞和 B 細胞分泌,TNF-β 和 TNF-α 的 胺基酸序列非常類似【99, 100】 ,而 TNF-β 又被稱為 lymphotoxin(LT-α) ; TNF-β(LT-α)會和膜蛋白 TNF-C(LT-β)以 heterodimer 的方式共同表 現(coexpress),成為 LT-αβ(lymphotoxin-αβ)的複合體,廣泛存在於 細胞膜的蛋白質上;LT-α1β2 可以和 TNF 家族的接受器—LTβR(lympho toxin-β receptor)結合。當淋巴細胞被活化,或一些細胞激素、趨化因 子,如:IL-4、IL-7 存在時,可以誘發 LT-αβ 的活化【101】 。 有研究認為 TNF-β 對於有高胰島素血症的冠狀血管疾病患者而 言,是一種危險因子,因為 TNF-β 會引發更複雜且敏感的代謝症候群徵 狀(metabolic syndrome) 【102】 。以微矩陣基因晶片(micro array)來探. - 25 -.
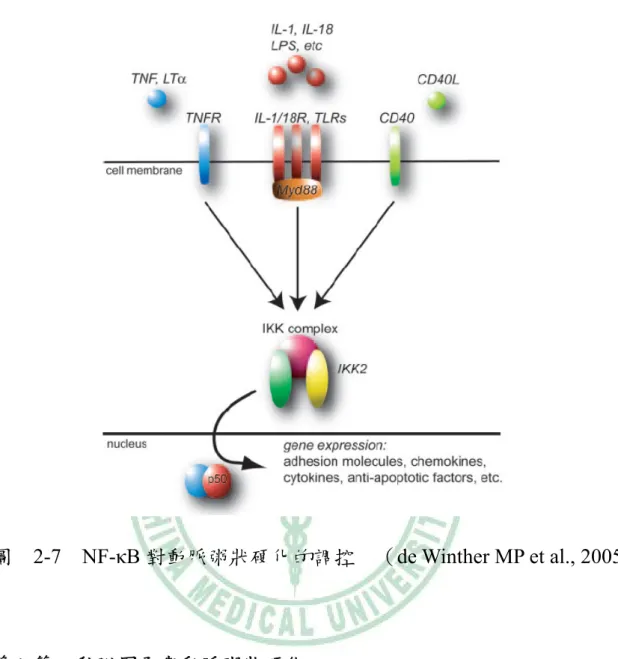
(40) 討人類基因和心肌梗塞的相關性中,發現 TNF-β 基因會因為族群不同而 對心肌梗塞有不同的關連性,日本人擁有 TNF-β 基因卻和心肌梗塞沒有 相關;但以德國人為受試者的實驗,則顯示 TNF-β 基因和心肌梗塞有相 關【101】 。TNF-β 基因踢除(TNF-β -/-)的小鼠相較於野生型(wild-type) 小鼠,TNF-β 基因踢除小鼠可以下降約 62 %的動脈粥狀硬化損傷型成 【100】 。如圖 2-7 所示,細胞激素如:TNF 和 IL-1 會透過活化 IKK(I-κB kinase)的活性,促使 I-κB 磷酸化而與核酸因子(nuclear factor-κB, NF-κB)分離,導致核酸因子(nuclear factor-κB, NF-κB)活化而進入細 胞核中與 DNA 結合,接著進行下游基因如:黏附因子、趨化因子表現 【96】 ;且 TNF 也會和 IL-1 藉由產生 ROS(reactive oxygen species) ,對 傷口產生氧化壓力和發炎作用【98】 。有研究指出,中藥藿香裡的 tilianin 可以有效的藉由抑制 NF-κB 活化,來降低由 TNF-α 所活化的血管黏附 因子 VCAM-1 的表現【103】 ;顯示 TNF-α 和 TNF-β 在動脈粥狀硬化的 發展中,對氧化壓力和發炎作用都扮演著調控的角色【98,100,103】。. - 26 -.
(41) 圖. 2-7 NF-κB 對動脈粥狀硬化的調控. 第八節. (de Winther MP et al., 2005). 黏附因子與動脈粥狀硬化. 細胞黏附因子就是表現在細胞表面的分子,作為此細胞和其他細胞 或細胞外基質黏附的媒介。約在 1993 年,研究者對於血管內皮細胞和 循環血液中白血球上的細胞黏附因子已有廣泛的認知【104】 。而這些可 溶性的黏附分子大致分為三類:. - 27 -.
(42) (1)Selectins:包含:噬酸性中性球(eosinophils) 、單核球、淋巴 球表現的 L-selectin(CD62L)、內皮細胞表現的 E-selectin(CD62E)、 內皮細胞和血小板表現的 P-selectin(CD62P)等。 (2)Immunoglobulin superfamily 為免疫球蛋白的超級家族,如:血 管細胞黏附因子(vascular cell adhesion molecule, VCAM-1)、細胞間黏 附因子(intercellular adhesion molecule, ICAM-1)【104】 。 (3)Integrins:大多分佈於淋巴球、單核球與顆粒球上,每種不同 的細胞帶有不同的 intergrins,包括: (A)VLA-4(α4β1: CD49d/CD29):分佈於噬酸性白血球 (eosinophils) 、淋巴球與單核球。 (B)LFA-1(αLβ2: CD11α/ CD18):分佈於多核型態白血球 (polymer-phonuclear leukocytes, PMN)、單核球和淋巴球。 (C)Mac(αMβ2: CD11b/CD18):分佈於多核型態白血球、單核 球、自然殺手細胞與 T 細胞【105】 。 在高血壓病患之血漿中的可溶性黏附因子,如:VCAM-1、ICAM-1 較 正常血壓的人高【106,107】 ,但 E-selectin 則和血壓正常者沒有顯著差異 【107】 。對於高血壓合併出現高膽固醇血症病患,高膽固醇血症會增強 血漿可溶性黏附因子的活性【106】 。高三酸甘油酯症年輕人和正常年輕. - 28 -.
(43) 人相比,其 VCAM-1、ICAM-1 濃度較高,且血管舒張力(brachial artery flow mediated vasodilation)亦不正常【108】 。細胞表面所表現的黏附因 子,除了參與細胞的遷移(migration) 、訊號傳遞和其他血管生理反應, 其亦可與內皮和血球細胞作用,是造成初期動脈粥狀硬化發展的重要步 驟【109】 。在動脈粥狀硬化損傷血管的內皮細胞模式中,發現 VCAM-1、 ICAM-1、E-selectin 與動脈粥狀硬化的生成有相關性。因此,偵測循環 血液中的黏附分子不只能代表內皮功能不良,更是早期動脈粥狀硬化的 指標【110】。 VCAM-1(CD106)專一性的分佈於血管內皮細胞上,可以辨識表 現在噬酸性白血球(eosinophils)上的 α4-integrin,此醣蛋白也會出現在 單核球和淋巴球上,但卻不會出現在中性顆粒球(neutrophils)上。抑 制 VCAM-1 和 α4-integrin 的結合,則可抑制嗜酸性白血球、淋巴球、單 核球黏附到血管內皮細胞,預期可有效治療多種發炎性疾病【105,111】 。. 第九節. 單核球趨化因子與動脈粥狀硬化. 循環血液中的單核球和 T 細胞會不斷黏附到血管內皮上,此為早期 動脈粥狀硬化生成的標記【112】 。單核球趨化因子(monocyte chemoattrac tant protein 1 ,MCP-1)對於黏附於血管內皮細胞的單核球之進入血管壁. - 29 -.
(44) 內膜扮演重要角色。許多的細胞都會分泌 MCP-1,如:內皮細胞、平滑 肌細胞、巨噬細胞,而這些細胞在動脈粥狀硬化的生成都佔有舉足輕重 的地位【113】 。 臨床的研究中,發現血漿中的 MCP-1 的濃度和心血管疾病如:高 脂血症的危險因子相關;且在急性心肌梗塞相較於穩定心絞痛的病患, 血中有較高的 MCP-1 濃度【114】。血管平滑肌細胞內的氧化脂質會透 過 NF-κB,調控 MCP-1 的基因表現【115】 。紐西蘭白兔的動物模式中, 給予 0.5 %膽固醇飲食相較於正常飲食兔子之胸主動脈 MCP-1 mRNA 和 蛋白質的表現量都較高,而在給予 estradiol 後則發現其可降低高膽固醇 飲食兔子胸主動脈 MCP-1 mRNA 和蛋白質的表現【116】;亦有研究指 出,補充紐西蘭白兔類黃酮物質如:naringin 和 naringenin,則可以降低 肝中 ACAT (acyl-CoA: cholesterol acyltransferase)的活性,以及胸主動 脈 VCAM-1 與 MCP-1 mRNA 的表現【117】。. - 30 -.
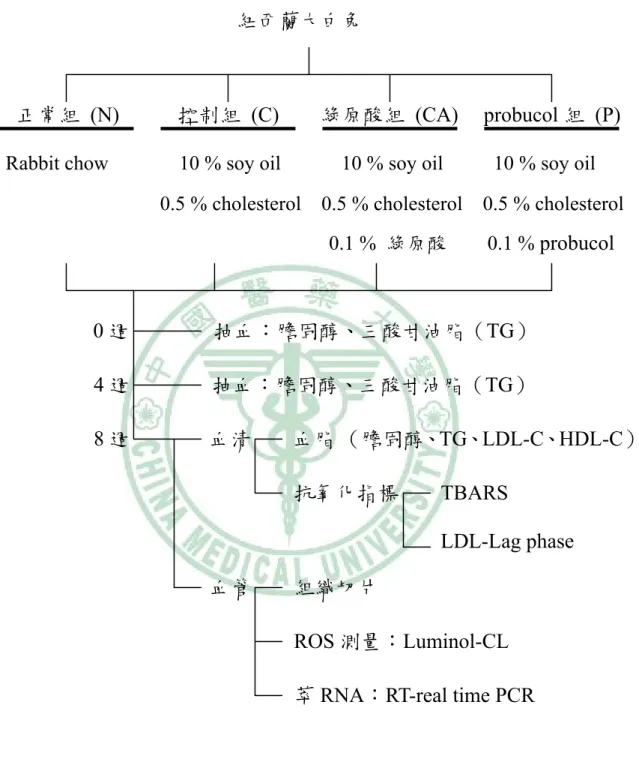
(45) 第三章 實驗設計. 先以體外試驗:捕捉 DPPH 自由基、抑制 LDL 氧化與總抗氧化能 力(trolox equivalent antioxidant capacity, TEAC)測定綠原酸和 probucol 的抗氧化能力,接著進行動物實驗。 以紐西蘭大白兔(New Zealand White rabbits, NZW rabbits)餵食高 油高膽固醇飲食導致高血脂症,且誘發血管動脈粥狀硬化的模式來進行 研究。 紐西蘭大白兔飼養於動物中心,飼養於十二小時交替的日夜循環, 室溫控制於 21±1 ℃、溼度約 40~60 %,每隻兔子單獨飼養於籠子中。 進入實驗期前,先以 rabbit chow 飼養一到兩個禮拜,使兔子適應 100 g/d 的飼料攝取量。24 隻紐西蘭大白兔體重約 2~2.5 kg,隨機分為四組,分 別為餵食 rabbit chow 的正常組(normal group,N);第二組為餵食含有 10 %大豆油、0.5 %膽固醇的高油高膽固醇組(high cholesterol diet group,C);第三組則是餵食高油高膽固醇組加 probucol(1 g/Kg diet) (high cholesterol + probucol diet group,P) ;第四組則是餵食高油高膽 固醇組加綠原酸(1 g/Kg diet) (high cholesterol + chlorogenic acid diet group,CA) ,且自由給予飲水。每天記錄飲食量,每週記錄兔子的體重,. - 31 -.
(46) 以作為飲食與兔子正常生長狀況的評估。實驗期為 8 週,於 0 週、4 週 進行耳靜脈抽血,進行抽血實驗前禁食 12 小時;兔子於 8 週利用 CO2 進行犧牲,犧牲前亦禁食 12 小時。犧牲後,取出的組織分別依實驗不 同,暫時存放於 DEPC 水或 PBS 進行清洗分裝,並立即冰入-80 ℃冰箱。 實驗流程簡圖如下:. 圖 3-1 體外實驗流程簡圖 Chlorogenic acid. 捕捉 DPPH 自由基. Probucol. 抑制 LDL 氧化. 總抗氧化能力測試 (TEAC). 進行體內試驗 備註:TEAC:Trolox equivalent antioxidant capacity. - 32 -.
(47) 圖 3-2 體內實驗流程簡圖 紐西蘭大白兔. 正常組 (N) Rabbit chow. 控制組 (C). 綠原酸組 (CA). 10 % soy oil. 10 % soy oil. probucol 組 (P) 10 % soy oil. 0.5 % cholesterol 0.5 % cholesterol 0.5 % cholesterol 0.1 % 綠原酸. 0.1 % probucol. 0週. 抽血:膽固醇、三酸甘油脂(TG). 4週. 抽血:膽固醇、三酸甘油脂(TG). 8週. 血清. 血脂 (膽固醇、TG、LDL-C、HDL-C) 抗氧化指標. TBARS LDL-Lag phase. 血管. 組織切片 ROS 測量:Luminol-CL 萃 RNA:RT-real time PCR. - 33 -.
(48) 第四章. 材料與方法. < 一 > 體外試驗-1:捕捉 DPPH 自由基(DPPH radicals scavenging ability). A 器材: DPPH(Sigma, MO, USA) B 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) C 原理: DPPH(1,1-diphenyl-2-picrylhydrazyl)可溶於酒精中,呈現深紫色, 被抗氧化劑捕捉後會呈現淺黃色,且在 517 nm 有最大吸光值,所以利 用其顏色變化的原理來評估物質的抗氧化性。 D 步驟: 將不同濃度的 chlorogenic acid 和 probucol 分別加入 DPPH 自由基之 甲醇溶液,混和 30 min 後,以 spectrophotometer 在波長 517 nm 測吸光 值。整個實驗流程均需避光進行。當吸光值愈低表示清除 DPPH 自由基 能力愈強,以未添加 chlorogenic acid 或 probucol 為控制組,以控制組吸 光值為 100 % 來作比較。紀錄抑制 50 % DPPH 自由基產生之 chlorogenic. - 34 -.
(49) acid 和 probucol 的濃度(IC50)。 E 計算方式: 清除率(%) = 控制組吸光值-實驗組吸光值 X 100 % 控制組吸光值. < 二 > 體外試驗-2:抑制 LDL 氧化作用. A 儀器: ELISA plate reader(μ QUANT, Bio-Tek, USA) B 步驟: 從健康人的血清中分離出 LDL,先進行蛋白質定量並經 PBS 稀釋 至 0.9 mg/ml 後均分至各試管中。之後各試管再加入不同濃度綠原酸和 probucol,並以 50 μM CuSO4 誘發氧化,再利用 EDTA 終止氧化。加入 2-propanol 沈澱蛋白質,並以 12000 rpm 離心 10 分鐘後,吸取上清液在 波長 234 nm 測吸光值,以未添加其他抗氧化劑之 CuSO4 誘發 LDL 氧化 組別做為控制組,以下列算式求的抑制百分比為橫軸,抗氧化劑濃度為 縱軸做出抑制曲線,求綠原酸或 probucol 抑制 50 %之 LDL 氧化時,所 需莫耳濃度即為 IC50。. - 35 -.
(50) C 計算: 抑制 LDL 氧化(%)=(控制組吸光值–實驗組吸光值)X 100 % 控制組吸光值. < 三 > 體外試驗-3:總抗氧化能力測試(Trolox equivalent antioxidant capacity , TEAC assay). A 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) B 試劑炮製: Peroxidase 由原來濃度 1000 unit/mg, 用二次水稀釋成 44 unit/mg;由 30% H2O2 原液取 57 μL 加二次水泡成 1L 的 500 μM H2O2;取 8.23. mg. ABTS 用 95 %酒精泡成 20 mL 的 750 μM ABTS 溶液。 C 步驟: 將 0.5 mL 之 44 unit/mg peroxidase、0.5 mL 之 500 μM H2O2 以及 0.5 mL 之 750 μM ABTS(2,2'-azino-bis(3-ethylbenzothiazoline -6-sulfonic acid))與 3 mL 的二次水混和均勻,於室溫反應 6 分鐘後生成穩定之藍 綠色的陽離子自由基 ABTS•+,再分別加入 0.5 mL 不同濃度的 trolox 及 chlorogenic acid 與之反應 6 分鐘,在波長 734 nm 下測量吸光值,利. - 36 -.
(51) 用 trolox 清除 ABTS•+陽離子自由基的能力來定義 chlorogenic acid 的 TEAC。利用相同方式,測定 probucol 的抗氧化力。TEAC 值愈大表抗 氧化能力愈佳。 D 計算: (1)利用 trolox 檢量線求得樣品的濃度 (2)TEAC 值=. 樣品的吸光值 0.028. 註. X 樣品的濃度. 註:0.028 是迴歸係數. < 四 > 飼料製備. A 藥品: Cholesterol(Sigma, MO, USA) Probucol(Sigma, MO, USA) Chlorogenic acid(Sigma, MO, USA) 大豆油(泰山,Taichung,Taiwan) 兔飼料(福壽牌,Taichung,Taiwan) B 步驟: 秤取十天份的 cholesterol, probucol, chlorogenic acid 與一般兔飼料. - 37 -.
(52) (rabbit chow) 。先將冷凍的飼料熱炒,逼出飼料內的水氣,使飼料較容 易吸附油脂,倒入秤好的 cholesterol、probucol 或 chlorogenic acid,接著 加入十天份的大豆油以低溫拌炒均勻,注意火候不要太大以免燒焦。炒 好放入夾鏈袋,標明製作日期、天份、飲食種類、實驗室負責人,儲存 於-20 ℃冰箱。. < 五 > 抽血. A 器材: 23 1/2 G 蝴蝶針(華杏,Meditop Corporation, Malaysia) 棉花(藤旺,彰化,台灣) 10 mL 針筒(特浦,高雄,台灣) 15 mL 離心管(Sastedt, Germany) 兔架 B 步驟: 抽血實驗需兩人以上方可進行。將兔子從籠中抱出,先進行安撫使 其情緒穩定,將其抱入兔架固定,使其身體無法動彈。一人用雙手固定 兔子的頭部,避免其因疼痛而亂動導致更嚴重傷害,且在固定頭部的同 時,注意兔子的呼吸,以及眼睛瞳孔狀況。若有眼睛翻白、瞳孔縮小的. - 38 -.
(53) 情形,則需趕快鬆開兔架使兔子回覆呼吸。先噴酒精且按摩兔子耳朵直 到血管清楚浮現,再將蝴蝶針整支插入血管內,以免因為兔子掙扎而針 脫落,整支插入後開始進行抽血。取出全血先冰於 4 ℃,待進一步處理。. < 六 > 分離血清(Isolation of serum). A 儀器: 往復式震盪恆溫水浴機(Reciprocal shaking baths, TKS, Taiwan ) 離心機(Himac CR21, HITACHI, Japan) 氮氣(吉源行, Taichung, Taiwan) B 步驟: 血清分離過程全程避光。從冰桶中取出兔子全血,立即放入 37℃水 浴槽,水浴 1.5 小時,取出水浴槽中的樣本放入 4 ℃冰箱中 1 小時,以 利血液凝結、血球沉澱;再反覆以 4 ℃、4000g 離心 15 分鐘,抽取上 清液。上清液即為血清,血清充氮氣並保存於 4 ℃或-20 ℃。. < 七 > 組織切片染色(Histochemistry). A 器材:. - 39 -.
(54) 包埋盒 10 % 福馬林 B 藥品泡製: 10 % 福馬林:取甲醛原液(原液濃度:37 %) 270.3 mL 加入 DDW 至 1 L。調配成 10 %福馬林溶液放置室溫即可使用。 C 步驟: 犧牲時,將血管放入 PBS 中稍微清洗,縱切下約 3~5 mm 血管放入 包埋盒中,將包埋盒完全浸置於 10 %的福馬林,固定組織。接著進行組 織脫水,用石蠟浸潤並包埋,組織內的水分會被石蠟所取代,組織經過 石蠟包埋成臘塊,有利於保存;接著用切片機切成約 3-5 μm 的石蠟切 片,並且用蘇木紫-伊紅(hematoxylin-eosin, HE)染色。蘇木紫可將細 胞核染為藍紫色,伊紅則可將細胞質染為淡粉紅色。利用 alphaimage 2200 軟體計算內膜增厚的面積,將內膜增厚(含血管管腔)之面積減去血 管管腔面積,即得到血管內膜增厚的面積。相較於控制組的內膜增厚百 分比計算方式,如下: 內膜增厚(%)=. 各組內膜增厚面積 控制組內膜增厚面積. - 40 -. X 100 %.
(55) < 八 > 血清總膽固醇測試(Total cholesterol level of serum). A 試劑: SMT calibrator(Merck, Germany) cholesterol reagent(Merck, Germany) LDL precipitant reagent(Merck, Germany) B 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) C 原理: 利用膽固醇酯解酶水解膽固醇酯,產生膽固醇和脂肪酸,再以膽固 醇氧化酶氧化膽固醇,進而產生 H2O2 和 cholestrol-3-one,最後利用 peroxidase 將 H2O2 和 4-aminoantipyrine 及 phenol 作用,產生 H2O 和紅 色的 quinonimine 呈色物質作為定量的依據。 D 步驟: 所有的試劑保存於 4 ℃皆可直接使用。取 10 μL 的血清加入 1000 μL 膽固醇試劑,取 10 μL 的二次水和標準液作為 blank 和 standard,混合均 勻後室溫靜置 10 分鐘,於波長 500 nm 測吸光值。. - 41 -.
(56) E 計算: cholesterol(mmol/L)=(樣本吸光值–blank 吸光值) x 6.03 Standard 吸光值 註:6.03 為 standard 的濃度(mmol/L). < 九 > 血清低密度脂蛋白測試(LDL-cholesterol level of serum). A 試劑: SMT calibrator(Merck, Germany) cholesterol reagent(Merck, Germany) LDL precipitant reagent(Merck, Germany) B 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) C 原理: 樣品加入 heparin 將大部分的 LDL 沉澱,離心之後 HDL 和 VLDL 仍可留在上清液,接著測量上清液殘餘的膽固醇,則總膽固醇減去上清 液膽固醇,就等於此樣品的 LDL-C 濃度。 D 步驟: (1) 沉澱法:. - 42 -.
(57) 取 100 μL 的血清加入 1000 μL 的 LDL 沉殿劑,混合均勻後室溫靜 置 15 分鐘,接著以 2500 g 離心 20 分鐘。在一小時內,抽取上清液進 行以下實驗。 (2) 酵素法: 取 100 μL 的上清液加入 1000 μL 膽固醇試劑,亦取 100 μL 的標準 液加入 1000 μL 膽固醇試劑做為 standard。混合均勻後室溫靜置 10 分 鐘,於 45 分鐘之內在波長 500 nm 測吸光值。 E 計算: (1) 計算上清液中的膽固醇 cholesterol(mmol/L)=(樣本吸光值–blank 吸光值)x 6.03. 註. Standard 吸光值 註:6.03 為 standard 的濃度(mmol/L) (2) 計算樣本 LDL-cholesterol LDL-cholesterol(mmol/L)=總膽固醇–上清液中的膽固醇. < 十> 血清高密度脂蛋白測試(HDL-cholesterol level of serum). A 試劑: SMT calibrator(Merck, Germany). - 43 -.
(58) cholesterol reagent(Merck, Germany) HDL precipitant reagent(Merck, Germany) B 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) C 原理: 利用 phosphotungstic acid 和鎂離子將乳糜微粒、VLDL 和 LDL 沉 澱。離心之後上清液僅存 HDL-C(high density lipoprotein- cholesterol, HDL-C),膽固醇含量同樣可用方法< 八 >測得。 D 步驟: (1) 沉澱法: 取 200 μL 的血清加入 500 μL 的 HDL 沉殿劑,混合均勻後室溫靜置 15 分鐘,接著以 2500 g 離心 20 分鐘。在二小時內,抽取上清液進行以 下實驗。 (2) 酵素法: 取 100 μL 的上清液加入 1000 μL 膽固醇試劑,亦取 100 μL 的標準 液加入 1000 μL 膽固醇試劑做為 standard。混合均勻後室溫靜置 10 分 鐘,於波長 500 nm 測吸光值。. - 44 -.
(59) E 計算:. HDL-cholesterol(mmol/L)=(樣本吸光值–blank 吸光值)x 6.03. 註. Standard 吸光值 註:6.03 為 standard 的濃度(mmol/L). < 十一 > 血清三酸甘油脂測試(Triglyceride level of serum). A 試劑: SMT calibrator(Merck, Germany) Triglyceride reagent(Merck, Germany) B 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) C 原理: 利用脂蛋白脂解酶(LPL)會分解三酸甘油酯的特性,而為 quinoneimine 為指標辨識物,酵素反應公式如下: (1)triglyceride (2)glycerol + ATP. LPL. glycerol + fatty acid GK. (3)glycerol-3-phosphate + O2. - 45 -. glycerol-3-phosphate + ADP GOP.
(60) dihydroxyaceton phosphate + H2O2 (4)2 H2O2 + amynoantipyrine + 4-chlorophenol. Peroxidase. quinoneimine + HCl + 4 H2O D 步驟: 所有的試劑保存於 4 ℃ 皆可直接使用。取 10 μL 的血清加入 1000 μL 膽固醇試劑,取 10 μL 的二次水和標準液作為 blank 和 standard,混合 均勻後室溫靜置 10 分鐘,於波長 550 nm 測吸光值。 E 計算: Triglyceride(mmol/L)=(樣本吸光值–blank 吸光值)x 2.63 Standard 吸光值 註:2.63 為 standard 的濃度(mmol/L). < 十二 > 血清脂質過氧化物 MDA 之測試(TBARS of serum). A 試劑: MDA(Alderich, WI, USA) 95 % Ethanol(景明化工, Taichung, Taiwan) TBA(Sigma, MO, USA) TCA(Sigma, MO, USA). - 46 -.
(61) CuSO4(Sigma, MO, USA) B 儀器: 往復式震盪恆溫水浴機(Reciprocal shaking baths, TKS, Taiwan) ELISA plate reader(μ QUANT, Bio-Tek, USA) 離心機(5415R centrifuge, Eppendorf, Hamburg, Germany) C 原理: 丙二醛(malondialdehyde, MDA)是脂質過氧化作用的終產物,而 TBA (thiobarbituric acid)會和 MDA 結合產生粉紅色的物質,此物質 在波長 532 nm 時有最大的吸光值。所以將血清用 CuSO4 誘發血清過氧 化,而 TCA(trichloroacetic acid)能和蛋白質結合而沉澱,使蛋白質不 至於干擾吸光值的測定。 D 步驟: 血清 25 μL 加入 60 μM 的 CuSO4 2.5 μL 促氧化,且加入 22.5 μL 的 PBS 混合均勻。Blank 的部份則用 95 % Ethanol 代替 sample;用標準品 MDA (1 g/mL)稀釋成所需要的濃度,做出檢量線,將配置好的檢量 線濃度、Blank 和 sample 抽取 50 μL ,依序加入 0.67 % TBA、20 % TCA 各 350 μL 混合均勻後,放入 67 ℃水浴槽水浴 30 分鐘,接著以 10000 rpm 離心 2 分鐘,小心吸取上清液 200 μL 放入 96 well,利用 ELISA plate reader. - 47 -.
(62) 測量其在波長 532 nm 的吸光值。做出檢量線,用檢量線推出 sample 中 MDA 的濃度,以 nmol/mL 表示。. < 十三 > 分離 LDL(Isolation of LDL). A 試劑: NaCl(Sigma, MO, USA) KBr(SHOWA, Tokyo, Japan) B 藥劑泡製: 0.9 % NaCl:秤取 0.9 g 的 NaCl 泡成 100 mL,並滅菌使用。 C 儀器: Vortex genie 2(BOHEMIA, N.Y., USA) 超高速離心機(Himac CS100FX, HITACHI, Japan) D 步驟: 整個實驗流程分為三天。第一天將 2 mL 血清置入 3 PC 管,加入 0.7 mL 的 0.9 % NaCl 混合均勻,放入超高速離心機,在 10 ℃以 100,000 rpm (Acc.:5,Dec.:7) 離心 3.5 小時,取出 3 PC 管,棄置最上層 0.7 mL 的上清液且充氮氣,保存於-20 ℃冰箱。第二天,從冰箱取出 3 PC 管, 在室溫下解凍或用手緊握加速其溶解,再重複第一天的所有步驟,同樣. - 48 -.
(63) 保存於-20 ℃冰箱。第三天,取出 3 PC 管,在室溫下解凍後,於各管加 入 166.8 mg 的 KBr 調整液體的濃度梯度,用 Vortex genie 2 將 KBr 混合 至完全溶解,放入超高速離心機,在 10 ℃以 100,000 rpm (Acc.:9, Dec.:7)離心 3.5 小時,小心吸取最上層 0.6 mL 的上清液,充氮氣保 存於 4 ℃冰箱。全程需避光進行。. < 十四 > 蛋白質定量(protein assay). A 試劑: Bradford reagent(BioRad, CA, USA) BSA(Bovine serum albumin)(Sigma, MO, USA) B 儀器: Vortex genie 2(BOHEMIA, N.Y., USA) ELISA plate reader(μ QUANT, Bio-Tek, USA) C 步驟: 以二次水稀釋 0.1 mg/mL 的 BSA 作出不同濃度的檢量線,取 10 μL 的樣品加 790 μL 二次水,接著加入 200 μL 的 Bradford reagent 混合均 勻,室溫靜置 10 分鐘後,每管抽 200 μL 放入 96 well,接著利用 ELISA plate reader 測量 590 nm 的吸光值。做出檢量線,用檢量線回推 sample. - 49 -.
(64) 中蛋白質的濃度,以 mg/mL 表示。. < 十五 > LDL 之脂質過氧化遲滯期測試(Lag time of LDL). A 試劑: CuSO4(Sigma, MO, USA) KCl(SHOWA, Tokyo, Japan) KH2PO4(SHOWA, Tokyo, Japan) Na2HPO4(SHOWA, Tokyo, Japan) B 藥劑泡製: PBS ( Phosphate buffer saline):秤取 9.35 g 的 NaCl、0.2 g 的 KH2PO4、0.38 g 的 Na2HPO4∙7 H2O,加入二次水泡成 1 L,混合均勻後, 調整其 pH 為 7.4,可依情況滅菌使用。 C 儀器: 往復式震盪恆溫水浴機(Reciprocal shaking baths, TKS, Taiwan) Micro centriphuge II(The Griffin Group , USA) 紫外光分光光譜儀(DU-640 spectrophotometer, Beckman Coulter, USA) 紫外光分光光譜儀(DU-800 spectrophotometer, Beckman Coulter,. - 50 -.
(65) USA) D 步驟: 蛋白質定量 LDL 的濃度,限定最終 LDL 濃度為 50 μg/mL, CuSO4 最終濃度為 5 μM,最後用 PBS 來補剩餘的體積至 1 mL,混合均勻後, 放入 37 ℃的水浴 10 分鐘。取出後,將所有的液體利用 Micro centriphuge 將所有液體 spin down,接著將所有液體移入石英管,放入分光光譜儀 中,以 37 ℃測量 234 nm 的吸光值,每 15 分鐘為間隔測量一次,共 1200 分鐘。. < 十六 > 組織均質(Homogenize). A 藥劑: Tris-Sucross(Sigma, MO, USA) B 藥劑泡製 Tris-Sucross buffer:秤取 3.152 g 的 Tris-HCl、85.575 g 的 Sucross、 0.3722 g 的 EDTA,加入二次水泡製成 1 L,準備使用。 C 儀器: 微量天秤(CP224S, Sartorius, Germany) T25 Basic 刀片式高速均質乳化機(T25 Basic, IKA , Germany). - 51 -.
(66) 離心機(5415R centrifuge, Eppendorf, Hamburg, Germany) D 步驟: 刀片式均質探頭、夾子、剪刀…等所有器械,皆須滅菌過後才能使 用。從-80 ℃取出樣品,秤取 100 mg 兔子主動脈,剪碎後加入 1.1 mL Tris-HCl buffer,接著放於冰上以均質機高速(20000 rpm/min)均質磨 碎,均質 3 分鐘後,吸取均質液至容量 1 mL 的 eppendorf 中,接著以 4000 g 在 4 ℃離心半個小時,小心抽取上清液,立刻進行下一實驗步驟 或充氮氣隔天始用。. < 十七 > 組織自由基測試(Luminol-CL levels of thoracic aorta). A 試劑: Luminol(Sigma, MO, USA) TBHP(turt- Butyl hydroperoxide, Aldrich, USA) B 藥品泡製: Luminol:秤取 0.0025 g 的 Luminol 加入 500 mL 的二次水,包上鋁 泊紙避光儲存於 4 ℃冰箱。 C 儀器: 超微弱化學發光儀 CLA-110 chemiluminescence(Chemiluminescence. - 52 -.
(67) TOHOKU, Kawasaki, Japan) D 步驟: 取以 Tris-HCl buffer 均質的上清液 400 μL 加入 200 μL luminol,上 下倒置混合均勻,避光靜置於室溫,培養 15 分鐘,放入超微弱化學發 光儀設定 37 ℃、17 分鐘;實驗開始後,到 100 秒時,加入 100 μL TBHP 盡量混合均勻,測量 Total CL count。以蛋白質定量樣本中蛋白質的濃 度,將 Total CL count 除以蛋白質的濃度,以 CL count /mg protein 來表 示。整個實驗需全程避光。. < 十八 > 分離兔子主動脈 total RNA(Isolation thoracic aorta of total RNA ). A 試劑: Trizol reagent(Invitrogen, CA, USA) Chloroform(BDH, Poole, England) Isopropanol(Sigma, MO, USA) DEPC(Sigma, MO, USA) 95% Ethanol(景明化工, Taichung, Taiwan) B 藥劑泡製:. - 53 -.
(68) DEPC-treat water:加入 100 μL 的 DEPC 原液至 1 L 的二次水 (DDW),調製成 0.01 %(v/v)的濃度,並隔夜滅菌後使用。 C 儀器: 微量天秤(CP224S, Sartorius, Germany) T25 Basic 刀片式高速均質乳化機(T25 Basic, IKA , Germany) 離心機(5415R centrifuge, Eppendorf, Hamburg, Germany) D 步驟: 刀片式均質探頭、夾子、剪刀…等所有器械,皆須滅菌過後才能使 用,或用 DEPC 水浸泡隔夜,以防止 RNA 分解。刀片式探頭裝上均質 機後,須以依序以酒精、二次水,重複洗兩遍以上,最後再以二次水沖 洗、擦乾備用。 從-80 ℃取出樣品,秤取 100 mg 兔子主動脈,剪碎後加入 1.5 mL Trizol,接著放於冰上以均質機高速(20000 rpm/min)均質磨碎,以維 持低溫狀態,均質 3 分鐘後,吸取均質液至 2 mL eppendorf 中,並靜置 於冰上 5 分鐘;接著,各管加入 0.4 mL 氯仿,手搖晃混合 15 秒後,靜 置於冰上 10 分鐘,再用 12000 g 在 4 ℃離心 15 分鐘,小心吸取 800 μL 澄清透明的上清液至另一 2 mL 的 eppendorf 中;加入 950 μL isopropanol 上下倒置混合均勻,靜置於冰上 10 分鐘,再以 12000 g 在 4 ℃離心 10. - 54 -.
(69) 分鐘;倒掉上清液留下底部白色的沉澱物,加入 1.5 mL 75 % DEPC 酒 精使白色沉澱物溶解,放置 4 ℃冰箱隔夜。利用 7500 g 在 4 ℃離心 5 分鐘,倒掉上清液留下沉澱物,使 75 % DEPC 酒精盡量蒸發,再加入 10~20 μL DEPC 水將沉澱物溶解,保存於-80 ℃冰箱。. < 十九 > Total RNA 定量(Quantification of total RNA). A 試劑: DEPC(Sigma, MO, USA) B 藥劑泡製: DEPC-treat water:加入 100 μL 的 DEPC 原液至 1L 的二次水 (DDW),調製成 0.01 % (v/v)的濃度,並隔夜滅菌後使用。 C 儀器: 分光光度計(HITACHI U-2000 spectrophotometer, HITACHI, Japan) D 步驟: 分離完 Total RNA,立即取 1 μL 分離出來的 Total RNA 加入 999 μL 混合均勻,並以 1000 μL DEPC 水做為 Blank,測量在 260 nm、280 nm 的吸光值。核酸在波長 260 nm 有最大吸收波,所以 OD 260 可間接用來 偵測核酸的量。蛋白質在波長 280 nm 有最大吸收波,所以 OD 280 可間. - 55 -.
(70) 接用來偵測蛋白質的量。 OD 260/OD 280 為 Total RNA 的純度,比值 要 1.8~2.0 以上較純。各管 OD 260 先扣掉 Blank 的吸光值後,Total RNA 濃度的算法如下: (RNA 濃度:1 OD 吸光值 = 40 μg/mL) Net total RNA 濃度(μg/mL)= OD 260 吸光值 x 40. < 二十 > 反轉錄(Reverse transcription). A 試劑: Reverse Transcription kit(Invitrogen, CA, USA) B 儀器: Thermal cycler PCR machine(iCycler, BioRad, USA) Micro centriphuge II(The Griffin Group , USA) C 步驟: 設定 total RNA 為 4.2 μg/管,則取適量的 sample RNA,用滅菌過的 二次水來補足不足的體積,加入 Oligo(dT) 、10 mM dNTP,用 pipement 抽吸均勻,放入 PCR machine 在 65 ℃下反應 5 分鐘,冷卻於 4 ℃;接 著加入 10X RT buffer、25 mM MgCl2 、0.1 M DTT、RNase OUTTM Recombinant RNase Inhibitor 混合均勻後,放入 PCR machine 在 42 ℃ 下 反應 2 分鐘,冷卻於 4 ℃;接著加入 SuperScriptTM Ⅱ RT 放入 PCR. - 56 -.
(71) machine 在 42 ℃下反應 50 分鐘,在 70 ℃下反應 15 分鐘,冷卻於 4 ℃; 接著加入 RNase H 放入 PCR machine 在 37 ℃下反應 20 分鐘,冷卻於 4 ℃。反轉錄後的產物,放在-20 ℃冰箱保存。. < 二十一 > 即時定量聚合酶鏈反應(Real-time PCR). A 試劑: SYBR Green supermix(BioRad, CA, USA) VCAM-1 Primer(Invitrogen, Taichung, Taiwan) MCP-1 Primer(Blossom,Taichung, Taiwan) TNF-α Primer(GeneMark,Bio Basic, Taichung, Taiwan) TNF-β Primer(Genemed synthesis, South San Francisco,CA, USA) β-actin Primer(Genemed synthesis, South San Francisco,CA, USA) B 儀器: Thermal cycler PCR machine(iCycler, BioRad, USA) Real-time machine(iCycler, BioRad, USA) Micro centriphuge II(The Griffin Group , USA) C 步驟: 200 μL 的 thin-wall PCR tube、tips 和二次水皆需經過滅菌。操作台. - 57 -.
(72) 先噴酒精,以去除其他核酸或序列不必要的干擾,並且帶手套操作一切 的實驗。 從-20 ℃冰箱取出 cDNA、各種基因的 sense 和 anti-sense primer 在 室溫下解凍,而 SYBR Green supermix 則需要避光解凍。SYBR Green supermix 含有特殊螢光染劑可以和雙股螺旋結合(600bp 以下適用)。 所有實驗盡量在維持在 4 ℃。除了 cDNA 外,將所需的所有試劑含:sense 和 anti-sense primer、滅菌過的二次水和 SYBR Green supermix,用 pipement 抽 吸 均 勻 , 因 為 是 非 常 微 量 的 實 驗 , 所 以 必 須 用 micro centriphuge 將殘留於管壁的試劑都離下來,不可有液體或氣泡殘留於管 壁。接著取適量加入 200 μL 的 thin-wall PCR tube,再加入 cDNA 同樣 用 pipement 抽吸均勻,接著用 micro centriphuge 將殘留於管壁的試劑都 離心下來後放入 thermal cycler. 設定溫度、時間和 cycle 數,先用 95 ℃, hotstart 3 分鐘,以 95 ℃進行 denature,使雙股 DNA 模版分開,接著不 同的基因有不同的溫度條件如下: VCAM-1:(50X)60.1 ℃,1 分鐘;72.0 ℃,50 秒 MCP-1: (40X)60.1 ℃,1 分鐘;72.0 ℃,50 秒 TNF-α:(40X)60.1 ℃,1 分鐘;72.0 ℃,50 秒 TNF-β:(40X)63 ℃,1 分鐘;72.0 ℃,50 秒. - 58 -.
(73) 跑完以上的步驟後,收集此時的吸光值,當雙股螺旋 DNA 放出的螢光 超過儀器設定偵測閥值的 cycle 數,稱為 Ct 值(threshold cycle)。Ct 值 越大表示此基因的 mRNA 表現較弱;反之,Ct 值越小表示此基因的 mRNA 表現較強;接著再經過 95 ℃,1 分鐘的過程;接著,以 55 ℃, 1 分鐘後,跑 melting curve 並收集此時的吸光值。 跑 melting curve 的方式如下:從 55 ℃開始,以每一個 cycle 增加 0.5 ℃的方式增加至 95 ℃。melting curvet 除了可以用來判斷 PCR 的產 物是否具有專一性,且可判斷整個實驗過程沒有污染物產生,也沒有 primer dimer 的產生,因為產物不專一則最大波峰不只一個,會產生高 低起伏如鋸齒狀的曲線,且波峰與波谷差異大。當所有的 melting curve 之最大波峰的尖端都指向同一溫度時,通常大於 80 ℃以上,且波峰的 peak 較尖、波峰橫跨距離較小時,代表此實驗結果沒有污染,且沒有 primer dimer 的產生;所以我們可以用 Ct 值來算出 target gene mRNA 的 表現量。本實驗以 β-actin 作為 housekeeping gene。mRNA 的表現量以 △Ct 來表示,Ct 值計算法如下: D 計算方式: △Ct = target gene Ct–housekeeping gene Ct. - 59 -.
(74) 表 3-1 即時定量聚合酶鏈鎖反應引子序列表. Name. Sequence. Length. Vascular cell adhesion. (F)GAACACTCTTACCTGTGCACAGC. 567. molecule-1(VCAM-1). (R)CCATCCTCATAGCAATTAAGGTGAG. Monocyte chemoattractant (F)GTCTCTGCAACGCTTCTGTGCCTG protein-1(MCP-1). (R)CAATGAAGTAGTAGTAGAGGGTGT. Tumor necrosis factor -α. (F)CCTCTCATCCTCCTCTAGCT. (TNF-α). (R)TGCAGGCCTACACCACCAT. Tumor necrosis factor-β. (F)CTCCGCCACGGCTTCTC. (TNF-β). (R)AACCACCTGCGAGTACACGAA. β-actin. (F)GCCGGGACCTGACAGACTAC. 369. 150. 81. 74. (R)CCTCTCAGCTGTGGTGGTGAA. 註:簡寫(F)表示 Forward sequence;(R)表示 Reverse sequence. - 60 -.
(75) < 二十二 > 統計與分析. 所有數據以平均值+標準誤(means + S.E.M)來表示,control 組與 normal 組兩組間差異以 one-way ANOVA 來分析,使用統計軟體 SAS 9.1,多項變異數分析中的鄧肯氏最小顯著差異法(Duncan’s multiple range test)來判別各組間之差異,在 p < 0.05 時具有顯著差異性。. - 61 -.
(76) 第五章. 第一節. 結果. Probucol 和綠原酸的抗氧化能力. 在 probucol 和綠原酸進行抗氧化力的體外試驗中,由表 4-1 可觀察 到 probucol 於 12.52±0.46 μM 即可抑制 50 %的 LDL 氧化作用,且綠原 酸於更低的劑量(7.95±0.19 μM) ,亦可達到同樣的效果。在抑制 DPPH 自由基的體外試驗中,也發現: probucol 於 1.45±0.05 mM 可以抑制 DPPH 自由基的產生,而綠原酸於 0.85±0.02 mM 時,也達到同樣的抑制 效果。接著,再以 TEAC 方法,將 probucol 和綠原酸和脂溶性的維生 素 E—trolox 的抗氧化力相比較,發現 probucol 的總抗氧化能力是 2.38± 0.54,綠原酸總抗氧化能力是 1.72±0.19。綜合以上三個實驗,顯示: probucol 和綠原酸在體外試驗中具有抗氧化力。. 第二節 實驗期間各組生長狀況的觀察. 實驗期間,我們給予每天 100 g 的飼料,並紀錄兔子每天的攝食情 形,且從零週開始測量每週的體重變化。由表 4-2 所示,進入實驗的兔 子體重大約為 2.2~2.4 公斤,實驗八週結束後的體重約為 2.8~3.2 公斤, 經統計之後並無顯著差異性;食物攝取量在各組間也沒有顯著的差異. - 62 -.
數據

+7




相關文件
Mannocarolose, β-glucan, Amino acid nitrogen, Crude protein, Crude ash and
the lymphocyte function-associated antigen 1, or LFA-1, was so named because antibodies recognizing this structure interfere with lymphocyte cell adhesion events and
附表 1-1:高低壓電力設備維護檢查表 附表 1-2:高低壓電力設備維護檢查表 附表 1-3:高低壓電力設備(1/13) 附表 2:發電機檢查紀錄表. 附表
No risk factor for necrosis could be identified except the realization of dental extraction before treat- ment initiation (P = .025) (Table 2): at 36 months, the incidence of
Higher immunoexpression of HIF-1 a, NOTCH1, ADAM-12, and heparin-binding epidermal growth factor like growth factor (HB-EGF) in epidermoid cells in compari- son with mucous cells
Orthokeratinized odontogenic cyst with an associated keratocystic odontogenic tumor component and ghost cell Table 1 Previous case reports of multiple orthokeratinized
The calcifying epithelial odontogenic tumor (CEOT), also known as Pindborg tumor, is a rare and typically benign odontogenic neoplasm [1].. Pin- dborg first described it as a
Histological sections showed tumor cells at a distance of 8 mm from the clinical and radiographic limit of the lesion, so the elimination of at least 1 cm of peripheral bone margin
