國
立
交
通
大
學
生物科技學系
碩
士
論
文
以螢光共振能量轉移揭示蛋白質 PGB1 分子內摺疊
機制
The Folding Mechanism of Protein G B1 Domain Revealed by
Fluorescence Resonance Energy Transfer
研 究 生〆鍾偉賢
指導教授〆張家靖 教授
以螢光共振能量揭示蛋白質 PGB1 分子內摺疊機制
The Folding Mechanism of Protein G B1 Domain Revealed by Fluorescence Resonance Energy Transfer
研 究 生〆鍾偉賢
Student〆Wei-Hsien Chung
指導教授〆張家靖
Advisor〆Chia-Ching Chang
國 立 交 通 大 學
生 物 科 技 學 系
碩 士 論 文
A ThesisSubmitted to Department of Biology Science and Technology College of Biology Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biology Science and Technology
June 2011
Hsinchu, Taiwan, Republic of China
以螢光共振能量轉移揭示蛋白質 PGB1 分子內摺疊機制
學生〆鍾偉賢
指導教授
〆張家靖
國立交通大學生物科技學系碩士班
摘要
蛋白質如何摺疊成特殊的結構,為目前學界不斷探討的重點。為了要能夠理解蛋白 質摺疊機制,研究單功能區蛋白質(single-domain protein)變成了一個很重要的指標。 本研究中,我們以 Protein G B1 Domain(PGB1)作為研究之標的蛋白質,利用本實驗 室所開發之蛋白質摺疊方法,Over-critical Folding Process,將 PGB1 由未摺疊態摺疊至 自然態並觀察摺疊中間的結構變化。PGB1 是一個小型蛋白質只有 56 個胺基酸,是 Protein G 的 IgG-binding domain。藉由觀察 PGB1 自身的 Trp43 螢光光譜和 acrylamide 焠熄光譜,可發現 PGB1 之摺疊是一個二狀態反應(two-state reaction)的摺疊過程々而 以動態光散射(dynamic light scattering)觀察分子大小的變化認為 PGB1 摺疊過程應具 有另一個中間態存在。藉由螢光共振能量轉移(Fluorescence Resonance Energy Transfer, FRET)分析各主要二級結構之間在摺疊過程中的變化,發現 PGB1 的 β-hairpin 2 與 α-helix 在摺疊初期就已互相靠近,但其 β-hairpin 1 顯然較前者晚形成々綜合以上結果顯示蛋白質 PGB1 的摺疊是一個多狀態反應(multi-state reaction)。本研究藉由 FRET 方法
The Folding Mechanism of Protein GB1by Fluorescence Resonance
Energy Transfer
Student: Wei-Hsien Chung
Adisor:Chia-Ching Chang
Department of Biology Science and Technology
National Chiao Tung University
Abstract
―How does a protein fold?‖ This has been questioned for long period. Studying on a single-domain protein can be a remark to understand the protein folding mechanism in general. Recently, we folded the Protein G B1 Domain (PGB1) by an over-critical folding process, which was developed by our lab, and studied the conformational changes in each folding state. PGB1 is a small protein with 56 residues, and is an IgG-binding domain of protein G. Because PGB1 contains the basic folding elements in a short sequence of amino acid without disulfide bonds, making it an excellent model for protein folding studies. The intrinsic Trp43 fluorescence of PGB1 and the acrylamide-quenching fluorescence showed that the PGB1 folding is a two-state reaction. However, the molecular diameter changes during the folding process indicated a folding intermediate exists in the PGB1 folding process. The FRET experiment, with Trp/IAEDANS as fluorophore pairs, showed that the β-hairpin 2 attaches to the α-helix long before the β-hairpin 1 formation. All the results together suggest the PGB1 folding process is a multi-state reaction. Overall, we were successful to reveal the folding process of PGB1 by FRET analysis.
謝誌
我不算是一個勤奮的好研究生,總是被周遭的事物給吸引了,老是被老師念說實驗 進度太慢了……即使我鑽研得不夠深,花的心思也不算多,但還是覺得很充實,讓我對 於分子層次的科學研究了解許多,每次都會對新技術的開發、新知識的挖掘感到嘆為觀 止。在實驗室度過不算長也不算短的兩年,我終於完成了這個研究題目,也熬到可以寫 謝誌的時候了! 這兩年有些苦悶的日子,要感謝和我共患難的一貣繼續在交大打拚的大學室友們, 朝彥、小光和 Dion,有他們在就會有樂子,因為我們幾個就是這麼的臭味相投,很多事 不用說出來,用幾個眼神就可以猜的到,因為我們幾個就是這麼的默契一百々而我也很 高興,能在離開交大登入國軍 online 之前,完成了我們幾個在大學一年級時候的約定, 整個感覺就是讚!祝福彼此繼續幸福快樂! 也更要感謝這兩年每天都會遇到的實驗室學長和夥伴們,互相幫忙扶持,互相加油 打氣,再辛苦也能撐過來。首先,我們有個老頑童鄭財木博士,他的待人處事值得我學 習,還有他的幽默的生活小故事總讓我笑破肚皮々李星原醫師和曾憲儀姊這對賢伉儷, 每天都在我們面前曬恩愛,好幸福喔!還有你們那些可愛的小孩,長大後一定和爸媽一 樣都是很棒的人!年紀不小了還很幼稚的朱學亮學長,我無法想像實驗室如果沒有你的 樣子,實驗室烤肉和火鍋一定是你揪的團,還有一貣度過的「輕鬆寫意」的日子,怎麼 會忘,以後有空一定會去找你喝喝茶。當然,還有和我一貣努力熬日子的同學娟姊、虹 瑋和昌瀚,一同分享生活、討論課業、分享樂子、尋找美食,以後還是會很懷念這些時 候有這些真心的朋友。 我的指導老師張家靖教授帶領我走過這兩年,一邊指導我們的課業也一邊關心我們 的生活,就是希望我們能夠在這兩年能過得充實也過得快樂舒適,還有機會讓我們能夠 出國開開眼界,這實在是個難忘的經驗。另外,更感謝老師很重視我的另一項興趣,平 面設計々因為如此,我的功力大大地進步了很多,人說要有壓力才會進步就是這個樣子, 因為我從沒想過我會在一個禮拜內就學會我本來根本就不會的網頁設計! 最後,謝謝永遠在背後支持的家人,你們是我驕傲的包袱,我要成為你們的榮耀! 還有一位大功臣,熊,妳總是能適時的督促我,有妳在我覺得我更勇敢,生活上很多的 事物和中心都和妳有關,我還要更努力更進步,還要一貣繼續為未來努力! 其實,要謝得還有很多很多,這時總會想貣陳之藩的那句話〆謝天吧!目錄
摘要 ... i Abstract ... ii 謝誌 ... iii 目錄 ... iv 表目錄 ... vii 圖目錄 ... viii 縮寫表 ... x 第一章 緒論 ... 1 1-1 研究動機與目的 ... 1 1-2 蛋白質結構 ... 1 1-3 蛋白質摺疊 ... 1 1-4 蛋白質摺疊方法 ... 2 1-5 蛋白質摺疊理論模型 ... 2 1-6 蛋白質 PGB1(Protein G B1 Domain) ... 4 1-7 螢光焠熄(Fluorescence Quenching) ... 6 1-8 四氯金酸(Chloroauric acid,HAuCl4) ... 71-9 螢光共振能量轉移(Fluorescence Resonance Energy Transfer) ... 8
第二章 實驗策略 ... 10
2-1 蛋白質策略 ... 10
2-2 實驗策略 ... 11
2-3 基因合成策略(含突變株) ... 12
第三章 實驗原理與步驟 ... 14
3-1 聚合酶連鎖反應 (Polymerase Chain Reaction,PCR) ... 14
3-2 IPTG 誘導蛋白質表現 (Protein Expression) ... 16
3-3 聚丙烯醯胺膠體電泳(Polyacrylamide Gel Electrophoresis,PAGE) ... 17
3-5 蛋白質摺疊 – 過臨界點摺疊法(Over-critical Folding Process) ... 18
3-6 等溫滴定微量熱法 (Isothermal Titration Calorimetry) ... 19
3-7 螢光光譜 (Fluorescence Spectrum) ... 20
3-8 螢光焠熄 (Fluorescence Quenching) ... 22
3-9 螢光共振能量轉移 (Förster Resonance Energy Transfer)... 24
3-10 圓二色光譜 (Circular Dichroism Spectrum) ... 25
3-11 動態光散射 (Dynamic Light Scattering) ... 27
第四章 實驗結果 ... 28 4-1 基因序列 ... 28 4-2 大量表現並純化 PGB1 蛋白質 ... 30 4-3 摺疊後 PGB1 功能測試 (ITC 實驗)... 31 4-4 蛋白質摺疊中間物之螢光光譜變化 ... 33 4-5 蛋白質摺疊中間物之圓二色光譜變化及二級結構比例分析 ... 35 4-6 蛋白質摺疊中間物之水合直徑分析結果 ... 38 4-7 螢光焠熄光譜變化 ... 39 4-8 螢光共振能量轉移受體接合結果 ... 44 4-9 螢光共振能量轉移之 Förster Distance (R0)變化 ... 46 4-10 螢光共振能量轉移螢光光譜 ... 48 4-11 HAuCl4吸收光譜及焠熄螢光光譜 ... 52 第五章 討論 ... 57 A. 摺疊後 PGB1 之功能測試 ... 57 B. 螢光光譜變化與同步輻射圓二色光譜... 57
C. Hydrophobic Collapse〆λmax變化與 Acrylamide 焠熄螢光光譜 ... 58
D. HAuCl4焠熄螢光光譜 ... 60
E. 蛋白質粒徑大小變化與 Trp43 包埋程度不同調... 60
G. 蛋白質 PGB1 摺疊為 multi-state reaction ... 61 H. 結語 ... 62 參考文獻 ... 63 附錄 ... 71 A. pET200-PGB1wt 基因定序圖譜 ... 71 B. pET200-PGB1G9C 基因定序圖譜 ... 72 C. pET200-PGB1G9C 基因定序圖譜 ... 73 D. 各使用之緩衝液之成分 ... 74 E. IAEDANS 接合方法 ... 76
表目錄
表一〆合成 PGB1 G9C 的 primer 組 ... 13
表二〆合成 PGB1 A23C 的 primer 組 ... 13
表三〆Over-critical Folding Process 各摺疊狀態緩衝液成分 ... 19
表四〆蛋白二級結構之圓二色光譜特徵峰 ... 27
表五〆各摺疊狀態的螢光波峰波長(λmax)。 ... 34
表六〆由 Acrylamide 焠熄 PGB1 wt 的 Trp43 螢光之結果總表 ... 43
表七〆PGB1 G9C 和 A23C 在各摺疊狀態下的 Trp 螢光量子產率以及 Förster Distance 47 表八〆PGB1 G9C-I 的光譜強度變化與螢光共振能量轉移效率 ... 49
表九〆PGB1 A23C-I 的光譜強度變化與螢光共振能量轉移效率。 ... 50
圖目錄
圖一〆蛋白質摺疊似一階態轉變示意圖 ... 3 圖二〆(a)蛋白質 PGB1 上負責與 Fc 區域鍵結的胺基酸々(b)PGB1 與 IgG 上的一個 Fc 區 域鍵結的結構圖(PDB〆1FCC) ... 4 圖三〆PGB1 以卡通(cartoon)模式表示的二級結構圖 ... 5 圖四〆acrylamide 結構式 ... 7 圖五〆HAuCl4結構式 ... 7 圖六〆IAEDANS 化學結構式 ... 9 圖七〆IAEDANS 固定在 Cys 上的化學反應 ... 9 圖 八〆蛋白質 PGB1 之胺基酸突變位置 ... 10 圖九〆解釋螢光發生機制的 Jablonski 圖[39] ... 20 圖十〆描述碰撞焠熄和螢光共振能量轉移的電子躍遷機制的 Jablonski 簡圖 ... 22 圖十一〆靜態焠熄分子機制簡圖。 ... 23 圖十二〆以 PGB1 fp 與 T7 rp 所挑選放大的 PGB1 wt 基因... 28 圖十三〆PGB1G9C 基因 ... 29 圖十四〆以 T7 fp 與 PGB1 rp 所挑選放大的 PGB1 A23C 基因 ... 29 圖十五〆蛋白質 PGB1(包括突變株)表現效果以及從上清液純化效果 ... 30圖十六〆Over-critical Refolding Process 摺疊後的 PGB1 wt 與 IgG 的 Isothermal Titration Calorimetrics 結果。(a)每次滴定作用的功率變化。(b)每次滴定作用的莫爾熱變化。32 圖十七〆10 M PGB1wt 在 over-critical refolding process 中不同的摺疊狀態的螢光光譜變 化,左上內插圖為各摺疊狀態的螢光波峰波長(λmax)。激發光波長為 295 nm。 ... 34 圖十八〆不同摺疊態下的 PGB1 的圓二色光譜 ... 35 圖十九〆PGB1 在 222 nm 波長的圓二色光譜變化 ... 36 圖二十〆PGB1 wt 在 M5狀態的同步輻射圓二色光譜 ... 36 圖二十一〆PGB1wt 與各突變株的同步輻射圓二色光譜 ... 37 圖二十二〆動態光散射之 GB1 wt 蛋白質分子水合直徑隨摺疊狀態變化圖 ... 38
圖二十三〆PGB1 wt 在 denatured 狀態下被 Acrylamide 焠熄螢光的 Stern-Volmer 關係圖, KSV=3.37×109 M-1s-1 ... 39
KSV=1.73×109 M-1s-1 ... 40 圖二十五〆PGB1 wt 在 M2 狀態下被 Acrylamide 焠熄螢光的 Stern-Volmer 關係圖, KSV=1.79×109 M-1s-1 ... 40 圖二十六〆PGB1 wt 在 M3 狀態下被 Acrylamide 焠熄螢光的 Stern-Volmer 關係圖, KSV=1.81×109 M-1s-1 ... 41 圖二十七〆PGB1 wt 在 M4 狀態下被 Acrylamide 焠熄螢光的 Stern-Volmer 關係圖, KSV=1.48×109 M-1s-1 ... 41 圖二十八〆PGB1 wt 在 M5 狀態下被 Acrylamide 焠熄螢光的 Stern-Volmer 關係圖, KSV=1.52×109 M-1s-1 ... 42 圖二十九〆Acrylamide 對各摺疊狀態的 PGB1 wt 的 kq變化圖 ... 42 圖三十〆IAEDANS(實線)和 PGB1(虛線)的吸收光譜 ... 44 圖三十一〆PGB1 G9C-I 吸收光譜... 45 圖三十二〆PGB1 A23C-I 吸收光譜 ... 45 圖三十三〆不同 pH 下的 IAEDANS 吸收光譜 ... 47 圖三十四〆PGB1 G9C-I 隨摺疊狀態改變的螢光光譜 ... 49 圖三十五〆PGB1 A23C-I 隨摺疊狀態改變的螢光光譜。 ... 50 圖三十六〆β-hairpin 1 – β-hairpin 2 (G9C)的距離變化 ... 51
圖三十七〆β-hairpin 2 – α-helix (A23C)的距離變化 ... 51
圖三十八〆HAuCl4 在不同 buffer 下的吸收光譜 ... 52 圖三十九〆在 Denatured 狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=4.66×103 M-1 53 圖四十〆在 M1狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=4.04×103 M-1 ... 53 圖四十一〆在 M2狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=2.78×103 M-1 ... 54 圖四十二〆在 M3狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=3.43×103 M-1 ... 54 圖四十三〆在 M4狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=6.49×103 M-1 ... 55 圖四十四〆在 M5狀態下,HAuCl4焠熄 PGB1 的螢光光譜,KSV=5.34×103 M-1 ... 55 圖四十五〆HAuCl4對各摺疊狀態的 PGB1 wt 的 kq變化圖 ... 56 圖四十六〆Acrylamide 對單 Trp 蛋白質的螢光焠熄能力(kq)與螢光波峰波長(λmax)之 關係圖 ... 59
縮寫表
Ala Alanine Met Methionine
Asn Asparagine Phe Phenylalanine
Cys Cysteine Pro Proline
Glu Glutamic acid Trp Tryptophan Gln Glutamate Tyr Tyrosine Gly Glycine
Lys Lysine
CD Circular Dichroism
FRET Förster/Fluorescence Resonance Energy Transfer
IAEDANS 5-[2-[(2-Iodo-1-oxoethyl)amino]ethylamino]-1-naphthalenesulfonic acid IgG Immunoglobulin G
ITC Isothermal Titration Calorimetry
kq Quenching Constant
λmax Emission Maximum
PGB1 Protein G B1 Domain
R0 Förster Distance
第一章 緒論
1-1 研究動機與目的 蛋白質是組成生物體的一個很重要的分子,是由一系列的胺基酸以不同比例 組成不同長度並各自擁有特定結構的巨分子。這些特定結構與其功能有很大的關 係,但是如何從一團混亂的狀態摺疊成特定的構型仍未明瞭々以最簡單的想法, 假若一個蛋白質有 100 個胺基酸,每個胺基酸之間具有兩個可以旋轉的鍵結,每 個鍵結擁有二到三個自由度,則這個蛋白質會有 4100~9100個可能的構型。但實際 上,蛋白質通常只有一個特定的構型,要排出如此多的可能構型勢必使蛋白質的 摺疊所需時間大大增加,但實際上蛋白質傴需毫秒甚至是微秒時間便可摺疊完畢, 因此人們對於蛋白質的摺疊充滿了好奇並積極地去了解其過程。故在此論文中, 我們利用螢光共振能量轉移這種技術直接地去觀察蛋白質內部在摺疊過程的變化, 去推敲蛋白質的摺疊過程。 1-2 蛋白質結構 蛋白質為多種胺基酸以不同比例所組成的生物巨分子 , 這些胺基酸的序列稱 為一級結構,二級結構主要是由主鏈中胺基之間的氫鍵形成的結構,亦包含局部 殘基(residues)之間的氫鍵々最常見的就是 α-helix、β-sheet 以及 turn。三級結 構則的是蛋白質的整體構型。四級結構則是多種蛋白質彼此以特定的方式形成的 複合體[1]。 1-3 蛋白質摺疊 蛋白質摺疊是一種物理現象指的是一條胺基酸序列鏈如何從一個在空間中隨 意排列的構型形成特定且具有功能的構型[1]。這些特定的構型通常為能量最低態, 當某蛋白質摺疊至其特有且具有功能的構型時,我們稱該構型狀態為自然態。摺 疊至自然態構型包含了溶劑的參與、鹽濃度的改變、溫度變化以及輔助分子等等。 一般認為蛋白質的胺基酸序列決定了其自然態構型[2]。1-4 蛋白質摺疊方法 直接稀釋法(Direct Dilution) 顧名思義是以單一步驟的溶液置換使蛋白質快速從高變性劑環境進入無變性 劑環境,從而使蛋白質摺疊,又稱作直接反應法(off-path)。以此方法必須將蛋 白質維持在非常低的濃度底下才能進行,蛋白質的摺疊效率隨著蛋白質濃度的增 加而下降[3]。
階段性熱平衡透析法(Stepwise Thermal Equilibrium Dialysis)
此方法是透過逐步緩慢的去除溶液中的變性劑,一般是利用透析法,由於變 性劑置換的過程十分緩慢,對蛋白質摺疊而言幾乎為靜止狀態,故又稱作準靜過 程(Quasi-static process)[4]。此法能夠大量地摺疊蛋白質至其自然態,避免造成 錯誤的摺疊或聚集[4-7]。 1-5 蛋白質摺疊理論模型 Framework Model 此理論由 Ptitsyn[8]所提出,蛋白質的摺疊是先形成二級結構,因此完成了整 個蛋白質結構的架構(framework)再進而形成三級結構。不穩定的二級結構區段 先形成再凝聚使整個胜肽鏈變的緊密,最後形成三級結構。
Hydrophobic Collapse Model
此理論認為蛋白質摺疊最主要的驅動力是由疏水性作用造成,親水環境驅使 疏水性胺基酸非特定的聚集。這項理論可以解釋為何蛋白摺疊速度如此快速。此 理論的實驗證明是蛋白質 barstar 的摺疊研究[9]。
Nucleation-condensation Growth Model
一個任意的蛋白質序列擁有的可能構型數量極為可觀,但其 摺疊 傴發生在一 個極短的時間內 變達到其自然態,故此理論利用蛋白質結構的 核化(nucleation) 來解釋如此快速的摺疊[10, 11]々如此一來,蛋白質所擁有的可能構型經過核化後
便能大量減少,因此加速了摺疊的速度。這些核化的區域是由彼此相鄰的胺基酸 殘跡組成的二級或三級結構,接著核加速了四周的結構成型,而這些周邊的結構 又使得核的結構更為穩定[12]。
The Diffusion-collision Model
1976 年由 Karplus 和 Weaver[13, 14]所提 出,認為蛋 白質是 許多 微 小區 間 ( micro-domain)的組合,在摺疊初期各個微小區間藉由擴散碰撞尋找到最穩定 的結構,接著再逐漸為調至自然態。
First-order-like Phase Transition
此理論是本實驗室於 2002 年提出[4],指出蛋白質在準靜態熱力學平衡的摺 疊模型與 Landau 的一階相變(first-order phase transition)相似,系統相圖(phase
diagram)中具有一個有限長度的 相變線(transition line),若此一相變線存在一次
微分不連續,則此稱為一階相變,通常此不連續出現在其端點,此點稱作特異點 (critical point)々此相變區隔出線兩旁的相區,此線上為兩相共存狀態,若繞過 此特異點,儘管相已改變,仍不會觀察到相轉變現象。而蛋白質摺疊穿過相轉變 區,則此蛋白質會發生沉澱或是玻璃態( glassy)的未摺疊態。故透過 stepwise thermal equilibrium dialysis 可以繞過此相轉 變區,使蛋白質正確的摺疊。
圖一〆蛋白質摺疊似一階態轉變示意圖
Φ(n1, n2, ..)表 示蛋 白 質摺 疊 狀態 (the order parameter ) 々而 n1、n2 代表 著 各項 會 影響 摺疊 狀 態
1-6 蛋白質 PGB1(Protein G B1 Domain) PGB1 的自然功能
Protein G 是一個多功能區蛋白質(mutlidomain protein),存在於 Group C 和
G Streptococcus 的細胞表面,一般認為該種細菌可以藉由此蛋白質與宿主的 免疫 球蛋白(Immunoglobulin G,IgG)結合而躲避宿主身體的防禦系統[15-19]々Protein G B1 IgG-Binding Domain (PGB1)是 Protein G 的一個功能區(domain)對 IgG 的 hFc 上的 CH2-CH3區域(即第二與第三個 Heavy Chain Constant Domain)有很
強的親和力[20, 21],無論是對於 monoconal 或 polyclonal IgG 都有很好的偵測或 分離特性[22, 23]々對 IgG 的 Fab 區域(the antigen-binding region)的親和力傴只 有對 Fc 區域的 10% [22, 23]。X-ray 結晶結構顯示一個 IgG 能與兩個 PGB1 鍵結 (圖二,PDB〆1FCC)[21]。 圖二〆(a)蛋白質 PGB1 上負責與 Fc 區域鍵結的胺基酸々(b)PGB1 與 IgG 上的一個 Fc 區域鍵結 的結構圖(PDB〆1FCC) CH3 CH2 PGB1 (a) (b)
PGB1 的蛋白質結構
蛋白質 PGB1 擁有 56 個胺基酸(包含 N 端的 Met),中間序列形成一個α-helix
(Ala23-Asn36)躺在由四個 β-strand (β1〆Met1-Leu9々β2〆Lys12-Ala20々β3〆 Glu42-Tyr45々β4〆Lys50-Thr55)組成的 β-sheet 之上々β1 和 β2 以及 β3 和 β4 分 別形成兩端的 β-hairpin (β-hairpin1〆Met1-Ala20々β-hairpin 2〆Glu42-Thr55)[24]。 由於不包含半胱胺(Cys)或脯胺酸(Pro)等會形成較複雜蛋白結構的胺基酸, PGB1 非常適合做為研究蛋白質摺疊研究。又由於 PGB1 只擁有 56 胺基酸,故常 被用作分子模擬的模型蛋白質。另外,將 近 95%的 GB1 胺基酸都參與了二級結構 的組成,並包覆了一個緊密的疏水核心,以及超過 70%骨幹上的 amide 和 carbonyl 都形成了氫鍵,因此擁有非常高熱穩定性[25], [26],無論是對熱或 urea 引貣的變 性都有很好的抵抗性[24],且可由熱變性中回復原有結構[26]。PGB1 的 β-hairpin 2 被發現能夠穩定地單獨存在於水溶液中[27],且此獨立出來的 β-hairpin 摺疊時 間約為 6 s [28]。 1 11 21 31 41 51 MTYKLILNGKTLKGETTTEAVDAATAEKVFKQYANDNGVDGEWTYDDATKTFTVTE -strand -helix 圖三〆PGB1 以卡通(cartoon)模式表示的二級結構圖
α-helix 以紅色表示,β-strand 或 β-sheet 則以藍色表示。(PDB〆1pga)
β1 β2
β4 β3
PGB1 的摺疊
PGB1 屬於小型 蛋白 質,因 此在 早期被 認 為此蛋 白質的 摺疊 會如 同其他 小型 蛋白質一樣遵循二狀態反應(two-state reaction)[24, 26, 29]。而此蛋白質的摺疊 是否為存在中間態(intermediate)目前還有些爭議[30-32]。但多數的研究結果都 指 出 蛋白 質 PGB1 的摺疊最初期存在一個與自然態類似的疏水核心。 氫氘交換 ( hydrogen-deuterium exchange ) 和 核 磁 共 振 光 譜 ( nuclear magnetic resonance spectrum)的研究指出 PGB1 的 β-hairpin 2 以及 α-helix 的中間區段與 N 端區段在 摺疊過程初期即不易進行氫氘交換且此區的胺基酸的動態(mobility)顯然較與其 他 區 域 來 的 慢 , 故 認 為 此 區 域 在 摺 疊 初 期 扮 演 了摺 疊 核 心 的 角 色 [33],
[34]。 以 continuous-flow fluorescence 測量 PGB1 上的 Trp43 螢光動態變化的研究結果推測 PGB1 的摺疊初期存在一個與自然態類似的疏水核心[35]。分子模擬也給予相同的 推論,S. Kmiecik 和 A. Kolinski [36]於 2008 年以 CA-CB side (CABS) chain model [37]與蒙地卡羅模擬(Monte Carlo simulation)得到 PGB1 的摺疊為多狀態反應 (multi-state reaction),摺疊由 α-helix 與 β-hairpin 之間的疏水胺基酸聚集形成摺 疊核心(Phe30、Trp43、Tyr45 和 Phe52),再由其他的疏水胺基酸繼續聚集形成 二級結構,最後達到自然態。另外,David Baker [38]等人於 2000 年針對 PGB1
的 β-sheet formation 的研究發現,在 β-hairpin1 與 α-helix 上的點突變對整體結構
影響最小,而在 β1、β3 和 β4 的點突變對整體影響較大,故認為可能會存在由三
個 β-strand 組成的 β-sheet 的中間態結構。
1-7 螢光焠熄(Fluorescence Quenching)
螢光焠熄(fluorescence quenching)一般泛指任何能造成螢光強度減弱的效應, 其中因分子交互作用造成的螢光焠熄包括了激發態反應(excited-state reaction)、 分子重排(molecular rearrangements)、能量轉移(energy transfer)、基態錯合物 (ground-state complex formation),以及碰撞焠熄(collisional quenching)[39]。 在此研究中,我們利用碰撞焠 熄 (又可稱作動態 焠 熄 dynamic quenching)對於 PGB1 內部的 Trp43 的螢光焠熄能力來比較在各個摺疊狀態下 Trp43 包埋於蛋白質 內部的程度。欲達到此目的,我們利用 acrylamide 這種中性的小分子焠熄劑來焠
熄 Trp43 的螢光。Acrylamide 能特別針對色胺酸的螢光進行焠熄的效應[40],此 分 子 很 小 且 為 中 性 能 夠 藉 由 擴 散 進 入 蛋 白 質 內 部 的 疏 水 核 心 [41, 42] 々 故 acrylamide 的螢光焠熄能力會受到擴散速率的影響,及越接近蛋白質內部的色胺 酸越不容易被焠熄,因此能夠利用此螢光焠熄能力的變化來了解色胺酸在包埋於 蛋白質內部的程度。 圖四〆acrylamide 結構式 1-8 四氯金酸(Chloroauric acid,HAuCl4) 化學式 HAuCl4,一般以 HAuCl4〃4H2O 存在,黃色針狀結晶,易溶入水。由 王水(濃鹽酸和濃硝酸按 3 比 1 混合制得)和純金反應製得。HAuCl4的腐蝕性能 極強,氧化性也極強,它能腐蝕一切金屬。但 HAuCl4 不穩定,加熱即可分解成
HCl,AuCl3和 AuCl。在水中以檸檬酸根還原 HAuCl4製得奈米金粒子[43, 44]。
HAuCl4 是一個平面分子,由中間一個金離 子外面四個 氯離子組成。HAuCl4 的吸
收光譜在 217 和 287 nm 各有一個特徵峰[45]。我們實驗室發現,HAuCl4可以焠
熄 Tryptophan 的螢光,但目前機制並不明瞭。
1-9 螢光共振能量轉移(Fluorescence Resonance Energy Transfer)
螢光共振能量轉移(fluorescence resonance energy transfer,FRET)是一種發 生在兩個很靠近的發光基團之間的能量轉移的機制々由德國科學家特奧多〄福斯 特(Theodor Föster)於 1946 年所發現,故此機制應正確地稱為福斯特共振能量 轉移(Föster Resonance Energy Transfer)[46],英文縮寫與此節標題同為 FRET々 Föster Resonance Energy Transfer 是較正確的英文名稱,但由於應用上大多是兩個 螢光發光基團所以許多研究偏好將 FRET 作 fluorescence resonance energy transfer 解釋。 當兩個擁有彼此重疊的 激發光譜與放射光譜的發光基團靠近到一定距離,其 中一個發光基團(供體)受外加的光能量激發後,通過偶極-偶極作用(dipole-dipole interaction)將供體激發態能量轉移到受體。這是一種非輻射能量躍遷的過程。產 生 FRET 現象後,供體螢光強度降低,而受體可以發射更強於本身的特徵螢光, 也可以不發螢光(螢光焠熄),同時也伴隨著螢光壽命的改變。另一個 FRET 最重 要的特點,FRET 只能發生在很短的距離,一般約在 10 nm 內,而且 FRET 的能 量轉移效率與發光基團之間的距離六次方有關,故 FRET 常被作為一種奈米等級 的光學「分子尺」。因此,常應用於生物巨分子之間的交互作用[47, 48](包括在 活細胞生理條件下[49, 50]),甚至是生物巨分子內部的結構變化[51, 52]。 在此研究中,我們以色胺酸(Trp)和 IAEDANS 作為螢光共振能量轉移的供 體和受體。色胺酸自然存在蛋白質 PGB1 上,且為該蛋白質唯一的色胺酸,位於
組成 β-hairpin 2 的 β-strand 3 上,簡稱為 Trp43(圖三)。IAEDANS [53]是一個螢
光發光基團,激發光波長約為 336 nm,放出的螢光波長約為 490 nm々其激發光 與色胺酸的螢光光譜(325~350 nm)有很好的重疊區域。IAEDANS 上的 iodoacetyl group 能與硫醇基(thiol)產生反應[54],使它的整個發光基團包括一個短碳鏈 轉 移到硫醇基上(圖七)。因此,我們將 PGB1 上的特定位置胺基酸突變為 Cys,再 將 IAEDANS 接合上去以觀察 PGB1 摺疊過程的內部距離變化。經由前面對蛋白 質 PGB1 的摺疊研究文獻的整理,我們知道 β-haripin 2 與 α-helix 在摺疊初期便形 成一個互相靠近的構型作為摺疊貣始的核心,而認為 β-haripin 1 應在摺疊過程中
是較慢形成的部分々所以我們 Cys 分別突變在 Gly9 和 Ala23 上的兩個位置(分別 為兩個突變蛋白質 PGB1 G9C 和 A23C),並將 FRET 受體接合在點突變上以觀察 α-helix、β-haripin 1 和 β-haripin 2 在摺疊過程中的變化。
圖六〆IAEDANS 化學結構式
圖七〆IAEDANS 固定在 Cys 上的化學反應
第二章 實驗策略
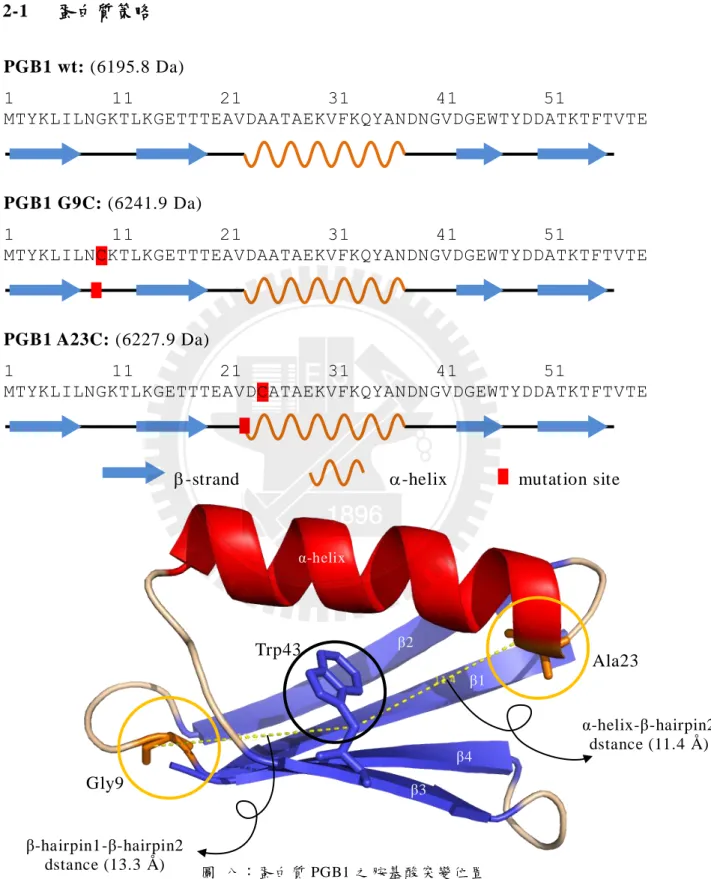
2-1 蛋白質策略 PGB1 wt: (6195.8 Da) 1 11 21 31 41 51 MTYKLILNGKTLKGETTTEAVDAATAEKVFKQYANDNGVDGEWTYDDATKTFTVTE PGB1 G9C: (6241.9 Da) 1 11 21 31 41 51 MTYKLILNCKTLKGETTTEAVDAATAEKVFKQYANDNGVDGEWTYDDATKTFTVTE PGB1 A23C: (6227.9 Da) 1 11 21 31 41 51 MTYKLILNGKTLKGETTTEAVDCATAEKVFKQYANDNGVDGEWTYDDATKTFTVTE-strand -helix mutation site
圖 八〆蛋白質 PGB1 之胺基酸突變位置
黑色圈者為 Trp43,將作為 FRET 的螢光 donor々橘色圈者為 Gly9 與 Ala23,將突變為 Cys 以便 接合 FRET 的螢光 acceptor。圖中之距離是以兩胺基酸的 Cβ為準。 Ala23 Gly9 Trp43 α-helix β2 β1 β4 β3 β-hairpin1-β-hairpin2 dstance (13.3 Å ) α-helix-β-hairpin2 dstance (11.4 Å )
2-2 實驗策略 以 acryamide 焠熄蛋白質螢光,求得 Trp43 的包埋程度 以動態光散射分析各摺疊中間態的 分子大小變化 觀察螢光的 λmax變化 利用螢光共振能量轉移直接觀察 PGB1 蛋白質內部的摺疊動態變化 以o v e r-c ri ti c a l fo ld in g p ro c e ss 摺 疊 各 蛋 白 質 螢 光 與 圓 二 色 光 譜 測 量 各 蛋 白 質 摺 疊 中 間 態,並觀察其穩定性和二級結構比例。 利用 ITC 確認摺疊後 PGB1 wt 的功 能是否與自然態相同 各突變點皆設計在非主要結構的 loop 上 以避免影響整體結構 G9C 用以觀察 β-hairpin1-β-hairpin2 距離 A23C 用以觀察 α-helix-β-hairpin2 距離
2-3 基因合成策略(含突變株)
由於該 DNA 序列,具有相當多的重複片段,為了避免重複的片段彼此產生 錯誤的黏合,所以將整個 PGB1 基因序列分成 1-85 和 78-174 兩段有互補的部分, 再各自直接合成長片段的 forward primer(fp)與 reverse primer(rp)〆1-36 fp, 33-85 rp 以及 78-137 fp,117-174 rp。其中各組的 fp 與 rp 有幾個鹼基能互補,再 用 klenow 補齊 3’端到 5’端的 overhanging 形成一完整的雙股 DNA。兩組的雙股 DNA 再以 Pst I 切出互補的 stick end 之後進行 ligation 形成完整的 PGB1 基因,最 後轉型到 pET200 載體並於 E. coli BL21(DE3)表現蛋白質。突變的 G9C 和 A23C 的基因序列也是直接合成再以相同手法組成完整基因。
表一〆合成 PGB1 G9C 的 primer 組
基因序列(5’ 3’) Tm (°C) 長度
1-36 (G9C)
forward primer CATATGACGTACAAGCTCATACTCAACTGCAAGACC 73.6 36
33-85 reverse primer TCTCTGCA^GTAGCACAATCAACAGCTTCAGTGGTC GTCTCCCCCTTCAGGGTC 74.6 53 78-137 forward primer C^TGCAGAGAAGGTCTTCAAGCAGTACGCTAACGAC AACGGGGTGGACGGGGAGTGGACG 77.0 58 117-174 reverse primer CTACTCCGTCACTGTGAAGGTCTTAGTCGCATCGTC GTACGTCCACTCCCCGTCCACC 77.0 58
框貣來處為突變處將被編譯為 Cys(原為 Gly)々粗體字表示為各組 primer 互補區々畫有底線處 為 Pst I 辨認區,「^」表為切位。
表二〆合成 PGB1 A23C 的 primer 組
基因序列(5’ 3’) Tm (°C) 長度
1-36
forward primer CATATGACGTACAAGCTCATACTCAACGGGAAGACC 73.6 36
33-85 (A23C) reverse primer TCTCTGCA^GTAGCTGCATCAACAGCTTCAGTGGTC GTCTCCCCCTTCAGGGTC 74.6 53 78-137 forward primer C^TGCAGAGAAGGTCTTCAAGCAGTACGCTAACGAC AACGGGGTGGACGGGGAGTGGACG 77.0 58 117-174 reverse primer CTACTCCGTCACTGTGAAGGTCTTAGTCGCATCGTC GTACGTCCACTCCCCGTCCACC 77.0 58
框貣來處為突變處將被編譯為 Cys(原為 Ala)々粗體字表示為各組 primer 互補區々畫有底線處 為 Pst I 辨認區,「^」表為切位 。
第三章 實驗原理與步驟
3-1 聚合酶連鎖反應 (Polymerase Chain Reaction,PCR)
聚合酶連鎖反應是一種分子生物技術,係在生物體外以 DNA 聚合酶(DNA polymerase)增加特定目標的 DNA 片段數量。此技術傴能夠將非常微量的 DNA 片段快速地複製增加至所需的數量,故被廣泛地應用於各種生物相關領域,例如 遺傳疾病檢查、物種鑑定、親子鑑定等等。PCR 技術是由美國科學家 Kary Mullis 於 1983 發明[55],並在七年後以此發明獲得諾貝爾化學獎。此技術發展至今已有
許多的變體,例如遞減聚合酶連鎖反應(Touchdown PCR)、反轉錄聚合酶連鎖反
應(Reverse Transcription PCR)、熱啟動聚合酶連鎖反應(Hot Start PCR)、即時 聚合酶連鎖反應(Real-time PCR)等等。
PCR 技術的基本原理是利用一種存在於水生嗜熱菌(Thermus aquaticus)的 DNA 聚合酶(Taq polymerase)在高溫環境下(~96°C)將高熱解旋成單股的 DNA 片段,藉由單股的 DNA 引子(primer)的標定而將特定的 DNA 片段複製增加[56]。 這基本原理可簡單地分成三個步驟〆( 1)變性反應(Denaturation),以高溫分離 DNA 的雙股形成單股的模板々(2)緩冷配對反應(Annealing),溫度降低至略低 於引子熔點溫度(指 50%的兩互補單股 DNA 相結合的最高溫度)而使引子與 DNA 模板配對々(3)引子延展反應(Elongation/Extension)〆引子由 5′端往 3′端延長形 成新的雙股 DNA。此三步驟為一循環,一般 PCR 反應由 20~30 個循環完成,當 完成 20 個循環,目標 DNA 將數量便達到原有的 2 的二十次方倍。 另外,引子的設計好壞會影響整個 PCR 反應的效果。一般 PCR 反應會由兩 段引子標定出欲複製增加的 DNA 片段,分別為前置引子(Forward primer)和反 置引子(Reverse primer)。引子係由人工合成的短片段 DNA,通常為 18~25 個 鹼基。引子太短便可能造成有多個能與模板 DNA 結合的位置而形成非特異性複 製々若太長會導致過高的熔點而使酵素活性降低。以下列出一般引子設計的原則〆 (1) 引子長度通常為 18~22 個鹼基。 (2) GC 比例為 40%~60%。 (3) 兩個引子的熔點相差不超過 5°C。 (4) 兩引子不可互補以免自相黏合。
(5) 注意模板 DNA 中所有可能與引子互補的區域並避開這些區域。 (6) 3′端的互補尤其重要。
實驗步驟〆
此實驗為利用 2-3 基因合成策略所提到的兩組 primer 合成兩個 PGB1 基因片 段 , 在 近 一 步 接 合 形 成 完 整 的 PGB1 基 因 DNA, 最 後 接 入 pET 200/D-TOPO vector。 成分 最後濃度 dNTP 10 M 10 M 1-36 fp/78-117 fp 0.2 M 10 M 33-85 rp/117-174 rp 0.2 M 10X reaction buffer 1X Klenow 1 (0.5 unit) Total Volume 30 l
10X reaction buffer〆500 mM Tris-HCl (pH 7.2), 100 mM MgSO4 and 1 mM DTT.
如上表標示成分及其比例加入混合。兩組 primer 先於先加熱至 94℃、2min。 再降溫至室溫(25°C)30 分鐘,再加入 Klenow(invitrogen, Carlsbad, CA)於溫 度 30℃下反應 15 min。反應完後,加熱至 75°C 約 10 分鐘,執行電泳並做 gel extract
(Biokit, Taiwan)。加入相同於樣品體積的 binding buffer 充分混勻,加熱至 60°C,
10 分鐘。將 kit 中的 column 與收集管結合,接著在將混勻溶液加入 column,並
離心一分鐘,接著加入 washing buffer(含酒精)600 l,並以離心機 (Beckman
Coulter Avanti® J-E Centrifuge, Fullerton, CA ) 14,000g 離心一分鐘。離心完後,捨 去收集管內的液體,再將新的 washing buffer 加入 column,並再以 14,000g 離心 一分鐘。將收集管內的液體移除,並再次離心 14,000g 離心三分鐘以去除剩餘之 酒精。最後,將 column 置入新的 eppendorf tube,並加入適量無菌水,離心沉澱, 即可獲得純化後產物。
60°C 以去除酵素活性,並進行 lean up 純化已被 digest 的基因片段。加入 ligase 將兩段基因片段接合,於 4℃下反應 overnight。最後,將接合產物(ligation product) 與 pET 200/D-TOPO vector(Invitrogen, Carlsbad, CA)於室溫下反應混合 30 min。
3-2 IPTG 誘導蛋白質表現 (Protein Expression)
Isopropyl-β-D-thio-galactoside (IPTG)誘 導的蛋白質 表現機 制是利用 IPTG
這種乳糖相似物誘導乳糖操縱子(lac operon),啟動表現下游的 T7 RNA 聚合酶 ( T7 RNA polymerase), 接 著 T7 RNA 聚 合 酶 活 化 載 體 上的 T7 促 進 子 (T7
promoter),使得目標蛋白質被大量表現。由於 T7 RNA 聚合酶的活性遠高於 E. coli
原有的 RNA 聚合酶, E. coli 自身基因轉錄圖敵不過 T7 表現系統而使整個細胞 的資源被用在表現目標蛋白質上。以上由 lac operon 調控 T7 表現系統機制的 E.
coli,是與噬菌體 DE3 熔源化的菌株(例如 E. coli BL21(DE3)),帶有 lacI 抑制基
因和位於 lacUV5 啟動子下的 T7 RNA 聚合酶基因。另外,IPTG 雖能被 E. coli 所利用,但並不會被代謝,所以可以使得蛋白質穩定地表現。
Isopropyl-β-D-thio-galactoside (IPTG)
實驗步驟〆
將 序 列 確 定 的 菌 株 放 入 1ml LB 培 養 液 ( 以 下 LB 培 養 液 皆 含 有 0.02% Kanamycin) 中 培 養 四小 時 。待 四 小 時培 養 後, 將 菌 液分 為 兩管 , 並各 加 入 新 鮮 的 2.5 ml LB 培養液,其中一管加入 IPTG 至總濃度為 1 mM,兩管皆於 37°C 隔 夜培養(12~16 小時)。隔夜培養後,抽取 1ml LB 菌液,以 12,000 g 離心一分鐘 分離細菌。分離後的細菌以 100 l sample buffer(4% SDS,0.2% Bromophenol Blue, 20% Glycerol,200 mM DTT,100 mM Tris-HCl,pH6.8)回溶,再以超音波震盪 器(UP200S,Hielscher,German)以功率 70%,單次震盪 0.5 秒,震盪細菌一分
鐘共五個週期,使細菌破碎。再將破碎的細菌以 15% SDS-PAGE 確認蛋白質的表 現。 將小量表現成功的菌株取出 60 l 加入 3ml LB 培養液隔夜培養活化。隔夜培 養後,將飽和菌液取出 60 l 再次加入 3ml LB 培養液培養四小時,此為第二次活 化。第二次活化後,取出 300 l 菌液加入 300 ml LB 培養液,先培養四小時後再 加入 1 mM IPTG 隔夜培養。隔夜培養後,將菌液以 9,000 g 離心 20 分鐘以沉澱細 菌,移除 LB 培養液後加入 15 ml His-tag binding buffer (20 mM Imidazole,50 mM NaCl,10 mM Tris-HCl,pH 7.2)回溶細菌。利用細胞破碎機(One Shot Model, Constant Systems Ltd,UK)將細菌打碎,再以 12,000 g 離心 30 分鐘分離目標蛋 白質々此蛋白質 PGB1 為水溶性,故分離後是存在於上清液中。取出上清液再以 His-tag Column 純化。
3-3 聚丙烯醯胺膠體電泳(Polyacrylamide Gel Electrophoresis,PAGE)
電泳(Electrophoresis)係指帶電粒子在電場作用下,向著該粒子自身相反電 性方向的電極移動的現象々由於各粒子的帶電量可能不同而使遷移速率不同,故 能使不同的粒子分離。此現象早於 1808 年就被發現,但一直到 1937 年才第一次 被當作粒子分離方法並且該科學家(瑞典科學家 Tiselius)以此貢獻在 1948 年獲 得諾貝爾獎。但由於該電泳所使用的介質分辨率不佳且整體儀器造價昂貴,促使 在五零年代開始有許多科學家尋找更佳的電泳介質。直到六零年代 Davis 等科學 家發展出以聚丙烯醯胺(Polyacrylamide)作為電泳介質的電泳儀器[57],使得此 技術變得更簡單方便,並擁有更佳的分辨率。 聚 丙 烯 醯 胺 膠 體 電 泳 是 一 種 不 連 續 膠 體 的 蛋 白 質 膠 體 電 泳 ( Gel
electrophoresis),由焦集膠體(Stacking gel)和分離膠體(Separating gel)所組
成。兩膠體皆由聚丙烯醯胺(Polyacrylamide)所組成,但兩者的膠體孔徑不同、 pH 值不同、緩衝液離子成分亦不同。這三種不同使得蛋白質混合液在焦集膠體中 進入分離膠體前壓縮成一條狹窄的區帶,而進入分離膠體後,蛋白質隨著空間中 的電場遷移,因各個蛋白質所帶淨電荷不相同造成不同的遷移速率,又因分離膠 體的孔徑很小造成不同分子大小和形狀的蛋白質表現出不同的遷移速率。到了六
(Sodium dodecyl sulfate,SDS)的帶電清潔劑作為蛋白質變性劑能夠使得蛋白質 變性而去除蛋白質本身形狀造成的遷移率差異,而且與 SDS 的結合使蛋白質表面 均勻帶電進而使蛋白質電泳的遷移速率只與其本身的分子量大小有關[58], [59]。 使用 SDS 的聚丙烯醯胺膠體電泳常簡稱做 SDS-PAGE。 3-4 蛋白質變性 (Protein Denaturation) 蛋白質變性係蛋白質的結構受到破壞或影響使 之與原來的構型不相同,這可 能造成許多的影響,例如聚集沉澱或失去其生理活性。蛋白質的變性可以發生在 蛋白質的各級結構〆在四級結構的變性,蛋白質的次級單元分離或是在空間排列 生發生錯亂々在三級結構的變性,主要影響的是各個胺基酸殘基的共價鍵(例如 雙硫鍵)、極性胺基酸彼此或與溶劑之間的偶極耦合交互作用(例如 Salt bridge), 以及非極性胺基酸之間的凡德瓦力和疏水性作用(Hydrophobic interaction)々在 二級結構的變性,蛋白質失去原有的胺基酸排列的模式包括α-helix、β-sheet、β-turn 等。不過,蛋白質變性並不包括其一級結構[60]。 穩定蛋白質結構最主要的作用力是疏水性作用、氫鍵( Hydrogen bonds)和 離子作用(Ionic interaction),故大多數的變性因素是影響這些作用力而變性蛋白 質,包括溫度、pH(酸鹼)、有機溶劑、變性劑、還原劑、機械力等等 。變性劑 (Denaturant)主要是破壞蛋白質內部的疏水性作用和氫鍵,常用的變性劑如 鹽酸 胍(Guanidine hydrochloride)和尿素(Urea)。而還原劑是將雙硫鍵還原為兩個 硫 氫 基 , 這 可 能 造 成 蛋 白 質 三 級 結 構 崩 毀 , 常 見 的 還 原 劑 有 1,4- 二 硫 蘇 糖 醇 (Dithiothreitol,DTT)和 2-巰基乙醇 (β-mercaptoethanol)。
3-5 蛋白質摺疊 – 過臨界點摺疊法(Over-critical Folding Process)
此蛋白質摺疊方法以階段性熱平衡透析法為基礎,以透析法使變性的蛋白質 在一個緩慢變化的環境逐步摺疊成自然態,此透析過程稱作準靜過程(quasi-static process)[4, 7]。我們認為在 蛋白質 摺疊 反 應相圖中 包含 了一個 相轉變 線(phase
臨界範圍,因此設計出一個繞過態轉變線的蛋白質摺疊方法,稱作過臨界點摺疊 法(over-critical folding process)[5, 6]。這樣的摺疊方法主要是藉由 urea 和 pH 值的逐步改變使蛋白質摺疊,並利用了 mannitol 作為蛋白質摺疊時的化學輔佐子 (chemical chaperone)[5, 61]以保護並穩定蛋白質的摺疊中間態。
實驗步驟〆
將蛋白質 PGB1 以高鹼性及高 urea 濃度的 denature buffer (8 M urea,0.1% mannitol,10 mM Tris,pH 12)變性後,利用 MWCO 為 3.5 kDa 的透析膜逐步透
析至 M1、M2、M3、M4和 M5等五個摺疊階段緩衝液(各成分如表三所示),每次 皆以 2,500 ml 摺疊緩衝液透析並至少二次。透析膜須以 0.1 M EDTA 水溶液於高 溫(> 80°C)浸泡約五分鐘以除去透析膜上殘存的金屬離子,再以二次去離子水 清洗後方可使用。 因為需要觀察各摺疊中間態的蛋白質,故在每一階段透析完成後,便從透析 膜中取出適量的蛋白質溶液並以吸收光譜測量濃度,其餘的蛋白質溶液收回透析 膜中繼續摺疊直至 M5狀態。
表三〆Over-critical Folding Process 各摺疊狀態緩衝液成分
Folding State Urea Mannitol Tris-HCl pH
M % mM Denature 8 0.1 10 12 M1 2 0.1 10 11.5 M2 1 0.1 10 11.5 M3 0 0.1 10 11.5 M4 0 0.1 10 8.8 M5 0 0 10 8.8
3-6 等溫滴定微量熱法 (Isothermal Titration Calorimetry)
等溫滴定微量熱法是一種物理技術用以偵測溶液中的反應熱動力參數變化,
包括結合常數(associate constant,Ka)、熱焓變化(enthalpy change,ΔH)和結
研究。等溫滴定微量熱儀的基本架構是以兩個相同的高效率熱傳導材料的反應槽 ( amouple) 放 置 於 絕 熱 套 內 , 一 為 樣 品 槽 , 另 一 為 參 考 槽 。 高 敏 感 的 熱 電 偶 (thermopile)電路將偵測兩反應槽的熱量差。參考槽會持續地施加穩定的熱功率, 當另一個研究樣品已精確的量滴定入樣品槽產生熱量變化(反應熱或是稀釋熱), 樣品槽的加熱器將對此產生的熱量施以一回饋熱功率,使樣品槽與參考槽再次達 到熱平衡。實驗的測量將持續地偵測熱量對時間的變化。 實驗步驟〆 將蛋白質樣品以孔徑 0.22 m 的濾膜過濾,0.2 ml 濃度 237 M 的 PGB1 放置
於 syringe,2.8 ml 濃度 6 M 的 IgG 放置於 ampoule,兩者緩衝液皆為 phosphate
buffer saline (pH 7.4)々攪拌器轉速設定在 60 rpm,功率放大器(amplifier)設定
於 30 W々實驗將 syringe 裡的樣品分為 20 滴注入 ampoule 中,每次間隔十五分 鐘,測量每次樣品注入時的熱量變化。將每次熱量變化對當下樣品比例作圖可以 得出 PGB1 與 IgG 的結合比例以及反應常數。 3-7 螢光光譜 (Fluorescence Spectrum) 圖九〆解釋螢光發生機制的 Jablonski 圖[39] 絕大部分的穩定有機化合物均屬單重態( singlet)分子,意即該類分子為基 態時電子均成對(故其總自旋量子數為零),而自旋重態數( spin multiplicity)為 1。(multiplicity2n1,n 為未配對電子數)一般的螢光分子都屬於單重態(singlet,
S),因此當分子吸收 UV-Vis 光區的電磁波時,電子會在約 10-15秒從單重態基態 S0(ground State)躍遷至具有相同自旋重態數的激發態(S1、S2、…),例〆 0 A 1
S
h
S
此式子中,h 為普朗克常數,A為吸收的光子頻率。若激發的電子躍遷到更高的 激發態如 S2,在 10-14~10-11秒的時間會經由內轉換(internal conversion)以無輻 射躍遷至能量稍低的同自旋多重度的 第一個激發態(S1)的最低振動態,接著很 快地以發光的方式釋放能量(hF)回到基態 S0,如下表示〆 2 1 0 FS
S
S
h
此時所發出的光便是一般所謂的螢光,該螢光頻率為F。一般來說,用來激發螢 光分子的能量會大於釋放出的螢光能量,因此F會小於A,及螢光波長會大於激 發波長。螢光壽命(lifetime〆螢光分子維持在激發態的時間。)大約在 10-9~10-7 秒。通常電子從 S2躍遷至 S1的內轉換過程非常快(10-14~10-11秒),從不同的振 動能階將能量以熱方式釋放而到達 S1的振動基態,此過程稱作內轉換或振動弛緩 (vibrational relaxation),故大多數的螢光來自於 S1。 另外,激發的電子可以從激發單重態 S1經由系間跨越(intersystem crossing) 無輻射躍遷至能量較低但具有不同自旋多重度的激發態(一般多為自旋三重態), 再以同於單重態的模式由內轉換無輻射躍遷至自旋多重態的振動基態,接著又以 光的形式將能量釋放而回到基態 S0,此時所發出的光稱為磷光(phosphorescence)。 不過,從自旋多重態的振動基態回到 S0的躍遷過程受到躍遷選擇規則限制,因此 磷光的壽命(lifetime)多半很長,而其量子產率(quantum yield〆吸收單位數量 光子後,能發出多少的光子々意即發光效率)遠小於螢光。綜合以上的電子躍遷 發光機制可以用 Jablonski 圖來解釋(圖九)。由於很多的生物分子在適當的激發 光下都能夠發出螢光,故在生物科技領域中螢光的應用非常廣泛。 實驗步驟〆 將蛋白質濃度調整至 10 M,總體積為 1 ml 放置 於 1 cm 厚的螢光比色管。設定實驗條件為〆excitation,295 nm々emission,300~500 nm々PMT voltage,700 V々 Scan rate,240 nm/min々excitation and emission slit ,5 nm。實驗所用的螢光光譜
3-8 螢光焠熄 (Fluorescence Quenching)
螢 光 焠 熄 意 指 任 何 造 成 樣 品 螢 光 強 度 減 弱 的 效 應 , 包 括 激 發 態 反 應 (excited-state reaction)、分子重排(molecular rearrangements)、能量轉移(energy transfer )、 形 成 基 態 錯 合 物 ( ground-state complex formation ), 以 及 碰 撞 焠 熄 (collisional quenching)[39]。在此部分,我們主要解釋碰撞焠熄(又可稱作動態
焠熄 dynamic quenching)和靜態焠熄(static quenching),而能量轉移將在下一章
節介紹。無論是碰撞焠熄或是靜態焠熄,都需要焠熄劑與螢光發光基團接觸,故 兩者的接觸程度將決定螢光焠熄的程度々藉由此一特性便能夠以螢光焠熄的程度 判斷發光基團曝露於溶劑的程度。
碰撞焠熄係指在螢光焠熄過程中,焠熄劑在溶液中擴散接觸處於激發態的發 光基團並使之以非輻射方式退回基態,兩者並不形成光化學反應。重原子的碰撞
焠熄劑,例如碘(iodide)和溴離子(bromide),增加了系間跨越(intersystem crossing)
可 能 性 而 使 得 螢 光 被 焠 熄 [62] 。 另 外 , 其 他 的 螢 光 焠 熄 劑 , 例 如 丙 烯 醯 胺 (acrylamide)[42]、銫離子(Cs+)[63]、咪唑(imidazole)[64],可能是接受了 螢光發光基團的電子而使螢光焠熄。碰撞焠熄的電子躍遷機制可以 Jablonski 圖表 示(圖十)。 圖十〆描述碰撞焠熄和螢光共振能量轉移的電子躍遷機制的 Jablonski 簡圖 i k 表示為除了碰撞螢光 焠熄和能量轉移之外的非輻射退躍遷過程。 靜態焠熄係指在焠熄劑與螢光發光基團接觸形成基態錯合物,使其吸收光後 立即回到基態而不發出螢光(圖十一)。此 焠熄過程使得部分的螢光發光基團無法 發光而造成螢光強度的減弱,因此螢光發光基團的壽命( lifetime)將不會改變,
這是靜態焠熄與碰撞焠熄最大的不同點之一。另外,改變實驗溫度亦可以區分兩 者〆溫度升高會使得碰撞焠熄劑的擴散係數(diffusion constant)增加而使焠熄作 用增強,反之,靜態焠熄劑與發光基團形成的基態錯合物因升溫而解離進而使焠 熄作用減弱。 圖十一〆靜態焠熄分子機制簡圖。 實驗上,焠熄劑的焠熄作用強度可以用 Stern-Volmer equation 表示〆 0 0 1 SV[ ] 1 q[ ] F K Q k Q F 方程式中,F0表示沒有焠熄劑存在時的螢光強度々F 表為焠熄劑存在時的螢光強 度々[Q]為所加的焠熄劑濃度々KS V則為 Stern-Volmer 焠熄常數々τ0表為螢光物初 始的螢光壽命々kq則為實際的螢光焠熄能力。實驗數據以 F0/F 對[Q]作圖,斜率 便是 KS V。碰撞焠熄與靜態焠熄皆可以形成線性的 F0/F vs. [Q]關係圖々得出 KS V 後,再除以該螢光物質最初始的螢光壽命,便獲得焠熄劑在當下環境對該螢光物 質的焠熄能力。 實驗步驟〆 將蛋白質濃度調整至 10 M,總體積為 1 ml 放置 於 1 cm 厚的螢光比色管。設
定實驗條件為〆excitation,295 nm々emission,300~500 nm々PMT voltage,700 V々 Scan rate,240 nm/min々excitation and emission slit ,5 nm。實驗所用的螢光光譜
儀為 F-7000,Hitachi,Tokyo。準備 5 M acrylamide 作為原液,每次取 2 l 加入 蛋白質溶液觀察螢光強度,重複數次。以 F/F0(焠熄的螢光強度除以原螢光強度) 對 焠 熄 劑 濃 度 作 圖 , 在 Stern-Volmer 方 程 式 獲 得 acrylamide 對 PGB1 的 Stern-Volmer 常數 KSV。而 acryamide 對 PGB1 的螢光焠熄能力 kq再由以下關係式 獲得〆 0 SV q K k
3-9 螢光共振能量轉移 (Förster Resonance Energy Transfer)
螢光共振能量轉移(Fluorescence Resonance Energy Transfer ,FRET),是描 述 兩個 發 光基團之 間 能量轉 移 的機制 々 由 德 國科學家特 奧多〄 福斯特(Theodor Föster)於 1946 年所發現[46]。螢光共振能量轉移是螢光焠熄機制的一種,其焠 熄效果(能量轉移效率)與供體和受體兩者之間的距離有以下的關係式〆
6 6 6 0 0 1 DA/ D / E F F R r R r E 表示為能量轉移效率,FD A與 FD各表示螢光能量供體有與沒有伴隨下的螢 光強度,R0和 r 各表示為 Föster distance 以及供體與受體之間的距離。R0可由以 下關係式得到〆 6 5 2 4 0 8 79 10. ( D ( )) R Q n J 6 Å ( ) R0主要受到螢光供體的螢光量子產率(QD)、折射率(n)和光譜重合區域(J(λ)) 所 影 響 。 光 譜 重 合 區 域 指 的 是 螢 光 能 量 供 體 的 標 準 化 發 射 光 譜 ( normalized fluorescence spectrum)與螢光能量受體以消光係數(extinction coefficient)表示的吸收光譜,兩者的重合面積〆 4
( ) D( ) A ( )
J
f ε d。因為螢光共振能量轉移與距離有很大的關係且該距離通常為 1~10 nm,故常 被用在觀察生物分子的動態研究上,例如蛋白質與蛋白質之間的交互作用[47, 65, 66],蛋白質與 DNA 之間的交互作用[67-69],DNA 或 RNA 構型的改變[70, 71], 以及蛋白質內部的構型變化[72-74]。
實驗步驟一〆測量 IAEDANS 接合率
將 PGB1 G9C 或 A23C 溶解在 Tris-HCl buffer (10 mM Tris-HCl,pH7.2)。加 入與蛋白質相同濃度的 DTT 使蛋白質的雙硫鍵還原,反應在室溫約九十分鐘。待 雙硫鍵還原反應結束後,緩慢滴入 IAEDANS 到攪拌中的蛋白質溶液中,IAEDANS 的最後濃度為蛋白質的五倍。使 IAEDANS 與蛋白質接合反應約兩個小時在室溫 並且需要避光。反應後,以大量而頻繁的透析將蛋白質與 IAEDANS 混合液置換 到 Tris-HCl buffer (10 mM Tris-HCl,pH7.2)並去除多餘的 IAEDANS。最後,將透 析後的溶液測量 UV 吸收光譜並以下列公式計算接合比率〆
280 336 10810 1448 107 5700 [ ] [ ] [ ] [ ] A P F A P F A280和 A336為 280 和 336 nm 的吸收值々[P]和[F]表示為蛋白質和 IAEDANS 在 溶液中的濃度。此公式是 PGB1 和 IAEDANS 的在 280 和 336 nm 的消光係數組合 而成。PGB1 在 280 nm 的消光係數為 10,810 M-1
cm-1,由 Gill 和 von Hippel [75] 的方法所預估得知々在 336 nm 的消光係數是依比例推算得知。IAEDANS[53]在 336 nm 的消光係數為 5,700 M-1cm-1,280 nm 的消光係數為依比例得知。代入 280 和 336 nm 的吸收值便可以以二元一次聯立方程式解出 PGB1 和 IAEDANS 的濃度, 進而得知兩者接合的比例。 實驗步驟二〆FRET 距離計算 將 PGB1 G9C-IAEDANS 與 A23C-IAEDANS 濃度調整為 10 M,以前述螢光 光譜測量相同的條件測量其螢光。此螢光強度為 FD A,將同樣濃度的 PGB1 G9C 與 A23C 的螢光強度為 FD,計算各突變蛋白質在各突變狀態下的螢光共振能量轉 移效率 E。能量轉移效率 E 與供體和受體兩者之間的距離 r 有以下的關係式〆
6 6 6 0 0 1 DA/ D / E F F R r R r E 表示為能量轉移效率,FD A與 FD各表示螢光能量供體有與沒有伴隨下的螢 光強度,R0各表示為 Föster distance。R0可由以下關係式得到〆 6 5 2 4 0 8 79 10. ( D ( )) R Q n J 6 Å ( ) QD表示供體的螢光量子產率,n 為折射率和 J(λ)為光譜重合區域所影響。光 譜 重 合 區 域 指 的 是 螢 光 能 量 供 體 的 標 準 化 發 射 光 譜 ( normalized fluorescence spectrum)與螢光能量受體以消光係數( extinction coefficient )表示的吸收光譜,兩者的重合面積〆 4
( ) ( )
D A
J
f ε d。3-10 圓二色光譜 (Circular Dichroism Spectrum)
當一束平面偏振光通過具有手性(chiral)的介質時,因該手性介質對於左右
Lambert Law [76]〆
( )
L( ) R( ) c l 其中
L( )
、
R( )
表示在各波長下手性介質對左右旋圓偏振光的消光係數,c 為手 性介質濃度,l 則為光路徑長(cm)。此現象又稱作圓二色性(Circular Dichroism)。 由於左右旋圓偏振光的吸收率不同,故一束平面偏振光通過手性介質後便會形成 一橢圓偏振光,該橢圓偏振光的橢圓率( Ellipticity,θ)可以由該物質的 左右旋 圓偏振光消光係數差( ) 轉換獲得〆 45 10 3297 2 ln ( ) . ( )
此轉換得到的橢圓率單位為 deg。手性介質對各個波長造成的吸光率差不同,將 各波長下形成吸光率差對該波長作圖便形成圓二色光譜,並從此光譜可以了解該 介質結構的手性特性。 蛋白質是由許多胺基酸以胜肽鍵連接並具有特定結構的生物聚分子,而在其 中胜肽鍵、芳香族胺基酸和雙硫鍵具有圓二色光特性。在遠紫外光波段(光波長 小於 250 nm)的圓二色光譜能夠揭示蛋白質在該溶液中的二級結構比例( 包括 α-helix、β-sheet、β-turn 等等),主要由胜肽鍵的電子躍遷引貣[77]。近紫外光波 段(光波長介於 250 nm 至 300 nm)的圓光二色光譜則能代表三級結構的變化, 主要由芳香族胺基酸(Phe、Tyr、Trp)的電子躍遷引貣[77]。 蛋白質的遠紫外光圓二色光譜中,α-helix 結構在 192 nm 有一正訊號(右旋), 在 222 和 208 nm 則表現負訊號(左旋)々β-sheet 結構在 216 nm 有一負訊號,在 195 nm 有一正訊號々β-turn 結構在 212 nm 有一正訊號,在 200 nm 則有一負訊號。 以上結構訊號整理如表四。 為 比 較 各 個 蛋 白 質 造 成 的 橢 圓 率 , 一 般 習 慣 將 橢 圓 率 表 示 為 平 均 殘 基 橢 圓 率 (Mean residue ellipticity, [θ]MRE)〆10 [θ]MRE r Mw c l n 其 中 Mw 為 蛋 白 質 分 子 量 , nr 為 組 成 單 體 或 殘 基 的 數 量 々 MRE 單 位 為 2 degcm /dmol。
表四〆蛋白二級結構 之圓二色光譜特徵峰
蛋白二級結構-圓二色光譜特徵峰 Secondary
Structures - band (nm) + band (nm)
-helix 222、208 192 -sheet 216 195 -turn 220、203 205 Polypro-II helix 190 210-230 Random coil 200 212 實驗步驟〆 使用 1 mm 厚度的比色管,蛋白質樣品濃度調整為 0.1~0.5 mg/ml,總體積至 少 200 l。通入氮氣以去除光電倍增管上的氧氣,並打開水冷卻系統(20°C)等 待 二 十 分 鐘 々 打 開 Xe Lamp power 並 開 啟 電 腦 , 進 入 量 測 軟 體 。 實 驗 參 數 〆 wavelength range,190~260 nm々scan speed,100 nm/min々accumulation,10s々 interval,0.2 nm々band width,1nm。使用機型為 AVIV model 410(AVIV,New Jersey, USA)。
3-11 動態光散射 (Dynamic Light Scattering)
雷射光射入含有目標分子的溶液中,分子與雷射光產生彈性散射,該散射光 的波長與原入射光相同且互相干涉並隨時間改變。散射光的改變與分子在溶液中 的布朗運動(Brownian motion)有關,意即這變化又與分子的大小有關々分子越 小則布朗運動越強烈,分子越大則反之。因此,利用 Stokes-Einstein 方程式可以 得知布朗運動(擴散)與分子大小的關係〆 6 B k T D r D 表示為擴散常數,T 為絕對溫度,η 為溶液黏滯性( viscosity),r 為球形分 子的半徑。此方程式計算得知的分子半徑包含分子外層的水合層,且以分子為球 形做假設。
第四章 實驗結果
4-1 基因序列
目標 PGB1 基因序列全長為 178 bp(如圖十三中的 PGB1 G9C 基因所示),接合入
pET200 基因載體中 E. coli BL21( DE3)系統表現蛋白質。接入 pET200 基因載體後 的 PGB1 基因可以利用 PGB1 fp 和 T7 rp 或 T7 fp 和 PGB1 rp 確認目標基因為正確 方向接入載體中(如圖十二和圖十四所示)。 圖十二〆以 PGB1 fp 與 T7 rp 所挑選放大的 PGB1 wt 基因 以 PGB1 fp 與 T7 rp 所挑選放大的 PGB1 wt 基因長度應為 245 bp(箭頭所指處),並且藉此確認 目標 PGB1 基因是正確 地接合入 pET200 基因載體中。
bp
500 400 300 200 100圖十三〆PGB1G9C 基因 PGB1 基 因全 長 為 178 bp( 箭 頭所 指 處), 利用 預 先 設計 好 的 PGB1 G9C fp 與 PGB1 rp 進行 PCR 反應以此送交基因定序。 圖十四〆以 T7 fp 與 PGB1 rp 所挑選放大的 PGB1 A23C 基因 以 T7 fp 與 PGB1 rp 所挑選放大的 PGB1 wt 基因長度應為 371 bp(箭頭所指處),並且藉此確認 目標 PGB1 A23C 基因是正確地接合入 pET200 基因載體中。
bp
500 400 300 200 100 M 1 2 3 4bp
500 400 300 200 1004-2 大量表現並純化 PGB1 蛋白質
將帶有 pET200-pgb1 基因的 E. coli BL21( DE3)以 IPTG 誘導表現至少八小時, 破菌後 12,000 離心 20 分鐘,再從上清液純化帶有 His-tag 的 PGB1。以 15% SDS-PAGE 確認表現和純化效果。大約 1.8 公升菌液可以得到約 40 mg 的蛋白質 PGB1々純化後的蛋白質純度至少 96%。各突變株皆以相同方式獲得。蛋白質 PGB1 的分子量約為 10 kDa。
圖十五〆蛋白質 PGB1(包括突變株)表現效果以及從上清液純化效果
圖(a)~(c)中,各 lane 的表示為〆M,Unstained Protein Marker (mid range) (3.5 L)々1,表 現 PGB1 wt 之破 菌 後上 清 液々2,通過 His-tag column 後 的 上清 液 々3, 純化 後 PGB1 wt (a)、A23C (b)或 G9C (c)。 以上 皆 是 15% SDS PAGE。
kDa
116.0 66.2 45.0 35.0 25.0 18.4 14.4 M 1 2 3 M 1 2 3 M 1 2 3 (a) (b) (c)4-3 摺疊後 PGB1 功能測試 (ITC 實驗)
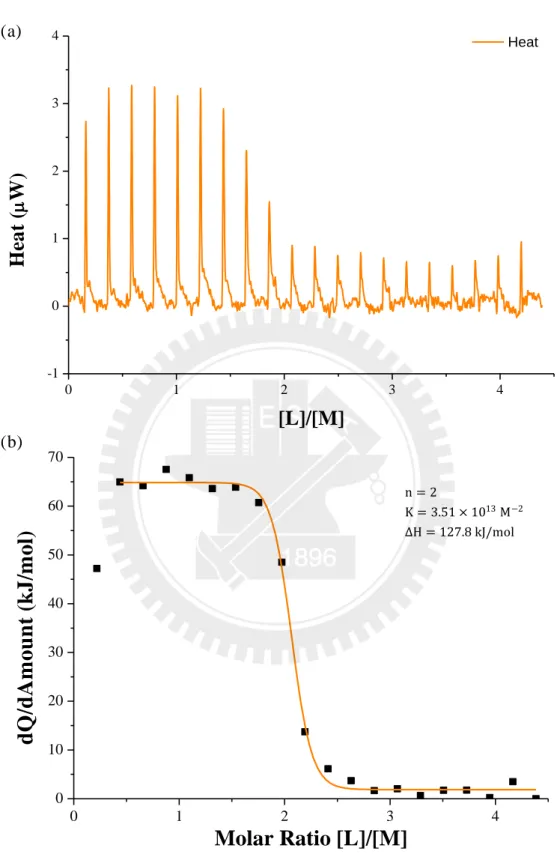
此實驗係 為了確 認摺 疊後之 蛋白質 PGB1 是否還具 有自然 與免 疫球蛋 白質 (IgG)結合的能力。自然狀態下,一個 IgG 能與兩個 PGB1 鍵結。PGB1 與 IgG 的鍵結與其三級結構有關。
以 47.4 nmole 摺疊後的 PGB1 wt 與 10.8 nmole rabbit IgG 進行滴定作用,觀 察每一次作用的熱量變化。所使用的緩衝液為 phosphate buffer saline (pH 7.4)。圖 十六(a)為每一次滴定作用所造成的熱量變化,總共有 20 次滴定作用。圖十六(b) 為每 次熱 量 的變 化 量與 各次 滴 定狀 態 下的 蛋白 質濃 度 比例 。 結果 顯示 摺 疊後 的 PGB1 wt 與 IgG 的結合比率為 2:1,與自然狀態摺疊的 PGB1 wt 狀態[21]相同々反 應常數為 ,莫爾反應熱為 。
0 1 2 3 4 -1 0 1 2 3 4
H
e
a
t
(
W
)
[L]/[M]
Heat 0 1 2 3 4 0 10 20 30 40 50 60 70d
Q
/d
A
m
o
u
n
t
(k
J
/m
o
l)
Molar Ratio [L]/[M]
n = 2 K = 3.51 × 1013 M−2 ΔH = 127.8 kJ/mol圖 十 六 〆 Over-critical Refolding Process 摺 疊 後 的 PGB1 wt 與 IgG 的 Isothermal Titration Calorimetrics 結果 。(a)每 次滴 定 作用 的 功率 變化 。 (b)每 次 滴定 作 用的 莫 爾熱 變 化。
(a)
4-4 蛋白質摺疊中間物之螢光光譜變化
利用 295 nm 激發光,我們能夠觀察 Trp 這個疏水性胺基酸環境的變化。隨著
Over-critical Folding Process 摺疊,蛋白質 PGB1 wt 的 Trp43 螢光整體逐漸增強(圖
十七)々傴在 M2進入 M3摺疊態時有個明顯的下降的現象,可能與 urea 保護了蛋 白質使所受的高 pH 變性的程度減低。在 8 M Urea 和 pH 12 環境(denatured 狀態) 底下變性的 PGB1 wt 的螢光強度或說 Trp43 的螢光量子產率(fluorescence quantum yield)傴約只有摺疊至 M5狀態下的三分之一々將環境中的 urea 逐漸地減少並維 持在 pH 11 的高鹼性,PGB1 wt 的螢光便逐漸地增強,當 pH 下降到 8.8 時,整體 的螢光強度立刻增強原有的一倍々M4狀態下地螢光略強於 M5狀態可能是因為有 了 mannitol 這種化學輔佐子穩定蛋白質結構使得 M4狀態下的 PGB1 wt 略較穩定 而有較強的螢光強度或螢光量子效率。 另外,從螢光光譜波峰波長(Emission Maximum,λma x)的變化為藍位移(圖 十七內插圖),意為波峰往低波長移動。在 denatured 狀態下,λma x位在較大的波 長 345.0±0.4 nm々M1狀態下,λmax為 341.4±0.4 nm々M2,341.4±0.4 nm々M3,341.6±0.4 nm々M4,341.2±0.4 nm々M5,340.8±0.4 nm。λma x變化與 Trp43 所處的周遭微環 境變化有關,當 Trp 在極性很強的環境中時,其螢光光譜波峰波長會出現在約 350 nm,而當 Trp 在非極性(疏水性)強的環境中,波峰的位置便會下降到約 325 nm々 因此,從 PGB1 wt 的 λma x變化可以得知 Trp43 周遭的環境在 denatured 環境下是 暴 露 在 高 極 性 環 境 中 , 隨 著 摺 疊 而 被 包 埋 入 蛋 白 質 的 疏 水 核 心 ( hydrophobic core)。
350 400 450 500 0.0 0.2 0.4 0.6 0.8 1.0 D M1 M2 M3 M4 M5 340 341 342 343 344 345 346
R
e
la
tiv
e
F
lu
o
r
e
sc
e
n
c
e
(
a
r
b
.
u
n
it
)
Wavelength (nm)
D M1 M2 M3 M4 M5 310 ma x (n m ) Folding State圖十七〆10 M PGB1wt 在 over-critical refolding process 中 不 同的 摺 疊狀 態 的螢 光光 譜 變化 , 左 上內插圖為各摺疊狀態的 螢光波峰波長(λma x)。激發光波長為 295 nm。
表五〆各摺疊狀態的螢光波峰波長(λmax)。
Folding State Emission Maximum λma x (nm) Denature 345.0±0.4 M1 341.4±0.4 M2 341.4±0.4 M3 341.6±0.4 M4 341.2±0.4 M5 340.8±0.4