中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/32626
82
0
0
全文
(2) 誌謝. 首先要感謝家人的支持和指導老師郭盛助、黃麗嬌、王繼平教授的教誨,使 我的畢業論文能趨於完善。在求學過程中,不僅僅是在實驗技巧上的鑽研,更重 要的是學習如何解決問題和增強邏輯思考能力,在這短短兩年半中,的確使我進 步成長許多。 還要感謝實驗室的好伙伴,首先最最感謝菊子學姐,在實驗上的指點迷津; 秉文學長在各方面的建議和指教;韻絜學姐、依瑩和洪喬在實驗室的幫忙,多虧 有了大家的協助,使我的論文能夠順利完成。最重要的是,有佩宜學姐、薰儀學 姐、婷翔、佩珊、峰哥和小鵲等好朋友的支持與鼓勵,陪我度過低潮。. i.
(3) 中文摘要 新合成 phenazine carboxylate 類的化合物 TCH 可以濃度依存性但並無明顯 的時間依存性的抑制 formyl-Met-Leu-Phe (fMLP) 所誘導嗜中性白血球超氧自由 基的生成(IC50 值為 4.5 ± 0.8 μM)。此抑制作用並非來自於影響細胞存活、超氧自 由基清除或直接抑制 NADPH oxidase 的活性。在相同的 IC50 值下 TCH 抑制嗜 中性白血球細胞中 p47phox 和 p22phox 的連結、Rac2 和 gp91phox 的連結、p47phox、 p21-activated kinase 1 (PAK1) 及 Vav 的 磷 酸 化 。 TCH 並 不 影 響 p38 mitogen-activated protein kinase 與 MAPK-activated protein kinase-2 的磷酸化。 TCH 可抑制細胞中 Akt 與 p47phox 的連結、Akt 的磷酸化及 Akt 酵素活性,但不 抑制細胞溶解液中的 Akt 酵素活性。TCH 可抑制 active human recombinant PDK1 酵素活性,但不影響 Akt 與 PDK1 的細胞膜轉移。TCH 會抑制 PKC-α、-δ與-ζ 和 p47phox 的連結作用,但不影響 PKC-β 和 p47phox 的連結。TCH 會抑制 PKC-α, 但不會抑制 PKC-β、-δ與-ζ的細胞膜轉移。TCH 也不會抑制 fMLP 刺激細胞引 起的 PKC 活性。TCH 不會增加細胞內 cyclic AMP 的含量。綜合以上結果,TCH 可能經由影響 PDK1/Akt、PKC 和 PAK1 訊息傳遞途徑來抑制 p47phox 的磷酸化, 經由影響 Vav/Rac2 訊息傳遞途徑來抑制 Rac2 的活化,進而抑制 NADPH oxidase 聚合及超氧自由基的生成。. ii.
(4) Abstract A. novel. synthetic. phenazine. carboxylate. derivative. TCH. inhibited. formyl-Met-Leu-Phe (fMLP)-stimulated superoxide anion generation in rat neutrophils in a concentration- but not a time-dependent manner with IC50 value about 4.5 ± 0.8 μM. This inhibitory effect was not owing to the decrease in cell viability, scavenging of generated superoxide anion or direct blockade of NADPH oxidase activity. Under the same IC50 value, TCH inhibited the interaction of p47phox and Rac2 with p22phox and gp91phox, respectively, and the phosphorylation of p47phox, p21-activated kinase 1 (PAK1) and Vav in fMLP-stimulated cells. TCH had no effect on the phosphorylation of p38 mitogen-activated protein kinase and MAPK-activated protein kinase-2. Pretreatment of cells with TCH inhibited the interaction of Akt with p47phox, the phosphorylation of Akt and Akt activity, whereas, it did not affect the Akt activity in the cell-lysates of fMLP-stimulated cells. TCH inhibited the enzymatic activity of active human recombinant PDK1, whereas, it had no effect on the recruitment of Akt and PDK1 to membrane. TCH attenuated the interaction of PKC-α、-δ and -ζ with p47phox, whereas, it did not affect the association of PKC-β with p47phox. TCH decreased the membrane recruitment of PKC-α but not of PKC-β、-δ and -ζ. TCH had neither block the PKC activity nor increase the cellular cyclic AMP levels of fMLP-stimulated cells. Taken together, TCH probably attenuate PDK1/Akt, PKC and PAK1 signalings and Vav pathway leading to the inhibition of p47phox phosphorylation and Rac2 activation, respectively, which in turn blockade the assembly of active NADPH oxidase and then superoxide anion generation.. iii.
(5) 目錄. 誌謝...............................................................................................................................iv 中文摘要......................................................................................................................ivi 英文摘要.......................................................................................................................iii 目錄.............................................................................................................................ivii 圖目錄............................................................................................................................ v 縮寫表..........................................................................................................................vii 第一章 緒論.................................................................................................................. 1 第二章 研究動機與目的............................................................................................24 第三章 實驗部分........................................................................................................25 第四章 結果與討論....................................................................................................34 第五章 結論................................................................................................................61 參考文獻......................................................................................................................62. iv.
(6) 圖目錄. 圖 1-1 PMN 在不同時期顆粒球分泌的物質. 14. 圖 1-2 NADPH oxidase 組成結構. 15. 圖 1-3 Cytochrome b558 的結構. 16. 圖 1-4 FPR 在化學刺激物下的訊息傳遞路徑. 17. 圖 1-5 PI3K class IA 和 IB 的結構. 18. 圖 1-6 Akt 的結構. 19. 圖 1-7 Akt 活化機制. 20. 圖 1-8 PAK 的種類和基本結構. 21. 圖 1-9 PAK 基本結構和活化過程. 22. 圖 1-10 MAPK 的活化路徑. 23. 圖 2-1 TCH 抑制嗜中性白血球產生超氧自由基的濃度依存性. 41. 圖 2-2 TCH 抑制嗜中性白血球產生超氧自由基的時間依存性. 42. 圖 2-3 TCH 對細胞存活率的影響. 43. 圖 2-4 TCH 對超氧自由基的清除作用. 44. 圖 2-5 TCH 對 xanthine-xanthine oxidase 產生超氧自由基的影響. 45. 圖 2-6 TCH 抑制 fMLP 刺激細胞產生超氧自由基之可逆性. 46. 圖 2-7 TCH 對cell-free系統NADPH oxidase 活性的影響. 47. 圖 2-8 TCH 對 fMLP 誘導 p47phox 活化之影響. 48. 圖 2-9 TCH 對 fMLP 誘導 Rac2 活化之影響. 49. 圖 2-10 fMLP 誘導 Akt 和 PDK1 細胞膜轉位的作用. 50. 圖 2-11 TCH 對 Akt 磷酸化、細胞膜轉位及與 p47phox 連結的影響. 51. v.
(7) 圖 2-12 TCH 對 Akt 活化的影響. 52. 圖 2-13 TCH 對 PDK1 活化的影響. 53. 圖 2-14 6BIO 對嗜中性白血球產生超氧自由基、Akt 和 PDK1 活化的影響 54 圖 2-15 TCH 對 PKC 活化的影響. 55. 圖 2-16 TCH 對 p38 MAPK、MK2 磷酸化的影響. 56. 圖 2-17 TCH 對 PAK 磷酸化的影響. 57. 圖 2-18 TCH 對 Vav 磷酸化的影響. 58. 圖 2-19 TCH 對 cyclic AMP 生成的影響. 59. 圖 2-20 結論. 60. vi.
(8) 縮寫表. 6BIO. (2'Z,3'E)-6-bromoindirubin-3'-oxime. AA. arachidonic acid. Akt. protein kinase B. aPKC. atypical PKC. ARF. ADP-ribosylation factor. CB. cytochalasin B. cPKC. classical or conventional protein kinase C. DAG. diacylglycerol. DHF. dihydroxyfumaric acid. DMSO. dimethyl sulphoxide. ERK. extracellular signal-regulated kinase. FAD. flavin adenine dinucleotide. FDA. fluorescein diacetate. fMLP. formyl-methionyl-leucyl-phenylalanine. FPR. formyl-peptide receptor. FPRL. fMLP receptor-like. GAP. GTPase-activating protein. GDI. guanine nucleotide dissociation inhibitor. GEF. guanine nucleotide exchange factor. GPCR. G-protein-coupled receptor. HBSS. Hanks’ balanced salt solution. IP3. inositol 1,4,5-trisphosphate. JNK. c-Jun N-terminal kinase. LY 294002. (4- morpholinyl)-8-phenyl-4H-1-benzopyran-4-one vii.
(9) MAPK. mitogen-activated protein kinase. MAPKAPK2/MK-2. MAPK-activated protein kinase-2. MKK/MEK. MAPK kinase. MKKK/MEKK. MAPK kinase kinase. NADPH. nicotinamide adenine dinucleotide phosphate. NBT. nitroblue tetrazolium. nPKC. novel PKC. PA. phosphatidic acid. PAK. p21-activated kinase. PB1. Phox and Bem1. PBD. p21-binding domain. PDK. 3-phosphoinositide-dependent protein kinase. PH. Pleckstrin-homology. PI3K. phosphoinositide 3-kinase. PI(3)P. phosphatidylinositol 3-phosphate. PI(3,4)P2. phosphatidylinositol 3,4-bisphosphate. PI(4,5)P2. phosphatidylinositol 4,5-bisphosphate. PI(3,4,5)P3. phosphatidylinositol 3,4,5-trisphosphate. PKA. protein kinase A. PKC. protein kinase C. PLA2. phospholipase A2. PLC. phospholipase C. PLD. phospholipase D. PMA. phorbol 12-myristate 13-acetate. PRR. proline-rich region. PS. phosphatidylserine. PVDF. polyvinylidene difluoride viii.
(10) PX SB 203580. Phox homology 4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4 -pyridyl)-1H-imidazole. SDS-PAGE. sodium dodecyl sulfate-polyacrylamide gel electrophoresis. SH2. Src homology 2. SH3. Src homology 3. SOD. superoxide dismutase. TPR. tetratricopeptide repeat. ix.
(11) 第一章. 緒論. 嗜中性白血球在免疫上扮演的角色. 人體對於微生物的攻擊所產生的保護性反應很類似軍事上的防禦。皮膚為抵 禦外敵入侵的第一防線,但病原菌若因皮膚受損或以其他路徑進入體內,則駐紮 在該處的防衛隊(吞噬細胞)必須立刻啟動防衛機制,此為先天性免疫系統 (innate immune system)。 多形核白血球 (polymorphonuclear leukocytes)為先天性免疫系統的成員,包 括嗜中性白血球、嗜酸性白血球和嗜鹼性白血球。其中嗜中性白血球約佔循環白 血球數的50-60%,細胞直徑約11-14 μm,缺乏粒腺體,核為多葉,細胞質富含 顆粒 (granules),在血液循環中只存活約7-10小時,為人體主要的噬菌系統。當 人體受到感染時,嗜中性白血球的數目會上升,並受到發炎物質如 chemokines 或 cytokines 的刺激,進行趨化作用 (chemotaxis) 到達患部。趨化作用的過程 裡,血液循環中的嗜中性白血球先向血管壁靠近,沿著內皮細胞滾動 (rolling), 透過 selectin 與內皮細胞連接,進而附著 (adhesion) 在內皮細胞上,再由內皮 細胞間隙滲出 (transmigration) 離開血管向著患部移動 (migration) (圖1-1)。隨 後,嗜中性白血球對病原菌進行吞噬作用 (phagocytosis),形成 phagosome,同 時伴隨著去顆粒化作用(degranulation),而 phagosome 會與分泌顆粒融合形成 phagolysosome,以殺死並分解細菌 (Segal, 2005)。 嗜中性白血球的殺菌作用分為兩類,一為非氧化性 (non-oxidative) ,二為 氧化性 (oxidative) 機制。非氧化性機制是利用分泌顆粒中多種酵素及殺菌蛋白 來 進 行 殺 菌 , 包 括 初 級 顆 粒 (primary/azurophilic granules) 中 的 lysozyme 、 β-glycerophsphatase、bactericidal/permeability-increasing protein (BPI)、elastase、 1.
(12) cathepsin G、protease 3、azurocidin、myeloperoxidase 等;次級顆粒 (secondary /specific granules) 中的 lactoferrin、lipocalin、lysozyme、LL37、MMP8、MMP9 和 MMP25;三級顆粒 (tertiary/gelatinase granules) 中的 lysozyme、gelatinase、 acetyltransferase等 (Nathan, 2006)。Chediak-Higashi syndrome 和 specific granule deficiency 為兩種罕見的遺傳疾病,因缺乏非氧化性機制造成白血球功能的異 常,導致病患容易造成較嚴重的感染。第二種殺菌作用,為氧化性機制是嗜中性 白血球活化膜上的 NADPH oxidase,此過程迅速消耗大量的氧分子還原成超氧 自由基 (superoxide anion),稱為呼吸爆發作用 (respiratory burst)。超氧自由基會 .. 代謝成H2O2、HO 和HOCl等產物,合稱為活性氧物質 (reactive oxygen species, ROS)。ROS會和細胞內的核酸、蛋白質、脂肪和醣類等作用,造成結構和生理 功能的改變。慢性肉芽腫病 (chronic granulomatous disease, CGD)是一種CYBB基 因突變因,NADPH oxidase異常無法正常生成超氧自由基,導致病患容易造成較 嚴重的感染。約70%患者為 gp91phox缺陷 ,25%是 p47phox的缺陷。但是NADPH oxidase過度活化也會造成正常組織器官的損傷,如動脈粥狀硬化 (atherosclerosis) 好發於慢性發炎反應。類風濕性關節炎 (rheumatoid arthritis)在關節部分發現大 量嗜中性白血球和巨噬細胞的聚集,因NADPH oxidase過度活化造成關節傷害 (El-Benna et al., 2009)。. NADPH oxidase NADPH oxidase 為蛋白質複合物,包括在細胞膜上的cytochrome b558 (由 gp91phox/NOX2 及 p22phox組成) 和存在細胞質中的 p47phox、p67phox、p40phox 和 small G protein Rac (Babior et al., 2002)。在細胞進行吞噬作用的同時也活化了 NADPH oxidase,使細胞質中的 p47phox、 p67phox、 p40phox 及 Rac 轉移到膜上, 與 cytochrome b558 結合形成完整具酵素活性的 NADPH oxidase。經由 NADPH 2.
(13) oxidase 的穿膜電子傳遞鏈 (transmembrane electron transport chain) (NADPH → FAD → 2 heme → O2),由 NADPH 提供電子來開啟反應,最後將氧分子還 原成超氧自由基 (NADPH + 2O2 → NADP+ +2O2.- + H+) (圖1-2) (Takeya and Sumimoto, 2003)。 NADPH oxidase 催化作用的核心是醣蛋白 gp91phox,存在嗜中性白血球細胞 膜和分泌顆粒的膜上。gp91phox 的 N-terminal 有六個穿膜的 α-helices,且第三 和第五的 helices 間有兩個 hemes,heme 以鐵離子與 His 殘基連接;而在細胞 質 中 的 C-terminal 為 ferredoxin-NADP+ reductase (FNR) 可 與 NADPH 和 FAD 結 合 ( 圖 1-3) (Hideki, 2008) 。 另 外 , 有 文 獻 指 出 若 阻 斷 gp91phox 的 C-terminal 後並不影響 NADPH oxidase 與其他蛋白質的結合,但降低 NADPH oxidase 的活性 (Burritt et al., 2003)。 p22phox 在 C-terminal 包含 proline-rich region(PRR)與 p47phox 的 Src homology 3 (SH3) domains 結合。另外,p22phox 的 PRR 有一段 polyproline II (PPII) helix,其氨基酸序列上的 Pro152、Pro156 和 Arg158 與 p47phox 的結合 有很重要的關係(Nobuhisa et al., 2006)。而在 cell-free system 中, p22phox 可 經由 phosphatidic acid (PA) 依存性的方式進行磷酸化,進而活化 NADPH oxidase (Regier et al., 1999)。 p47phox 或稱neutrophil cytosolic factor 1 (NCF1)由 390 個氨基酸構成,其結 構上包含有 N-terminal 的 phox homology (PX) domain、兩個 SH3 domains、 autoinhibitory region (AIR) 和 C-terminal 的 PRR。p47phox 在非活化狀態下,經 由 AIR 將兩個 SH3 domain 的活性部位遮蔽起來;而 SH3 domain 將 PX domain 活性部位遮蔽起來 (Hiroaki et al., 2001)。當細胞受到刺激時,AIR 可經 由多處 Ser 的磷酸化 (S303、S304、S359、S370)改變了 p47phox 的構形,使 SH3 domain 暴露出來與膜上 p22phox 的 PRR 結合 (El Benna et al., 1994; Inanami et al., 1998; Johnson et al., 1998);而 p47phox 的 PRR 則與 p67phox 的 SH3 domain 3.
(14) 結合 (Sumimoto et al., 2005)。另外,在 AIR磷酸化的同時,低濃度的 AA (1-5 μM) 可以幫忙 p47phox 將 PX domain 暴露出來,具有更好的活性。在 cell-free system 或完整的細胞中,高濃度的 AA (50-100 μM) 可直接使 p47phox 與 p22phox 完整 地結合進而活化 NADPH oxidase (Shiose et al., 2000)。PX domain 可與位在膜上 的 phosphoinositides (PI(3,4)P2)或 phospholipids(PA、phosphatidylserine (PS)) 結合. (Karathanassis et al., 2002; Stahelin et al., 2003) 。 另 外 , 在. formyl-methionyl-leucyl-phenylalanine (fMLP) 或 phorbol 12-myristate 13-acetate (PMA)刺激下,許多蛋白質激酶(protein kinase)會對p47phox 磷酸化,包括p38 Mitogen-activated protein kinase (MAPK)和extracellular signal-regulated kinase 1/2 (ERK1/2),還有protein kinase B (PKB/Akt)、protein kinase C (PKC)、p21-activated kinase (PAK)等都會增強NADPH oxidase的活性 (El-Benna et al., 2009)。相反的, protein kinase A (PKA)和casein kinase II 對p47phox磷酸化則有可能造成其活性下 降 (Bengis-Garber et al., 1996; Park et al., 2001)。 p67phox (NOX activator)由 526 個氨基酸構成,其結構上的 N-terminal 具有 四個 tetratricopeptide repeat (TPR) motifs,而 C-terminal 有兩個 SH3 domains 和介於 SH3 domain 間的 Phox and Bem1 (PB1) domain。TPR motifs 可與 Rac 結合, PB1 domain 則會辨識 p40phox 的 PB1 domain (Sumimoto et al., 2005),而 靠近 C-terminal 的 SH3 domain 則與 p47phox 的 PRR 連結。所以 Rac 存在 下,可促進 p67phox 和 gp91phox 的結合。 p67phox 也參與 NADPH 所提供的電 子傳給氧分子。有研究指出,當細胞缺乏 cytochrome b558 時, p47phox 和 p67phox 便無法轉移至膜上。而 p67phox 在沒有 p47phox 時也無法轉移到膜上。但p47phox 在沒有 p67phox 仍然可以轉移到膜上。可能是 p47phox 活化後產生 p67phox 的結 合位置,使 p67phox 轉移至膜上 (Roos et al., 2003)。 p40phox 由 339 個氨基酸構成,其結構在 N-terminal 有 PX domain 和 SH3 domain,而在 C-terminal 有 PB1 domain。p40phox 的功能為促進 p47phox 和 4.
(15) p67phox 在膜上的結合(Kuribayashi et al., 2002)。p40phox 的 PX domain 可與膜 上的 PI(3)P 結合,因此 p67phox 便能藉此與細胞膜連接 (Ellson et al., 2001)。但 目前對 p40phox 功能的了解仍然有限。有些學者認為 p40phox 會活化 NADPH oxidase,但也有些學者認為 p40phox 會抑制 NADPH oxidase (Sathyamoorthy et al., 1997)。 Rac (21 kDa) 是 Rho-family small GTPase 的成員,包含三種 isoforms (Rac1、Rac2和Rac3)。Rac2 主要表現在在嗜中性白血球,而 Rac1 主要表現 在巨噬細胞。Rac2 在活化的過程中為獨立的,並不受 p47phox 或 p67phox 的影 響,並將 C-terminal的 Cys189 上 geranylgeranylation 以便與細胞膜結合。未活 化態的 Rac-GDP 存在細胞質中,且與 guanine nucleotide dissociation inhibitors (GDIs) 結合,抑制 GDP 和 GTP 的交換。當細胞受刺激活化時,Rac-GDP 與 GDI 分開,並在 P-Rex1 (185 kDa) 的調節下, GDP 交換 GTP 而成活化態, 並轉移到膜上與 cytochrome b558 和 p67phox 的 TPR 結合,最後活化 NADPH oxidase (Chuang et al., 1993;Bokoch, 1994;Koga et al., 1999;Sarfstein et al., 2004) 。 而 P-Rex1 為 guanine nucleotide exchange factors (GEFs) , 可 受 到 phosphatidylinositol-3,4,5-trisphosphate (PIP3) 和 Gβγ subunits 的調節。. NADPH oxidae活化的細胞訊息傳遞路徑 已知formylpeptide receptors (FPR)接受刺激後,會經由 phosphoinositide 3-kinase (PI3K)和 phospholipase C (PLC)、PAK、PKC、Rac和MAPK (Montserrat et al., 1992; Stoyanov et al., 1995)訊息傳遞途徑來活化NADPH oxidae (圖1-4)。. FPR. 5.
(16) FPR為 G protein-coupled receptor (GPCR) 與 chemoattractant 結合,會使其 構形發生變化,並可促使 Giα 的 GTP 與 GDP 交換,造成 Gβγ subunits 與 Giαsubunit 分離。當 Giα-GTP 的 GTP 被水解為 GDP,可和 Gβγ subunit 再結 合回到不活化態 (Simon et al., 1991)。表現在人類嗜中性白血球的有 FPR 和 FPRL1 (FPR-like 1)兩種,且 FPR 對 fMLP的親合度較佳。fMLP 刺激細胞所產 生的生物功能,包括趨化作用、去顆粒作用和超氧自由基生成。. PI3K PI3K 為一種 lipid kinase 主要對phosphoinositide 結構上inositol的3'-OH進行 磷酸化。依受質的不同可分三類:class I (IA、IB)、II 和 III。PI3K 都具有 C2 domain 和 catalytic domain ,且都經由 helical domain (PIK domain) 連接。Class I 可作用 於. phosphatidylinositol. (PI) 、 phosphatidylinositol. 4-phosphate. (PI(4)P) 和. phosphatidylinositol 4,5- bisphosphate (PI(4,5)P2);class II 可作用於 PI 和 PI(4)P; class III 可作用於PI (Koyasu, 2003) 。 Class IA PI3K由adaptor subunit 和catalytic subunit 複合體形成的heterodimer。其 中catalytic subunit 有 p110α、β 和δ 三種。p110α 和β 分佈較廣,而 p110δ 主 要表現在白血球。而adaptor subunit 有 p85α、p85β、p55α、p55γ 和 p50α 五種。 其中 p85α 、p85β 皆具有二個 SH2 domain 可與 p110 的N-terminal region 連接, 以維持 p110 的穩定( 圖 1-5)。SH2 domain 可與phospho-tyrosine結合,在tyrosine kinase 活化時可促進 p110 聚集到膜上,造成 p110 的活化 (Wymann and Pirola, 1998)。Class IB PI3K只有一種,由 p110γ catalytic subunit 和p101 adaptor subunit 形 成heterodimer。Class IB 只存在哺乳動物,主要表現在白血球中。Class IB 主要受到 GPCR 所調節,Gβγ 作用到 p101 並活化 p110γ (Furman et al., 1998; Vanhaesebroek and Waterfield, 1999) 。Class IA 或 IB 皆具有 Ras-binding domain ,因此活化態的 6.
(17) Ras-GTP 也會活化 Class IA 或 IB (圖1-5)。Class II 及 class III 的活化機轉尚未完 全釐清 (Koyasu, 2003)。 活 化 PI3K 生 成 的 PI(3,4)P2 或 PI(3,4,5)P3 , 會 誘 使 下 游 蛋 白 激 酶 phosphoinositide-dependent kinase (PDK) 和 Akt利用其結構上的PH domain 與之 結 合 , 由 細 胞 質 中 轉 移 至 細 胞 膜 上 , 方 便 PDK 對 Akt 的 磷 酸 化 作 用 (Vanhaesebroeck and Alessi, 2000;Koyasu, 2003) (圖1-7)。此外,Gβγ亦可經由促進 Shc adaptor protein 的活化及與growth-factor-receptor-bound protein 3 (Grb3)、Sos 形成複合物去活化Ras,進而活化p85 (Koyasu, 2003; Wymann, 2003)。PI(3,4,5)P3和 PI(3,4)P2,也可分別與p40phox和p47phox的PX domain結合 (Kanai et al., 2001)。 PI3K 與Akt 可藉由影響actin polymerization調控嗜中性白血球的趨化作用(Inoue and Meyer, 2008)。 PDK (63 kD) 為 serine/threonine kinase,有 PDK1 和 PDK2 兩種。PDK1 在 結構上 N-terminal 有 catalytic domain,C-terminal 有 PH domain ,可對 Akt1的 Thr308磷酸化並活化Akt1 (Alessi et al., 1997)。有研究指出,人類的PDK1在Ser241 的磷酸化會促進其活性 (Casamayor et al., 1999)。 Akt (57-60 kDa) 亦為 serine/threonine kinase。Akt 參與細胞生長、存活和轉 錄因子調控。Akt 有三種 isoforms 即 PKBα/Akt1 、PKBβ/Akt2 和 PKBγ/Akt3 (圖. 1-6)。Akt的 N-terminal 有 PH domain,中間為 kinase domain (又稱 activation loop), C-terminal 則是 regulatory domain (又稱 hydrophobic motif, HM)。Akt1的 活化,包含由PDK1對kinase domain上的Thr308進行磷酸化,而PDK2則會對HM 上 的Ser473進行磷酸化 (Toker and Newton, 2000)。至於PDK2為何,至今仍眾說紛紜。 有 文 獻 指 出 , 嗜 中 性 白 血 球 的 PDK2 可 能 是 MAPK-activated protein kinase-2 (MAPKAPK2或MK2)。另外,有學者提出三種方式會對 Ser473磷酸化:(1) PDK1 (2) PDK2 (3) Akt 自體磷酸化 (Rane et al., 2001)。且 Akt可直接磷酸化 p47phox 而參與 NADPH oxidase 的活化 (Chen et al., 2003) 。 7.
(18) PAK PAK為serine/threonine kinase,根據蛋白結構與活性調節方式可分為二個族 群:第一類包括PAK1、PAK2和PAK3,而第二類為PAK4、PAK5和PAK6 (圖1-8)。 第一類 PAK的N-terminal 有p21-binidng domain (PBD) (又稱Cdc42/Rac-interactin binding domain 或CRIB) 和autoinhibitory domain (AID)為重疊部分,PBD可與活 化態的Rac和Cdc42 GTPase結合 (Manser et al., 1995;Knaus et al., 1998;Mira et al., 2000;Jaffer and Chernoff, 2002)。第一類 PAK的N端含有2個 SH3-binding motif 和1個非典型的 SH3-binding site,此2個 SH3-binding site 可與 Nck 和 Grb2 結合,非典型 SH3-binding site 則可與 Pak-interacting exchange factor (PIX) 結 合 (Bokoch et al., 1996; Manser et al., 1998; Puto et al., 2003)。第一類 PAK的 C-terminal有 catalytic domain可對下游 p47phox、p67 phox蛋白分子進行磷酸化作用 (Ahmed et al, 1998; Martyn et al., 2005)。第二類 PAK 則缺乏N-terminal的 AID和 SH3-binding site,所以其活性調控的方式與第一類 PAK不相同。第二類 PAK可 與Cdc42結合,但是不與Rac結合,且與GTPase結合後並不會増加其活性 (Abo et al., 1998; Cotteret and Chernoff, 2006)。第一類的PAK對於嗜中性白血球超氧自由 基生成、趨化作用以及吞噬作用有關(Bokoch, 2003;Martyn et al., 2005)。 不活化狀態的PAK以dimer的形態存在於細胞質中。PAK的活化過程 ( 圖 1-9) ,當small GTPase 與 PBD結合後,PAK構形受改變,使其AID與catalytic domain分開,促使activation loop 的autophosphorylation,進而增加 PAK 的活性 (Zenke et al., 1999; Lei et al., 2000)。第一類 PAK 的 activation loop 中,在Thr423 的磷酸化對於PAK的活化極為重要 (Gatti et al., 1999)。另外,Ser144、Ser199 和 Ser204的磷酸化亦可促進PAK的活性 (Gatti et al., 1999; Chong et al., 2001)。Rac 可藉由PAK影響p47phox 磷酸化作用 (Knaus et al., 1995)。此外,PAK可經由促進 8.
(19) 其下游蛋白的磷酸化,例如 myosin light chain kinase、LIM kinase,進而影響細 胞的趨化作用 (Daniels and Bokoch, 1999; Bokoch, 2003)。. PKC PKC為serine/threonine kinase,屬於 AGC kinase family當中一員到目前共發 現 11 種 異 構 物 。. PKC 結 構 的 N-terminal 是. regulatory region ( 含 有. pseudosubstrate、C1與C2 domain),C-terminal為catalytic region (含有C3 與 C4 domain) , 兩 者 間 經 由 一 個 hinge bond 連 接 。 其 中 , pseudosubstrate 序 列 有 autoinhibition的作用,C1 domain 為DAG 和 PMA的結合位置 (Bell and Burns, 1991);C2 domain 為鈣離子和 PS 的結合位置 (Igarashi et al., 1995);C3 domain 為 ATP 結合的位置;C4 domain 為與受質結合的位置 (Silinsky and Searl, 2003)。依 regulatory region結構上的不同可將PKC分為classical PKC (cPKC) 、 novel PKC (nPKC)和atypical PKC (aPKC) 三個族群。cPKC包括PKC-α、-βI、-βII 和 -γ,於 regulatory region具 pseudosubstrate、C1和C2 domain,需要PS、DAG 和 鈣離子的存在下才可以活化。nPKC包括 PKC-δ、-ε、-η 和-θ,在regulatory region 具有 pseudosubstrate和C1 domain,需要PS、DAG,但不需要鈣離子即可活化。 aPKC包括 PKC-ζ、-τ 和-ι/λ,其regulatory region 只有 pseudosubstrate domain、 nuclear localization signals 和 nuclear export signals , 因 此 不 需 要 鈣 離 子 和 DAG,但可受到磷脂類 (PS、PA 和 ceramides) 或轉移到細胞核受刺激而活化 (Newton, 2003)。 PKC 成熟至活化的過程:(1) 首先經轉譯作用剛合成尚未磷酸化且未成熟 (immature)的 PKC先停留在細胞質中;(2) PDK1 可使位於PKC catalytic region的 Thr500磷酸化;(3) PKC 進行 autophosphorylation 將本身C-terminal的Thr641 和 Ser660磷酸化,導致構形改變,此時為安定且成熟 (mature) 的 PKC而存在於細 9.
(20) 胞 質 中 (Bornancin and Parker, 1997; Edward et al., 1999) 。 此 構 形 變 化 使 N-terminal的pseudosubstrate 序列和 Thr500結合並將C 端的catalytic region遮 蔽,因而抑制 PKC 的活性 (Gao et al., 2001);(4)當訊息刺激細胞時,細胞膜上 脂質水解產生 DAG和IP3 ,增加細胞內鈣離子的濃度。而使 PKC 藉由 C1 domain 與 DAG 結合以及C2 domain 和鈣離子結合,導致 PKC轉移到細胞膜 並且活化 (Newton, 2003; Di Mari et al., 2005)。PDK1 活化 PKC的過程中,並不 需要 PI3K 的參與,因此不論是否具有PH domain 的PDK1都能活化 PKC (Newton, 2003)。 人類嗜中性白球中,具有 PKC-α、-βI、-βII、-δ 和 -ζ 等 isoforms (Majumdar et al., 1993;Kent et al., 1996)。而大鼠的嗜中性白球中具有 PKC-α、-β、-δ、-ε、 -θ、-μ、-ι/λ 和 -ζ,且 PKCλ 和 PKCζ 含量很少 (Tsao and Wang, 1997)。 PKC-α、-βI、-βII、-δ 和 -ζ 皆與呼吸爆發有關。在嗜中性白血球細胞中,cPKC、 PKCδ 和 PKCζ 會對 p47phox 進行磷酸化 (El-Benna et al., 1996;Dang et al., 2001;Yamamori et al., 2004) 增加 NADPH oxidase 的活性 (Regier et al., 1999; Fontayne et al., 2002)。. MAPK. MAPK 為 Ser/Thr kinase。MAPK 可依其訊息傳遞路徑之不同而分成幾類: p38 MAPK (p38α/β/γ/δ 四種 isoforms)、ERK (ERK1/2 二種 isoforms)、c-Jun NH2-terminal kinase (JNK) (JNK1/2/3 三種 isoforms)、ERK3、ERK4 及 ERK5 (Johnson and Lapadat, 2002)。其中,以 p38 MAPK、ERK1/2 與 JNK 活化的機 轉研究最清楚 (Roux and Blenis, 2004)。嗜中性白血球表現有 ERK1/2 (Torres et al., 1993),而 p38 MAPK 以 α 與 δ 兩種 isoforms 為主,β 與 γ 含量極少 (Hale et al., 1999)。 10.
(21) MAPK 活化的過程,主要是一連串的 kinase cascade,即 MAPK kinase kinase (MKKK/MEKK)、MAPK kinase (MKK/MEK) 及 MAPK 三個階段 (Kyriakis and Avruch, 2002) (圖 1-10)。當細胞受到刺激,經由 small G protein 活化 MKKK, 然後對 MKK 進行磷酸化作用。活化的 MKK 會對下游的 MAPK 結構上的 T-X-Y 進行 Tyr/Thr 雙磷酸化作用 (English et al., 1999; Krens et al., 2006)。在 JNK 為 T-P-Y,在 ERK 為 T-E-Y,在 p38 MAPKs 為 T-G-Y。嗜中性白血球受到 fMLP 刺激後,會快速引起 ERK 和 p38 MAPK 磷酸化,並參與 p47phox 的磷酸化 (El-Benna et al., 1996)。此外,在嗜中性白血球中,ERK2 及 p38 MAPK 亦會參 與 fMLP 引起的 p67phox 磷酸化 (Dang et al., 2003)。 在 NADPH oxidase 活化的過程中,有報告指出 p38 MAPK 的角色較 ERK1/2 為更重要 (Rane et al., 1997)。亦有報告指出,ERK1/2 並不參與調節 fMLP 所引起的 NADPH oxidase 活化 (Yu et al., 1995; Zu et al, 1998)。因而,ERK 是否參與調節嗜中性白血球 NADPH oxidase 活化,至今尚無定論。有文獻指出, p38 MAPK 的下游蛋白 MK-2 (Stokoe et al., 1992; Kummer et al., 1997) 為真正影 響 NADPH oxidase 活化的調控因子 (Coxon et al, 2003)。. PLC 哺乳動物細胞中具有五類的 PLC,分別為 PLC-β (1-4)、-γ (1-2)、-δ (1, 3, 4)、 -ζ 及 -ε (Song et al., 2001; Saunders et al., 2002)。PLC 的蛋白結構包括具有 X 及 Y domain 形成一個 catalytic domain、PH domain、EF-hand domain 及 C2 domain (Katan, 1998)。因具有 EF-hand 與 C2 domain,所以需要鈣離子才可以活化 PLC (Rhee, 2001; Wing et al., 2003)。PLC-β、-γ、-δ 因具有 PH domain,所以可結合 到含有 inositol 的磷脂質上。不同的 PLC 與不同的磷脂質結合,PLCβ 可與 PI(3)P 結合,PLCγ 與 PI(3,4,5)P3 結合 (Ferguson et al., 1995),PLCδ 與 PI(4,5)P2 11.
(22) 結合 (Razzini et al., 2000),PLCδ1 可與 IP3 結合,以迴饋調節酵素的活性 (Essen et al., 1996)。PH、SH2 與 SH3 domain 皆可促使 PLC 由細胞質轉移至 細 胞 膜 上 (Rhee, 2001)。 PLCβ 和 PLCγ 分 別 受 細 胞 內 的 Gβγ 和 tyrosine kinases 調控。而 PLCδ 會由鈣離子調控,鈣離子和 PLCδ 的 catalytic domain 結合使其構形產生變化 (Okada et al., 2005)。PLCε 可能是藉由 Ras 來調節 (Fukami, 2002)。 活化的 PLC 可水解細胞膜上的 PI(4,5)P2 生成 IP3 及 DAG, 為主要的二 級 訊 息 傳 遞 因 子 。 DAG 可 活 化 PKC 或 代 謝 成 PA , 進 而 影 響 下 游 訊 息 (Andresen et al., 2002)。IP3 則作用到內質網上的 IP3 receptors,促進鈣離子由內 質網釋出,迅速增加細胞質內鈣離子濃度 (Berridge, 1989; Taylor, 1998)。細胞外 的鈣離子可經由二種機制進入細胞。一為排空內質網中儲存的鈣離子後,可促使 Ca2+-influx factor 擴散至細胞膜上,造成鈣離子通道打開,讓細胞外鈣離子持續 內流 (Randriamampita and Tsien, 1993; Hallett and Pettit, 1997)。二為當排空內質 網中儲存的鈣離子後,原存在於內質網的 STIM1 與位於細胞膜上的 store-operated Ca2+ channel 產生作用,讓鈣離子內流 (Brechard and Tschirhart, 2008)。因此,細 胞內鈣離子濃度增加可分成二個階段,第一階段源於內質網釋放儲存的鈣,第二 個階段則源於細胞外鈣離子內流。. Cyclic AMP Adenylyl cyclase可促進ATP轉換成cyclic AMP。而生成的cyclic AMP可經由 phosphodiesterases水解。細胞內cyclic AMP的含量多寡取決於Adenylyl cyclase及 phosphodiesterases兩種酵素活性之間的平衡。而增加細胞內cyclic AMP的含量, 可抑制嗜中性白血球超氧自由基的生成(Cronstein et al., 1985)。已知fMLP刺激嗜 中性白血球經由釋出adenosine作用到A2 adenosine receptor,來活化adenylyl 12.
(23) cyclase生成cyclic AMP (Spisani et al., 1996)。增加細胞內cyclic AMP會降低Akt的 活性(Kim et al., 2001)。也有報告指出,增加細胞內cyclic AMP會抑制Ca2+進到細 胞裡 (Schudt et al., 1991)。而fMLP刺激嗜中性白血球產生超氧自由基的作用是 需要增加細胞內Ca2+。因此藥物影響細胞內cyclic AMP也會改變超氧自由基的生 成。. 13.
(24) 圖 1-1 PMN 在不同時期顆粒球分泌的物質 (節錄自 Soehnlein et al., 2009) 14.
(25) (1). (2). 圖 1-2 (1) NADPH oxidase 組成結構的簡圖(節錄自 Takeya and Sumimoto, 2003) (2) NADPH oxidase組成結構的詳圖(節錄自 Dale et al., 2008) 15.
(26) C. 圖1-3 (A) gp91phox /Nox2的結構 (B) Bis-heme 嵌在 gp91phox中 (C) cytochrome b558 的結構 (節錄自Sumimoto, 2008). 16.
(27) 圖1-4 FPR在化學刺激物下的傳遞路徑 (節錄自Rabiet et al., 2007) 17.
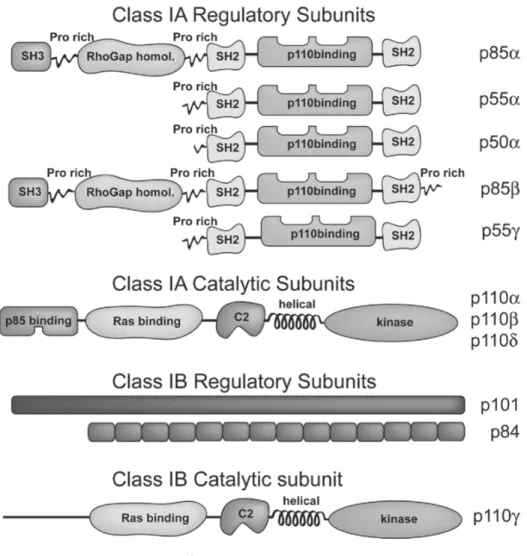
(28) 圖 1-5 PI3K class IA 和 IB 的結構 (圖片來源 http://www.biochemsoctrans.org/bst/035/1109/bst0351109f01.htm). 18.
(29) 圖 1-6 Akt 的結構(節錄自 Nicholson and Anderson, 2002). 19.
(30) 圖 1-7 Akt 活化機制 (節錄自 Nicholson and Anderson, 2002). 20.
(31) 圖1-8 PAK的種類和基本結構 (節錄自 Arias-Romero and Chernoff, 2008). 21.
(32) 圖1-9 (A) PAK 基本結構. (B) PAK活化過程 (節錄自 Chong et al., 2001). 22.
(33) 圖1-10 MAPK的活化機制 (節錄自Johnson and Lapadat, 2002). 23.
(34) 第二章. 研究動機與目的. 嗜中性白血球為免疫機制的重要防線,利用生成的超氧自由基可殺死入侵的 病源菌。但生成過多的超氧自由基也會造成組織、器官的損傷。因此抑制超氧自 由基的生成,有助於改善嗜中性白血球過度活化所引起的病變。但在臨床上缺乏 此類的抗發炎藥物。研究嗜中性白血球活化生成超氧自由基的機制,有助於研發 抑制嗜中性白血球活化的抗發炎藥物。參與嗜中性白血球生成超氧自由基的細胞 內訊息傳遞機制非常複雜,目前的了解仍然有限。對特定訊息傳遞途徑有專一作 用的工具藥不足,在機制探討上也受限。經初步篩選結果發現新合成化合物TCH 可抑制嗜中性白血球生成超氧自由基。本論文在探討TCH抑制嗜中性白血球細胞 生成超氧自由基的作用機制,如對特定訊息傳遞路徑具有專一性的抑制效果,可 作為藥理研究的工具藥。並期待能發展成為因過量超氧自由基造成疾病的治療用 藥。. 24.
(35) 第三章 實驗部分. (一) 實驗材料 Hanks ' balanced salt solution (HBSS) 購 自 Invitrogen (Carlsbad, CA, USA)。Anti-RhoA (sc-418)、anti-p110γ(sc-1404)、anti-phospho-Akt1/2/3 (Ser473) (sc-7985)、anti-Akt1/2/3 (sc-8312)、anti-p22phox (sc-2078)、anti-p47phox (sc-17845 for IP)、anti-gp91phox (sc-20782 for IP)、anti-ERK2 (sc-154)、anti-p38 MAPK (sc-7149)、 anti-PKCα(sc-208)、anti-PKCβI (sc-209)、anti-PKCβII (sc-210)、anti-PKCδ (sc-213) anti-PKC ζ (sc-216) 、 anti-vav (sc-132) 與 anti-phospho-vav(Tyr174) (sc-16408-R) 等抗體皆購自Santa Cruz Biotechnology (Santa Cruz, CA, USA)。 Anti-phospho-PAK1 (Thr423)/PAK2 (Thr402) (#2601) 、 anti-PAK1 (#2602), anti-phospho-Akt (Thr308) (#9275) 、 anti-phospho-PDK1 (Ser241) (#3602) 、 anti-phospho-p38 MAPK (Thr180/Tyr182) (#9211)、anti-phospho-MK-2 (Thr222) (#3044)、anti-phospho-GSK3-α/β(Ser21/9) (#9331) 與GSK3 fusion protein (#9237) 等抗體皆購自 Cell Signaling Technology (Beverly, MA, USA)。Anti-phosphoserine (4H4). (#ALX-804-165-C100) 與 (2'Z,3'E)-6-bromoindirubin-3'-oxime. (6BIO). (#ALX-430-156-M001)購自 Alexis (Carlsbad, CA, USA)。Polyvinylidene difluoride 轉漬膜、Immunoblot Western Chemiluminescent HRP Substrate 與anti-gp91phox (#07-024)、anti-p67phox (#07-502)、anti-p47phox (#07-500)、anti-Rac2 (#07-604) 等 抗體皆購自Millipore (Bedford, MA, USA)。Anti-Rac1 (#R56220) 抗體購自BD Biosciences (Palo Alto, CA, USA)。Cyclic AMP 購自Cayman Chemical (Ann Arbor, MI, USA)。Akt/PKB kinase activity assay kit (#EKS-400A)和 PKC kinase activity assay kit (#EKS-420A)購自Assay Designs (Cambridge, UK)。PDK1 kinase activity 25.
(36) assay kit (#CY-1180)購自MBL International。PAK-p21-binding domain (PBD) protein agarose beads (#RT02) 購自Cytoskeleton Inc. (Denver, CO, USA)。Active PDK1 (#P14-10H-05) 和active Akt1 (#P16-10G-05) 購自SignalChem。Dextran 500、 Ficoll-Paque 及 protein A sepharose (#17-0780-01) 購自GE Healthcare (Piscataway, NJ, USA)。二級抗體購自Jackson ImmunoResearch Laboratories (West Grove, PA, USA)。2-(4- Morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY 294002) 購 自 Biomol. Research. 4-(4-Fluorophenyl)-. Laboratories. (Plymouth. Meeting,. PA,. 2-(4-methylsulfinylphenyl)-5-(4-pyridyl)-1H-imidazole. 203580). (SB 及. 2-[1-(3-dimethylaminopropyl)-1H-indol-3-yl]-3-(1H-indol-3-yl)-maleimide 109203X). USA) 。. 購. 自. Merck. (Taipei,. Taiwan). 。. (GF. 1,4-Diamino-. 2,3-dicyano-1,4-bis(2-aminophenylthio) butadiene (U0126) 購自Promega (Madison, WI, USA)。其他的化學藥品皆購自Sigma-Aldrich (St. Louis, MO, USA)。化學藥 品溶於DMSO中,DMSO在培養液中的濃度小於0.1% (v/v)。. (二) 分離大鼠的嗜中性白血球 以腹腔注射麻醉劑 pentobarbital (60 mg/kg) 麻醉大鼠 (Sprague Dawley, 300-350 g) 。 待 大 鼠 昏 迷 後 , 由 其 腹 腔 動 脈 抽 取 全 血 且 與 含 有 100 mM ethylenediaminetetraacetic acid (EDTA) 的針筒混合。再將血液與 5% dextran 混 合後靜置。待大部分的紅血球沉降後,收集試管上層的血液。收集的血液用 Ficoll-PaqueTM PLUS離心 400 g,20℃, 30 分鐘,以分離白血球 (Wang et al., 1994)。在試管底層的嗜中性白血球懸浮在HBSS其含有 10 mM HEPES (pH 7.3) 及 4 mM NaHCO3 中離心 (800 g,4℃) 10 分鐘。將懸浮細胞先以低張溶液 (0.05% (w/v) NaCl)打破殘餘的紅血球,再以等體積的高張溶液 (1.75% (w/v) 26.
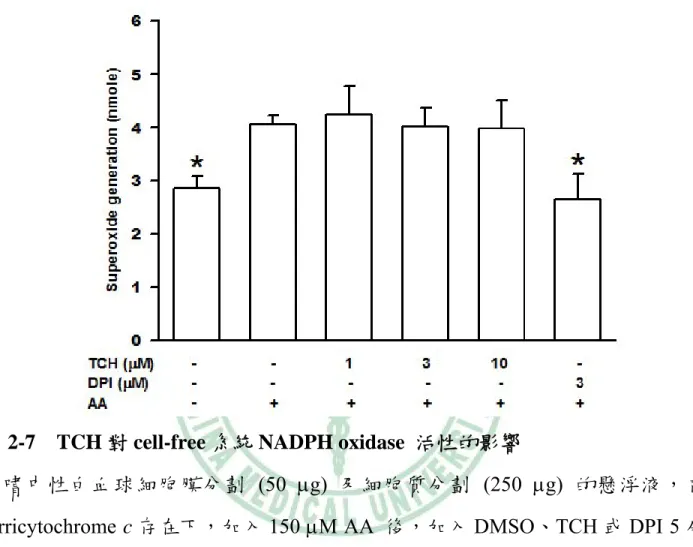
(37) NaCl含 0.25% (w/v) bovine serum albumin) 調回等張。可製備純度及存活率皆≧ 95% 的嗜中性白血球懸浮液。. (三)測量 TCH 對細胞之存活率. 利用 fluorescein diacetate (FDA;測量存活之細胞)染色的方式來檢驗 TCH 是 否對細胞存活率造成影響,以確保 TCH 抑制超氧自由基的生成並非因細胞傷害 或死亡所造成的。 嗜中性白血球懸浮於 HBSS 溶液中(1 × 106 cells/0.5 ml),於 37°C 下預熱 3 分鐘,加入 DMSO (control) 或 TCH 反應 15 分鐘後,以五倍體積 HBSS 稀釋。 離心 (500 g,4°C) 5 分鐘,再以 phosphate-buffered saline (PBS)清洗一次,將細 胞懸浮於 50 μl 的 0.1% (w/v) bovine serum albumin-PBS 中 block 30 分鐘,再加入 FDA (10 μg/ml)於室溫下進行染色 30 分鐘,以 PBS 清洗三次,懸浮於 400 μl 1% (v/v) parafromaldehyde-PBS,以 Flow cytometry 進行偵測。. (四)測量 TCH對嗜中性白血球產生超氧自由基的作用 嗜中性白血球懸浮液 (2 × 106 cells/0.4 ml HBSS) 先與 cytochrome c (0.5 mg/ml) 、cytochalasin B (5 μg/ml) 和 CaCl2 (1 mM) 於試管中混合,置於 37℃中 預熱 3 分鐘。在加入不同濃度的 DMSO、TCH 或 3 μM diphenylene iodonium (DPI) 作用五分鐘後,以 1 μM fMLP 刺激細胞 10 分鐘,加入 100 U/ml superoxide dismutase (SOD)中止反應。另外含有 100 U/ml SOD 的細胞懸浮液當 作背景值 (blank)。將反應後的細胞懸浮液,在離心去除細胞後,將上清液吸取 300 μl 至 96 孔盤中,以 550 nm 波長讀取吸光值 (Wang et al., 2003),並計算 TCH 對超氧自由基抑制的程度。 27.
(38) (五)測量 TCH 對超氧自由基的清除作用 測量 dihydroxyfumaric acid (DHF) 自發性氧化反應。試管中含 0.891 mM DHF 及 0.274 mM nitroblue tetrazolium (NBT),且以含有 60 U/ml 的SOD作為 對照組。在加入 TCH 室溫下反應 15 分鐘後,將溶液轉移至 96 孔盤,讀取 560 nm吸光值的變化 (Goldberg and Stern, 1977)。. (六)測量 TCH 對 xanthine-xanthine oxidase 產生超氧自由基的影響 將 phosphate buffer (50 mM Na2HPO4 (pH 7.4)、0.1 mM EDTA)中加入 20 mU/ml xanthine oxidase、DMSO (control)或 TCH、ferricytochrome c在37℃下培 養5分鐘,以 0.15 mM xanthine 開啟反應,以分光光譜儀 550 nm 連續讀取 4 分鐘吸光值的變化 (Selloum, et al., 2001; Wang et al., 1994)。. (七)TCH影響細胞生成超氧自由基之可逆性 實驗組嗜中性白血球懸浮液 (2 × 106 cells/0.4 ml HBSS) 先與 cytochalasin B (5 μg/ml) 及 CaCl2 (1 mM) 於試管中混合,並置於 37℃中預熱 3 分鐘。加 入 TCH或DMSO作用5分鐘,加入0.8 ml HBSS 稀釋,以HBSS清洗兩次。而對 照組細胞懸浮液與 cytochrome c (0.5 mg/ml) 、cytochalasin B (5 μg/ml) 和 CaCl2 (1 mM) 於試管中混合,置於37℃中預熱 3 分鐘。加入TCH或DMSO作用5分鐘。 兩組細胞懸浮液以 1 μM fMLP 刺激細胞 10 分鐘,加入 100 U/ml SOD中止反 應。另外以含有 100 U/ml SOD的細胞懸浮液當作背景值 (blank)。離心取上清液 300 μl至96孔盤中,以550 nm讀取吸光值。 28.
(39) (八)測量 TCH 對 NADPH oxidase 活性的影響 含嗜中性白血球細胞膜分劃 (50 μg 蛋白質的量) 及細胞質分劃 (250 μg 蛋 白質的量) 的懸浮液,在 ferricytochrome c 存在下,加入 150 μM arachidonic acid (AA)後,再加入 DMSO (control)、TCH 或 3 μM DPI 共浴 3 分鐘,以 200 μM NADPH 開啟反應。利用分光光譜儀在 550 nm 下,記錄 10 分鐘的吸光值的變化。 超氧自由基產量的計算方式:superoxide anion (nmol) = ΔA × 47.4 × 1.5 ml。. (九)免疫沈澱分析 嗜中性白血球(2 × 107 cells/0.5 ml HBSS)置於 37°C中預熱 3 分鐘,加入藥 物反應 5 分鐘後,以1 μM fMLP刺激。細胞反應後加入冰HBSS減緩反應,離心 (800 g,4℃) 2 分鐘。將細胞溶解於MLB buffer (25 mM HEPES pH 7.5、150 mM NaCl、1% Igepal CA-630、10 mM MgCl2、1 mM EDTA、2% glycerol、1 mM NaF、 1 mM Na3VO4、1 mM phenylmethylsulfonyl fluoride與10 μg/ml aprotinin、pepstatin A、leupeptin 及 antipain) ,靜置於冰上10分鐘。待離心(1,000 g,4°C) 10分鐘後, 收集上清液。 取 500 μg 上清液蛋白質與 anti-p47phox 抗體於 4 °C 下翻轉混合反應過夜。隔 日加入 protein A separose beads 於 4°C 下翻轉混合反應 2 小時。Sepharose 再以 MLB buffer 清洗二次,加入 5 x Laemmli sample buffer,煮沸 15 分鐘。蛋白質以 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) 展開, 並轉移至 polyvinylidene difluoride (PVDF) 膜上。以含 5% 脫脂奶粉的 TBST buffer (10 mM Tris-HCl (pH 7.5) 、150 mM NaCl 和 0.1% Tween 20)填充後,用 anti-p47phox、anti-phosphoserine、anti-PKC 或 anti-p22phox 抗體偵測。以 Luminescent 29.
(40) Image Analyzer (Fujifilm LAS-3000) 照相,並以 MultiGauge software 分析。. (十)細胞膜轉移作用 嗜中性白血球(2 × 107 cells/0.5 ml HBSS)置於 37°C 中預熱 3 分鐘,加入藥 物反應 5 分鐘後,以 1 μM fMLP 刺激。加入 isolation buffer (0.34 μM sucrose、 10 mM Tris-HCl (pH 7.0)、1 mM phenylmethylsulfonyl fluoride、1 mM NaF、1 mM Na3VO4 及 10 μg/mL leupeptin、antipain、aprotinin 和 pepstatin A)。於冰浴中,以 超音波打破細胞終止反應後離心 (800 g,4℃) 10 分鐘。取上清液進行超高速離 心 (130000 g,4℃) 30 分鐘。取沉澱物為細胞膜分劃。以 Lowry method 定量蛋 白後,10% 的 SDS-PAGE 展開,並轉移至 PVDF 膜上。以含 5% 脫脂奶粉充 填後,用 anti-Akt 及 anti-phospho-PDK1 抗體反應來辨識。同時利用 anti-Gβ抗 體辨識作為 loading control。. (十一)測量蛋白質的磷酸化 在 37℃下,嗜中性白血球 (5 × 106 cells)與藥物反應後,再以 1 μM fMLP 刺激,最後加入 5 x Laemmli sample buffer (250 mM Tris-HCl pH 6.8、50% glycerol、10% SDS) 打破細胞終止其反應。離心並收集上清液,以去除多餘的 殘渣,用 Lowry method 定量蛋白。蛋白質以 10% 的 SDS-PAGE 展開,轉移 至 PVDF 膜上。以含 5% (w/v) 脫脂奶粉的 TBST Buffer (10 mM Tris-HCl (pH 7.5) 、150 mM NaCl 和 0.1% Tween 20) 充填空隙後,用 anti-phospho-p38 MAPK 、anti-phospho-MK2、anti-phospho-Akt (Ser473)、anti-phospho-Akt (Thr308) 或anti-phospho-PAK(PAK1 (Thr423)/PAK2 (Thr402) 抗體辨識。辨識後的 PVDF 膜,利用 stripping buffer (62.5 mM Tris-HCl (pH 6.8)、100 30. mM β-mercaptoethanol.
(41) 和 2% SDS) 在 50℃ 清洗 15 分鐘後,再次以含 5% (w/v) 脫脂奶粉的 TBST Buffer 充填膜後。並以 anti-p38 MAPK、anti-Akt、 anti-PAK1或anti-Vav1抗體 辨識作為loading control。. (十二)測量 TCH 對 Rac2 活化 在 37℃反應後的嗜中性白血球 (2 ×107 cells) 懸浮液,以 5 倍體積的 HBSS 減緩反應後,離心 (500 g,4℃) 5 分鐘。將細胞以 0.5 ml 的 MLB 溶液(10 mM MgCl2、25 mM HEPES、1% Igepal CA-630、150 mM NaCl、1 mM EDTA、2% glycerol)將細胞打破,靜置於冰上 10 分鐘。離心(10000 g,4℃) 10 分鐘,收集 上清液,以 Bradford method 定量蛋白至 400 μg,加入 PAK-GST-beads 在 4℃下, 反應一小時。之後,將殘留的抗體清洗乾淨後,加入 5 x Laemmli sample buffer。 再以西方墨點法偵測 Rac2。利用 total Rac2 細胞 lysate 作為 loading control。. (十三)PDK1 kinase 活性之測量 利用 PDK1 activity assay kit 測量。將DMSO、TCH或3 μM 6BIO先與active PDK1 (5 munit/assay)在30℃反應5分鐘。取 100 μl 反應液,加入 62.5 μM ATP 在30°C反應 30 分鐘,清洗後,加入anti-phospho-AKT (T308) 抗體於室溫反應 30分鐘。清洗後,加入HRP 連結的二次抗體在室溫反應 30分鐘。清洗後,在加 入Substrate Reagent 於室溫反應5分鐘,加入Stop Solution,以450 nm偵測。. (十四)Akt 和 PKC kinase 活性之測量. 嗜中性白血球(4 × 106 cells/0.5 ml HBSS) 與藥物反應後,再以 1 μM fMLP 31.
(42) 刺激。細胞反應液以冰的HBSS停止反應,離心(800 g,4°C) 2 分鐘,移去上清 液。將細胞溶解於於lysis buffer (20 mM MOPS (pH 7.2)、1% NP-40、5 mM EGTA、 2 mM EDTA、50 mM β-glycerophosphate、1 mM dithiothreitol、50 mM NaF、1 mM Na3VO4、1 mM benzamidine、1 mM phenylmethylsulfonyl fluoride、10 μg/ml的 aprotinin、pepstatin A、leupeptin 及 antipain)後,靜置於冰上10分鐘。經離心 (10,000 g,4°C) 10 分鐘後,取上清液以Akt/PKB 或是PKC kinase activity assay kit 進行分析。或細胞經fMLP刺激,取得細胞溶解液。經離心取上清液,與DMSO 或藥物反應後,以Akt/PKB kinase activity assay kit進行分析。或human recombinant active Akt1 (30 ng/assay) 與DMSO、TCH或rottlerin反應後。取 30 μl 反應液,加 入 250 μg/ml ATP在30°C反應60分鐘,然後加入Phosphospecific Substrate 抗體於 室溫反應60分鐘。清洗後,加入HRP連結的二次抗體在室溫反應 30分鐘。清洗 後,在加入TMB Substrate於室溫反應60分鐘後,加入Stop Solution,以450 nm偵 測。. (十五) Cyclic AMP 之測量 嗜中性白血球(2 × 10. 6. cells/0.2 ml HBSS)置於 37°C 中預熱 3 分鐘,加入. DMSO、TCH 或 isoproterenol (3 μM)反應 5 分鐘,以 0.1 M HCl 停止反應,置於 冰上 20 分鍾。以超音波震盪打破細胞,離心(1,000 g,4°C) 10 分鐘,取上清液, 加入 KOH 中和。加入 acetic anhydride 混合均勻,利用 cyclicAMP EIA kit 進行 分析。將反應液加入 tracer 和 antiserum 抗體在室溫反應過夜,在清洗後,加入 Ellman’s Reagent 在室溫反應 1 小時,以 405-420 nm 偵測。. (十六) 統計分析. 32.
(43) 實驗數據以 mean ± S.D.來呈現。兩組數據比較時,以 student t-test 分析差 異。多組數據比較時,則是先以 ANOVA 分析後,由 Bonferroni t-test 來比較差 異。p 值小於 0.05 視為具有顯著差異性。. 33.
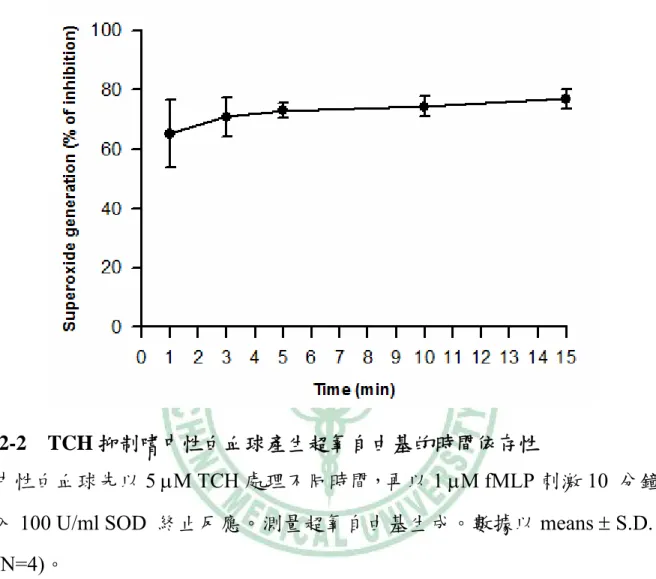
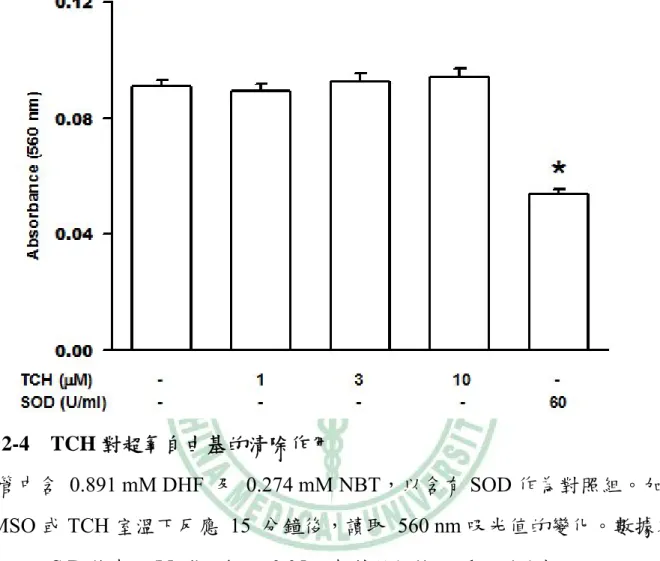
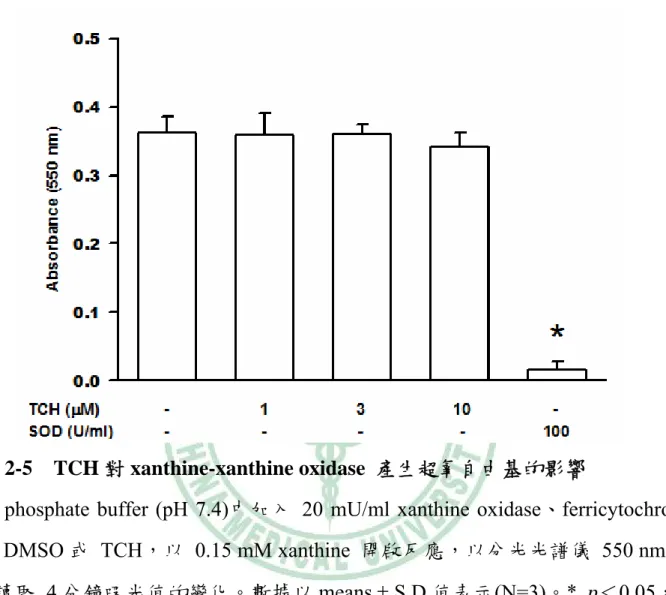
(44) 第四章 結果與討論. 一、TCH 抑制超氧自由基生成. 嗜中性白血球先與不同濃度(1-30 μM)的 TCH 反應 5 分鐘後,以 1 μM fMLP 刺激超氧自由基生成。結果 TCH 可以濃度依存性(concentration-dependent)抑制 fMLP 所誘導的超氧自由基之生成,在 1 μM 時即有顯著性差異,其 IC50 值為 4.5 ± 0.8 μM。而已知 NADPH oxidase 的抑制劑 DPI 在 3 μM 可抑制 80% (圖 2-1)。 而若以 5 μM 的 TCH 先與細胞反應不同的時間,發現其反應時間長短對於其抑 制效果並無大的差異,在 1 分鐘的抑制作用為 65%,至 15 分鐘時的抑制作用為 75% (圖 2-2)。後續的實驗中細胞在刺激前均先與 TCH 反應 5 分鐘。利用 FDA 染色測試 TCH 是否會影響細胞存活,結果顯示分別在 DMSO、10 μM 和 20 μM 的濃度下細胞的存活率仍有 95%、94% 和 93% (圖 2-3),因此排除 TCH 因降低 細胞存活而抑制超氧自由基生成之可能性。 TCH (1-10 μM)對 DHF 的自體氧化(autoxidation)並無顯著影響,因此 TCH 不同於 SOD,不具有超氧自由基清除者的作用(圖 2-4)。另外 xanthine oxidase 與 TCH (1-10 μM)反應 5 分鐘後,以 xanthine 開啟反應,結果也顯示 TCH 不會減 少超氧自由基生成(圖 2-5)。若細胞經 10 μM TCH 處理後,可經由 HBSS 清洗細 胞而使 TCH 喪失抑制超氧自由基生成的能力(圖 2-6)。偵測 TCH 是否直接影響 NADPH oxidase 的活性,可藉由 AA 促進聚集 NADPH oxidase 的細胞質和細胞 膜組成蛋白而引起超氧自由基生成之特性。在 AA 活化 NADPH oxidase 後,再 加入 TCH (1-10 μM),發現 TCH 不會抑制 NADPH oxidase 的活性 (圖 2-7)。相 反地,抑制劑 DPI 則可明顯地抑制 NADPH oxidase 的活性。實驗結果顯示,TCH 34.
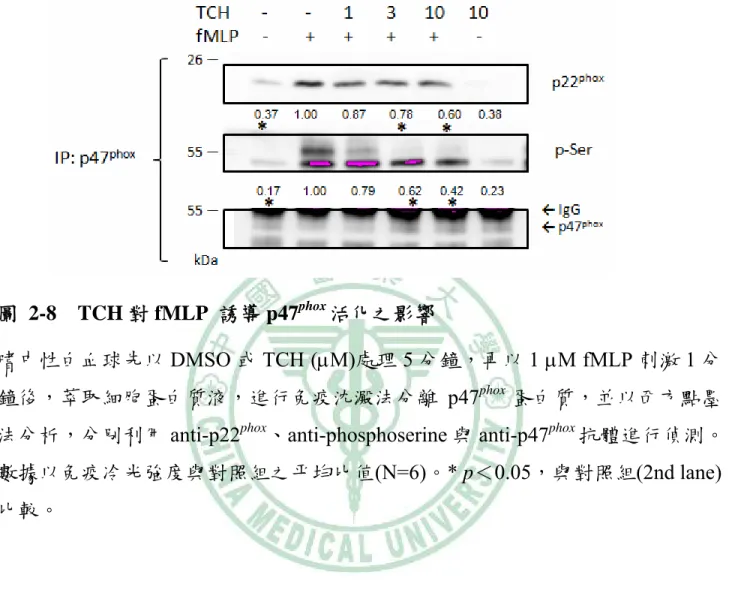
(45) 抑制 fMLP 所誘導的超氧自由基之生成,可能是抑制細胞內訊息傳遞路徑而來。. 二、TCH 對 NADPH oxidase 聚集之影響. 在 NADPH oxidase 活化時,位於細胞質中的組成蛋白 p40phox、p47phox、p67phox 和 Rac2 會轉移至細胞膜與 flavocytochrome b558 (p22phox 和 gp91phox)連結。因此, 利用 p47phox 抗體做免疫沈澱分析,顯示 TCH 可抑制 p47phox 與 p22phox 的結合,在 3 μM 和 10 μM TCH 的抑制百分比分別為 59%和 77%,IC50 為 4.5 ± 4.0 μM。另 外,在 p47phox 的 serine 磷酸化可促進 p47phox 和 p22phox 間的連結(Ago et al., 2003)。而 fMLP 可明顯地增加 p47phox 結構上的 serine 磷酸化作用,此作用亦 被 TCH 所抑制,在 3 μM 和 10 μM TCH 分別有 55%和 75%的抑制作用,IC50 為 4.4 ± 3.0 μM (圖 2-8 )。 Rac2 為 NADPH oxidase 活化的重要蛋白質。Rac2 活化時可與 GTP 結合進 而轉移至細胞膜上(Bokoch, 2005)。利用 gp91phox 抗體做免疫沈澱分析,顯示在 3 μM 和 10 μM TCH 分別有 58%和 79%的抑制 Rac2 和 gp91phox 間的連結作用,IC50 為 2.3 ± 1.5 μM。由於 PAK-PBD protein agarose 與 Rac-GTP 結合,可用來偵測 活化的 Rac2。結果顯示 TCH 可抑制 Rac2 的活化 (在 10 μM TCH 時可抑制 69%),IC50 為 9.2 ± 0.6 μM (圖 2-9)。 綜合以上結果,推測 TCH 是影響細胞內訊息傳遞,進而抑制 p47phox 和 p22phox 以及 Rac2 和 gp91phox 間的連結,導致 NADPH oxidase 產生超氧自由基的活性受 抑制。. 三、TCH 對 Akt 活化之影響. 35.
(46) 有文獻指出,在人類嗜中性白血球,PI3K 參與超氧自由基的生成作用 (Didichenko et al., 1996)。PI3Ks 可分成 Class I (IA 與 IB)、II 和 III,目前研究較清 楚的是 Class I (Vanhaesebroeck et al., 2005)。在 fMLP 刺激下,Class IB 當中的 PI3Kγ扮演超氧自由基生成的重要角色(Condliffe et al., 2005)。PI3K 的活化促進 PI(3,4,5)P3 的生成,其可幫助 Akt 和 PDK1 轉移至細胞膜上。而大鼠嗜中性白血 球在 fMLP 的刺激下,5 秒即有明顯的 Akt 和 PDK1 轉移至細胞膜上,在 15 秒 時達到高峰之後隨即下降(圖 2-10)。 在 fMLP 刺激下,Akt 會對 p47phox 磷酸化,增強 NADPH oxidase 的活性 (El-Benna et al., 2009)。在圖 2-11 (1)顯示,fMLP 可明顯地促進 Akt 與 p47phox 間的連結作用,TCH 可濃度依存性地降低此一連結,IC50 為 7.3 ± 1.7 μM。而完 全活化的 Akt 是在 catalytic site 上的 Thr308 和 hydorphobic motif 上的 Ser473 都 被磷酸化。PDK1 可磷酸化 Thr308,而磷酸化 Ser473 是受到 PDK2 的作用。在 人類嗜中性白血球中,MK-2 被認為是 PDK2 (Rane et al., 2001; Burelout et al., 2007)。fMLP 刺激大鼠嗜中性白血球,會促進 Akt Thr308 和 Ser473 的磷酸化。 藉由西方墨點法, TCH 在 3 μM 和 10 μM 抑制 fMLP 所誘導的 Akt (Thr308) 磷 酸化分別為 24%和 40%。而 Akt (Ser473) 的磷酸化,在 10 μM TCH 抑制程度亦 有 51%。PI3K 抑制劑 LY 294002 (10 μM)能有效地抑制 fMLP 所誘導的 Akt 磷酸 化作用(圖 2-11 (2))。因此可推測 TCH 會影響 Akt 的活性。藉由 Akt activty assay kit 偵測 Akt 的活性,發現 TCH 有濃度依存性抑制 Akt 活性,IC50 為 7.9 ± 3.2 μM (圖 2-12 (1))。而在 fMLP 刺激的細胞溶解液中,TCH 無法抑制 Akt 活性(圖 2-12 (2))。此結果可推測,TCH 影響 Akt 活性為抑制其上游路徑。但是,TCH (3 μM 和 10 μM)可抑制 active human recombinant Akt1,抑制程度分別為 22%和 33%, 而 Akt1 活性的抑制劑 rottlerin (20 μM)也有 90%的抑制程度(圖 2-12 (3))。此差異 可能是由於細胞的萃取液中,含有 Akt1/2/3 三種 isoform 的活性,而 TCH 則只. 36.
(47) 抑制 Akt1 的活性,而其他兩種 isoform 的活性也許還存在,有待進一步研究。 此結果顯示 Akt 可能參與 TCH 抑制 p47. phox. 磷酸化。. 四、TCH 對 PDK1 活性之影響. PDK1 為一持續性活化與自體磷酸化之酵素,PDK1 蛋白 Ser241 的磷酸化 對於其活性極為重要(Casamayor et al., 1999)。TCH (5 μM 和 10 μM)可抑制 active human recombinant PDK1,抑制程度分別為 65%和 70% (IC50 為 7.2 ± 2 μM),而 PDK1 活性的抑制劑 3 μM 6BIO 也有 86%的抑制程度(圖 2-13 (1))。當嗜中性白 血球以 fMLP 刺激時,可增加細胞膜上 Akt 和 phospho-PDK1 (Ser241),但是 TCH 不會減少細胞膜上 Akt 與 phospho- PDK1 的含量 (圖 2-11 (3);圖 2-13 (2))。推 測 TCH 可能並不會影響 PI3K 生成 PIP3,因此 Akt 和 phospho-PDK1 仍可轉移到 細胞膜上。此結果顯示 TCH 抑制 Akt 的活化可能來自於抑制 PDK1 的活性。. 五、6BIO 對嗜中性白血球之影響 為了證明 PDK1、Akt 與 p47phox 確實與超氧自由基的生成有關,所以利用已 知 PDK1 活性的抑制劑 6BIO 來探討。經由 PDK1 activty assay kit 方法確認 6BIO 抑制 active human recombinant PDK1 的 IC50 為 0.8 ± 0.5 μM (圖 2-14 (1)),與文 獻相近(Zahler et al., 2007)。以西方墨點法檢測 6BIO 抑制 Akt 磷酸化的程度,顯 示 6BIO (3 μM 和 10 μM) 抑制 Akt Thr308 磷酸化的程度分別為 35%和 60% (IC50 為 9.2 ± 3.5 μM),而 LY 294002 (10 μM)有 79%的抑制程度。而 6BIO (3 μM 和 10 μM)抑制 Akt Ser473 磷酸化的程度分別為 53%和 66% ( IC50 為 6.7 ± 3.6 μM),而 LY 294002 (10 μM)有 99%的抑制程度 (圖 2-14 (2))。至於為何 6BIO 會 抑制 Akt Ser473 的磷酸化?有報告指出 Akt 在 Thr308 磷酸化後,會對 Ser473 37.
(48) 位置自體磷酸化 (Rane et al., 2001;Dong and Liu, 2005) ,或者是 6BIO 的專一性 不佳,也抑制了 PDK2 的活性。在 fMLP 誘導的大鼠嗜中性白血球生成超氧自由 基的實驗中,6BIO (10 μM 和 30 μM)抑制超氧自由基程度分別為 27%和 56% ( IC50 為 22.7 ± 3.6 μM),而 LY 294002 (10 μM)和 DPI (3 μM)分別有 31%和 89% 的抑制程度(圖 2-14 (3))。此結果顯示抑制 PDK1/Akt 訊息傳遞途徑會影響超氧自 由基的生成。. 六、TCH 對 PKC 活化之影響. 大鼠的嗜中性白球中具有 PKC-α、-β、-δ、-ε、-θ、-μ、-ι/λ 和 -ζ,且 PKCλ 和 PKCζ 含量很少 (Tsao and Wang, 1997)。到底是哪一個PKC isoform促進 p47phox的磷酸化作用,至今仍不確定。而在cell-free系統與PKC isoforms基因剔除 (knockout)的HL60細胞與小鼠嗜中性白血球中,發現PKC-α、-β、-δ 與 -ζ 可參 與NADPH oxidase的活化 (Reeves et al., 1999; Dang et al., 2001; Dekker et al., 2000; Brown et al., 2003; Korchak et al., 2007)。PKC活化的最主要特徵是原本位於 細胞質中的PKC會轉移至細胞膜。TCH會抑制PKC-α細胞膜轉移作用,但卻不會 抑制PKC-β、-δ與-ζ的細胞膜轉移作用 (圖2-15 (1))。PKC與p47phox連結有助於對 p47phox磷酸化。TCH會抑制PKC-α、-δ與-ζ和p47phox的連結作用,但是卻不會抑 制PKC-β 和p47phox的連結 (圖2-15 (2))。當中的原因有待進一步探討。 利用 PKC activity assay kit 偵測 TCH 對 fMLP 促進 PKC 活性的影響,顯 示 PKC 活性並不會被 TCH 所抑制。而 PKC 抑制劑 GF 109203x (1 μM)可抑制 fMLP 所引起的 PKC 活性 (圖 2-15 (3))。但由於 PKC activity assay kit 所偵測的 為所有 PKC isoform。因此,究竟影響大鼠嗜中性球產生超氧自由基為哪一個 isoform,需在經由對 PKC-α、-β、-δ與-ζ具專一性的抑制劑加以探討。. 38.
(49) 七、TCH 對 p38 MAPK 和 MK2 磷酸化之影響. MAPKs 包括 ERK、 p38 MAPK 和 JNK 三種。有文獻指出,p38 MAPK 與 ERK 可以促進 p47phox 的磷酸化作用 (El Benna et al., 1996)。但是,以抑制劑探 究 p38 MAPK 和 ERK 在 fMLP 刺激超氧自由基生成的角色,卻發現只有 p38 MAPK 抑制劑 SB 203580 能抑制 fMLP 所引起的超氧自由基之生成作用,而 ERK 抑制劑 U0126 則無法抑制超氧自由基之生成 (Zu et al., 1998; Kuan et al., 2005)。 MAPKs 的活化主要是來自於上游活化激酶一連串的磷酸化。活化 p38 MAPK 的 磷酸化是受到上游激酶 MKK3/6 活化的作用,然後對下游的 MK-2 磷酸化並活 化 (Krens et al., 2006)。本研究顯示,fMLP 可促進 p38 MAPK 與 MK-2 的磷酸 化。SB 203580 能明顯地抑制 p38 MAPK 與 MK-2 的磷酸化作用,而 TCH 的濃 度提高至 10 μM 仍無法抑制 p38 MAPK 與 MK-2 的磷酸化 (圖 2-16)。此結果顯 示 TCH 並沒有抑制 p38 的活性。在人類嗜中性白血球中,MK-2 被認為是 PDK2 (Rane et al., 2001; Burelout et al., 2007),可對 Akt Ser473 進行磷酸化。p38 MAPK 與 MK-2 也被指出會促進 p47phox 的磷酸化作用。但是在大鼠嗜中性白血球中 MK-2 是否扮演 PDK2 的角色,以及 p38 MAPK 與 MK-2 是否可直接對 p47phox 磷 酸化,仍有待進一步的研究。. 八、TCH 對 PAK 磷酸化影響 活化的 Rac 可與 PAK 的 PBD domain 結合,促進 PAK 的活化 (Burbelo et al., 1995)。Rac 可藉由 PAK 活化對 p47phox 磷酸化 (Knaus et al., 1995)。第一類 PAK (包括 PAK1、PAK2 和 PAK3)在 activation loop 結構上的 Thr423 磷酸化,對於 PAK 的活化極為重要 (Gatti et al., 1999)。TCH (10 μM)抑制 PAK1 和 PAK2 磷酸. 39.
(50) 化分別為 62%和 63% (IC50 值分別為 3.8 和 4.0 μM) (圖 2-17)。TCH 是否影響 PAK 的活性有待進一步的研究。. 九、TCH 對 Vav 磷酸化影響 Vav 為 Rho-GEF 有三種 isoform (Vav1/2/3)。Vav 可經由 tyrosine 的磷酸化調 節活性。在嗜中性白血球中,Vav 對 Rac 來說較為重要 (Kim et al., 2003)。TCH (3 μM 和 10 μM)抑制 Vav (Tyr174)磷酸化分別為 47%和 68% (IC50 為 4.3 ± 3.0 μM),而 Src kinase family 抑制劑 PP1 (10 μM) 有 94%的抑制程度 (圖 2-18)。以 結果顯示 TCH 可能經由抑制 Vav 磷酸化來降低 Rac 的活化,進而抑制 PAK 的 活化。所以 Vav/Rac/PAK 傳遞途徑可能參與 TCH 抑制 p47phox 磷酸化的作用。. 十、TCH 對細胞內 cyclic AMP 濃度之影響 已知增加細胞內 cyclic AMP 會抑制 fMLP 刺激嗜中性白血球超氧自由基的 生成(Cronstein et al., 1985)。TCH 不會影響嗜中性白血球細胞內 cyclic AMP 的含 量。而已知的β-adrenoceptor 促進劑 isoproterenol (3 μM)可大量促進細胞內 cyclic AMP 的生成 (圖 2-19)。此結果顯示 TCH 不會經由增加 cyclic AMP 來抑制嗜中 性白血球生成超氧自由基。. 40.
(51) 圖表(結果). 圖 2-1 TCH 抑制嗜中性白血球產生超氧自由基的濃度依存性 嗜中性白血球懸浮液 (2 × 106 cells/0.4 ml HBSS) 先與 cytochrome c (0.5 mg/ml) 、cytochalasin B (5 μg/ml) 和 CaCl2 (1 mM) 於試管中混合。在加入 DMSO、TCH 或 DPI 作用 5 分鐘後,以 1 μM fMLP 刺激細胞 10 分鐘,加入 100 U/ml SOD 中止反應。測量超氧自由基生成。數據以 mean ± S.D.表示(N=5)。 *, p<0.05 與對照組(2nd column)比較。. 41.
(52) 圖 2-2. TCH 抑制嗜中性白血球產生超氧自由基的時間依存性. 嗜中性白血球先以 5 μM TCH 處理不同時間,再以 1 μM fMLP 刺激 10 分鐘後, 加入 100 U/ml SOD 終止反應。測量超氧自由基生成。數據以 means ± S.D.值表 示(N=4)。. 42.
(53) 圖 2-3 TCH 對細胞存活率的影響 在嗜中性白血球懸浮液(1 × 106 cells/0.5 ml)中加入 DMSO 或 TCH 處理 15 分鐘。 細胞以 FDA 進行染色,再以 Flow cytometry 偵測螢光值之變化。細胞存活率是 以 FDA positive 之細胞數與所有細胞數比值來計算。數據以 means ± S.D.值表示 (N=3)。. 43.
(54) 圖 2-4 TCH 對超氧自由基的清除作用 試管中含 0.891 mM DHF 及 0.274 mM NBT,以含有 SOD 作為對照組。加入 DMSO 或 TCH 室溫下反應 15 分鐘後,讀取 560 nm 吸光值的變化。數據以 means ± S.D.值表示(N=4)。* p<0.05,與對照組(1st column)比較。. 44.
(55) 圖 2-5 TCH 對 xanthine-xanthine oxidase 產生超氧自由基的影響 將 phosphate buffer (pH 7.4)中加入 20 mU/ml xanthine oxidase、ferricytochrome c、DMSO 或 TCH,以 0.15 mM xanthine 開啟反應,以分光光譜儀 550 nm 連 續讀取 4 分鐘吸光值的變化。數據以 means ± S.D.值表示(N=3)。*, p<0.05,與 對照組(1st column)比較。. 45.
(56) 圖 2-6 TCH 抑制 fMLP 刺激細胞產生超氧自由基之可逆性 實驗組嗜中性白血球懸浮液 (2 × 106 cells/0.4 ml HBSS) 先與TCH或DMSO作用 5分鐘,以HBSS清洗兩次。而對照組細胞懸浮液與TCH或DMSO作用5分鐘。兩 組細胞懸浮液以 1 μM fMLP 刺激細胞 10 分鐘,加入 100 U/ml SOD中止反 應。測量超氧自由基生成。數據以mean ± S.D.表示(N=3)。*, p<0.05。. 46.
(57) 圖 2-7 TCH 對 cell-free 系統 NADPH oxidase 活性的影響 含嗜中性白血球細胞膜分劃 (50 μg) 及細胞質分劃 (250 μg) 的懸浮液,在 ferricytochrome c 存在下,加入 150 μM AA 後,加入 DMSO、TCH 或 DPI 5 分 鐘,以 200 μM NADPH 開啟反應。測量超氧自由基生成。數據以 means ± S.D. 值表示(N=4)。* p<0.05,與對照組比較(2nd column)。. 47.
(58) 圖 2-8 TCH 對 fMLP 誘導 p47phox 活化之影響 嗜中性白血球先以 DMSO 或 TCH (μM)處理 5 分鐘,再以 1 μM fMLP 刺激 1 分 鐘後,萃取細胞蛋白質液,進行免疫沈澱法分離 p47phox 蛋白質,並以西方點墨 法分析,分別利用 anti-p22phox、anti-phosphoserine 與 anti-p47phox 抗體進行偵測。 數據以免疫冷光強度與對照組之平均比值(N=6)。* p<0.05,與對照組(2nd lane) 比較。. 48.
(59) (1). (2). 圖 2-9 TCH 對 fMLP 誘導 Rac2 活化之影響 嗜中性白血球先藥物處理 5 分鐘後,(1)以 fMLP 刺激 1 分鐘,萃取細胞蛋白質 液,進行 anti-gp91phox 抗體免疫沈澱法,並以西方點墨法分析。數據以免疫冷光 強度與對照組之平均比值(N=4)。* p<0.05,與對照組(2nd lane)比較;(2) 以 fMLP 刺激 30 秒,萃取細胞蛋白質液,加入 PAK-PBD protein agarose beads 在 4℃下, 反應 1 小時。再以西方墨點法偵測 Rac2,且利用 total Rac2 細胞 lysate 作為 loading control。數據以免疫冷光強度與對照組之平均比值(N=4)。* p<0.05,與對照組 (2nd lane)比較。. 49.
(60) 圖 2-10 fMLP 誘導 Akt 和 PDK1 細胞膜轉位的作用 嗜中性白血球以 DMSO 或 1 μM fMLP 刺激不同時間。以超音波打破細胞終止反 應,取上清液進行超高速離心。細胞膜分劃以西方墨點法偵測 Akt 和 PDK1。以 anti-Gβ 抗體辨識作為 loading control 。數據以免疫冷光強度與對照組(1st column) 之平均比值表示(N=3)。. 50.
(61) (1). (2). (3). 圖 2-11 TCH 對 Akt 磷酸化、細胞膜轉位以及與 p47phox 連結的影響 (1)嗜中性白血球先以 DMSO 或 TCH (μM)處理 5 分鐘後,再以 1 μM fMLP 刺激 30 秒,進行 anti-Akt 抗體免疫沈澱法,並以西方點墨法分析。數據以免疫冷光 強度與對照組之平均比值(N=3)。* p<0.05,與對照組(2nd lane)比較。(2) 細胞 以 TCH 或 LY 294002 處理 10 分鐘,再以 fMLP 刺激 1 分鐘,並以西方點墨法偵 測 phospho-Akt (Ser473)、phospho-Akt (Thr308)和 Akt。數據以免疫冷光強度與 對照組之平均比值(N=4)。* p<0.05,與對照組(2nd lane)比較。(3) 細胞以 TCH 處理 5 分鐘,再以 1 μM fMLP 刺激 15 秒。分離細胞膜分劃,以西方點墨法偵測 Akt。以 anti-Gβ抗體辨識作為 loading control。數據以免疫冷光強度與對照組之 平均比值(N=4)。* p<0.05,與對照組(2nd lane)比較。. 51.
(62) (1). (2). (3). 圖 2-12 TCH對Akt活化的影響 (1) 嗜中性白血球以 DMSO 或 TCH 處理 5 分鐘後,再以 1 μM fMLP 刺激 1 分鍾, 萃取細胞蛋白質液,以 Akt/PKB kinase activity assay kit 分析酵素活性。數據以 means ± S.D.值表示(N=5)。 * p<0.05,與對照組(2nd column)比較。(2)細胞以 DMSO 或 fMLP 刺激 1 分鐘。細胞蛋白萃取液以 DMSO 或 TCH 處理 5 分鐘,利 用 Akt/PKB kinase activity assay kit 分析酵素活性(N=3)。(3) 在 30℃下,active human recombinant Akt1 以 TCH 處理 5 分鐘或 rottlerin 處理 10 分鐘,以 Akt/PKB kinase activity assay kit 偵測其活性。數據以 means ± S.D.值表示(N=3)。* p< 0.05,與對照組(1st column)比較。. 52.
(63) (1). (2). 圖 2-13 TCH 對 PDK1 活化的影響 (1) Active human recombinant PDK1 以 TCH 或 3 μM 6BIO 在 30℃下處理 5 分 鐘,再以 PDK1 kinase activity assay kit 偵測活性。數據以 means ± S.D.值表示 (N=4)。* p<0.05,與對照組(1st column)比較。(2) 嗜中性白血球以 TCH 前處理 5 分鐘,再以 1 μM fMLP 刺激 15 秒。分離細胞膜分劃,以西方點墨法偵測 phospho-PDK1。同時利用 anti-Gβ抗體辨識作為 loading control。數據以免疫冷光 強度與對照組之平均比值(N=4)。* p<0.05,與對照組(2nd lane)比較。. 53.
(64) (1). (2). (3). 圖 2-14 6BIO對嗜中性白血球產生超氧自由基、Akt和PDK1活化的影響 (1) Active human recombinant PDK1以6BIO在 30℃下處理5分鐘,再以PDK1 kinase activity assay kit 偵測活性。數據以means ± S.D.值表示(N=2)。(2) 嗜中性 白血球以6BIO處理5分鐘或LY 294002處理10分鐘,再以 1 μM fMLP刺激1分 鐘。以西方點墨法偵測phospho-Akt (Ser473) 、phospho-Akt (Thr308) 和Akt。數 據以免疫冷光強度與對照組之平均比值(N=4)。* p<0.05,與對照組(2nd lane)比 較。(3)嗜中性白血球懸浮液以6BIO、DMSO、DPI 或 LY 294002作用5分鐘後, 以fMLP 刺激細胞 10 分鐘。測量超氧自由基生成。數據以mean ± S.D.表示 (N=3)。*, p<0.05,與對照組 (2nd column) 比較。. 54.
(65) (1). (2). (3). 圖 2-15 TCH 對 PKC 活化的影響 嗜中性白血球以 TCH 處理 5 分鐘,再以 1 μM fMLP (1) 刺激 1 分鐘。分離細胞 膜分劃,以西方點墨法偵測 PKC 和β actin。數據以免疫冷光強度與對照組之平 均比值(N=4)。* p<0.05,與對照組(2nd lane)比較;(2)刺激 30 秒後,萃取細胞 蛋白質液,進行 anti-p47phox 抗體免疫沈澱法,以西方點墨法分析。數據以免疫冷 光強度與對照組之平均比值(N=4)。* p<0.05,與對照組(2nd lane)比較。(3) 細 胞先以 DMSO、TCH 處理或 GF109203X 處理後,再以 fMLP 刺激 1 分鍾,萃取 細胞蛋白質液,以 PKC kinase activity assay kit 分析酵素活性。數據以 means ± S.D. 值表示(N=5)。* p<0.05,與對照組(2nd column)比較。 55.
(66) 圖 2-16 TCH對p38 MAPK、MK2磷酸化的影響 嗜中性白血球 (5 × 106 cells)以TCH處理5分鐘或10 μM SB 203580處理10分鐘, 再以 1 μM fMLP刺激1分鐘。以西方墨點法偵測phosph-p38、phosph-MK2和p38。 數據以免疫冷光強度與對照組之平均比值(N=3)。*P < 0.05,對照組 (2nd lane) 比較。. 56.
(67) 圖 2-17 TCH 對 PAK 磷酸化的影響 嗜中性白血球以 TCH 處理 5 分鐘,再以 fMLP 刺激 15 秒。以西方墨點法 anti-phospho-PAK1 (Thr423)/PAK2 (Thr402)抗體偵測。轉漬膜再以 anti-PAK1 抗 體偵測。數據以免疫冷光強度與對照組之平均比值(N=4)。*P < 0.05,對照組(2nd lane)比較。. 57.
(68) 圖 2-18 TCH 對 Vav 磷酸化的影響 嗜中性白血球以 TCH 處理 5 分鐘或 10 μM PP1 處理 10 分鐘,再以 fMLP 刺激 30 秒。進行西方點墨法並利用 anti-phospho-Vav (Tyr174)抗體偵測。轉漬膜再以 anti-Vav1 抗體偵測。數據以免疫冷光強度與對照組之平均比值(N=4)。*P < 0.05, 對照組(2nd lane)比較。. 58.
(69) 圖 2-19 TCH 對 cyclic AMP 生成的影響 嗜中性白血球以 DMSO、TCH 或 isoproterenol 處理後,吸取上清液,利用 cyclic AMP EIA kit 進行偵測。數據以 means ± S.D.值表示(N=3)。*p < 0.05 與 DMSO 組(1st column)比較。. 59.
(70) 圖 2-20. 結論. 60.
(71) 第五章 結論. 新合成 phenazine carboxylate 類的化合物 TCH 可以抑制 fMLP 所誘導嗜中 性白血球超氧自由基的生成。此抑制作用並非來自於影響細胞存活、超氧自由基 清除或直接抑制 NADPH oxidase 的活性,可能是影響細胞內的訊息傳遞機制。 而 p38 MAPK/MK-2 傳遞途徑及 cyclic AMP 可能並不參與 TCH 抑制超氧自由基 生成的機制。TCH 可能經由影響 PDK1/Akt、PKC 和 PAK1 訊息傳遞途徑來抑制 p47phox 的磷酸化,經由影響 Vav/Rac2 訊息傳遞途徑來抑制 Rac2 的活化,進而抑 制 NADPH oxidase 聚合及超氧自由基的生成 (圖 2-20)。TCH 可直接抑制 PDK1 的活性,而已知可直接抑制的 PDK1 的活性的藥物不多,TCH 有可能做為探討 PDK1 活性的工具用藥。. 61.
(72) 參考文獻 Abo, A., Qu, J., Cammarano, M.S., Dan, C., Fritsch, A., Baud, V., Belisle, B., Minden, A. (1998) PAK4, a novel effector for Cdc42Hs, is implicated in the reorganization of the actin cytoskeleton and in the formation of filopodia. EMBO J. 17, 6527-6540. Ahmed, S., Prigmore, E., Govind, S., Veryard, C., Kozma, R., Wientjes, F.B., Segal, A.W., Lim, L. (1998) Cryptic Rac-binding an p21(Cdc42Hs/Rac)-activated kinase phosphorylation sites of NADPH oxidase component p67(phox). J. Biol. Chem. 273, 15693-15701. Alessi, D.R., James, S.R., Downes, C.P., Holmes, A.B., Gaffney, P.R., Reese, C.B., Cohen, P. (1997) Characterization of 3-phosphinositide-dependent protein kinase which phosphorylates and activates protein kinase Balpha. Curr. Biol. 7, 261-169. Andresen, B.T., Rizzo, M.A., Shome, K., Romero, G. (2002) The role of phosphatidic acid in the regulation of the Ras/MEK/Erk signaling cascade. FEBS Lett. 531, 65-68. Arias-Romero, L.E., Chernoff, J. (2008) A tale of two Paks. Biol. Cell 100, 97-108. Babior, B.M. (1998) Activation of the leukocyte NADPH oxidase by phorbol ester requires the phosphorylation of p47phox on serine 303 or 304. J. Biol. Chem. 273, 9539-9543. Babior, B.M., Curnutte, J.T., Kipnes, R.S. (1975) Biological defense mechanisms. Evidence for the participation of superoxide in bacterial killing by xanthine oxidase. J. Lab. Clin. Med. 85, 235-244. Babior, B.M., Lambeth, J.D., Nauseef, W. (2002) The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 397: 342-344. Behn-Krappa, A., Newton, A.C. (1999) The hydrophobic phosphorylation motif of conventional protein kinase C is regulated by autophosphorylation. Curr. Biol. 9, 728-737. Bell, R.M., Burns, D.J. (1991) Lipid activation of protein kinase C. J. Biol. Chem. 266, 4661-4664. Bengis-Garber, C., Gruener N. (1996) Protein kinase A downregulates the phosphorylation of p47 phox in human neutrophils: a possible pathway for inhibition of the respiratory burst. Cell Signal 8, 291. Berridge, M.J., Irvine, R.F. (1989) Inositol phosphates and cell signaling. Nature 341, 62.
數據



+7

相關文件
國際學術機構 QS 世界大學排名(QS World University Rankings)或泰晤士高等教育世界 大學排名(Times Higher Education World
畢業於國際學術機構 QS 世界大學排名(QS World University Rankings) 或泰晤士高等教育世界 大學排名(Times Higher Education World. University
1、尚未有合適方法對成品本身進行佐劑去 吸附處理,故目前成品放行規格中 total protein 非以成品本身為檢品執行檢測,亦未 包含 viral protein
• • Linear Mode : Provide Wild Mass Range Linear Mode : Provide Wild Mass Range – – Protein MW determination Protein MW
北京中醫藥大學 天津中醫藥大學 上海中醫藥大學 遼寧中醫藥大學 南京中醫藥大學 浙江中醫藥大學 廈門大學 福建中醫藥大學 江西中醫藥大學 暨南大學
An alternative activated-state model was generated by substituting the single ligand-biased receptor refinement protocol with the following procedure: (1) rotation of TM VI by
Conserved complexes are connected sub- graphs within the bacteria-yeast alignment graph, whose nodes represent orthologous protein pairs and edges represent conserved
We will calculate the relationship points as their features and find the maximum relation protein spot pair as basic information for image matching.. If we cannot find any referable
