行政院國家科學委員會專題研究計畫 期中進度報告
脂蛋白元 A5 之功能研究(1/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-283- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 高照村 共同主持人: 林淑華 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 6 月 1 日
行政院國家科學委員會補助專題研究計畫
□ 成 果 報 告
V 期中進度報告
脂蛋白元 A5 之功能研究(1/3)
計畫類別:V 個別型計畫 □ 整合型計畫
計畫編號:NSC 93 - 2314- B- 002 -283 -
執行期間: 93 年 8 月 1 日至 94 年 7 月 31 日
計畫主持人:高照村
共同主持人:林淑華
計畫參與人員:
成果報告類型(依經費核定清單規定繳交):V 精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:國立臺灣大學醫學院醫事技術學系暨研究所
中 華 民 國 94 年 5 月 31 日
中文摘要 高三酸甘油酯血症是否為動脈粥狀硬化之危險因子目前仍有爭議。雖然空腹時高三酸 甘油酯之濃度可作為心血管疾病之危險指標,但如以其他種危險因子加以調整後,則其關 係性便減少了。然而仍陸續有許多數據支持高三酸甘油酯血症是心血管疾病之獨立危險因 子。位在第十一對染色體上的脂蛋白元AI-CIII-AIV之基因被認為是高脂血症特別是高三酸 甘油酯血症之一相關之基因,可是有些報告則指出脂蛋白元AI及CIII之多型性在調控脂肪 濃度之角色上只扮演有限的功能。最近有人發現了一個稱為脂蛋白元A5 的新基因,老鼠 如被轉殖入人類的脂蛋白元A5基因後,則其血中三酸甘油酯濃度會降為原來的三分之一。 而我們也發現在脂蛋白元A5基因上一新的變異c.553G>T與高三酸甘油酯血症有很高的關 聯性,而且此變異會引起脂蛋白元A5第185位置上的甘胺酸變成半胱胺酸,或許與高三酸 甘油酯血症之成因有關。本年度計劃便是研究脂蛋白元A5基因如何調控三酸甘油酯之代 謝,先建構能表現脂蛋白元A5的質體,使產生脂蛋白元A5蛋白質及其抗體並設立其定量方 法。 英文摘要
The exact role of hypertriglyceridemia as a risk factor for atherosclerosis is controversial. Although high fasting triglyceride levels are generally predictive of cardiovascular risk,
multivariate adjustment for other risk factors diminishes this association. Nevertheless, a growing body of evidence supports hypertriglyceridemia as an independent cardiac risk factor. The
apolipoprotein AI-CIII-AIV gene cluster on chromosome 11 has been identified as a candidate region for hyperlipidemia and in particular for hypertriglyceridemia. But some reports suggested that the apolipoprotein AI and CIII polymorphisms play only a very limited role in mediating variation in lipid concentrations. Pennacchio et al revealed a new apolipoprotein A5 gene. Mice expressing a human apolipoprotein A5 transgene showed a decrease in plasma triglyceride concentrations to one-third of those in control mice. We also found that a novel variant, c.553G>T, in the apolipoprotein A5gene that was associated with hypertriglyceridemia. In contrast to some other polymorphisms, which occur in non-coding regions of the gene, this variant occurs within the coding region and causes the change of amino acid sequence (a substitution of a cysteine for a glycine residue). In the first year, we repared plasmids of mature apo A5 and expressed the apo A5 protein. Polyclonal and monoclonal antibodies were produced then the ELISA was established.
實驗結果 第一節 脂蛋白元A5互補DNA的選殖 一、反轉錄-聚合酶鏈反應(RT-PCR) 由HepG2細胞抽取出總RNA後,以該RNA為模板,oligo(dT)18為引子進行反轉錄 反應,得到單股cDNA後,再以此單股cDNA為模板,以針對人類脂蛋白元A5cDNA的專一 性序列為引子進行PCR,進而合成雙股cDNA。為了能純化出高量的重組脂蛋白元A5,我 們選擇利用大腸桿菌進行大量的蛋白質表現。但由於脂蛋白元A5的N端具有20個胺基酸 的訊息胜肽,必須先將其切除後才能成為成熟的蛋白,而原核細胞無法自行進行這項
修飾動作;因此在選殖時,所設計出的引子(a5clone-F、a5clone-R,見Table 1)將 針對此20個胺基酸序列與以剔除。為了方便而後的蛋白質純化步驟,我們選擇使用 pET-15b質體,在成熟脂蛋白元A5的N端增加了6個Histidine(His-tag)的序列,同時 於成熟脂蛋白元A5與His-tag的中間含有thrombin之切點,以便純化蛋白後可將 His-tag切除,因此在設計引子時也增加了NdeⅠ之切點使之能接上質體;此段只含成 熟脂蛋白元A5序列的PCR產物大小為1059 bp(Fig.1,lane 1)。另一方面為了建構出 能於哺乳細胞表現的質體,我們也設計另一組引子(a5cDNA1、a5cDNA2,見Table 1), 針對全長的脂蛋白元A5進行PCR,此段包含脂蛋白元A5所有coding sequence的PCR產物 大小為1196 bp(Fig.1,lane 2)。 二、TA cloning
由RT-PCR所得的產物進行純化之後,接下來利用yT&A cloning kit接至TA質體中。 其中只含成熟脂蛋白元A5的質體命名為TA-a5sig,全長3787 bp(Fig.2,lane 1); 而含全長脂蛋白元A5的質體命名TA-a5full,全長3924 bp(Fig.2,lane 3)。經由轉 型至大腸桿菌後,抽取出少量質體DNA,利用限制酶切割產物進行初步確認;其中 TA-a5sig經NdeⅠ限制酶切割後會產生276、1041及2470 bp的片段(Fig.2,lane 2), 而TA-a5full經XmnⅠ限制酶切割後會產生129、864及2931 bp的片段(Fig.2,lane 4)。 三、選殖至表現載體 TA-a5sig及TA-a5full經直接定序確認基因序列正確無誤後,再將脂蛋白元A5序列 以限制酶切下,分別殖入表現載體pET-15b及pCR3TM -Uni中,並稱為pET-a5WT(Fig.3, lane 1)及pCR-a5WT(Fig.3,lane 3)。其中pET-a5WT為在細菌表現的質體,內含 His-tag、thrombin切點及成熟脂蛋白元A5;而pCR-a5WT則為在哺乳細胞表現的質體, 內含全長的脂蛋白元A5。同樣經由轉型至大腸桿菌後,抽取出少量質體DNA,利用限制 酶切割產物進行初步確認;pET-a5WT以XmnⅠ限制酶切割後會產生129、1628、1934及 3058 bp的片段(Fig.3,lane 2),而經ApaLⅠ限制酶切割後會產生1229、1972及2923 bp的片段(Fig.3,lane 4)。初步確認無誤後再經直接定序進行最終確認。 第二節 重組脂蛋白元A5的表現、純化及確認 一、重組脂蛋白元A5的表現 利用大量製備抽取出質體後,將pET-a5WT轉型至BL21(DE3)大腸桿菌中,並以IPTG 誘導重組脂蛋白元A5的表現,將其培養收集之後以超音波振盪破菌,分離出上清液及 沉澱物,再以SDS-PAGE配合Coomassie blue染色加以觀察分析;如同預期,由細菌大 量表現出來的人類重組脂蛋白元A5分子量大小約為41 kDa,且結果顯示大部分存在於 細菌的沉澱物中(Fig.4,lane 4)。 二、重組脂蛋白元A5的純化及確認 將細菌的沉澱物以含尿素(Urea)的Denaturing buffer溶解後,利用鎳離子鍵結 反應的管柱,以含高濃度imidazole的Elution buffer,將含His-tag的重組脂蛋白元 A5引流純化出來(Fig.4,lane 5);之後以Vivaspin 20離心管將蛋白質濃縮並把溶 劑置換成PBS。所得到的重組脂蛋白元A5利用thrombin可將His-tag切除,而成為大小 約39 kDa的蛋白,並以此做為初步的確認(Fig.4,lane 6)。最後把His-tag切除的 重組脂蛋白元A5轉印至PVDF膜後,送交做蛋白質N端的胺基酸定序,進行最終的確認。 於定序的過程中,由於第九個胺基酸為色胺酸,其苯環容易被循環反應中的酸性試劑 破壞,因此判讀時出現無訊號的結果;但因為其他胺基酸的序列均如預期,且先前質 體DNA也已確定序列的正確性,因此認定所表現出的蛋白質確實為重組的脂蛋白元A5。
確認後的重組蛋白便經由動物免疫的方式產生抗脂蛋白元A5的抗體,所產生天竺鼠抗 脂蛋白元A5抗體以ELISA測量抗體效價,結果約為943 k。 第三節 脂蛋白元A5在細胞內與培養液中的表現 利用本實驗室先前所選殖好的野生型脂蛋白解脂酶質體DNA(pCR3-L252L),與本 次實驗中所選殖的野生型脂蛋白元A5質體DNA(pCR-a5WT),共同轉染至HEK 293細胞, 首先利用西方墨點法觀察脂蛋白元A5於細胞內與培養液中的表現。由結果得知,取50 µg 的pCR3-L252L及10 µg的pCR-a5WT,以0.26 kV/960µF條件下轉染每毫升8×106的細胞, 經48小時小時培養後,於細胞溶解產物及培養液中,都可發現成熟脂蛋白元A5(39 kDa) 的表現;由此可知,脂蛋白元A5的確會將N端的訊息胜肽切除,並分泌至細胞外。另一 方面,於陰性對照組(只轉染pCR3-L252L或未經轉染的HEK 293細胞)中則沒有任何脂 蛋白元A5的表現(Fig.5)。 第四節 脂蛋白解脂酶於培養液中的活性與表現量分析 一、活性時效分析 由脂蛋白解脂酶的活性時效分析圖(Fig. 6),可以知道轉染pCR3-L252L後的HEK 293 細胞在培養第48小時後,會出現最高的活性表現;然而隨著培養時間的延長,因為細 胞數、代謝物毒素的增加及培養液越來越少的情況下,細胞活性逐漸下降。因此選擇 轉染後第48小時當作蛋白表現的最佳時間。另一方面,與pCR-a5WT共同轉染時,在同 時具有脂蛋白元A5表現的存在下,對於脂蛋白解脂酶活性的改變,並沒有造成明顯的 影響。 二、活性與表現量分析 在活性分析方面,轉染相同量的pCR3-L252L,加入Heparin後於第48小時收集培養 液,可以發現即使與pCR-a5WT共同轉染而同時具有脂蛋白元A5表現的存在下,脂蛋白 解脂酶的活性並沒有明顯的改變(Fig.7);而沒有轉染質體DNA的HEK 293細胞則幾乎 沒有活性。在表現量的濃度分析方面,以ELISA測量同樣為第48小時的培養液,脂蛋白 元A5的存在與否,對於脂蛋白解脂酶的表現量亦未造成明顯的影響;而沒有轉染質體 DNA的HEK 293細胞也無脂蛋白解脂酶的表現(Fig.8)。 第五節 脂蛋白元A5於培養液中的表現量分析 為確定培養液中脂蛋白元A5的實際表現量,我們以ELISA的方法測定其濃度 ,標準 曲線則是利用已知濃度的重組脂蛋白元A5求得(Fig.9)。結果顯示,在有pCR-a5WT共 同轉染的培養液中,脂蛋白元A5的表現量達4.68(±0.78) µg/ml;而在只轉染 pCR3-L252L或未經轉染的HEK 293細胞中,則沒有脂蛋白元A5的表現(Fig.10)。 討論 動物體內負責合成脂蛋白元 A5 的組織為肝臟細胞,而根據對脂蛋白元 A5 胺基酸序列 的分析,顯示脂蛋白元 A5 具有訊息蛋白的切點,表示脂蛋白元 A5 於肝細胞中被製造出後 會分泌出細胞外,van der Vliet 等人也確實於老鼠的血液中偵測到了脂蛋白元 A5 的存在。 在本篇報告中,我們分別建構出於細菌及哺乳細胞表現的脂蛋白元 A5 質體,首先利用大 腸桿菌大量表現並純化出重組脂蛋白元 A5,藉此製造出抗脂蛋白元 A5 的多株抗體,而後 再於 HEK 293 細胞株中轉染脂蛋白元 A5 質體,並利用抗體以西方墨點法偵測脂蛋白元 A5 於細胞的表現;結果顯示利用轉染方式所表現出的脂蛋白元 A5,其訊息胜肽同樣會經細胞
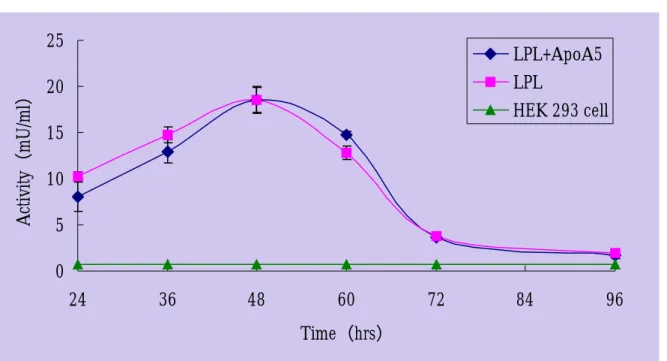
的修飾後切除,並且釋放至細胞外,而這樣的蛋白很可能是保有原來的生理活性的。另一 方面,於轉染脂蛋白元 A5 質體時,我們也共同轉染了含野生型脂蛋白解脂酶的質體至 HEK 293 細胞中;從脂蛋白解脂酶活性的時效分析可知,脂蛋白解脂酶於轉染後的第 48 小時會 有最高活性的表現量,因此選擇以第 48 小時的細胞培養液做為接下來進一步的分析。然而 結果顯示,脂蛋白元 A5 的存在與否,無論對於脂蛋白解脂酶的活性或蛋白表現濃度,甚 至於活性的時效分析上,都未造成任何明顯的影響。
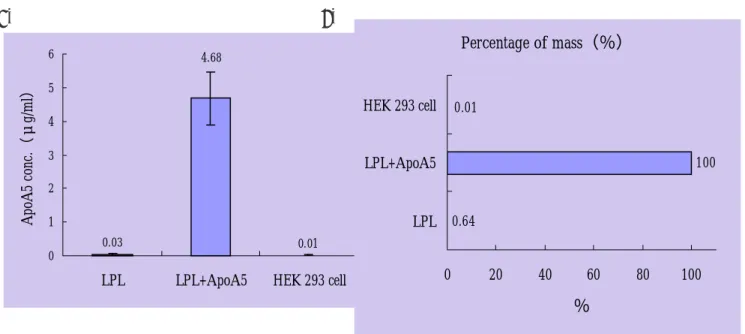
在 van der Vliet 等人的研究中指出,正常小鼠血清中的脂蛋白元 A5 濃度約為 1 µg/ml, 而於注入 apoa5 腺病毒載體的小鼠中,脂蛋白元 A5 表現量提高了約 20 倍。另外在 Pennacchio 等人的實驗中,apoa5 基因剔除小鼠的三酸甘油酯濃度是正常小鼠的 4 倍,且 異型合子的濃度介於同型合子與野生型小鼠之間;由此可知,脂蛋白元 A5 的表現量對三 酸甘油酯濃度的作用是呈現劑量效應關係的,且濃度在 1 µg/ml 之內就可造成影響。在本 篇報告中,我們以 ELISA 的方法測量了細胞培養液中的脂蛋白元 A5 濃度;測量結果發現, 以轉染方式於細胞培養液中所分泌出的脂蛋白元 A5 濃度為 4.68(±0.78)µg/ml。雖然其 脂蛋白元 A5 濃度未高達正常小鼠血中濃度的 20 倍,且目前為止人類血液中脂蛋白元 A5 的正常濃度值仍未有報告,但就先前實驗結果的推導歸納,我們仍可認為如此程度的脂蛋 白元 A5 表現量是十分充足的,因此脂蛋白元 A5 確實不會改變脂蛋白解脂酶的活性或蛋白 表現濃度。
脂蛋白元 A5 的生理功能與作用機轉究竟為何仍有許多其他的可能性。在 van der Vliet 等人的研究中,注入 apoa5 腺病毒載體的小鼠不僅三酸甘油酯的濃度減低,其膽固醇的量 也是明顯下降的;然而這種現象在 Pennacchio 等人的實驗中並未出現,其實驗中的 APOA5 基因轉殖小鼠膽固醇濃度仍維持在正常值。造成這種差異的可能原因是小鼠血液中脂蛋白 元 A5 的表現程度;在 van der Vliet 等人實驗中,注射 apoa5 腺病毒載體的小鼠脂蛋白元 A5 表現量比正常小鼠高了 20 倍,因此脂蛋白元 A5 可能必須在非常高表現量的情況下才 會對膽固醇濃度造成影響,而這也可以解釋 Pennacchio 等人實驗中的 apoa5 基因剔除小 鼠,其膽固醇濃度仍為正常的原因。此外,由他們先前的實驗結果顯示,脂蛋白元 A5 在 小鼠中位於 HDL 上,因此在高量脂蛋白元 A5 的存在下,其可能藉由競爭的方式,抑制了 膽固醇藉由脂蛋白元或 HDL 由細胞釋放出來的過程,也因此降低了血中膽固醇的濃度。
至於脂蛋白元 A5 在三酸甘油酯代謝的調控方面,van der Vliet 等人也提出了各種的可 能性。其中之一便是經由促進脂蛋白解脂酶的活性而提高三酸甘油酯的水解,因此增加了 TRLs 的清除;但本篇報告的實驗結果中,否認了此一可能性。另外脂蛋白元 A5 也可能會 影響肝臟清除脂蛋白的效率,例如類似或增強脂蛋白元 E 的功能,做為脂蛋白與肝細胞表 面接受器的配合基,進而促進 VLDL 的清除並降低三酸甘油酯的濃度。再者,於 Pennacchio 等人實驗中也發現,APOA5 基因轉殖小鼠血液中的脂蛋白元 C3 下降了 40%,而 apoa5 基因剔除小鼠的脂蛋白元 C3 則上升了 90%,且這樣的差異僅在脂蛋白元 C3 的蛋白量上 看到改變,脂蛋白元 C3 的轉錄層次上則無影響,因此脂蛋白元 A5 對於三酸甘油酯濃度的 影響,可能是經由改變脂蛋白元 C3 所造成;但是另一方面,Pennacchio 等人也比較了先 前 apoc3 基因剔除小鼠的實驗結果,他們發現 APOA5 基因轉殖小鼠的三酸甘油酯量為 apoc3 基因剔除小鼠的一半,因此,單純脂蛋白元 A5 對脂蛋白元 C3 的影響無法完全解釋 這樣的結果,所以除此之外脂蛋白元 A5 應該還有其他的作用機制存在。
Table 1:實驗過程中所需之寡核酸引子序列 a5cDNA1 5’-GCAGATAATGGCAAGCATGG-3’ a5cDNA2 5’-GCCACTTTCAAGGACTGAAC-3’ a5clone-F 5’-CGGAGTTCATATGCGGAAAGGCTTCTGGGACT-3’ a5clone-R 5-CGGAATTCATATGCTCAGGGGTCCCCCAGATG-3’ M13 promoter 5’-GTAAAACGACGGCCAGTG-3’ T7 promoter 5’-TAATACGACTCACTATAG-3’ a5E4-2500F 5’-AGCAACTGAAGCCCTACAC-3’ a5E4-2950F 5’-TTTCCGCCAGGACACCTAC-3’
Fig. 1:人類脂蛋白元 A5 之 RT-PCR 電泳圖
M: 1 kb DNA ladder
lane 1: 成熟脂蛋白元 A5 之 RT-PCR 產物(1059 bp) lane 2: 全長脂蛋白元 A5 之 RT-PCR 產物(1196 bp)
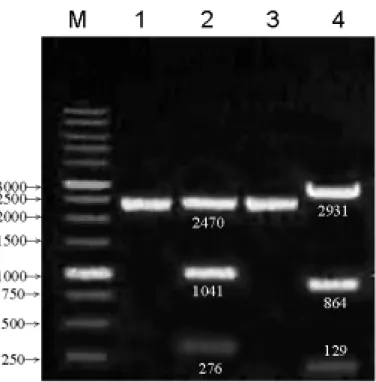
Fig. 2:TA-a5sig及 TA-a5full的限制酶圖譜
M: 1 kb DNA ladder
lane 1: 未經限制酶切割 Supercoiled form 的 TA-a5sig(3787 bp) lane 2: 經 NdeⅠ限制酶切割後的 TA-a5sig(276、1041、2470 bp) lane 3: 未經限制酶切割 Supercoiled form 的 TA-a5full(3924 bp) lane 4: 經 XmnⅠ限制酶切割後的 TA-a5full(129、864 及 2931 bp)
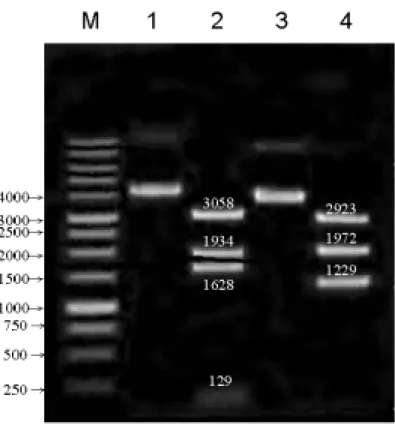
Fig. 3:pET-a5WT及 pCR-a5WT的限制酶圖譜 M: 1 kb DNA ladder
lane 1: 未經限制酶切割 Supercoiled form 的 pET-a5WT(6749 bp)
lane 2: 經 XmnⅠ限制酶切割後的 pET-a5WT(129、1628、1934、3058 bp) lane 3: 未經限制酶切割 Supercoiled form 的 pCR-a5WT(6124 bp)
Fig. 4:重組脂蛋白元 A5 之 SDS-PAGE(Coomassie blue 染色)
M: Prestained protein marker
lane 1: 未加入 IPTG 誘導前的細菌上清液 lane 2: 未加入 IPTG 誘導前的細菌沉澱溶解物 lane 3: 加入 IPTG 誘導後的細菌上清液
lane 4: 加入 IPTG 誘導後的細菌沉澱溶解物
lane 5: 經 Ni2+-column 純化後的重組脂蛋白元 A5(41 kDa) lane 6: 經 thrombin 切割後的重組脂蛋白元 A5(39 kDa)
47.5 kDa → 32.5 kDa →
Fig. 5:脂蛋白元 A5 的西方墨點分析圖
細胞經轉染後培養,於第 48 小時收集培養液或細胞溶解產物。
lane 1: 經 Ni2+-column 純化後的重組脂蛋白元 A5(41 kDa) lane 2: 經 thrombin 切割後的重組脂蛋白元 A5(39 kDa) lane 3: 經 pCR3-L252L 及 pCR-a5WT共同轉染細胞後的培養液 lane 4: 只轉染 pCR3-L252L 的培養液 lane 5: 未經轉染的 HEK 293 細胞培養液 lane 6: 經 pCR3-L252L 及 pCR-a5WT共同轉染細胞後的細胞溶解產物 lane 7: 只轉染 pCR3-L252L 的細胞溶解產物 lane 8: 未經轉染的 HEK293 細胞溶解產物 1 2 3 4 5 6 7 8
Fig. 6:脂蛋白解脂酶活性表現的時效分析 野生型脂蛋白解脂酶及脂蛋白元 A5 之質體 DNA 以電擊方式轉染至 HEK 293 細胞株中, 按一定時間收集細胞培養液測得的活性。各活性數據均以 3 次個別轉染結果之平均值加減 標準偏差(Mean±S.D.)表示。 0 5 10 15 20 25 24 36 48 60 72 84 96 Time(hrs) Activity(mU/ml) LPL+ApoA5 LPL HEK 293 cell
Fig. 7:脂蛋白解脂酶活性的分析圖(A)及比較圖(B) 野生型脂蛋白解脂酶及脂蛋白元 A5 之質體 DNA 以電擊方式轉染至 HEK 293 細胞株中, 第 48 小時收集培養液測得的脂蛋白解脂酶活性。各活性數據均以 3 次個別轉染,經β-gal 質體校正感染效率後之平均值加減標準偏差(Mean±S.D.)表示。 (A) 各型細胞所表現脂蛋白解脂酶之活性 (B) 活性之百分比,以只轉染脂蛋白解脂酶質體 DNA 所表現的蛋白活性為 100%,與共 同轉染脂蛋白元 A5 質體 DNA 相比較。 A. B. 18.52 18.57 0.72 0 5 10 15 20 25
LPL LPL+ApoA5 HEK 293 cell
Activity(mU/ml) Percentage of activity(%) 100 100.27 3.89 0 20 40 60 80 100 LPL LPL+ApoA5 HEK 293 cell %
Fig. 8:脂蛋白解脂酶表現量的分析圖(A)及比較圖(B) 野生型脂蛋白解脂酶及脂蛋白元 A5 之質體 DNA 以電擊方式轉染至 HEK 293 細胞株中, 第 48 小時收集培養液測得的脂蛋白解脂酶濃度。各濃度數據均以 3 次個別轉染,經β-gal 質體校正感染效率後之平均值加減標準偏差(Mean±S.D.)表示。 (A) 各型細胞所表現脂蛋白解脂酶之濃度 (B) 濃度之百分比,以只轉染脂蛋白解脂酶質體 DNA 所表現的蛋白濃度為 100%,與共 同轉染脂蛋白元 A5 質體 DNA 相比較。 A. B. 0.3 7.65 7.5 0 1 2 3 4 5 6 7 8 9
LPL LPL+ApoA5 HEK 293 cell
LP L c onc .( μ g/ m l) Percentage of mass(%) 100 98.03 3.92 0 20 40 60 80 100 LPL LPL+ApoA5 HEK 293 cell %
Fig. 9:脂蛋白元 A5 之 ELISA 標準曲線圖 以已知濃度的重組脂蛋白元 A5 做序列稀釋,經 ELISA 測定後所得之標準曲線圖。縱座標 為扣除背景值後的吸光值。 y = 0.0012x - 0.0006 R2 = 0.9968 0 0.2 0.4 0.6 0 100 200 300 400 500 600 conc.(ng/ml) A410
Fig. 10:脂蛋白元 A5 表現量的分析圖(A)及比較圖(B) 野生型脂蛋白解脂酶及脂蛋白元 A5 之質體 DNA 以電擊方式轉染至人類 293 細胞株中,第 48 小時收集培養液測得的脂蛋白元 A5 濃度。各濃度數據均以 3 次個別轉染,經β-gal 質 體校正感染效率後之平均值加減標準偏差(Mean±S.D.)表示。 (A) 各型細胞所表現脂蛋白元 A5 之濃度 (B) 濃度之百分比,以共同轉染脂蛋白元 A5 質體 DNA 表現的蛋白濃度為 100%,與只 轉染脂蛋白解脂酶質體 DNA 相比較。 A. B. 0.03 0.01 4.68 0 1 2 3 4 5 6
LPL LPL+ApoA5 HEK 293 cell
ApoA5 conc.(μg/ml) Percentage of mass(%) 0.64 100 0.01 0 20 40 60 80 100 LPL LPL+ApoA5 HEK 293 cell %