國 立 交 通 大 學
生物科技研究所
碩士論文
利用定點突變對氧化鯊烯環化酵素
的假設活性區進行功能性分析
Functional Analysis of Putative Active Site
Residues in the Oxidosqualene-Lanosterol
Cyclase by Site-Directed Mutagenesis
研 究 生 : 虞美婷
指導教授 : 吳東昆 博士
利用定點突變對氧化鯊烯環化酵素的
假設活性區進行功能性分析
Functional Analysis of Putative Active Site Residues
in the Oxidosqualene-Lanosterol Cyclase
by Site-Directed Mutagenesis
研 究 生 : 虞美婷 Student : Mei -Ting Yu
指導教授 : 吳東昆 博士 Advisor : Dr. Tung- Kung Wu
國立交通大學
生物科技研究所
碩士論文
A Thesis
Submitted to Department of Biological Science and Technology College of Science
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biological Science and Technology July, 2005
Hsinchu, Taiwan, Republic of China
利用定點突變對氧化鯊烯環化酵素的假設活性區進行功能性分析 研究生 : 虞美婷 指導教授 : 吳東昆老師 摘要 氧化鯊烯環化酵素(Oxidosqualene cyclase,OSC),催化直鏈狀的 氧化鯊烯經單一步驟環化生成四環或五環的三萜類。不同物種(氧化) 鯊烯環化酵素活性區上的差異,誘導酵素進行不同的摺疊,形成產物 的多樣性及複雜性。 為了解氧化鯊烯環化酵素假設活性區之胺基酸與受質環化/重組 催化機制的關係,以同屬之鯊烯環化酵素(Squalene-Hopene cyclase, SHC)的 X-ray 晶體結構為模板,經序列比對推估出 OSC 的 29 個可能 活性區位置,以定點突變進行環化重組的機制研究。經質體交換 (plasmid shuffle)初步篩選發現有 9 個突變點(OSCF104T、OSCH146A、
OSCW232A、OSCW232R、OSCH234A、OSCM532A、OSCW587A、OSCF699A 與
OSCY707A)無法存活,可能代表這些突變位置是催化反應的重要影響
點。培養此 9 個含 OSC 突變株的酵母菌,經由產物分離與結構鑑定, 以推測這些胺基酸在環化機制上所扮演的催化角色。
在 OSCW232A、OSCH234A
突變株有新產物發現,OSCW232A與實驗室
另一個突變株 OSCH234Y 產物相同,OSCH234A 另多了一個未知物,進
一步純化分離,NMR 鑑定未知物結構為 Protosta-20,24-dien-3s-ol。藉 由建構 OSCW232A、OSCH234A產物圖譜,推論 OSCW232A、OSCH234A在 催化反應中調節環化的重組步驟,可能扮演穩定碳陽離子及脫氫的功 能性角色。 為了更進一步探討 OSCW232在環化機制的功用,進行飽和突變實 驗(Saturated mutagenesis),將此位置胺基酸突變成其餘 19 種氨基酸, 觀察產物變化探討催化的功能性。利用氣相層析/質譜比對,發現有 5 個突變產物,除了 OSCW232A原本的 3 個突變產物,另有 2 個未知物 滯留時間 26 min 及 26.5 min,目前正進一步分離此 2 個未知物,同時
Functional Analysis of Putative Active Site Residues in the Oxidosqualene-Lanosterol Cyclase by Site-Directed Mutagenesis Student: Mei-Ting Yu Advisor: Dr. Tung-Kung Wu
Abstract
Oxidosqualene cyclases (OSCs) constitute a family of enzymes which catalyze the conversion of linear oxidosqualene into polycyclic tetracyclic and pentacyclic triterpenoids in a one-step enzyme-catalyzed reaction. The OSC-catalyzed cyclization /rearrangement reaction represents one of the most remarkable and fascinating biotransformations found in nature. In order to clarify the function of the amino acid residues located on the putative active site cavity surface of oxidosqualene-lanosterol cyclase of yeast Saccharomyces cerevisiae, site-directed mutagenesis coupled with plasmid shuffle experiments were carried out. Following the assaying of twenty-nine alanine-scanning mutants for the complementation of yeast viability, nine inactive mutants(OSCF104T, OSCH146A, OSCW232A, OSCW232R, OSCH234A, OSCM532A, OSCW587A, OSCF699A and OSCY707A) were further analyzed for product profile. Further product isolation and characterization of each mutant showed that OSCW232A mutant produced Protosta-12,24-dien-3s-ol, Lanosterol and Parkeol as end products, exhibiting the same product profile as that of OSCH234Y mutant. In parallel, the OSCH234A mutant produced Protosta-12,24-dien-3s-ol, Lanosterol, Parkeol, and Protosta-20,24-dien -3s-ol as end products.
To completely characterize the functional role of W232 in OSC activity, the saturated mutagenesis experiments at W232 position were performed. Following the analysis of each mutant products, two additional new products with retention time (rt.) at 26.0 and 26.5 min were observed on GC-MS spectrometry. The EI low mass spectrum of rt. 26 min product showed a molecular ion at m/z 426 and fragment peaks at 357 ([M-C5H9]+) and 313 ([M-C8H15-H2]+), suggesting an intact tetracyclic ring. In
summary, the results suggest a role for W232 in stabilizing a cationic intermediate, probably after protosteryl cation formation, and in determining the deprotonation position for diverse product profile.
謝 誌
即將告別二年的碩士生活,當中有期許、有挫折、有失落,一一 的完成克服,感覺上完成了人生的一個階段,即將要展開人生的下一 關挑戰。兩年研究所生涯在我浩瀚的人生當中也許只佔小小的比例, 但卻讓我獲益良多。不同與以往的生活經歷,科學研究及做實驗佔了 生活的大部分時間,實驗總是會遇到不同的困難,計畫永遠趕不上變 化,我從中學習到積極面對事情、堅持到底的精神。我覺得科學研究 者必須具備理性與感性,面對許多挑戰與困難,必須要有很強的熱 情、無比的毅力與永不服輸的精神,以及冷靜的邏輯思考與細心的分 析。在研究所兩年的日子裡,學到許多科學概念與實驗技術,週遭的 人事物實在令我獲益不少。 首先要感謝我的指導教授吳東昆博士,感謝老師二年前以無比的 勇氣收了不曾摸過 pipette 的我,老師對實驗設計、科學理念以及實 驗技術上的細心指導帶給我不少的啟發,從老師身上我也學到了處事 積極的態度與獨立思考的能力。另外也要感謝應化系李耀坤教授與淡 江化學系的鄭建中博士,於口試期間對於論文細心的審閱與修改,並 且提供寶貴的經驗與建議使此論文能夠更盡完整,並感謝清大貴儀中 心彭菊蘭小姐,在 NMR 分析上的幫忙。 兩年的研究生活,感謝程翔學長亦師亦兄的照顧及在實驗上的指 導,還要特別感謝媛婷學姊不厭其煩的教導實驗技巧與建議,學姊總 是用她超乎常人的樂觀激勵我,她 color 般閃亮積極的態度推動了這 本論文的產生,不知何時,感覺上有了亦師亦友的情誼。 還有同窗的夥伴吳家四千金-希希、小八、晉豪,雖然晉豪很早 就單飛躍升為中草藥一哥,還是要感謝豪哥在 GC/MS 上的協助。小 八總是以她犀利及獨到的見解在我實驗陷入膠著時給予幫助,而小希 以她北極企鵝般的冷笑話使我的生活酷涼有勁,真不愧是我冷笑話軍 團的成員。也要感謝大仔碩一時在生物晶片方面的指導,使我眼界開 拓不少。還有令宗、宏明、大鳥、天才源、駿哥、文暄、大景、怡親、 浩宇等可愛的學弟妹們,在實驗上的協助及閒暇之餘所帶來的歡樂。目錄
中文摘要 ………...… i 英文摘要 ……….……….…ii 謝誌………...iii 目錄 ……….…iv 圖目錄 ………...…...….……vi 表目錄 ……….………ix 第一章 序論 1-1 固醇類及三萜類化合物對生物體的重要性…..………...…1 1-2 三萜類環化酵素家族的簡介 ………...………..…6 1-3 氧化鲨烯環化酵素家族的簡介 ………...….………...13 1-3-1 氧化鲨烯-羊毛硬脂醇環化酵素 (OSC) ……….….…..15 1-3-2 氧化鲨烯-環阿屯醇環化酵素 (CAS ) ………..27 1-3-3 鲨烯-蛇麻烯環化酵素 ( SHC ) .……….……32 1-4 (氧化)鲨烯環化酵素之胺基酸序列比對 ...………..36 1-5 研究目的 ..………...……….…….42 第二章 實驗材料與方法 2-1 實驗材料 ( Material ): 2-1-1 化學藥品與材料 ………...…. 49 2-1-2 儀器 ………..…. 50 2-1-3 菌株與載體 ………..…..……. 50 2-1-4 溶液與緩衝液 ………..………..…....51 2-1-5 培養基 ………..……….……… 52 2-2 實驗方法 ( Method ): 2-2-1 重組質體的建構 ( The Construction of recombinants )..542-2-2 質體 DNA 轉入活細胞中……… 61 2-2-3 質體交換 / 死/活篩選 ..…...……….. ……….63 2-2-4 麥角固醇補充篩選 ...………....64 2-2-5 酵母菌的培養 ………..……….65 2-2-6 脂質萃取 ………...65 2-2-7 薄層色層分析及管柱液相色層分析 ………….……... 65 2-2-8 硝酸銀矽膠管柱液相色層分析……….67 2-2-9 建構突變點電腦模擬圖 ……….……70 第三章 結果與討論 3-1 篩選目標 ……….………...…….71 3-2 質體交換 / 反向篩選 ...………...………72 3-3 產物分析與探討 ……….…...…...76 3-4 W232 的細部探討………...86 3-4-1 由序列比對探討 Trp232 在催化上角色……….86 3-4-2 W232 saturated mutagenesis 突變株的建構與產物分析.91 3-5 藉電腦模擬圖佐証突變效應影響的產物圖譜………..104 第四章 結論與未來展望 ……….………….…..116 第五章 參考文獻 ………...119 附 錄……….……….123
圖目錄
圖 1-1 固醇類生合成途徑 ………...……….…...5 圖 1-2 環化酵素家族在不同物種間的產物專一性………...………..7 圖 1-3 氧化鯊烯的環化機制-經不同碳陽離子中間物有產物多樣..9 圖 1-4 高等植物中的 CAS 與 LUS 環化假設機制………...12 圖 1-5 Johnson Model………..…………...….….….16 圖 1-6 Griffin 提出 Aromatic Hypothesis 理論模組………...17 圖 1-7 酵素催化環氧化物開環及環化起始的假設模組… ……...18 圖 1-8 不同物種 CAS1 基因與 ERG7 基因之高度保留胺基酸…19 圖 1-9 酵母菌(S.c) OSC Val454 突變株可能進行的環化過程…...20 圖 1-10 人類 OSC X-ray晶體結構……….…..21 圖 1-11 人類 OSC與膜結合時的X-ray晶體結構………..22 圖 1-12 OSC催化直鏈的氧化鯊烯環化成四環羊毛硬脂醇的機制..23 圖 1-13 Asp455與Cys456及Cys533氫鍵拉扯誘導Epoxide開環…...24 圖 1-14 OSC酵素活性區的芳香族基團 ..…....…………..…...26 圖 1-15 人類OSC表面活性區胺基酸………..……….26 圖 1-16 CAS與OSC間有高度保留性的胺基酸…………..….…..….28 圖 1-17 不同物種氧化鯊烯-環阿屯醇環化酵素在阿拉伯芥 CAS H477 有高度保留..………...………...……….………....29 圖 1-18 阿拉伯芥 CAS 正常途徑合成物及突變點生成的產物.…..29 圖 1-19 同源性的鯊烯環化酵素與氧化鯊烯-羊毛硬脂醇環化酵素有 類似的環化機制……….……….32 圖 1-20 SHC 之 X-射線晶體結構圖………..….…...33 圖 1-21 以 OSC 抑制劑說明環化起始反應…………..…………34 圖 1-22 OSC、CAS、SHC 的序列比對圖……….37 圖 1-23 Q-W 區域在環化酵素家族內之分佈情形………..41 圖 1-24 不同物種環化酵素活性區些微差異形成物種專一性產物...44 圖 1-25 實驗動機之假設………..47 圖 1-25 實驗策略流程……….….48圖 2-1 QuickChange Site-Directed Mutagenesis 選殖策略 ………55 圖 2-2 以切接選殖建構雙點突變的策略……….……..59 圖 3-1 以 SHC X-ray 結構為模板,分子模擬 OSC 可能活性區位 置對照圖,及所建構的突變株……….……..……...71 圖 3-2 質體交換/反向篩選策略圖……….………….73 圖 3-3 9 個突變株轉入 TKW14C2 在 TLC 片的初步產物分布...76 圖 3-4 以 GC-MS 分析突變產物,建構 W232A、H234A 產物圖 78 圖 3-5 產物進行乙醯化反應………...……….80 圖 3-6 以 GC-MS 確認經銀染矽膠管柱層析分離的 H234-unknown acetate 純度……….…..…81 圖 3-7 H234A-unknown 1 HNMR………..……….….81 圖 3-8 H234A-unknown 13CNMR………82 圖 3-9 H234A-unknown HSQC………82 圖 3-10 H234A-unknown 專一的結構訊息………...83 圖 3-11 新產物 Protosta-20,24-dien-3ß-01 的結構圖……….84 圖 3-12 由產物圖譜推估 W232、H234 參與的環化重組途徑.…89 圖 3-13 Trp232 及 His234 在酵母菌氧化鲨烯-羊毛硬脂醇環化酵素 (OSC)電腦模擬圖中與產物之間的空間關係圖………9 0 圖 3-14 140L W232I-rt.26 min 未知物通銀染純化送測 NMR 前的 GC-MS………....94 圖 3-15 銀染 TLC 片以乙醚為展開液的產物展佈順序…………95 圖 3-16 rt.26 min 未知物乙醯化前後 MS 圖……….…………97 圖 3-17 標準品乙醯化前的 MS 圖譜對照圖………...…….98 圖 3-18 標準品乙醯化的 MS 圖譜對照圖………...………….99 圖 3-19 Protosta-12,24-dien-3ß-ol 乙醯化前後 MS 圖………..…100
資訊………..………102 圖 3-21 Trp232Ala/Tyr510Ala 雙點突變產物與控制組的 GC-MS 比對圖………114 圖 3-22 Trp232Ala/Tyr510Lys 雙點突變產物與控制組的 GC-MS 比對圖………114 圖 3-23 Trp232Ala/His234Ala 雙點突變產物與控制組的 GC-MS 比對圖………115 圖 3-24 His234Ala/Tyr510Ala 雙點突變產物與控制組的 GC-MS 比對圖………115
表目錄
表 1-1 酵母菌 OSC Val454 突變株產物………..20
表 1-2 阿拉伯芥 CAS 已被報導的定點突變產物及比例分布…30 表 2-1 Quick-Change Site-Directed Mutagenesis 所用材料條件...57 表 2-2 Quick-Change Site-Directed Mutagenesis 使用之聚合酵素 放大程式………57 表 2-3 Quick-Change Site-Directed Mutagenesis PCR 產物處理...58 表 2-4 接合反應前以適當限制酵素處理質體 DNA 的條件.…….60 表 3-1 OSC 假設活性區建構的突變點進行質體交換/反向篩選 將酵母菌 CBY57 篩出的突變株,轉入酵母菌 TKW14C-2 進行產物分析……...………74 表 3-2 利用序列比對得到 OSC、CAS、SHC 活性區及重要的 鄰近胺基酸的對應表………...87 表 3-3 W232 saturated mutagenesis 突變株建構及功能性互補結果 91 表 3-4 以 GC-MS 的標準品比對建構 W232 saturated mutagenesis 產物圖譜以及利用 GC 算出產物比例………93 表 3-5 W232 飽和突變株依產物比例分布的分類……….104 表 3-6 突變點側鏈上胺基酸基團與各碳陽離子中間物的距離...106
第一章 序 論
1-1 固醇類及三萜類化合物對生物體的重要性
固醇類的組成通常是四環~六環的固性結構,外有一個 8 個碳的 側鏈,且在 C-3 有一羥基(-OH)者稱之。自然界的固醇類,例如:膽 固醇、麥角固醇、β-麥胚固醇、植物固醇,以及其生合成代謝途徑中 的上游產物與衍生物-三萜類天然物(Triterpenes),他們的作用機制、 生合成途徑、代謝調節,一直是近幾十年來的重要研究課題。 許多研究指出,這些固醇類及三萜類天然物,在生物體具有重要 的 生 理 功 能 。 自 然 界 固 醇 類 的 生 合 成 , 是 由 兩 個 碳 的 乙 醯 輔 酶 -A(Acetyl-CoA)開始合成,在幾個步驟的反應縮合後,經速率決定步 驟HMG-CoA 還原酶(HMG-CoA reductase)催化生成 3-甲基-3,5-羥 基戊酸(Mevalonate),再經一連串的 ATP 水解參與反應,生成異戊二 烯類的中間物(Isoprenoid intermediates),隨後六個五碳的異戊二烯單 元體經過縮合及還原反應,產生疏水性的鯊烯(Squalene),從鯊烯開 始,固醇類生合成中間物變為未磷酸化、強疏水性、油脂狀的化合物, 因此在生物體內需細胞外固醇攜帶蛋白(sterol carrier protein)來維持 運送的溶解度。鯊烯再經氧化代謝生成氧化鯊烯,而後經由環化及一 連串反應,合成其最終產物[1]。在不同物種間,會分別利用鯊烯或氧 化鯊烯作為環化起始物。細菌、一些原核生物及低等植物會利用鯊烯 環化生成蛇麻烯(Hopene),而在高等植物、真菌、動物…等會由氧化 鯊烯進行環化,經一連串反應生成植物固醇(phytosterol)、麥角固醇 (Ergosterol)、膽固醇(圖 1-1)。 過去文獻指出固醇類生合成途徑的中間物參與很多重要的生理 反 應[2]。 如 異 戊 二 烯 類 中 間 物(Isoprenoid intermediates) 是 泛 醌(Ubiquinone)、多萜醇(Dolicol) 、血基質(Heme)及植物脂質萜類、胡 蘿蔔素(β-carotene)…等數種重要化合物的前驅物。血基質與紅血球的 形 成 、 攜 氧 及 合 成 數 種 血 基 質 蛋 白 質 有 關(如 : 細 胞 色 素 P450 (cytochrome P450)、電子傳遞鏈上的蛋白複合體…等)。泛醌,亦為輔 酶Q(Coenzyme Q),廣泛存於大部分的細胞中,參與能量代謝的電子 傳遞,可傳遞電子協助氧化還原反應[1],目前己被製成健康食品,其 具有抗氧化、強化免疫的功效,可抑制氧化型低密度脂蛋白,預防心 血管疾病。多萜醇與內質網中蛋白質的醣化有關,負責作為 N-醣苷 鍵(N-linked glycosides)的多醣攜帶者。植物固醇(phytosterol)可與食物 中的膽固醇競爭其與膽酸的結合,因此可阻礙膽固醇吸收,降低心血 管疾病的風險,且植物固醇可提高免疫能力,幫助自然殺手細胞(NK cell)運作。鯊烯是一種三萜類化合物,為一種強力的攜氧物質,由於 其結構的特殊性,與水作用可產生 3 個氧,因此主要的功能為細胞活 化及生氧作用,可提高內臟功能,避免細胞缺氧,以及調節膽固醇總 量,預防心血管疾病。此外,在動物體內產生的膽固醇參與體內許多 新陳代謝的重要生理調控,它是細胞膜上重要組成份,調控細胞膜的 流動性,影響胞內外物質的滲透,並且可藉此調控膜上蛋白質或膜結 合酵素,使其有最適活性去進行訊息傳遞、代謝反應與催化作用[3]。 另外,膽固醇也是膽汁、固醇類荷爾蒙、胞內生理代謝調控的重要物 質。膽固醇在體內代謝生成膽汁而幫助脂質及脂溶性維生素的吸收, 它也是體內五種固醇類激素的前驅物:糖皮質固醇(Glucocorticoids; 如:皮質醇 Cortisol)、礦物皮質固醇(Mineralocorticoids;如:醛固酮 Aldosterone) 、 雄 性 激 素 (Androgens) 、 雌 激 素 (Estrogens) 與 黃 體 酮 (Progestins),透過這些激素,使人體可面對不同的生理壓力,它們可
份、礦物質吸收再利用與排出,使其代謝平衡,並影響男女性徵成熟
與發育。而人體也可利用膽固醇,自行合成脂溶性維生素 D,維生素
D 是一群具有激素樣功能的固醇,繼而影響鈣質吸收,血鈣與骨鈣迴
饋循環平衡,刺激基因表現與骨質的強度[1]。此外,膽固醇也是目前
正被積極研究的Lipid raft(脂質筏)的組成分。Lipid raft 是指細胞膜上 一塊固性區域,當細胞膜上膽固醇比例增加,細胞膜的流動性會減 少,許多研究發現,細胞膜上有一塊區域富含膽固醇,因而較不具流
動性,就像筏在流動性的膜上移動,故此稱為脂質筏[4]。Lipid raft 普
遍存於所有細胞膜上,許多文獻的研究指出,其可能與訊息傳遞、發 炎反應、細胞移動(Migration)、神經傳導…等反應有關,如:阿茲海 默症(Alzheimer,s disease)[5],目前很多文獻在探討阿茲海默症與Lipid raft 的相關性,實驗証實當 Lipid raft 被破壞或組成改變,β-amyloid (β-類澱粉樣蛋白)在腦中的的含量會減少,阿茲海默症的成因就是因為 這種蛋白不正常的沈積在腦中而造成的,目前許多研究都傾向此疾病 是某些物質透過 Lipid raft 引發訊息傳遞,繼而造成此種絲狀病變蛋 白形成的假說[4, 6]。而膽固醇則是Lipid raft 的主要組成份,因此,針 對固醇類的研究更是日益重要[5]。此外,在酵母菌的實驗也發現,部 份具特定結構的固醇類,以及其相應的激素,對於細胞分裂增生週期 的調控,有密切的相關性。因此,固醇類及三萜類天然物在酵母菌、 動、植物細胞一直被廣泛的注意著[7, 8]。 固醇類在生命體具有一自然的迴饋調節機制,使體內的固醇含量 達一恆定,假若此調節機制失效,又或人體長期攝食高膽固醇飲食, 會使得血液中膽固醇含量升高,而人體血液裡主要負責膽固醇運輸的 脂蛋白-LDL(Low density lipoproteins;低密度脂蛋白)也會相應增加, 此時若人體受到自由基攻擊,又或處於高氧化壓力之下,血液循環中
的 LDL 會被修飾化,也就是進行乙醯化作用或 LDL apo B (LDL apolipoprotein B,低密度脂蛋白表面脂蛋白 B)被氧化形成 ox-LDL(氧 化型低密度脂蛋白),此時會被循環中的巨噬細胞(Macrophages)所帶 有的清道夫受器(Scavenger receptor)所辨識而引發內噬作用,移入血 管內皮的巨噬細胞攝取了這些修飾型的 LDL,而這些修飾型的 LDL 並無法參與細胞內膽固醇正常代謝的迴饋調節機制,因而他們所攜帶 的膽固醇會過量聚積在巨噬細胞內,造成巨噬細胞轉形變成泡沫細胞 (foam cell),泡沫細胞在內皮細胞的沈積釋出生長因子,刺激平滑肌 的分化及血管壁上鈣化斑痕的形成,使得血管變狹窄阻塞、彈性變 差,有粥狀斑塊出現,因而造成了高血壓、心臟病、動脈粥狀硬化、 中風…等心血管疾病的發生。而在黴菌當中,其終產物麥角固醇 (Ergosterol)為黴菌胞膜上的重要組成,也是不可或缺的存活因子,因 此,目前許多先進的分子生物技術已被大量用來研究生命體中固醇類 及三萜類天然物的重要性,了解其代謝上的反應途徑,而能用以發展 抗黴菌及降膽固醇的藥物[9, 10]。因為心血管疾病為國人十大死因之 一,也是許多已開發國家的主要致死因素。另一方面,由於院內感染、 癌症治療、器官移植或其他治療,導致免疫力降低引發的黴菌感染日 益嚴重,故此,抗黴菌及降膽固醇的藥物的研究一直被持續進行著[11]。 近年來,固醇類生合成代謝途徑中的環化酵素--氧化鯊烯環化酵 素(Oxidosqualene Cyclase),已逐漸成為抗黴菌及降膽固醇的藥物研發 標的。先前所發展的藥物及相關研究的抑制劑,多針對固醇類生合成 途徑的限速步驟酵素 HMG-CoA Reductase 做抑制,但此方式會影響 其下游的異戊二烯中間物與三萜類化合物的生成,繼而影響具重要生 理功能的二次代謝物的形成與調節,如:泛醌、多萜醇…等。所以,
受抑制所引起的副作用[11]。此外,由於此環化酵素的環化反應,涉及 了包括幾十個鍵的斷裂、形成及碳骨架的重排、去質子化步驟(終止 脫除反應),如此高效率的一步環化反應,更強烈的引發我們想去探 究的興趣。
S
S
t
t
e
e
r
r
o
o
l
l
B
B
i
i
o
o
s
s
y
y
n
n
t
t
h
h
e
e
t
t
i
i
c
c
P
P
a
a
t
t
h
h
w
w
a
a
y
y
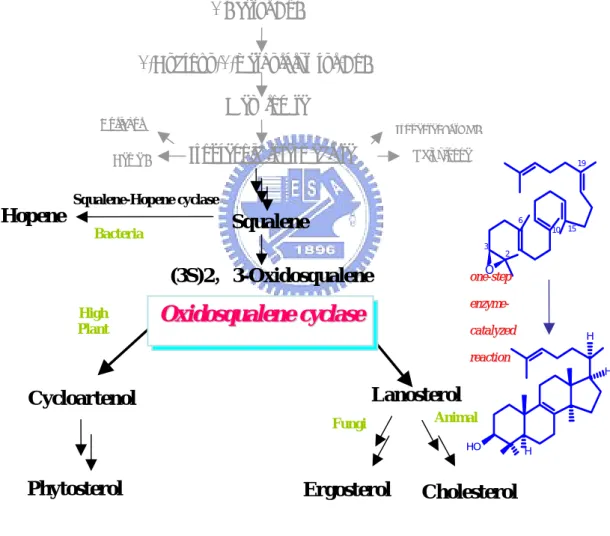
3 Acetyl CoA O 2 3 6 10 15 19 HO H 3-Hydroxy-3-methylglutaryl-CoA MevalonateDolichol Isopentenyl tRNA
Isoprenoid intermediate Ubiquinone
Heme A Squalene-Hopene cyclase Hopene Squalene Bacteria (3S)2,3-Oxidosqualene Lanosterol Cycloartenol High Plant Fungi Animal O Oxxiiddoossququaalleennee cycyccllaassee H H o onnee--sstteepp e ennzzyymmee- -c caattaallyyzzeedd r reeaaccttiioonn
Phytosterol Ergosterol Cholesterol
1-2 三萜類環化酵素家族的簡介
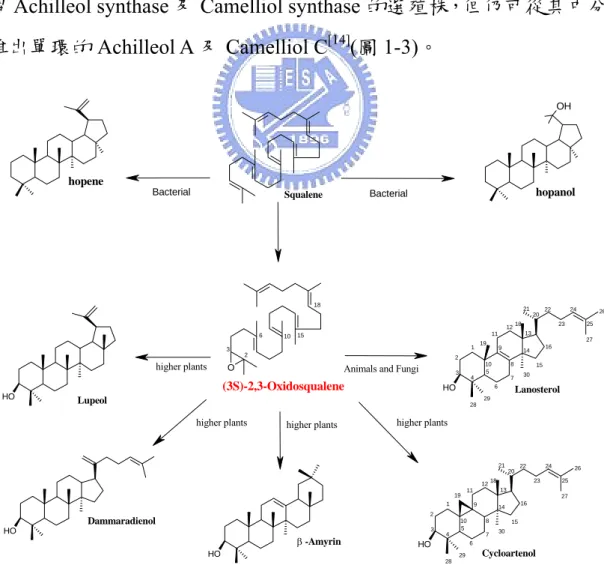
近半世紀以來,三萜類環化酵素家族所催化鯊烯(Squalene)或氧 化鯊烯((3S)-2,3-oxidosqualene)環化形成多環多烯類(terpenes)的催化 機制,一直是分子生物家及化學家認為最迷人及最具挑戰性的生化轉 化反應之一。三萜類化合物(Triterpenoid)是一群由三十個碳前驅物衍 生而來的多烯類產物,目前,由天然來源、酵素立體結構相關的環化 作用轉化而來的有近200 個不同多烯類骨架的三萜類化合物[12]。這些 具多烯類骨架的三萜類化合物,是由自然界三萜類環化酵素家族,經 單一步驟酵素催化反應,藉酵素與受質間的鏡像或立體選擇性環化及 骨架重排,生成了如此多樣的產物,而這些產物泛存於自然界的各物 種中形成產物多樣性。 在不同物種間依其所使用之酵素,將直鏈多烯類的鯊烯或環氧化 物的氧化鯊烯以單一生物轉化過程,催化多個不對稱立體中心的環化 及 1,2-氫化基(hydride)、甲基轉移,而獲得不同結構的多環脂醇或三 萜類化合物[13]。整個環化過程包括雙鍵或 Epoxide(環氧物)質子化 (Protonation)以活化誘導開環、環化(Cyclization)、甲基與氫鍵之重組 (Rearrangement) 、 脫 除 (Elimination) 之 終 止 反 應 。 這 些 同 源 性 (Homologous)的三萜類環化酵素,可將直鏈狀的鯊烯或氧化鯊烯(OS) 轉化成多樣產物,且在蛋白質序列上有相似的胺基酸保留區域位置。 這些物種催化生成的物種專一性產物(圖 1-2),依反應的產物骨 架的複雜性區分,有6-6-6-5 四環、6-6-6-6-5 五環、6-6-6-6-6 五環及 單環、雙環、三環與六環的三萜類化合物[12]。例如在動物、真菌、甲 基營生菌(methylotrophic bacterium) ,氧化鯊烯會藉氧化鯊烯-羊毛硬 脂 醇 環 化 酵 素(Oxidosqualene-Lanosterol Cyclase;OSC) 催 化 生 成中,氧化鯊烯可被環阿屯醇合成酵素(Cycloartenol Synthase;CAS) 環
化生成 6-6-6-5 四環的環阿屯醇(cycloartenol),或是藉羽扇醇合成酵
素(Lupeol Synthase;LUS ) 合成 6-6-6-6-5 五環的羽扇醇(Lupeol),或 是藉香桂素合成酵素(Amyrin Synthase;AMS) 生成 6-6-6-6-6 五環的 α- 香桂素(α-Amyrin) 或 ß-香桂素(ß-Amyrin),又或藉其他環化酵素 環化形成四環~六環的產物。而在較低等的植物、部分原蟲類(Protozoa) 及細菌中,藉由鯊烯-蛇麻烯環化酵素(Squalene - Hopene Cyclase;SHC)
將直鏈的鯊烯環化生成6-6-6-6-5 五環的蛇麻烯(Hopene), 或藉其他
環化酵素反應生成蛇麻醇(Hopanol) 和里白醇(diplopterol) 等環狀產 物。另外在 Achillea odorata 及 Camellia sasanqua 之中,雖然無法獲 得Achilleol synthase 及 Camelliol synthase 的選殖株,但仍可從其中分 離出單環的Achilleol A 及 Camelliol C[14](圖 1-3)。 O 2 3 6 10 15 18 2 3 19 1 4 5 7 8 9 10 11 13 14 12 15 16 18 20 21 22 24 23 6 O H 27 28 29 30 25 26 2 3 19 1 4 5 7 8 9 10 11 13 14 12 15 16 18 20 21 22 24 23 6 O H 27 28 29 30 25 26 O H O H O H OH Lupeol (3S)-2,3-Oxidosqualene Lanosterol Cycloartenol Dammaradienol -Amyrin hopene hopanol Bacterial Squalene
Animals and Fungi higher plants
higher plants higher plants higher plants Bacterial
β
這些三萜類環化酵素家族催化生成的產物多樣性,開始是藉環化 酵素中所含殘基(Residue),使得親核性的雙鍵或 Epoxide 藉親電性作 用 進 行 開 環 起 始 反 應 , 經 一 連 串 的 碳 陽 離 子- 烯 烴 環 化 作 用 (cation-olefin cyclization)生成高能帶正電的碳陽離子中間物(cation intermediates)。鯊烯在其環化酵素當中,會依全椅形的形式(all-chair form)摺疊,而氧化鯊烯在氧化鯊烯環化酵素中,經不同物種環化酵 素上殘基的誘導,會依椅形-船形-椅形(chair-boat-chair)或椅形-椅形-椅形(chair-chair-chair) 二種方式摺疊,繼而進行上述的開環形成碳陽 離子中間物的反應[15]。 由於相同反應受質-氧化鯊烯在酵素中不同的摺疊方式,會造成 立體構形相異的反應機制,經質子化及一連串的雙鍵電子轉移後,會 生成二種類型的碳陽離子中間物: (一) chair-boat-chair 會生成原脂醇 碳陽離子中間物(Protosteryl Cation intermediates),之後經甲基、氫化 基轉移的重排重組作用,在不同位置脫氫或藉水分子作用,中止反應 而形成環阿屯醇、羊毛硬脂醇和南瓜子雙烯脂醇(Cucurbitidienol)。(二) chair- chair –chair 會形成達瑪烯碳陽離子中間物(Dammarenyl Cation intermediates),在不同酵素中,有些繼續誘導環化,形成 6-6-6-6-5
及 6-6-6-6-6 五環的碳陽離子中間物,繼而再經甲基、氫化基轉移的
重排重組,各自環化生成達瑪雙烯醇(Dammaradienol)、羽扇醇、α-香桂素及ß-香桂素…等[16-18]。
而另一方面,三萜類環化酵素所催化的環化反應其反應本身的複 雜性,吸引許多研究學者來了解其酵素所催化反應的反應機制,及探 討其酵素本身的結構與功能間的關係。早期對環化酵素機制的研究, 是利用反應受質的類似物進行催化,藉由分析酵素轉化生成結構的構 形範圍及產物的特性,推估其作用機制[13]。由於反應受質類似物其立 體或電子上的差異,可能對酵素的受質結合區或活性區參與環化的殘 基 有 斥 力 的 交 互 作 用 , 繼 而 影 響 了 受 質 在 酵 素 的 結 合 摺 疊 , 如:11-fluorooxidosqualene(11-氟氧化鯊烯)的電子斥力會使其不傾向 椅形-船形-椅形的重排。而 14-fluorooxidosqualene(14-氟氧化鯊烯)其 碳14 上所接的取代基(-F substituent)可能對碳 10 上的甲基有斥力交互 作用,或是對鄰近的酵素受質結合的口袋區有排斥作用,使得受質在 酵素中失去正確摺疊形式而喪失Epoxide 活化開環的起始作用[14]。可 利用此方法找出活性區上的殘基涉及的環化機制及影響哪一環的形 成…等。 此外,一些對三萜類環化酵素的研究發現,環化酵素家族於高等 生物中經分子演化下,酵素受質由較低等的鯊烯演化成需氧參與的較 高等的氧化鯊烯。先前的研究對這些環化酵素提供的一些反應機制上 的資訊。這些環化酵素利用其受質結合區的胺基酸誘導受質有正確的 定位而進行摺疊,將環狀的氧化鯊烯或具對稱性的直鏈多烯類的鯊 烯,能夠在各自的環化酵素中摺疊環化成具多個不對稱立體中心的多 環脂醇或三萜類化合物,整過環化過程包含 Epoxide 或雙鍵質子化活 化誘導開環、共價鍵斷裂與形成的環化、氫化基、甲基轉移的重排重 組(rearrangement)、脫除(elimination)而至中止反應[19, 20]。在酵素中經 環化重組形成的過渡性高能碳陽離子中間物,最後會藉質子被抽取
物。酵素受質結合區與活性區胺基酸,藉其上的電子散佈及斥力而與 受質有交互作用,當胺基酸改變,活性區上的立體構形受扭曲,會衝 擊受質在酵素中的摺疊,活性區的電子散佈改變,會修飾環化、碳陽 離子重排及脫氫的鍵形成之反應過程。 進一步來說,這些環化酵素在蛋白質序列上有相似的序列及胺基 酸保留區,但因演化上的可塑性,使其構形序列上胺基酸因演化的過 程而增加改變,生成新的催化特性,而引起自然界中的多樣性產物分 佈。這些胺基酸廣泛變化,改變了酵素活性區胺基酸與受質間的交互 作用,或反應過程的碳陽離子中間物,因此衝擊產物結構的形成,故 而各物種的環化酵素有其產物專一性[21]。例如:環化酵素 CAS 與 LUS 同樣存在於高等植物中,因演化造成酵素活性區胺基酸的變化,使相 同的反應受質氧化鯊烯,有不同的誘導摺疊形成,經酵素上質子的捕 抓與給予、電子穩定過渡態中間物,CAS 會催化生成四環碳陽離子 中間物,而 LUS 在產物的 D、E 環間有進一步的電子穩定作用,會 繼續環化生成五環的碳陽離子中間物,而後各自再依活性區胺基酸的 電子分佈,利用氫鍵拉扯效應、質子在鄰近的胺基酸位置與受質間轉 移,電子穩定作用協助甲基、氫化基轉移重排,分別形成四環的環阿 屯醇及五環的羽扇醇(圖 1-4)。
O 6 10 15 14 18 19 22 23 Asp N N H H H O O His + O H H N NH His + O 6 10 14 15 18 19 22 23 Asp H O O N N H H His + O H 20 B Aromatic group + O H 17 18 B Aromatic group + O H 19 20 B Aromatic group + O H 17 18 20 19 21 22 圖 1-4 : 高等植物中的 CAS 與 LUS,會催化相同受質氧化鯊烯,演 化過程形成的活性區胺基酸差異,會使受質在酵素中進行不同摺疊方 式,進行不同的環化過程而有其產物專一性。上圖為 CAS 開環活化 引起環化形成的假設機制;下圖為 LUS 的推估機制,兩者因活性區 上胺基酸的差異,環化生成各自專一的產物[12]
1-3 氧化鯊烯環化酵素家族的簡介
氧 化 鯊 烯 環 化 酵 素 是 指 真 核 生 物 中 轉 化 氧 化 鯊 烯 (Oxidosqualene;OS)生成下游的代謝前驅物,如:固醇類、膜組成物、 固醇類激素或其他二級代謝物。這些氧化鯊烯環化酵素會催化氧化鯊 烯進行複雜的生物轉換反應,屬於自然界三萜類環化酵素家族的一 類,主要利用 OS 做為反應受質。其下包含許多環化酵素,如:氧化鯊 烯-羊毛硬脂醇環化酵素(OSC)、氧化鯊烯-環阿屯醇環化酵素(CAS)、 羽扇醇合成酵素(LUS)、香桂素合成酵素(AMS)…等。 氧化鯊烯環化酵素的研究已持續了數十年。Robinson 發現酵素必 須將受質的碳原子折疊在膽固醇的相似位置才可進行催化反應[22]。而Bloch 與 Cornforth 利用混入實驗(incorporation)直接証明羊毛硬脂醇 合成的反應機制中,酵素內有催化甲基與氫化基的轉移的重組反應 [23]。Corey 與 Bloch 証明了哺乳類中氧化鯊烯環化酵素合成羊毛硬脂 醇的反應受質是2,3-氧化鯊烯而不是鯊烯[24]。Barton 更進一步証明真 核生物是利用 3(S)-2,3-氧化鯊烯做為環化的反應受質,而非 3(R)-鏡 像異構物[23, 25, 26]。Robustell 也利用受質類似物進行氧化鯊烯環化酵素 生物轉化的機制研究,以不同位置取代基的受質類似物,驗証酵素上 鄰近基團的功用[26, 27]。Corey 藉由合成氧化鯊烯環化酵素所催化生成 的可能中間物,探討此單一步驟高效率的將不具立體中心的氧化鯊烯 環化生成具多個不對稱立體中心的反應機制[28]。而 Ruzicka 與其研究 團隊,也利用生命期短、離子性的中間物,建構出立體化學的假說[29]。
Corey 和 Matsuda 也以實驗証明 Epoxide 親電性活化開環,需要酸性 基團作為質子供應者誘導Epoxide 活化開環[30, 31]。研究發現 OS 置於 中性及弱酸溶液中在室溫可穩定存放一天,因此強酸被認為是開環的
留且為催化所必需的基團[32]。故此,Asp 被認為作為質子供應者誘導 Epoxide 開環,繼而進行環化反應。而氧化鯊烯環化酵素依各物種酵 素上的差異而有不同的摺疊機制及產物骨架的複雜性。 由先前研究的驗證,對氧化鯊烯環化酵素的機制有一概略的了 解,此類酵素有鏡像及立體化學專一性的特性,藉單一步驟的生物轉 化反應,可生成具十幾個立體中心的化合物。其酵素的環化反應先經 酵素上酸性基團誘導開環而質子化,而後經酵素上胺基酸基團電子分 佈引導摺疊,進行環化、重排等反應而形成高能的碳陽離子中間物, 之後經酵素-受質間質子的牽引交換,或是藉由水分子參與反應進行 提供-OH 基團,或水分子做為酵素與受質間的中介者而有氫鍵效應的 拉扯作用,繼而中止反應,藉脫氫作用形成雙鍵,又或被提供-OH 基 團形成diol 產物。目前有近 100 種氧化鯊烯環化酵素的產物被報導[33, 34],這些產物廣泛的骨架差異,是經類似的催化機制,但因酵素活性 區或受質結合區的差異,導致多種的環狀碳骨架的形成,經環擴張與 甲基、氫化基的轉移,產生不同碳陽離子中間物,最後依不同的碳陽 離子-脫除(cation-quenching)步驟中止反應。
1-3-1 氧化鯊烯-羊毛硬脂醇環化酵素 (OSC)
在固醇類代謝途徑中,氧化鯊烯-羊毛硬脂醇環化酵素會催化氧 化鯊烯環化生成羊毛硬脂醇,繼而在真菌與動物分別代謝成其重要的 生理物質-麥角固醇(Ergosterol)與膽固醇。在酵母菌(Saccharomyces cerevisiae)中 OSC 是由 ERG7 基因所轉譯出來的膜蛋白,由 2196 個
鹼基對轉譯成 731 個胺基酸序列,蛋白質分子量為 83.7KD。由於此 環化酵素分子量大、會與膜結合的特性,使得酵素純化不易,故此多 年來對結構與功能機制上的探討,所知仍有限。因此,對 OSC 環化 反應機制的研究,主要是利用分生技術及反應受質類似物進行探討。 先前對羊毛硬脂醇生合成的環化反應研究,通常會透過下述3 種方式 探討:(1)羊毛硬脂醇合成酵素藉由受質氧化鯊烯的結構類似物進行作 用的研究。(2)對不同物種的羊毛硬脂醇合成酵素做蛋白質序列比對 及基因選殖,得到一些結構上的資訊,及序列結構與機制上的相關 性。(3)以定點突變的酵素活性研究,判斷酵素上一些胺基酸基團在 環化機制上的重要性。 針對OSC 受質專一性和反應的立體化學特性,在 1987 年 Johnson 提出了一個理論模組(Johnson Model)(圖 1-5),認為環化反應藉由反應 活性區域上的酸性殘基(acidic residue)對(3Z)-2,3-氧化鯊烯的環氧基 (epoxide)或是鯊烯的雙鍵提供質子,促使起始反應,隨後進行碳陽離 子-烯烴多環環化作用(cation-olefin polycyclization)。受質之立體化學 上,酵素利用具相位選擇性(facing selective)的負電荷,穩定過渡狀態 (transition state)的正電高能量的碳陽離子。Johnson Model 提出了 OSC
以酵素活性區帶負電胺基酸穩定環化過程的高能中間物的假說[35,
36]。在1992 年 Griffin 也提出了一個 Aromatic Hypothesis 理論模組[37]
因此,認為這些富含電子芳香族基團,可能會利用碳陽離子-π 電子作 用(cation-π interaction)的穩定效應,引導受質折疊在適當位置,並且 能穩定高能的正電過渡態中間物,使得之後的甲基、氫化基團能順利 的被誘導轉移。 O Enz-AH 2 3 6 7 10 11 15 18 23 14 19 + O O -O -O O -O Enz-AH 2 3 6 7 10 11 15 18 23 14 19 + O Enz-AH 2 3 6 7 10 11 15 18 23 14 19 + O O -O -O O -圖1-5 : Johnson Model 指出酵素活性區以帶負電胺基酸穩定高能 中間物[35, 36]
O Enz-AH 2 3 6 7 10 11 15 18 23 14 19
+
N O H N N N O Enz-AH 2 3 6 7 10 11 15 18 23 14 19+
O Enz-AH 2 3 6 7 10 11 15 18 23 14 19+
N O H N N N圖1-6 : Griffin 提出 Aromatic Hypothesis 理論模組,酵素利用活性 區高度保留的芳香族胺基酸基團,透過碳陽離子-π 共振作
用穩定帶正電的高能產物[37]
整合先前對氧化鯊烯環化酵素催化反應的研究,可得到一些資 訊:(1)原脂醇碳陽離子中間物(Protosterol cation intermediate)結構上為
碳17ß-側鏈,顯示 OSC 催化生成羊毛硬脂醇,在碳 20 上有立體化學 的控制,且碳 17 側鏈位於酵素緊密鍵結袋中[24]。(2)原脂醇碳陽離子 中間物在酵素中的確是以 B 環形成船形的立體構形摺疊環化,雖然 船形構形在能量上較不傾向,但可能是藉酵素反應區上胺基酸電子散 布所引導生成的。[38](3)反應受質氧化鯊烯碳 10 上所帶的甲基,對受 質能否在酵素中正確摺疊有決定性的影響,且可決定碳10、碳 11 雙 鍵加成作用的正確相位選擇性(facial selectivity)[39] 。(4)氧化鯊烯形成 四環原脂醇碳陽離子中間物的反應過程,是先生成五碳的 C 環,之 後藉 C 環的環擴張以形成六碳環後再繼續環化[31]。(5)氧化鯊烯的環
化起始,只能發生在受質有正確結合在酵素鍵結區,及有正確的摺疊
引起環化形成,環化起始的 Epoxide 開環活化反應,是藉高度保留的
Asp 基團將 Epoxide 的氧質子化而引起[31]。(6)A 環的關閉與 Epoxide 鍵斷裂有協調作用。(7)原脂醇碳陽離子中間物與產物-羊毛硬脂醇之 間的氫化基、甲基轉移的重組機制,是受酵素內部低活化能的熱動力
學驅動,使其有最小的能量需求,而停在碳8、碳 9 位置脫氫形成雙
鍵[31]。
透過一些受質類似物與 OSC 做親和性標定(Affinity labeling)、定
點突變及一些由受質類似物推測反應機制的實驗[40],酵母菌 OSC 的 His146、Trp232、His234、Asp456 被認為是催化機制上所必需的[41]。 這些研究提出了一個假設模組(圖 1-7),認為 Asp456 及 His146 與酵 素催化 Epoxide 開環引起環化起始的反應有關,推測 His146 藉氫鍵 拉扯效應增強 Asp456 的酸性,使其做為質子供應者促使 Epoxide 質 子化而開環。Trp232 與 H234 被認為可能位於受質接觸區域,利用富 含電子的側鏈穩定原脂醇碳陽離子中間物[41]。 圖1-7 : 酵素催化環氧化物開環及環化起始的假設模組[41]
利用序列比對針對物種高度保留的位置進行定點突變及導向演 化的實驗中發現,CAS 的 Ile481 位置[42],在不同物種的 CAS1 基因均
為 Ile,對應於不同物種間的 ERG7 基因均為 Val,而 CAS 的 Tyr410
位置也有相同情況,在不同物種的 ERG7 基因均為 Thr。在 CAS 定點 突變的研究發現,對這些高度保留位置突變成 ERG7 的胺基酸,產物
會由正常的環阿屯醇變為羊毛硬脂醇。而CAS Ile481 及 Tyr410 以序
列比對之後,對應於 OSC 為 Val454 及 Thr384(圖 1-8),因此,一些
定點突變實驗被用以探討OSC Val454 及 Thr384 位置所參與的催化反
應(表 1-1)。這些研究認為 Val454 涉及 B 環的形成,而與脫氫的中止
反應有關[42, 43](圖 1-9)。
O 20 O H H + O H H H 8 9 11 + O H H O H Lanosterol O H OH O H O H H H 8 9 11 + Achilleol A Parkeol Lanost-24-ene-3ß,9α- diol Squalene O 20 O H H + O H H H 8 9 11 + O H H O HO Lanosterol H Lanosterol O H OH O H O H H H 8 9 11 + Achilleol A Parkeol Lanost-24-ene-3ß,9α- diol Squalene 圖1-9 : 酵母菌(S.c) OSC Val454 突變株可能進行的環化過程[43, 44] 酵母菌(S.c) OSC 突變株 Lanostrol % Parkeol % Lanost-24-ene -3ß,9α-diol % Achilleol A % OSCV454A 95 -- -- 5 OSCV454G 83 -- -- 17 OSCV454L 100 -- -- -- OSCV454I 100 -- -- -- OSCT384Y 79 11 10 -- OSCT384Y V454L 20 60 20 -- OSCT384Y V454I 13 64 23 --
人類的氧化鯊烯-羊毛硬脂醇環化酵素之 X-ray 晶體結構、酵素的 活性區域及活性區胺基酸基團可能參與的反應機制,目前已在 2004 年 Nature 期刊被正式發表了[45]。此 OSC 結構(圖 1-10)的確定,有助 於提供不同物種的環化酵素的基本模板的依據,並在結構與反應機制 上提供相關的訊息。 從人類 OSC 的 X-ray 晶體結構可發現,此酵素插入膜的部分有 一直徑 25Å 的通道,形成酵素活性位置的結合凹槽。這個通道被認 為允許反應受質-氧化鯊烯進入疏水性的活性區域,且會對於此活性 區域會有分隔作用使受質的出入口被壓縮封起。受質的通道會藉OSC 的 Tyr237、Cys233 及 Ile524 這些胺基酸側鏈構形的改變,或是藉被 扭曲拉緊的環狀區段 516~524 及 697~699 的重排作用而被完成[46](圖 1-11)。 圖 1-10 : 人類 OSC X-ray 晶體結構,圖中黑色部分為與抑制劑結合, 指出受質結合的反應活性位置[45]
圖 1-11 : 人類 OSC 與膜結合時的構形,圖中黑色部分為抑制劑 Ro 48-8071 結合在中心反應活性結合區,藉此表示催化時反應 物與酵素活性區胺基酸的相關位置[45] 同屬於三萜類環化家族的SHC 常被用以做為了解 OSC 環化機制 的結構模組,但此同源性結構模組有其限制性。SHC 是利用演化程 度較早的鯊烯為受質,藉由組織胺酸(Histidine)的殘基增強酸活化打 開雙鍵的起始環化作用,在B 環為椅形的構形,且藉著 SHC Trp169 的作用,利用碳陽離子-π 電子共振的穩定效應,引起 D 環擴張,繼 而引導E 環環化而形成五環的蛇麻烯(Hopene)[47]。先前許多關於OSC
較不傾向的船形構形摺疊,環化反應中止在D 環[48]。OSC 與 SHC 的 環化反應在機制上有明顯的差異性,故此,利用人類 OSC 的 X-ray 晶體結構,對此高度立體選擇性的環化機制,會有較清楚的論點[45]。 先前一些實驗及研究已經証明 OSC 催化氧化鯊烯一步環化生成 四環的羊毛硬脂醇,是透過活性區上的 Asp455 酸催化開環,使 Epoxide 的氧被質子化而有環化起始反應,同時也建構了酵素中催化 環化反應過程[30] (圖 1-12)。 O Enz-AH 6 10 15 14 18 19 22 23 + O 6 10 3 15 19 O H 6 + A Ring O H 10+ B Ring O H 15 + C Ring O H H H H 17 20 9 8 + D Ring H H H O H Lanosterol Squalene O Enz-AH 6 10 15 14 18 19 22 23 + O 6 10 3 15 19 O 6 10 3 15 19 O H 6 + A Ring O H 10+ B Ring O H 15 + C Ring O H H H H 17 20 9 8 + D Ring H H H O H Lanosterol Squalene 圖1-12 :OSC 催化直鏈的氧化鯊烯環化生成四環羊毛硬脂醇的機制[30]
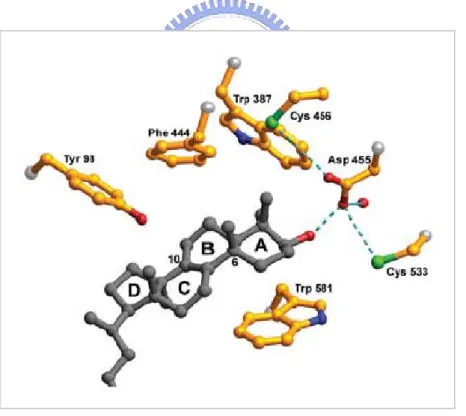
因此,利用人類 OSC 的 X-ray 晶體結構,可發現 OSC 是利用活
性區的Cys456 及 Cys533 與 Asp455 有氫鍵的交互作用,藉此氫鍵的
拉扯效應增強 Asp455 的酸性,幫助酸誘導開環的起始環化。當一個 催化循環完成,Asp455 會透過水分子及 Glu459 的羧酸基團,或是藉 最後脫氫步驟的質子轉移再質子化。Trp387、Phe444、Tyr503、Trp581 的位置具保留性,會利用其側鏈富含電子的特性,透過碳陽離子-π 共振的交互作用,在適當的位置及位向來穩定 A 環形成與 B 環形成 所生成的碳6、碳 10-四級碳陽離子中間物。Tyr98 藉著推動氧化鯊烯 碳10 上的甲基至分子平面之下,可迫使 B 環有能量不傾向的船形構 形的摺疊,也就是透過其立體空間大的側鏈,阻礙 B 環形成能量較 傾向的椅形構形[45] (圖 1-13)。
圖 1-13 : Asp455 與 Cys456 及 Cys533 的氫鍵拉扯效應幫助 Epoxide 誘導開環; Trp387、Phe444、Trp581 穩定 A 環形成與 B 環
His232 與 Phe696 藉由側鏈基團富含電子的 π 電子交互作用,穩 定C 環形成時,依反-馬可尼可夫(anti- Markovnikov)機制形成的二級 碳陽離子。且可藉碳陽離子-π 電子共振的交互作用,穩定碳 20-碳陽 離子的正電。His232 對碳 8/碳 9-碳陽離子是最靠近的鹼基團,因此 能協助碳 8/碳 9 的脫氫反應。OSC 一連串的環化步驟會中止在五個 碳的D 環所形成的碳 20-四級原脂醇碳陽離子,因為 OSC 缺乏像 SHC 的Trp169 芳香族官能基,SHC 藉著 Trp169 的 π 電子,穩定碳 17-二 級碳陽離子中間物,使其有較長的生命期而更進一步的環化形成 E 環。碳 20-碳陽離子中間物藉著骨架重排被轉換生成碳 8/碳 9-碳陽離 子,這些碳陽離子的轉移,是因為酵素活性區在碳 8/碳 9 附近有 7 個 芳香族胺基酸基團--Trp192、Trp230、His232、Tyr237 Tyr503、Phe521、 Phe696,這些在碳 8/碳 9 附近的高密度 π 電子,促使碳陽離子中間物 骨架的重排(圖 1-14)。高度保留的 His232,其鹼性殘基靠近碳 8/碳 9-碳陽離子,能夠接受質子而催化環化反應中最後的專一性脫氫作
用。而Tyr503 側鏈上的-OH 基團,會與 His232 有氫鍵交互作用的拉
扯效應,使得 His232 能有最適當的位置,催化最後的脫氫作用而中
止反應,且 His232 也許會透過鄰近的水分子的交互作用,影響催化
圖 1-14 : 酵素活性區在碳 8/碳 9 附近有 7 個芳香族胺基酸基團 --Trp192、Trp230、His232、Tyr237 Tyr503、Phe521、Phe696, His232 與 Tyr503 有氫鍵拉扯效應,使得 His232 能有最適
當的位置,催化最後的脫氫作用而中止反應,且 His232 會 透過和水分子的交互作用,影響催化反應[45]。 圖 1-15 : 氧化鯊烯環化 酵素與其產物-羊毛硬脂 醇 形 成 複 合 物 的 結 構 圖。圖中所顯示的胺基 酸基團為距離產物小於 5Å 的位置,水分子只有 在 Asp455 及 His232 附 近有被觀察到[45]
1-3-2 氧化鯊烯-環阿屯醇環化酵素 ( CAS )
氧化鯊烯-環阿屯醇環化酵素在脂醇生合成的途徑中,催化氧化 鯊烯環化生成植物固醇的前驅物—環阿屯醇(Cycloartenol),繼而代謝 合成植物固醇(Phytosterol)。在阿拉伯芥(Arabidopsis thaliana) 的環阿 屯醇環化酵素(CAS, EC 5.4.99.8),由 759 個胺基酸所組成,分子量為 86 kDa。 氧化鯊烯-環阿屯醇環化酵素(CAS)與氧化鯊烯-羊毛硬脂醇環化 酵素(OSC)在環化的機制上有相當相似的反應過程。它們同樣以氧化 鯊烯作為反應受質,在各自的專一性酵素中,同樣依椅形-船形-椅形 的摺疊形式在酵素中進行催化,環化生成碳 20-原脂醇碳陽離子中間 物,且有相同的甲基、氫化基轉移的重組機制,只有在最後的脫氫終 止反應時有不同位置的脫氫作用,氧化鯊烯-環阿屯醇環化酵素(CAS) 催化碳19 上的氫脫除而生成環阿屯醇,氧化鯊烯-羊毛硬脂醇環化酵 素則催化碳8 上的氫脫除而生成羊毛硬脂醇。而利用序列比對的方式 對照兩者假設的活性區位置,發覺只有少數的胺基酸基團不同,這些 位置很有可能就是造成兩者在脫氫作用上不同的物種專一性反應。因 此,目前很多研究就是利用序列比對不同物種的 CAS1 基因及 ERG7 基因的胺基酸序列,找出各自的高度保留性胺基酸,將其彼此做突 變,希望藉此找出酵素結構上與催化機制的相關性[49, 50]。 一些探討 CAS 催化反應的研究指出[51](表 1-2) (圖 1-18),Ile481 及 Tyr410 在不同物種的氧化鯊烯-環阿屯醇環化酵素有高度保留性 (圖 1-16),對應於不同物種的氧化鯊烯-羊毛硬脂醇環化酵素為高度保留的 Val 及 Thr,而將氧化鯊烯-環阿屯醇環化酵素 Ile481 突變成 Val
及 Tyr410 突變成 Thr,產物會有羊毛硬脂醇生成,顯示透過突變效
的功能,而在CAS H477 突變點[52],也可發現有導向演化的結果出現 (圖 1-17)。
圖1-16 : 紅色箭頭所指為不同物種的氧化鯊烯-羊毛硬脂醇環化酵素
及氧化鯊烯-環阿屯醇環化酵素具有高度保留性,此可能
圖 1-17 : 不同物種的氧化鯊烯-環阿屯醇環化酵素在阿拉伯芥 CAS H477 有高度保留性,但在氧化鯊烯-羊毛硬脂醇環化酵素 有些為Gln,有些為 Cys[52] O Squalene O H Lanosterol O H Cycloartenol O H Parkeol O H H Achilleol A O H H Camelliol C CAS 1 WT CAS 1 Mutant CAS 1 Mutant CAS 1 Mutant CAS 1 Mutant O Squalene O H Lanosterol O H Cycloartenol O H Parkeol O H H Achilleol A O H H Camelliol C CAS 1 WT CAS 1 Mutant CAS 1 Mutant CAS 1 Mutant CAS 1 Mutant 圖1-18 : 阿拉伯芥 CAS 正常途徑合成的產物及突變點生成的產物
阿拉伯芥CAS 突變株 Cycloartenol % Lanosterol % Parkeol % 9ß-∆7-
Lanosterol Achilleol A % Camelliol C%
CASI481 99 -- 1 -- -- -- CASI481L 83 1 16 -- -- -- CASI481V 55 24 21 -- -- -- CASI481A 12 54 15 -- 13 6 CASI481G 17 23 4 -- 44 12 CASY410T -- 65 2 33 -- -- CASY410C -- 75 -- 24 1 -- CASH477N -- 88 12 -- -- -- CASH477Q -- 22 73 5 -- -- CASI481V/ Y410T -- 78 < 1 22 -- -- CASI481V/ H477N/ Y410T -- 78 -- 22 -- -- CASI481V/ H477Q/ Y410T -- 78 -- 22 -- --
統整這些突變點的研究結果而有一些論述。Ile481 位於活性區域 的頂端位置,非常靠近已被公認的受質鍵結區域上,與環化反應之去
質子作用有關,影響A 環形成,當 Ile 突變成立體空間障礙較小的
Val,在空間上與羊毛硬脂醇合成酵素有相似的位置,會改變原先在 碳19 的脫氫作用[53]。
CAS Tyr410 與 His257 有氫鍵配對的交互作用,其靠近在碳 19
的位置,協助碳 19 上的氫脫除反應而形成一個碳 19 的三角環生成。 Tyr410 可能屬於保留序列區域,且位於分子模擬活性區域表面的另一 面,對於形成 Dammarenyl Cation 中間產物極為重要[54]。在胺基酸 序列上 Tyr410 的前一個胺基酸為甘胺酸 ( Glycine ),後一個胺基酸 為天門冬醯胺 (Asparagine),Gly-Tyr-Asn Motif 活性功能區在酵素催 化過程中扮演重要的角色。當 Tyr410 突變成半胱胺酸 (Cysteine),
突變株 CASTyr410Cye 失去 CAS 活性使得產物由環阿屯醇轉變為 75
% 羊毛硬脂醇、24 % 9β-lanosta-7, 24-dien-3β-ol 以及 1% Achilleol A。半胱胺酸在空間上與電子的分佈上與酥胺酸相似,這樣的性質對
於動物及真菌的羊毛硬脂醇的生成極為重要[54]。
而His257 與 Asp483 被認為是 CAS 催化活性上所必須的,Asp483 被認為[52]作為Lewis acid 幫助 Epoxide 開環,協助環化的起始作用。 His257 被認為是作為活性區域的鹼基團,協助碳 19 上的氫與活性區 上的酸基有一質子的來回移動,幫助碳19 的脫氫作用。 CAS His477 雖不位於受質鍵結的活性區域,但在利用電腦模擬 探討雙點突變空間上的結構相關性,認為 His477 位於活性區域外面 的 N 端附近,在空間中相當接近受質鍵結區域,與 Tyr410 有氫鍵的 交互作用,藉著對 Tyr410 的氫鍵拉扯效應,影響環化機制終止反應 前的脫氫作用,因而會生成羊毛硬脂醇,造成一導向演化的結果[51]。
1-3-3 鯊烯-蛇麻烯環化酵素 ( SHC )
細菌的鯊烯環化酶(SHC)與 OSC、CAS 同屬於自然界中的三萜類 環化酵素家族,SHC 與真核生物之 OSC 有類似的反應機制(圖 1-19)。與真核的氧化鯊烯環化酵素比較,細菌的鯊烯環化酵素以鯊 烯為反應受質,被認為是在演化的過程中出現在較早的厭氧時期。真 核細胞以有氧參與的氧化鯊烯反應生成具高度立體選擇性的環狀產 物。因此,只有在演化出現光合作用的生命體後才能發展出將鯊烯環 氧化成為氧化鯊烯的過程。此外,鯊烯環化酵素在反應機構和形態上 都是以較簡單的方式進行。當它進行環化作用時是以”椅形-椅形-椅 形”的構形存在,而有能量上的趨向性使其所需較小的酵素助力。細 菌的鯊烯環化酵素具有較低的受質專一性。環化反應不但可以環化一 般自然界的鯊烯受質,而且對於氧化鯊烯的光學異構物和一般的多萜 醇也可以進行反應。相反地,真核的氧化鯊烯環化酵素則具有很高的 受質結構專一性。 圖1-19 : 同源性的鯊烯環化酵素與氧化鯊烯-羊毛硬脂醇環化酵素,目前在 A. acidocaldarius 中鯊烯-蛇麻烯環化酵素(SHC, EC 5.4.99.x),其X-ray晶體結構、酵素蛋白的活性區域與可能的反應機 制,已經正式發表在 1997 年 Science 期刊[56],這些結構的資訊, 可用作其他同源性環化酵素家族的基本模板依據,並在反應機制與結 構 功 能 上 提 供 重 要 之 相 關 性( 圖 1-20) 。 比 對 Alicyclobacillus acidocaldarius 的 SHC 與人類的 OSC,在蛋白質重要區域的胺基酸 序列有 20 %的相同性 ( Identity ),且有明顯的同源性。希望透過對 SHC結構上的相關資訊,推估OSC環化過程的催化機制。 圖1-20 : SHC 之 X-射線晶體結構圖。C:胺基酸的 COOH 端;N: NH2 端;L:抑制劑接合位置;E:酵素表面唯一的非極性 區域,約 1600 Å2 為受質進入的通道 ( Channel Entrance )。 紅色與黃色之滾筒狀緞帶構形為 α-Helix 的結構;綠色為 β 結構;紫色為 QW-Motifs 之重複區域[56]
先 前 有 研 究 利 用 固 醇 類 抑 制 劑Ro48-8071探討Alicyclobacillus acidocaldarius 的SHC酵素結構與催化功能的相關性[57]。研究指出 Ro48-8071 在 A. acidocaldarius 的 SHC 之 X-射線晶體結構中,與 酵素蛋白相結合的活性區域位置(圖1-21),以及藉由酵素動力學之實 驗數據證明 SHC 與 Ro48-8071 間的抑制關係,繼而探討各活性區 位置與催化上所參與的作用。 Ro48-8071 N H + O O Br F 圖1-21 : 以OSC抑制劑說明環化起始反應。藉由目前有效的 OSC 抑 制劑 Ro48-8071 (灰色) 與 SHC 接合的分子模擬圖,證明 Asp376 為提供質子起始環化反應
研究指出SHC 之蛋白質序列第 374 至 380 個胺基酸以及受質
鍵結凹槽區之芳香族胺基酸 Trp312、Phe365、Trp489、Tyr495、
Phe601、Tyr609 與 Tyr612 為保留區域[56]。在 SHC (A.c) 反應結合區 域,Asp376 提供質子起始環化反應。Glu45 和 Glu95 在氫鍵牽引的 拉扯效應下,利用水分子來吸引質子。His451 和 Asp376 形成氫鍵, 加上 Tyr495 和 Asp376 以一個水分子相維繫,共同增強 Asp376 所 擔任之路易士酸的角色協助起始環化反應。Tyr420 可能與蛇麻烯的 B 與 C 環之形成有關[56]。Trp489 在 A 環形成過程中,以胺基酸側鏈 的 π 電子基團穩定 C-10 碳陽離子。Phe601 可穩定 C-18 位置之碳陽 離子中間物。Asp377 和 Asp374 形成一個空間距離近的氫鍵(2.6 Å),利用胺基酸側鏈所攜帶負電穩定鯊烯的 C-10 碳陽離子[58]。 Trp312 和 Trp489 以及芳香族胺基酸 Phe365、Tyr495、Phe601、 Tyr609 與 Tyr612,利用富含電子的苯環及 Indole 側鏈,經由碳陽離 子- π 電子共振作用(Cation-π interaction) 穩定碳陽離子中間物,加速 環化作用,使得鯊烯能順利被催化重組[59]。突變株 SHCH451A 與 SHCW489A 無新產物之產生,但在 SHC 活性影響方面,突變的結果 卻導致酵素活性下降;在突變株 SHCF601A與 SHCY420A中,除了酵素 活性明顯下降之外,有新產物四環的 17-Isodammara-20 和五環的 Diplopterol 生成。因此,受質鍵結區域之芳香族胺基酸不但可以穩定 過渡態陽離子中間產物,也可能決定產物之生成[49]。
1-4 (氧化)鯊烯環化酵素之胺基酸序列比對
環化酵素家族催化具高度立體選擇性的反應,形成產物專一性,在 反應機制上也有很高的平行性。一些研究指出,環化酵素家族催化的反 應具有下述的特性: (1)具有特殊的蛋白質構型以利於反應受質的接合。 (2)有嚴格的反應機制。反應受質必須結合至酵素上正確的受質結合 區,以及有正確的反應摺疊構形,才能催化開環而有環化起始反應發 生。(3)活性區域之胺基酸有碳陽離子-π電子交互作用以穩定過渡態的中 間產物,因此,可以預防早期環化重組的過程被截斷,確保產物順利生 成[60]。 以生物演化之觀點而言,若序列彼此間皆保留相同之胺基酸,此類 胺基酸可能因為在演化過程中具備重要之功能,因而在不同種類但功能 相似之酵素間被保存下來,如環化酵素家族中芳香環的胺基酸;反之, 序列間相異之胺基酸可能因為演化上較不重要而產生變異,或者是物種 的差異而有所不同,但這些差異有可能就是不同物種之酵素間控制催化 機制的關鍵處。故此,為了探求進一步的資訊,利用相似功能的同一酵 素家族之胺基酸序列比對,得知彼此間的相同性、相似性,及官能基性 質相近之胺基酸在比對序列時會排列於相對應之位置上,用來探討酵素 結構上與功能的相關性。利用 Clustal W 系統將存於 National Center For Biotechnology Information (NCBI) 基 因 庫 中 之 環 化 酵 素 家 族 的 蛋 白 質 序 列 : A.
acidocaldarius 的 SHC、阿拉伯芥( A. Thaliana )之 CAS 與酵母菌 ( S. cerevisiae ) 之 OSC、牛肝(Bovine liver) 之 OSC 及人類(H. sapiens) 之
1 50 H. sapienes OSC MTEGTCLRRRGGPYKTEPATDLGR--WRLNCERGR---QTWTYLQDERAGREQTG 50 B. Liver OSC MTEGTCLRRRGGPYKTEPATDLSR--WRLSNQVGR---QTWTYSQEEDPVREQSG 50 A. thaliana CAS -MWKLKIAEGGSPWLRTTNNHVGRQFWEFDPNLGTPEDLAAVEEARKSFSDNRFVQKHSA 59
S. cerevisiae OSC -MTEFYSDTIG---LPKTDPRLWR---LRTDELGR---ESWEYLTPQQAANDPPS 45
A.acidocaldarius SHC ---MAEQLVEAP-- 9
H. sapienes OSC LEAYALGLDTKNYFKDLP---KAHTAFEGALN-GMTFYVGLQAED-GHWTGDY 98
B. Liver OSC LEAHLLGLDTKSFFKDLP---KAHTACRGALN-GVTFYAALQTED-GHWAGDY 98
A. thaliana CAS DLLMRLQFSRENLISPVLPQVKIEDTDDVTEEMVETTLKRGLDFYSTIQAHD-GHWPGDY 118
S. cerevisiae OSC TFTQWLLQDPK-FPQPHPERNK----HSPDFSAFDACHN-GASFFKLLQEPDSGIFPCQY 99 A.acidocaldarius SHC ---AYARTLDRAVEYLLSCQKDE-GYWWGPL 36
H. sapienes OSC GGPLFLLPGLLITCHVA---RIPLPAGYREEIVRYLRSVQLP-DGGWGLHIEDKSTVFGT 154
Phe104T
B. Liver OSC GGPLFLLPGLLITCHVA---NIPLPAGYREEIIRYLRSVQLP-DGGWGLHIEDKSTVFGT 154
A. thaliana CAS GGPMFLLPGLIITLSITGALNTVLSEQHKQEMRRYLYNHQNE-DGGWGLHIEGPSTMFGS 177
S. cerevisiae OSC KGPMFMTIGYVAVNYIAG---IEIPEHERIELIRYIVNTAHPVDGGWGLHSVDKSTVFGT 156
A.acidocaldarius SHC LSNVTMEAEYVLLCHILD----RVDRDRMEKIRRYLLHEQRE-DGTWALYPGGPPDLDTT 91
His146A
H. sapienes OSC ALNYVSLRILGVGPDDPD--LVRARNILHKKGGAVAIPSWGKFWLAVLNVYSWEGLNTLF 212
B. Liver OSC ALNYVSLRILGVGPDDPD—LVRARNLLHKKGGAVFIPSWGKFWLAVLNVYSWEGLNTLF 212
A. thaliana CAS VLNYVTLRLLGEGPNDGDGDMEKGRDWILNHGGATNITSWGKMWLSVLGAFEWSGNNPLP 237
S. cerevisiae OSC VLNYVILRLLGLPKDHPV--CAKARSTLLRLGGAIGSPHWGKIWLSALNLYKWEGVNPAP 214
Trp232A/R
H. sapienes OSC PEMWLFPDWAPAHPSTLWCHCRQVYLPMSYCYAVRLSAAEDPLVQSLRQELYVEDFASID 272
B. Liver OSC PEMWLFPDWMPAHPSTIWCHCRQVYLPMAYCYSTRLSAEEGPLVQSLRQELYLEDYSCID 272
A. thaliana CAS PEIWLLPYFLPIHPGRMWCHCRMVYLPMSYLYGKRFVGPITSTVLSLRKELFTVPYHEVN 297
S. cerevisiae OSC PETWLLPYSLPMHPGRWWVHTRGVYIPVSYLSLVKFSCPMTPLLEELRNEIYTKPFDKIN 274 A.acidocaldarius SHC PEIMFLGKRMPLNIYEFGSWARATVVALSIVMSRQPVFPLPERARVP--ELYETDVPPRR 207
His234A
H. sapienes OSC WLAQRNNVAPDELYTPHSWLLRVVYALLN---LYEHHHS-AHLRQRAVQKLYEHIVAD 326
B. Liver OSC WAAHRNSVAPDDLYTPHSWLLHVVYAILN---LYERHHS-TSLRQWATQKLYEHIAAD 326
A. thaliana CAS WNEARNLCAKEDLYYPHPLVQDILWASLHKIVEPVLMRWPG-ANLREKAIRTAIEHIHYE 356
S. cerevisiae OSC FSKNRNTVCGVDLYYPHSTTLNIANSLVV---FYEKYLRNRFIYSLSKKKVYDLIKTE 329
A.acidocaldarius SHC RGAKGG---GGWIFDALDRALHG---YQKLSVHPFRRAAEIRALDWLLERQ 252
H. sapienes OSC DRFTKSISIGPISKTINMLVRWYVDGPASTAFQEHVSRIPDYLWMGLDGMKMQGTNGSQI 386
B. Liver OSC DRFTKCISIGPISKTINMLVRWHVDGPASAVFQEHVSRIPDYLWLGLDGMKMQGTNGSQI 386
A. thaliana CAS DENTRYICIGPVNKVLNMLCCWVED-PNSEAFKLHLPRIHDFLWLAEDGMKMQGYNGSQL 415
S. cerevisiae OSC LQNTDSLCIAPVNQAFCALVTLIEEGVDSEAFQRLQYRFKDALFHGPQGMTIMGTNGVQT 389
A.acidocaldarius SHC AGDGSWGGIQPP-WFYALIALKILDMTQHPAFIKGWEGLELYGVELDYGGWMFQASISPV 311
H. sapienes OSC WDTAFAIQALLEAGGHHRPEFSSCLQKAHEFLRLSQVPDNPP-DYQKYYRQMRKGGFSFS 445
B. Liver OSC WDTAFAIQALLEARAQHRPEFWSCLRKAHEYLRISQVPDNFP-DYQKYYRHMSKGGFSFS 445
A. thaliana CAS WDTGFAIQAILAT--NLVEEYGPVLEKAHSFVKNSQVLEDCPGDLNYWYRHISKGAWPFS 473
S. cerevisiae OSC WDCAFAIQYFFVAGLAERPEFYNTIVSAYKFLCHAQFDTECV---PGSYRDKRKGAWGFS 446
H. sapienes OSC TLDCGWIVSDCTAEALKAVLLLQEK--CPHVTEHIPRERLCDAVAVLLNMRNPD----GG 499
B. Liver OSC TLDCGWIVADCTAEALKSILLLQEK--CPFVSNHVPRERLFDTVAVLLSLRNPD----GG 499
A. thaliana CAS TADHGWPISDCTAEGLKAALLLSKVP-KEIVGEPIDAKRLYEAVNVIISLQNAD----GG 528
S. cerevisiae OSC TKTQGYTVADCTAEAIKAIIMVKNSPVFSEVHHMISSERLFEGIDVLLNLQNIGSFEYGS 506
A.acidocaldarius SHC FDNVYYPDVDDTAVVVWALNTLRLP---DERRRRDAMTKGFRWIVGMQSSN----GG 416
Met532A
H. sapienes OSC FATYETKRGGHLLELLNPSEVFGDIMIDYTYVECTSAVMQALKYFHKRFPEHRAAEIRET 559
B. Liver OSC FATYETKRGGHLLELLNPSEVFGDIMIDYTYVECTSAVMQALKTFHKQFPDHRAGEIRET 559
A. thaliana CAS LATYELTRSYPWLELINPAETFGDIVIDYPYVECTSAAIQALISFRKLYPGHRKKEVDEC 588
S. cerevisiae OSC FATYEKIKAPLAMETLNPAEVFGDIMVEYPYVECTDSSVLGLTYFHKYF-DYRKEEIRTR 565
A.acidocaldarius SHC WGAYDVDNTSDLPNHIPFCDFG--EVTDPPSEDVTAHVLECFG---SFGYDDAWKV 467
H. sapienes OSC LTQGLEFCRRQQRADGSWEGSWGVCFTYGTWFGLEAFACMGQTYRDGTACAEVSRACDFL 619
B. Liver OSC LEQGLQFCRQKQRPDGSWEGSWGVCFTYGAWFGLEAFACMGHTYHNGVACAEISRACDFL 619
A. thaliana CAS IEKAVKFIESIQAADGSWYGSWAVCFTYGTWFGVKGLVAVGKTLKN---SPHVAKACEFL 645
S. cerevisiae OSC IRIAIEFIKKSQLPDGSWYGSWGICFTYAGMFALEALHTVGETYEN---SSTVRKGCDFL 622
A.acidocaldarius SHC IRRAVEYLKREQKPDGSWFGRWGVNYLYGTGAVVSALKAVGIDTREP----YIQKALDWV 523
Trp587A
H. sapienes OSC LSRQMADGGWGEDFESCEERRY--LQSAQSQIHNTCWAMMGLMAVRHPDIEAQ--ERGVR 675
B. Liver OSC LSRQMADGGWGEDFESCKQRRY--VQSAQSQIHNTCWALMGLMAVRHPDVAAL--ERGVS 675
A. thaliana CAS LSKQQPSGGWGESYLSCQDKVYSNLDGNRSHVVNTAWAMLALIGAGQAEVDRKPLHRAAR 705
S. cerevisiae OSC VSKQMKDGGWGESMKSSELHSY--VDSEKSLVVQTAWALIALLFAEYPNKEVI--DRGID 678
Tyr707A Phe699A
H. sapienes OSC CLLEKQLPNGDWPQENIAG-VFNKSCAISYTSYRNIFPIWALGRFSQLYPERALAGHP- 732
B. Liver OSC YLLEKQLPNGDWPQENISG-VFNKSCAISYTSYRNVFPIWTLGRFSRLHPDPALAGHP- 732
A. thaliana CAS YLINAQMENGDFPQQEIMG-VFNRNCMITYAAYRNIFPIWALGEYR--CQVLLQQGE-- 759
S. cerevisiae OSC LLKNRQEESGEWKFESVEG-VFNHSCAIEYPSYRFLFPIKALGMYSRAYETHTL--- 731
A.acidocaldarius SHC YLVETQRPDGGWDEPYYTGTGFPGDFYLGYTMYRHVFPTLALGRYKQAIERR--- 631
圖1-22 : OSC、CAS、SHC 的序列比對圖。以 SHC 蛋白結構為模板,
依 SHC 活性區域 29 個胺基酸位置,利用分子模擬與胺基酸
序列比對,對應於人類OSC、牛肝 OSC、酵母菌 OSC、阿拉
伯芥CAS 環化酵素結構上之胺基酸位置 在環化家族酵素家族的蛋白質序列,都有一段獨特的胺基酸重複區 域,其序列通式為 [ (K/R)(G/A) X2-3 (F/Y/W)(L/I/V) X3QX2-5GXW ],一 般稱為 Q-W 活性功能區域(Q-W Motif),在 SHC 重複了八次、而在 OSC、CAS、AMS 與 LUS 中各重複了五次[61] (圖 1-23)。一般認為環化 酵素的Q-W Motif 與酵素催化過程涉及的十幾個鍵的斷裂、生成,以及 和引起的高度放熱反應所釋放的焓(enthalpy)有關,可能是反應受質在 酵素內環化重組所形成鍵能的改變,藉由電子轉移過程吸收反應熱而不 至於破壞酵素結構[62]。而在這些環化酵素中,Tyr 及 Trp 有高度保留性, 配合酵素環化過程可能生成許多高能帶正電中間物的特性,一般認為, 這些胺基酸可能因富含電子的側鏈,能穩定催化過程所生成的高能中間 物,而在演化上被高度保留下來,此推測也符合 Griffin 所提出的
SHC (A.a.) QW6 17 RAVEYLLSCQKDEGYW 32 OSC (S.c.) QW6 79 NGASFFKLLQEPDSGIF 95
CAS (A.t.) QW6 99 RGLDFYSTIQAHDGHW 114 AMS (P.g.) QW6 102 RAVHFFSALQASDGHW 117 LUS (A.t.) QW6 101 RGVRYFTALQASDGHW 116
SHC (A.a.) QW5c 63 KIRRYLLHEQREDGTW 78 OSC (S.c.) QW5 127 ELIRYIVNTAHPVDGGW 143
CAS (A.t.) QW5 139 EMRRYLYNHQNEDGGW 164 AMS (P.g.) QW5 152 EILRYIYCHQNEDGGW 167 LUS (A.t.) QW5 151 EMLRHIYCHQNEDGGW 166
SHC (A.a.) QW5b 243 RALDWLLERQAGDGSW 258 SHC (A.a.) QW5a 335 KAGEWLLDRQITVPGDW 363 SHC (A.a.) QW4 402 KGFRWIVGMQSSNGGW 417 SHC (A.a.) QW3 470 RAVEYLKREQKPDGSW 485 OSC (S.c.) QW3 568 IAIEFIKKSQLPDGSW 583
CAS (A.t.) QW3 591 KAVKFIESIQAADGSW 606 AMS (P.g.) QW3 594 NAVRYLEDTQMPDGSW 609 LUS (A.t.) QW3 591 KAVGFIQDNQTPDGSW 606
SHC (A.a.) QW2 528 KALDWVEQHQNPDGGW 533 OSC (S.c.) QW2 617 KGCDFLVSKQMKDGGW 632
CAS (A.t.) QW2 640 KACEFLLSKQQPSGGW 655 AMS (P.g.) QW2 643 KAVEFLLKSQMDDGGW 658 LUS (A.t.) QW2 640 NGVHFLLTTQRDDGGW 655
SHC (A.a.) QW1 576 RGVQYLVETQRPDGGW 591 OSC (S.c.) QW1 675 RGIDLLKNRQEESGEW 690
CAS (A.t.) QW1 702 RAARYLINAQMENGDF 718 AMS (P.g.) QW1 705 RAAKLLINSQMEDGDF 720 LUS (A.t.) QW1 702 RAAKLIINSQLENGDF 717
圖1-23 : Q-W 區域在環化酵素家族內之分佈情形
A. acidocaldarius ( A.a.) SHC ; S. cerevisiae (S.c.) OSC;A. thaliana ( A.t.) CAS ; P. ginseng ( P.g.) AMS ;A. thaliana

![圖 1-6 : Griffin 提出 Aromatic Hypothesis 理論模組,酵素利用活性 區高度保留的芳香族胺基酸基團,透過碳陽離子-π 共振作 用穩定帶正電的高能產物 [37]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/28.892.157.742.171.467/活性區高度保留芳香族胺基酸基團透過碳陽離子π共振作定帶正電.webp)
![圖 1-8 不同物種 CAS1 基因與不同物種 ERG7 基因高度保留胺基酸 [41]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/30.892.176.699.529.1081/圖18不同物種CAS1基因與不同物種ERG7基因高度保留胺基酸41.webp)
![表 1-1 : 酵母菌(S.c) OSC Val454 突變株產物 [43, 44]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/31.892.135.765.159.1023/表11酵母菌ScOSCVal454突變株產物4344.webp)
![圖 1-11 : 人類 OSC 與膜結合時的構形,圖中黑色部分為抑制劑 Ro 48-8071 結合在中心反應活性結合區,藉此表示催化時反應 物與酵素活性區胺基酸的相關位置 [45] 同屬於三萜類環化家族的 SHC 常被用以做為了解 OSC 環化機制 的結構模組,但此同源性結構模組有其限制性。SHC 是利用演化程 度較早的鯊烯為受質,藉由組織胺酸(Histidine)的殘基增強酸活化打 開雙鍵的起始環化作用,在 B 環為椅形的構形,且藉著 SHC Trp169 的作用,利用碳陽離子-π 電子共振的穩定效應,](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/33.892.168.737.144.725/環化機制結構模組但此同源性結構模組有其限SHC構形電子共振效應.webp)
![圖 1-14 : 酵素活性區在碳 8/碳 9 附近有 7 個芳香族胺基酸基團 --Trp192、Trp230、His232、Tyr237 Tyr503、Phe521、Phe696, His232 與 Tyr503 有氫鍵拉扯效應,使得 His232 能有最適 當的位置,催化最後的脫氫作用而中止反應,且 His232 會 透過和水分子的交互作用,影響催化反應 [45] 。 圖 1-15 : 氧化鯊烯環化 酵素與其產物-羊毛硬脂 醇 形 成 複 合 物 的 結 構 圖。圖中所顯示的胺基 酸基團為距離產物小於](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/37.892.208.704.121.428/PheHis有氫鍵拉扯效應使交互作用影響催化反應氧化鯊烯環化酵素小於.webp)
![圖 1-16 : 紅色箭頭所指為不同物種的氧化鯊烯-羊毛硬脂醇環化酵素 及氧化鯊烯-環阿屯醇環化酵素具有高度保留性,此可能 是催化差異的重要位置 [51]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8243583.171423/39.892.170.718.212.925/醇環化酵及氧化鯊烯環阿屯醇環化酵素具有高度保留性此可異的重要.webp)
