國 立 交 通 大 學
生物科技研究所
碩士論文
探討霍氏格里蒙菌中熱穩定性溶血素之原
始型與突變型生化活性與生物物理特性的
差異
Biological and Biophysical Characterization of
Wild-type and Mutated Thermostable Direct
Hemolysin from Grimontia hollisae
研究生:吳宜芳
Student: Yi-Fang Wu
指導教授:吳東昆 博士
Advisor: Prof. Tung-Kung Wu Ph.D
探討霍氏格里蒙菌中熱穩定性溶血素之原始
型與突變型生化活性與生物物理特性的差異
Biological and Biophysical Characterization of
Wild-type
and
Mutated
Thermostable
Direct
Hemolysin from Grimontia hollisae
研究生:吳宜芳
Student: Yi-Fang Wu
指導教授:吳東昆 博士
Advisor: Prof. Tung-Kung Wu Ph.D
國 立 交 通 大 學
生物科技研究所
碩士論文
A Manuscript of Dissertation
Submitted to Department of Biological Science and Technology
College of Biological Science and Technology
National Chiao Tung University
in partial Fulfillment of the Requirements for
the Degree of Master of Philosophy
in
Biological Science and Technology Hsinchu,
Taiwan, Republic of China
I
中文摘要
熱穩定溶血素(Thermostable direct hemolysin, TDH)在腸炎弧菌中被認為是 最主要的毒性因子。霍氏格里蒙菌所產生的熱穩定溶血素已經成功被表現、純化 及被定義其生化特性。然而由於我們所取得的菌株(G. hollisae BCRC 15890), 其所產生的 TDH 與發表在 NCBI 上的 TDH(G. hollisae 9041)在其胺基酸序列 上有所出入。最主要的差異是位在胺基酸 53, 59 以及 63 號的這三個位置。同時 我們也發現兩者在生化特性上有很大的差異,最明顯的就是在 G. hollisae BCRC 15890 與 V. parahaemolyticus 產生的 TDH 同樣擁有相似的生化性質,而 G. hollisae 9041 則否。當蛋白質加熱至 85oC 以上再經快速降溫過程到 4 度時,此 蛋白質的溶血活性可以回復至先前未經處理時的百分之八十以上;但當蛋白質加 熱在 60o C 與 80oC 之間,快速冷卻後此時的蛋白質卻不具有溶血活性,這一種現 象我們稱之為 Arrhenius effect。此效應存在至今已約 100 年,到現在一直無法被 解釋為何蛋白質有如此特殊的現象。在這裡我們試圖利用此兩種不同性質的蛋白 質來探討令人難以解釋的現象。 首先我們利用分生技術將 53, 59,以及 63 三個位置分別做單點、雙點以及三 點突變。包括野生型總共可以獲得八種蛋白質。接著我們利用 DSC 去探討各個 蛋白質的相轉變溫度。接著利用 CD 觀察各個蛋白質經由升溫所導致相轉變的過 程中其二級結構是如何變化的。最後利用紅血球及 AGS 細胞株觀察各個蛋白質 其對溶血以及對細胞的影響。我們獲得的初步結論是,單一點突變的蛋白質會使 得蛋白質結構較不穩定,但其依然保有 Arrhenius effect;雙點突變 53 及 59 或 59 及 63 位置會使得蛋白質喪失原本具有 Arrhenius effect 的現象,而三點突變的蛋 白質亦會具有相同的結果。而雙點突變在 53 及 63 位置在升溫過程時其二級結構 變化會介於具有 Arrhenius effect 蛋白與不具有 Arrhenius effect 蛋白之間。我們認 為 Arrhenius effect 的有無關鍵與蛋白質結構的穩定度有一定的關聯性。由 CD 結 果得知,擁有 Arrhenius effect 的蛋白質在高溫狀況下其二級結構相較於喪失
Arrhenius effect 蛋白質還要鬆散,所以在經由快速降溫的過程中蛋白質得以重新 摺疊成具有溶血功能的構形;在喪失 Arrhenius effect 的蛋白質中我們發現這些 蛋白質在加熱 60o C 以上時,其二級結構的構形皆會維持在α-helix 且其含量不受 溫度的影響。我們認為這些蛋白質結構在高溫下並沒有被打開,所以在冷卻後其 構形依然維持在 60o C 的纖維化型態(fiber form)。有趣的是,這樣的結果與先前 的報導不盡相同。在細胞毒性以及溶血活性方陎皆以雙點突變 53 以及 63 最為嚴 重。這些蛋白質對於細胞及紅血球有不同程度的傷害性,就先前的研究指出其可 能與結合到細胞膜表陎上的特定物質有關係或與蛋白質形成四聚體(tetramer)的 能力有關係。綜合上結果得知,就 G. hollisae TDH 而言,當其 53, 59,及 63 此三 個位置經突變效應後,確實在其生化特性或結構方陎會受到不同程度的影響。
III
Abstract
The recombinant thermostable direct hemolysin from Grimontia hollisae (G.h-rTDH) exhibited an “Arrhenius effect”, from which it was detoxified by heating at approximately 60-70oC, but reactivated its functional activity by additional heating above 80oC, coupled with a rapid cooling treatment. In order to characterize this paradoxical phenomenon, we compared the tdh gene from the difference species of
Grimontia hollisae listed in a database and used sequence alignment to identify the
critical residues. In addition, we used site-directed mutagenesis to construct the corresponding TDH mutants and obtained various mutated proteins via chromatographic purification. In this study herein, we investigated the individual or collective mutational effect on Tyr53, Thr59, and Ser63 positions of G.h-rTDH to characterize the Arrhenius effect, hemolytic activity, and the biophysical properties of various mutants, respectively. In contrast to the G.h-rTDH wild-type (G.h-rTDHWT) protein, no Arrhenius effect was detected from the G.h-rTDHY53H/T59I and
G.h-rTDHT59I/S63T double-mutants, as well as the G.h-rTDHY53H/T59I/S63T triple-mutant. These mutants also exhibited a different hemolytic activity compared to that observed for the G.h-rTDHWT protein. The differential scanning calorimetry (DSC) results consistently showed that the Arrhenius effect-losing and -retaining mutants exhibited higher and lower endothermic transition temperatures (Tm) than that of the
G.h-rTDHWT, respectively. Circular dichroism (CD) measurements indicated the partial decrease of β–sheet structures near the endothermic transition temperature, consistent with the conformational change of the Arrhenius effect-losing and -retaining protein. Moreover, we also evaluated its toxic effects by assessing their cytotoxic activities against in AGS cells.
謝誌 (Acknowledgement) 兩年多的日子裡,作息規律而踏實,學業由陌生到瞭解。時光匆匆,很快的 碩士生涯結束了,在這裡我學到了老師們以及每位學長姐的經驗與努力。 首先,要先感謝我的指導教授吳東昆老師,感謝老師給我進入實驗室學習的 機會,讓我在兩年的碩士生涯裡,收穫良多。並且由衷的感謝老師在百忙之中指 導我完成論文以及期間給予最大的幫助與鼓勵,讓我在碩士班期間得以順利完成 學業。感謝擔任口試委員的李耀坤老師和廖亦翰老師,感謝您們在百忙之中抽空 前來指導學生,因為您的細心審閱以及寶貴的建議讓我的論文更臻完美,感謝您! 而我的實驗之所以能順利的完成,在這一個過程中,必須要誠心地感謝許多 參與其中並且不吝給予幫忙的很多人。感謝生科系張家靖老師、學亮學長、虹瑋 學妹在 DSC 上給予的幫助與指導。感謝應化系李耀坤老師、黃靜萍博士、貴儀 中心李蘊明小姐、在 CD 上給予的協助,讓我得以在最短的時間內順利完成這部 分的實驗成果。感謝文亮老師在細胞實驗上給予的協助,謝謝您! 接著,感謝本組最資深以及默默指導我們的裕國學長,感謝您為 TDH 組建 立如此深厚的基礎,讓我們得以更快進入狀況,在我們遭遇問題時也與我們共度 難關,讓我們的實驗更加順利!感謝默默撐起 TDH 組一片天的聖慈學姊,感謝您 總是默默的為我們每一個人付出。您,不論在實驗或在生活上總是給予我很大很 大的幫忙;您,在我心中是不可或缺的極重要角色;您的不拘小節以及傳統女性 的犧牲奉獻精神是我最佳的生活導師,感謝您!感謝 Milli 學姊,感謝您在英文上 給予的協助。感謝幽默又充滿智慧的晏任學長,感謝您在實驗與生活上的照顧與 幫忙,感謝您在我低潮時給予的寶貴建議,讓我很快的重新進入正軌,有你真好。 感謝俞靜學妹、彥伶學妹、婉婷學妹,感謝你們幫忙分擔 TDH 的繁重工作,謝 謝你們。這段期間,追隨學長姐們的步伐,讓我感受到身為研究學者的我們,對 於學術應有的熱忱與堅持,有這種感覺,真的很棒!也因為有你們的相伴讓我在 實驗上不孤單寂寞。謝謝你們! 感謝程翔學長、媛婷學姐、晉豪學長、晉源學長、文鴻學長,謝謝您們對實 驗室以及學弟妹們無私的付出與掏心傾力的指導,因為有你們實驗室才會運作的 如次順暢與井然有序,你們真的很重要。非常感謝程翔學長在這期間的照顧與幫 忙,感謝您總是給我信心與鼓勵,讓我有繼續向前走的動力,感謝您陪伴我度過 了不順遂的實驗生活,感謝有您!感謝天昶學長以及已經畢業的禕婷學姊、亦諄 學姊感謝你們在我剛進實驗室時帶領青澀的我認識實驗室,使我能迅速的熟悉並 且融入當中。感謝一起奮鬥的同學們,靜婷、書涵、亦齊,謝謝你們常常提醒迷
V 兒、欣樺、富生,感謝你們分擔實驗室的雜事,讓我們可以全心全意專注在實驗 上,也謝謝你們讓實驗室充滿了歡笑與年輕的活力!感謝依娟學姐、亦榮學長、 建成學長、小新學長、阿伯、冠倫和宗翰學弟,感謝您們在實驗上對我的幫忙! 最後,最重要的,我要感謝我的家人,謝謝你們全力支持我做我想做的事情, 謝謝你們當我心靈上的避風港,你們總是給我許多許多的溫暖與感動,謝謝你們 為我付出那麼多,我愛你們!! 謝謝在這期間支持我、關心我的朋友們,很開心這段回憶中有你們相伴!! 宜芳 2010/07/28
Table of Contents
Chapter 1. General Introduction. ... 1
1.1 Vibrionaceae family ... 1
1.1.1Virio species ... 1
1.1.2 Vibrio parahaemolyticus ... 2
1.1.3Grimontia hollisae. ... 3
1.2 Thermostable direct hemolysin (TDH) ... 3
1.2.1Arrhenius effect ... 7
Chapter 2. Global Research Goals and Design ... 7
Chapter 3. Materials and Methods ... 9
3.1 Bacterial and materials... 12
3.2Construction, expression, and purification of G.h-rTDHWT protein from G. hollisae. ... 12
3.3 Construction, expression, and purification of mutant G.h-rTDHY53A/T59B/S63C protein from G. hollisae ... 15
3.4Assay of hemolytic activity. ... 16
3.5 Analyze thermostability of the G.h-rTDH protein ... 17
3.6Compare hemolytic activity for G.h-rTDHY53A/T59B/S63C. ... 17
3.7 MALDI-TOF-TOF MS analysis ... 17
3.8Difference scanning calorimetry (DSC). ... 18
3.9 Circular dichroism (CD) spectroscopy ... 19
3.10Cell line. ... 19
3.11Morphology examination ... 20
3.12MTTassay ... 20
3.13Thioflavin T floresence assay. ... 21
Chapter 4. Results ... 22
4.1 Cloning, sequence analysis and identification of the G.hollisae tdh gene. ... 22
4.2 Expression, purification, determination and hemolytic activity of G.h-rTDHWT. ... 23
4.3 Identification of G.h-rTDHWT by MALDI-TOF-TOF MS spectrometry. ... 25
4.4 Expression, purification, and hemolytic activity determination of mutated G.h-rTDHY53A/T59B/S63C. ... 27
4.5 The temperature-dependent hemolytic activity analysis and thermostability studies of G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T . ... 28
4.6 Comparison of hemolytic activity for G.h-rTDHY53A/T59B/S63C. ... 30
4.7 Comparison of the hemolytic activity of G.h-rTDHY53A/T59B/S63C mutants at 37oC, 70oC, 90oC. . 31
4.8 Analysis of G.h-rTDHY53A/T59B/S63C thermostability by difference scanning calorimetry... 32
4.9 Analyze the secondary structure change of various G.h-rTDHY53A/T59B/S63C mutants by circular dichroism spectroscopy. ... 34
4.10 Morphology examination and MTT assay for G.h-rTDHY53A/T59B/S63C of the cytotoxicity and cytoviability effect on AGS cells . ... 41
4.11Thioflavin T floresence assay. ... 45
VII
Table of Figures
Figure 1.DNA hybridization test with the tdh gene probe ... 6
Figure 2. Effect of heat treatment on conformation of TDH. ... 8
Figure 3. Model of heat-induced conformation change of TDH. ... 8
Figure 4.The flow chart lists the strategy ... 11
Figure 5. The sequence alignment of tdh gene. ... 23
Figure 6. Purification and characterization of the G.h-rTDHWT protein. ... 24
Figure 7.MALDI-TOF-TOF MS analyze ... 26
Figure 8. SDS-PAGE、native PAGE and hemolytic assay analyze. ... 28
Figure 9. Thermostability assay of G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T. ... 29
Figure 10.Hemolytic activity of G.h-rTDH mutant ... 30
Figure 11. The thermostability assay of G.h-rTDH mutant... 32
Figure 12. The DSC result. ... 34
Figure 13.(A) The CD spectrum of G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T ... 36
Figure 13. (B) The CD spectrum of G.h-rTDHY53H/T59I/S63T, G.h-rTDHY53H/T59I and G.h-rTDHT59I/S63T which are deficiency the Arrhenius effect. ... 37
Figure 13.(C) The CD spectrum ofG.h-rTDHY53H, G.h-rTDHT59I and G.h-rTDHS63T which are process the Arrhenius effect ... 38
Figure 13.(D) The CD spectrum of G.h-rTDHWT, G.h-rTDHY53H/T59I/S63T, and G.h-rTDHY53H/S63T. ... 39
Figure 13.(E) The CD spectrum of all mutated G.h-rTDHY53A/T59B/S63C proteins ... 40
Figure 14. Cell morphology ... 44
Figure 15.MTT assay results of various G.h-rTDHY53A/T59B/S63C mutants. ... 45
Figure 16.Thioflavin T floresence assay result ... 46
Table of Tables
Table 1.Pathogenic Vibrio species associated with human infections ... 2Tabel 2. Sequences of PCR primers and PCR condition for PCR amplification of tdh gene.. ... 14
1
Chapter 1
General Introduction
1.1. Vibrionaceae family
The Vibrionaceae are a family of Proteobacteria. They are gram-negative organisms and belong to facultative anaerobes, capable of fermentation. The family Vibrionaceae currently comprises six validly published genera: Aliivibrio, Salinivibrio,
Enterovibrio, Grimontia, Photobacterium, and Vibrio.
1.1.1 Vibrio species
Many Vibrio species are pathogenic to humans and have been implicated in food-borne disease. Vibrio infections are more frequently encountered in coastal states, apparently due to the greater consumption of raw or uncooked shellfish. Vibrio spp. distinct from Vibrio cholerae and Vibrio mimicus, do not grow in media that lack added sodium chloride, and are referred to as “halophilic”.
The Vibrio species with the most medical significance include Vibrio cholera, Vibrio
parahaemolyticus, Vibrio vulnificus, and Grimontia hollisae. Major clinical
manifestations of infection by this type of organism include primary septicemia, gastroenteritis, and wound infections1. Primary septicemia and gastroenteritis are usually caused by ingesting contaminated seafood. Generally, the symptoms of gastroenteritis include diarrhea, vomiting, and abdominal pain, where antibiotics are needed when infection is present. Moreover, primary septicemia is also a disease of the circulation system, caused by invasion of the bacteria into the blood vessels from hepatic portal vein or the intestinal lymph system. Patients with primary septicemia might have fever, chills, hypotension on presentation, nausea, vomiting, diarrhea,
abdominal pain, and skin lesions2. In addition, if injured skin comes into contact with the contaminated marine life or seawater, it could cause wound infections, edema, erythema, and blain.
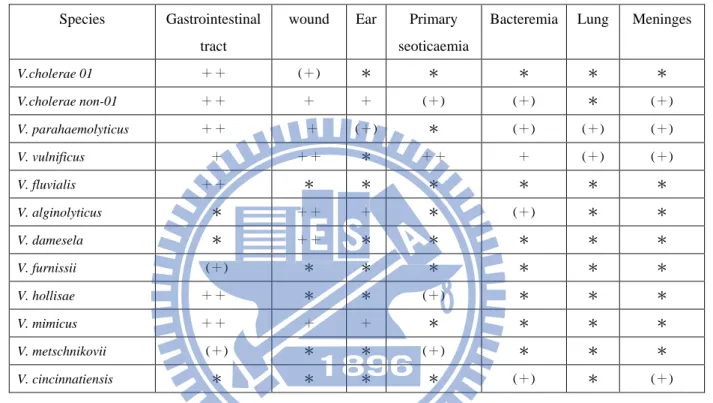
Table 1. Pathogenic Vibrio species associated with human infections1.
NOTE : ++, most common site of infection ; +, other sites of infection ; (+), rare sites of infection ; *, infection remains to be firmly establish.
1.1.2 Vibrio parahaemolyticus
V. parahaemolyticus, a common pathogenic bacterium of food-borne
gastroenteritis in people, is frequently isolated from a variety of marine organisms consumed as seafood. V. parahaemolyticus was first described as the cause of gastroenteritis in Japan, and was first found in the United States by Baross and Liston (1968) in the estuarine waters of Puget Sound3. Between 1971 and 1978, V.
Species Gastrointestinal tract
wound Ear Primary seoticaemia
Bacteremia Lung Meninges
V.cholerae 01 ++ (+) * * * * * V.cholerae non-01 ++ + + (+) (+) * (+) V. parahaemolyticus ++ + (+) * (+) (+) (+) V. vulnificus + ++ * ++ + (+) (+) V. fluvialis ++ * * * * * * V. alginolyticus * ++ + * (+) * * V. damesela * ++ * * * * * V. furnissii (+) * * * * * * V. hollisae ++ * * (+) * * * V. mimicus ++ + + * * * * V. metschnikovii (+) * * (+) * * * V. cincinnatiensis * * * * (+) * (+)
3
and lobster infection, which might be caused by the consumption of raw or insufficiently heated seafood, or by consumption the properly cooked food but still be contaminated after its cooking4,5. V. parahaemolyticus has a worldwide distribution in estuarine and coastal environments, and has been isolated not only from many species of fish, shellfish, and crustaceans, but also from the cases of gastroenteritis in humans. Among the pathogenic factors, V. parahaemolyticus TDH has been considered as a major virulence factor in the case of gastroenteritis6.
1.1.3 Grimontia hollisae
Grimontia hollisae, a species of Vibrionaceae, which was first described by Hickman
et al. and recently reclassified by Thompson et al. causes moderate to severe cases of gastroenteritis including diarrhea and abdominal pains7,8. This organism was reported as typically not growing on TCBS agar or MacConkey agar, but does grow well on sheep blood agar and marine agar8. Grimontia hollisae was found with much greater frequency in samples taken from clinical cases of gastroenteritis, acute diarrhea, bacteremia, and septicemia9-13. Although epidemiological evidence8,9 and the halophilic nature of the vibrio suggested that G. hollisae is an organism of marine origin, to the best of our knowledge, the isolation of G. hollisae from the marine environment is not the only way for acquisition. Some Vibrio species can also cause infections in humans and have been isolated from a variety of intestinal and extra- intestinal sites14.
1.2 Thermostable direct hemolysin (TDH)
Many important pathogenic factors of Vibrio, such as hemolysins, proteases, hemagglutinins, and other hydrolytic exoenzymes, have been reported to contribute to virulence, or to facilitate the disease process15,16. Among these factors, hemolysin is
known to be the most fearful virulence factor, which is involved in the gastrointestinal disorders caused by V. parahaemolyticus. V. parahaemolyticus is commonly isolated from the cases of gastroenteritis in humans, and thermostable direct hemolysin (TDH), has further been considered as a major virulence factor in the cases of gastroenteritis. This protein has been confirmed to contain hemolytic, cytotoxic, enterotoxic, lethal (in mice), and cardiotoxic activities17.
TDH is a pore-forming toxin of approximately 2 nm in size on erythrocyte membranes and causes colloidal osmotic lysis. TDH consists of 165 amino acid residues to perform a variety of biological activities including hemolytic activity, cytotoxicity, cardiotoxicity, and enterotoxicity. TDH with toxic effects was identified from a variety of Vibrio species, including V. cholera non-O1, V. parahaemolyticus, V.
mimicus, V. alginolyticus, and G. hollisae, and was proposed as a major virulence
factor of V. parahaemolyticus18. Although scientists have originally studied its hemolytic properties, TDH has been long suspected to be an enterotoxin involved in most cases of V. parahaemolyticus diarrhea. V. parahaemolyticus has been recognized as an agent of gastroenteritis associated with the consumption of seafood, but not all strains of this species are considered to be truly pathogenic. An important virulence factor that has been considered in V. parahaemolyticus gastroenteritis is TDH. TDH is one putative virulence factor that has been epidemiologically associated with disease, and it is one of the hemolysins produced by V. parahaemolyticus that produces beta-type hemolysis on a special blood agar medium, Wagatsuma agar9,19. Almost all
V. parahaemolyticus strains isolated from clinical specimens demonstrated this
hemolytic activity, which has been called the Kanagawa phenomenon (KP), only 1-2% of strains from nonclinical sources are KP positive19,20. Accordingly, this
5
secretion, as well as cytotoxicity in a variety of cell types21. The effects of the toxin on human amniotic membrane cells (FL cells) have been characterized by loss of viability and by some morphological changes, including the disappearance of microvilli from the cell surface, degeneration of the cytoplasm, and disintegration of the nucleus22. Furthermore, the Ca2+-independent cytotoxicity of TDH in a human embryonic cell line (Int407) was also reported, where it caused damage to plasma membranes and lysosomes, as well as induced a cellular degeneration in the form of large transparent blebs23. Although the TDH virulence factor has been studied, the mechanism of virulence responsible for these activities has not been fully elucidated.
TDH is detoxified by aggregation into fibrils, formed after being heated at 60-70 °C, which can be reversibly refolded into the toxic native form by being rapidly cooled after unfolding at higher temperatures24. Transmission electron microscopy further indicates the nature of the fibrillar structure of TDH (TDHi). These fibril formed structures show both the property of the nucleation-dependent elongation, and the fluorescent incensement from its thioflavin T fluorescence. Formation of β-rich structures of TDH were also observed in the presence of lipid vesicles containing ganglioside GT1b25, a putative TDH receptor. Congo red, which is a well-known dye sensitive for detection of amyloid fibrils, was found to inhibit the hemolytic activity of TDH in a dose-dependent manner. These findings support the idea that the conformational change in TDH structure, with the increase in its β-sheet content, in a cellular membrane, may be associated with its cytotoxicity24.
In 1988, it was reported that a vibrio isolated from the intestine of a coastal fish was identified as G. hollisae by its biochemical characteristics and by reaction with a gene probe for the thermostable direct hemolysin of V. parahaemolyticus (Figure 1). The hemolysin produced by the isolate from the coastal fish had traits identical to those of the thermostable direct hemolysin-like hemolysin produced by a clinical
strain of G. hollisae26 (Figure 1B). The hemolysis characteristics of TDH are fascinating and also thought to be responsible for the virulence of G. hollisae. G.
hollisae has been reported only in clinical cases (largely from diarrheal stools). G. hollisae has also been reported to be isolated from the coastal environment. This
finding is important in that it presents evidence to support previous suggestions that G.
hollisae infection originated from ingestion of contaminated food or contact with the
environmental reservoir. Recently, patients with severe gastroenteritis and hypovolemic shock symptoms have been identified to be infected with G. hollisae and no other enteric pathogens, suggesting a likely underestimation of the incidence of G.
hollisae infections2726.
FIGURE 1: (A) DNA hybridization test with the tdh gene probe. A: Kanagawa phenomenon-positive V. parahaemolyticus; B: Kanagawa phenomenon-negative V.
parahaemolyticus; C: G. hollisae ATCC 33564; D: G. hollisae KUMA871. (B)
Ouchterlony immunodiffusion analysis of TDH-like hemolysin produced by G.
hollisae. A: anti-TDH rabbit serum; B: purified TDH; C: concentrated culture
7
1.2.1 Arrhenius effect
As early as 1907, staphylococcal alpha-toxin was found to show the Arrhenius effect, and the mechanism of this effect has been studied by several researchers28. For staphylococcal alpha-toxin, Arrhenius reported that it was inactivated by heating at 70oC, but was reactivated by heating at 100oC28. This phenomenon has been described as the Arrhenius effect, and several workers have studied this Arrhenius effect through the investigation of staphylococcal alpha-toxin, proposing the existence of some substance which interacts with alpha-toxin at 60oC28. On the other hand, some reports indicated that active alpha-toxin aggregates at 60oC to an insoluble, nontoxic form, whereas at higher temperatures soluble active toxin is released. Crude hemolysin of V. parahaemolyticus shows an Arrhenius effect similar to that of staphylococcal alpha-toxin28-31. In the thermostable direct hemolysin (TDH), a major virulence factor of V. parahaemolyticus is detoxified by heating at approximately 60-70oC, but is reactivated by additional heating above 80oC. This paradoxical phenomenon has been shown in several strains of V. parahaemolyticus even though it still remained unexplained for almost 100 years24. The previous study demonstrated that the Arrhenius effect in the TDH from V. parahaemolyticus is related to structural changes from a soluble form into a fibrils form. The native TDH (TDHn) is transformed into the nontoxic fibrils rich in β-strands by incubation at 60oC (TDHi). The TDHi fibrils are dissociated into unfolded conformations by further heating above 80oC (TDHu). The rapid cooling of TDHu results in the refolding of the protein into the toxic TDHn, whereas the protein is trapped in the TDHi structure by a slow cooling of TDHu (Figure 2). TDHi, with fibrillar structure has no hemolytic activity at 37oC, consistent with the Arrhenius effect. When TDHi fibrils are incubated above 80oC they dissociate into unfolded states, which can further refold into toxic TDHn upon rapid cooling to 37oC (Figure 3). This is an unusual phenomenon because the
formation of inactive protein aggregation is generally irreversible.
FIGURE 2: Effect of heat treatment on the conformation of TDH. (A) Relative hemolytic activities of TDH, measured at 37°C after various heat treatments (n = 5 per group) (B) CD spectrum of TDH at 37°C after rapid cooling(∆). The spectrum is identical to that of TDHn (dashed line). (C) CD spectrum of TDH at 37°C after slow cooling (O). The spectrum was identical to that of TDHi at 57.5°C (solid line)24.
FIGURE 3: Model of heat-induced conformational change of TDH. (A) Rapid heating and cooling. (B) Slow heating and cooling24.
9
Chapter 2
Global Research Goals and Design
TDH widely distributed in the strain of Grimontia hollisae and a few Vibrio species, has a variety of biological activities in animals, including hemolytic activity, cytotoxicity, and enterotoxicity in mice32. The information of physiochemical and biophysical properties of G.h-rTDH, however, have not been well reported. In this study, we aim to analyze the physiochemical and biophysical characterization of
G.h-rTDH. First, the G.h-rTDH was cloned from a commercial Grimontia hollisae
strain, BCRC 15890. The amino acid sequence of the cloned G.h-rTDH was compared with that of the published tdh gene, Grimontia hollisae 9041. From the results of sequence alignment, three distinct amino acid changes, i.e. Tyr53His53, Thr59Ile59, and Ser63Thr63, between tdh gene from G. hollisae BCRC 15890 and G. hollisae 9041, were observed and examined, attributed with the physiochemical and biophysical characteristics.
As mentioned previously, the TDH of V. parahaemolyticus has the ability to revert to a native form via a rapid-cooling treatment after it was unfolded at high temperatures, without any assistance of other enzymes or chemical compounds24.
G.h-rTDHWT also can display the Arrhenius effect, as the protein physiochemical characterization of V. parahaemolyticus TDH has revealed. Interestingly the
G.h-rTDHY53H/T59I/S63T lost this Arrhenius effect. It is reasonable to speculate on the interesting points between the recovery ability and the protein structure.
The functional analysis of the amino acid residues is the most essential study to characterize the relationship between the protein function and critical amino acids. The thermostability of G.h-rTDH might change when mutants are created. In other
words, the substitution of the amino acid residues might influence the secondary structure directly, and further affect its tertiary structure. Otherwise, it might not change the protein conformation but only permit the energy unfavorable for recovery to its native form. Briefly, by analyzing the changes in activity of
G.h-rTDHY53A/T59B/S63C (A=Y or H; B=T or I; C=S or T) mutants, the secondary structure and biophysical relationships will be better understood and elucidated.
Previous reports revealed an effect of a particular mutation on TDH from which a mutant toxin of TDH was formed from V. parahaemolyticus, R7, which has a single amino acid substitution of serine for glycine 62, constructed to show the deficiency in its hemolytic activity33. Thus, in my experimental design, G.h-rTDHY53A/T59B/S63C may be influencing its hemolytic activity and cytotoxicity via mutagenesis effects. In this work herein, we will study the toxic characterization of G.h-rTDHWT and
G.h-rTDHY53A/T59B/S63C and propose to understand the mechanism of the Arrhenius effect with the strategy shown in Figure 4.
11
FIGURE4. The flow chart describes a strategy for analyzing the relationship between protein secondary structure and biophysical characterization when the Tyr53, Thr59, or Ser63 positions of G.h-rTDHWT were substituted into various combinations of amino acid substitutions.
Chapter 3
Materials and Methods
3.1 Bacterial strains and materials
Grimontia hollisae (BCRC 15890) was obtained in a freeze-dried form from the
Culture Collection and Research Center (CCRC, Hsin-Chu, Taiwan). The bacteria were cultured in a Tryptic Soy Broth (TSB, Difco, Detroit, MI) medium, which was supplemented with 1.5% NaCl and incubated at 37°C overnight, with shaking (180 cycles/min). This strain showed the hemolytic phenomenon on agar plates containing 5% sheep erythrocytes. Phenyl Sepharose 6 Fast Flow was purchased from Amersham Pharmacia Biotech (Piscataway, NJ, USA).
3.2 Construction, expression, and purification of G.h-rTDH
WTprotein
from G. hollisae
G. hollisae were cultured in 3 mL Tryptic soy broth (TSB) medium with 3 %
sodium chloride (NaCl) at 37oC with continuous shaking for 12 h. Cultures were harvested by centrifugation at 10,000 x g for 1 min at room temperature. The supernatant was removed and the genomic DNA was extracted from the pellets using QIAamp DNA Mini Kit, following the manufacturer’s protocol (Qiagen). According to the information derived from the database from National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov) databases, the tdh gene was cloned from the Grimontia hollisae strain with two primers. The PCR conditions were similar to the general protocol previously published by our research group (Table 2)34.
13
autosequencer, according to the manufacturer’s protocol (Applied Biosystems, Foster City, CA). A recombinant plasmid harboring the tdh gene was transformed into
Escherichia coli BL21 (DE3) (pLys S) cells by heat shock. Transformants were
cultivated at 37°C with rotary shaking in Luria-Bertani Broth (Difco) supplemented with 50 μg/mL kanamycin, and the culture was incubated for another 16 h. The cells were then harvested by centrifugation at 6,000 x g for 30 min, and resuspended in 15 mL of 20 mM Tris-HCl buffer (pH 7.0). The mixture was sonicated, and the cell debris was removed by centrifugation at 12,000 x g for 30 min at 4°C. Then the crude protein solution was loaded onto a Phenyl-Sepharose 6 Fast Flow column prequilibrated with 20 mM Tris-HCl buffer (pH 7.0) and eluted with a linear 0% to 50% ethylene glycol gradient. Fractions exhibiting G.h-rTDH activity were pooled, and added with NaCl to 200 mM concentration. The G.h-rTDH was again applied to a Phenyl-Sepharose 6 Fast Flow column with 20 mM Tris-HCl buffer (pH 7.0) containing 200 mM NaCl and then eluted with 4-fold volumes of a step gradient consisting of 200, 100, 50 and 20 mM NaCl in 20 mM Tris-HCl (pH 7.0), respectively, and sole equilibrating buffer without any salt concentration. Pure protein eluted with 20 mM Tris-HCl buffer (pH 7.0). Then, TDH protein was dialyzed against 10 mM phosphate-buffered saline buffer (PBS, pH 7.0) overnight for a hemolytic activity assay.
Table 2. Sequences of PCR primers and PCR condition for PCR amplification of tdh gene.
Gene
G. hollisae (BCRC 15890) PCR primer sequence 34 tdh F 5’- ATGAAATACAGACATCT -3’ R 5’- TTATTGTTGAGATTCAC -3’ PCR Condition34 Temperature (oC) Time Denaturation 94 oC 5 min Denaturation 94 oC 15 sec 35 cycles Annealing 58°C 1 min Extension 72°C 1 min Extension 72°C 10 min
15
3.3 Construction,
expression,
and
purification
of
mutant
G.h-rTDH
Y53A/T59B/S63C protein from G. hollisae
The mutant G.h-rTDHY53A/T59B/S63C was constructed by site-directed mutagenesis, using the recombinant plasmid harboring the tdh gene as template (tdh gene in pCR®2.1-TOPO®) and using two primers. The PCR condition, purification, and expression method of the mutant G.h-rTDHY53A/T59B/S63C protein were the same as that for G.h-rTDHWT (Table 2). The primers for construction of mutant
G.h-rTDHY53A/T59B/S63C plasmid are shown in Table 3.
Table 3. The primers sequence for construct of G.h-rTDHY53A/T59B/S63C plasmid
tdh gene tempelate
(tdh gene in pCR®2.1-TOPO®)
PCR primer sequence
tdhY53H tdhWT F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer) R 5’-AAAgATgTTCACggACAATCAgTCTTCACA-3’ tdhT59I tdhY53H/T59I F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
R 5’-TACAAAgATgTTTATggACAATCAgTCTTCACA-3’ tdhS63T tdhY53H/S63T F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
R 5’-TACAAAgATgTTTATggACAATCAgTCTTCACA-3’ tdhY53H/T59I tdhY53H/T59I/S63T F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
R 5’-ATAACgTCAggTTCTAAATggTTAACATCC-3’ tdhY53H/S63T tdhY53H/T59I/S63T F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
R 5’-ggACAATCAgTCTTCACAACgTCAggTACT-3’ tdhT59I/S63T tdhWT F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
R 5’-ggACAATCAgTCTTCATAACgTCAggTACTAAA-3’ tdhY53H/T59I/S63T tdhY53H F 5’-GTAAAACGACGGCCAG-3’ (M13 forward primer)
3.4 Assay of hemolytic activity
Hemolytic activity was determined on the human erythrocytes. Human erythrocytes were first washed with 100 mM PBS buffer (pH 7.0) 3 times, and then resuspended to a final concentration of 4% (v/v) in PBS buffer. For the hemolytic activity assay, 0.1 mL of 0.1% Triton X-100, which caused complete release of hemoglobin from erythrocytes and resulted absorbance change at 570 nm, was used as a positive control. Aliquots of 0.1 mL of 100 mM PBS buffer (pH 7.0) were used as negative controls. Different concentrations of the protein solution (0.1 mL) were added to the solution of human erythrocytes (0.1 mL). After incubation at 37°C for 1 h, the reaction mixtures were centrifuged at 800 x g for 5 min, and the 0.1 mL supernatant was packed. The amount of hemoglobin released from the disrupted erythrocytes was quantified by spectrophotometry on an ELISA reader at 540 nm. The 100% hemolysis activity was defined as the 570 nm absorption, with the hemoglobin released from erythrocytes treated with 0.1% Triton X-100. The equation for hemolytic activity assay is as follows:
Hemolytic activity (%) =
(protein O.D570 value – negative value)/(positive value – negative value) x 100
In parallel, the G.h-rTDH was subjected to 10% native polyacrylamide gel electrophoresis, and then embedded onto the agar plate containing 5% sheep erythrocytes. The blood agar plate was incubated at 37°C for an appropriate amount of time to visualize a suitable signal. In addition, G.h-rTDH proteins were also electrophoresed on a 15% SDS-PAGE and stained with Coomassie brilliant blue for
17
3.5 Analyze thermostability of the G.h-rTDH protein
The effect of temperature on hemolytic activity of purified G.h-rTDH was determined by incubating 1 μM of the purified protein in 0.1 M PBS buffer (pH 7.0) for 30 min at different temperatures (4°C, 16°C, 25°C, 30°C, 37°C, 45°C, 50°C, 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C, 90°C, 95°C and 100°C), and then assayed for residual hemolytic activity on 4% human erythrocytes. The hemolytic activity assay method and the equation were described in section.3.4.
3.6 Compare hemolytic activity for G.h-rTDH
Y53A/T59B/S63CThe G.h-rTDHY53A/T59B/S63C mutant was prepared at 200 μg/mL by the described xpression and purification method in section 3.3. A dilution series of protein with half concentration dilutions were incubated with 0.1 mL of 4% human erythrocytes at 37°C for 1 h. After centrifugation at 3,000 rpm for 5 min, the supernatant was measured at 570 nm. The hemolytic activity assay method and the equation were described in section 3.4.
3.7 MALDI-TOF-TOF MS analysis
The SDS-PAGE band corresponding to G.h-rTDHWT was subjected to in-gel trypsin digestion. The in-gel trypsin digestion experiment, excision of protein bands from polyacrylamide gels and the gel particles were prepared for in-gel digestion and washed with 50 μL wash buffer (10mM NH4HCO3, 50% ACN) for 15 min. All
remaining liquid was removed, and 100 μL ACN was added to cover the gel particles for 20 min. When the gels shrink and stick together, 3 μL of trypsin (20 ng/mL) was added to the gel and incubated for 1 h at 4oC, and then incubated at 37oC overnight. The reaction was stopped with 1% TFA, and 10 min sonication, supernatant recovered, then the sample was directly mixed with MALDI matrix (CHCA, 20 mg/mL in 50%
ACN, 0.1% TFA), and analyzed using an autoflex III (BRUKER). After a default calibration, all MS spectra were recorded in positive reflector mode within a mass range of m/z 500–4000. For an initial MS scan, 4 subspectra with 200 shots per subspectrum were accumulated for each spot using a random search pattern. Spectral peaks were included in the acquisition list for the MS/ MS run of the result-dependent experiment, if they met threshold criteria (S/N above 6). For MS/MS experiments, 2,000 shots per spectrum were accumulated. Subsequently, all acquired MS/MS spectra were searched against the Swiss-Prot database using the MASCOT search engine (biotools, v3.1). Search parameters for peptide and MS/MS mass tolerance were 100 ppm and 100 ppm, respectively, with the allowance for one missed cleavage made from the trypsin digest. The search mode was carried out to identify the variable modification of oxidation (M), and Carboxymethyl (C) groups at the C terminus. Proteins were identified by PMF and MS/MS with MASCOT, which corresponds to p<0.05.
3.8 Difference scanning calorimetry (DSC)
DSC measurements were performed using the DSC-Q10 (TA instruments). The DSC-Q10 was run without feedback and 10 min equilibration times at 25oC were used as previously described. The protein was scanned from 25oC to 95oC at a heating rate of 0.5 oC/min. A pan containing 10 mM Kpi buffer, pH 7.0 was used as a reference. The sample and reference cells of optical operational volume of 0.5 mL were used. Protein samples were concentrated to 0.36 mg/mL in 10 mM Kpi buffer (K2HPO4, pH
7.0). DSC data were corrected for instrument baselines and normalized for scan rate and protein concentration. Data obtained for TDH protein were analyzed with the TA
19
3.9 Circular dichroism (CD) spectroscopy
Circular dichroism (CD) spectra were recorded with a J-815 spectropolarimeter (JASCO, Tokyo, Japan) equipped with a thermoelectric temperature controller. Data were processed with software provided by JASCO. Measurements were taken in a quartz cuvette with a path length of 1 mm, and scanned in the interval of 0.2 nm at a rate of 50 nm/min. The data from 6 individual replicates were averaged. The protein concentration was 0.18 mg/mL in 10 mM Kpi buffer (pH 7.0) for the measurement of far-UV (190-250 nm) CD spectrum. The experimental temperatures were 37°C, 50-60°C, 70°C, 75°C, 80°C, 85°C, 90°C, 95°C, and its Tm value, respectively (the Tm
value was determined from DSC data). Before measurement, the sample was pre-equilibrated at each experimental temperature for 5 min. The mean residue ellipticity, [Θ], was calculated by using the following relationship:
[Θ] was expressed in degrees squared centimeters per decimole.
3.10
Cell line
The AGS cell line (BCRC 60102) is a human stomach adenocarcinoma cell taken from a 54-year-old female Caucasian. Cells were maintained in RPMI Nutrient Mixture (Gibco) supplemented with fetal calf serum (10%, v/v, Gibco) and penicillin streptomycin (1%, v/v, Gibco). Cells were incubated at 37°C in an incubator of 5%
[Θ]
MRW = [Θ] / ( 10 x Cr x l )Cr is the mean residue molar concentration l is the cell path in cm
Cr = ( n x 1000 x cg ) / Mr
n is the number of peptide bonds ( residue ) cg is macromolecule concentration ( g / mL ) Mr is the molecular weight of species ( KDa )
CO2 in air. Every 2 to 3 days the culture medium was renewed, and doubling time of
this cell line was 20 h.
3.11
Morphology examination
The AGS cells were first cultured in a 6-well plate overnight. Before examination, 100 mM PBS buffer (pH7.0) was used to wash the cells twice, then cells were mixed with 10 g/mL G.h-rTDH in RPMI complete medium for 30 min at 26oC. Subsequently, the images of the cell morphology were recorded with the 200x magnification by an Olympus digtal camera.
3.12
MTT assay
For cytoviability, the MTT assay has been widely used as a colorimetric assay for measuring the activity of mitochondrial dehydrogenase that reduce MTT, a tetrazolium dye: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, substrate to form formazan, which is generated only by living cells but not dead cells. In this assay, AGS cells were inoculated in 200 uL of complete growth medium at a concentration of 2 x 104 cells in a 96-well microtiter plates. Plates were incubated for 24 h at 37oC in an atmosphere of 5% CO2 in air. Different concentrations of
G.h-rTDH were added and treated for 1 h before the supernatant was removed via a
centrifugation separation. Fresh MTT solution (5 mg/mL) was dissolved in PBS and filtered by a 0.22 filter, and was diluted 10-fold in a complete medium. A 100 ul aliquot of the above solution was added to each well. Plates were wrapped by the aluminum foil and incubated for a further 5 h at 37 oC. MTT was then removed from the wells, and the formazan crystals were dissolved in 200 ul of Dimethyl sulfoxide
21 experiments.
3.13
Thioflavin T florescence assay
Thioflavin T (ThT) is a benzothiazole compound that possesses the light-emitting component of leuciferin. It can be utilized to visualize the amyloid beta content of protein in the solution. ThT binds to amyloid fibrils and the florescence intensity can identify the fibril content by florescence spectra. A 10 g sample of G.h-rTDH protein was respectively and sequentially heated to 37oC, 70oC and 90oC for 15 min and then subjected to rapid cooling in ice water for 15 min.
Before measuring the florescence of the fibril content in the solution, ThT and Tris-HCl buffer were premixed. The ThT/Tris solution was as described: ThT powder was dissolved in distilled water and the final concentration was 1 mg/mL (3.14 mM). Then, 1.6 l of ThT solution was added to 1 mL 50mM Tris-HCl, pH7.0 buffer. Before measurement, an aliquot of 200 l ThT/Tris solution was mixed with thermally pre-treated protein, the fiber forming protein containing beta sheets became immediately bound to ThT. The florescence of Thioflain T was measured at 460-600 nm when excited at 450 nm using fluorescence spectrophotometry (Hitachi, F-7000). The Thioflavin T was obtained from Sigma (St. Louis, MO).
Chapter 4
Results
4.1 Cloning, sequence analysis and identification of the G. hollisae tdh
gene
The nucleic acid sequences of tdh gene from various Vibrio species were aligned and analyzed to find the highly conserved sequences. The primers for the conserved gene were thus designed, and used in the polymerase chain reaction (PCR) to amplify the putative tdh gene from G. hollisae BCRC 15890 genomic DNA. The construct plasmid carry out tdh-mutated gene was obtained from the lab. The PCR amplified G.
hollisae tdh gene is 570-bp in size and encodes a polypeptide of 189 amino acids
involving a signal peptide with 24 amino acids in the N-terminal region, and a mature protein of 165 residues, with a predicted molecular mass of 18,616.9 Da. Notably, three distinct amino acid changes, i.e. Tyr53His53, Thr59Ile59, and Ser63Thr63, were observed from the sequence alignment between the PCR amplified tdh gene from G. hollisae BCRC 15890 genomic DNA (assigned as
G.h-rTDHWT) and the historically published tdh gene from G. hollisae 9041 in the NCBI data bank (assigned as G.h-rTDHY53H/T59I/S63T). Amino acids sequence alignment between the two tdh genes is shown in Figure 5.
23
FIGURE 5. The sequence alignment of putative tdh genes between the PCR amplicon
tdh gene from G. hollisae BCRC 15890 genomic DNA (assigned as G.h-rTDHWT) and the historically published tdh gene, from G. hollisae 9041 as described in the NCBI data bank (assigned as G.h-rTDHY53H/T59I/S63T). The sequence of the signal peptide with 24 amino acids in the N-terminal is shown in red. The distinct amino acid change between G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T are red boxed.
4.2 Expression, purification, determination and hemolytic activity of
G..h-rTDH
WTThe wild type G. hollisae tdh gene G.h-rTDHWT was cloned into the plasmid pCR®2.1-TOPO® and subsequently transformed into Escherichia coli
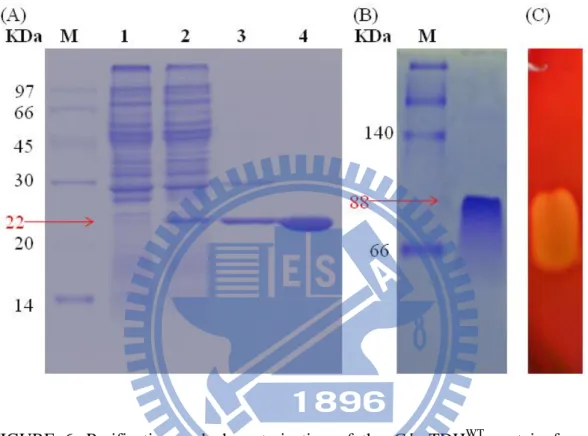
BL21(DE3)(pLysS) cells for protein expression. Following the incubation for 16 h at 37oC, the harvested cells were sonicated for the expressed protein purification using a Phenyl-Sepharose 6 Fast Flow column. After the first round of chromatographic purification for the separation of impurities from crude extraction, the homogenous protein with a molecular mass of approximately 22 kDa, as resolved by 15% SDS-PAGE was collected from the subsequent purification on the same Phenyl-Sepharose 6 Fast Flow column (Figure 6). A single band at approximately 90
kDa was observed by 10% native-PAGE, and the hemolytic activity of this protein band suggested that it is an active tetrameric protein under physiological conditions (Figure 6). MALDI-TOF MS spectrum of peptide mapping via a trypsin digestion further confirmed the identity of G. hollisae TDH.
FIGURE 6. Purification and characterization of the G.h-rTDHWT protein from G.
hollisae. (A) The crude protein without tdh gene insertion in a pCR®2.1®-TOPO plasmid was obtained in the BL21(DE3)pLysS strain (lane 1). The crude protein containing the expressed G.h-rTDHWT in the BL21(DE3)pLysS strain was also included (lane 2). The homogenous protein with a molecular mass of ~22 kDa was obtained via two Phenyl Sepharose 6 Fast Flow chromatography runs (lane 3 and lane 4). (B) Native-PAGE of purified G.h-rTDHWT, with a molecular mass of ~90 kDa. (C) Hemolytic activity of G.h-rTDHWT.
25
4.3 Identification of G.h-rTDH
WTby MALDI-TOF-TOF MS
spectrometry
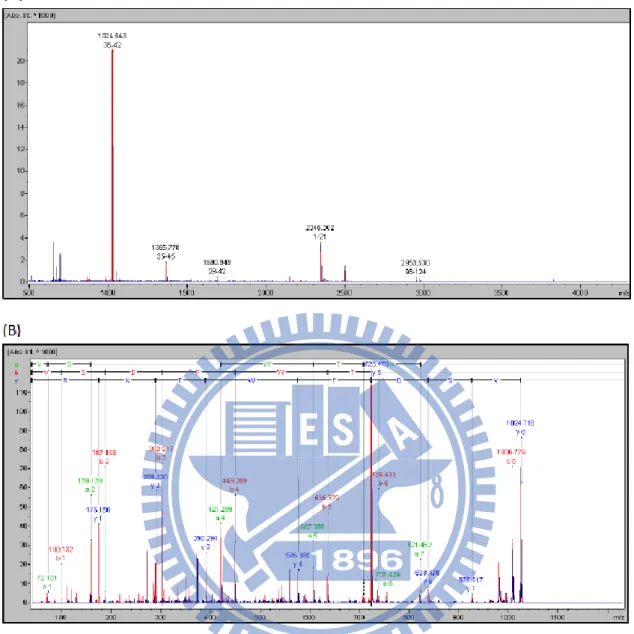
The purified protein was then subjected to a MALDI-TOF MS spectrometry for the internal amino acid determination to confirm the identity of purified protein. The SDS-PAGE band corresponding to G.h-rTDHWT was first applied to an in-gel trypsin digestion as described by Rosenfeld et al.35. The cutting sites of trypsin are lysine
(Lys, K) and arginine (Arg, R), respectively. After digestion, the resulting peptide mixtures were analyzed by MALDI-TOF MS. Among these peptide fragments shown in Figure 7, via a database search for the mass spectrum of the peptide and its fragment ions, the tandem mass spectrum of mono-charged precursor was observed at m/z 1024.543, 1365.788, 1690.949, 2346.302, 2953.530. Following the analysis of the highest signal by MS/MS, the sequence of this fragmental was determined to be
35
FIGURE 7. (A) MALDI-TOF-TOF MS spectrum and peptide mapping of G.h-rTDH protein. (B) Tandem mass spectrum of a signal charged tryptic peptide at m/z 1024.543, was deduced from the mass differences in the y-fragment ion series and partial b-ion and a-ion.
27
4.4 Expression, purification, and hemolytic activity determination
of mutated G.h-rTDH
Y53A/T59B/S63CIn order to understand the role of these distinct amino acid changes between the
tdh gene from the G. hollisae BCRC 15890 strain, and the historically published tdh
gene from NCBI data bank from G. hollisae 9041, seven G.h-rTDH mutants specific for different combinations of Tyr53, Thr59, or Ser 63 positions were constructed and purified. The procedures for expression, purification, identification, and hemolytic activity assays of these G.h-rTDHY53A/T59B/S63C mutants were identical with that of Gh-rTDHWT. These seven G.h-rTDHY53A/T59B/S63C mutants include G.h-rTDHY53H,
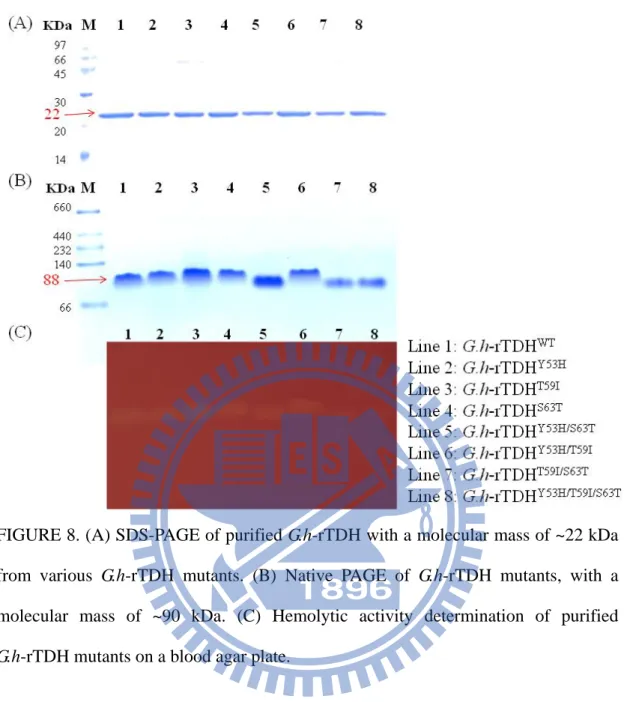
G.h-rTDHT59I, G.h-rTDHS63T, G.h-rTDHY53H/T59I, G.h-rTDHY53H/S63T, G.h-rTDH T59I/S63T, and G.h-rTDHY53H/T59I/S63T. As shown in the 15% SDS-PAGE data, all mutated
G.h-rTDHY53A/T59B/S63C exhibited a homology protein of approximately 22 KDa. The native-PAGE coupled with the blood agar plate assay further indicated that the corresponding protein bands with molecular mass of ~90KDa possess the hemolytic activity among these G.h-rTDHY53A/T59B/S63C mutants.
FIGURE 8. (A) SDS-PAGE of purified G.h-rTDH with a molecular mass of ~22 kDa from various G.h-rTDH mutants. (B) Native PAGE of G.h-rTDH mutants, with a molecular mass of ~90 kDa. (C) Hemolytic activity determination of purified
G.h-rTDH mutants on a blood agar plate.
4.5 The temperature-dependent hemolytic activity analysis and
thermostability studies of G.h-rTDH
WTand G.h-rTDH
Y53H/T59I/S63TTo investigate the thermostability and optimal temperature for hemolytic activity of purified G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T, a suspension of 5% human erythrocytes was incubated with G.h-rTDH protein for 1 h at different temperatures ranging from 4.0-100oC (Figure 9). Interestingly, the hemolytic activity of
29
additional heating over 85oC coupled with a rapid cooling treatment. Under incubation below 55 oC for 30 min, the G.h-rTDHWT protein still retained over 80% of its full activity (Figure 9 (A)). In comparison, the G.h-rTDHY53H/T59I/S63T lost the entire hemolytic activity via a heating above 60oC, and no recovering activity was observed after a rapid cooling treatment (Figure 9 (B)).
FIGURE 9. Thermostability assay of G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T mutants. The relative hemolytic activities were measured at 37oC under various temperature treatments. According to the results, G.h-rTDHWT has the Arrhenius effect, whereas the G.h-rTDHY53H/T59I/S63T did not. Data are presented as the means for triplicate experiments. Error bars represent the standard deviations (SD)
4.6 Comparison of hemolytic activity for G.h-rTDH
Y53A/T59B/S63CIn order to investigate the biophysical characterization of various combination mutants on Tyr53, Thr59, or Ser63 positions of G.h-rTDH, the hemolytic activity of various mutants on human erythrocytes were further studied. In Figure 10, the hemolytic activities of various G.h-rTDHY53A/T59B/S63C mutants were examined on a 96 well plate. Decreased hemolytic activities were displayed from G.h-rTDHY53H/T59I,
G.h-rTDHT59I/S63T, and G.h-rTDHY53H/T59I/S63T mutants. Moreover, the
G.h-rTDHY53H/S63T mutant showed more dominant hemolytic activity than that observed for G.h-rTDHWT. The hemolysis ability of G.h-rTDHS63T and G.h-rTDHT59I mutants were very similar, and also more dominant than that observed for
G.h-rTDHWT. The concentration of various G.h-rTDHY53A/T59B/S63C higher 25 g/mL caused 100% hemolysis, while lower than 0.39 g/mL, their hemolytic activity was below the detectable level and had no effect on human erythrocytes.
31
4.7 Comparison of the hemolytic activity of G.h-rTDH
Y53A/T59B/S63Cmutants at 37
oC, 70
oC, 90
oC
As illustrated above, the G.h-rTDHWT lost its hemolytic activity under 60-80 oC but recovered its function via continuing to heat at 90oC coupled with a rapid cooling. This paradoxical phenomenon is referred to as “Arrhenius effect”. G.h-rTDHWT was first treated with different temperature heating conditions at 37oC, 70oC, or 90oC, coupled by a rapid cooling treatment, respectively. The protein structure or its biophysical characteristics were changed via these three various pretreatments with different significance. The protein structure was changed from a native form into a fiber form, then transformed to the unfold state, and recovered it to its native form via a induced cooling treatment. This conformational change may affect the biophysical properties. Some mutants, G.h-rTDHY53H, G.h-rTDHT59I, G.h-rTDHS63T,
G.h-rTDHY53H/S63T, also exhibited this unusual phenomena, but the G.h-rTDHY53H/T59I, and G.h-rTDHT59I/S63T, as well as G.h-rTDHY53H/T59I/S63T lost this Arrhenius effect, even with the concentration elevated to 10 g/mL (Figure 11).
FIGURE 11. The hemolytic activity of various G.h-rTDHY53A/T59B/S63C mutants after different temperature pre-treatments, coupled with a rapid cooling treatment. Data are presented as the means for triplicate experiments. Error bars represent standard deviations (SD).
4.8 Analysis of G.h-rTDH
Y53A/T59B/S63Cthermostability by difference
scanning calorimetry
DSC measurements involve the heating of a sealed sample of protein solution at a constant rate of temperature increase. As long as no other process occurs in the solution that releases or takes up heat, the temperature of the solution will rise monotonically with the instrumental heating (electrical power input). The DSC profile provides much valuable information, including the temperature at which the maximum excess heat capacity occurs, called “Tmax”; the area under the curve,
obtained by integration using the software supplied with the instrument, gives “the enthalpy of denaturation”; the width of the peak at half of the maximum excess heat
33
co-operativity of the protein structure36. The calorimetric scan of TDHWT andother TDH mutationsin 10 mM Kpi (pH 7.0) were characterized by a single peak. The transition temperature of those proteins from low to high were 51.8oC (G.h-rTDHT59I), 52oC (G.h-rTDHY53H), 55.3oC (G.h-rTDHS63T), 56.3oC (G.h-rTDHY53H/S63T), 56.6oC (G.h-rTDHWT), 57.1oC (G.h-rTDHT59I/S63T), 58oC (G.h-rTDHY53H/T59I), 58.4oC (G.h-rTDHY53H/T59I/S63T), respectively. Those proteins without Arrhenius effect including G.h-rTDHY53H/T59I/S63T, G.h-rTDHY53H/T59I and G.h-rTDHT59I/S63T have the top three Tm values, and obtained a wider FWHM, indicating that during the unfolding
process by increasing temperature the more entropy is needed to disrupt the intramolecular interaction with those proteins, and more stable and compacted structures than others were conserved in these mutants. The G.h-rTDHT59I and
G.h-rTDHY53H have the lowest Tm values, indicating that these proteins were more
FIGURE 12. The DSC result. Corrected DSC thermograms of G.h-rTDHY53A/T59B/S63C in 10 mM Kpi buffer at pH 7.0.
4.9 Analyze
the
secondary
structure
change
of
various
G.h-rTDH
Y53A/T59B/S63Cmutants by circular dichroism spectroscopy
The change of secondary/tertiary structure caused by the thermal denaturation of
G.h-rTDH in Kpi buffer (pH 7.0) was examined by circular dichroism (CD) spectra.
At temperatures below 50°C, all proteins exist in a native state characterized by a -rich secondary structure with a pronounced minimum at 218 nm (Figure 13 (E)). Between 50-60oC, proteins underwent an intermediate state as shown in (Figure 14 (E)). Interestingly, the melting temperature-curve (the Tm value was determined by
previous DSC instrument) of CD spectra of G.h-rTDHY53H/T59I/S63T was compared to that of G.h-rTDHWT (Figure 14 (A)). However, G.h-rTDHY53H/T59I/S63T protein does not transit from a -rich structure into a -rich structure until 75oC, whereas G.h-rTDHWT
35
G.h-rTDHY53H/T59I/S63T has a higher activation energy than that of G.h-rTDHWT in their phase transition processes. The increase of temperature from 60-95oC, the -helix-rich structure content of G.h-rTDHY53H/T59I/S63T was still retained, whereas that of G.h-rTDHWT was decreased vividly. Interestingly, all G.h-rTDH proteins were not completely denatured even at 95oC. In 10 mM Kpi buffer (pH 7.0), the temperature-induced conformational change of G.h-rTDHY53H/T59I/S63T occurs in a two-state manner, while that of G.h-rTDHWT is in a three-state manner. For other
G.h-rTDHY53A/T59B/S63C mutants, the similar spectrum changes, as that of
G.h-rTDHY53H/T59I/S63T, (in either the far-UV CD spectra or DSC data) were observed in G.h-rTDHT59I/S63T and G.h-rTDHY53H/T59I mutants, from which both of them lost its Arrhenius effect after continuing heating, followed by a rapid cooling treatment (Figure 13 (B)). However, all proteins that have an Arrhenius effect, except for
G.h-rTDHY53H G.h-rTDHT59I and G.h-rTDHS63T mutants, must undergo a transition state prior to the formation of an unfolded state after a thermal denaturation treatment. In the far-UV CD spectra, the complete collapse of its secondary structure was characterized in the G.h-rTDHY53H G.h-rTDHT59I and G.h-rTDHS63T mutants at temperatures above 56oC and 60oC, respectively (Figure 13 (C)). The
G.h-rTDHY53H/S63T CD spectrum is observed to occur between G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T (Figure 13 (D)).
FIGURE 13. (A)The CD spectrum of G.h-rTDHWT and G.h-rTDHY53H/T59I/S63T and the
G.h-rTDHWT show three-state and G.h-rTDHY53H/T59I/S63T show two-state manner when heat the temperature.
37
FIGURE 13. (B) The CD spectrum of G.h-rTDHY53H/T59I/S63T, G.h-rTDHY53H/T59I and
FIGURE 13. (C) The CD spectrum of G.h-rTDHY53H, G.h-rTDHT59I and G.h-rTDHS63T which are process the Arrhenius effect.
39
FIGURE 13. (D) The CD spectrum of G.h-rTDHWT, G.h-rTDHY53H/T59I/S63T, and
G.h-rTDHY53H/S63T. The G.h-rTDHY53H/S63T pattern is between G.h-rTDHWT and
FIGURE 13. (E) The CD spectrum of all mutated G.h-rTDHY53A/T59B/S63C proteins in 10mM Kpi buffer, pH 7.0. Red box showed the no Arrhenius effect proteins.
41
4.10
Morphology
examination
and
MTT
assay
for
G.h-rTDH
Y53A/T59B/S63Cof the cytotoxicity and cytoviability effect on
AGS cells
To investigate the cytotoxicity of purified G.h-rTDHY53A/T59B/S63C proteins on mammalian cells, the human stomach epithelial cell line, AGS, served as an in vitro model. The morphological change of AGS cells could be visually assessed in the absence or presence of G.h-rTDHY53A/T59B/S63C exposure for 30 min with 10 μg/mL
G.h-rTDHY53A/T59B/S63C at 26oC. Among the result of these mutants, the morphology of AGS cells were changed, including the membrane blebbing, the cell detachment, loss of cell cytoplasm with cell shrinkage, and the reduction of nuclei size.
G.h-rTDHY53A/T59B/S63C mutants have different levels of cytotoxicity on the AGS cell line and the cell morphology has different degrees of damage during the treated duration, as shown in Figure 14. When AGS cells were exposed to the
G.h-rTDHY53H/S63T mutant, cell morphology has a dramatic change. When AGS was exposed to other mutants like G.h-rTDHY53A/T59B/S63C except for G.h-rTDHY53H/T59I/S63T and G.h-rTDHT59I/S63T, the cell morphology had obvious changes within the initial 5 min, and the complete dramatic change in the morphology was observed in 30 min. In addition, we used the MTT assay (Figure 16) to analyze the cytoviability when cell was treated with different concentrations of G.h-rTDHY53A/T59B/S63C (100 μg/mL and different serial dilutions in ten-fold increments) in 1 h at 37oC. Accordingly, the inhibitory concentration (IC50) value of G.h-rTDHY53H/T59I (defined as the drug
concentration at which cell growth was inhibited by 50%) at 10 μg/mL was the lowest observed, and the IC50 value of G.h-TDHY53H/S63T inhibitory concentration (IC50) at 2
43
FIGURE 14. Cell morphology. AGS cell line was exposed G.h-rTDHY53A/T59B/S63C 10ug/mL in 30min at 26oC. (I) G.h-rTDHT59I/S63T (H) G.h-rTDHY53H/S63I (A) G.h-rTDHY53H,T59I,S63T S63T (G) G.h-rTDHY53H/T59I
45
FIGURE 15. MTT assay results of various G.h-rTDHY53A/T59B/S63C mutants. The cytoviability decreased in proportion to different concentrations of G.h-rTDH (100 μg/mL and ten-folded serial dilutions) in 1 h at 37o
C. Data are presented as the means for triplicate experiments. Error bars represent the standard deviations (SD)
4.11 Thioflavin T florescence assay
Thioflavin T fluorescence assay was used to determine the fibrils
contents of various TDH mutants, which are generated from a
thermal-pretreatment of TDH mutants at either 37
oC, 70
oC, or 90
oC
coupled with a rapid cooling, respectively. According to the result of ThT
assay,
the
filber
contents
of
G.h-rTDH
T59I,
G.h-rTDH
S63T,
G.h-rTDH
Y53H/T59I,
G.h-rTDH
Y53H/S63T,
G.h-rTDH
T59I/S63T,
and
G.h-rTDH
Y53H/T59I/S63Tgenerated from a 90
oC thermal pre-treatment
coupled with rapid cooling are higher than that of TDH mutations
generated from a 70
oC thermal pre-treatment coupled with rapid cooling.
Comparably, the fibrils contents of G.h-rTDH
WTand G.h-rTDH
Y53Hare
similar at either fiber generation from a 90
oC thermal pre-treatment or a
70
oC thermal pre-treatment, coupled with a rapid cooling, respectively.
FIGURE 16. Thioflavin T florescence assay result. Those curves are presented as the means for triplicate experiments.
G.h-rTDHY53H G.h-rTDHWT G.h-rTDHWT G.h-rTDHY53H G.h-rTDHT59I G.h-rTDHS63T G.h-rTDHY53H/T59I G.h-rTDHY53H/S63T G.h-rTDHT59I/S63T G.h-rTDHY53H/T59I/S63T 37oC 70oC 90oC