國立臺灣大學生命科學學院植物科學研究所 碩士論文
Institute of Plant Biology College of Life Science National Taiwan University
Master Thesis
阿拉伯芥中 LecRK-V.2 和 LecRK-VII.1 蛋白質在處理 細菌鞭毛蛋白後與辨識鞭毛蛋白受體 FLS2 和其共同
受體 BAK1 的蛋白質複合體形成之分析 Analysis of LecRK-V.2 and LecRK-VII.1 complex formation with FLS2 and BAK1 upon flagellin perception
in Arabidopsis 麥賢偉 Hsien Wei Mai
指導教授:金洛仁 博士 Advisor: Laurent Zimmerli, Ph.D.
中華民國 105 年 7 月
July, 2016
摘要
在阿拉伯芥植物中有許多 L 型態凝集素受體激酶(LecRKs)參與病原 菌辨識機制中引起細胞內對抗生物性跟非生物性的逆境防禦機制。此 種受體激酶有細胞外的豆科植物凝集素區域,穿膜區域和細胞內的絲 氨酸/蘇氨酸激酶區域。在我們實驗室先前研究發現 LecRK-V.2 和 LecRK-VII.1 都參與在病原菌防禦機制當中。此兩基因突變株比起野生
型 Col-0 阿拉伯芥對抗 Pseudomonas syringae pv. tomato DC3000 (Pst
DC3000)此病的能力來得弱;然而,此兩基因過量表現轉殖株則相較
野生型對抗 Pst DC3000 的抗性更強,並且我們發現此兩基因突變株在
氣孔的免疫機制上有明顯的缺陷。在此,我們利用阿拉伯芥原生質體
過量表現此兩基因蛋白質再利用共免疫沉澱法去偵測兩蛋白質對於鞭
毛蛋白辨識受體 FLS2 和其複合體蛋白質 BAK1 是否有與此免疫複合
體產生結合的情形。我們也在酵母菌雙雜合系統實驗發現 LecRK-V.2
和 LecRK-VII.1 蛋白質可以透過內部激酶區域與 FLS2 及 BAK1 蛋白
質內部激酶區域產生結合的現象。為了更加瞭解此兩基因參與的植物
抗病菌的防禦機制調控,在此研究,我們發現此兩基因參與在細菌鞭
毛蛋白引起植物體免疫防禦機制的 FLS2-BAK1 複合體當中。
關鍵字:
阿拉伯芥、LecRK-V.2、LecRK-VII.1 、 病原菌辨識受體、防禦
機制、FLS2、BAK1
。Abstract
In Arabidopsis, several L-type lectin receptor kinases (LecRKs) were found to
function as or together with pattern-recognition receptors (PRRs) in recognition of
stress signals and initiation of plant defense responses towards abiotic and biotic
stresses. LecRKs possess an extracellular legume-lectin domain, a transmembrane
domain and an intracellular serine/threonine kinase domain. Our previous analyses
show that LecRK-V.2 and LecRK-VII.1 are involved in the pattern-triggered immunity
(PTI) defense response. Both lecrk-V.2 and lecrk-VII.1 mutant plants are indeed more
susceptible to bacteria Pseudomonas syringae pv. tomato DC3000 than Col-0
wild-type plants. LecRK-V.2 and LecRK-VII.1 overexpression lines demonstrate
increased resistance to Pst DC3000. In addition, both lecrk-V.2 and lecrk-VII.1 mutants
are defective in stomatal immunity. Here, we show by co-immunoprecipitation in
Arabidopsis protoplast that LecRK-V.2 and LecRK-VII.1 associated with the PRR
FLS2 and the co-receptor BAK1 before and after perception of the bacterial
pathogen-associated molecular pattern (PAMP) flagellin. Moreover, LecRK-V.2 and
LecRK-VII.1 could interact with FLS2 and BAK1 through their kinase domains when
analyzed by the in vitro yeast-two-hybrid assay. In this study, I clarified the role of
LecRK-V.2 and LecRK-VII.1 in the FLS2-BAK1 complex upon flagellin perception.
Keywords:FLS2、LecRK-V.2、LecRK-VII.1、BAK1、stomatal closure、Pst DC3000、
PTI.
Contents
摘要 ... II
Abstract ... IV
Contents ... VI
Introduction ... 1
Material and Methods ... 9
Biological Material and Plant Growth Conditions ... 9
Co-immunoprecipitation Assay in Arabidopsis Protoplast. ... 9
Yeast Two-Hybrid Assays ... 11
Results ... 14
LecRK-V.2 and LecRK-VII.1 associate with FLS2 in vivo ... 14
LecRK-VII.1 could directly interact with FLS2 through their kinase domain, but LecRK-V.2 could not. ... 15
LecRK-V.2 and LecRK-VII.1 associate with BAK1 in vivo ... 16
In vitro, BAK1 can directly interact with LecRK-V.2 and LecRK-VII.1 through their kinase domain ... 17
In vitro and in vivo association analyses of LecRK-V.2 and LecRK-VII.1 ... 18 Each LecRKs may not affect the association of FLS2 with the other LecRK20
Discussion ... 22
Conclusions and Future Perspectives ... 27
Figures ... 30
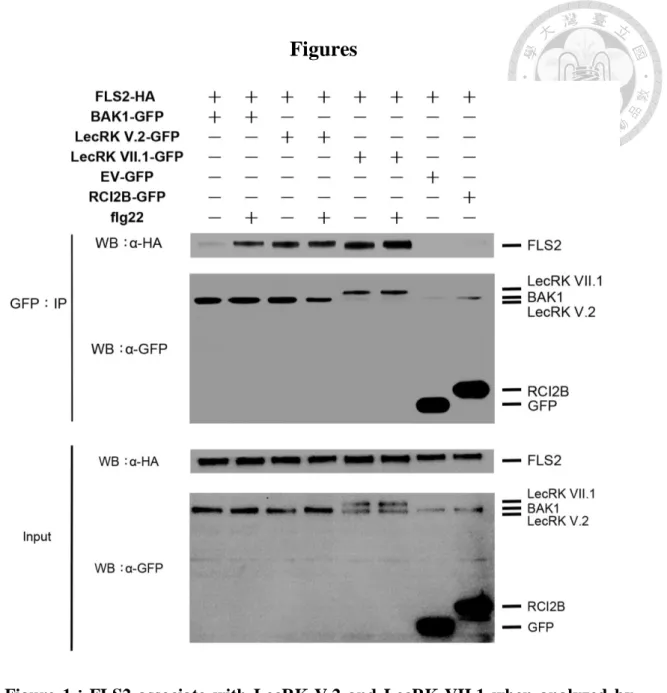
Figure 1:FLS2 associate with LecRK-V.2 and LecRK-VII.1 when analyzed
by co-immunoprecipitation in Arabidopsis protoplasts. ... 30
Figure 2:In vitro FLS2 direct interaction with LecRK-VII.1 kinase domains
but not LecRK-V.2. ... 32
Figure 3:BAK1 associates with LecRK-V.2 and LecRK-VII.1 when analyzed
by co-immunoprecipitation in Arabidopsis protoplasts. ... 33
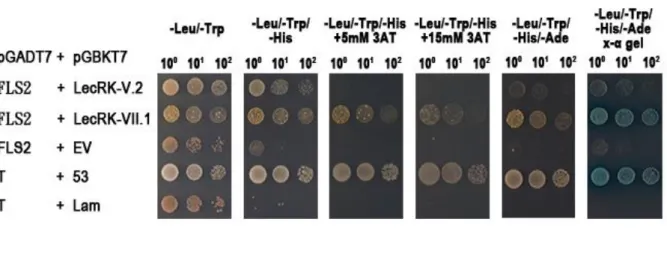
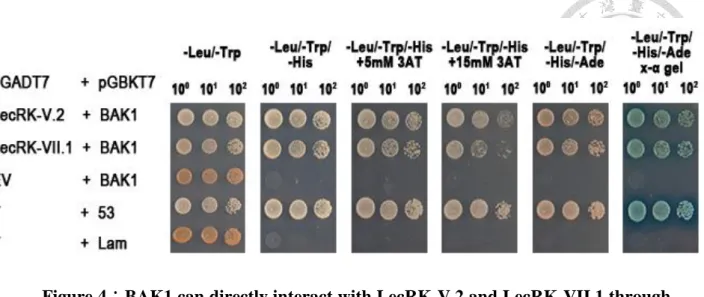
Figure 4:BAK1 can directly interact with LecRK-V.2 and LecRK-VII.1
through its kinase domain. ... 35
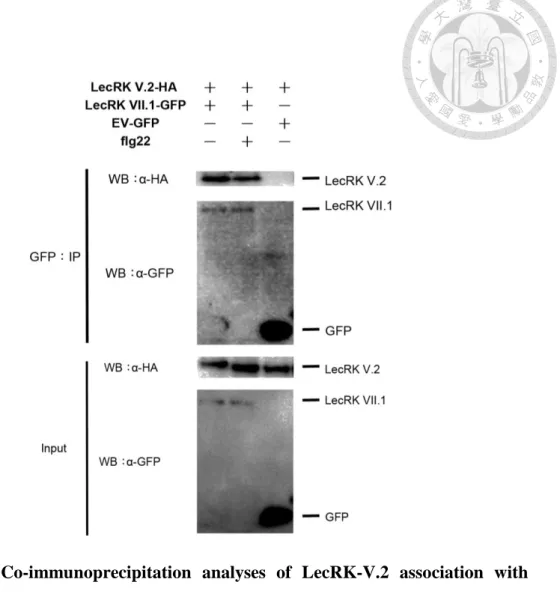
Figure 5: Co-immunoprecipitation analyses of LecRK-V.2 association with
LecRK-VII.1 in Arabidopsis protoplasts. ... 36
Figure 6: LecRK-V.2 cannot directly interact with LecRK-VII.1 through
their kinase domain when analyzed with the yeast two hybrid assay. ... 38
Figure 7: FLS2 associates with BAK1 and LecRK-VII.1 in the lecrk-V.2
mutant background. ... 39
Figure 8: FLS2 associates with BAK1 and LecRK-V.2 in the lecrk-VII.1
mutant background. ... 40
Figure 9:Proposed model ... 41
Tables ... 43
Table 1 :Primers for kinase domain constructs ... 43
Table 2 :List of abbreviations ... 44
Supplementary Figures ... 45
Supplemental Figure 1: LecRK-V.2 and LecRK-VII.1 are necessary for the resistance against to Pst.DC3000. ... 45
Supplemental Figure 2: LecRK-V.2 and LecRK-VII.1 are necessary for stomatal immunity. ... 46
Supplemental Figure 3:The PTI apoplastic responses and regulations in lecrk-V.2 and lecrk-VII.1 mutant lines. ... 47
Supplemental Figure 4:LecRK-V.2-GFP and LecRK-VII.1 –GFP fusion proteins localized on the membrane in Arabidopsis mesophyll protoplast. 49 References... 52
Introduction
Pathogen-triggered plant defense immunity
Plant growth and reproduction are affected by many pathogens known as
plant-associated microbes. Mainly, two pathways are typical of the innate immune
responses that plants activate upon infection with plant-associated microbes. The first
branch is triggered by the recognition of highly conserved pathogen /microbe-associated
molecular patterns (PAMPs/MAMPs) by host cell transmembrane proteins that function
as pattern recognition receptors (PRRs), which in turn, activate
PAMP/MAMP-triggered immunity (PTI/MTI). The second branch recognizes type III
effectors inside the plant cell via nucleotide-binding site-leucine-rich repeat (NB-LRR)
resistance (R) proteins. This leads to the activation of the effector-triggered immunity
(ETI) (Jones and Dangl 2006).
Membrane receptors recognize microbe-associated molecular patterns in plants
Membrane-localized receptors which are is also called PRRs can recognize the
conserved molecules characteristic of an entire group of microbes known as MAMPs.
MAMPs include bacterial lipopolysaccharides (LPS), peptidoglycans (PGN), fungal
chitin, quorum sensing factors, flagellin, and even danger signals (damage-associated
molecular patterns; DAMPs) which are released by plants during microbial infection
(Monaghan and Zipfel 2012). Arabidopsis flagellin-sensing 2 (FLS2) is one of the
best-characterized PRR in plants. It encodes a leucine-rich repeat-receptor-like kinases
(LRR-RLK) that recognizes bacterial flagellin or its active peptide derivative flg22
(Gomez-Gomez and Boller 2000). Upon flg22 perception, FLS2 instantaneously
interacts with another LRR-RLK, the brassinosteroid (BR) insensitive 1-associated
kinase 1 (BAK1) (Chinchilla et al. 2007, Heese et al. 2007). Botrytis-induced kinase 1
(BIK1), a plasma membrane-localized receptor-like cytoplasmic kinase (RLCK), is
rapidly phosphorylated upon flg22 perception in an FLS2- and BAK1-dependent
manner (Lu et al. 2010, Zhang et al. 2010). BIK1 functions as a kinase substrate of
BAK1 and forms a complex with FLS2 and BAK1 in transducing flagellin signaling
(Lu, Wu, Gao, Zhang, Shan and He 2010).
Flagellin triggers plant defense responses
After exposure to flg22 or other MAMPs, the plant cell undergoes a rapid activation
of the MAP kinase (MPK) cascade (Nuhse et al. 2000), a burst of reactive oxygen
species (ROS) controlled by the NADPH oxidase RbohD (Blume et al. 2000, Nuhse et
al. 2007), and a transient influx of calcium ion from the apoplast (Blume, Nurnberger,
Nass and Scheel 2000, Lecourieux et al. 2002), with the latter being essential for the
ROS production. The plasma-membrane-associated kinase BIK1, which is a direct
substrate of the PRR complex, directly interacts with and phosphorylates RBOHD upon
PAMP perception (Kadota et al. 2014). BIK1 phosphorylates in different residues of
calcium-dependent protein kinases. Both MAMP-induced BIK1 activation and
BIK1-mediated phosphorylation of RBOHD are calcium independent. Importantly,
phosphorylation of these residues is critical for the MAMP-induced ROS burst and
antibacterial immunity (Kadota, Sklenar, Derbyshire, Stransfeld, Asai, Ntoukakis,
Jones, Shirasu, Menke, Jones and Zipfel 2014, Li et al. 2014).
Lectin receptor kinases are involved in plant defense innate responses
Arabidopsis possesses many receptor-like kinases (RLKs), and one class of RLKs is
the lectin receptor kinase which contain an extracellular lectin domain that are
predicted to bind to carbohydrates, a transmembrane domain, and an intracellular
kinase domain. Lectin receptor kinases are classified by their extracellular lectin
motifs. The first group is called G-type lectins because it contains a GNA-related
motif. Beside lectin GNA-related motif, the extracellular domains of this group also
contain cysteine-rich EGF-like (epidermal growth factor) and PAN
(plasminogen-apple-nematode) motifs that both function in protein homodimerization
(Naithani et al. 2007, Van Damme et al. 2007). G-type lectins group contain around 40
RLK genes in Arabidopsis. But the function of G-type lectins is unknown. The second
group includes calcium-dependent lectins which are also abbreviated as C-type lectins,
and this group is mostly found in mammalian cells but only one gene which encoding a
C-type lectins protein in Arabidopsis has been found until now (Bouwmeester and
Govers 2009, Cambi et al. 2005). The third group regroups lectin proteins with a
legume-like or L-type lectin domain in their extracellular domains. Proteins in this
group are also called LecRKs. There are 45 genes encoding L-type lectin receptor
kinases in plants. LecRKs may be involved in the binding of potential
ligands-oligosaccharide. LecRKs may also recognize small hydrophobic ligands, such
as PAMPs and plant hormones (Andre et al. 2005, Bouwmeester and Govers 2009,
Van Damme, Nakamura-Tsuruta, Smith, Ongenaert, Winter, Rouge, Goldstein, Mo,
Kominami, Culerrier, Barre, Hirabayashi and Peumans 2007).
LecRKs are involved in many biotic and abiotic stress
LecRKs are involved in both biotic and abiotic stress (Vaid et al. 2013). In abiotic
stress, LecRK-I.3 gene is differentially up-regulated during salt stress (He et al. 2004).
The LecRK-V.1 functions as a positive regulator in responsive to abscisic acid (ABA)
and is also involved in osmotic and salt stress (Deng et al. 2009). In biotic stress,
LecRK-VI.2 provides resistance against the hemibiotrophic bacteria Pseudomonas
syringae and the necrotrophic bacteria Pectobacterium carotovorum (Singh et al.
2012). The Arabidopsis LecRK-V.5 gene was found to reverse the stomatal closure
induced by bacterial infection and relieve the plant from CO2 deficiency (Arnaud et al.
2012, Desclos-Theveniau et al. 2012). LecRK-IX.1 and LecRK-IX.2 are functional
analogs in the resistance regulation of Phytophthora infection and are involved in the
plant cell death in Arabidopsis (Wang et al. 2015). LecRK-I.9 is involved in the
binding of extracellular ATP and signaling of ATP-induced calcium response,
mitogen-activated protein kinase activation, and gene expression (Cao et al. 2014,
Choi et al. 2014). Hence, LecRKs are known to be involved in the plant defense
response, signaling, and even in extracellular ligand binding.
LecRK-V.2 and LecRK-VII.1 are related to flg22-triggered defense responses
T-DNA insertion mutant lines of LecRKs with high expression levels under biotic
stress from previous microarray studies (Bouwmeester and Govers 2009) were ordered
from the Arabidopsis Biological Resource Center (ABRC). Afterward, T-DNA
insertion mutant plants were dipped in a solution of hemibiotrophic Pseudomonas
syringae pv. tomato DC3000 (Pst DC3000) bacteria and symptoms were evaluated 3
days later. From this screening, two lecrk mutants, namely leck-V.2 and lecrk-VII.1
demonstrated increased susceptibility to Pst DC3000 bacteria (Supplemental Figure 1).
Lecrk-V.2 and lecrk-VII.1 showed defective stomatal closure compare to wild-type
Col-0 after infection with Pst DC3000 (Supplemental Figure 2). But lecrk-V.2 and
lecrk-VII.1 T-DNA insertion mutant line showed no difference compare with wild-type
Col-0 in the other PTI pathway, such as the production of extracellular ROS, the
deposition of callose, the activation of MAP kinase (MPK) cascades, and expression of
defense-related genes (Supplemental Figure 3). The transient expression in
Arabidopsis protoplast to clarify the localization of LecRK-V.2 and LecRK-VII.1
proteins. LecRK-V.2 and LecRK-VII.1 proteins were indeed localized on the plasma
membrane of mesophyll Arabidopsis protoplast (Supplemental Figure 4). Previous
experiments showed that LecRK-V.2 and LecRK-VII.1 only interact with FLS2 in the
presence of flg22 by performing bimolecular fluorescence complementation (BiFC)
assay (Supplemental Figure 5).
LecRK-V.2 and LecRK-VII.1 are involved in FLS2-BAK1 complex
In order to clarify the roles of LecRK-V.2 and LecRK-VII.1 in the flagellin-triggered
immunity response, my works analyzed the relationship among LecRK-V.2,
LecRK-VII.1, FLS2, and BAK1. Towards this goal, the relationships of association
among these proteins were performed by the co-immunoprecipitation (Co-IP) of
proteins in Arabidopsis protoplasts. Both LecRKs associated with the FLS2-BAK1
complex with and without the perception of flagellin. LecRK-V.2 cannot interact with
FLS2, but LecRK-VII.1 can interact with FLS2 directly through their kinase domains
by performing the yeast two-hybrid. Both LecRKs can interacted with the BAK, but
LecRK-V.2 cannot interact with LecRK-VII.1 through their kinase domains. It is
wondered that if both LecRKs will affect each other. Hence, I tested the association
between FLS2, BAK1, and LecRK-VII.1 in lecrk-V.2 mutant protoplasts by
performing the Co-IP assay. It was found that LecRK-V.2 didn’t affect the association
between FLS2, BAK1, and LecRK-VII.1, respectively. Further, I tested the association
between FLS2, BAK1, and LecRK-V.2 in lecrk-VII.1 mutant protoplasts by
performing the Co-IP assay. The results indicated that the LecRK-VII.1 didn’t affect
the association between FLS2, BAK1, and LecRK-V.2, respectively either. So we
proposed that LecRK may affects the interaction between FLS2 and the other LecRK,
but they are still in the FLS2-BAK1 complex before and after the treatment with flg22.
In this study, we found that LecRK-V.2 and LecRK-VII.1 associate with FLS2 and
BAK1 before and after the treatment of flg22. FLS2 can interact with LecRK-VII.1
through their intracellular kinase domain but FLS2 cannot interact with LecRK-V.2.
BAK1 can interact with LecRK-V.2 and LecRK-VII.1 through their intracellular
kinase domain. LecRK-V.2 cannot interact with LecRK-VII.1 through their
intracellular kinase domain.
Material and Methods
Biological Material and Plant Growth Conditions
Arabidopsis thaliana ecotype Col-0 plants were grown in commercial potting soil
and perlite (ratio-3:2) at 22–24°C day and 17–19°C night temperature under short day
(9 hours under the light / 15 hours under the dark) condition of light illumination
period. The lighting was supplied at an intensity of ∼100μE m−2s−1 by fluorescence
white light tubes. T-DNA insertion mutant lecrk-V.2 and lecrk-VII.1 lines were
obtained from the Arabidopsis Biological Resource Center (ABRC) (the T-DNA
insertion site).
Co-immunoprecipitation Assay in Arabidopsis Protoplast.
For Co-IP assay, we constructed the plasmid of FLS2-HA, BAK1-HA,
LecRK-V.2-HA, BAK1-GFP, LecRK-V.2-GFP, LecRK-VII.1-GFP, GFP-empty vector,
RCI2B-GFP. FLS2, BAK1, and LecRK-V.2 PCR amplified products were cloned into
pCR8 TOPO TA cloning vector (Invitrogen) and then cloned into pEG100 vector by
the ph.D student, Shweta Bagul, and the postdoctoral researcher, Mathieu Gauthier.
BAK1-GFP, LecRK-V.2-GFP, LecRK-VII.1-GFP, and RCI2B-GFP constructs were
PCR amplified from wide type Col-0 cDNAs, and PCR products were then cloned into
pCR8 TOPO. Then, these plasmids were finally recombined into the
Gateway-compatible expression vectors pEarlyGate103 (ordered from ABRC).
GFP-empty vector construct was modified from pEarlyGate103 with removing
gateway cassette by using XhoI restriction enzyme. Plasmids of FLS2-HA and
BAK1-GFP or FLS2-HA and LecRK-V.2-GFP, or FLS2-HA and LecRK-VII.1-GFP or
FLS2-HA and GFP-empty vector, or FLS2-HA and RCI2B-GFP were co-transformed
into Arabidopsis protoplasts by a polyethylene glycol method as described previously
(Yoo et al. 2007).
We separated 150 μL protoplasts from a total volume of 500 μL of total
protoplasts for running of input proteins. The rest of total proteins were extracted in an
extraction buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl,10 mM DTT, 10 mM EDTA
pH8.0, 1 mM Na3VO4, 10% glycerol, and 1% IGEPAL CA-630 [Sigma-Aldrich]) with
protease inhibitor cocktail (Roche). The solution was incubated with gentle shaking at
4 °C for at least 30 minutes and then centrifuged in the speed of 14,000 g for 15 min at
4°C. The supernatant was incubated with 15 μL GFP-Trap○R _MA (ChromoTek,
GmbH) for 4 hours to 16 hours at 4°C, and GFP-Trap was diluted with dilution buffer
(10 mM Tris-HCl, pH 7.5, 100 mM NaCl, 0.5% EDTA,pH8.0) before the incubation
with the supernatant. After the incubation at 4°C, the beads were centrifuged and
washed four times with the washing buffer (10 mM Tris-HCl, pH 7.5, 100 mM NaCl,
0.5% IGEPAL CA-630 [Sigma-Aldrich]). Proteins were separated by 10% SDS–PAGE
gel and then transferred to a polyvinylidine fluoride membrane (ImmobilonP;
Millipore). GFP and HA fusion proteins were detected by immunoblotting with
anti-GFP and anti-HA primary antibodies, respectively.
Yeast Two-Hybrid Assays
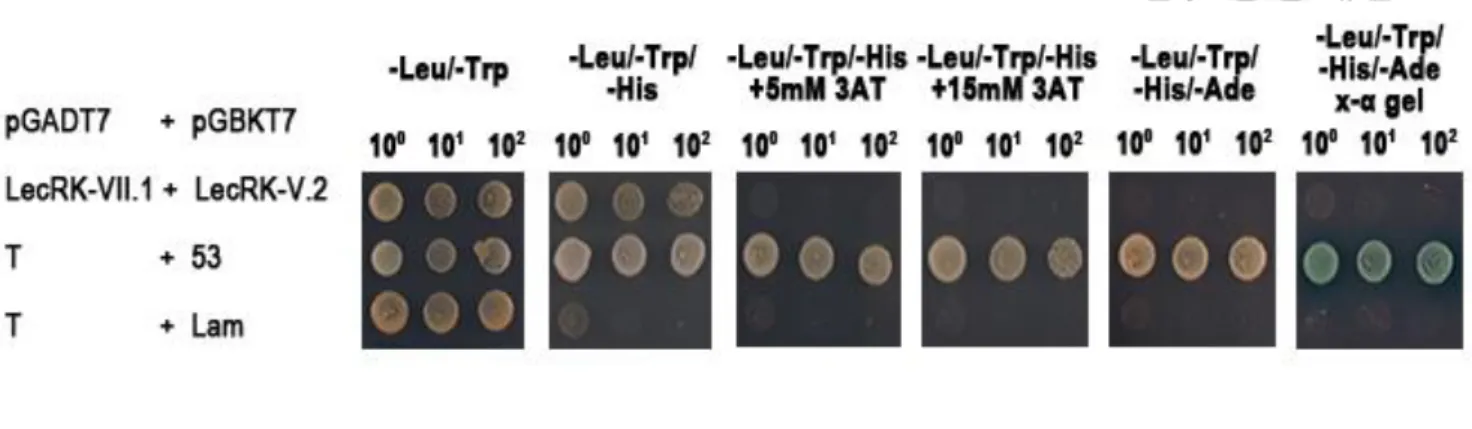
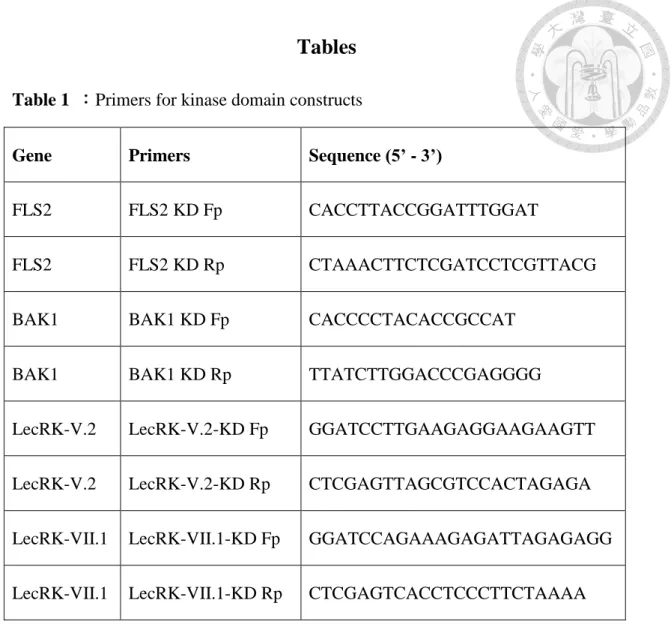
Yeast two-hybrid assays were performed as described in the manual of Matchmaker
Gold Yeast Two-Hybrid Systems (Clontech) and the user manual of Yeast Protocol
Handbook (Clontech). Briefly, we amplified the kinase domains of FLS2, BAK1,
LecRK-V.2, and LecRK-VII.1 fragments from pCR8-FLS2, pCR8-BAK1,
pCR8-LecRK-V.2, and pCR8-LecRK-VII.1 by using the kinase domain forward and
reverse primers (primers are listed in S1 Table). The PCR amplified products were
subcloned into pGBKT7 and pGADT7 vectors to construct different bait and prey
constructs (primers are listed in S1 Table). Then, different pairs of bait and prey
constructs were co-transformed into yeast AH109 strain by lithium acetate (LiAc)
method. From Clontech Yeast Protocol Handbook, they used and suggested users to
adopt the LiAc method (Ito et al. 1983), which then modified into much more simple
and more reproducible protocols (Gietz et al. 1992, Hill et al. 1991, Schiestl and Gietz
1989). In this LiAc transformation method, yeast competent cells were cultured in
small amounts form the colonies of streaked YPD plates in fresh and incubated for
12-16 hours with shaking (180–200 rpm) in 28°C. The next day, we transferred enough
of the overnight culture into the fresh medium and incubated at 28°C for 3–5 hours
with shaking (180–200 rpm) to produce an OD600 concentration of yeast cultures of 0.4
– 1.0. The yeast cultures were then centrifuged at 1000~1500 g for 5 minutes in 4°C
and washed with sterilized water for 2 times and suspended in the LiAc solution. The
transformed DNA were prepared by mixture of 200 ng plasmid DNA per each
reaction, and the mixture of DNA and yeast was mixed with Polyethylene glycol
(PEG) and the appropriate amount of LiAc and then incubated at 28°C for half an hour.
After the incubation, the heat shock method, which allows the DNA to enter the cells,
was used for 15 minutes. The yeast cells were then centrifuged at 500 g for 5 minutes
and then resuspended into the LiAc solution. The yeast cells were then plated on the
synthetic dropout (SD) medium, or DDO medium (minimal media double dropouts,
SD medium with-Leu/-Trp), to select for transformants containing the introduced
plasmids for 3 days. Transformed colonies were dropped onto TDO medium (Triple
dropout medium: SD/–His/–Leu/–Trp), TDO/3-AT medium (SD/–His/–Leu/–Trp
supplemented with 3-amino-1,2,4-triazole from Avocado Lot No. H7734B), QDO
medium (minimal media quadruple dropouts, SD medium with-Leu/-Trp/-Ade/-His),
and QDO/X medium (SD medium with-Leu/-Trp/-Ade/-His) containing 4 mg mL-1
X-α-Gal to test for possible interactions between FLS2 and LecRK-V.2 or FLS2 and
LecRK-VII.1 or BAK1 and LecRK-V.2 or BAK1 and LecRK-VII.1 according to their
growth status.
Results
LecRK-V.2 and LecRK-VII.1 associate with FLS2 in vivo
Former members of the Zimmerli laboratory found that both lecrk-V.2 and
lecrk-VII.1 mutant plants are more susceptible to Pst DC3000 than Col-0 wild-type,
while LecRK-V.2 and LecRK-VII.1 overexpression lines are more resistant. We thus
speculated that LecRK-V.2 and LecRK-VII.1 are involved in the flagellin-triggered
immunity defense response. Flagellin is a bacterial MAMP that can be recognized by
the plant PRR, FLS2. We thus asked whether LecRK-V.2 and LecRK-VII.1 associate
with FLS2. Toward this goal, I performed Co-IP analyses in Arabidopsis protoplasts.
In this assay, the plasmid of FLS2-HA was co-transformed with BAK1-GFP,
LecRK-V.2-GFP, LecRK-VII.1-GFP, the GFP-empty vector, or RCI2B-GFP.
Immunoprecipitation was conducted with GFP-Trap beads to immunoprecipitate either
BAK1, LecRK-V.2, LecRK-VII.1, the GFP-empty vector, or RCI2B proteins and
western blot analyses were performed with anti-GFP and anti-HA antibodies. Using
this approach, I showed that BAK1 associates with FLS2 after the treatment of flg22,
but not before the treatment of flg22, confirming previous studies (Chinchilla et al.
2006, Chinchilla, Zipfel, Robatzek, Kemmerling, Nurnberger, Jones, Felix and Boller
2007), and thus suggesting that our experimental conditions were appropriate (Figure
1). By contrast, LecRK-V.2 and LecRK-VII.1 could associate with FLS2 before and
after treatment with flg22 (Figure 1). These results suggest that LecRK-V.2 and
LecRK-VII.1 are already in the FLS2 complex even before treatment with flg22. Upon
perception of flg22, FLS2 will recruit BAK1 into the complex and still associate with
LecRK-V.2 and LecRK-VII.1. These data suggest that LecRK-V.2 and LecRK-VII.1
are part of the FLS2 protein complex that are involved in the flg22-triggered immunity
response.
LecRK-VII.1 could directly interact with FLS2 through their kinase domain, but
LecRK-V.2 could not.
To further understand the mechanism and interaction between FLS2, LecRK-V.2, and
LecRK-VII.1, I used the yeast two-hybrid assay with both LecRK proteins as baits. The
kinase domain of FLS2 containing amino acids 219-1173, the kinase domain of
LecRK-V.2 containing amino acids 299-656, and the kinase domain of LecRK-VII.1
containing amino acids 309-686 were used to perform the yeast two-hybrid assay.
LecRK-V.2 could not directly interact with FLS2 through its kinase domain in yeast
two-hybrid assay (Figure 2). However, the kinase domain of LecRK-VII.1 could
interact with the kinase domain of FLS2 when analyzed with the yeast two-hybrid
system (Figure 2). PGBKT7-53 that encodes the murine p53 and pGADT7-T that
encodes the SV40 large T-antigen were used as positive control, because they are
known to interact in the yeast two-hybrid assay (Li and Fields 1993, Mokdad-Gargouri
et al. 2001). pGBKT7-Lam (which encodes the lamin proteins) and pGADT7-T were
used as negative controls (Li and Fields 1993).
LecRK-V.2 and LecRK-VII.1 associate with BAK1 in vivo
BAK1 is a membrane protein and is the co-receptor of FLS2 to sense the MAMP
flagellin (Chinchilla, Bauer, Regenass, Boller and Felix 2006, Chinchilla, Zipfel,
Robatzek, Kemmerling, Nurnberger, Jones, Felix and Boller 2007). During perception
of flagellin, BAK1 is recruited to the FLS2 complex and associate with FLS2 (Sun et al.
2013b). I therefore tested the interaction between BAK1 and LecRK-V.2 or
LecRK-VII.1 with the Co-IP approach in Arabidopsis protoplasts. In this assay, the
plasmid of BAK1-HA was co-transformed with LecRK-V.2-GFP, or LecRK-VII.1-GFP,
or the GFP-empty vector, or RCI2B-GFP. Immunoprecipitation was conducted with
GFP-Trap beads to immunoprecipitate LecRK-V.2, LecRK-VII.1, GFP-empty vector, or
RCI2B proteins and immunoblot analyses were performed with anti-GFP and anti-HA
antibodies. BAK1 associated with both LecRK-V.2 and LecRK-VII.1 before and after
flg22 treatment (Figure 3). As a negative control, I analyzed the association of BAK1
with RCI2B-GFP or only GFP-empty vector by immunoprecipitating equal amounts of
RCI2B-GFP or GFP-empty vector with GFP-Trap beads and by analyzing the presence
of BAK1-HAusing anti-HA immunoblotting. BAK1 could not be detected, suggesting
that they do not associate with GFP at the plasma membrane. Taken together, these data
suggested that BAK1 already associated with LecRK-V.2 and LecRK-VII.1 before
perception of flg22. After the treatment of flg22, BAK1 still associated with LecRK-V.2
and LecRK-VII.1. LecRK-V.2 and LecRK-VII.1 may be involved in the flg22-triggered
innate immunity response through association with the FLS2/BAK1 complex.
In vitro, BAK1 can directly interact with LecRK-V.2 and LecRK-VII.1 through
their kinase domain
Co-IP assays in Arabidopsis protoplast suggested that BAK1, LecRK-V.2, and
LecRK-VII.1 are in the same protein complex before and after treatment with flg22. To
further analyze the interaction mechanism between BAK1, LecRK-V.2, and
LecRK-VII.1, The yeast two-hybrid assay was used with the kinase domain of BAK1 as
bait. To perform the yeast two-hybrid assay, the clones that contain the kinase domain
of BAK1 from the amino acid 158 to 663, the kinase domain of LecRK-V.2 from the
amino acid 299 to 656, and the kinase domain of LecRK-VII.1 from the amino acid 309
to 686 were used. BAK1 was found to directly interact with both LecRK-V.2 and
LecRK-VII.1 through their kinase domains when analyzed with the yeast two-hybrid
assay (Figure 4). As with previous results, pGBKT7-53 and pGADT7-T were used as
positive control, because they interact in the yeast two-hybrid assay (Li and Fields 1993,
Mokdad-Gargouri, Belhadj and Gargouri 2001). The negative control was performed by
using pGBKT7-Lam (which encodes lamin proteins) and pGADT7-T (Li and Fields
1993).
In vitro and in vivo association analyses of LecRK-V.2 and LecRK-VII.1
LecRK-V.2 and LecRK-VII.1 are involved in the FLS2/BAK1 complex
independently of flagellin. Next, I focused on elucidating the association and the
interaction between LecRK-V.2 and LecRK-VII.1. First, co-transient expression in
Arabidopsis protoplasts and Co-IP of proteins were used. In this assay,
LecRK-V.2-HA was co-transformed with LecRK-VII.1-GFP, the GFP-empty vector,
and RCI2B-GFP. Immunoprecipitation was conducted with GFP-Trap beads to
immunoprecipitate the GFP-tagged proteins and immunoblot analyses were performed
with anti-GFP and anti-HA antibodies. LecRK-V.2 could associate with LecRK-VII.1
before and after flg22 treatment (Figure 5). As a negative control, I analyzed the
association of LecRK-V.2 with RCI2B-GF or the GFP-empty vector by
immunoprecipitating equal amounts of RCI2B-GFP and the GFP-empty vector with
GFP-Trap beads and by analyzing the presence of BAK1-HA using anti-HA
immunoblotting. LecRK-V.2 could not be detected, suggesting that LecRK-V.2 does
not associate non-selectively with GFP at the plasma membrane. Second, in order to
know whether LecRK-V.2 can directly interact with LecRK-VII.1, the yeast
two-hybrid assay was performed with LecRK-V.2 proteins as baits. To perform this
assay, the clones containing the kinase domain of LecRK-V.2 from the amino acid 299
to 656, and the kinase domain of LecRK-VII.1 from the amino acid 309 to 686 were
used. In this assay, pGBKT7-53 and pGADT7-T were used as positive control (Li and
Fields 1993, Mokdad-Gargouri, Belhadj and Gargouri 2001). A negative control was
performed by using pGBKT7-Lam and pGADT7-T (Li and Fields 1993). LecRK-V.2
did not directly interact with LecRK-VII.1 through its kinase domain when analyzed
with the yeast two-hybrid assay (Figure 6). Taken together these data suggest that
LecRK-V.2 associate with LecRK-VII.1, but they cannot directly interact with each
other through their kinase domain.
Each LecRKs may not affect the association of FLS2 with the other LecRK
To test whether one LecRK affects the other LecRK association with FLS2, the
co-transient expression assay in Arabidopsis protoplasts of Col-0, lecrk-V.2, and
lecrk-VII.1 T-DNA insertion mutant plants was performed. In this Co-IP assay, the
FLS2-HA construct was used to co-transform with BAK1-GFP, LecRK-V.2-GFP,
LecRK-VII.1-GFP, or the GFP-empty vector. Immunoprecipitation was conducted
with GFP-Trap beads to immunoprecipitate the GFP-tagged proteins and immunoblot
analyses were performed with anti-GFP and anti-HA antibodies. In lecrk-V.2 mutant
protoplasts, the association between FLS2 and LecRK-VII.1 was still observed (Figure
7). Similarly, the association between FLS2 and LecRK-V.2 was still observed in the
lecrk-VII.1 mutant background (Figure 8). Together these data suggest that FLS2 and
LecRK-V.2 or LecRK-VII.1 are still in the same complex before and after treatment
with flg22 even when one of the LecRK is missing. These data suggested that the
association of FLS2 and the LecRKs is independent from the presence of the other
LecRK. From the data of BiFC, it shows that FLS2 cannot interact directly with
LecRK-VII.1 in the lecrk-V.2 mutant protoplasts and vice-versa. We proposed that one
LecRK affects the direct interaction between FLS2 and the other LecRK, but they are
still in the same FLS2 PRR-complex.
Discussion
LecRK-V.2 and LecRK-VII.1 associate with FLS2
FLS2 and BAK1 interact and form an immune-complex to induce the
flagellin-triggered immunity pathway upon perception of flagellin (Chinchilla, Zipfel,
Robatzek, Kemmerling, Nurnberger, Jones, Felix and Boller 2007). Previous studies
from the Zimmerli laboratory showed that LecRK-V.2 and LecRK-VII.1 are involved in
the resistance against Pst DC3000 and are also involved in the regulation of stomatal
immunity (Supplemental Figure 1 and 2). We hypothesized that LecRK-V.2 and
LecRK-VII.1 participate in the FLS2 PRR complex to modulate the flagellin-triggered
immunity responses. First, we focused on the flagellin receptor, FLS2. Because
LecRK-V.2 and LecRK-VII.1 are membrane-localized proteins, they may affect
flagellin-triggered immunity pathway by interacting with FLS2. From previous BiFC
data, it was shown that FLS2 can interact with LecRK-V.2 and LecRK-VII.1 only after
treatment with flg22 but no interactions were observed prior to flg22 treatment. By
contrast, this work shows that FLS2 associates with LecRK-V.2 and LecRK-II.1 before
and after treatment with flg22 when analyzed by Co-IP in Arabidopsis protoplast. It is
known that BAK1 interacts with FLS2 in a ligand-dependent manner (Chinchilla,
Zipfel, Robatzek, Kemmerling, Nurnberger, Jones, Felix and Boller 2007). We propose
that LecRK-V.2 and LecRK-VII.1 are part of the FLS2 receptor complex already
before flg22 elicitation, but direct interact with FLS2 is flg22 dependent. It is known
that BIK1 interacts with FLS2, but dissociate in the presence of flagellin and BIK1 will
be phosphorylated after flg22 treatment (REF needed). After the dissociation from the
FLS2 complex, BIK1 will directly phosphorylate NADPH oxidase RbohD at specific
sites in a calcium-independent manner to enhance ROS generation (Kadota, Sklenar,
Derbyshire, Stransfeld, Asai, Ntoukakis, Jones, Shirasu, Menke, Jones and Zipfel
2014, Laluk et al. 2011, Li, Li, Yu, Zhou, Liang, Liu, Cai, Gao, Zhang, Wang, Chen
and Zhou 2014). Our data showed that LecRK-V.2 and LecRK-VII.1 can interact with
FLS2 through their kinase domain. LecRK-V.2 and LecRK-VII.1 may act like BIK1
and have the modification, such as phosphorylation, with FLS2, and then affect the
NADPH oxidase, ion-pumps or stomatal membrane proteins, such as Open Stomata 1
(OST1) (Montillet et al. 2013, Shang et al. 2016), to regulate stomatal immunity.
BAK1 associates and interacts through kinase domain with LecRK-V.2 and
LecRK-VII.1
BAK1 is another important receptor in Arabidopsis. Notably, BAK1 is involved in
hormone signaling and also in MTI (Chinchilla et al. 2009, Heese, Hann,
Gimenez-Ibanez, Jones, He, Li, Schroeder, Peck and Rathjen 2007). BAK1 interacts
with FLS2 after the perception of flg22 (Chinchilla, Bauer, Regenass, Boller and Felix
2006, Chinchilla, Zipfel, Robatzek, Kemmerling, Nurnberger, Jones, Felix and Boller
2007, Sun, Li, Macho, Han, Hu, Zipfel, Zhou and Chai 2013b). Previous BiFC data
suggest that BAK1 can interact with LecRK-V.2 and LecRK-VII.1 independently of
flg22 treatment. By Co-IP however, BAK1 associates with LecRK-V.2 and
LecRK-VII.1 before and after elicitation with flg22. BAK1 is known to interact with
BRI1 in the presence of brassinosteroids (BRs) and dissociates upon flagellin
perception (Kim et al. 2013, Sun et al. 2013a). Both LecRKs interact with BAK1 prior
to and after treatment with flg22, so both LecRKs are not only important for the
flagellin-sensing receptor, FLS2, but also important for the flagellin-sensing
co-receptor, BAK1. BAK1 unidirectionally phosphorylates BIR2 and BAK1 and BIR2
interactions are kinase-activity dependent (Blaum et al. 2014, Halter et al. 2014). From
our yeast two-hybrid results, BAK1 can directly interact with LecRK-V.2 and
LecRK-VII.1 through its kinase domain. BAK1 and both LecRKs may thus
phosphorylate each others and regulate the downstream signals during perception of
flg22. Further studies and researches are needed to clarify this point.
LecRK-V.2 cannot interact with LecRK-VII.1
Because LecRK-V.2 and LecRK-VII.1 are both membrane-localized proteins and
they associate or interact with FLS2 and BAK1, we asked whether LecRKV-2 and
LecRK-VII.1 associate or directly interact together. From previous protein-protein
interaction experiments, we knew that LecRK-V.2 and LecRK-VII.1 associate with
FLS2 and BAK1 independently of flg22 perception. We thus expected that both
LecRKs will associate together prior to and after treatment with flg22. By in vivo
Co-IP assay, LecRK-V.2 was shown to associate with LecRK-VII.1. It indicates that
both LecRKs are involved in the same FLS2 PRR complex. The kinase domains
interaction of membrane proteins may involve in the phosphorylated modification
(Blaum, Mazzotta, Noldeke, Halter, Madlung, Kemmerling and Stehle 2014, Halter,
Imkampe, Mazzotta, Wierzba, Postel, Bucherl, Kiefer, Stahl, Chinchilla, Wang,
Nurnberger, Zipfel, Clouse, Borst, Boeren, de Vries, Tax and Kemmerling 2014,
Kadota, Sklenar, Derbyshire, Stransfeld, Asai, Ntoukakis, Jones, Shirasu, Menke,
Jones and Zipfel 2014, Li, Li, Yu, Zhou, Liang, Liu, Cai, Gao, Zhang, Wang, Chen and
Zhou 2014). But LecRK-V.2 cannot interact directly through their kinase domain with
LecRK-VII.1 by performing in vitro yeast two-hybrid, suggesting that both LecRKs
may not phosphorylate each other.
LecRK-V.2 and LecRK-VII.1 are involved in FLS2-BAK1 complex
In summary, LecRK-V.2 and LecRK-VII.1 affect the PTI pathway such as the
stomatal closure by interacting with FLS2 or BAK1 upon perception of flg22.
LecRK-V.2 and LecRK-VII.1 may have the modification with FLS2 and BAK1 and
then affect the downstream stomatal closure and the resistance against Pst DC3000
bacteria and even the perception of flg22. In this thesis works, I clarified the
relationships between FLS2, BAK1, LecRK-V.2, and LecRK-VII.1. These new data
helped us to unravel the complex model of LecRK-V.2 and LecRK-VII.1 involvement
in the FLS2/BAK1 immune-complex.
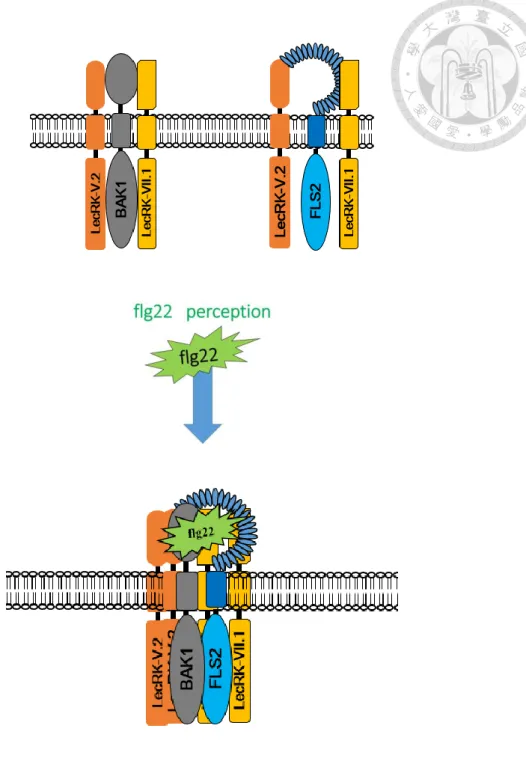
There is proposed model to conclude our results (Figure 9). LecRK-V.2 and
LecRK-VII.1 interact with BAK1 but not FLS2 before the treatment of flg22. But both
LecRKs associate with FLS2 and BAK1 before the treatment of flg22. Upon
perception of flg22, FLS2 recruits LecRK-V.2, LecRK-VII.1, and BAK1. The
involvement of both LecRKs and FLS2/BAK1 complex may play the key role to
regulate stomatal immunity when the perception of flg22.
Conclusions and Future Perspectives
LecRKs contain extracellular lectin domain, transmembrane domain, and
intracellular kinase domain (Bouwmeester and Govers 2009, Senchou et al. 2004). The
extracellular lectin domain is suggested to be critical for monosaccharide binding or
ligand binding and then trigger the downstream signals (Vaid, Macovei and Tuteja
2013). LecRK-I.9 can bind to extracellular ATP and is required for the ATP-induced
calcium response, mitogen-activated protein kinase activation, and gene expression
(Cao, Tanaka, Nguyen and Stacey 2014, Choi, Tanaka, Cao, Qi, Qiu, Liang, Lee and
Stacey 2014). From previous studies, the extracellular domain of BAK1 and FLS2
interactions can proceed independently of intracellular domain interactions upon the
perception of flg22 (Koller and Bent 2014). The extracellular domains of FLS2 and
BAK1 are important for defense signaling activation independently of intracellular
kinase domain. So we proposed that the extracellular domain of LecRK-V.2 and
LecRK-VII.1 may play the key role in the perception of flg22 together with FLS2. We
can try to isolate only both LecRKs kinase domains and check their sensing of flg22
will be affected or not.
Upon perception of flg22, BIK1 is phosphorylated and dissociates from the
FLS2/BAK1 complex. Then BIK1 phosphorylates the NADPH oxidase, RbohD, at
specific sites in a calcium-independent manner to enhance ROS generation to trigger
plant immunity against pathogens (Kadota, Sklenar, Derbyshire, Stransfeld, Asai,
Ntoukakis, Jones, Shirasu, Menke, Jones and Zipfel 2014, Laluk, Luo, Chai, Dhawan,
Lai and Mengiste 2011, Li, Li, Yu, Zhou, Liang, Liu, Cai, Gao, Zhang, Wang, Chen
and Zhou 2014). Besides, BAK1 can unidirectionally phosphorylate BIR2 and
negatively regulate MTI (Blaum, Mazzotta, Noldeke, Halter, Madlung, Kemmerling
and Stehle 2014, Halter, Imkampe, Mazzotta, Wierzba, Postel, Bucherl, Kiefer, Stahl,
Chinchilla, Wang, Nurnberger, Zipfel, Clouse, Borst, Boeren, de Vries, Tax and
Kemmerling 2014). It was found that both LecRKs can directly interact with FLS2
after the treatment of flg22. Both LecRKs can interact with BAK1, and both LecRKs
were involved in stomatal immunity. Hence, both LecRKs may be involved in the
phosphorylated modification between FLS2, BAK1, and BIK1, and then affect
stomatal closure. It would be interesting to perform the kinase assay to clarify the
phosphorylated modification between FLS2, BAK1, and both LecRKs. This should
help to understand the functional relationships between members of the FLS2 complex
during stomatal immunity.
Figures
Figure 1:FLS2 associate with LecRK-V.2 and LecRK-VII.1 when analyzed by
co-immunoprecipitation in Arabidopsis protoplasts.
FLS2-HA was co-transformed with BAK1-GFP, LecRK-V.2-GFP, LecRK-VII.1-GFP,
GFP empty vector, and RCI2B-GFP into Arabidopsis protoplasts. Total proteins
(Input) were subjected to immunoprecipitation (IP) with GFP-Trap beads followed by
immunoblot analysis with anti-HA to detect FLS2-HA. EV-GFP and RCI2B-GFP were
used as negative control to confirm that FLS2-HA did not stick to GFP beads or
associate with GFP at the plasma membrane. The protoplasts were treated with or
without 1 μM flg22 for 10 minutes. Experiments were repeated three times with the
similar results.
Figure 2:In vitro FLS2 direct interaction with LecRK-VII.1 kinase domains but
not LecRK-V.2.
Yeast two-hybrid assays with the kinase domains of FLS2, LecRK-V.2, and
LecRK-VII.1. Three consecutive dilutions on selection media lacking the amino acids
leucine, tryptophan, histidine and adenine are shown; growth on medium lacking
leucine and tryptophan assures proper growth of transformed yeast. For the selection of
the interacted yeasts, we use the selective dropout media lacking the amino acids
leucine, tryptophan, and histidine but containing 5 mM, and 15 mM 3-aminotriazole
(3-AT); and the selective dropout media lacking the amino acids leucine, tryptophan,
histidine and adenine but containing X-α-Gal. These experiments were repeated three
times with the similar results.
Figure 3:BAK1 associates with LecRK-V.2 and LecRK-VII.1 when analyzed by
co-immunoprecipitation in Arabidopsis protoplasts.
BAK1-HA was co-transformed with LecRK-V.2-GFP, LecRK-VII.1-GFP, GFP empty
vector, and RCI2B into Arabidopsis protoplasts. Total proteins (Input) were subjected
to immunoprecipitation (IP) with GFP-Trap beads followed by immunoblot analysis
with anti-HA to detect BAK1-HA. EV-GFP and RCI2B-GFP were used as negative
control to confirm that BAK1-HA did not stick to GFP beads or associate with GFP at
minutes. EV, empty vector. Experiments were repeated three times with the similar
results.
Figure 4:BAK1 can directly interact with LecRK-V.2 and LecRK-VII.1 through
its kinase domain.
Yeast two-hybrid assays with the kinase domains of BAK1, LecRK-V.2, and
LecRK-VII.1. Three consecutive dilutions on selection media lacking the amino acids
leucine, tryptophan, histidine and adenine are shown; growth on medium lacking
leucine and tryptophan assures proper growth of transformed yeast. For the selection of
the interacted yeasts, we use the selective dropout media lacking the amino acids
leucine, tryptophan, and histidine but containing 5 mM, and 15 mM 3-aminotriazole
(3-AT) are shown; the selective dropout media lacking the amino acids leucine,
tryptophan, histidine and adenine but containing X-α-Gal. These experiments were
repeated three times with the similar results.
Figure 5: Co-immunoprecipitation analyses of LecRK-V.2 association with
LecRK-VII.1 in Arabidopsis protoplasts.
LecRK-V.2-HA was co-transformed with LecRK-VII.1-GFP, GFP empty vector, and
RCI2B-GFP into Arabidopsis protoplasts. Total proteins (Input) were subjected to
immunoprecipitation (IP) with GFP-Trap beads followed by immunoblot analysis with
anti-HA to detect LecRK-V.2-HA. EV-GFP and RCI2B-GFP were used as negative
control to confirm that LecRK-V.2-HA did not stick to GFP beads or associate with
GFP at the plasma membrane. The protoplasts were treated with or without 1 μM flg22
for 10 minutes. EV, empty vector. Experiments were repeated three times with the
similar results.
Figure 6: LecRK-V.2 cannot directly interact with LecRK-VII.1 through their
kinase domain when analyzed with the yeast two hybrid assay.
Yeast two-hybrid assays with the kinase domains of LecRK-V.2 and LecRK-VII.1.
Three consecutive dilutions on selection media lacking the amino acids leucine,
tryptophan, histidine and adenine are shown; growth on medium lacking leucine and
tryptophan assures proper growth of transformed yeast. For the selection of the
interacted yeasts, we use the selective dropout media lacking the amino acids leucine,
tryptophan, and histidine but containing 5 mM, and 15 mM 3-aminotriazole (3-AT) are
shown; the selective dropout media lacking the amino acids leucine, tryptophan,
histidine and adenine but containing X-α-Gal. These experiments were repeated three
times with the similar results.
Figure 7: FLS2 associates with BAK1 and LecRK-VII.1 in the lecrk-V.2 mutant
background.
FLS2-HA was co-transformed with BAK1-GFP and LecRK-VII.1-GFP into
Arabidopsis protoplasts. Immunoprecipitation was conducted against anti-GFP
antibodies to immunoprecipitate the FLS2 proteins and immunoblot analyses were
performed with anti-GFP and anti-HA antibodies. The protoplasts were treated with or
without 1 μM flg22 for 10 minutes. Experiments were repeated three times with the
similar results.
Figure 8: FLS2 associates with BAK1 and LecRK-V.2 in the lecrk-VII.1 mutant
background.
FLS2-HA was co-transformed with BAK1-GFP and LecRK-V.2-GFP into Arabidopsis
protoplasts. Immunoprecipitation was conducted against anti-GFP antibodies to pull
down the FLS2 proteins and western blot analyses were performed with anti-GFP and
anti-HA antibodies. The protoplasts were treated with or without 1 μM flg22 for 10
minutes. Experiments were repeated three times with the similar results.
Figure 9:Proposed model
LecRK-V.2 and LecRK-VII.1 can associate with BAK1 and FLS2 before the treatment
of flg22. Upon the perception of flg22, both LecRKs and BAK1 can interact and
associate with FLS2 and form the immune-complex.
Tables
Table 1 :Primers for kinase domain constructs
Gene Primers Sequence (5’ - 3’)
FLS2 FLS2 KD Fp CACCTTACCGGATTTGGAT
FLS2 FLS2 KD Rp CTAAACTTCTCGATCCTCGTTACG
BAK1 BAK1 KD Fp CACCCCTACACCGCCAT
BAK1 BAK1 KD Rp TTATCTTGGACCCGAGGGG
LecRK-V.2 LecRK-V.2-KD Fp GGATCCTTGAAGAGGAAGAAGTT
LecRK-V.2 LecRK-V.2-KD Rp CTCGAGTTAGCGTCCACTAGAGA
LecRK-VII.1 LecRK-VII.1-KD Fp GGATCCAGAAAGAGATTAGAGAGG
LecRK-VII.1 LecRK-VII.1-KD Rp CTCGAGTCACCTCCCTTCTAAAA
Table 2 :List of abbreviations
Abbreviation Full name
PAMPs/MAMPs Pathogen/microbe-associated molecular patterns
PRRs Pattern recognition receptors
PTI/MTI PAMPs/MAMPs-triggered immunity
LPS Lipopolysaccharides
PGN Peptidoglycans
FLS2 Flagellin-sensing 2
BAK1 Brassinosteroid insensitive 1-associated kinase 1
BIK1 Botrytis-induced kinase 1
RLKs Receptor-lie kinase
Pst DC3000 Pseudomonas syringae pv. tomato DC3000
BiFC Bimolecular fluorescence complementation
Co-IP Co-immunoprecipitation
LiAc Lithium acetate
PEG Polyethylene glycol
SD medium Synthetic dropout medium
3-AT 3-amino-1,2,4-triazole
Supplementary Figures
Supplemental Figure 1: LecRK-V.2 and LecRK-VII.1 are necessary for the
resistance against to Pst.DC3000.
(A) The disease symptoms were observed 2-3 days after dip-inoculation with Pst
DC3000. (B) The bacterial growth were assessed 2-3 days after dip-inoculation with 5
x 106 cfu•ml-1 Pst DC3000 in Col-0 and lecrk mutants. Bacterial titers were evaluated
at 3d after inoculation (dpi). Values are the means ± SD of three biological replicates
(n = 9). Asterisks indicate a significant difference to Col-0 wild-type control based on
a t test (P <0.01). This experiments was repeated by Ph.D student, Shweta Bagul and
the other former members in the Zimmerli laboratory.
Supplemental Figure 2: LecRK-V.2 and LecRK-VII.1 are necessary for stomatal
immunity.
(A and B) The stomata aperture from the epidermal peels of Col-0, lecrk-V.2, and
lecrk-VII.1 mutant lines. Epidermal peels were exposed to MES buffer (Mock) or 5 x
106 cfu•mL-1 Pst DC3000 for 1.5 or 3 hours. (C) The stomatal aperture from the
epidermal peels of Col-0, lecrk-V.2, and lecrk-VII.1 mutant lines after 3 hours of
incubation in flg22 with 0 (Mock), 0.5, 1 or 5 μM, respectively.
Supplemental Figure 3:The PTI apoplastic responses and regulations in lecrk-V.2
and lecrk-VII.1 mutant lines.
(A and B) The reactive oxygen species productions in Arabidopsis leaves of Col-0,
lecrk-V.2 and lecrk-VII.1 mutant lines after treatment with 1 μM flg22 as relative light
units (RLU). The bak1-4 mutant line is used as negative control. Values are the means
± SE (n = 6). (C) Callose deposition in Col-0, lecrk-V.2 and lecrk-VII.1 mutant leaves
9 hours after infiltration with 10 mM MgSO4 (Mock) or 1 x 108 cfu•mL-1 Pst DC3000
hrcC. Data are means ± SD of callose deposits per square millimeter (n = 9).
Differences among Col-0, lecrk-V.2 and lecrk-VII.1 mutant were not significantly
different base on a t test (P < 0.01). Bar = 400μm. (D) FRK1 expression levels in
10 mM MgSO4 (Mock) or 1 x 108 cfu•mL-1 Pst DC3000 hrcC (h). Transcripts levels
determined by qRT-PCR and normalized to both EF-1 and UBQ10 were compared to
Col-0 mock at 45 minutes (defined value of 1). Three independent biological replicated
performed in triplicate were average ± SD (n = 9). Differences among Col-0,
lecrk-V.2 and lecrk-VII.1 mutant were not significantly different base on a t test (P <
0.01).
Supplemental Figure 4:LecRK-V.2-GFP and LecRK-VII.1 –GFP fusion proteins
localized on the membrane in Arabidopsis mesophyll protoplast.
LecRK-V.2-GFP and LecRK-VII.1–GFP fusion proteins were driven by the
cauliflower mosaic virus 35S promoter and transiently expressed in Arabidopsis
mesophyll protoplast. (A and D) GFP fluorescence channel. (B and E) Plasma
membrane marker pm-rk CD3-1007-mCherry fluorescence localization. (C and F)
Overlapping images of the GFP, plasma membrane marker and chlorophyll (red)
autofluorescence. (G) Protoplast expressing the GFP vector only as a GFP control.
Supplemental Figure 5:LecRK-V.2 and LecRK-VII.1 interact with FLS2 only
after the treatment with flg22.
In vivo, the interactions between FLS2 and LecRK-V.2 or FLS2 and LecRK-VII.1 are
determined by performing bimolecular fluorescence complementation (BiFC) analyses.
LecRK-V.2-GFP and LecRK-VII.1–GFP fusion proteins were under the control of the
cauliflower mosaic virus (CaMV) 35S promoter. FLS2-YFPN and BAK1-YFPC,
FLS2-YFPN and LecRK-V.2-YFPC, FLS2-YFPN and LecRK-VII.1-YFPC were
transiently co-transformed into Arabidopsis protoplast and with (+) or without (-) the
treatment of 1μM flg22 for 10 minutes. The red and yellow signals are
autofluorescence and YFP, respectively. There were the image of bright field and the
merge which is combined the autofluorescence, YFP, and bright field image.
References
Andre, S., Siebert, H.C., Nishiguchi, M., Tazaki, K. and Gabius, H.J. (2005) Evidence for lectin activity of a plant receptor-like protein kinase by
application of neoglycoproteins and bioinformatic algorithms. Biochim Biophys Acta, 1725: 222-232.
Arnaud, D., Desclos-Theveniau, M. and Zimmerli, L. (2012) Disease resistance to Pectobacterium carotovorum is negatively modulated by the Arabidopsis Lectin Receptor Kinase LecRK-V.5. Plant Signal Behav, 7: 1070-1072.
Blaum, B.S., Mazzotta, S., Noldeke, E.R., Halter, T., Madlung, J., Kemmerling, B.
and Stehle, T. (2014) Structure of the pseudokinase domain of BIR2, a regulator of BAK1-mediated immune signaling in Arabidopsis. J Struct Biol, 186: 112-121.
Blume, B., Nurnberger, T., Nass, N. and Scheel, D. (2000) Receptor-mediated increase in cytoplasmic free calcium required for activation of pathogen defense in parsley. The Plant Cell, 12: 1425-1440.
Bouwmeester, K. and Govers, F. (2009) Arabidopsis L-type lectin receptor kinases:
phylogeny, classification, and expression profiles. J Exp Bot, 60: 4383-4396.
Cambi, A., Koopman, M. and Figdor, C.G. (2005) How C-type lectins detect pathogens. Cell Microbiol, 7: 481-488.
Cao, Y., Tanaka, K., Nguyen, C.T. and Stacey, G. (2014) Extracellular ATP is a central signaling molecule in plant stress responses. Current Opinion in plant biology, 20: 82-87.
Chinchilla, D., Bauer, Z., Regenass, M., Boller, T. and Felix, G. (2006) The
Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. The Plant Cell, 18: 465-476.
Chinchilla, D., Shan, L., He, P., de Vries, S. and Kemmerling, B. (2009) One for all: the receptor-associated kinase BAK1. Trends in Plant Science, 14:
535-541.
Chinchilla, D., Zipfel, C., Robatzek, S., Kemmerling, B., Nurnberger, T., Jones, J.D., Felix, G. and Boller, T. (2007) A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature, 448: 497-500.
Choi, J., Tanaka, K., Cao, Y., Qi, Y., Qiu, J., Liang, Y., Lee, S.Y. and Stacey, G.
(2014) Identification of a plant receptor for extracellular ATP. Science (New York, N.Y.), 343: 290-294.
Deng, K., Wang, Q., Zeng, J., Guo, X., Zhao, X., Tang, D. and Liu, X. (2009) A Lectin Receptor Kinase Positively Regulates ABA Response During Seed Germination and Is Involved in Salt and Osmotic Stress Response. Journal of Plant Biology, 52: 493-500.
Desclos-Theveniau, M., Arnaud, D., Huang, T.Y., Lin, G.J., Chen, W.Y., Lin, Y.C. and Zimmerli, L. (2012) The Arabidopsis lectin receptor kinase
LecRK-V.5 represses stomatal immunity induced by Pseudomonas syringae pv.
tomato DC3000. PLoS Pathogens, 8: e1002513.
Gietz, D., St Jean, A., Woods, R.A. and Schiestl, R.H. (1992) Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research, 20:
1425.
Gomez-Gomez, L. and Boller, T. (2000) FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Molecular Cell, 5: 1003-1011.
Halter, T., Imkampe, J., Mazzotta, S., Wierzba, M., Postel, S., Bucherl, C., Kiefer, C., Stahl, M., Chinchilla, D., Wang, X., Nurnberger, T., Zipfel, C., Clouse, S., Borst, J.W., Boeren, S., de Vries, S.C., Tax, F. and Kemmerling, B.
(2014) The leucine-rich repeat receptor kinase BIR2 is a negative regulator of BAK1 in plant immunity. Curr Biol, 24: 134-143.
He, X.J., Zhang, Z.G., Yan, D.Q., Zhang, J.S. and Chen, S.Y. (2004) A
salt-responsive receptor-like kinase gene regulated by the ethylene signaling pathway encodes a plasma membrane serine/threonine kinase. Theor Appl Genet, 109: 377-383.
Heese, A., Hann, D.R., Gimenez-Ibanez, S., Jones, A.M., He, K., Li, J., Schroeder, J.I., Peck, S.C. and Rathjen, J.P. (2007) The receptor-like kinase
SERK3/BAK1 is a central regulator of innate immunity in plants. Proceedings of the National Academy of Sciences of the United States of America, 104:
12217-12222.
Hill, J., Donald, K.A. and Griffiths, D.E. (1991) DMSO-enhanced whole cell yeast transformation. Nucleic Acids Research, 19: 5791.
Ito, H., Fukuda, Y., Murata, K. and Kimura, A. (1983) Transformation of intact yeast cells treated with alkali cations. Journal of Bacteriology, 153: 163-168.
Jones, J.D. and Dangl, J.L. (2006) The plant immune system. Nature, 444: 323-329.
Kadota, Y., Sklenar, J., Derbyshire, P., Stransfeld, L., Asai, S., Ntoukakis, V., Jones, J.D., Shirasu, K., Menke, F., Jones, A. and Zipfel, C. (2014) Direct
regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity. Molecular Cell, 54: 43-55.
Kim, B.H., Kim, S.Y. and Nam, K.H. (2013) Assessing the diverse functions of BAK1 and its homologs in arabidopsis, beyond BR signaling and PTI responses. Mol Cells, 35: 7-16.
Koller, T. and Bent, A.F. (2014) FLS2-BAK1 extracellular domain interaction sites required for defense signaling activation. PloS One, 9: e111185.
Laluk, K., Luo, H., Chai, M., Dhawan, R., Lai, Z. and Mengiste, T. (2011) Biochemical and genetic requirements for function of the immune response regulator BOTRYTIS-INDUCED KINASE1 in plant growth, ethylene signaling, and PAMP-triggered immunity in Arabidopsis. The Plant Cell, 23:
2831-2849.
Lecourieux, D., Mazars, C., Pauly, N., Ranjeva, R. and Pugin, A. (2002) Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. The Plant Cell, 14: 2627-2641.
Li, B. and Fields, S. (1993) Identification of mutations in p53 that affect its binding to SV40 large T antigen by using the yeast two-hybrid system. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 7: 957-963.
Li, L., Li, M., Yu, L., Zhou, Z., Liang, X., Liu, Z., Cai, G., Gao, L., Zhang, X., Wang, Y., Chen, S. and Zhou, J.M. (2014) The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity.
Cell Host & Microbe, 15: 329-338.
Lu, D., Wu, S., Gao, X., Zhang, Y., Shan, L. and He, P. (2010) A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity. Proceedings of the National Academy of Sciences of the United States of America, 107: 496-501.
Mokdad-Gargouri, R., Belhadj, K. and Gargouri, A. (2001) Translational control of human p53 expression in yeast mediated by 5'-UTR-ORF structural interaction.
Nucleic Acids Research, 29: 1222-1227.
Monaghan, J. and Zipfel, C. (2012) Plant pattern recognition receptor complexes at the plasma membrane. Current Opinion in Plant Biology, 15: 349-357.
Montillet, J.L., Leonhardt, N., Mondy, S., Tranchimand, S., Rumeau, D., Boudsocq, M., Garcia, A.V., Douki, T., Bigeard, J., Lauriere, C., Chevalier, A., Castresana, C. and Hirt, H. (2013) An abscisic
acid-independent oxylipin pathway controls stomatal closure and immune defense in Arabidopsis. PLoS Biology, 11: e1001513.
Naithani, S., Chookajorn, T., Ripoll, D.R. and Nasrallah, J.B. (2007) Structural modules for receptor dimerization in the S-locus receptor kinase extracellular domain. Proceedings of the National Academy of Sciences of the United States of America, 104: 12211-12216.
Nuhse, T.S., Bottrill, A.R., Jones, A.M. and Peck, S.C. (2007) Quantitative phosphoproteomic analysis of plasma membrane proteins reveals regulatory mechanisms of plant innate immune responses. The Plant Journal : for cell and molecular biology, 51: 931-940.
Nuhse, T.S., Peck, S.C., Hirt, H. and Boller, T. (2000) Microbial elicitors induce activation and dual phosphorylation of the Arabidopsis thaliana MAPK 6. The Journal of Biological Chemistry, 275: 7521-7526.
Schiestl, R.H. and Gietz, R.D. (1989) High efficiency transformation of intact yeast cells using single stranded nucleic acids as a carrier. Current Genetics, 16:
339-346.
Senchou, V., Weide, R., Carrasco, A., Bouyssou, H., Pont-Lezica, R., Govers, F.
and Canut, H. (2004) High affinity recognition of a Phytophthora protein by Arabidopsis via an RGD motif. Cell Mol Life Sci, 61: 502-509.
Shang, Y., Dai, C., Lee, M.M., Kwak, J.M. and Nam, K.H. (2016) BRI1-Associated Receptor Kinase 1 Regulates Guard Cell ABA Signaling Mediated by Open Stomata 1 in Arabidopsis. Molecular Plant, 9: 447-460.
Singh, P., Kuo, Y.C., Mishra, S., Tsai, C.H., Chien, C.C., Chen, C.W.,
Desclos-Theveniau, M., Chu, P.W., Schulze, B., Chinchilla, D., Boller, T.
and Zimmerli, L. (2012) The lectin receptor kinase-VI.2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity. The Plant Cell, 24: 1256-1270.
Sun, Y., Han, Z., Tang, J., Hu, Z., Chai, C., Zhou, B. and Chai, J. (2013a) Structure reveals that BAK1 as a co-receptor recognizes the BRI1-bound brassinolide. Cell Res, 23: 1326-1329.
Sun, Y., Li, L., Macho, A.P., Han, Z., Hu, Z., Zipfel, C., Zhou, J.M. and Chai, J.
(2013b) Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science (New York, N.Y.), 342: 624-628.
Vaid, N., Macovei, A. and Tuteja, N. (2013) Knights in action: lectin receptor-like kinases in plant development and stress responses. Molecular Plant, 6:
1405-1418.